Embed Size (px)
Citation preview

Current Biology, Vol. 14, 801–805, May 4, 2004, 2004 Elsevier Ltd. All rights reserved. DOI 10.1016/j .cub.2004.04.037
Involvement of Putative SNF2 Chromatin RemodelingProtein DRD1 in RNA-Directed DNA Methylation
quences can induce methylation of homologous pro-moter regions and transcriptional gene silencing [4, 5,10–14]. To study RNA-directed DNA methylation of a
Tatsuo Kanno,1 M. Florian Mette,2 David P. Kreil,3
Werner Aufsatz,1 Marjori Matzke,1,*and Antonius J.M. Matzke1
1Gregor Mendel Institute of Molecular Plant Biology tissue-specific promoter, we established a two-compo-nent transgene system based on the seed-specific ��Austrian Academy of Sciences
UZAII, Althanstrasse 14 promoter in Arabidopsis (Figure 1A; and SupplementalExperimental Procedures in the Supplemental DataA-1090 Vienna
Austria available with this article online). In this system, silencing(Figures 1B and 1C) and methylation (Figure 2A) of an2 Institute of Plant Genetics and Crop Plant
Research ��GFP reporter gene are triggered by an �� promoterhairpin RNA that is transcribed from an inverted DNACorrensstrasse 3
D-06466 Gatersleben repeat by an unrelated constitutive promoter. To identifyproteins of the silencing machinery, we mutagenizedGermany
3 Department of Genetics/Inference Group seeds of a homozygous, silenced ��GFP line (termedDT7-3) by using ethyl methane sulfonate (EMS). TheUniversity of Cambridge
Cambridge, CB2 3EH treated seeds were germinated, and the resulting M1plants were allowed to self fertilize to produce M2 seeds.United KingdomWe detected silencing-defective mutants by screeningM2 seeds for green fluorescence (Figure 1B). From poolsof M2 seeds harvested from approximately 50,000 M1Summaryplants, 59 putative mutants defective in RNA-directedDNA methylation (drd mutants) were recovered andIn plants, the mechanism by which RNA can induceplaced into three complementation groups. These mu-de novo cytosine methylation of homologous DNA istants were recessive, as indicated by resilencing thepoorly understood. Cytosines in all sequence contexts��GFP target gene in seeds obtained by backcrossingbecome modified in response to RNA signals [1, 2].to wild-type DT7-3 plants (Figure 1C). We report hereRecent work has implicated the de novo DNA methyl-on drd1.transferases (DMTases), DRM1 and DRM2, in estab-
