Embed Size (px)
Citation preview

INVESTIGATION OF Puccinia graminis RESISTANCE GENES IN
BARLEY
By
ROBERT SAXON BRUEGGEMAN
A dissertation submitted in partial fulfillment of
The requirements for the degree of
DOCTOR OF PHILOSOPHY
WASHINGTON STATE UNIVERSITY
Department of Crop and Soil Sciences
May 2009

To the Faculty of Washington State University
The members of the Committee appointed to examine the dissertation of ROBERT S
BRUEGGEMAN find it satisfactory and recommend that it be accepted
______________________________
Andris Kleinhofs
_______________________________
Stephen Jones
_______________________________
Diter von Wettstein
_______________________________
Scot Hulbert
ii

ACKNOWLEDGMENTS
I would like to thank Dr. Andris Kleinhofs for his guidance and teaching for the past eleven
years. Dr. Kleinhofs has been an exceptional advisor guiding me in all aspects of my scientific
development while allowing me the freedom to develop and explore new ideas and methods.
Over the years I have come to know him as a leader in science and a fishing friend who taught
me how to tie knots. I would also like to thank my committee members: Dr. Stephen Jones, Dr.
Diter von Wettstein and Dr. Scot Hulbert. Thank you for your excellent advise, instruction and
challenging questions. A special thanks goes to Dr. Brian Steffenson. This work was not possible
without his expertise and the rust reaction assays he has performed for us over the years. I would
also like to thank Dr. Arnis Druka for the years of instruction in the lab.
I would also like to thank all the present and past members of the WSU barley genomics
group. The diversity of friends I have now extends across the globe. I learned from each, either
through their instruction or my teaching.
iii

INVESTIGATION OF THE Puccinia graminis RESISTANCE GENES IN BARLEY
Abstract
by Robert Saxon Brueggeman, Ph. D. Washington State University
May 2009
Andris Kleinhofs
The barley stem rust resistance gene Rpg1 was cloned and characterized by a map-based
approach. It was shown to have a novel R-gene structure containing tandem kinase domains. To
determine the origin of Rpg1 we analyzed alleles from several barley land races (Hordeum
vulgare) and wild barley (H. vulgare subsp. spontaneum) accessions. Our allele analysis
determined that the functional Rpg1 gene is rare among the landraces and was absent from the
wild accessions examined. During the 1990s two new races of Puccinia graminis f. sp. tritici
(Pgt) designated QCCJ and TTKSK, virulent on Rpg1, emerged in North America and Eastern
Africa, respectively. Analysis of barley accessions for QCCJ and TTKSK reaction identified the
barley line Q21861 as the best resistance source and was designated the rpg4 gene. Q21861 also
contains resistance against Puccinia graminis f. sp. secalis (Pgs) designated the Rpg5 gene. The
two resistance genes initially co-segragated to a region on barley chromosome 5H(7). In an
attempt to quickly identify an rpg4/Rpg5 candidate gene, five Rpg1 gene family members were
identified and mapped. One was tightly linked to the rpg4/Rpg5 locus but segregated away in a
high-resolution mapping population. Recombinant analysis of the high-resolution population
revealed that rpg4 and Rpg5 were distinct genes. We cloned and partially characterized a 70 kbp
genomic region from barley containing the rpg4 and Rpg5 genes. An Rpg5 candidate gene was
iv

identified and confirmed by allele sequencing. The predicted RPG5 protein from the resistant
line Q21861 contains a Nucleotide-Binding-Site, Leucine-Rich-Repeat region and a protein
kinase domain. A reverse genetics approach, Virus Induced Gene Silencing was used to confirm
the Rpg5 gene. A gene encoding an actin depolymerizing factor-like protein (ADF2) was
identified as rpg4. The Adf2 gene (rpg4), in combination with Rpg5, appears to be essential for
resistance against the Pgt races, but not for the Pgs isolate. One possible hypothesis for the Adf2
gene function is that it might be activated or inactivated by RPG5 upon pathogen recognition.
Thus making the ADF2 protein inaccessible for the fungus to redirect actin networks to establish
a haustoria-plant interface to obtain nutrients from the plant.
v

TABLE OF CONTENTS
Page
ACKNOWLEDGEMENTS........................................................................................................ iii
ABSTRACT................................................................................................................................ iv
PREFACE................................................................................................................................. viii
DEDICATION..............................................................................................................................x
PROLOGUE .................................................................................................................................1
A. DEVELOPMENT OF STEM RUST RESEARCH ................................................................1
1. Puccinia graminis .......................................................................................................2
2. Stem Rust in North America.......................................................................................4
3. Identification of Barley Stem Rust Resistance Genes ................................................6
4. Resistance Genes ......................................................................................................11
5. Rust Resistance Genes; Cloning and Characterization.............................................13
B. Rpg1 CLONING AND CHARACTERIZATION ................................................................17
1. Rpg1 Map-Based Cloning.........................................................................................18
2. Gene Family Characterization ..................................................................................19
3. Rpg1 Allele Characterization....................................................................................22
4. Rpg1 Functional Analysis .........................................................................................24
vi

C. MAP BASED CLONING OF rpg4 AND Rpg5 ...................................................................26
1. Cloning and Characterization of Rpg5........................................................................27
2. Interaction of rpg4 and Rpg5 for P.g. f. sp. tritici Resistance ....................................29
REFERENCES ...........................................................................................................................34
CHAPTERS
1. The barley stem rust-resistance gene Rpg1 is a novel disease-resistance gene with
homology to receptor kinases. ....................................................................................................44
2. The barley serine/threonine kinase gene Rpg1 providing resistance to stem rust
belongs to a gene family with five other members encoding kinase domains............................51
3. Allele sequencing of the barley stem rust resistance gene Rpg1 identifies regions
relevant to disease resistance. .....................................................................................................64
4. The stem rust resistance gene Rpg5 encodes a protein with nucleotide-binding-site,
leucine-rich, and protein kinase domains....................................................................................74
5. The rpg4/ Rpg5 stem rust resistance locus in barley; resistance genes and
cytoskeleton dynamics. ...............................................................................................................81
SUMMARY AND FUTURE DIRECTION ...............................................................................87
vii

Preface
This thesis is based on the following published manuscripts, which are referred to by the
respective chapters. Research from chapter 1 was carried out while I was an undergraduate.
Chapter 2 is reflective of my masters thesis work. Chapters 3-5 are reflective of my doctoral
work.
Chapter 1 (R. Brueggeman is the main contributor)
Brueggeman R., Rostoks N., Kudrna D., Kilian A., Han F., Chen J., Druka A., Steffenson B. and
Kleinhofs A. (2002) The barley stem rust-resistance gene Rpg1 is a novel disease-resistance gene
with homology to receptor kinases. Proc. Natl. Acad. Sci. USA 99(14):9328-9333.
Chapter 2 (R. Brueggeman is the main contributor)
Brueggeman R., Drader, T., and Kleinhofs A. (2006) The barley serine/threonine kinase gene
Rpg1 providing resistance to stem rust belongs to a gene family with five other members
encoding kinase domains. Theor. Appl. Genet. 113:1147-1158.
Chapter 3 (A. Mirlohi and R. Brueggeman contributed equally to this work)
Mirlohi A., Brueggeman R., Drader T., Nirmala J., Steffenson B.J. and Kleinhofs A. (2008)
Allele sequencing of the barley stem rust resistance gene Rpg1 identifies regions relevant to
disease resistance. Phytopathology 98:910-918.
viii

Chapter 4 (R. Brueggeman is the main contributor)
Brueggeman R., Druka A., Nirmala J., Cavileer T., Drader T., Rostoks N., Mirlohi A.,
Bennypaul H., Gill U., Kudrna D., Whitelaw C., Killian A., Han F., Sun Y., Gill K., Steffenson
B. and Kleinhofs A. (2008) The stem rust resistance gene Rpg5 encodes a protein with
nucleotide-binding-site, leucine-rich, and protein kinase domains. Proc Natl Acad Sci USA
105(39):14970-14975.
Chapter 5 (R. Brueggeman is the main contributor)
Brueggeman R., Steffenson B.J. and Kleinhofs A. (2009) The rpg4/Rpg5 stem rust resistance
locus in barley; Resistance genes and cytoskeleton dynamics. Cell Cycle, 8(7);977-981.
ix

x
DEDICATION
I dedicate this thesis to my loving wife Leah
Thank you!
and our three beautiful boys
Max, Saul and Gus

PROLOGUE
A. DEVELOPMENT OF STEM RUST RESEARCH
Rust fungi infect and damage most crop plants but those infecting the small grain crops
wheat, barley, rye and oats have caused the largest economic losses and pose the biggest threat to
world food security. One of the all time most devastating diseases of the small grains is stem rust
also known as black rust caused by the biotrophic fungal pathogen Puccinia graminis.
Historically and prehistorically dating as far back as the beginning of cultivating grain crops in
the Fertile Crescent region of the Middle East, stem rust epidemics have probably been one of
the major factors contributing to crop failure and famine. It is believed that stem rust has been a
problem since the beginning of cultivating grain crops in the Fertile Crescent region. The earliest
record of plant diseases is found in the Bible. It is written that while still in the desert of Sinai,
some thirty-five centuries ago, Moses had warned the people of Israel that if they failed to
observe the commandments of Jehovah, the Lord would punish them by destroying their field
crops with smut and with rust (Deut, 28:22) indicating that rust was a major concern at the time.
Now it is assumed that, in part, it was due to stem rust (Chester, 1946). Puccinia graminis spores
have been identified in archeological sites in Israel containing grain bins that date as far back as
1300 BC (Kislev, 1982), approximately the same period as this first historic record of rust on
wheat.
Wheat stem rust was also a serious disease in ancient Greece and during the Roman
Empire. It was observed and recognized as early as the times of Aristotle (384-322 B.C.).
Aristotle documented that the rust was brought on by the warm vapors indicating that ancient
scientists made the correlation between a warm moist environment and severe disease onset
(Chester, 1946). The ancient Romans also sacrificed red animals to the rust god Robigo in the
1

spring hoping that they could appease the god and he would spare their wheat crops of the
devastating red rust (Chester, 1946). Historical weather records, indicating a string of unusually
moist years, suggest that rust epidemics may have caused a food crisis contributing to the fall of
the Roman Empire.
1. Puccinia graminis
The Italian Scientist Felice Fontana was the first to describe wheat stem rust in detail in
the 1760s. Then in 1797 Christian Hendrik Persoon named it Puccinia graminis. By the 1850s
the Tulasne brothers, Charles and Louis, had discovered that a number of autoacious (single
host) rust fungi produced up to five different spore types or stages during their lifecycle. They
also observed and described that the red (urediniospores) and black (teliospores) were different
spore stages of Puccinia graminis, but the reason basidiospores of Puccinia graminis could not
infect its grass host remained a mystery until Anton deBarry’s work in 1865. Anton deBarry was
interested in the observation that the basidiospores of Puccinia graminis were unable to infect
wheat plants. He was aware of the farmer’s beliefs that barberry increased the onset of wheat rust
early in the season and greatly affected the severity of the disease. Using this knowledge he
successfully inoculated barberry with Puccinia graminis basidiospores and observed the
remaining spore stages, which occur on the alternate host (Chester, 1946). This heteroecious
nature of stem rust was a very important discovery in phytopathology and once the heteroecious
nature of rusts was described in stem rust it was established in many other known rust fungi and
their secondary hosts were identified.
The function of the spore stages were not completely understood until John Craige’s
studies in 1927 (Chester, 1946). Puccinia graminis was determined to be a heteroecious,
macrocyclic rust with a life cycle involving five spore stages and two hosts. The two most
2

important spore stages concerning disease management are the asexual urediniospore (dikaryotic
n+n) and the sexual aeciospore (n+n). The uredinium is produced on the grass host after
infection resulting from inoculation by either a urediniospore or aeciospore. The urediniospore is
very stable in the environment and can be wind disseminated over long distances. Urediniospores
then act as secondary inoculum reinfecting the grass host species cycling as secondary inoculum
during the summer on the primary hosts including barley and wheat. When environmental
conditions are favorable, the generation time from spore to secondary inoculum is approximately
10-15 days. The rapid production of inoculum, coupled with the long distances the spores can be
disseminated, makes stem rust a very explosive and destructive disease where sufficient
inoculum, the right environmental conditions and a susceptible host is grown over large tracts of
land. These three components make up the disease triangle in epidemiological terms and dictate
the severity of an epidemic.
Late in the growing season, the stem rust fungus begins producing teliospores on the
stems of maturing grass host. Teliospores lay dormant in the straw during the winter providing
the only means for long term survival of the obligate biotroph in the absence of a living host. In
the spring, the teliospores germinate and produce four haploid basidiospores, the result of
karyogamy and meiosis. Basidiospores can not infect the primary grass hosts but rather infect the
alternate host Berberis (barberry) or, Mahonia spp. The infection on the alternate host results in
the development of pycnia, occurring with the fusion between a pycniospore and receptive
hyphae of opposite mating types. Plasmogamy results in a dikaryotic hyphae which produce
aecium with dikaryotic aeciospores. Although new races can emerge by mutation events in the
absence of barberry, the secondary host functions to produce greater diversity of new races
because the pycnial stage preceding aecial development allows for genetic recombination.
3

Aeciospores are then wind disseminated back to the grass host, thus completing the sexual stages
of the life cycle. In order to stop the early season inoculum comming from the secondary host,
barberry iradication programs began as early as the 1600s in France and the 18th century in the
United States.
2. Stem Rust in North America
Stem rust caused devastating epidemics to wheat and barley production in the
Midwestern United States and Prairie Provinces of Canada until the mid twentieth century.
Efforts to develop rust resistance in wheat varieties were initiated in Kansas in 1911 soon
followed by numerous other states, beginning the so called “Boom and Bust” cycles of wheat
breeding and production in the Midwest. R-genes deployed in the new wheat varieties would
protect the crop (the Boom) until races virulent on these R-genes became the predominant races
causing large-scale stem rust epidemics again (the Bust). Stem rust epidemics prior to the 1950s
during a single growing season resulted in wheat yield losses as high as 200 million bushels in
the US alone. Then in the 1950s wheat breeders began pyramiding several resistance genes into
their varieties. This practice provided more durable resistance ending the occurrence of the major
stem rust epidemics (Peterson, 2001). The losses on barley are not well documented but the
epidemics were similar to that found on wheat (Steffenson, 1991). Stem rust was also managed
using genetic resistance in barley, but only a single source of resistance was deployed and has
protected midwestern barley cultivars since 1942.
The small grain hosts of stem rust, barley, oats, rye and wheat are all non-native crops
brought to North America by the European colonists. Barberry was also brought to North
America for its practical uses including production of yellow dye and edible berries used in jams
4

and wine. The fast growing thorny hedges were also used to coral domestic animals. Farmers
made the connection between the alternate host barberry and the early and more severe onset of
rust within their wheat and barley fields much earlier than the scientific community. Farmers
urged the passage of laws as early as the 1660,s requiring the destruction and banning of
barberry in wheat growing regions of Rouen, France (Chester, 1946).
In the US barberry bans were enacted as early as 1726 in Connecticut and soon after in
other states (Chester, 1946). Then after the severe stem rust epidemic of 1916 the US
implemented the barberry eradication program in 1918. This impacted the control efforts of rust
in two major ways. First it reduced the amount of early season inoculum coming in from the
barberry plants found in fence rows effectively delaying disease onset by 10 days. Second, it
removed the sexual stage of the rusts lifecycle thus limiting its genetic diversity making
recombinant forms and new races less likely giving breeders more time because R-genes
deployed were effective for longer periods (Roelfs, 1982).
After over half a century of managing stem rust through genetic resistance it was believed
by many that stem rust was a thing of the past. Norman Borlaug’s statement sums this up quite
well “ Isn’t it quite possible that the greatest ally of the pathogen is our short memory of the
disastrous stem rust epidemics from 1951-1954 across much of the wheat producing regions of
North America” (Peterson, 2001). The misconception that stem rust has been defeated has
fostered complacency and currently restricts research on this shifty enemy. Stem rust research
has received less and less funding by the powers that be, leading to the dependency on too few
resistant sources in the US and the major wheat and barley producing regions worldwide. The
recent emergence of a new highly virulent race of stem rust in Eastern Africa and its quick
5

migration through Middle Eastern Asia has sent plant scientist scrambling to identify and deploy
new sources of resistance in both wheat and barley.
3. Identification of Barley Stem Rust Resistance Genes
Barley is an important crop in the Upper Midwestern region of the US and Prairie
Provinces of Canada with over 15 million metric tons produced annually. However, barley has a
much lower rank than wheat as a food crop for human consumption thus justifiably gets
substantially less attention than wheat in the field of stem rust research. Stem rust was
historically as much a problem on Midwestern barley as wheat, but has not caused a widespread
epidemic on barley for over 70 years due to the fortuitous deployment of a single durable
resistance gene, Rpg1 (Roelfs, 1978). Rpg1 was the first stem rust resistance gene identified and
was first described from an unimproved line imported by the USDA from the Canton of Lucerne
in Switzerland in 1914. It was shown to be a single dominant gene originally designated the T
gene (Powers and Hines, 1933), later changed to Rpg1 for Reaction to Puccinia graminis 1
(Sogaard and von Wettstein-Knowles, 1987). However, it was a different source of resistance
that gave rise to the first released barley cultivar containing the Rpg1 gene. This source of
resistance originated from a farmer named Sam Lykken from Kindred, North Dakota. During the
severe stem rust epidemic year of 1935 Sam was surveying his severely rusted barley field
planted with the susceptible variety Wisconsin 37, when among the entirely rusted and shriveled
field he observed a single green healthy plant. He harvested seed from this plant and increased
the seed for several generations. By 1941 Sam had enough seed for his own planting and 1,200
bushels to sell. This single plant selection was released as the cultivar (cv.) Kindred in 1942 and
remained the dominant cultivar in the Red River valley for over a decade (Steffenson, 1991).
6

Kindred together with the two barley selections, Chevron and Peatland, made from the
unimproved line obtained from the 1914 USDA imported germplasm from Switzerland were the
only sources of stem rust resistance used in barley breeding in the Midwestern USA and Canada.
With the cloning of the barley Rpg1 gene, it was shown that all three cultivars, Kindred, Chevron
and Peatland, posses an identical Rpg1 allele (Brueggeman et al., 2002 [chapter 1]). Wisconsin
37, the cultivar planted in the field from which Sam Lykken recovered the progenitor of Kindred,
contains an allele that could not have given rise to the resistant allele by a spontaneous mutation
event. This molecular evidence confirmed that the source of Kindred barley probably came from
a seed contamination as proposed earlier by Steffenson (Steffenson, 1992).
Rpg1 resistance is classified as durable due to its successful control of stem rust since the
release of Kindred in 1942. This remarkable durability held up in the 40s and 50s even through
epidemics that devastated the wheat crop (Steffenson, 1992). Considering the potential
variability of the fungus and the vast hectares (~ 1.8 million) over which cultivars with this
resistance gene are grown (Steffenson, 1992), Rpg1 fulfills the requirements to be defined as
durable (Johnson, 1984). In the field Rpg1 has conferred resistance to most pathotypes of
Puccinia graminis f. sp. tritici (Pgt), but there are exceptions. The genes conferring resistance to
these exceptional stem rust races virulent on Rpg1 was the focus of my doctoral research.
Aside from Rpg1 there are seven other identified sources of resistance against P.
graminis from barley, but none of these have been deployed in commercial cultivars. A complete
list of Rpg genes includes Rpg1, Rpg2, Rpg3, rpg4, Rpg5, rpg6, rpgBH, and RpgU . Only Rpg1,
rpg4 and Rpg5 have been cloned (Brueggeman et al., 2002 [chapter 1]; Brueggeman et al., 2008
[chapter 4]) and investigated at the molecular level. Rpg2, identified from Hietpas-5 (CIho 7124)
(Patterson et al., 1957), Rpg3, from PI382313 (Jedel, 1990), rpgBH from Black Hulless (CIho
7

666) (Steffenson et al., 1984) and RpgU from Peatland (Fox and Harder, 1995) all confer low
levels of stem rust resistance making these genes difficult to phenotype therefore non of these
resistance loci have been mapped (Sun and Steffenson, 2005).
A new pathotype of Pgt, designated race QCCJ, was identified on Midwestern barley
cultivars containing Rpg1 during the 1989 growing season (Roelfs et al., 1991). This race
increased in prevalence and became one of the most common virulence types in the US causing
minor epidemics (Roelfs et al., 1993). The widespread distribution of race QCCJ and the
susceptibility of barley grown on the Northern Great Plains warranted expectations of a coming
large epidemic, but it has never materialized. Barley production was threatened by race QCCJ
prompting the evaluation of over 18,000 barley accessions from the USDA National Small
Grains collection for sources of resistance. The best source of resistance against race QCCJ was
discovered in the unimproved barley line Q21861 from the International Maize and Wheat
Improvement Center (CMMYT) (Jin et al., 1994a). Genetic studies revealed that resistance to
race QCCJ in line Q21861 was conferred by a single recessive gene given the designation rpg4.
A recessive resistance gene is unusual among disease resistance genes suggesting that resistance
may be the result of a non-functional gene or null allele. The rpg4 gene is also temperature
sensitive providing resistance at lower temperatures (17-22°C), but completely ineffective at
temperatures above 27°C (Jin et al., 1994b). The rpg4 gene was genetically mapped to the long
arm of barley chromosome 5H(7) (Borovkova et al., 1995).
A new source of resistance to Pgt race QCCJ was recently discovered from a Hordeum
bulbosum introgression into the barley cv. Golden Promise (susceptible) background. (Fetch et
al., 2009) The rpg6 gene was identified in the translocation from H. bulbosum and maps to
chromosome 6H. Similar to rpg4, the rpg6 gene is also recessive.
8

