Embed Size (px)
Citation preview

S U R G E R YVolume 139 Number 6
JUNE 2006
Surgical research review
Is all fat the same? The role of fat inthe pathogenesis of the metabolicsyndrome and type 2 diabetes mellitusErik Hansen, MD, Tahar Hajri, PhD, and Naji N. Abumrad, MD, FACS, Nashville, Tennessee
From the Department of Surgery, Vanderbilt University School of Medicine, Nashville, Tennessee
SURGERY 711
THE PROBLEM OF OBESITY HAS REACHED epidemic propor-tions. Sixty-five percent of US adults are said to beoverweight (body mass index, >25 kg/m2).1 Globally,the overweight population is estimated to be 1 billionpeople.2 This epidemic has led to a resurgence of in-terest in adipose tissue metabolism, physiology, andpathophysiology. The adipocyte is no longer under-stood merely as a repository of fat but rather is a dy-namic, metabolically active factory that producesmetabolic substrates, hormones, cytokines, and adi-pokines that exert their actions locally, systemically,and even centrally at the hypothalamus to regulateoverall energy homeostasis. Increasingly, we under-stand that obesity is not only a problem of too muchfat, per se, but also a much more far-reaching dysregu-lation of metabolism that affects adipose tissue andother organs, such as the liver, muscle, and pancreas.
FAT STORES
Adipose tissue is considered the natural organfor large stores of fat. A limited pool of fatty acids
Accepted for publication October 20, 2005.
Reprint requests: Naji N. Abumrad, MD, FACS, D-4313 MedicalCenter North, 1161 21st Ave South, Vanderbilt University Schoolof Medicine, Nashville, TN 37232. E-mail: [email protected].
Surgery 2006;139:711-6.
0039-6060/$ - see front matter
� 2006 Mosby, Inc. All rights reserved.
doi:10.1016/j.surg.2005.10.018
can be found in other organs (such as the liver,muscle, and pancreas), where they are considered‘‘nontoxic’’ and are integral to cell function. Theidentification of non-differentiated cells (preadi-pocytes) confers to adipose tissue the potential toacquire new mature fat cells that appears to bepermanent. The differentiation of the preadipocyteinto an adipocyte is a complex event that is initiatedmainly by peroxisome proliferator-activated recep-tor-c and CCAAT/enhancer binding proteins.3
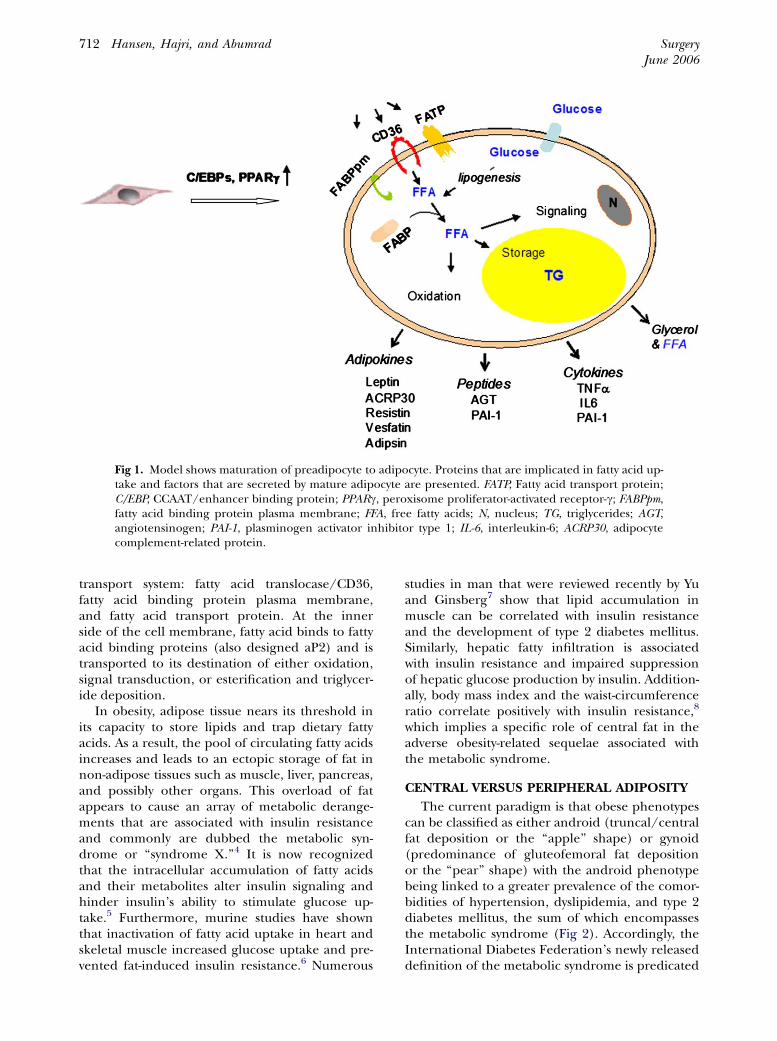
During this process, these nuclear factors triggerthe expression of multiple genes that are involvedin various steps of the adipogenesis, such as CD36(also called fatty acid translocase), fatty acid trans-port proteins, fatty acid binding proteins, and lip-oprotein lipase. Maturation of the adipocyte isassociated with the gain of new functions that in-clude lipogenic capacity and the appearance ofcytoplasmic lipid droplets, acquisition of insulinsensitivity and increased glucose uptake, and theexpression and secretion of numerous bioactivemolecules (Fig 1). Transport of fatty acids throughthe lipid bilayer is a major step that regulates fattyacid utilization. Although passive diffusion hasbeen described as the only mechanism of fattyacid transport through the cell membrane, theidentification of new proteins in the plasma mem-brane added another important component infatty acid transport. Several candidate proteinshave been identified to play a role in this