Northern blot analysis demonstrated that the synthe-lishing RNA-directed methylation of the constitutivesis of �� promoter double-stranded RNA (not shown)nopaline synthase promoter [3], as well as the DMTaseand processing to �� promoter short RNAs occurredMET1 [4] and the putative histone deacetylase HDA6normally in the drd1 mutant (Figure 2B). Hence, release[5] in maintaining or enhancing CpG methylation in-of silencing was not due to impaired production of RNAduced by RNA. Despite the identification of enzymessignals. Methylation-sensitive restriction enzymes andthat catalyze epigenetic modifications in response tobisulfite sequencing were used for the examination ofRNA signals, it is unclear how RNA targets DNA forDNA methylation of the target �� promoter in the drd1methylation. A screen for mutants defective in RNA-mutant. Non-CpG methylation (analyzed by the restric-directed DNA methylation identified a novel putativetion enzymes abbreviated F, S, Ps, Pa, E, and B) waschromatin-remodeling protein, DRD1. This protein be-absent in leaf DNA isolated from the drd1 mutant, andlongs to a previously undefined, plant-specific sub-only trace amounts were detected in seed DNA. By con-family of SWI2/SNF2-like proteins most similar to thetrast, CpG methylation (analyzed by the enzyme abbrevi-RAD54/ATRX subfamily. In drd1 mutants, RNA-inducedated H) was largely unaffected in leaves and seeds ofnon-CpG methylation is almost eliminated at a targetdrd1 plants, as demonstrated by the distinctive doublepromoter, resulting in reactivation, whereas methyla-band that is also observed in wild-type DT7-3 plantstion of centromeric and rDNA repeats is unaffected.(Figure 2A). The results obtained from the Southern blotThus, unlike the SNF2-like proteins DDM1/Lsh1 [6, 7]analysis were confirmed by bisulfite sequencing, whichand ATRX [8, 9], which regulate methylation of repeti-revealed a dramatic decrease in CpNpG and CpNpNtive sequences, DRD1 is not a global regulator of cyto-methylation in the drd1 mutant accompanied by approx-sine methylation. DRD1 is the first SNF2-like proteinimately wild-type DT7-3 levels of CpG methylation (Fig-implicated in an RNA-guided, epigenetic modificationures 3 and S1). Thus, non-CpG methylation induced byof the genome.RNA in the �’ promoter silencing system requires DRD1.By contrast, neither CpG nor non-CpG methylation was
Results and Discussion detectably reduced in centromeric or rDNA repeats inthe drd1 mutant (Figure 4). Although it is not yet known
RNA-directed DNA methylation requires a double- whether RNA triggers cytosine methylation of these re-stranded RNA that is processed by a Dicer-like activity petitive sequences in plants, these results suggest thatinto short RNAs that are 21–26 nucleotides in length DRD1 acts locally, not globally, to regulate levels of non-[10–12]. Double-stranded RNAs containing promoter se- CpG methylation.
The drd1 mutation was mapped (Supplemental Data)and found to correspond to a putative chromatin remod-*Correspondence: [email protected]
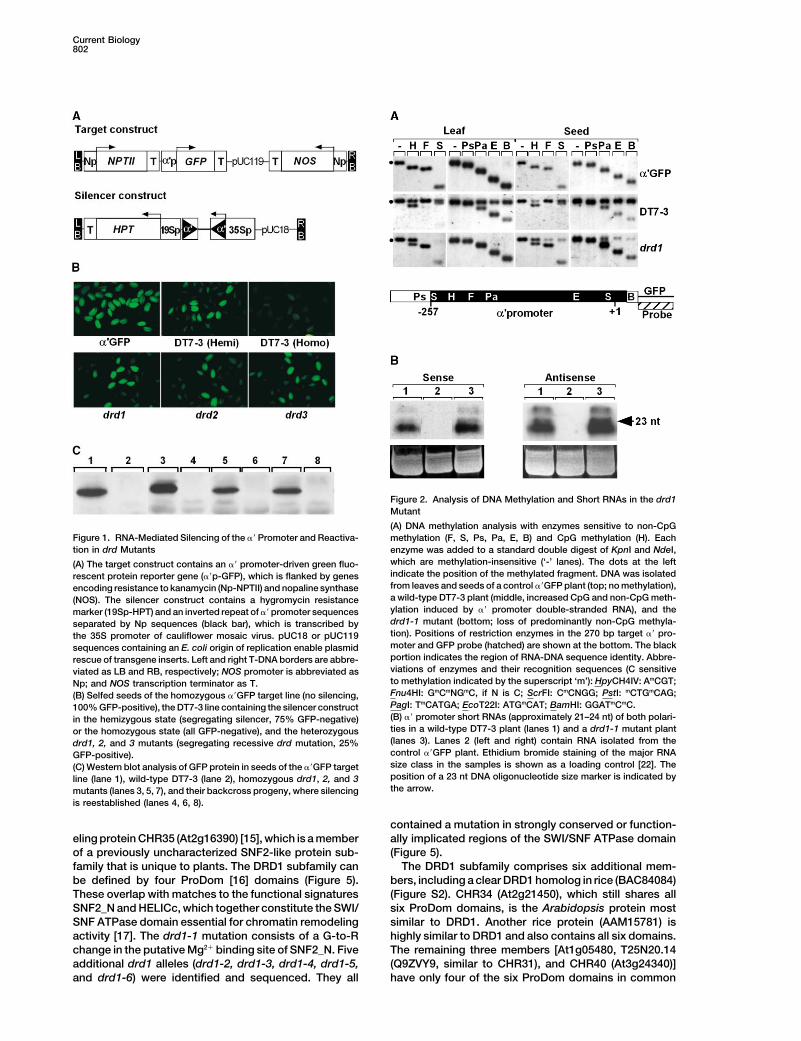
Current Biology802
Figure 2. Analysis of DNA Methylation and Short RNAs in the drd1Mutant