Line Q21861 also contains resistant to an isolate of rye stem rust, Puccinia graminis f. sp.
secalis (Pgs) isolate 92-MN-90. Although Pgs 92-MN-90 was first isolated from rye hence the
name f. sp. secalis, it is closely related to Pgt and this forma specialis of stem rust infects barley
causing damage to the crop in some years (Steffenson, 1992). The single dominant resistant gene
was originally designated RpgQ, later changed to Rpg5 and was genetically mapped to the long
arm of chromosome 5H(7) very closely linked to rpg4 (Druka et al., 2000). Initially it was
believed that rpg4 and Rpg5 could be the same gene despite the different inheritance, recessive
vs dominant respectively.
A new threat to barley production worldwide is stem rust race TTKSK (original isolate
synonym Ug99), a pathotype first characterized from Uganda in 1999 (Pretorius et al., 2000).
Race TTKSK is considered highly virulent because it carries a unique combination of virulence
genes and is capable of attacking most barley cultivars, including those having Rpg1 (Steffenson
and Jin, 2006), and over 70% of the world’s wheat cultivars (Singh et al., 2008). Race TTKSK
has not reached North America but poses a threat to wheat and barley production in the US and
Canada because of its wide range of virulence and the ability of stem rust to spread rapidly over
long distances. Since the time it was initially discovered in 1999 (Pretorius et al., 2000), TTKSK
has spread in Eastern Africa, then disseminated over the Red Sea into Yemen, and has been
confirmed in Iran and possibly Pakistan (Nazari et al., 2009). It will be a short time before it
spreads into the major cereal production areas of southern Asia and ultimately will find its way
to North America.
A major global effort through the Borlaug Global Rust Initiative (BGRI) has been
launched to breed wheat varieties resistance to pathotype TTKSK and great progress has been
made (http://www.globalrust.org/about_us.cfm?m=2). However, relative to wheat very little
9

funding has been allocated to the barley breeding efforts due to its lower rank as a human food
crop. It has been overlooked by the BGRI that in countries where TTKSK now exists (Ethiopia,
Eritrea, and Yemen), barley is one of the most important food crops (Zemede, 1996) and these
resource poor farmers subsisting on barley should be a focus of the initiative. To effectively
control stem rust in these regions and elsewhere, it is important that both wheat and barley crops
are resistant since they are often cultivated in close proximity. This practice is important to keep
the Pgt population in check, otherwise the probability of virulent mutants arising on the
susceptible crop with the ability to start an epidemic on the previously resistant crop is greatly
increased (Schafer and Roelfs, 1985).
The barley line Q21861 was found to be one of the most resistant barleys to race TTKSK
at both the seedling stage in greenhouse and at the adult plant stage in the field (Steffenson,
personal communication). Understanding the genetics of race TTKSK resistance in Q21861 is
important for developing the most efficient strategy for deploying this source of resistance. It has
been determine that race TTKSK resistance in Q21861 is conferred by a single locus
cosegragating with rpg4 and Rpg5 (Steffenson, personal communication; Brueggeman et al.,
unpublished).
Line Q21861 is a good source of stem rust resistance but other sources of resistance
especially against race TTKSK need to be identified and transferred into adapted breeding lines.
Evaluation of Hordeum germplasm identified several sources of resistance to race TTKSK in
various landraces (H. vulgare) and wild barley (H. vulgare subsp. spontaneum) accessions
(Steffenson and Jin 2006). Sequence analysis of rpg4 and Rpg5 alleles from many of these lines
indicated that the majority contain functional alleles, however there is polymorphism among the
Rpg5 alleles. The rpg4/Rpg5 locus probably confers the race TTKSK resistance in these
10

landraces and wild barley accessions. Some Hordeum germplasm with resistance to race TTKSK
and a predicted non-functional Rpg5 allele were identified, suggesting that these lines may
contain novel sources of resistance (Brueggeman et al., unpublished). Allelism and mapping
studies are underway to confirm and characterize these new sources of stem rust resistance
(Brian Steffenson, personal communication).
It has been fortunate for Midwestern barley growers and breeders that the first barley
stem rust resistance source discovered was so remarkably durable. However, unfortunate that so
much reliance was placed on this single gene. It has long been known that reliance on a single or
limited numbers of stem rust resistance genes is dangerous because of the high probability of a
pathogen shift or the introduction of a virulent race. After 70+ years of exclusive use the new
virulent race TTKSK may bring the Rpg1 era to an end and barley production may once again be
threatened in the Midwestern United States and Prairie Provinces of Canada.
4. Resistance Genes
Plant R-genes mediate resistance to a diverse variety of pathogens including fungi,
bacteria, nematodes, viruses, insects and oomycetes. In 1955 Flor postulated that for each R-gene
there is a matching avirulence (Avr) gene in the pathogen, and that the presence of both is
required to elicit a resistance response in the plant. This gene-for-gene hypothesis was later
translated into a receptor-ligand model where the R-protein functions as the receptor that
recognizes or interacts with the corresponding Avr protein ligand. The protein-protein interaction
acts as the trigger initiating the signaling cascades resulting in defense responses leading to
resistance.
The lack of data showing that R-proteins directly interact with their corresponding Avr
ligands coupled with the identification of plant proteins that directly interact with both Avr and
11

R-proteins has led to an alternate hypothesis: the guard hypothesis (Van der Biezen and Jones,
1998). In this model the R-proteins (‘the guards’) monitor other plant proteins (‘the guardees’)
that are the target of modifications by the pathogen’s Avr effector proteins. The modification of
the guardee by the effector is detected by the R-protein triggering the defense response. This
model suggests that R-proteins maybe present in complexes containing the guardee and may not
have a direct protein-protein interaction with the Avr effector protein.
Advances in molecular techniques and tools have facilitated the cloning of ~ seventy
plant disease R-genes in the past three decades. These R-genes have been grouped into different
classes according to their protein domain structure (Martin et al., 2003). The majority of R-genes
belong to the NBS-LRR family of genes characterized by a N-terminal nucleotide-binding site
(NBS) and a C-terminal leucine rich region (LRR). Many of the NBS-LRR R-proteins have
different N-terminal domains. Most fall into one of two classes, the Toll and interleukin-1
receptors (TIR) or the putative leucine-zipper (LZ) motifs (Hammond-Kosack and Jones, 1997).
The only known function for this class of genes in plants is involvement in disease
resistance. However, in Arabidopsis a NBS-LRR gene confers susceptibility to the fungus
Cochliobolus victoriae (Lorang et al., 2007). Some NBS-LRR R-proteins have been shown to
interact directly with pathogen Avr proteins and the LRR is the critical component determining
pathogen specificity (Dodds et al., 2006). NBS-LRR genes also function to guard the effector
targets detecting modification by pathogen Avr proteins (Jones and Dangle, 2006).
The proteins encoded by the second major class of R-genes all contain at least one
serine/threonine protein kinase (STPK) domain. The STPK domain suggests that these proteins
function in plant signal transduction pathways. The seven members of the STPK group confer
resistance to bacterial and fungal pathogens and have been cloned from wide taxa of plant
12

species. This class of R-genes include the tomato Pto gene, which confers resistance to the
bacterial pathogen Pseudomonas syringae (Martin et al., 1993), the Arabidopsis PBS1 gene
conferring resistance to the bacterial pathogen Pseudomonas syringae (Swiderski and Innes
2001), the rice Xa21 and Xa26 genes which confer resistance to the bacterial pathogen
Xanthomonas oryzae (Song et. al., 1995; Sun et al., 2004), the recently cloned wheat Yr36 gene
conferring resistance to Puccinia striiformis (Fu et al., 2009) and the barley Rpg1 and Rpg5
genes (Brueggeman et al., 2002 [chapter 1]; Brueggeman et al., 2008 [chapter 4]). Rpg1 has a
novel R-gene structure containing two tandem STPK domains whereas the other STPK R-genes
contain a single protein kinase domain that can be connected to other protein domains
characteristic of R-genes; Xa21 and Xa26 have LRR-STPK structures (Song et. al., 1995; Sun et
al., 2004) and Rpg5 has NBS-LRR-STPK structure (Brueggeman et al., 2008 [chapter 4]). The
recently cloned Yr36 gene has a novel structure with a lipid binding domain attached to the
STPK domain (Fu et al., 2009).
5. Rust Resistance Genes; Cloning and Characterization
Sixty years ago J.B.S. Haldane predicted that polymorphisms in proteins would govern
host-pathogen interaction and this variability was the result of the rapid rate of microbial
evolution. His knowledge of the variability of stem rust resistance in wheat spurred his
suggestion that host plants could only react by generating high levels of polymorphism in their
immune system genes (Haldane J.B.S., 1949). Harold Flor was a pioneer of rust resistance gene
research and provided evidence to support these speculations. In the 1940s he crossed many flax
rust races then determined the disease reaction on a large number of flax lines when infected
with the progeny of his crosses. Flor’s flax rust research determined that the rust pathogen
13

Melampsori lini pathogenicity was genetically inherited as single dominant genes (Flor H.H,
1946). Later Flor proposed the gene for gene hypothesis discussed earlier (Flor H.H, 1956).
Moving into the molecular genetics era flax rust resistance has maintained its status as the
most extensively studied rust resistance systems. Five resistance loci from Flax have been
mapped (L, M, P, N, and K) (Ellis et al., 1988) and four R-genes have been cloned and
characterized. (L, M, P and N) (Anderson et al., 1997; Dodds et al., 2001; Lawrence et al.,
1995). All four characterized Flax R-genes code for TIR-NBS-LRR domain proteins. It was
determined that the race specificity between the different L alleles was determined by
polymorphisms within the LRR domain suggesting that the LRR domain could be responsible
for Avr binding/detection (Ayliffe et al., 1999).
The subject of R-genes and their underlying mechanisms can not be fully understood
without the identification and characterization of the pathogens Avr protein; proteins that directly
or indirectly interact with the plant R-gene to illicit the resistance response. Many Avr proteins
have been identified from a large taxonomy of pathogens and most encode small peptides from
very diverse classes of proteins (Abramovitch and Martin, 2004; Catanzanti et al., 2007). The
first rust Avr gene was cloned from flax rust by a map-based cloning approach (Dodds et al.,
2004). This Avr gene was designated the flax AvrL567 gene because its protein product is
recognized by L5, L6 and L7 and was later determined to interact directly with all three
corresponding R-proteins in vivo and in yeast two-hybrid assays (Dodds et al., 2006). AvrL567
was shown to be expressed in the haustoria and encodes a 127 aa secreted protein that is
delivered to the host cells across the plasma membrane (Dodds et al., 2004).
Approximately 10% of the R-genes cloned are rust resistance genes from the small cereal
grains. These cloned R-genes are the maize Rp1 gene conferring resistance against P. sorghi
14

(Collins et al., 1999), the wheat leaf rust resistance genes Lr10, Lr21 and Lr34 (Feuillet et al.,
2003; Huang et al., 2003; Krattinger et al., 2009) conferring resistance against Puccinia triticina,
the wheat stripe rust resistance gene Yr36 (Fu et al., 2009) conferring resistance against Puccinia
striiformis and the barley stem rust resistance genes Rpg1, rpg4 and Rpg5 described here.
Rp1 and Rp3 of maize are complex loci consisting of clusters of closely related NBS-
LRR genes (Collins et al., 1999; Webb et al., 2003). The Rp3 gene has been delimited to a
complex locus containing a NBS-LRR gene family consisting of nine genes but the specific gene
conferring the resistance has not been identified (Webb et al., 2003). These complex loci have a
tendency to mispair and recombine in meiosis creating haplotypes that are structurally variable.
For example, Rp1 haplotypes in different maize lines vary in the number of Rp1 genes they carry
from a single gene to greater than 50 (Smith et al., 2004). Recombinant genes and haplotypes
have been identified that have different reaction to pathotypes than the original parental types
(Hulbert, 1997). In addition to typical race specific resistance genes like Rp1-D, recombinant
genes have been identified that give an extensive hypersensitive reaction to inoculation with any
rust isolate, including non-host species like the wheat rusts (Hu et al., 1996). Unequal
recombination within these complex loci appears to be an important process to generate new
combinations of NBS-LRR genes resulting in more diversity and new pathotype specificities.
The wheat R-genes Lr10 and Lr21 confer race specific resistance to Puccinia triticina.
Both genes were cloned by map-based cloning strategies and determined to code for typical
NBS-LRR R-genes. These rust resistance loci also appear to be part of complex loci containing
several copies of NBS-LRR genes from the same and different families (Huang et al. 2001)
similar to the Mla resistance locus in barley which contains three distinct NBS-LRR gene
families, cosegregating with Mla (Wei et al., 1999). The recently cloned wheat leaf rust
15

resistance gene Lr34 has been shown to code for a putative ABC transporter. The current model
for the function of the transporters is to translocate toxic compounds into the apoplast (Krattinger
et al., 2009). Lr34 has conferred durable resistance against leaf rust for 50 years but is also
associated with resistance against stripe rust and powdery mildew. This source of durable adult
resistance when deployed with other adult plant resistance genes provides a resistance level near
immunity and has been deployed in wheat cultivars worldwide.
Map-based cloning was also used to recently clone the Yr36 gene that confers resistance
to wheat stripe rust (Puccinia striiformis) (Fu et al., 2009). Yr36 confers broad-race resistance
(slow rusting) to stripe rust at relatively high temperatures (25-35oC). New races of stripe rust are
virulent on most of the known race-specific resistance sources due to the rapid pathogen
evolution but the slow rusting genes providing partial adult resistance are generally durable. The
Yr36 gene has been identified and contains two protein domains, a STPK and a START lipid
binding domain. The authors proposed the possibility that the START domain may bind a lipid
from or redirected by the pathogen which then initiates the signaling cascade via the protein
kinase domain resulting in resistance (Fu et al., 2009).
It is important to note that no rust resistance genes are listed here from rice, the model
cereal grain. Rice is not afflicted by rust thus is a non-host species. This has been the focus of
controversial molecular genetics research of rust, initiated by the nobel laureate Norman
Borlaug. It is his belief or as he stated “his dream” that the mechanisms that underlie the
immunity in rice could be determined then transferred to wheat (Borlaug, 2000). A substantial
amount of research funding maybe directed toward this line of research which could probably be
better directed at the understanding of the mechanisms that underlay resistance in barley and
wheat, the major cereals actually afflicted by the disease. It should be kept in mind that R-gene
16

complexes or mechanisms although made up of many similar components and classes of proteins
may react to the pathogen in different ways, especially across species that behave as host and
non-host.
B. Rpg1 CLONING AND CHARACTERIZATION
In order to characterize the barley rust resistance genes and the underlying mechanisms
involved in resistance first they must be identified and cloned. Dr. Andris Kleinhofs initiated the
cloning of stem rust resistance genes in 1995 soon after his lab developed an extensive genetic
map of barley. The development of the Steptoe/Morex map (Kleinhofs et al., 1993) prompted the
desire to utilize this resource to clone an agronomically important gene by a map-based
approach. At this time map-based cloning was in its fledgling stages especially in a large grass
genome such as barley. The Rpg1 gene was chosen as a target for several reasons most
importantly that it was a remarkably durable resistance gene present in the cv. Morex, one of the
parents in the mapping population. It was also a quantitatively or simply inherited gene easily
phenotyped and had recently been mapped to the telomeric end of barley chromosome 7H(1).
This was fortunate because the syntenic region in rice was also being sequenced and prompted
the hope of exploiting rice/barley synteny to identify the gene quickly. While synteny with rice
did provide valuable molecular markers, which advanced the fine mapping in barley (Kilian et
al., 1997), a candidate Rpg1 gene was not found in the rice chromosome syntenic region (Han et
al., 1999). This intensified the effort in Dr. Kleinhofs’ lab and after the development of many
molecular tools the arduous task of cloning the first stem rust resistance gene was completed in
2002 (Brueggeman et al., 2002 [paper I])
17

The large (>5,000 Mb), repetitive (>80%) barley genome makes map-based cloning very
labor intensive and time consuming yet has been proven as the most effective strategy for
identifying barley R-genes. Mlo was the first barley R-gene cloned by map-based cloning using
a yeast artificial chromosome (YAK), library (Buschges et al., 1997). The subsequent
construction of a bacterial artificial chromosome (BAC) library from the barley cv. Morex (Yu et
al., 2000) was the pivotal tool in the more recent cloning of the barley powdery mildew
resistance genes Mla1 and Mla6 (Halterman et al., 2001; Zhou et al., 2001) and the stem rust
resistance gene Rpg1, rpg4 and Rpg5 (Brueggeman et al., 2002 [chapter 1]; Brueggeman et al.
2008 [chapter 4]). The development of the barley Steptoe/Morex Bin maps in conjunction with
molecular tools (EST libraries (Close et al., 2001), BAC clone libraries (Yu et al., 2000), and the
sequence of related organisms (Sasaki and Burr, 2000)) has made map-based cloning in barley a
much more efficient strategy. Utilizing syntenous regions of the sequenced genomes of rice and
Brachypodium for molecular marker development, high quality genetic maps and the cv. Morex
BAC library we have developed physical contigs across the rust resistance regions of Rpg1
(Brueggeman et al., 2002 [paper 1]), rpg4/Rpg5 (Druka et al., 2002; Brueggeman et al., 2008
[paper 4]) and the spot blotch resistance locus Rcs5 (Drader et al., personal communications).
1. Rpg1 Map-Based Cloning
A high resolution genetic map was constructed using 17 critical recombinants selected
from 8,518 gametes by genotyping with co-dominant flanking markers. A 330 kilobase BAC
contig was developed across the locus and the physical map positioned the gene between two
cross overs a genetic distance of .21 cM and a physical distance of 110 kb (Brueggeman et al.,
2002 [chapter 1;Fig 1]). Two STPK genes were identified in the region by low copy probe
development and sequencing. Allele sequencing from resistant and susceptible lines and a
18

fortuitous recombinant (AMS170) combining parts of resistant and susceptible alleles of Rpg1
confirmed the identity of the correct candidate gene (Brueggeman et al., 2002 [chapter 1;Fig 3])
The Rpg1 gene has a unique gene structure encoding two tandem kinase domains within a
genomic sequence of 4,466 bp including 14 exons producing an in silico translated protein of
837 aa (~94.5 kDa) (Brueggeman et al., 2002 [chapter 1;Fig 2]). The RPG1 protein appears to be
a typical kinase except for the tandem kinase domains, a structure not previously reported in
plant disease resistance genes. The only report of structural and perhaps functional similarity
comes from the animal Janus Kinases (JAKs), which consists of two tandem kinase domains and
function in signal transduction pathways activated by diverse cytokine receptors (O’Shea and
Leonard, 1998).
2. Gene Family Characterization
It has been reported that resistance to specific races of a pathogen can be conferred by
different alleles at the same locus as with the Flax L locus (Dodds et al., 2004) or diverse
members of resistance gene families at different loci as is the situation with Mla1 and Mla6
(Halterman et al., 2001; Zhou et al., 2001, Huang et al., 2001). In an attempt to quickly identify
other candidate stem rust resistance genes that confer different resistance specificity than Rpg1,
particularly candidate rpg4 or Rpg5 genes, we analyzed the cv. Morex genome for Rpg1 gene
family members by nucleotide hybridization assays with a Rpg1 specific probe (Brueggeman et
al., 2006 [chapter 2]). The initial hybridizations to cv. Morex BAC and cDNA libraries identified
three additional genes belonging to the Rpg1 gene family. The Rpg1 structure containing two
tandem kinase domains had not been previously reported, but all three initially identified Rpg1
gene family members, designated ABC1036, ABC1037 and ABC1040, contained the dual kinase
domain structure (Brueggeman et al., 2006 [chapter 2;Fig. 1]). Of these, ABC1037 had the
19

highest homology with Rpg1 and was very closely linked to Rpg1 (ca. 50 kb) suggesting the
locus was the result of unequal recombination giving rise to a tandem duplication. ABC1037 is
expressed and contains high homology to Rpg1 at the kinase domains but diverges at the 5’ end
and has no known function.
ABC1036 and ABC1040 are highly homologous one to another and tightly linked on
chromosome 5H(7) probably due to another tandem duplication, but they are highly diverged
from Rpg1 at the kinase 2 domain. The two loci at 7H(1) and 5H(7) suggest an ancient
duplication followed by a translocation event then divergence leading to the two loci containing
the tandem kinase genes.
Hybridization with ABC1036 and ABC1040 specific probes identified two additional
genes, ABC1041 and ABC1063. These genes had relatively low homology to Rpg1, but were
highly homologous to the protein kinase domain 2 of ABC1036 and ABC1040 and contain only
a single kinase domain (Brueggeman et al. 2006 [chapter 2;Fig 1 and 4]). ABC1041 mapped
very close to the rpg4/Rpg5 genes but segregate away in the high-resolution population used to
clone rpg4 and Rpg5.
The presence of three tandem kinase domain genes and their scattered location in the cv.
Morex genome suggest that this tandem kinase structure has an ancient origin. A search of the
sequenced plant genomes Aabidopsis, Oryzae, and draft sequence of Brachypodium did not
identify any Rpg1 orthologs with this dual kinase structure, but genes with high amino acid
homology to the individual kinase domains were identified. PCR using barley Rpg1 specific
primers on wheat genomic DNA produced sequences with high homology to Rpg1, but the PCR
bands were very heterogeneous and assembling the ~1kb overlapping PCR fragment into
contiguous unigene sequences was impossible (The PCR primers used are described in Mirlohi
20

et al., 2008 [Chapter 3; Table 2]. We cloned the heterogeneous PCR products from wheat and
sequenced many independent clones. Single nucleotide polymorphism (SNP) analysis of the
PCR derived clones identified at least eight and possibly more copies of Rpg1 in wheat
(Brueggeman et al., unpublished). A Triticum monococcum BAC library was screened using a
barley Rpg1 specific probe and a BAC contig was assembled containing three tandem repeats of
dual kinase Rpg1 homologous genes. Mapping of these Rpg1 orthologs placed them in the
syntenic region of the wheat genome. Therefore, the tandem kinase gene structure in plants is not
unique to Hordeum. The detection of Rpg1 orthologs in wheat (Brueggeman et al., unpublished)
shows that the Rpg1 gene has an ancient evolution occurring before the wheat barley split ~13
mya (Wolfe et al., 1989).
Our attempts at identifying the rpg4/Rpg5 genes or other candidate resistance genes were
initially unsuccessful, but we did identify a family of genes with homology to Rpg1. Genetic
mapping did not identify a putative function for any of the genes, however one family member,
ABC1041, was tightly linked to the rpg4/Rpg5 locus. After cloning Rpg5 by a map-based
approach amino acid comparisons revealed that the Rpg1 gene family members ABC1041 and
ABC1063 were highly homologous to the Rpg5 kinase domain (60% and 61% aa identity,
respectively, and 76% aa similarity). The nucleic acid homology between ABC1063 and Rpg5 is
sufficiently high that cross hybridization could occur. We certainly could have identified Rpg5
with our assays, screening BAC and cDNA libraries, however the Rpg5 allele from cv. Morex is
missing the kinase domain. Thus in hind sight, screening the susceptible cv. Morex cDNA and
BAC libraries to identify Rpg5 was a futile effort.
21