SurgeryJune 2006
712 Hansen, Hajri, and Abumrad
Fig 1. Model shows maturation of preadipocyte to adipocyte. Proteins that are implicated in fatty acid up-take and factors that are secreted by mature adipocyte are presented. FATP, Fatty acid transport protein;C/EBP, CCAAT/enhancer binding protein; PPARc, peroxisome proliferator-activated receptor-c; FABPpm,fatty acid binding protein plasma membrane; FFA, free fatty acids; N, nucleus; TG, triglycerides; AGT,angiotensinogen; PAI-1, plasminogen activator inhibitor type 1; IL-6, interleukin-6; ACRP30, adipocytecomplement-related protein.
transport system: fatty acid translocase/CD36,fatty acid binding protein plasma membrane,and fatty acid transport protein. At the innerside of the cell membrane, fatty acid binds to fattyacid binding proteins (also designed aP2) and istransported to its destination of either oxidation,signal transduction, or esterification and triglycer-ide deposition.
In obesity, adipose tissue nears its threshold inits capacity to store lipids and trap dietary fattyacids. As a result, the pool of circulating fatty acidsincreases and leads to an ectopic storage of fat innon-adipose tissues such as muscle, liver, pancreas,and possibly other organs. This overload of fatappears to cause an array of metabolic derange-ments that are associated with insulin resistanceand commonly are dubbed the metabolic syn-drome or ‘‘syndrome X.’’4 It is now recognizedthat the intracellular accumulation of fatty acidsand their metabolites alter insulin signaling andhinder insulin’s ability to stimulate glucose up-take.5 Furthermore, murine studies have shownthat inactivation of fatty acid uptake in heart andskeletal muscle increased glucose uptake and pre-vented fat-induced insulin resistance.6 Numerous
studies in man that were reviewed recently by Yuand Ginsberg7 show that lipid accumulation inmuscle can be correlated with insulin resistanceand the development of type 2 diabetes mellitus.Similarly, hepatic fatty infiltration is associatedwith insulin resistance and impaired suppressionof hepatic glucose production by insulin. Addition-ally, body mass index and the waist-circumferenceratio correlate positively with insulin resistance,8
which implies a specific role of central fat in theadverse obesity-related sequelae associated withthe metabolic syndrome.
CENTRAL VERSUS PERIPHERAL ADIPOSITY
The current paradigm is that obese phenotypescan be classified as either android (truncal/centralfat deposition or the ‘‘apple’’ shape) or gynoid(predominance of gluteofemoral fat depositionor the ‘‘pear’’ shape) with the android phenotypebeing linked to a greater prevalence of the comor-bidities of hypertension, dyslipidemia, and type 2diabetes mellitus, the sum of which encompassesthe metabolic syndrome (Fig 2). Accordingly, theInternational Diabetes Federation’s newly releaseddefinition of the metabolic syndrome is predicated

SurgeryVolume 139, Number 6
Hansen, Hajri, and Abumrad 713
Fig 2. Development of an android phenotype (truncal/central obesity) leads to insulin resistance.A gynoid habitus (gluteofemoral obesity) does not confer the same risk of insulin resistance.