(A) DNA methylation analysis with enzymes sensitive to non-CpGFigure 1. RNA-Mediated Silencing of the �� Promoter and Reactiva- methylation (F, S, Ps, Pa, E, B) and CpG methylation (H). Eachtion in drd Mutants enzyme was added to a standard double digest of KpnI and NdeI,
which are methylation-insensitive (‘-’ lanes). The dots at the left(A) The target construct contains an �� promoter-driven green fluo-indicate the position of the methylated fragment. DNA was isolatedrescent protein reporter gene (��p-GFP), which is flanked by genesfrom leaves and seeds of a control ��GFP plant (top; no methylation),encoding resistance to kanamycin (Np-NPTII) and nopaline synthasea wild-type DT7-3 plant (middle, increased CpG and non-CpG meth-(NOS). The silencer construct contains a hygromycin resistanceylation induced by �� promoter double-stranded RNA), and themarker (19Sp-HPT) and an inverted repeat of �� promoter sequencesdrd1-1 mutant (bottom; loss of predominantly non-CpG methyla-separated by Np sequences (black bar), which is transcribed bytion). Positions of restriction enzymes in the 270 bp target �� pro-the 35S promoter of cauliflower mosaic virus. pUC18 or pUC119moter and GFP probe (hatched) are shown at the bottom. The blacksequences containing an E. coli origin of replication enable plasmidportion indicates the region of RNA-DNA sequence identity. Abbre-rescue of transgene inserts. Left and right T-DNA borders are abbre-viations of enzymes and their recognition sequences (C sensitiveviated as LB and RB, respectively; NOS promoter is abbreviated asto methylation indicated by the superscript ‘m’): HpyCH4IV: AmCGT;Np; and NOS transcription terminator as T.Fnu4HI: GmCmNGmC, if N is C; ScrFI: CmCNGG; PstI: mCTGmCAG;(B) Selfed seeds of the homozygous ��GFP target line (no silencing,PagI: TmCATGA; EcoT22I: ATGmCAT; BamHI: GGATmCmC.100% GFP-positive), the DT7-3 line containing the silencer construct(B) �� promoter short RNAs (approximately 21–24 nt) of both polari-in the hemizygous state (segregating silencer, 75% GFP-negative)ties in a wild-type DT7-3 plant (lanes 1) and a drd1-1 mutant plantor the homozygous state (all GFP-negative), and the heterozygous(lanes 3). Lanes 2 (left and right) contain RNA isolated from thedrd1, 2, and 3 mutants (segregating recessive drd mutation, 25%control ��GFP plant. Ethidium bromide staining of the major RNAGFP-positive).size class in the samples is shown as a loading control [22]. The(C) Western blot analysis of GFP protein in seeds of the ��GFP targetposition of a 23 nt DNA oligonucleotide size marker is indicated byline (lane 1), wild-type DT7-3 (lane 2), homozygous drd1, 2, and 3the arrow.mutants (lanes 3, 5, 7), and their backcross progeny, where silencing
is reestablished (lanes 4, 6, 8).