3. Rpg1 Allele Characterization
Sequence analysis of the Rpg1 gene from seven stem rust resistant North American
barley cultivars yielded no polymorphism at the nucleotide level suggesting that North American
germplasm contains a single source of Rpg1 probably originating from the unimproved bulked
seed lot obtained by the USDA from Switzerland in 1914 (Brueggeman et al., 2002 [chapter 1]).
In order to trace the origin of the Rpg1 resistance gene from this bulked seed lot we analyzed the
Rpg1 alleles from eight land races (Hordeum vulagare) from Switzerland. We also analyzed
alleles from eight wild barley (H. vulgare ssp. spontaneum) accessions originating from diverse
geographic regions to attempt to gain insight into the origin of the Rpg1 gene.
A set of 74 barley landraces from Switzerland and 318 ecogeographically diverse H.v.
spotaneum wild barley accessions were tested with Pgt race MCCF. Eight barley lines, four
resistant and four susceptible to race MCCF, from each of the landraces and wild barleys were
initially selected. Then a comprehensive evaluation of known stem rust resistance was brought to
our attention. This study revealed that Pgt race HKHJ was specific for Rpg1-mediated stem rust
resistance while race MCCF was shown to be avirulent on barley containing the rpg4/Rpg5 locus
(Sun and Steffenson, 2005). Only one Swiss landrace and none of the H.v. spontaneum lines
used in this study showed resistance to race HKHJ. The Swiss line resistant to race HKHJ proved
to have an Rpg1 gene identical to that previously sequenced from North American barley
cultivars (Mirlohi et al., 2008 [chapter 3; Fig. 3]) and probably comes from the same source as
the one imported to USA in 1914. As expected by the HKHJ phenotypes none of the other lines
had a functional Rpg1 gene. The lines that were resistant to race MCCF and susceptiple to race
HKHJ were subsequently genotyped at the rpg4/Rpg5 locus. The resistance correlates with
functional rpg4/Rpg5 alleles suggesting it is due to the rpg4/Rpg5 genes (Brueggeman et al.
22

unpublished). These data indicate that a functional Rpg1 gene is rare in the H.v. spontaneum and
Swiss land race populations, however a larger collection need to be characterized to confirm this.
A GTT insertion was present in all seven barley (landrace or wild) accessions containing
a defective Rpg1-like gene making it a common allelic variation. This insertion results in an S to
R substitution at amino acid position 319 and an F insertion at position 320 leading to virulence
by both races HKHJ and MCCF (Mirlohi et al., 2008 [chapter 3;Fig 3]). We do not know if the
Rpg1 gene with a GTT insertion carries out some other function or confers resistance to other
Pgt races, but its presence in a diverse population and its status as an apparently functional gene
in the wild barley population suggests that its function possibly has a selective advantage in the
wild. The wild barleys had Rpg1 alleles that encoded intact proteins with the GTT insertion
although they are non-functional in known stem rust resistance, but still have
autophosphorylation activity confirming previous observations that autophosphorylation is
necessary but not sufficient for resistance (Nirmala et al., 2006 ). Most of the cultivated barleys
with the GTT insertion have acquired mutations that produce a truncated protein thus appear to
be psuedogenes.
Recent in vitro assays using the Yeast-two-Hybrid system in our lab has demonstrated
that the GTT insertion within the pseudokinase domain does cause interference with protein-
protein interaction between RPG1 and at least three proteins identified as interactors (Nirmala,
personal communication). This exemplifies the importance of identifying natural allelic variation
and the information that these alleles can provide in determining amino acids or domains that are
functionally important. These allelic variations can be as valuable as induced mutants when
elucidating the molecular function of a gene.
23

4. Rpg1 Functional Analysis
The cloning and characterization of the durable stem rust resistance gene Rpg1 has
greatly enhanced the understanding of the underlying mechanisms of stem rust resistance. In the
years following the cloning of Rpg1 in 2002, we have come a long way in elucidating the Rpg1
mediated mechanisms of resistance. However, a great deal of work still remains.
The validation of the Rpg1 gene (Brueggeman et al., 2002 [chapter 1]) was subsequently
confirmed by Agrobacterium mediated stable transformation of the susceptible cv. Golden
Promise (Horvath et al., 2003). Golden Promise had been shown to be missing the Rpg1 gene
presumed to be either deleted or highly diverged (Brueggeman et al., 2002 [chapter I]). Several
independent transformants reacted to the pathogen with a much higher resistance than the cv.
Morex from which the gene originated. Quantitation of the Rpg1 mRNA levels from transgenic
plants containing different copy numbers of Rpg1 (1-5 copies), showed various levels of Rpg1
mRNA, but there was no correlation between Rpg1 mRNA levels and resistance to Pgt race
MCCF (Horvath et al. 2003). The potential for other genes within the background of various
cultivars even the susceptible cultivars that affect the level of resistance conferred by a race
specific R-gene may have utility when breeding for resistance. It is also important to identify this
factor or factors to gain a further understanding of why or how they are enhancing the Rpg1
mediated resistance. These genes also may provide further insight into elucidating the resistance
mechanisms.
Expression analysis of Rpg1 mRNA showed that it is constitutively and uniformly
expressed in all plant organs and developmental stages except the leaf epidermis where it
exhibited a 30-fold higher level than in the whole leaf (Rostoks et al., 2004). P graminis
penetrates the barley leaf through the stomata so the elevated Rpg1 expression in the leaf
24

epidermis possibly indicates that the fungus is detected at the leaf surface. It was determined that
the RPG1 protein is mostly cytoplasmic with small but significant amounts associated with
membranes (Nirmala et al., 2006). This membrane association is similar to the animal janice
kinases (JAKs), which share the dual kinase structure with Rpg1 and provide a model for the
Rpg1 mechanism. The JAKs were originally determined to be cytosolic proteins, but recent
analysis has shown that they are loosely bound to the plasma membrane (Behrmann et al., 2004)
and associate with transmembrane cytokine receptors (Aaronson and Horvath, 2002). The
function of the membrane bound fraction of RPG1 protein maybe similar to JAKs by associating
with a transmembrane receptor (Aaronson and Horvath, 2002). Recent data using a technique for
detection of in vivo RPG1 phosphorylation may substantiate this speculation (Nirmala, personal
communication).
Although the structural similarity of the RPG1 protein and the animal JAKs is probably
the result of convergent evolution and the Rpg1 association with a surface receptor is speculation
there are other similarities shared between the two systems. Both RPG1 and the JAKS have a
functional kinase domain and a nonfunctional pseudokinase (Nirmala et al., 2006). The role of
the pseudokinase domain in RPG1 function appears to be important in protein-protein
interaction. Mutating two adjacent lysine’s (K152 and K153) of the pseudokinase domain did not
inactivate the RPG1 protein kinase auto catalytic activity, but did result in a susceptible disease
reaction of the stable transformant (Nirmala et al., 2006). The wild-type RPG1 interacts with ten
different proteins in the yeast two-hybrid system, but the K152 and K153 pseudokinase mutant is
known to lose interaction with at least three of these proteins (Nirmala, personal
communication). When the RPG1 pseudokinase was completely deleted it also abolished all
interactions with proteins that interacted with the wild type RPG1.
25

Similarity between the two systems is also evident by the degradation of the RPG1
protein approximately 24 hours after avirulent pathogen challenge (Nirmala et al., 2007). This
degradation was determined to occur via the ubiquitin-proteasome pathway and it was also
shown that JAK2 also degrades after activation via the ubiquitin-proteasome pathway
(Ungureanu et al., 2002).
Another similarity between the RPG1 kinase and JAKs is the interaction with
transcription factors, but this similarity is also shared with resistance pathways activated by an R-
gene with only a single kinase domain. Four genes identified as downstream Pto interactors by
the yeast two-hybrid system were also identified to interact with RPG1. These genes were Pti1, a
serine/threonine protein kinase involved in the HR response, and the transcription factors Pti4,
Pti5, and Pti6 that function in the expression of pathogenesis-related (PR) genes (Zhou et al.,
1997). The Pti4/5/6 transcription factors are related to the ethylene-response-element binding
proteins and bind the GCC box found in the promoter regions of many pathogenesis related
genes (Zhou et al., 1997). The JAKs interact and phosphorylate the Signal Transducers and
Activators of Transcription (STAT) factors (Leonard, 2001).
C. MAP-BASED CLONING OF rpg4 AND Rpg5
The Rpg1 gene has conferred remarkable durability against stem rust but the new races that have
emerged, QCCJ and TTKSK, are highly virulent on barley containing Rpg1. The rpg4/Rpg5
locus has been shown to confer resistance to both of these exceptional races warranting the
cloning and characterization of this locus and the genes underlying this important source of stem
rust resistance. We have cloned the rpg4/Rpg5 locus and shown that these two genes are distinct
and both code for novel resistance proteins. The Rpg5 gene confers resistance to an isolate of
Pgs and rpg4 confers resistance to Pgt. We have discovered that while resistance to Pgs isolate
26

92-MN-90 is conferred by Rpg5 alone, resistance against the Pgt races QCCJ, MCCF and
TTKSK requires both Rpg5 and rpg4.
1. Cloning and Characterization of Rpg5
High-resolution genetic mapping provided initial localization of the rpg4/Rpg5 locus to a
small region of chromosome 5H(7) (Druka et al., 2000; Han et al., 1997; Kilian et al., 1997). A
70 kb region between markers ARD5016 and ARD5112 was cloned and sequenced
(Brueggeman et al., 2008 [chapter 4; Fig. 1]). Sequence annotation revealed five candidate
genes, two encoding predicted NBS-LRR proteins (HvRGA1 and HvRGA2), two actin
depolymerizing-like factors (HvADF2 and HvADF3) and a protein phosphatase 2C-like protein
(HvPP2C) (Brueggeman et al., 2008 [chapter 4; Fig. 1]). The sequence was derived from the
susceptible cv. Morex BAC clones therefore the equivalent sequence was needed from the
resistant line Q21861. The Q21861 sequence was generated by PCR approaches utilizing the cv.
Morex sequence and development of a Q21861 lambda phage library. Annotation of the Q21861
sequence revealed that the HvPP2C gene was missing and replaced by a STPK gene just 3 prime
of the HvRGA2 gene. Allele sequencing of the candidate genes HvRGA1, HvADF2 and
HvADF3 from susceptible barley lines showed that they were essentially identical to those found
in the resistant line Q21861 eliminating them as candidate Rpg5 genes. The HvRGA2 allele from
Q21861 was left as the only candidate Rpg5 gene conferring resistance to Pgs isolate 92-MN-90
and was confirmed by sequencing alleles from several susceptible genotypes.
It was determined by RT-PCR and northern blot analysis that the NBS-LRR and STPK
domains were transcribed in a single mRNA, a unique gene structure among plant disease
resistance genes (Brueggeman et al., 2008 [chapter 4; Fig 3]). Annotation using cDNA to gDNA
comparisons predicted the Rpg5 gene to contain seven exons in a total genomic sequence of
27

8,504 bp coding for an 1,378-aa (151.6-kDa) protein (Brueggeman et al., 2008 [chapter 4; Fig.
2]).
A technique optimized for the study of barley functional genomics is Viral Induced Gene
Silencing (VIGS). VIGS is a form of post translational gene silencing (PTGS) utilizing infectious
virus as a vector to introduce dsRNA into a host. VIGS is used to silence or reduce the transcript
of an endogenous plant gene by use of a virus genomic RNA with an inserted anti-sense or
inverted repeat with homology to the target gene. The viral infection induces the formation of
dsRNA triggering the plant defense response and the degradation of the target gene transcript
resulting in a loss of function. VIGS has been developed for barley using the Barley Stripe
Mosaic Virus (BSMV) as a vector (Holzberg et al., 2002). We utilized this method for reverse
genetics to confirm the NBS-LRR-STPK gene as Rpg5. Using QRT-PCR we demonstrated a
correlation between silencing the candidate Rpg5 gene and the conversion of the resistant line
Q21861 to a susceptible phenotype when inoculated with Pgs isolate 92-MN-90 (Brueggeman et
al., 2008 [chapter 4; Fig 3 and 4]).
Present work in our lab has identified parallels between the Rpg1 mechanism of
resistance and the Pto mechanism. The Pto gene of tomato conferring resistance to races of P.
syringae carrying the avrPto or AvrPtoB genes (Martin et al., 1993; Kim et al., 2003) contains a
single kinase domain with homology to the PKDs of Rpg1 (29% a.a. identity and 48% a.a.
similarity to PKD2 of Rpg1) and Rpg5 (36% aa identity and 53% aa similarity). The Pto
resistance function requires the presence of a second gene Prf, which is a classic NBS-LRR gene
(Salmeron et al., 1996). Elucidation of the barley stem rust Rpg1 mediated resistance pathway is
underway yet Rpg1 has not been shown to require an NBS-LRR gene for resistance. However,
in-vivo co-immuno precipitation experiments using a full length Rpg1 specific antibody have
28

identified a putative NBS-LRR gene by MASS Spec analysis (Nirmala, personal
communication). Interestingly, the Rpg5 gene contains all three domains and has aa homology to
Pto and Prf (the NBS-LRR homology to PRF is 26% aa identity and 42% aa similarity). Pto
requires two genes, an NBS-LRR and protein kinase, for resistance and Rpg1 probably does as
well, but the Rpg5 gene is a novel R-gene containing the components of both genes in a single
transcript. It may be possible that having all three domains in a single transcript could facilitate
the identification of not only the rust Avr protein or proteins in this system by yeast-two hybrid
or co-immuno precipitation but also could be valuable in identifying components down stream of
Rpg5 in the resistance pathway or components of the resistance complex. Rpg5 maybe used as a
tool to better understand these mechanisms with all three components present in a single protein.
This may not only facilitate the understanding of stem rust resistance pathways but resistance
pathways in general.
2. Interaction of rpg4 and Rpg5 for P.g. f. sp. tritici Resistance
Recombinant analysis of the 70 kb rpg4/Rpg5 region delimited the rpg4 gene to a 1kb
interval encoding only the HvAdf2 gene (Brueggeman et al., 2008 [chapter 4; Fig. 1]). HvAdf2
(rpg4) encodes an apparently functional Adf gene containing three exons in a total sequence of
906 bp coding for an 147 amino acid (16.17-kDa) protein. Current evidence suggests that the
rpg4 gene does not act independently, but rather that it is dependent on the Rpg5 gene for
function (Brueggeman et al., 2009 [chapter 5]). The Pgt QCCJ and TTKSK susceptible barley
cv. Harrington and line Sm89010 contain an rpg4 allele identical at the amino acid level to the
rpg4 allele in the resistant line Q21861. This initially suggested that HvAdf2 was not the rpg4
gene. However, analysis of allele sequences of rpg4 and Rpg5 coupled with SNP analysis of the
critical recombinants showing that the genes were distinct, suggested that the susceptibility of
29

these barleys was due to a non-functional Rpg5 gene. The susceptible cv. Harrington and line
Sm89010 contain truncated Rpg5 alleles missing the protein kinase domain. The
Harrington/Q21861 recombinant #18 (HQ18) combines an apparently functional cv. Harrington
rpg4 allele (which is identical at the amino acid level to the Q21861 rpg4 allele) with the
functional Q21861 Rpg5 gene. HQ18 is resistant to Pgt QCCJ and TTKSK, indicating that
Harrington is susceptible to the Pgt races due to a lack of a functional Rpg5 gene (Brueggeman
et al., 2009 [chapter 5]).
Phenotype analysis of three populations (Morex, Steptoe, and Robust/Q21861) revealed
1:3 segregation for resistance:susceptibility, demonstrating the recessive nature of rpg4 (Jin et
al., 1994a). The three Pgt susceptible barley cultivars (Morex, Steptoe, and Robust) have
identical Adf2 alleles at the amino acid level and differ from the resistant Q21861 allele by three
amino acids (Q39H, A101T, S135G). Three Pgt susceptible progeny from the Steptoe/Q21861
population have recombination within the Adf2 gene and contain a functional Rpg5 allele. These
recombinant Adf2 genes contain Q39 (Q21861-like) with T101 and G135 (Steptoe-like), showing
that the amino acids at positions 101 and/or 135 are important for reaction to Pgt QCCJ and
TTKSK. All Adf2 alleles tested were expressed at the transcription level, indicating that these
two amino acid polymorphisms determine susceptibility/resistance to Pgt QCCJ and TTKSK.
These data suggest that Pgt resistance should behave as a single dominant gene in
populations where only the Rpg5 gene is segregating in the presence of a non-polymorphic rpg4
gene (Harrington/Q21861 and Sm89010/Q21861). This hypothesis is currently being tested in
the Harrington/Q21861 and Sm89010/Q21861 F1 and F2 populations and by down-regulating the
two alternative rpg4 alleles using the BSMV-VIGS method (Brueggeman et al., unpublished).
30

The rpg4 gene is genetically recessive, suggesting that it may have a negative or neutral
function on the fungus’ ability to establish itself within the plant. One hypothesis is that the
fungus captures the dominant ADF2 protein function to facilitate feeding itself via the haustoria
and that the fungus is unable to utilize the rpg4 encoded ADF2 protein to establish or maintain
its feeding structure. Establishment of the fungal haustorial apparatus and/or haustorium function
probably has a dependency on ADFs. Stem rusts are biotropic fungi that invade plants in non-
destructive manner developing the haustoria after penetration of mesophyll cell wall. The
haustoria is associated with host plasma membrane by invagination of the plasma membrane
creating the haustoria-plant cell interface that consists of the haustorial membrane, an extra-
haustorial matrix, and the host plasma membrane (Schulze-Lefert and Panstruga 2003). It has
been shown that in the compatible interaction between barley and powdery mildew the
haustorium is associated with actin filament rings and actin filaments closely follow the
haustorium when invaginating the plasma membrane (Opalski et al., 2005). This is similar to
what occurs in the symbiotic interaction between a mycorrhizal fungus and tobacco root cells
(Genre and Bonfante, 1998), suggesting similarity between how pathogenic and symbiotic fungi
may establish the fungus-host interface. All these processes are dependant on cytoskeletal
restructuring which is an ADF dependant process.
The rpg4 gene by itself is not sufficient to confer resistance, perhaps due to failure to
detect the fungus via Rpg5. An appealing hypothesis is that the RPG4 protein function in
compatible or incompatible situations can be explained by differential phosphorylation at the two
important amino acids T101 in the Q21861 ADF protein or S135 in the Steptoe ADF protein. In
either case it suggests that rpg4 may act downstream of Rpg5 and is possibly activated or
deactivated by direct or indirect phosphorylation of RPG4 by the RPG5 kinase activity. If the
31

ADF protein is present from the Rpg4 susceptible allele in the heterozygous or homozygous
state, RPG5 may be unable to activate or deactivate the protein thus the fungus can utilize it to
establish itself within the plant. This hypothesis explains rpg4 as a recessive gene.
RPG5 could also be the guard protein monitoring the ADF2 protein for any modifications
by the fungus effector; however, this would not explain why rpg4 is recessive. RPG5 should still
be able to guard the ADF2 protein even when the rpg4 gene is present in a heterozygous state.
The temperature sensitivity of the rpg4 mediated resistance must also be addressed (Sun
et al., 1994b). The classic interpretation of temperature sensitivity is that the protein structure is
stable and functional at the lower temperature, but may become unstable at higher temperatures.
It is possible that the ADF protein is temperature sensitive and the elevated temperatures affect
the actin depolymerizing or polymerizing process. However, it could also be other proteins or
components within the pathway that are temperature dependent. Since Rpg5 is required for the
rpg4 mediated resistance response it could possibly be the RPG5 protein acting in a temperature
dependant manner. It has been demonstrated that an ADF is required for cold hardiness in wheat
and seems to be required for the cytoskeleton rearrangement that under lays this cold tolerance
(Ouellet et al., 2001). This ADF protein appears to be phosphorylated by a 52kDa protein kinase
activity which is temperature dependent suggesting the kinase protein in this system acts in a
temperature dependant manner. Post pathogen recognition, RPG5 may act upon the ADF for
activation or deactivation via phosphorylation and RPG5 phosphorylation activity maybe
temperature dependant when phosphorylating ADF2 or other substrates.
Plant genomes have limited numbers of Adf genes as demonstrated by 12 Adf genes
present in the sequenced genomes of Arabidopsis and rice (Feng et al., 2006). This suggests that
ADF proteins have several different functions and this may also be true of rpg4 (HvAdf2). It is
32

reasonable to speculate that biotrophic fungi evolved to utilize one or more of these limited
number of ADF proteins to establish or maintain their haustorial feeding apparatus. It may
become possible to analyze the limited number of ADF proteins present in plants to specifically
detect which ADF proteins the biotrophic fungi are utilizing to establish themselves within the
host. This makes the rpg4/Rpg5 resistance mechanism of particular research interest because it
maybe a very important system to begin understanding how the fungus establishes and maintains
its biotrophic life style. This scientific knowledge could be invaluable in our efforts to manage
not only stem rust, but also many diseases caused by biotrophic fungal pathogens.
33

REFERENCES
Aaronson D.S. and Horvath C.M. (2002) A road map for those who don't know JAK-STAT.
Science 296:1653-1655.
Abramovitch R.B. and Martin G.B. (2004) Strategies used by bacterial pathogens to suppress
plant defenses. Curr Opin in Pl Bio 7:356-364.
Anderson P.A., Lawrence G.J., Morrish B.C., Ayliffe M.A., Finnegan E.J. and Ellis J.G.
(1997) Inactivation of the flax rust resistance gene M associated with loss of repeated unit
within the leucine-rich repeat coding region. Plant Cell 9:641-651.
Ayliffe M.A., Frost D.V., Finnegan E.J., Lawrence G.J., Anderson P.A. and Ellis J.G.
(1999) Analysis of alternative transcripts of the flax L6 rust resistance gene. Plant J.
17:287-292.
Borlaug N. E. (2000) Ending world hunger: The promise of biotechnology and the threat of
antiscience zeolotry. Plant Physiology 124:487-490.
Borovkova I.G., Steffenson B.J., Jin Y., et al., (1995) Identification of molecular markers
linked to the stem rust resistance gene rpg4 in barley. Phytopathology 85:181-185.
Behrmann I., Smyczek T., Heinrich P.C., Schmitz-Van de Leur H., et al. (2004) Janus kinase
(Jak) subcellular localization revisited. J. Biol. Chem. 279:35486-35493.
Buschges R., Hollricher K., Panstruga R., Simmons G., Wolter M., et al. (1997) The Barley
Mlo Gene: A novel control element of plant pathogen resistance. Cell 88:695-705.
Catanzariti A.-E., Dodds P.N. and Ellis J.G. (2007) Avirulence proteins from haustoria-
forming pathogens. FEMS Microbiol Lett 269:181-188.
Chester K. S. (1946) The Nature and Prevention of the Cereal Rusts as Exemplified in the Leaf
Rust of Wheat. Waltham, MA:Chronica Botanica.
34