on racial- and gender-specific criteria for centralobesity.9
Regional differences in fat depots are intriguingand far-reaching and involve processes such aslipolysis, lipogenesis, fatty acid uptake, and theexpression and secretion of hormones and inflam-matory factors. Relative to visceral adipose tissue,subcutaneous fat, for example, exhibits a higherbasal rate of lipolysis, hormone-sensitive lipase ac-tivity, anti-lipolytic action of insulin, expression ofinsulin receptor substrate 1 expression, lipoproteinlipase activity, and fatty uptake by preadipocytes.10
Additionally, expression of leptin (a pleiotropic hor-mone that, among other actions, regulates satietythrough its hypothalamic action) correlates posi-tively with the area of subcutaneous adipose tissue.Conversely, visceral adiposity exhibits a greater lipo-lytic response to catecholamines10 and a bluntedresponse to insulin’s anti-lipolytic effect withincreasing doses of insulin.11 This response mayaccount for the hypertriglyceridemia seen in themetabolic syndrome.
Visceral fat predominates in the productionof certain adipokines (eg, resistin, adiponectin),
inflammatory cytokines (eg, interleukin 6), andplasminogen activator inhibitor 1.10 This observa-tion provides a link between the low-grade, chronicinflammatory state of obesity and insulin resis-tance, in that resistin can induce insulin resistancein mice, and tumor necrosis factor-a (TNF-a) andinterleukin-6 interfere with insulin activity directlyby altering intracellular signaling and indirectlyby influencing leptin concentration. Furthermore,the increase in glucose and non-esterified fattyacid concentrations seen with diabetes mellituscan lead to b-cell loss, which further facilitatesthe onset of diabetes mellitus.12 Some investigatorshave postulated that the increase in visceral fat-derived, portally delivered non-esterified fattyacids to the liver is a primary mechanism in thedevelopment of impaired hepatic glucose metabo-lism. Ironically, visceral fat secretes the newlyidentified hormone, visfatin that exhibits insulin-like properties, which is a fact that, at best, ap-pears to mitigate the role of visceral fat in thedevelopment of insulin resistance.13 Visceral adi-pose tissue also expresses the angiotensinogengene,14 which provides a potential mechanism

SurgeryJune 2006
714 Hansen, Hajri, and Abumrad
for the hypertension that is seen in the metabolicsyndrome.
Visceral and subcutaneous fats differ also intheir morphologic condition and adipogeniccapacity. In obese premenopausal women, thegynoid habitus is associated with smaller and morenumerous fat cells than the android habitus, andvisceral adipocytes are smaller than subcutaneousadipocytes.15 Fat cells that are harvested fromobese patients who are undergoing gastric bypassexhibit 2 distinct populations of preadipocytes,rapidly and slowly replicating subtypes, both ofwhich were identified in subcutaneous, mesen-teric, and omental fat biopsy specimens, albeit indiffering proportions by depot.16 Relative to mes-enteric or subcutaneous fat, the slowly replicatingsubtype comprises a statistically greater proportionof preadipocytes, has a lower lipid-accumulatingcapacity, is more susceptible to TNF-a–inducedapoptosis, andexpresses lessCCAAT/enhancerbind-ing proteins a (adipogenic transcription factor). Infact, both preadipocyte subtypes accumulate lessfat in omental tissue than in subcutaneous tissue.Although not proved, it seems a logical conclusionthat visceral adipose tissue is ‘‘over-loaded’’ morereadily, thereby leading to untoward sequelae.
Studies that used animal models have shownthat the association of visceral fat with the meta-bolic syndrome is not absolute, however.17,18 Trans-genic expression of human growth hormone in thehypothalamus of the rat17 induced late onset obe-sity in the male rat that is mostly visceral in natureand due to adipocyte hyperplasia and not hyper-trophy, which suggests that the visceral fat in theseanimals has a greater storage potential because ofa greater number of adipocytes. However, the ani-mals have normal fasting blood glucose, enhancedinsulin sensitivity, and a lack of intrahepatocellularor intramyocellular fatty deposits. This evidenceindicates that abdominal obesity cannot be thesole, fundamental cause of the metabolic syndrome.Moreover, Korach-Andre et al18 questioned thecausal relationship between visceral adiposity andinsulin resistance in the muscle of rats underdietary and pharmacologic treatments.