contained a mutation in strongly conserved or function-ally implicated regions of the SWI/SNF ATPase domaineling protein CHR35 (At2g16390) [15], which is a member
of a previously uncharacterized SNF2-like protein sub- (Figure 5).The DRD1 subfamily comprises six additional mem-family that is unique to plants. The DRD1 subfamily can
be defined by four ProDom [16] domains (Figure 5). bers, including a clear DRD1 homolog in rice (BAC84084)(Figure S2). CHR34 (At2g21450), which still shares allThese overlap with matches to the functional signatures
SNF2_N and HELICc, which together constitute the SWI/ six ProDom domains, is the Arabidopsis protein mostsimilar to DRD1. Another rice protein (AAM15781) isSNF ATPase domain essential for chromatin remodeling
activity [17]. The drd1-1 mutation consists of a G-to-R highly similar to DRD1 and also contains all six domains.The remaining three members [At1g05480, T25N20.14change in the putative Mg2� binding site of SNF2_N. Five
additional drd1 alleles (drd1-2, drd1-3, drd1-4, drd1-5, (Q9ZVY9, similar to CHR31), and CHR40 (At3g24340)]have only four of the six ProDom domains in commonand drd1-6) were identified and sequenced. They all
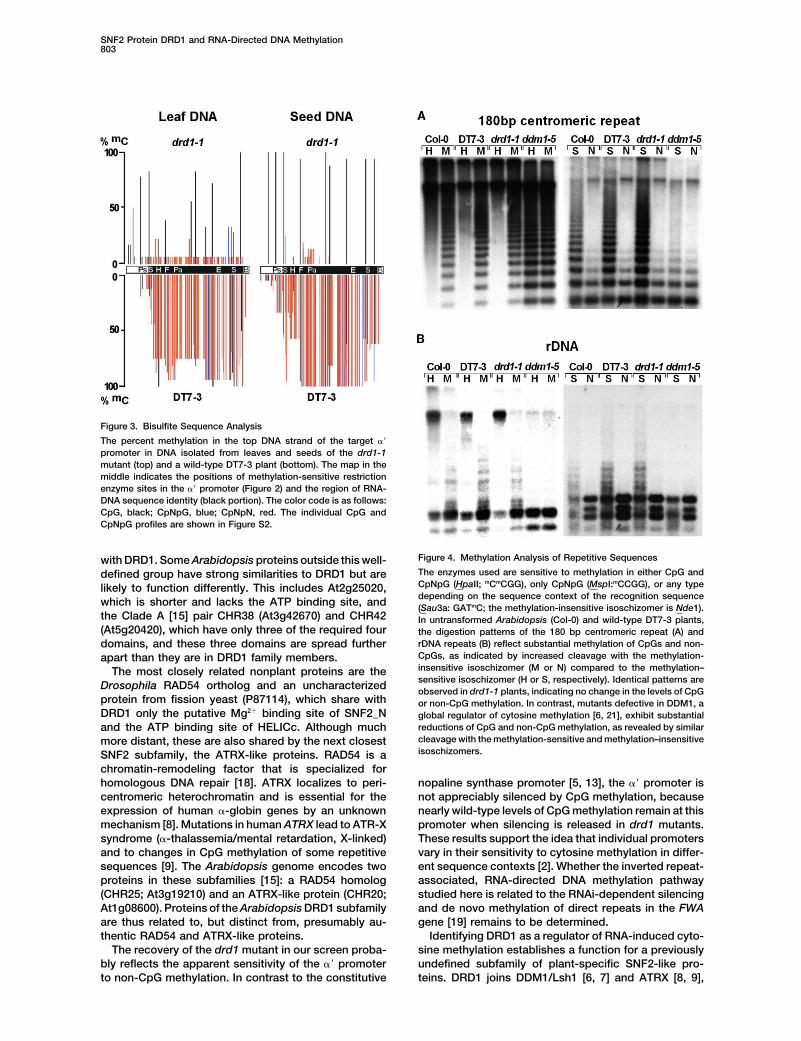
SNF2 Protein DRD1 and RNA-Directed DNA Methylation803
Figure 3. Bisulfite Sequence Analysis
The percent methylation in the top DNA strand of the target ��
promoter in DNA isolated from leaves and seeds of the drd1-1mutant (top) and a wild-type DT7-3 plant (bottom). The map in themiddle indicates the positions of methylation-sensitive restrictionenzyme sites in the �� promoter (Figure 2) and the region of RNA-DNA sequence identity (black portion). The color code is as follows:CpG, black; CpNpG, blue; CpNpN, red. The individual CpG andCpNpG profiles are shown in Figure S2.