Collins N., Drake J., Ayliffe M., Sun Q., Ellis J., Hulbert S. and Pryor T. (1999) Molecular
characterization of the maize Rp1-D rust resistance haplotype and its mutants. The Plant
Cell 11:1365-1376.
Close T.J., Wing R., Kleinhofs A. and Wise R. (2001) Genetically and physically anchored
EST resources for barley genomics. Barley Genetics Newsletter 31:29-30.
Dodds P.N., Lawrence G.J. and Ellis J.G. (2001) Six amino acid changes confined to the
leucine-rich repeat B-strand/B-turn motif determine the difference between the P and P2
rust resistance specificities in flax. Plant Cell 13:163-178.
Dodds P.N., Lawrence G.J., Catanzariti A.-M., Ayliffe M., and Ellis J. (2004) The
Melampsora lini AvrL567 avirulence genes are expressed in haustoria and their products
are recognized inside plant cells. The Plant Cell 16:755-768.
Dodds P.N., Lawrence G.J., Catanzariti A.-M., Teh T., Ching I., Wang A., et al. (2006)
Direct protein interaction underlies gene-for-gene specificity and co-evolution of the flax
resistance genes and flax rust avirulence genes. Proc Natl Acad Sci USA 103:8888-8893.
Druka A., Kudrna D., Han F., Kilian A., Steffenson B., Frisch D., Tomkins J., Wing R. and
Kleinhofs A. (2000) Physical mapping of barley stem rust resistance gene rpg4. Mol Gen
Genet 264:283-290.
Ellis J.G., Lawrence G.J., Peacock W.J. and Pryor A.J. (1988) Approaches to cloning plant
genes conferring resistance to fungal pathogens. Annu. Rev. Phytopathol. 26:245-263.
Feng Y., Liu Q., and Xue Q. (2006) Comparative study of rice and Arabidopsis actin-
depolymerizing factors gene families. J. Plant Physiol. 163:69-79.
Fetch T., Johnston P.A. and Pickering R. (2009) Chromosomal Location and Inheritance of
Stem rust transferred from Hordeum bulbosum into cultivated barley (H. vulgare).
35

Phytopathology 99:339-343.
Feuillet C., Travella S., Stein N., Albar L., Nublat A. and Keller B. (2003) Map-based
isolation of the leaf rust disease resistance gene Lr10 from hexaploid wheat (Triticum
aestivum L.) genome. Proc Natl Acad Sci USA 100(25):15253-15258.
Fox S. L. and Harder D. E. (1995) Resistance to stem rust in barley and inheritance of
resistance to race QCC. Can. J. Plant. Sci. 75:781-788.
Flor H.H. (1946) Genetics of pathogenicity in Melampsora lini. J. Agric. Res. 73:335-357.
Flor H.H. (1956) The complementary system in flax and flax rust. Adv. Genet. 8:29-54.
Fu D., Uauy C., Distelfeld A., Blechl A., Epstein L., Chen X., Sela H., Fahima T. and
Dubcovsky J. (2009) A kinase-START gene confers temperature-dependent resistance to
wheat stripe rust. Science www.sciencexpress.org/19 February 2009/.
Genre A. and Bonfante P. (1998) Actin versus tubulin configuration in arbuscule-containing
cells from mycrorrizal tobacco roots. New Phytol 140:745-52.
Haldane J.B.S. (1949) Disease and evolution. Ric. Sci. Suppl. A 19:68-76.
Halterman D., Zhou F., Wei F., Wise R.P. and Schulze-Lefert P. (2001) The Mla6 coil-coil,
NBS-LRR protein confers AvrMla6-dependent resistance specificity to Blumeria
graminis f. sp. hordei in barley and wheat. The Plant Journal 25(3):335-348.
Hammond-Kosack K.E. and Jones J.D.G. (1997) Plant disease resistance genes. Annual
Review of Plant Physiology and Plant Molecular Biology 48:573-605.
Han F., Kilian A., Chen J.P., Kudrna D., Steffenson B., Yamamoto K., Matsumoto T.,
Sasaki T. and Kleinhofs A. (1999) Sequence analysis of a rice BAC covering the
syntenous barley Rpg1 region. Genome 42:1071-1076.
Holzberg S.S., Brosio P., Gross C. and Pogue G.P. (2002) Barley stripe Mosaic Virus-induced
36

gene silencing in monocot plant. The Plant Journal 30:315-327.
Horvath H., Rostoks N., Brueggeman R., Steffenson B., von Wettstein D. and Kleinhofs A.
(2003) Genetically engineered stem rust resistance in barley using the Rpg1 gene. Proc.
Natl. Acad. Sci. USA 100:364-369.
Hu G., Richter T.E., Hulbert S.H. and Pryor A.J. (1996) Disease lesion mimicry caused by
mutations at the rust resistance gene Rp1. The Plant Cell 8:1367-1376.
Huang L. and Gill B. S. (2001) An RGA-like marker detects all known Lr21 leaf rust-resistance
gene family members in Aegilops tauschii and wheat. Theor. Appl. Genet. 103:1007-
1013.
Huang L., Brooks S.A., Li W., Fellers J.P., Trick H.N. and Gill B.S. (2003) Map-based
cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread
wheat. Genetics 164:655-664.
Hulbert S.H. (1997) Structure and evolution of the rp1 complex conferring rust resistance in
maize. Annu. Rev. Phytopathol. 35:293-310.
Jedel P.E. (1990) A gene for resistance to Puccinia graminis f. sp. tritici in PI 382313. Barley
Genet. Newsl. 20:43-44.
Jin Y., Steffenson B.J. and Fetch T.G.J. (1994a) Sources of resistance to pathotype QCC of
Puccinia graminis f. sp. tritici in barley. Crop Sci 34:285-8.
Jin Y., Steffenson B.J. and Miller J.D. (1994b) Inheritance of resistance to pathotypes QCC
and MCC of Puccinia graminis f. sp. tritici in barley line Q21861 and temperature effects
on the expression of resistance. Phytopathology 84:452-5.
Johnson R. (1984) A critical analysis of durable resistance. Ann Rev Phytopathol 22:309-330.
Jones J.DG. and Dangl J.L. (2006) The plant immune system. Nature 444:323-329.
37

Kilian A., Chen J., Han F., Steffenson B. and Kleinhofs A. (1997) Towards map-based
cloning of the barley stem rust resistance genes Rpg1 and rpg4 using rice as an
intergenomic cloning vehicle. Plant Molecular Biology 35:187-195.
Kim Y.J., Lin N.C. and Martin G.B. (2002) Two distinct Pseudomonas effector proteins
interact with the Pto kinase and activate plant immunity. Cell 109:589–598.
Kislev M.E. (1982) Stem rust of wheat 3300 years old found in Isreal. Science 216:993-994.
Kleinhofs A, Kilian A, Saghai Maroof M, et al. (1993) A molecular, isozyme and
morphological map of the barley (Hordeum vulgare) genome. Theor. Appl. Genet.
86:705-712.
Krattinger S.G., Lagudah E.S., Spielmeyer W., Singh R.P., Julio H.-E. et al. (2009) A
putative ABC transporter confers durable resistance to multiple fungal pathogens in
wheat. Science www.sciencexpress.org/19 February 2009/.
Lawrence G.J., Finnegan E.J., Ayliffe M.A. and Ellis J.G. (1995) The L6 gene for flax rust
resistance is related to the Arabidopsis bacterial resistance gene RPS2 and the tobacco
viral resistance gene N. Plant Cell 7:1195-1206.
Leonard W.J. (2001) Role of Jak kinases and STATs in cytokine signal transduction. Int. J
Hematol. 73(3):271-277.
Lorang J.M., Sweat T.A. and Wolpert T.J. (2007) Plant disease susceptibility conferred by a
“resistance” gene. Proc. Natl. Acad. Sci. USA 104(37):14861-14866.
Martin G.M., Brommonschenkel S.H., Chunwongse J., Frary A., Ganal M.W., et al. (1993)
Map-based cloning of a protein kinase gene conferring disease resistance in tomato.
Science 262:1432-1436.
Martin G.B, Bogdanove A.J. and Sessa G. (2003) Understanding the function of plant disease
38

resistance proteins. Annu Rev Plant Biol 54:23-61.
Nazari, K., Mafi M., , Yahyaoui M., Singh R. P. and Park R. F. (2009) Detection of Wheat
Stem Rust (Puccinia graminis f. sp. tritici) Race TTKSK (Ug99) in Iran. Plant Dis.
93:317.
Nirmala J., Brueggeman R., Maier C., Clay C., Rostoks N., Kannangara C.G., Wettstein D.
von, Steffenson B. and Kleinhofs A. (2006) Subcellular localization and functions of the
barley stem rust resistance receptor-like serine/threonine-specific protein kinase Rpg1.
Proc. Natl. Acad. Sci. USA 103:7518-7523.
Nirmala J., Dahl S., Steffenson B.J., Kannangara C.G., Wettstein D. von, Chen X. and
Kleinhofs A. (2007) Proteolysis of the barley receptor-like protein kinase RPG1 by a
proteasome pathway is correlated with Rpg1-mediated stem rust resistance. Proc. Natl.
Acad. Sci. USA 104:10276-10281.
Opalski K.S., Schultheiss H., Kogel K.-H. and Huckelhoven R. (2005)The receptor-like MLO
protein and the RAC/ROP family G-protein RACB modulate actin reorganizatio in barley
attacked by the biotrophic powdery mildew fungus Blumeria graminis f. sp. hordei. The
Plant J 41:291-303.
O'Shea J.J. and Leonard W.J. (1998) Jaks and STATs: biological implications. Annu. Rev.
Immunology 16:293-322.
Ouellet F., Carpentier E., Cope J., Monroy A.F. and Sarhan F. (2001) Regulation of a wheat
actin-depolymerizing factor during cold acclimation. Plant Physiol. 125:360-368.
Patterson F.L., Shands R.G., and Dickson J.G. (1957) Temperature and seasonal effects on
seedling reactions of barley varieties to three races of Puccinia graminis f. sp. tritici.
Phytopathology 47:395-402.
39

Peterson P.D. (2001) Stem rust of wheat from enemy to modern foe. St Paul, MN:APS press.
Powers, L. and Hines, L. (1933) Inheritance of reaction to stem rust and barbing awns in barley
crosses. J. Agric. Res. 46:1121-1129.
Pretorius Z.A., Singh R.P., Wagoire W.W. and Payne T. S. (2000) Detection of virulence to
wheat stem rust resistance gene Sr31 in Puccinia graminis. f. sp. tritici in Uganda. Plant
Dis. 84:203.
Roelfs A.P., Long D.L. and Roberts J.J. (1993) Races of Puccinia graminis in the United
States during 1991. Plant Dis 77:129-32.
Roelfs A.P., Casper D.H., Long D.L. and Roberts J.J. (1991) Races of Puccinia graminis in
the United States in 1989. Plant Dis 75:1127-30.
Roelfs A.P. (1978) Estimated losses caused by rust in small grain cereals in the United States
1918-76. U.S. Dept. Agri. Misc. Pub. #1363.
Roelfs A.P. (1982) Effects of barberry eradication on stem rust in the United States. Plant
Disease 66:177-181.
Rostoks N., Steffenson B.J. and Kleinhofs A. (2004) Structure and expression of the barley
stem rust resistance gene Rpg1. Physiology and Molecular Plant Pathology 64:91-101.
Salmeron J., Oldroyd G., Rommens C., Scofield S. and Kim H. (1996) Tomato Prf is a
member of the leucine-rich repeat class of plant disease resistance genes and lies
embedded within the Pto kinase gene cluster. Cell 86:123-133.
Sasaki T. and Burr B. (2000). International Rice Genome Sequencing Project: The effort to
completely sequence the rice genome. Curr. Opin. Plant Biol. 3:138–141.
Schafer J.F. and Roelfs A.P. (1985) Estimated relation between number of urediniospores of
40

Puccinia graminis f. sp. tritici and rates of occurrence of virulence. Phytopathology
75:749-750.
Schulze-Lefert P. and Panstruga R. (2003) Establishment of biotrophy by parasitic fungi and
reprogramming of host cells for disease resistance. Annual Review of Phytopathology
41:641-667.
Singh R.P., Hodson D.P., Huerta-Espino J., Jin Y., Njau P., Wanyera R., et al. (2008) Will
stem rust destroy the world's wheat crop? Advances in Agronomy 98:271-309.
Søgaard, B. and Wettstein-Knowles P. von (1987) Barley: genes and chromosomes. Carlsberg
Res. Commun. 52:123-196.
Smith S.M., Pryor A.J. and Hulbert S.H. (2004) Allelic and haplotypic diversity at the Rp1
rust resistance locus of maize. Genetics 167:1939-1948.
Song W.Y., Wang G.L., Chen L.L., Kim H.S, Pi L.Y. Gardner J. et al. (1995) A receptor
kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 270:1804-
1806.
Steffenson B.J., and Jin Y. (2006) Resistance to race TTKS of Puccinia graminis f. sp. tritici in
barley. Phytopathology 96:S110.
Steffenson B.J. (1992) Analysis of durable resistance to stem rust in barley. Euphytica 63:153-
167.
Steffenson, B.J. (1991) Stem rust of barley: History, calamity, and control. Proc. 28th Barley
Improvement Conference 3-8.
Steffenson B.J., Wilcoxson R.D. and Roelfs A.P. (1984) Inheritance of resistance to Puccinia
graminis f. sp. secalis in barley. Plant Dis. 68:762-763.
Sun X., Cao Y., Yang Z., Xu C., Li X., Wang S., et al. (2004) Xa26, a gene conferring
41

resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-
like protein. The Plant Journal 37:517-527.
Sun Y. and Steffenson B.J. (2005) Reaction of barley seedlings with different stem rust
resistance genes to Puccinia graminis f. sp. tritici and P. g. f. sp. secalis. Can J Plant
Pathol 27:80-89.
Swiderski M.R. and Innes R.W. (2001) The Arabidopsis PBS1 resistance gene encodes a
member of a novel protein kinase subfamily. The Plant Journal 26:101-112.
Ungureanu D., Saharinen P., Juntilla I., Hilton D.J. and Silvennoinen O. (2002) Regulation
of Jak2 through the Ubiquitin-proteasome pathway involves phosphorylation of Jak2 on
Y1007 and interaction with SOCS-1. Molecular and Cellular Biology 22:3316-3326.
Van Der Bizen E.A. and Jones J.D.G. (1998) Plant disease-resistance proteins and the gene-
for-gene concept. Trends Biochem Sci 23:454-456.
Webb C.A., Richter T.E., Collins N.C., Nicolas M., Trick H.N., Pryor T. and Hulbert S.H.
(2003) Genetic and molecular characterizatio of the maize rp3 rust resistance locus.
Genetics 169:381-394.
Wei F., Gobelman-Werner K., Morroll S. M., Kurth J., Mao L., Wing R., Leister D.,
Schulze-Lefert P. and Wise R. P. (1999) The Mla (powdery mildew) resistance cluster
is associated with three NBS-LRR gene families and suppressed recombination within a
240-kb DNA interval on chromosome 5S (1HS) of barley. Genetics 153:1929-1948.
Wolfe K.H., Gouy M., Yang Y.W., Sharp P.M. and Li W.H. (1989) Date of the monocot-
dicot divergence estimated from chloroplast DNA sequence data. Proc Natl Acad Sci
86:6201-6205.
Yu Y., Tomkins J., Waugh R., Frisch D., Kudrna D., Kleinhofs A., Brueggeman R., et al.
42

(2000) A bacterial artificial chromosome library for barley (Hordeum vulgare L.) and
identification of clones containing putative resistance genes. Theor Appl Genet,
101:1093-1099.
Zemede A. (1996) Barley in Ethiopia: the link between botany and tradition. In: Barley
Research in Ethiopia: Past work and Future prospects. Proceedings of the first Barley
Research Review Workshop.
Zhou J., Tang X. and Martin G.B. (1997) The Pto kinase conferring resistance to tomato
bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-
related genes. EMBO J. 16:3207-3218.
Zhou F., Kurth J., Wei F., Elliott C., Vale G., Yashiaoui N., Keller B., Somerville S., Wise
R. and Schulze-Lefert P. (2001) Cell-autonomous expression of barley Mla1 confers
race-specific resistance to the Powdery Mildew fungus via a Rar1-independent signaling
pathway. The Plant Cell 13:337-350.
43

44
CHAPTER ONE
The barley stem rust resistance gene Rpg1 is a novel disease resistance gene
with homology to receptor kinases.
R. Brueggeman1*, N.Rostoks1, D. Kudrna1, A. Kilian2, F. Han3, J. Chen4, A. Druka1, B.
Steffenson5, A. Kleinhofs1,6
1 Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-
6420, USA; 2 Current address: Center for the Application of Molecular Biology to International
Agriculture, GPO Box 3200, Canberra, ACT 2601, Australia; 3 Current address: Pioneer Hi-Bred
Intl. Inc., 7300 NW 62nd Ave., Johnston, IA 50131, USA; 4 Current address: USDA-ARS Plant
Stress and Germplasm Lab., 3810 4th St., Lubbock, TX 79415, USA; 5 Department of Plant
Pathology, 495 Borlaug Hall, 1991 Upper Buford Circle, St. Paul, MN 55108-6030, USA; 6School of Molecular Biosciences, Washington State University, Pullman, WA 99164-4234,
USA
Communicated by Diter von Wettstein
Biological Sciences, Genetics
Corresponding author: Andris Kleinhofs, Dept. Crop and Soil Sciences, Washington State
University, Pullman, WA 99164-6420, FedEx address Johnson Hall 201, Tel: 1-509-335-4389
Fax: 1-509-335-8674, E-mail: [email protected]
As Published In
Proceedings of the National Academy of Sciences (2002) 99(14):9328-9333
* R. Brueggeman is the main contributor

The barley stem rust-resistance gene Rpg1 is a noveldisease-resistance gene with homology toreceptor kinasesR. Brueggeman*, N. Rostoks*, D. Kudrna*, A. Kilian*†, F. Han*‡, J. Chen*§, A. Druka*, B. Steffenson¶,and A. Kleinhofs*�**
*Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6420; ¶Department of Plant Pathology, 495 Borlaug Hall, 1991Upper Buford Circle, St. Paul, MN 55108-6030; and �School of Molecular Biosciences, Washington State University, Pullman, WA 99164-4234
Communicated by Diter von Wettstein, Washington State University, Pullman, WA, May 13, 2002 (received for review March 25, 2002)
Stem rust caused by Puccinia graminis f. sp. tritici was among themost devastating diseases of barley in the northern Great Plains ofthe U.S. and Canada before the deployment of the stem rust-resistance gene Rpg1 in 1942. Since then, Rpg1 has provideddurable protection against stem rust losses in widely grown barleycultivars (cvs.). Extensive efforts to clone Rpg1 by synteny with riceprovided excellent flanking markers but failed to yield the genebecause it does not seem to exist in rice. Here we report themap-based cloning and characterization of Rpg1. A high-resolutiongenetic map constructed with 8,518 gametes and a 330-kb bacterialartificial chromosome contig physical map positioned the genebetween two crossovers �0.21 centimorgan and 110 kb apart. Theregion including Rpg1 was searched for potential candidate genesby sequencing low-copy probes. Two receptor kinase-like geneswere identified. The candidate gene alleles were sequenced fromresistant and susceptible cvs. Only one of the candidate genesshowed a pattern of apparently functional gene structure in theresistant cvs. and defective gene structure in the susceptible cvs.identifying it as the Rpg1 gene. Rpg1 encodes a receptor kinase-likeprotein with two tandem protein kinase domains, a novel structurefor a plant disease-resistance gene. Thus, it may represent a newclass of plant resistance genes.
The barley stem rust-resistance gene Rpg1 confers resistanceto many pathotypes of the stem rust fungus Puccinia graminis
f. sp. tritici. North American barley cultivars (cvs.) with Rpg1were first released in 1942 and since that time there have beenno significant losses as a result of stem rust except for a minorepidemic caused by pathotype QCC in 1989–91 (1). The Rpg1resistance is dominant and considered durable (2), because it hasremained effective for a long time in widely grown cvs. across thestem rust-prone northern Great Plains (1). The earliest knownsource of stem rust resistance in the U.S. was derived from anunimproved cv. imported by the U.S. Department of Agriculturefrom Canton Lucerne, Switzerland, in 1914. Two selections fromthis seed lot became the cvs. Chevron and Peatland. Peatland hasbeen the main source of stem rust resistance in most Canadiancvs. since 1947 (1). A potentially different source of resistancewas discovered during the severe stem rust epidemic of 1935 bySam Lykken, a farmer from Kindred, North Dakota, whoidentified a single healthy plant in a heavily rusted field ofWisconsin 37 barley (3). This plant was saved and gave rise to thecv. Kindred. Kindred and cvs. derived from it have dominatedthe barley hectarage in the northern Great Plains of the U.S.from 1942 to the present (1). Both sources of barley stem rustresistance are the results of the Rpg1 gene (4, 5).
The Rpg1 gene was mapped to the short arm of barleychromosome 1(7H) with morphological and molecular markers(6, 7). Our initial efforts at map-based cloning of the Rpg1 genewere focused on using rice as an intergenomic cloning vehicle (8,9). Despite the excellent synteny between barley chromosome1(7H) short arm and rice chromosome 6 short arm and theexcellent alignment of flanking markers, the Rpg1 gene was not
found in the syntenous position in rice (10). The development ofa barley bacterial artificial chromosome (BAC) library allowedus to initiate a chromosome walk toward Rpg1 (11). A resistancegene analog (RGA), B9 (12), and barley homologues of themaize rust-resistance gene Rp1-D (pic20) (13) were proposed asthe barley stem rust-resistance gene Rpg1. We mapped both B9and the pic20 homologues at high resolution and found that theydid not cosegregate with Rpg1. The pic20 probe, however, dididentify a complex RGA locus near Rpg1 and helped to extendthe BAC contig (14).
Numerous genes that confer resistance to a variety of plantpathogens have been cloned and characterized (15–17). Thegenes that seem to be involved in signal transduction and behavein a gene for gene manner (18) have been classified into fourgroups. The majority resembles intracellular receptors and con-tains a predicted nucleotide binding site�leucine-rich repeat(NBS�LRR) structure. Another group encodes an LRR putativeextracellular receptor and a membrane anchor. Two additionalgroups contain serine�threonine kinase domain without anyobvious ligand-binding domain (i.e., Pto) and receptor kinaseswith LRR extracellular ligand-binding, transmembrane, andserine�threonine kinase domains (i.e., Xa21) (16). A recentlyreported tomato Verticillium wilt-resistance gene Ve seems toencode a new group with cell-surface glycoprotein, signals forreceptor-mediated endocytosis, and leucine zipper or PESTsequences [PEST, region rich in proline (P), glutamate (E),serine (S), and threonine (T); ref. 19].
Here we report map-based cloning and partial characteriza-tion of the barley stem rust-resistance gene Rpg1. The Rpg1 geneidentity was confirmed by high-resolution genetic and physicalmapping, sequencing of multiple alleles from resistant andsusceptible lines, a fortuitous recombination that combined partsfrom the resistant and susceptible parents, and exclusion of otherpotential candidate genes by sequence analysis. The Rpg1 geneencodes a receptor kinase-like protein with two tandem protein
Abbreviations: BAC, bacterial artificial chromosome; cv., cultivar; LRR, leucine-rich repeat;RT, reverse transcription; cM, centimorgan.
Data deposition: The sequences reported in this paper have been deposited in the GenBankdatabase [accession nos. AF509747 (Rpg1 cDNA); AF509748–AF509765 (Rpg1 genomicsequences from barley lines Morex, OSU6, Q21861, 80-TT, Bowman, Chevron, Leger,Kindred, Peatland, Steptoe, Lion, Wisconsin 38, Moravian III, OSU15, SM89010, Dicktoo,Gobernadora, and AMS170, respectively); AF509744–AF509746 (NRG031 genomic se-quence from barley lines Morex, Steptoe, and OSU6, respectively); AF509766–AF509779(genomic sequences from low copy probes isolated from the Rpg1 genomic region NRG019,NRG021, NRG026, NRG027, NRG046, NRG048, NRG050, RSB347, RSB409A, RSB409B,RSB410, RSB414, and RSB416, respectively)].
†Present address: Center for the Application of Molecular Biology to International Agri-culture, GPO Box 3200, Canberra ACT 2601, Australia.
‡Present address: Pioneer Hi-Bred International Inc., 7300 NW 62nd Avenue, Johnston, IA50131.
§Present address: U.S. Department of Agriculture�Agricultural Research Service Plant Stressand Germplasm Lab, 3810 4th Street, Lubbock, TX 79415.
**To whom reprint requests may be addressed. E-mail: [email protected].
9328–9333 � PNAS � July 9, 2002 � vol. 99 � no. 14 www.pnas.org�cgi�doi�10.1073�pnas.142284999
45