Conversely, selective over-expression of 11b�hydroxysteroid dehydrogenase type 1 (the enzymeresponsible for the formation of active glucocorti-coids) in mouse adipose tissue increased the levelof corticosterone and induced visceral obesity thatwas associated with pronounced insulin-resistanceand hyperlipidemia.19 In humans and mice, eleva-tion of serum retinol binding protein 4 is seen inobesity. Serum retinol binding protein 4 impairsinsulin signaling in muscle and increases hepatic
glucose production, likely through an effect onphosphoenolpyruvate kinase.20 Taken together,these observations suggest that abdominal obesitymay not be the sole, fundamental cause of themetabolic syndrome. The onset of metabolic syn-drome is most likely the result of biochemical fac-tors either secreted or regulated by visceral fat thatacts on vulnerable target organs.
LIPODYSTROPHY
The various forms of lipodystrophy (acquiredand congenital) provide further support for thenotion that the obesity-related metabolic syn-drome is perhaps more an issue of storage capacitythan fat quantity or location (lipodystrophy), ingeneral, is a condition whereby a pathologic loss offat leads to a constellation of symptoms like themetabolic syndrome. Despite the heterogeneity ofthe various lipodystrophies, the resultant impair-ment in insulin sensitivity common to all formsleads to type 2 diabetes mellitus.
STRESS AND INFLAMMATION
The question now is how this putative triglycerideoverload leads to the constellation of symptomsseen in the metabolic syndrome. The answer may liein the body’s stress response. Weisberg et al21 andXu et al22 showed that, in obese humans, macro-phages can account for up to 40% of the tissuemass. Accumulation of macrophages in the subcuta-neous tissue appeared to plateau at an earlier levelthan that in visceral fat; these macrophages accountfor most of the overall secretion of TNF-a. Theseresults lead to a reconsideration of the role of theadipocytes (and preadipocytes) alone and to takeinto account the role of macrophage infiltrationinto adipose tissue as a source of the inflammatorymolecules.23
In 1990, Daniel et al24 discovered a group ofmolecules termed F2-isoprostanes, which arederived from free radical–catalyzed peroxidation ofarachidonic acid. These investigators documenteda very strong correlation between increased F2-isoprostanes and body mass index,25 which arefindings that were reproduced by Keany et al26 in2800 subjects who were enrolled in the Framing-ham study. Additional indirect support is obtainedfrom the very strong association of obesity withplatelet-activation, with more than a 3-fold increasein the excretion of platelet-derived thromboxanesin the urine of obese women.27 Furthermore,Ozcan et al28 studied murine models of obesity(diet-induced and genetic [obese mice]) and dem-onstrated a link between the stress of the endoplas-mic reticulum and the inhibition of insulin action

SurgeryVolume 139, Number 6
Hansen, Hajri, and Abumrad 715
that is mediated by c-Jun N-terminal kinase andinositol-requiring kinase-1a.
TARGETED LIPECTOMY
When the evidence for the role of ‘‘ectopic fat’’is considered and especially the differences invarious fat depots and the particularly detrimentalsequelae of central obesity in regard to coronaryand cerebrovascular disease, it seems logical tofocus on the ectopic visceral fat stores in thesetting of obesity as a means of mitigating riskfactors for stroke, heart attack, and even diabetesmellitus. Although the lipodystrophic modelwould suggest that excess fat removal could leadto a state of insulin resistance, perhaps there is arole for the selective removal of adipose tissue inthe setting of obesity-associated insulin resistance.
We have collected preliminary data in the dogmodel of the role of visceral fat on glucose metab-olism. Catheters in the hepatic vein, portal vein,and femoral artery were used to sample bloodduring hyperinsulinemic-euglycemic clamps. Westudied mongrel dogs under a stepwise hyperinsu-linemic-euglycemic clamp and achieved 4- and10-fold basal levels of insulin, respectively, beforeand after removal of visceral fat (‘‘omentectomy’’).Low-dose insulin infusion is meant to shut offhepatic glucose production; higher insulin infu-sion maximizes peripheral (skeletal muscle) glu-cose uptake. The removal of visceral fat resulted ina reduction in basal hepatic glucose production bynearly 40%. Furthermore, during the high insulininfusion, omentectomy resulted in a greater than2-fold increase from basal in peripheral glucose useby insulin-dependent tissue (predominantly skele-tal muscle). In like fashion, Barzilai et al29 showedthat in Zucker Diabetic Fatty rats, removing visceralfat delayed the onset of diabetes mellitus. Con-versely, Klein et al30 found no improvement in insu-lin resistance, glucose, blood pressure, or othercardiovascular risk factors in 15 obese women be-fore and after liposuction removal of subcutaneousabdominal fat in amounts equivalent to 9% of totalbody mass or 18% of total fat mass.