Figure 4. Methylation Analysis of Repetitive Sequenceswith DRD1. Some Arabidopsis proteins outside this well-The enzymes used are sensitive to methylation in either CpG anddefined group have strong similarities to DRD1 but areCpNpG (HpaII; mCmCGG), only CpNpG (MspI:mCCGG), or any typelikely to function differently. This includes At2g25020,depending on the sequence context of the recognition sequencewhich is shorter and lacks the ATP binding site, and (Sau3a: GATmC; the methylation-insensitive isoschizomer is Nde1).
the Clade A [15] pair CHR38 (At3g42670) and CHR42 In untransformed Arabidopsis (Col-0) and wild-type DT7-3 plants,(At5g20420), which have only three of the required four the digestion patterns of the 180 bp centromeric repeat (A) and
rDNA repeats (B) reflect substantial methylation of CpGs and non-domains, and these three domains are spread furtherCpGs, as indicated by increased cleavage with the methylation-apart than they are in DRD1 family members.insensitive isoschizomer (M or N) compared to the methylation–The most closely related nonplant proteins are thesensitive isoschizomer (H or S, respectively). Identical patterns areDrosophila RAD54 ortholog and an uncharacterized observed in drd1-1 plants, indicating no change in the levels of CpG
protein from fission yeast (P87114), which share with or non-CpG methylation. In contrast, mutants defective in DDM1, aDRD1 only the putative Mg2� binding site of SNF2_N global regulator of cytosine methylation [6, 21], exhibit substantial
reductions of CpG and non-CpG methylation, as revealed by similarand the ATP binding site of HELICc. Although muchcleavage with the methylation-sensitive and methylation–insensitivemore distant, these are also shared by the next closestisoschizomers.SNF2 subfamily, the ATRX-like proteins. RAD54 is a
chromatin-remodeling factor that is specialized forhomologous DNA repair [18]. ATRX localizes to peri- nopaline synthase promoter [5, 13], the �� promoter is
not appreciably silenced by CpG methylation, becausecentromeric heterochromatin and is essential for theexpression of human �-globin genes by an unknown nearly wild-type levels of CpG methylation remain at this
promoter when silencing is released in drd1 mutants.mechanism [8]. Mutations in human ATRX lead to ATR-Xsyndrome (�-thalassemia/mental retardation, X-linked) These results support the idea that individual promoters
vary in their sensitivity to cytosine methylation in differ-and to changes in CpG methylation of some repetitivesequences [9]. The Arabidopsis genome encodes two ent sequence contexts [2]. Whether the inverted repeat-
associated, RNA-directed DNA methylation pathwayproteins in these subfamilies [15]: a RAD54 homolog(CHR25; At3g19210) and an ATRX-like protein (CHR20; studied here is related to the RNAi-dependent silencing
and de novo methylation of direct repeats in the FWAAt1g08600). Proteins of the Arabidopsis DRD1 subfamilyare thus related to, but distinct from, presumably au- gene [19] remains to be determined.