kinase domains, which is a novel structure for a plant disease-resistance gene. It does not contain a strong membrane-targetingmotif and known receptor sequences, suggesting a mechanism ofaction similar to the tomato Pto gene (20, 21). The barley Rpg1gene may be useful for durable stem rust-resistance in otherspecies.
Materials and MethodsMaterials. Barley crosses used to produce recombinant gametesare given in Table 1, which is published as supporting informa-tion on the PNAS web site, www.pnas.org. Most of the crosseswere produced by, and for, the North American Barley GenomeProject and are described at http:��www.css.orst.edu�barley�nabgmp�nabgmp.htm. The F2 materials were generated in thisstudy. FN53 is a fast neutron-induced brachytic dwarf (brh1.ae)derived from cv. Steptoe (22). MD2 is a barley genetic stockcarrying multiple dominant morphological markers (23). Resis-tant (24) and susceptible (25) barley lines used are described insupporting information on the PNAS web site. Libraries usedinclude the cv. Morex BAC library (11) and cDNA libraries madefrom various tissues and with different biotic and abiotic stresses(26) (http:��wheat.pw.usda.gov�ggpages�bgn�31�close.htm).
Molecular Marker Development. Low-copy probes suitable forgenetic mapping and hybridization-based BAC library searchingwere identified from small insert libraries developed fromindividual BAC clones. The BAC clone DNA was digested withfour- or six-base recognition restriction enzymes and subclonedinto a plasmid vector. The subclones were hybridized with32P-labeled barley total genomic DNA probe. Subclones that didnot show any or very little hybridization signal were selected andtested further by hybridization to Southern blots of HindIII-digested barley genomic DNA. The clones generated with six-base recognition enzymes were further digested with severalrestriction enzymes to develop small inserts. The digested DNAwas electrophoresed, blotted, and transferred to filters forhybridization with barley total genomic DNA probe (27). DNAfragments that failed to hybridize were selected as low-copyprobes.
Genetic Mapping. Recombinants between the Rpg1 f lankingmarkers ABG704 and ABG077, derived from 8,518 gametes(Table 1), were used for high-resolution mapping. The geneticdistances are expressed in number of crossovers. The stem rustphenotype of progeny was determined as described and mappedin reference to the molecular markers (7).
BAC Contig Development. BAC clones were isolated from the 6.3Xcv. Morex BAC library (11) by hybridizing the arrayed BACcolony filters with 32P-labeled probes. BAC DNA was isolated byusing the standard alkaline lysis procedure (28). Low-copyprobes were restriction-enzyme mapped on the most-extendingBAC clone, and the most-extending probes were hybridized tothe BAC library to identify further extending BAC clones. Theprocess was repeated as needed to complete the BAC contig.
Sequencing and Sequence Analysis. DNA was sequenced with theBigDye terminator system on ABI Prizm 377 DNA sequencer(Applied Biosystems) at the Bioanalytical Center, WashingtonState University, Pullman. The region delimited by the cross-overs flanking Rpg1 (Fig. 1) was screened for putative genes bysequencing low-copy clones. Small insert sublibraries were de-veloped with four- and six-base restriction endonucleases essen-tially as described above for low-copy probe development exceptthat more restriction enzymes were used and higher coveragewas achieved. All of the low-copy clones were sequenced andanalyzed with the BLASTX and BLASTN algorithms (29). Se-quences without significant matches in BLAST searches were
screened for ORFs and putative genes with the gene predictionsoftware GENEMARK.HMM (http:��opal.biology.gatech.edu�GeneMark�eukhmm.cgi).
Genomic DNA was isolated with a modified CTAB method(30), and specific fragments were amplified by PCR. PCRreactions of 50 �l contained 100 ng of genomic DNA, 0.2 mMof each dNTP, 25 pmols of each primer, 2.5 �l of RedTaq DNApolymerase (Sigma), and 5 �l of 10� RedTaq reaction buffer.Amplification was performed in a PTC-100 programmablethermal controller (MJ Research, Cambridge, MA) at 95°C for4 min; 35 cycles of 95°C for 1 min, 62°C for 1 min, and 72°C for1 min; followed by 7 min at 72°C. The PCR fragments wereelectrophoresed on 1% agarose gels and eluted from the agarose.The PCR fragments were sequenced with the same forward andreverse primers used to amplify the fragments. Primers weredesigned from the Morex sequence to amplify about 1-kboverlapping fragments that covered the entire gene (Table 2,which is published as supporting information on the PNAS website). Internal primers were designed to sequence across gaps andlow-quality sequence. DNA fragments including sequencechanges critical to the interpretation of the results were rese-quenced with different DNA preparations and different primerpairs. Sequences were assembled with the GENETIC COMPUTERGROUP program (31) supported by the Visualization, Analysis,and Design in the Molecular Sciences center at Washington StateUniversity.
RNA Isolation and Reverse Transcription (RT)-PCR. Total leaf RNAfrom 21-day-old Morex, Steptoe, and ASM170 plants was iso-lated with the Totally RNA kit (Ambion, Austin, TX). Integrityand quantity of the RNA samples were checked by formaldehydedenaturing agarose gel electrophoresis. Approximately 1 �g ofthe total RNA was used for RT-PCR reactions with SuperScriptOne-Step RT-PCR system (Invitrogen) under the manufactur-er’s recommended standard conditions. RT reactions wereprimed with gene-specific primers. Northern blots were done asrecommended by Amersham Pharmacia Biotech (http:��www.apbiotech.com�technical�documentation�dna�protein�tt159.htm).
ResultsHigh Resolution Genetic Mapping of the Rpg1 Gene. The Rpg1 locuswas previously mapped to the telomeric end of barley chromo-some 1(7H)S (9, 10). A high-resolution genetic map, based on8,518 gametes (Table 1), was developed with recombinants from
Fig. 1. A genetic and physical map of barley stem rust-resistance gene Rpg1region on chromosome 1(7H). (A) high-resolution genetic map based on 8,518gametes showing critical markers and the number of total crossovers (c.o.)separating them. (B) Physical map encompassing Rpg1 based on BAC clonecontig. The distance from Pic20A to RSB308 was estimated to be �270 kb.
Brueggeman et al. PNAS � July 9, 2002 � vol. 99 � no. 14 � 9329
GEN
ETIC
S

the region between the Rpg1 f lanking markers ABG704 andABG077. The interval between MWG035S40 (BB172#13) andthe barley homologue of the maize Rp1-D gene (pic20A) delim-ited the Rpg1 region, contained 17 recombinants, and repre-sented a genetic distance of �0.21 centimorgan (cM) (Fig. 1).
Physical Mapping of the Rpg1 Region. A BAC clone contig wasestablished for the Rpg1 region by chromosome walks frompic20A and 19j10LRR loci (Fig. 1). The probe 19j10LRR wasobtained from L. D. Holappa (M. K. Walker-Simmons Labora-tory, Pullman, WA). A BLASTX search revealed it to be ‘‘similarto a non-LTR [long terminal repeat] retroelement reversetranscriptase’’ (S � 170, 3e-41). Hybridization to barley poly-morphism filters revealed many bands, some polymorphic, whichwere mapped. A 2.1-kb band, generated with the restrictionenzyme NcoI, mapped one crossover distal to Rpg1 and coseg-regated with MWG035S40 and other markers. The probe wasused to isolate �2,000 BAC clones from the cv. Morex library.A BAC clone, 022c13, containing the mapped band was iden-tified after digestion with NcoI and hybridization with19j10LRR. A chromosome walk was initiated from this BACclone and led to an overlap with BAC clones isolated from thepic20A walk.
Identification of Candidate Genes. Three putative candidate geneswere identified by draft sequencing in the region between thetwo closest Rpg1 f lanking crossovers (Fig. 1). Two were receptorkinase-like genes and one was a truncated phophoenolpyruvatekinase-like gene that was not considered further. The firstcandidate gene was a 5-kb HindIII low-copy clone designatedRSB228. This clone, when used as a probe, was polymorphic inall crosses, and in DraI restriction enzyme-digested Morex andSteptoe genomic DNA yielded three and two polymorphicbands, respectively (Fig. 4, which is published as supportinginformation on the PNAS web site). All bands cosegregated inall recombinants except one, AMS170, which contained oneMorex and one Steptoe band and was susceptible to stem rust(rpg1). Sequence analysis of RSB228 revealed that it containedORFs with high amino acid homology to several receptorkinase-like proteins from Arabidopsis and the SRK receptorkinase-like gene family. Thus, this gene was a strong candidateto be Rpg1, and subsequent sequence analysis of stem rust-resistant (Rpg1) and susceptible (rpg1) cv. alleles confirmed thisconclusion (data presented below). A second receptor kinase-like gene, NRG031, was identified and sequenced from Morex,OSU6 (both resistant), and Steptoe (susceptible). Alleles from
resistant and susceptible lines were very similar, excludingNRG031 from Rpg1 candidacy.
The candidate genes were identified by sequencing of theapproximately 35 kb immediately distal to Rpg1. This sequencedregion consisted primarily of long terminal repeat retrotrans-posons that are a common feature of the intergenic space inbarley (32) and in other grass species, such as maize (33). For theabout 70 kb remaining, sublibraries were constructed with the4bp recognition endonucleases Sau3AI and TaqI and the 6-bprecognition restriction endonucleases HindIII and EagI. Approx-imately 18� coverage of this region was screened for low-copyclones, sequenced, and analyzed with the BLASTX and BLASTNalgorithms. Sequences with putative ORFs were hybridized toSouthern blots of HindIII-digested genomic DNA. Putativegenes were completely sequenced from cv. Morex BAC DNA. Intotal, 80 kb of nonredundant putatively low-copy sequence wasgenerated and examined with BLAST searches as well as with thegene prediction software GENEMARK.HMM. No other putativegenes were identified.
Characterization of the Candidate Genes. The Morex (resistant)allele of candidate gene 1 (RSB228) seemed to encode afunctional gene. The Steptoe (susceptible) allele contained40-bp differences, three of which introduced stop codons in the4th, 8th, and 13th exons, indicating a defective gene. TheSteptoe � Morex recombinant AMS170 allele had a crossoverwithin the gene in the 436–524-aa region including parts of exons7 and 8. The distal part of the AMS170 allele contained the twostop codons in the 8th and 13th exons from Steptoe, explainingthe stem rust-susceptible phenotype (rpg1). The other part of theAMS170 gene had the Morex haplotype and eliminated allproximal genes from Rpg1 candidacy (Fig. 1).
The genomic and cDNA sequence comparisons predicted thegene to contain 14 exons in a total sequence of 4,466 bp coding foran 837-aa (94.5 kDa) protein (Fig. 2). A BLASTP search of thenonredundant protein database showed the best homology tonumerous hypothetical receptor kinase-like proteins. The best hit tocharacterized function proteins was to the SRK family of self-incompatibility S-receptor kinases. The highest homology was tothe kinase domain regions. The N-terminal part of the proteinsequence exhibited low homology to several predicted Arabidopsisproteins with unknown function. A BLASTN search with the Rpg1coding sequence of the nonredundant database revealed no signif-icant homologies. The deduced protein sequence contained twotandem pfam00069 protein kinase domains. Program SIGNALPVERSION 2.0 (http:��www.cbs.dtu.dk�services�SignalP�) indicatedthat the predicted protein sequence does not contain a signal
Fig. 2. Rpg1 gene, cDNA, and translated protein structure. (A) Genomic and cDNA structures with introns (gray) and exons (black) are shown to scale. The mRNA5� and 3� untranslated regions are hatched. Exons are numbered above the drawing. Predicted TATA box, translation start site, and stop codon are shown below.The Rpg1 mRNA structure is shown below the gene. The mRNA was reconstructed with the sequence of 5� truncated cDNA clone HVSMEf55g12 and sequencesof the RT-PCR products from cv. Morex leaf total RNA. (B) Organization of the protein domains is shown to scale with predicted boundaries indicated above.pkinases 1 and 2 denote two tandem protein kinase domains. The asterisk indicates an asparagine residue within a putative N-glycosylation site NFSE.
9330 � www.pnas.org�cgi�doi�10.1073�pnas.142284999 Brueggeman et al.
47

peptide. The PREDICTPROTEIN tool at the European BioinformaticsInstitute (http:��www.ebi.ac.uk��rost�predictprotein) identified aputative transmembrane domain from amino acid 321 to 331,although with low probability. Alternatively, the TMPRED programat http:��www.ch.embnet.org�software�TMPRED�form.htmlfound 3 putative transmembrane domains, amino acids 23–47,amino acids 316–334, and amino acids 626–643, although only the316–334-aa domain was predicted with a score higher than thresh-old. Future work will be required to determine whether thepredicted RPG1 protein is membrane-bound or intracellular.
Candidate gene 2 (NRG031) was homologous to putativereceptor kinase-like genes. Eukaryotic GENEMARK.HMM version2.2a (http:��opal.biology.gatech.edu�GeneMark�eukhmm.cgi)gene prediction program predicted 2 exons in a total sequenceof 981 bp coding for a 327-aa protein. Sequence analysis of allelesfrom resistant (Morex and OSU6) and susceptible (Steptoe)lines showed differences at only two amino acid positions amongthe three alleles sequenced. Morex and Steptoe had isoleucine,whereas OSU6 had threonine in position 4. At position 241,OSU6 and Steptoe had glycine, but Morex had alanine. Becauseall amino acid differences found in the susceptible allele werealso present in one or the other of the resistant alleles, this genewas removed from further candidacy.
Analysis of Rpg1 cDNA. Hybridization of the RSB228 probe to 30arrayed cDNA filters representing 10 different tissue librariesand �500,000 clones identified four cDNA clones. Sequenceanalysis revealed three different expressed genes homologous toRpg1. One partial cDNA clone, HVSMEf 055g12, was identicalto the genomic sequence, whereas another came from a verysimilar member of the same gene family (ABC1037) locatedproximal to Rpg1 and excluded from candidacy by geneticmapping. The third clone came from a more distantly relatedmember of the Rpg1 gene family and mapped to chromosome7(5H). The HVSMEf 055g12 cDNA was used to predict the 3�end of the Rpg1 gene and intron–exon structure for exons 9–14.The homologous, nearly full-length cDNA ABC1037 was used topredict a preliminary intron–exon structure of the 5� part of theRpg1 gene. Sequences of RT-PCR products generated from cv.Morex leaf total RNA with primers corresponding to predictedexons allowed for the construction of apparently the completecoding sequence of the Rpg1 gene. Even though the region fromexon 1 to 3 forms an uninterrupted ORF, RT-PCR productsindicated the presence of two introns. Translation of the recon-structed cDNA sequence suggested that the coding sequencewas complete, because an in-frame stop codon was detectedfour codons upstream of the first predicted methionine. Thegene prediction programs GENSCAN (http:��genes.mit.edu�GENSCAN.html) and FGENESH (http:��www.softberry.com), aswell as NEURAL NETWORK PROMOTER PREDICTION (http:��www.fruitf ly.org�seq�tools�promoter.html) localized the puta-tive transcription start site of the gene about 400 bp upstream ofthe translation start site.
Northern blots of Morex, Steptoe, and ASM170 total orpoly(A) RNA failed to yield positive bands with Rpg1 probealthough positive results were obtained with an actin probe (datanot shown). This finding suggested that the gene was expressedat low levels.
Comparison of Rpg1 Alleles from Resistant and Susceptible BarleyLines. Sequence and RFLP analysis of 19 susceptible and 9resistant barley lines confirmed that the RSB228 probe was theRpg1 gene. The Morex gene sequence was compared withresistant cv. alleles from Kindred, Chevron, Peatland, Q21861,Leger, Bowman, 80-TT-29, and OSU6. All sequences, exceptOSU6, were identical, suggesting that the candidate gene wasfunctional and probably from the same source (see Discussion).The resistant Hordeum vulgare subsp. spontaneum line OSU6
allele had a 3-bp insertion that resulted in a serine to arginineconversion and an insertion of a phenylalanine in the 6th exonat amino acid position 320 (Fig. 3 and Fig. 5, which are publishedas supporting information on the PNAS web site). This was theonly nucleic acid difference between OSU6 and Morex thatresulted in an amino acid difference. Other transitions occurredin the 7th exon and 8th and 13th introns, and a 30-bp deletionwas observed in the 13th intron. Thus, the Rpg1 gene seemed tobe very highly conserved and functional in all resistant linesanalyzed.
In addition to Steptoe and AMS170 discussed above, thealleles from susceptible barley lines Lion, MoravianIII, Wiscon-son38, SM89010, Dicktoo, Gobernadora, and OSU15 weresequenced. The susceptible allele sequences formed threegroups, all seeming to encode defective proteins (Figs. 3 and 5).Group I cvs. Lion, MoravianIII, Steptoe, and Wisconsin38alleles, compared with Morex, had 42 bp differences translatinginto three stop codons and 8-aa changes. The 5� end, includingall of the first exon (6 bp), the first intron, and the first 38 bp ofexon 2, of the alleles sequenced from Group II cvs. Dicktoo,Gobernadora, and SM89010 were very similar among themselvesbut were not homologous to the Morex allele. The sequence afterthis point to amino acid 160 is still quite diverged with several bpsubstitutions resulting in amino acid differences and a 96-bpdeletion in exon 3. These differences indicated that the genewould not be functional. The allele from OSU15, the onlyrepresentative of Group III, was very similar to the OSU6sequence. However, it contained a single bp deletion in the 7thexon resulting in a frame shift that introduced a stop codon atamino acid position 358 and a predicted truncated protein.
Other susceptible barley lines (Baronesse, Betzes, GoldenPromise, Harrington, Klages, Oderbrucker, OWB-D, OWB-R,and 80-tt-30) were analyzed and failed to hybridize with theRSB228 probe. PCR amplification, with the primers describedpreviously, also failed. These lines seemed to have a highlydiverged or deleted Rpg1 gene.
Synteny with Rice and Other Cereals. Hybridization of the RSB228probe with rice cvs. Nipponbare and M201 genomic DNA was
Fig. 3. A diagram representation of the derived amino acid sequencealignment of Rpg1 alleles from resistant (Rpg1) and susceptible (rpg1) cvs. Thedifferences from the cv. Morex sequence are indicated. “Morex” includesadditional resistant cultivars Q21861, Kindred, Chevron, Peatland, Leger,Bowman, and 80-TT-29. “OSU6” is a resistant H. vulgare subsp. spontaneumline PBI004-7-0-015 (accession no. 8321). Group 1, Sequence from susceptiblecultivars Steptoe, Lion, Wisconsin38, and MoravianIII. Group 2, Sequencesfrom susceptible cultivars SM89010, Dicktoo, and Gobernadora. “OSU15” is asusceptible H. vulgare subsp. spontaneum line Wadi Qilt 23–38. AMS170 is adoubled haploid line with a recombination in the Rpg1 gene. *, A 3 bpinsertion that introduces a serine to arginine conversion and inserts a phe-nylalanine at amino acid positions 320 and 321. §, Base pair substitutions ingroup 1 cultivars which converts amino acid codons to stop codons at aminoacid positions 192, 524, and 788. ¶, A single base pair substitution results inlysine to glutamine conversion. **, Highly diverged with multiple amino acidsubstitutions, a small deletion, and a large deletion. §§, A single base pairdeletion introduces a frame shift that results in a stop codon. ¶¶, Crossoverregion in the recombinant AMS 170 was narrowed to a 295-bp region betweenthe lysine at amino acid position 436 of Morex and the stop codon at aminoacid position 524 of Steptoe.
Brueggeman et al. PNAS � July 9, 2002 � vol. 99 � no. 14 � 9331
GEN
ETIC
S