CONCLUSION
The proposition therefore is that fat, in and ofitself, may not be the fundamental problem, butrather it may be the out-stripping of the adipocytesability to handle a given triglyceride load that resultsin ‘‘ectopic fat storage’’31 and subsequent inflamma-tion that lead to the metabolic syndrome.32 Obesity,and specifically visceral obesity, could be considereda problem of excess triglycerides that leads to ec-topic fat deposition, which could potentially initiate
a stress response that leads to inflammation, excesscirculating triglycerides, the under production ofbeneficial adipokines such as adiponectin, and theover production of deleterious adipokines such asresistin, the sum of which leads to insulin resistanceand ultimately the metabolic syndrome. Under-standing the significant role that visceral fat playsin the metabolic syndrome, either because of itsmore limited capacity to handle an increasing tri-glyceride load or its preferential production ofdeleterious cytokines, in conjunction with our ownanimal data combined with that of Barzilai et al,29
shows that targeted lipectomy can mitigate the detri-mental effects of ectopic fat positively. To that end,we feel compelled to ask whether it would notmake sense to offer omentectomy alone as treat-ment for type 2 diabetes mellitus. This approacheffectively would remove a major source of portallydelivered non-esterified fatty acids and possiblycytokines and adipokines and result in a reductionin basal glucose production and a concomitantimprovement in glucose use by skeletal muscle,thus offering a potential cure for some of the majormetabolic derangements of type 2 diabetes mellitusand the metabolic syndrome. The procedure couldbe performed easily by trained laparoscopic sur-geons as an outpatient procedure and would obviatethe need for the chronic medical treatment ofdiabetes mellitus.
REFERENCES
1. Flegal KM, Carroll MD, Ogden CL, Johnson CL. Prevalenceand trends in obesity among US adults, 1999-2000. JAMA2002;288:1723-7.
2. Obesity and Overweight. World Health Report TRS916.Geneva/Rome. Diet, nutrition, and the prevention ofchronic diseases. World Health Organization; 2003.
3. Farmer SR. Regulation of PPARgamma activity duringadipogenesis. Int J Obes (Lond) 2005;29(suppl):S13-6.
4. Reaven GM. The insulin resistance syndrome: definitionand dietary approaches to treatment. Annu Rev Nutr2005;25:391-406.
5. Shulman GI. Cellular mechanisms of insulin resistance.J Clin Invest 2000;106:171-6.
6. Hajri T, Han XX, Bonen A, Abumrad NA. Defective fattyacid uptake modulates insulin responsiveness and meta-bolic responses to diet in CD36-null mice. J Clin Invest2002;109:1381-9.
7. Yu YH, Ginsberg HN. Adipocyte signaling and lipid homeo-stasis: sequelae of insulin-resistant adipose tissue. Circ Res2005;96:1042-52.
8. Clausen JO, Borch-Johnsen K, Ibsen H, Bergman RN,Hougaard P, Winther K, et al. Insulin sensitivity index,acute insulin response, and glucose effectiveness in apopulation-based sample of 380 young healthy Caucasians:analysis of the impact of gender, body fat, physical fitness,and life-style factors. J Clin Invest 1996;98:1195-209.
9. The IDF consensus worldwide definition of the metabolicsyndrome. 2005.

SurgeryJune 2006
716 Hansen, Hajri, and Abumrad
10. Lafontan M, Berlan M. Do regional differences in adipocytebiology provide new pathophysiological insights? TrendsPharmacol Sci 2003;24:276-83.
11. Meek SE, Nair KS, Jensen MD. Insulin regulation ofregional free fatty acid metabolism. Diabetes 1999;48:10-4.
12. Wellen KE, Hotamisligil GS. Inflammation, stress, and dia-betes. J Clin Invest 2005;115:1111-9.
13. Fukuhara A, Matsuda M, Nishizawa M, Segawa K, Tanaka M,Kishimoto K, et al. Visfatin: a protein secreted by visceralfat that mimics the effects of insulin. Science 2005;307:426-30.
14. Misra A, Vikram NK. Clinical and pathophysiological conse-quences of abdominal adiposity and abdominal adiposetissue depots. Nutrition 2003;19:457-66.
15. Garaulet M, Perex-Llamas F, Fuente T, Zamora S, Tebar FJ.Anthropometric, computed tomography and fat cell data inan obese population: relationship with insulin, leptin, tu-mor necrosis factor-alpha, sex hormone-binding globulinand sex hormones. Eur J Endocrinol 2000;143:657-66.