Identifying DRD1 as a regulator of RNA-induced cyto-thentic RAD54 and ATRX-like proteins.The recovery of the drd1 mutant in our screen proba- sine methylation establishes a function for a previously
undefined subfamily of plant-specific SNF2-like pro-bly reflects the apparent sensitivity of the �� promoterto non-CpG methylation. In contrast to the constitutive teins. DRD1 joins DDM1/Lsh1 [6, 7] and ATRX [8, 9],

Current Biology804
Figure 5. Domain Structure of DRD1 and Po-sitions of drd1 Mutations and Known Func-tional Signatures
The four ProDom domains defining the DRD1family are shown in bold (I, III, IV, VI). Thesix domains present in DRD1 and its closesthomologs (I-VI) are PD423058, PD217428,PD000441, PD210292, PD690098, andPD039514. A putative binding site for Mg2�
is at 492–495 (mutated in drd1-1) and for ATP,it is at Q803, R807 (mutated in drd1-2), andR810 (mutated in drd1–5). NLS, predicted bi-partite nuclear localization signal. The ‘#’ signindicates a premature stop codon.
Anti-rabbit Ig, HRP-Linked whole AB (Amersham, Vienna, Austria)which maintain CpG methylation, on the list of SNF2-was used for the second antibody. The result was visualized onlike proteins that are important for DNA methylation. TheKodak X-OMAT film (Kodak GmbH, Vienna, Austria). The film wasapparent preference of DRD1 for non-CpG methylationscanned and the signals were quantified with ImageMaster 1D
and the lack of close homologs in animals distinguish (Amersham, Vienna, Austria).it from these other proteins. Whether DRD1 is involvedin RNA-directed de novo methylation or acts to maintain RNA AnalysisRNA-induced non-CpG methylation remains to be deter- �� promoter double-stranded (ds) RNA and short RNA were isolated
from fresh rosette leaves of plants just starting to flower and ana-mined. However, the heavy loss in drd1 plants of CpNpNlyzed on Northern blots as previously described [5, 10, 13], withmethylation, which is not efficiently maintained in theslight modifications. A 32P-labeled �� promoter DNA probe was usedabsence of the RNA trigger [10, 13], suggests a directfor dsRNA detection, and 32P-labeled sense or antisense RNA
relationship between DRD1 activity and RNA signals. probes were used for small RNA detection on Northern blots, re-Given the relatedness of DRD1 to RAD54, it is intriguing spectively.to consider possible mechanistic similarities betweenRNA-directed DNA methylation and homologous DNA DNA Analysis
Total Arabidopsis DNA for methylation analysis was isolated fromrepair. In each case, the respective chromatin-remodel-dry mature seeds and fresh rosette leaves of BC1F3 drd1-1 plants,ing factor could facilitate a homology search on duplexfourth-generation wild-type DT7-3 plants, and fourth-generationDNA, nucleosome displacement, and DNA unpairing��GFP plants with a DNeasy Plant Maxi Kit (Qiagen, Hilden, Ger-
and thereby allow heteroduplex formation and recruit- many) according to the manufacturers instructions. Target �� pro-ment of enzyme complexes. In the RNA-directed DNA moter methylation was analyzed with methylation-sensitive restric-methylation pathway, this could create an RNA-DNA tion enzymes and Southern blot hybridization, according to
previously published protocols [5, 10, 13]. A 32P-labeled sense RNAhybrid that attracts DMTases, thus accounting for thecorresponding to the first 220 bp of the GFP protein coding regionextraordinary specificity of cytosine methylation, whichwas used for probing the Southern blots. Methylation in the centro-is largely restricted to the region of RNA-DNA sequencemeric and rDNA repeats was analyzed as previously described [21].