negative. Searching two rice cv. Nipponbare BAC librariesrepresenting 11X and 15X genomes yielded no positive clones,suggesting there are no homologues detectable by hybridizationpresent in rice. A BLASTN search of the Chinese rice genomedatabase (http:��210.83.138.53�rice�) representing the rice sub-species indica genome sequence also failed to uncover homol-ogous regions. A TBLASTX search of the ‘‘other’’ expressedsequence tag (EST) database yielded positive hits with wheat inaddition to barley. Other than within the Triticeae, fair homol-ogy was observed with ESTs from Sorghum propinquum (S �179, 4e-46) and Sorghum bicolor (S � 93, 9e-45), Medicagotrancatula (S � 80, 3e-40), Lycopersicon esculentum (S � 75,4e-37), and Zea mays (S � 99, 3e-35).
DiscussionA candidate gene for the barley stem rust-resistance gene Rpg1was identified by map-based cloning. The Rpg1 gene identity wasconfirmed by high-resolution genetic and physical mapping,sequencing of multiple alleles from resistant and susceptiblelines, a fortuitous recombination that combined parts from theresistant and susceptible parents, and exclusion of other poten-tial candidate genes by sequence analysis. Conceptual translationof the sequence showed amino acid homology to the S receptorkinase gene SRK protein family. SRK (S locus receptor kinase)encodes a plasma membrane-spanning receptor serine�threonine kinase specific to the stigma epidermis (34, 35). It ispresumed to be the determinant of self-incompatibility specific-ity in the stigma (36). Another interesting homology was to themaize kinase interacting kinase KIK1 gene predicted to encodea receptor kinase-like protein with an N-terminal signal peptide,an extracellular ligand-binding domain, a membrane-spanningdomain, and a cytoplasmic serine�threonine protein kinasedomain (37). The KIK1 protein was suggested to transfer a signaldownstream by means of a kinase-associated protein phospha-tase (KAPP) (37). Another protein with homology to the Rpg1gene product was the GA-induced LRR receptor kinase-likeprotein OsTMK from rice, which was also suggested to interactwith a KAPP protein (38).
Receptor kinase-like gene products are defined by the pres-ence of a potential N-terminal hydrophobic signal peptide, anN-terminal domain with a transmembrane region, and a C-terminal intracellular protein kinase domain (39, 40). The twolargest subclasses of plant receptor kinases are those whoseextracellular domain contains LRRs or similarity to the S-locusglycoprotein. Some extracellular domains of plant receptorkinases are unique. The cloned rice Xa21 gene (41), whichconfers resistance to Xanthomonas oryzae pv. oryzae and belongsto a receptor kinase-like gene family, showed homology to Rpg1in the kinase domain. However, the Xa21 gene contains the LRRextracellular domain whereas the Rpg1 gene has an N-terminaldomain that does not resemble any previously described recep-tor. In that sense, Rpg1 might be more similar to the tomato Ptogene (20), which confers resistance to Pseudomonas syringae pv.tomato. The Pto gene, also showing similarity to Rpg1 in thekinase domain, is a serine�threonine kinase without an identi-fiable ligand-binding domain (21). However, the Pto gene prod-uct and the corresponding avirulence gene product of thepathogen, avrPto, have been shown to interact directly (42). ThePto threonine 204 residue implicated in interaction with avrPto(43) is conserved in both kinase domains of the RPG1 protein(positions 302 and 612) as it is in a number of other kinasesincluding Xa21, KIK1, SRK, and OsTMK. Thus, the significanceof this observation is not clear.
The Rpg1 gene product is unique because it contains twotandem protein kinase domains. This finding has not beenpreviously reported in cloned plant pathogen-resistance genesand seems to be uncommon. Some Arabidopsis-predicted genesencode two kinase domains, e.g., AC007234�4 and AC069159�10.
Examples of characterized genes encoding tandem kinase do-mains are the mammalian Janus kinases that, in addition to twokinase domains, also have plasma membrane-binding and SH2domains (44). In the case of the mammalian ribosomal proteinS6 kinase isoforms, which are involved in activation of mitogen-activated kinase cascade, both of the kinase domains are cata-lytically active although they exhibit different phosphotransfer-ase activities (45). Analysis of the protein kinase catalyticaldomains from 65 different kinases found 9 invariant amino acidresidues (46). The most conserved amino acid residues found inthe core of the kinase catalytic domain were also conserved inRPG1 kinase domain 2 but not in kinase domain 1, indicatingthat this domain may not be active.
The origin of the Rpg1 gene bred into barley cvs. in the U.S.and Canada can be traced to two apparently different sources,one an introduction of an unimproved line from Switzerland,which yielded the resistant cvs. Chevron and Peatland, andanother from a single resistant-plant selection made by a farmerfrom Wisconsin 37 barley that became the cv. Kindred. Oursequencing results suggested that both sources of Rpg1 are thesame. Wisconsin 37 seed was no longer available, but analysis ofWisconsin 38, a sister line to Wisconsin 37, and the parentsOderbrucker (Wisconsin Pedigree 5) and Lion showed that evena rare recombination could not account for the resistant allelein Kindred, because Oderbrucker had an apparent deletion andLion a susceptible allele nearly identical to the Steptoe allele.These results suggested that the resistant plant identified in thefarmer’s field of Wisconsin 37 was most probably the result of anadmixture derived from Peatland or Chevron.
Other possible different sources of resistance were from thecv. Q21861 and the H. vulgare subsp. spontaneum line OSU6. Thepedigree of Q21861 is not known, but the allele sequence wasidentical to Morex and thus, probably from the same source. TheOSU6 line probably represented a unique source of the resis-tance gene, but still only had minor nucleotide substitutions anda 3-bp insertion. Interestingly, the 3-bp insertion is also seen inall of the susceptible alleles sequenced, suggesting that they arosefrom this type of resistance gene rather than from the one thatgave rise to the Morex-type gene. Thus, the Rpg1 gene in allresistant lines seemed to be highly conserved. On the other hand,the rpg1 alleles from susceptible cvs. varied from highly con-served to highly diverged or even absent. Thus, the gene does notseem to be essential in absence of the disease.
Physical and genetic mapping of the Rpg1 region indicated aregion of high recombination proximal to the Rpg1 gene and aregion of low recombination distal to the gene (Fig. 1). Approx-imate calculations suggested that the region between markerspic20A and ABC1037 had a physical�genetic distance ratio ofapproximately 350 kb�cM similar to that calculated for theadjoining proximal pic20 region (14). The physical�geneticdistance ratio for the ABC1037 to 19j10LRR region was �4Mb�cM, the predicted barley genome average (30). The regiondefined by the markers MWG035S40, BB172#13, and19j10LRR showed even higher physical�genetic distance ratio,because these three probes cosegregated in the 8,518 gametepopulation but did not form overlapping BAC clone contigs.Thus, the physical�genetic ratio cannot be calculated but must bein excess of 25 Mb�cM based on 1 recombinant and a minimumphysical distance of 300 kb. Similar divergent ratios from 0.18 to5 Mb�cM of physical to genetic distance were observed in theMla cluster on chromosome 5(1H) (47).
We have suggested that map-based cloning in large grassgenomes such as barley could be facilitated by using rice as anintergenomic cloning vehicle (9). Despite the excellent genecolinearity with the Rpg1 f lanking markers, we did not find acandidate gene in rice (10). Screening of two rice cv. Nippon-bare BAC libraries representing 11X and 15X genome cover-age failed to find positive clones. A BLASTN search of the
9332 � www.pnas.org�cgi�doi�10.1073�pnas.142284999 Brueggeman et al.
49

Chinese rice subspecies indica genome database was alsonegative. These results suggest that a homologous gene de-tectable by Southern hybridization does not exist in the ricegenomes tested. Expressed sequence tags with good homologyto the Rpg1 coding sequence were found in wheat and with fairhomology in sorghum, maize, barrel medic, and tomato. Thesedata suggest that Rpg1 homologues may have a function inother species.
In summary, we cloned the barley stem rust-resistance geneRpg1 by map-based cloning. The gene seems to encode areceptor kinase-like protein with two tandem kinase domainsbut no recognizable receptor and membrane anchor domains.Tandem arrangement of kinase domains is a novel feature ofplant disease-resistance genes. Structural similarity to tomatoPto protein supports the hypothesis that the RPG1 protein could
potentially recognize the avirulence gene product of the Pucciniagraminis f. sp. tritici pathotype MCC and function in a signaltransduction pathway.
Note. Preliminary evidence indicates that the Rpg1 genomic fragmenttransformed into the susceptible cv. Golden Promise confers resistanceto the barley stem rust pathotype MCC. Data were based on the analysisof 12 different transgenic plant families, 10 of which produced resistantprogeny.
We are grateful to Tom Fetch, Jr., Yongliang Sun, and Chris Kavanaughfor technical assistance in determining the phenotypes for the diseasereaction. This is Scientific Paper No. 0205-01 from the College ofAgriculture and Home Economics Research Center, Washington StateUniversity, Project 0196. Research was supported by U.S. Department ofAgriculture National Research Initiative Grant 9901325.
1. Steffenson, B. J. (1992) Euphytica 63, 153–167.2. Johnson, R. (1984) Annu. Rev. Phytopathol. 22, 309–330.3. Lejeune, A. J. (1951) in Barley Improv. Conf. Rpt., ed. Parker, J. H. (Midwest
Barley Improvement Association, Minneapolis).4. Powers, L. & Hines, L. (1933) J. Agric. Res. 46, 1121–1129.5. Miller, J. D. & Lambert, J. W. (1955) Agron. J. 47, 373–377.6. Jin, Y., Steffenson, B. J. & Franckowiak, J. D. (1993) Crop Sci. 33, 642–643.7. Kilian, A., Steffenson, B. J., Maroof, M. A. S. & Kleinhofs, A. (1994) Mol.
Plant–Microbe Interact. 7, 298–301.8. Kilian, A., Kudrna, D. A., Kleinhofs, A., Yano, M., Kurata, N., Steffenson, B.
& Sasaki, T. (1995) Nucleic Acids Res. 23, 2729–2733.9. Kilian, A., Chen, J., Han, F., Steffenson, B. & Kleinhofs, A. (1997) Plant Mol.
Biol. 35, 187–195.10. Han, F., Kilian, A., Chen, J. P., Kudrna, D., Steffenson, B., Yamamoto, K.,
Matsumoto, T., Sasaki, T. & Kleinhofs, A. (1999) Genome 42, 1071–1076.11. Yu, Y., Tomkins, J. P., Waugh, R., Frisch, D. R., Kudrna, D., Kleinhofs, A.,
Brueggeman, R. S., Muehlbauer, G. J., Wise, R. P. & Wing, R. A. (2000) Theor.Appl. Genet. 101, 1093–1099.
12. Leister, D., Kurth, J., Laurie, D. A., Yano, M., Sasaki, T., Devos, K., Graner,A. & Schulze-Lefert, P. (1998) Proc. Natl. Acad. Sci. USA 95, 370–375.
13. Ayliffe, M. A., Collins, N. C., Ellis, J. G. & Pryor, A. (2000) Theor. Appl. Genet.100, 1144–1154.
14. Rostoks, N., Zale, J. M., Soule, J., Brueggeman, R., Druka, A., Kudrna, D.,Steffenson, B. & Kleinhofs, A. (2002) Theor. Appl. Genet., 10.1007�s00122-002-0902-8.
15. Hammond-Kosack, K. E. & Jones, J. D. G. (1997) Annu. Rev. Plant Physiol.Plant Mol. Biol. 48, 575–607.
16. Richter, T. E. & Ronald, P. C. (2000) Plant Mol. Biol. 42, 195–204.17. Hulbert, S. H., Webb, C. A., Smith, S. M. & Sun, Q. (2001) Annu. Rev.
Phytopathol. 39, 285–312.18. Flor, H. H. (1947) J. Agric. Res. 74, 241–262.19. Kawchuck, L. M., Hachey, J., Lynch, D. R., Kulcsar, F., van Rooijen, G.,
Waterer, D. R., Robertson, A., Kokko, E., Byers, R., Howard, R. J., et al. (2001)Proc. Natl. Acad. Sci. USA 98, 6511–6515.
20. Martin, G. M., Brommonschenkel, S. H., Chunwongse, J., Frary, A., Ganal,M. W., Spivey, R., Wu, T., Earle, E. D. & Tanksley, S. D. (1993) Science 262,1432–1436.
21. Sessa, G. & Martin, G. B. (2000) Microbes Infect. 2, 1591–1597.22. Franckowiak, J. D. (1995) Barley Genet. Newslett. 24, 56–59.23. Wolfe, R. I. & Franckowiak, J. D. (1990) Barley Genet. Newslett. 20, 117–121.24. Steffenson, B. J., Miller, J. D. & Jin, Y. (1993) Plant Dis. 77, 626–629.
25. Vivar, H. (1987) Rachis 6, 17–19.26. Close, T. J., Wing, R., Kleinhofs, A. & Wise, R. (2001) Barley Genet. Newslett.
31, 29–30.27. Sambrook, J., Fritsch, E. F. & Manniatis, T. (1989) Molecular Cloning: A
Laboratory Manual (Cold Spring Harbor Lab. Press, Plainview, NY).28. Birnboim, H. & Doly, J. D. (1979) Nucleic Acids Res. 7, 1513–1523.29. Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J. H., Zhang, Z., Miller,
W. & Lipman, D. J. (1997) Nucleic Acids Res. 25, 3389–3402.30. Kleinhofs, A., Kilian, A., Maroof, M. A. S., Biyashev, R. M., Hayes, P., Chen,
F. Q., Lapitan, N., Fenwick, A., Blake, T. K., Kanazin, V., et al. (1993) Theor.Appl. Genet. 86, 705–712.
31. Devereux, J., Haeberli, P. & Smithies, O. (1984) Nucleic Acids Res. 12, 387–395.32. Rostoks, N., Park, Y.-J., Ramakrishna, W., Ma, J., Druka, A., Shiloff, B. A.,
SanMiguel, P. J., Jiang, Z., Brueggeman, R., Sandhu, D., et al. (2002) Funct.Integr. Genomics 2, 51–59, 10.1007�s10142-002-0055-5.
33. SanMiguel, P., Tikhonov, A., Jin, Y.-K., Motchoulskaia, N., Zakharov, D.,Melake-Berhan, A., Springer, P. S., Edwards, K. J., Lee, M., Avramova, Z. &Bennetzen, J. L. (1996) Science 274, 765–768.
34. Stein, J. C., Howlett, B., Boyes, D. C., Nasrallah, M. E. & Nasrallah, J. B. (1991)Proc. Natl. Acad. Sci. USA 88, 8816–8820.
35. Stein, J. C., Dixit, R., Nasrallah, M. E. & Nasrallah, J. B. (1996) Plant Cell 8,429–445.
36. Takasaki, T., Hatakeyama, K., Suzuki, G., Watanabe, M., Isogai, A. & Hinata,K. (2000) Nature (London) 403, 913–916.
37. Braun, D. M., Stone, J. M. & Walker, J. C. (1997) Plant J. 12, 83–95.38. van der Knaap, E., Song, W.-Y., Ruan, D.-L., Sauter, M., Ronald, P. C. &
Kende, H. (1999) Plant Physiol. 120, 559–569.39. Walker, J. C. (1994) Plant Mol. Biol. 26, 1599–1609.40. Torji, K. U. (2000) Curr. Opin. Plant. Biol. 3, 361–367.41. Song, W.-Y., Wang, G.-L., Chen, L.-L., Kim, H.-S., Pi, L.-Y., Gardner, J.,
Holsten, T., Wang, B., Zhai, W.-X., Zhu, L.-H., et al. (1995) Science 270,1804–1806.
42. Scofield, S. R., Tobias, C. M., Rathjen, J. P., Chang, J. H., Lavelle, D. T.,Michelmore, R. W. & Staskawicz, B. J. (1996) Science 274, 767–768.
43. Frederick, R. D., Thilmony, R. L., Sessa, G. & Martin, G. B. (1998) Mol. Cell2, 241–245.
44. Rane, S. G. & Reddy, E. P. (1994) Oncogene 9, 2415–2423.45. Fisher, T. L. & Blenis, J. (1996) Mol. Cell. Biol. 16, 1212–1219.46. Hanks, S. K., Quinn, A. M. & Hunter, T. (1988) Science 241, 42–52.47. Wei, F., Gobelman-Werner, K., Morroll, S. M., Kurth, J., Mao, L., Wing, R.,
Leister, D., Schulze-Lefert, P. & Wise, R. P. (1999) Genetics 153, 1929–1948.
Brueggeman et al. PNAS � July 9, 2002 � vol. 99 � no. 14 � 9333
GEN
ETIC
S
50

CHAPTER TWO The barley serine/threonine kinase gene Rpg1 providing resistance to stem rust belongs to a
gene family with five other members encoding kinase domains
R. Brueggeman1*, T. Drader2, A. Kleinhofs1,2
1Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-
6420, USA; 2School of Molecular Biosciences, Washington State University, Pullman, WA 99164-4234,
USA
Corresponding author: Andris Kleinhofs, Dept. Crop and Soil Sciences, Washington State
University, Pullman, WA 99164-6420, FedEx address Johnson Hall 201, Tel: 1-509-335-4389
Fax: 1-509-335-8674, E-mail: [email protected]
As Published In
Theoretical and Applied Genetics (2006) 113:1147-1158
* R. Brueggeman is the main contributor
51

Theor Appl Genet (2006) 113:1147–1158 DOI 10.1007/s00122-006-0374-3
123
ORIGINAL PAPER
The barley serine/threonine kinase gene Rpg1 providing resistance to stem rust belongs to a gene family with Wve other members encoding kinase domains
R. Brueggeman · T. Drader · A. Kleinhofs
Received: 5 April 2006 / Accepted: 15 July 2006 / Published online: 3 August 2006© Springer-Verlag 2006
Abstract The barley (Hordeum vulgare L.) stem rust(Puccinia graminis f. sp. tritici) resistance gene Rpg1encodes a serine/threonine protein kinase with twotandem kinase domains. The Rpg1 gene family wasidentiWed from the cv. Morex and consists of Wve addi-tional members with divergent homology to Rpg1. Allfamily members encode serine/threonine kinase-likeproteins with at least one predicted catalytically activekinase domain. The Wve family members weresequenced from cDNA and genomic DNA and geneti-cally mapped. The family member most closely relatedto Rpg1, ABC1037, is located on chromosome 1(7H)bin 01, very near (»50 kb) but not co-segregating withRpg1. Two others, ABC1036 and ABC1040, are closelyrelated to each other and tightly linked on chromo-some 7(5H) bin 07. ABC1041 mapped to chromosome7(5H) bin 13, tightly linked to the rust resistance genesrpg4 and Rpg5 providing resistance to barley stem rustpathotype QCC and rye stem rust pathotype 92-MN-90, respectively, but segregated away in a high-resolu-
tion population. ABC1063 was localized to chromo-some 4(4H) bin 6. An interesting Rpg1 allele thatappears to be the result of unequal recombinationbetween Rpg1 and ABC1037 was characterized. Noknown resistance loci cosegregated with any familymembers, however characterization of the Rpg1 familyhas provided insight into the evolution of this novelgene family and may present tools for understandingthe functional domains of Rpg1. The genetic mapping,gene structures, and analysis of amino-acid sequencesof the Rpg1 gene family members are presented.
Introduction
The barley stem rust resistance gene Rpg1 is a domi-nant gene that confers resistance to most, but not all,pathotypes of the stem rust fungus Puccinia graminis f.sp. tritici. Rpg1 was cloned by a map-based approachfrom the resistant cv. Morex (Brueggeman et al. 2002).A genomic fragment containing the complete Rpg1gene transformed into the highly susceptible cv.Golden Promise conferred resistance to the barleystem rust fungus pathotype MCC, demonstrating thatthe gene isolated was the functional Rpg1 gene (Horv-ath et al. 2003). In silico translation of the Rpg1sequence revealed an 837 amino acid (a.a.) proteinwith tandem protein kinase domains (PKDs), a novelresistance gene structure (Brueggeman et al. 2002).Analysis of the two Rpg1 kinase domains suggestedthat PKD2 was catalytically active while the PKD1may not function in autophosphorylation but may beimportant for resistance. This was conWrmed by site-directed mutagenesis and kinase activity assays (Nir-mala et al. 2006). Analysis of Rpg1 alternate splice
Communicated by B. Keller
Electronic Supplementary Material Supplementary material is available to authorised users in the online version of this article at http://dx.doi.org/10.1007/s00122-006-0374-3.
R. Brueggeman (&) · A. KleinhofsDepartment of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6420, USAe-mail: [email protected]
T. Drader · A. KleinhofsSchool of Molecular Biosciences, Washington State University, Pullman,WA 99164-4234, USA
52