16. Tchkonia T, Tchoukalova YD, Giorgadze N, Pirtskhalava T,Karagiannides I, Forse RA, et al. Abundance of two humanpreadipocyte subtypes with distinct capacities for replica-tion, adipogenesis, and apoptosis varies among fat depots.Am J Physiol Endocrinol Metab 2005;288:E267-77.
17. Bains RK, Wells SE, Flavell DM, Fairhall KM, Strom M,Le Tissier P, et al. Visceral obesity without insulin resis-tance in late-onset obesity rats. Endocrinology 2004;145:2666-79.
18. Korach-Andre M, Gao J, Gounarides JS, Deacon R, Islam A,Laurent D. Relationship between visceral adiposity andintramyocellular lipid content in two rat models of insulin re-sistance. Am J Physiol Endocrinol Metab 2005;288:E106-16.
19. Masuzaki H, Paterson J, Shinyama H, Morton NM, MullinsJJ, Seckl JR, et al. A transgenic model of visceral obesityand the metabolic syndrome. Science 2001;294:2166-70.
20. Yang Q, Graham TE, Mody N, Preitner F, Peroni OD, Zabo-lotny JM, et al. Serum retinol binding protein 4 contributesto insulin resistance in obesity and type 2 diabetes. Nature2005;436:356-62.
21. Weisberg SP, McCann D, Desai M, Rosenbaum M, LeibelRL, Ferrante AW Jr. Obesity is associated with macrophage
accumulation in adipose tissue. J Clin Invest 2003;112:1796-808.
22. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, et al.Chronic inflammation in fat plays a crucial role in the devel-opment of obesity-related insulin resistance. J Clin Invest2003;112:1821-30.
23. Wellen KE, Hotamisligil GS. Obesity-induced inflamma-tory changes in adipose tissue. J Clin Invest 2003;112:1785-8.
24. Daniel VC, Minton TA, Brown NJ, Nadeau JH, MorrowJD. Simplified assay for the quantification of 2,3-dinor-6-keto-prostaglandin F1 alpha by gas chromatography-massspectrometry. J Chromatogr B Biomed Appl 1994;653:117-22.
25. Block G, Dietrich M, Norkus EP, Morrow JD, Hudes M,Caan B, et al. Factors associated with oxidative stress inhuman populations. Am J Epidemiol 2002;156:274-85.
26. Keaney JF Jr, Larson MG, Vasan RS, Wilson PW, Lipinska I,Corey D, et al. Obesity and systemic oxidative stress: clinicalcorrelates of oxidative stress in the Framingham Study.Arterioscler Thromb Vasc Biol 2003;23:434-9.
27. Davi G, Guagnano MT, Ciabattoni G, Basili S, Falco A,Marinopiccoli M, et al. Platelet activation in obese women:role of inflammation and oxidant stress. JAMA 2002;288:2008-14.
28. Ozcan U, Cao Q, Yilmaz E, Lee AH, Iwakoshi NN, OzdelenE, et al. Endoplasmic reticulum stress links obesity, insulinaction, and type 2 diabetes. Science 2004;306:457-61.
29. Barzilai N, She L, Liu BQ, Vuguin P, Cohen P, Wang J, et al.Surgical removal of visceral fat reverses hepatic insulin resis-tance. Diabetes 1999;48:94-8.
30. Klein S, Fontana L, Young VL, Coggan AR, Kilo C, PattersonBW, et al. Absence of an effect of liposuction on insulinaction and risk factors for coronary heart disease. N EnglJ Med 2004;350:2549-57.
31. Heilbronn L, Smith SR, Ravussin E. Failure of fat cell prolifer-ation, mitochondrial function and fat oxidation results inectopic fat storage, insulin resistance and type II diabetesmellitus. Int J Obes Relat Metab Disord 2004;28(suppl):S12-21.
32. Garg A. Acquired and inherited lipodystrophies. N Engl JMed 2004;350:1220-34.



















![DiabeticCataract—Pathogenesis,Epidemiology andTreatment · 2019. 7. 31. · 1 diabetes mellitus [17, 18] due to the extensive swelling of cortical lens fibers [18]. A study performed](https://img.pdfslide.us/doc/110x75/6113d69688aa870125206466/diabeticcataractapathogenesisepidemiology-andtreatment-2019-7-31-1-diabetes.jpg)