similarity [1, 5, 13]. The lack of DRD1 homologs outside For bisulfite sequencing, DNA preparations were treated with so-of the plant kingdom may mean that RNA-directed DNA dium bisulfite, and subsequent steps were carried out according tomethylation occurs only in plants. Alternatively, RAD54 standard protocols [5, 12]. Degenerate primers for amplification after
bisulfite treatment were: 5�-AYGYGATAGAAAAYAAAATATAG-3�or ATRX-like proteins may serve this function in otherand 5�-CTTTACTCATTRTTATATCTCC-3� for the �� promoter toporganisms.strand. The number of PCR amplification products sequenced wasas follows: drd1 leaf, 18; drd1 seed, 16; DT7-3 leaf, 16; and DT7-3Experimental Proceduresseeds, 21. Methylation appears to be slightly higher with bisulfitesequencing, which samples a subset of PCR amplification products,Plant Transformationthan with methylation-sensitive restriction enzymes, which assessThe floral dip method [20] was used for transforming Arabidopsismethylation in the entire DNA sample. The general changes in meth-thaliana plants (ecotype Columbia) with Agrobacterium tumefaciensylation detected with both approaches are the same. Bisulfite se-harboring either the target or silencer constructs (Supplemental Ex-quencing detected negligible methylation in DNA isolated fromperimental Procedures). Transformants were selected on Murashigeseeds and leaves of the ��GFP target line (data not shown).and Skoog medium (Invitrogen GmbH, Lofer, Austria) containing
either 50 �g/ml kanamycin (target construct) or 20 �g/ml hygromycinSupplemental Data(silencer construct). The plant lines used in this study were theSupplemental Data including additional Experimental Procedures,homozygous ��GFP target line and the corresponding double trans-two figures, and references are available with this article online atformant line (DT7-3), which contains both the target and the silencerhttp://www.current-biology.com/cgi/content/full/14/9/801/DC1/.constructs.
GFP Visualization and Western Blot Analysis AcknowledgmentsGFP fluorescence of mature seeds was visualized under the fluores-cence microscope MZ FLIII (Leica, Wetzlar, Germany) with a GFP2 We thank Johannes van der Winden for DNA sequencing, Adriana
Danielopol for assistance with plant work, and Ortrun Mittelstenfilter. The image was captured with a Color CoolView Camera (Pho-tonic Science, Kent, UK) and optimized with Adobe Photoshop. Scheid for som8/ddm1-5 seeds. We are grateful to Cereon Geno-
mics for marker information. This work was supported by grantsGFP protein was detected on Western blots with the ECL AdvanceWestern Blotting Detection kit (Amersham, Vienna, Austria) as rec- to M.M. and A.J.M.M. from the Austrian Fonds zur Forderung der
wissenschaftlichen Forschung (grant numbers Z21-MED andommended by the manufacturer. Living Colors A.v. antibody (Clon-tech, BD Biosciences, Belgium) was used for the first antibody, and P-15611-B07) and the European Union (Contract QLK3-2000-

SNF2 Protein DRD1 and RNA-Directed DNA Methylation805
00078). D.P.K. acknowledges support of a research fellowship from the Rad51-ssDNA nucleoprotein filament. Nat. Struct. Biol. 10,182–186.the Medical Research Council, United Kingdom (grant number G81/
555). 19. Chan, W.-L., Zilberman, D., Xie, Z., Johansen, L., Carrington,J., and Jacobsen, S. (2004). RNA silencing genes control denovo DNA methylation. Science 303, 1336.
Received: February 26, 2004 20. Clough, S., and Bent, A. (1998). Floral dip: a simplified methodRevised: March 15, 2004 for Agrobacterium-mediated transformation of ArabidopsisAccepted: March 15, 2004 thaliana. Plant J. 16, 735–743.Published: May 4, 2004 21. Vongs, A., Kakutani, T., Martienssen, R., and Richards, E.J.
(1993). Arabidopsis thaliana DNA methylation mutants. Science260, 1926–1928.References
22. Papp, I., Mette, M.F., Aufsatz, W., Daxinger, L., Schauer, S.,Ray, A., van der Winden, J., Matzke, M., and Matzke, A.J.M.1. Wassenegger, M. (2000). RNA-directed DNA methylation. Plant(2003). Evidence for nuclear processing of plant microRNA andMol. Biol. 43, 203–220.short interfering RNA precursors. Plant Physiol. 132, 1382–1390.2. Matzke, M., Aufsatz, W., Kanno, T., Daxinger, L., Papp, I., Mette,
M.F., and Matzke, A.J.M. (2004). Genetic analysis of RNA-medi-ated transcriptional gene silencing. Biochim. Biophys. Acta1677, 129–141.