1148 Theor Appl Genet (2006) 113:1147–1158
123
forms identiWed one predicted protein with a putativetransmembrane domain and signal peptide (Rostokset al. 2004); however, the majority of the Rpg1 proteinis cytoplasmic (Nirmala et al. 2006).
Rpg1 has signiWcant a.a. homology (NCBI BLASTP)to the PKD of the Brassica napus S-receptor kinasefamily with the highest homology to the gene familymember Srk15 (35% a.a. identity with Rpg1 PKD1 and34% a.a. identity with PKD2) (Cabrillac et al. 1999).Srk15 encodes a plasma membrane-spanning receptorkinase that is speciWc to the stigma epidermis (Steinet al. 1991, 1996) and presumed to be the determinantof self-incompatibility (Takasaki et al. 2000).
The Rpg1 serine/threonine (S/T) protein kinase (PK)belongs to the cd00180 domain family (http://www.ncbi.nlm.gov/structure/cdd/cddsrv.cgi?uid = cd00180&version =v2.06) placing Rpg1 into the RLK/Pelle super family ofS/T PKs. The RLK/Pelle family includes receptor-likekinases (RLKs) and receptor-like cytoplasmic kinases(RLCKs) (Shiu and Bleeker 2003). Alignment of char-acterized plant S/T PKDs showed that they range insize from 250–300 amino acid residues and are not con-served uniformly but are made up of alternatingregions of high and low conservation. Eleven majorconserved subdomains (I–XI) are evident and impor-tant for catalytic activity (S1). Nine highly conserveda.a. in kinase catalytic domains are expected to playimportant roles in adenosine triphosphate (ATP) bind-ing and phosphotransfer, involved in signal transduc-tion (Hanks et al. 1988). The nine invariant a.a.indicative of a catalytically active PK, were identiWedand used to predict the inactive PKD1 and activePKD2 present in Rpg1. These predictions were con-Wrmed by autophosphorylation assays (Nirmala et al.2006).
The past two decades has culminated in the cloningand partial characterization of more than 40 diseaseresistance (R) genes (Martin et al. 2003). The majorityof R-genes belong to a class of proteins containing anucleotide-binding site and leucine rich repeats (NBS-LRR) whereas only four have been characterized asPKs. Disease resistance systems have been identiWedthat require both PK and NBS-LRR genes. Thetomato Pto gene confers resistance to races of Pseudo-monas syringae pv. tomato that carry the avrPto orAvrPtoB gene (Martin et al. 1993; Kim et al. 2002).The Pto resistance function requires the presence ofthe NBS-LRR gene Prf (Salmeron et al. 1996). Pto islocated in the cytoplasm and contains no recognizablereceptor domain placing it into the RLCK class ofgenes. The Arabidopsis PBS1 gene, which confersresistance to the bacterial pathogen Pseudomonassyringae pv. phaseolicola (Swiderski and Innes 2001), is
another RLCK whose resistance function requires asecond gene, RPS5, which encodes an NBS-LRR gene.It was shown that the Arabidopsis PBS1 is cleaved fol-lowing infection by the pathogen eVector protein Avr-PphB protease and RPS5 may act as the guard forPBS1 (Shao et al. 2003).
RLKs and RLCKs are abundant in the Arabidopsisgenome, which contains more than 610 family mem-bers (Shiu and Bleeker 2003). Many of the ArabidopsisRLK/RLCK subfamilies are present in clusters. Theevolution of plant genomes is shaped by the occurrenceof multiple gene duplication events that may occurthrough unequal recombination. Like many plantgenes, most of the cloned plant R genes are membersof families of related genes, indicating that gene dupli-cation and subsequent diversiWcation are common pro-cesses in plant R gene evolution (Richter and Roland2000). Previously identiWed R genes related to Rpg1,Pto and Xa21, are part of complex gene loci containingtandem arrays of closely related genes, some of whichconfer diVerent or altered resistance speciWcities (Mar-tin et al. 1994; Song et al. 1997). Genetic studies of theXa21 and Pto gene families have provided insight intothe function and evolution of R genes.
The Pto gene is part of a small gene family consist-ing of at least six members located on tomato chromo-some 5 (Martin et al. 1993). It is presumed thatduplication and diversiWcation of the Pto gene familyhas led to the generation of alternative recognitionspeciWcities. The Pto gene family member Fen encodesa 318 a.a. S/T PK sharing 87% a.a. identity with Pto.Fen does not confer resistance to any tested pathogens,but mediates a response similar to HR in tomato plantstreated with the organophosphorous insecticide, fen-thion (Martin et al. 1994).
The cloned rice Xa21 gene (Song et al. 1995), whichconfers resistance to Xanthomonas oryzae pv. oryzaebelongs to a RLK gene family and has homology toRpg1 at the kinase domains (32% a.a. identity and 52%a.a. similarity to Rpg1 PKD2 and 28% a.a. identity and47% a.a. similarity to Rpg1 PKD1). The Xa21 multi-gene family contains at least eight members (Ronaldet al. 1992; Song et al. 1995). Most of these memberswere mapped to a single complex locus on rice chromo-some 11 that contains at least nine major resistancegenes (Ronald et al. 1992). Analysis of the coding andXanking sequences of seven Xa21 gene family mem-bers identiWed recombination, duplication, and trans-position as the major events contributing to theevolution of this complex resistance locus (Song et al.1997). It has been shown that recombination betweenseparate Xa21 family members has occurred during theevolution and that transposon insertions resulted in

Theor Appl Genet (2006) 113:1147–1158 1149
123
truncated genes with alternative gene structure anddiVering resistance speciWcity (Song et al. 1997).
In addition to Rpg1 our laboratory has been work-ing on the map-based cloning of two other barley stemrust resistance genes, rpg4 and Rpg5 (Druka et al.2000). In this study we identiWed and partially charac-terized the Rpg1 barley stem rust resistance gene fam-ily with the goal of identifying other rust resistancegenes particularly the rpg4 and Rpg5 genes. The Rpg1gene family members in cv. Morex were identiWed byutilizing the Rpg1 gDNA probe to isolate homologousbarley sequences by cross-hybridization and by using insilico database mining. The Rpg1 gene family memberswere mapped and each was sequenced from both geno-mic and cDNA sources to determine their gene struc-ture. One member was closely linked to Rpg1 andanother was closely linked to the rpg4/Rpg5 locus. TheRpg1 barley gene family members did not co-segregatewith any other mapped barley disease resistance locibut they could function in disease resistance and possi-bly rust resistance pathways yet to be discovered. Thiswork also uncovered and partially characterized anapparent unequal recombination event that produced achimeric gene between Rpg1 and its closely linked fam-ily member ABC1037.
Materials and methods
Southern analysis
Barley genomic DNA isolation, Southern blotting andhybridization were as described (Kleinhofs et al. 1993).Approximately 25 ng of cDNA insert was labeledwith32P dCTP (New England Nuclear) using the RTSRadPrime DNA labeling system (Gibco BRL) accord-ing to manufacturers instructions.
Library screening
Arrayed barley cv. Morex cDNA libraries or the barleycv. Morex BAC library (Yu et al. 2000) were initiallyscreened using the Rpg1 gDNA probe RSB228 (Bru-eggeman et al. 2002). Clones identiWed from the cDNAlibraries were used as new probes to re-screen thecDNA libraries to identify further diverged familymembers and to hybridize the Morex BAC library toidentify genomic clone equivalents for each cDNA.For BAC conWrmation blots, DNA was digested withrestriction enzymes following the manufacturer’s rec-ommendations (New England Biolabs, MBI Fermen-tas, Gibco BRL) and transferred to nylon membranes(New England Nuclear) by the alkaline transfer proce-
dure. Hybridizations were at 65°C and the Wnal washwas at 65°C with 0.5 SSC 0.1% SDS.
Genetic mapping
Low resolution genetic mapping used the Steptoe £Morex “minimapper” population consisting of 35 linesselected from the original USBGP (US Barley GenomeProject) 150 Hordeum bulbosum-derived doubled-hap-loid line population (Kleinhofs et al. 1993; Kleinhofsand Graner 2001). The “minimapper” lines wereselected to allow placement of probes to speciWc bins onthe Steptoe £ Morex map (http://wheat.pw. usda.gov/cgi-bin/graingenes/report.cgi?class = mapdata;query =Barley*;name = Barley,+BinMap + 2005). High resolu-tion mapping at the Rpg1 locus utilized 33 recombi-nants selected from approximately 8,000 gametesbetween the Rpg1 Xanking markers ABG704 andABG077 as described in Brueggeman et al. (2002).High resolution mapping at the rpg4/Rpg5 locus utilized55 recombinants selected from approximately 5,000gametes between the rpg4/Rpg5 Xanking markers Aga5and ABG391 as described in Druka et al. (2000).
CAPS marker development
The ABC1063 gene was non-polymorphic using RFLPmarkers therefore Cleaved AmpliWed PolymorphicSequence (CAPS) markers were developed. TheMorex cDNA sequence was utilized to design primersto amplify »1 kb overlapping fragments of the genefrom Morex and Steptoe. The PCR fragments wereseparated on a 1£ TE, 1% agarose gel with low EDTA(0.002 M) and run at 100 V. The PCR fragments wereexcised from the gel and placed in trimmed Wlter tips(Rainin RT-L22F) inside 1.5 ml tubes and frozen.After thawing, the DNA was eluted by centrifuging at5,000 rpm for 10 min. The fragments were directlysequenced and compared to the Morex sequence inorder to identify single nucleotide polymorphisms(SNPs) that could be utilized for CAPS marker devel-opment. An AciI SNP was identiWed, primers devel-oped and used to map the gene.
Cloning and sequencing
The Morex BAC clones detected by hybridization withthe Rpg1 gene family cDNAs were isolated, DNAextracted, digested with HindIII, Southern-blotted andhybridized with the Rpg1 gene family cDNA probes.Representative BAC clones for each gene weredetected, digested with HindIII, EagI, SacI and XbaIand shotgun-cloned into the plasmid pBluescript II KS
54

1150 Theor Appl Genet (2006) 113:1147–1158
(Stratagene) by standard techniques (Sambrook et al.1989). Plasmid clones containing the Rpg1 family mem-bers were conWrmed by Southern blot analysis andsequenced using the BigDye terminator system on anABI Prizm 377 DNA sequencer (PE Biosystems) at theBioanalytical Center, Washington State University, Pull-manWA. The larger subclones, >3 kb, were sequencedusing the EZ:TN <KAN-2> Transposon Insertion Kit(Epicenter) and in some cases primers were designedand the BAC clone DNA and large insert subcloneDNA was sequenced directly. Primers were designedacross restriction enzyme junctions to conWrm that thesequence across these junctions was properly assembled.For a complete list of plasmids used for sequencing seeElectronic Supplementary Material (S2).
RNA isolation and RT-PCR
Total leaf RNA from 14-day-old Morex plants was iso-lated using a Trizol RNA method (Chomczynski andSacchi 1987) with modiWcations (R. Wise, Iowa StateUniversity, personal communication). Integrity andquantity of the RNA samples were checked by formal-dehyde denaturing agarose gel electrophoresis. Approx-imately 1 �g of total RNA was used for RT-PCRreactions using M-MLV Reverse Transcriptase (Pro-mega) under the manufacturer’s recommended standardconditions. RT-PCR fragments were directly sequencedusing the low EDTA elution method described above.
Northern analysis
About 500 ug of total leaf RNA was used to isolatepolyA mRNA using the oligotex mRNA Mini kit(Qiagen). Three to Wve micrograms of polyA mRNAwere denatured in 50% v/v formamide, 6.1% formalde-hyde, 1£ MOPS buVer (40 mM MOPS, pH 7.0, 10 mMsodium acetate, 1 mM EDTA) for 15 min at 55°Cimmediately before loading. Samples were resolvedwith a 1% w/v agarose, 6.7% v/v formaldehyde gel in1£ MOPS buVer for 2 h at 4 V/cm. Northern transferwas performed following the manufacturer’s recom-mendations (Perkin Elmer Genescreen Plus). Mem-brane was soaked in 2£ SSC and prehybridized (5£SSC, 5£ Denhardts, 1% SDS, 10% Dextran sulfate,50% Formamide) for 3 h at 42°C in a hybridizationtube using »3 ml solution per 10 cm2 of membrane.The cDNAs were labeled using {�-32P]-dCTP andALL-IN-ONE Random Prime DNA Labeling Mix(Sigma-Aldrich). Unincorporated nucleotides wereremoved using a performa DTR gel Wltration cartridge(Edge Biosystems). Hybridization (5£ SSC, 5£ Den-hardts, 1% SDS, 10% Dextran sulfate, 50% Formam-
ide) was done at 42°C overnight. Filters were rinsedtwice in 2£ SSC/0.1% SDS, 5 min at room temp oncein 0.2£ SSC/0.1% SDS, 10 min at room temp, once in0.2£ SSC/0.1% SDS, 30 min at 42°C and twice in 0.2£SSC/0.1% SDS, 30 min at 65°C. Autoradiography wasat ¡80°C for 7 days.
5� RACE
5� RACE was performed using three methods: (1) 5�
RACE System for Rapid AmpliWcation of cDNAEnds (Invitrogen); (2) Primer switching method; and(3) Nested PCR/DpnI method. The 5� RACE forRapid AmpliWcation of cDNA Ends kit was usedunder Invitrogen’s recommended standard condi-tions. Primer switching method was performed asdescribed by Rosenberg et al. (2003). The PCR/DpnImethod was performed using two gene-speciWcnested reverse primers designed from the mostextending 5� cDNA sequence of each gene. DNA wasisolated from a library of phagemid particles contain-ing cDNA derived from cv. Morex seedling shootscloned into the Lambda Uni-ZAP XR vector (Strata-gene) (provided by T. J. Close, UC Riverside, CA,USA). The Wrst round of ampliWcation contained20 ng of lambda DNA, 0.2 mM dNTP mix, 25 pmol ofT7 primer, 25 pmol of gene-speciWc reverse primer,2.5 �l of RedTaq DNA polymerase (Sigma), and 5 �lof 10£ RedTaq reaction buVer in 50 �l volume.AmpliWcations were performed in a PTC-1000 pro-grammable thermocycler (MJ Research) at 95°C for4 min, followed by 10 cycles of 95°C for 1 min, 60°Cfor 1 min, and 72°C for 2 min; followed by 60°C for3 min and 72°C for 5 min. The ampliWed DNA wasthen digested with 10 U of the Dam methylation-dependent restriction enzyme DpnI (New EnglandBioLabs). The second round of ampliWcation con-tained 2.5 �l of DpnI digested Wrst round PCR reac-tion product, 0.2 mM dNTP mix, 25 pmol of SKprimer, 25 pmol of gene-speciWc nested reverseprimer, 2.5 �l of RedTaq DNA polymerase (Sigma),and 5 �l of 10£ RedTaq reaction buVer in a 50 �l vol-ume. Second round ampliWcations were at 95°C for4 min, followed by 35 cycles of 95°C for 1 min, 60°Cfor 1 min, and 72°C for 2 min followed by 72°C for5 min. The PCR reaction was then loaded onto a lowEDTA 1% agarose gel and directly sequenced usingthe second round primers as described previously.
Phylogenetic analysis
A phylogenetic tree (CLUSTALX Phyllip NJ plotbootstrapped 1,000£) of the PK was developed using
123 55

Theor Appl Genet (2006) 113:1147–1158 1151
123
the derived a.a. sequences from the Rpg1 gene familyPKDs and published a.a. sequences of Rpg1, Pto, Xa21and SRK 15.
Results
IdentiWcation of Rpg1 gene family members
Hybridization of the the Rpg1 gDNA probe RSB228(Brueggeman et al. 2002) to ten-arrayed barley cDNAlibraries (Wrst 10 listed at http://www.genome.ari-zona.edu/genome/barley.html) representing approxi-mately 500,000 clones identiWed one Rpg1 cDNA (acc.# AY115550) (Brueggeman et al. 2002), and two othercDNA clones designated ABC1036 (acc. # DQ469715)and ABC1037 (acc. # DQ469714). Sequence analysis ofthese cDNA clones revealed that ABC1037 was closelyrelated to Rpg1 and ABC1036 was more diverged fromRpg1. Utilizing the ABC1036 and ABC1037 nucleotidesequences, a BLASTN search of the Triticeae ESTdatabase (NCBI) identiWed two new barley cv. MorexESTs (BG366053, BG300587). Sequencing of the ESTcDNA clones revealed two new genes, ABC1040 andABC1041. An additional ABC1037 EST (BI956515)was also identiWed. Using ABC1036, ABC1037,ABC1040, and ABC1041 as hybridization probes, 21cDNA clones were identiWed from the ten cDNAlibraries (S3). A BLASTN search of the barley ESTdatabase (NCBI) with the ABC1041 sequence identi-Wed two additional ESTs (BF621830 and 628226) rep-resenting one more gene, ABC1063. Hybridization ofthe original ABC1063 cDNA clone to the 10 cDNAlibraries identiWed 20 cDNAs all representingABC1063 transcripts (S3).
The Rpg1 family of genes were separated into threehomology-based groups: (1) Rpg1 and ABC1037; (2)ABC1036 and ABC1040; (3) ABC1041 and ABC1063.The gene structures and homology relationships areshown in Fig. 1. The group 1 gene, ABC1037, containstwo tandem protein kinase domains and a N-terminaldomain with good homology to Rpg1. ABC1036 andABC1040 are very closely related to each other, butfurther diverged from Rpg1. Both ABC1036 andABC1040 contain the two tandem PKDs, but not theN-terminal region homologous to the 110 a.a. N-termi-nus of Rpg1. ABC1041 and ABC1063 contain only thePKD2 and are moderately homologous to each otherand quite diverged from Rpg1 (Fig. 1). ABC1041 andABC1063 are included as Rpg1 gene family membersdue to their homology with the ABC1036 andABC1040 PKD2, which is quite diverged from theRpg1 and ABC1037 pKD2. These results suggest that
the diVerent two tandem kinase domain genes mighthave arisen by diVerent evolutionary pathways.
Genetic and physical mapping
The Rpg1 family members mapped to three of theseven barley chromosomes (Fig. 2). ABC1037 mappedon chromosome 1(7H) bin 01 and co-segregated withRpg1 in a low-resolution mapping population, but wasproximal to Rpg1 by two crossovers out of 8,518gametes in a high-resolution Rpg1 mapping population(Brueggeman et al. 2002). Rpg1 and ABC1037 resideon the same Morex BAC clone 607p19 and are sepa-rated by »50 kb. ABC1036 and ABC1040 co-segre-gated at low resolution and mapped to chromosome7(5H) bin 07. They have not been mapped at high reso-lution, but hybridization analysis showed they were notlocated on overlapping BAC clones. ABC1041mapped to chromosome 7(5H) bin 13 and co-segre-gated with the barley stem rust resistance genes rpg4and Rpg5 at low resolution, but in a high-resolutionmapping population (Druka et al. 2000) segregated dis-tal of the rpg4/Rpg5 locus. ABC1063 mapped to chro-mosome 4(4H) bin 06.
The barley cv. Morex BAC library (Yu et al. 2000)was screened with the Rpg1 family cDNA probesdescribed above and a total of 28 BAC clones wereidentiWed. Southern blot analysis conWrmed that BAC
Fig. 1 Diagram of the predicted Rpg1 protein domains and align-ment with family members. The cv. Morex Rpg1 protein pre-dicted functional domains were aligned with the Wve Rpg1 familymembers. The Sm89010 line is included to help visualize the un-equal recombination that appears to have taken place betweenRpg1 and ABC1037. The boundaries of the three protein do-mains are indicated on Rpg1. The a.a. identity between proteindomains of diVerent members and Rpg1 is presented as % iden-tity. The % a.a. identity between adjacent family protein domainsis in parentheses
N-terminal domain
Rpg1 p kinase 1 p kinase 2
ABC1037 p kinase 1 p kinase 2
p kinase 1 p kinase 2ABC1036
ABC1040
ABC1041
ABC1063
p kinase 1 p kinase 2
p kinase 2
p kinase 2
Protein kinase 1 Protein kinase 2
42% (86%) 73% (73%) 67% (67%)
59% (57%) 28% (28%)
59% (98%) 28% (92%)
28% (74%)
29% (75%)
Sm89010 p kinase 1 p kinase 2
42% 98% 98%
56

1152 Theor Appl Genet (2006) 113:1147–1158
123
clones were selected for all Wve family members (S3)and comparison with genomic southerns revealed thatall hybridizing fragments were identiWed (Fig. 3).
cDNA sequence analysis
The cDNA clones identiWed by database mining andhybridization (S3) were sequenced from 3� and 5� ends.The end sequences were placed into contigs represent-ing Wve unigene sequences. A clone from each of thegene contigs representing the most full-length cDNA
was sequenced (Table 1). Clones that did not appear torepresent full-length cDNAs were extended using RT-PCR facilitated by predicted gene structure deducedfrom the genomic sequence. The genomic gene struc-tures were predicted by sequence comparison with theRpg1 cDNA sequence and using the FGENESH geneprediction program (http://www.softberry.com). Theequivalent genomic sequences were obtained fromBAC subclone libraries (Table 1). Analysis of thecDNA clone sequences indicated that all containeduninterrupted open reading frames and may representfunctional genes. The cDNA sequences were alignedwith their respective genomic sequences to identify theexon/intron junctions (Fig. 4). The exon/intron junc-tions were highly conserved between Rpg1 andABC1037 and between Rpg1 and ABC1036 andABC1040 PKD1. The PKD2 exon/intron junctionswere conserved among the ABC1036, ABC1040,ABC1041 and ABC1063 genes, but not with Rpg1.These results are in line with expectations based on thehomology analysis (Fig. 1).
Comparison of the most 5� ABC1037 cDNA sequencewith the genomic sequence suggested that it did notrepresent a full length mRNA. We were not able toextend this sequence using several 5�-RACE proce-dures. However, a probable full-length mRNAsequence was predicted using RT-PCR and primersdesigned from the genomic sequence. This procedureidentiWed the putative transcription start site (TSS) toposition ¡52 to ¡161 based on the predicted AUGstart methionine for ABC1037. The predicted size ofthe ABC1037 transcript was conWrmed by Northernblot analysis (Fig. 5).
Fig. 2 Genetic map of the Rpg1 gene family members. The barley Steptoe £ Morex minimapper population was used to locate the genes on the genetic linkage BIN map. The position of the Rpg1 family members, Rpg1, rpg4 and Rpg5 are highlighted. Dis-tances between bin markers (on the right) are shown in cM (on the left). The BIN num-bers are in the circles. BIN locations and markers are de-scribed in Kleinhofs and Gra-ner (2001)
Chromosome 1(7H) Chromosome 7(5H)Chromosome 4(4H)
cM Bin Marker
ABG7041
2
3
4
ABC1037ABG320
Cen 1
5
6
7
8
9
10
11
12
cM Bin Marker
MWG6341
2
3
Cen 4
4
5
6
7
8
9
10
11
cM Bin Marker
MWG920.1A1
2
3
4Cen 7
5
6
7
8
9
10
11
12
ABC151A
ABG380ksuA1A
ABC255
ABG701
Amy2
RZ242ABC310BABC305
ABG461A
Tha
JS103.3
Ole1BCD402B
ABG484
BCD808B
ABC1063bBE54ABCD453BABG319A
KFP221
ABG397
ABG319C
Bmy1Tel4L
12
13
131415
DAK133
ABG705AABG395Ltp1WG530
ABC324ABC1036,ABC1040ABC302ABCD926
ABG473
MWG514BWG908
AGB496ABG390ABC1041ABG463Tel7L
0.0
9.4
18.6
rpg4, Rpg5
Rpg1
24.9
34.3
50.9
64.6
77.4
88.998.0
105.7
117.2
137.6
4.8
15.6
27.1
33.8
45.3
66.275.0
84.595.5
105.6
117.0
132.5
142.3
3.511.6
37.7
45.5
56.2
68.5
75.9
87.1
100.4
132.5142.1
156.8169.5
187.7
Fig. 3 Southern blots of genomic and BAC DNA hybridizedwith the Rpg1 gene family member cDNA probes. a GenomicDNA was digested with HindIII, separated on gels and hybrid-ized with cDNA probes from the family members indicated.b BAC DNA from clones used for sequencing hybridized withthe cDNA probe as above. ABC1036 is represented by two BACclones because the entire genomic equivalent of the gene was notfound on a single BAC clone. Some of the faint bands are due tocross hybridization between family members. The approximatesize in kilobases is shown to the left
AB
C10
37
AB
C10
36
AB
C10
40
AB
C10
41
AB
C10
63
AB
C10
37
AB
C10
36
AB
C10
40
AB
C10
41
AB
C10
63
10
5
1.5
AB
C10
36
57