3. Cao, X., Aufsatz, W., Zilberman, D., Mette, M.F., Huang, M.,Matzke, M., and Jacobsen, S.E. (2003). Role of DRM and CMT3methyltransferases in RNA-directed DNA methylation. Curr.Biol. 13, 2212–2217.
4. Jones, L., Ratcliff, F., and Baulcombe, D. (2002). RNA-directedtranscription gene silencing in plants can be inherited indepen-dently of the RNA trigger and requires MET1 for maintenance.Curr. Biol. 11, 747–757.
5. Aufsatz, W., Mette, M.F., van der Winden, J., Matzke, M., andMatzke, A.J.M. (2002). HDA6, a putative histone deacetylaseneeded to enhance DNA methylation induced by RNA. EMBOJ. 21, 6832–6841.
6. Jedelloh, J., Stokes, T., and Richards, E. (1999). Maintenanceof genomic methylation requires a SWI2/SNF2-like protein. Nat.Genet. 22, 94–97.
7. Dennis, K., Fan, T., Geiman, T., Yan, Q., and Muegge, K. (2001).Lsh, a member of the SNF2 family, is required for genome-widemethylation. Genes Dev. 15, 2940–2944.
8. Gibbons R.J., Pellagatti A., Garrick D., Wood W.G., Malik N.,Ayyub H., Langford C., Boultwood J., Wainscoat J.S., HiggsD.R. (2003). Identification of acquired somatic mutations in thegene encoding chromatin-remodeling factor ATRX in the�-thalassemia myelodysplasia syndrome (ATMDS). Nat. Genet.34, 446–449.
9. Gibbons, R.J., McDowell, T.L., Raman, S., O’Rourke, D.M., Gar-rick, D., Ayyub, H., and Higgs, D.R. (2000). Mutations in ATRX,encoding a SWI/SNF-like protein, cause diverse changes in thepattern of DNA methylation. Nat. Genet. 24, 368–371.
10. Mette, M.F., Aufsatz, W., van der Winden, J., Matzke, M., andMatzke, A.J.M. (2000). Transcriptional silencing and promotermethylation triggered by double-stranded RNA. EMBO J. 19,5194–5201.
11. Sijen, T., Vijn, I., Rebocho, A., van Blokland, R., Roelofs, D., Mol,J., and Kooter, J. (2001). Transcriptional and posttranscriptionalgene silencing are mechanistically related. Curr. Biol. 11,436–440.
12. Hamilton, A., Voinnet, O., Chappell, L., and Baulcombe, D.(2002). Two classes of short interfering RNA in RNA silencing.EMBO J. 21, 4671–4679.
13. Aufsatz, W., Mette, M.F., van der Winden, J., Matzke, A.J.M.,and Matzke, M. (2002). RNA-directed DNA methylation in Arabi-dopsis. Proc. Natl. Acad. Sci. USA 99, 16499–16508.
14. Melquist, S., and Bender, J. (2003). Transcription from an up-stream promoter controls methylation signaling from an in-verted repeat of endogenous genes in Arabidopsis. Genes Dev.17, 2036–2047.
15. Plant Chromatin Database. (http://www.chromdb.org); see theproposed Clade A.
16. Servant, F., Bru, C., Carrere, S., Courcelle, E., Gouzy, J., Peyruc,D., and Kahn, D. (2002). ProDom: automated clustering of ho-mologous domains. Brief. Bioinform. 3, 246–251.
17. Lusser, A., and Kadonaga, J. (2003). Chromatin remodeling byATP-dependent molecular machines. Bioessays 25, 1192–1200.
18. Alexeev, A., Mazin, A., and Kowalczykowski, S.C. (2003). Rad54protein possesses chromatin-remodeling activity stimulated by





