Theor Appl Genet (2006) 113:1147–1158 1153
123
The ABC1036 and ABC1040 cDNAs (acc. # sDQ469715 and DQ469716, respectively) alsoappeared to be incomplete. 5� RACE identiWed theTSS at position +551 bp relative to the Rpg1 genomicsequence. This placed the TSS of ABC1036 andABC1040 at the beginning of subdomain I of thePKD1. This result seemed improbable since it wouldresult in the translation start site 5� of subdomain IIIof the PKD1 and a truncated PKD1. Several attemptsto identify additional 5� cDNA sequences using prim-ers based on genomic sequences failed. Northern blotanalysis predicted mRNAs of »2.9 kb (Fig. 5) while
the cDNA sequence analysis predicted a »2.57 kbtranscript.
The ABC1041 and ABC1063 cDNAs (acc. # sDQ469717 and DQ469718, respectively) appeared to befull-length messages containing a putative TSS and a fulllength 5� UTR. In silico translation of the cDNAsequences suggested that the coding sequences were com-plete since in-frame stop codons were present upstreamof the Wrst predicted start methionine indicating a 5�
UTR. This data was supported by Northern blot analysisshowing that the mRNAs were »1.7 kb and »1.5 kb,respectively, as predicted by the sequences (Fig. 5).
Fig. 4 Rpg1 gene family member exon/intron structures. The in-trons (gray) and exons (black or striped) are shown to scale. Ex-ons with sequence conservation as compared to Rpg1 are in blackand are numbered above the drawing according to the Rpg1 geneexons. Exons with low homology to Rpg1 are striped. The exon
numbering speciWc for each gene is shown below the gene. Smalldashed lines joining intron/exon junctions indicates conservationof these exon splice sites. Large introns are indicated by inter-rupted back slashes and approximate sizes are shown in kilobases
Kb 1 2 3 4 5
Rpg11 2 3 4 5 6 7 8 9 10 11 12 13 14
1 2 3 4 5 6 7 8 9 10 11
3 4 5 6
3 4 5 6
ABC1037
ABC1036
ABC1040
ABC1041
ABC1063
~12.5 ~5.5 ~0.71 2 3 4 5 6 7 8 9 10 11 12
~ 8 ~10
~7.2 ~ 8
1 2 3 4 5 6 7 8 9 10
1 2 3 4 5 6 7 8 9 10
~ 2.3
~1.1
1 2 3 4 5 6
1 2 3 4 5 6
Table 1 Rpg1 gene family cDNA and genomic plasmids and GenBank accession numbers
cDNA plasmid clones are designated with library they were isolated from and the address. All BAC clones are from the cv. Morex BAClibrary. The ABC1036 and ABC1040 full-length cDNA sequences were completed by RT-PCR fragment sequencing
Rpg1 familymember
cDNA clone usedfor mapping
cDNA Genbankaccession number
BACclones usedfor sucloning
gDNA Genbank accession number
ABC1037 HVSMEn0003o03 DQ469714 290m12 DQ351213ABC1036 HVSMEg0085c17 DQ469715 666a11701k24 DQ469712ABC1040 HVSMEi0028b15 DQ469716 623E8 DQ469713ABC1041 HVSMEb0017j15 DQ469717 572b18 DQ352178ABC1063 HVSMEa0001a21 DQ469718 435B24 DQ386610
58

1154 Theor Appl Genet (2006) 113:1147–1158
123
Genomic sequence analysis
A 2 kb sequence 5� of the ABC1037 TSS was analyzedand compared with the Rpg1 promoter sequence usingNCBI BLAST 2 program. No apparent or signiWcantsimilarity between the two gene promoter regions wasdetected. A search for small almost identical stretchesof nucleotides was also negative.
The ABC1037, ABC1036 and ABC1040 genes con-tained very large introns from 8,057–12,516 bp (Fig. 4).BLAST searches of the ABC1037 genomic sequencerevealed four transposon-like elements, two within the3rd intron and two within the 11th intron (Fig. 4). TheWrst transposon-like element in the third intron had92% nucleic acid (n.a.) homology to a transposon “tha-los MITE” element annotated from Hordeum vulgare(acc. # AY146587). The second transposon-like ele-ment in the third intron had 84% n.a. identity to anunclassiWed class II DNA transposon from Triticumaestivum “George_1611A10_1.1” (acc. # CR626934).Transposon like elements present in the 11th intronhad »92% n.a. identity to transposon “Xobar” and92% n.a. identity to transposon “Sherlock” respec-tively from Hordeum vulgare (acc. # AY661558).BLASTX and BLASTN searches of the fourth intronof ABC1036 and both the fourth and ninth introns ofABC1040 revealed insertions of retrotransposon-likeelements with high (82–84%) n.a. homology to a LTRretrotransposon Leojyg-184G9-1 from Hordeum vulg-are, (acc. # AY268139.1) (Fig. 4). The Leojyg-184G9-1-like retro element found in the ninth intron ofABC1040 had a second retrotransposon-like element
nested within it with 95% n.a. homology to the Copia/Ty1-like retrotransposon “Horpia-2 from Hordeumvulgare, (acc. # AF427791). A BLASTX search of the10th intron of ABC1036 also revealed a polyproteinlike sequence representing a possibly uncharacterizedbarley retroelement with 55% a.a. identity to a reversetranscriptase from a retrotransposable-like element ofMedicago truncatula (acc. # ABD28426.1) (Fig. 4).
Characterization of the Rpg1 family gene structure
The Rpg1 gene structure was divided into three proba-ble functional domains: (1) the N-terminal domainfrom a.a. 1 to 129, (2) PKD1 from 130 to 393 a.a., and(3) PKD2 from 438 to 710 a.a. (cv. Morex Rpg1 num-bering, acc. # AF509748). Amino acid sequence homol-ogy within these functional domains was comparedbetween diVerent Rpg1 family members and Rpg1(Fig. 1). The ABC1037 gene was most closely relatedto Rpg1, sharing 64% overall a.a. identity. ABC1037has a gene structure similar to Rpg1, with two tandemS/T kinase domains that share 73 and 67% a.a. identitywith Rpg1, respectively, and a 98 a.a. N-terminalregion with 49% homology to the N-terminus of Rpg1.Genomic and cDNA sequence analysis of ABC1037indicated a functional gene predicted to contain 12exons in a total genomic sequence of 22,183 bp encod-ing a 694 a.a. protein (Fig. 4). A BLASTN andBLASTX search with the ABC1037 N-terminal codingsequence of the non-redundant database (NCBI)revealed no other signiWcant homologies besides theRpg1 homology.
Two other Rpg1 family members, ABC1036 andABC1040, were very closely related to one another,91% overall a.a. identity, and have two protein kinasedomains, but were only moderately homologous toRpg1, »59% a.a. identity at PKD1 and »28% identityat PKD2 (Fig. 1). ABC1036 and ABC1040 have con-served intron/exon junctions that are conserved withRpg1 only in the PKD1 domain. Genomic and cDNAsequence analysis indicated that ABC1036 andABC1040 may not be functional and could representtranscribed pseudogenes. ABC1036 and ABC1040contain ten exons in a total sequence of 21,924 and18,253 bp (Fig. 4), respectively, coding for similar 642and 632 a.a. predicted proteins.
More distantly related Rpg1 family membersincluded ABC1041 and ABC1063. These genes areclosely related (75% overall a.a. identity), but map todiVerent chromosomes. ABC1041 contains six exons ina total sequence of 4,562 bp (Fig. 4) coding for a 425a.a. predicted protein. ABC1063 also contains sixexons in a total sequence of 2,934 bp (Fig. 4) coding for
Fig. 5 Northern blot of the Rpg1 gene family members. Lane 1 isMillenium Marker (Ambion) stained with methylene blue. Lanes2–6 are cv. Morex mRNA hybridized with Rpg1 family membercDNA probes (lane 2 is ABC1036, lane 3 is ABC1037, lane 4 isABC1040, lane 5 is ABC1041 and lane 6 is ABC1063). Markerband sizes are in kilobases to the left
1 2 3 4 5 69
654
32.5
21.5
1
0.5
59

Theor Appl Genet (2006) 113:1147–1158 1155
123
a 372 a.a. predicted protein. These genes lack the Rpg1N-terminal domain and PKD1 (Fig. 1). The ABC1041and ABC1063 homology to Rpg1 in the PKD2 was low(28 and 29% a.a. identity, respectively), but theyappeared to be functional as judged by the presence ofall nine a.a. conserved in functional PKs (S1).
Phylogenetic relationships of the PK domains
Phylogenetic analysis of the Rpg1 family memberPKDs suggested that the ABC1037 and Rpg1 tandemkinase domains arose by duplication of a single progen-itor PKD. The resulting gene with two tandem PKDswas then duplicated giving rise to the two genes(Fig. 6). The ABC1036 and ABC1040 PKD1 is closelyrelated to the Rpg1 and ABC1037 PKD1, but thePKD2 is not, suggesting that the two PKDs evolvedseparately and were brought together by an unknowntransposition event. This tandem kinase progenitorgene then presumably underwent a localized duplica-tion, giving rise to ABC1036 and ABC1040.
Oryza sativa protein kinase homologs
A tandem kinase gene (ABA92216.1) with homology toRpg1 and ABC1037 was identiWed using BLASTPsearch NCBI (39 and 38% a.a. identity, respectively).However, there is a single kinase gene annotated fromrice (ABA94724.1) with higher homology to each of thekinase domains of Rpg1 (51 and 50% a.a. identity toPKD1 and PKD2, respectively) and ABC1037 (38 and41% a.a. identity to PKD1 and PKD2, respectively).ABC1036 and ABC1040 do not have tandem kinasegene homologues from rice. However, two separate sin-gle kinase genes (ABA94724.1 and AAO18450.1) havebeen identiWed with high homology to each of the
tandem kinase domains of ABC1036 and ABC1040 (61and 60% for PKD1, respectively and 65 and 70% forthe respective PKD2). The rice gene with the highesthomology to ABC1041 (90% a.a. identity) is the samesingle kinase gene (AAO18450.1) previously identiWedwith the highest homology to the PKD2 of bothABC1036 and ABC1040, which is consistent with thephylogenetic analysis and a.a. alignments. The best hitwith ABC1063 is a unique rice kinase (NP 914370.1)with 76% a.a. identity. There is no known functionfor the rice PK genes with homology to the Rpg1 genefamily.
An example of unequal recombination
Sequence analysis of the Rpg1 alleles from the suscep-tible cultivars Sm89010, Dicktoo, and Gobernadorawere previously characterized as having a highlydiverged 5� coding region (Brueggeman et al. 2002).The N-terminal region of this allele was highly homolo-gous (86% a.a. identity) to the 5� region of ABC1037while the 3� region including PKD1 and PKD2 washighly homologous (98%) to Rpg1 indicating that anunequal recombination event between Rpg1 andABC1037 probably gave rise to the Sm89010 allele.The predicted unequal recombination event occurredwithin the third intron of Rpg1 and the third intron ofABC1037. The recombinant gene is spliced together atthe PKD1 subdomain I (S1). RT-PCR, using primersthat span the chimeric junction, and subsequentsequence analysis indicated that the gene is transcribedand the recombinant intron is spliced out to form aprobable functional gene (data not shown). The func-tion of the Sm89010 Rpg1 allele is unknown, but it doesnot function in stem rust pathotype MCC resistance asindicated by susceptibility of the Sm89010 line.
Fig. 6 Phylogenetic tree of the barley Rpg1 gene family member protein kinase do-mains (PKDs) based on pre-dicted amino acid sequence and related protein kinases. Rpg1 (acc. # AAM81972), Pto (acc. # A49332), Xa21 (acc. # 220341A) and SRK 15 (acc. # CAB41879). The dend-ogram was created using Clu-stalX program by neighbor-joining method (Saitou and Nei 1987) and was boot-strapped 1,000 times. The number values represent the best branch location based on the 1,000 matrices
0.05
1036 PKD1
1040 PKD1
1037 PKD1
Rpg1 PKD1
1000
1000
1037 PKD2
Rpg1 PKD2
712
996
SRK15
1036 PKD2
1040 PKD2
1041 PKD2
1063 PKD2
1000
851
Pto
1000
Xa21
527
818
1000
60

1156 Theor Appl Genet (2006) 113:1147–1158
123
Discussion
Our goal was to characterize the Rpg1 gene familymembers in order to determine if family members wereresponsible for resistance against other rust diseasesand to evaluate their evolutionary relationships. Onemember, ABC1041, mapped very close, but did not co-segregate, with the rpg4/Rpg5 genes. AlthoughABC1041 was not rpg4 or Rpg5, we cannot exclude itsparticipation with these resistance genes or pathwayssince kinases are sometimes required together withNBS-LRR genes to provide disease resistance as dem-onstrated by the Pto/Prf and PBS1/RPS5 gene pairsdescribed in the Introduction.
The ABC1037 gene, closely linked to and veryhomologous to Rpg1, probably arose by a recent dupli-cation event. Duplication and subsequent divergence isthe major theme believed to have shaped the evolutionof plant disease resistance genes (Richter and Roland2000). We are not aware of any disease resistance func-tion for ABC1037 although it appears to be a func-tional gene. ABC1037 was apparently involved in afairly recent unequal recombination event giving riseto a chimeric gene with an ABC1037 N-terminal region(a.a. 1–110) and a C-terminal region from Rpg1 (a.a.140–837). The chimeric gene was not functional inresistance to the barley stem rust pathotype MCC andits function, if any, is unknown although it is expressedat the mRNA level. It could possibly function to pro-vide resistance to as yet unidentiWed rust pathotypes orother pathogens. Evidence exists that recombinationbetween gene family members or diVerent alleles ofthe same gene may be involved in the creation of novelresistance speciWcities (Richter et al. 1995; Dodds et al.2001).
It is tempting to speculate that the N-terminalregions of the Rpg1 and ABC1037 genes are importantfor R gene speciWcity, possibly representing a novelreceptor domain or a domain responsible for interac-tion with a receptor. BLASTN and BLASTX searchesusing the N-terminal domains of ABC1037 and Rpg1did not reveal homology to any known receptors. Adomain swapping study is underway to further explorethe N-terminal receptor hypothesis.
Some of the Rpg1 family members appeared to bethe products of recent duplication events and subse-quent divergence suggesting that the newly emerged orduplicated genes may have taken on a subset of theoriginal function. The new genes may have other rustresistance speciWcity as described for the M and N lociof Xax and the Rp1 locus in Maize (Richter et al. 1995;Dodds et al. 2001) or resistance against other patho-gens or completely novel functions. However, genetic
mapping did not associate the Rpg1 family memberswith any known functions. Although all of the geneswere transcribed, their translation was not tested andpossible functions, if any, are unknown.
The Rpg1 family members ABC1036 and ABC1040are diverged from Rpg1, but very closely related toeach other indicating a recent duplication event. BothABC1036 and ABC1040 have uncommonly largefourth introns, containing sequences with high homol-ogy to Leojyg-like retrotransposon inserted at thesame region suggesting that the duplication occurredafter the retroelement insertion. The ninth intron ofABC1040 contains another Leojyg-like retrotransponelement with a Horpia 2-like retrotransposon elementnested within it that is not present within the ninthintron of ABC1036 suggesting this insertion eventoccurred after duplication or the element was deletedfrom ABC1036 after duplication.
The PKD2 containing Rpg1 family members,ABC1041 and ABC1063, had relatively little homologyto Rpg1 (Fig. 1). These two genes were placed into theRpg1 gene family based on their high homology to thePKD2 of the family members ABC1036 and ABC1040.ABC1041 and ABC1063 are also diverged from oneanother and mapped to diVerent chromosomes. Theyare expressed at the transcription level and containprobable functional kinases, but their function, if any,is not known.
Most of the Rpg1 family members (ABC1036,ABC1037, ABC1040, and ABC1041) contained unusu-ally large introns ranging from »3 to »12.6 kb in lengththat are not present in Rpg1 (Fig. 3). These are amongthe largest plant introns reported to date. cDNAsequences show that these large introns are splicedout during mRNA maturation. The largest intronscontained repetitive retrotransposon- and transpo-son-like sequences. These repetitive element inser-tions are probably responsible for the expansion ofthese large introns. Repetitive element duplicationand insertion has been implicated for the expansionof large grass genomes (Bennetzen 2000). In rice atthe Xa21 locus, transposable elements and retrotrans-posable elements are a major source of variability andcan even create novel resistance functions as demon-strated with Xa21D. Transposable elements may alsoinXuence the rate of recombination (Richter andRonald 2000) or facilitate duplication events by serv-ing as the site for homologous unequal recombination(Ellis et al. 1995).
Analysis of the nine highly conserved a.a. in S/T PKs(as reviewed in Introduction) showed that the Rpg1PKD1 had four a.a. and ABC1037 PKD1 had three a.a.that were not conserved. Otherwise, all the other
61

Theor Appl Genet (2006) 113:1147–1158 1157
123
PKDs had all nine a.a. conserved suggesting that theyare functional PKs and could play a role in signal trans-duction.
The 5� RACE products obtained for ABC1036 andABC1040 consistently resulted with a TSS at the PKD1subdomain I. This TSS would result in a translationstart site after subdomain III and would translate atruncated PKDI. It is possible that the 5� RACE resultsfor ABC1036 and ABC1040 are due to a mRNA sec-ondary structure that terminated the reverse transcrip-tion reaction at that point prematurely. On the otherhand, it is possible that there are additional small exonsupstream of large introns that were not detected in thisstudy. The mRNA size estimated from Northern blot(2.9 kb) is not very diVerent from the sequence pre-dicted size of 2.57 kb; therefore, it is possible that this isthe correct TSS. The ABC1040 Northern blot alwayshybridized weakly and showed considerable smearingsuggesting that there could be multiple TSSs or thisgene may be pseudogene with an unstable mRNAtranscripts (Fig. 4).
Protein kinases with tandem kinase domains appearto be fairly rare. The RLK/Pelle is one of the largestgene families in Arabidopsis with >600 members yetonly two tandem kinase genes have been identiWed (S.Shiu, personal communication) (At1g11300, acc. #NM_101003 and At2g32800, acc. # NM_128840). BothAt1g11300 and At2g32800 genes are represented byfull-length cDNA sequences in the NCBI database andboth tandem PKDs belong to the same subfamily asRpg1 (cd00180 domain family). At2g32800 is a tandemkinase gene with similar structure to Rpg1. At1g11300kinase domains are not adjacent to one another buthave b-lectin and s-/DUF26 domains at the N-terminusas well as between the kinase domains representing ab-lectin/DUF26/ kinase gene duplication expressed in asingle transcript. Searching the NCBI rice genomedatabase with the Rpg1 and ABC1037 a.a. sequences(BLASTP) identiWed one tandem kinase gene withhomology to Rpg1 and ABC1037. However, it seemsunlikely that the rice tandem kinase gene and Rpg1and ABC1037 have a common tandem kinase progeni-tor. It has been reported that of the >1131 rice RLK/Pelle family members 27 have two kinase domains(Shiu et al. 2004) Given that tandem kinase genes arerare in plant and animal systems, it is interesting thatfour of the Rpg1 family gene members have tandemPKDs that appear to have arisen by two independentmeans (Fig. 6).
An interesting class of proteins with tandem kinasedomains has been characterized in animal systems.Janus Kinases (JAKs) appear to be similar to Rpg1 notonly with the presence of tandem PKDs, but also with
one PKD functional and the other a pseudokinase(Luo et al. 1997; Pellegrini and Dusanter-Fourt 1997;Nirmala et al. 2006). There is evidence that the pseudo-kinase, JH2 domain, although not catalytically active,plays a role in regulating JAK activity. This is evidentby the E695-K mutation in a JAK protein found in thehyperactive Drosophila HopT42 mutant. This residueoccurs in the JH2domain and is conserved among allJAK family members. The HopT42 mutant hyper-phosphorylates and hyperactivates D-Stat transcrip-tion factors when over-expressed in Drosophila cells(Luo et al. 1997). The non-autophosphorylating PKD1in Rpg1 also appears to be essential for function sincemutations within this domain abolish disease resistance(Nirmala et al. 2006).
The JAKs do not have their own receptor, butrather interact with a cytokine receptor (Behrmannet al. 2004). The N-terminal regions of the JAK pro-teins are variable and have been implicated in theinteraction with the cytokine receptors (Pellegrini andDusanter-Fourt 1997). Similarly Rpg1 may interactwith yet to be identiWed receptor(s).
In summary, Wve new Rpg1 gene family memberswere mapped, sequenced and characterized. Two wereclosely linked to the rust resistance loci Rpg1 andRpg5/rpg4. An apparent unequal recombination eventbetween Rpg1 and ABC1037 was characterized. Theresulting chimeric allele did not exibit the Rpg1 resis-tance speciWcity. Although our attempt at identifyingother rust resistance genes in the barley genome byhomology to Rpg1 was not immediately realized, thiswork may facilitate the identiWcation of resistancegenes identiWed in the future. To determine if the Rpg1N-terminal domain is important for resistance speciWc-ity a reciprocal recombination (i.e. Rpg1 N-terminaldomain and ABC1037 PK domains) was constructedand is being tested.
Acknowledgments This is ScientiWc Paper No. 0302-06 from theCollege of Agricultural, Human, and Natural Sciences ResearchCenter, Washington State University, Pullman, WA 99164, Pro-ject 0196. Research was supported by USDA-NRI grant # 2004-35301-14635 and the U.S. Barley Genome Project.
References
Behrmann I, Smyczek T, Heinrich P, Schmitz-Van de Leur H,Komyod W, Giese B, Muller-Newen G, Haan S, Haan C(2004) Janus kinase (JAK) subcellular localization revisited.J Biol Chem 279(34):35486–35493
Bennetzen J (2000) Transposable element contributions to plantgene and genome evolution. Plant Mol Biol 42:251–269
Brueggeman R, Rostoks N, Kudrna D, Kilian A, Han F, Chen J,Druka A, SteVenson B, Kleinhofs A (2002) The barley stemrust-resistance gene Rpg1 is a novel disease resistance gene
62

1158 Theor Appl Genet (2006) 113:1147–1158
123
with homology to receptor kinases. Proc Natl Acad Sci USA99:9328–9333
Cabrillac D, Delorme V, Garin J, RuYo-Chable V, Giranton J,Dumas C, Gaude T, Cock J (1999) The S15 self-incompati-bility haplotype in Brassica oleracea includes three S genefamily members expressed in stigmas. Plant Cell 11:971–986
Chomczynski P, Sacchi N (1987) Single-step method of RNA iso-lation by acid Guanidinium thiocyanate-phenol-chloroformextraction. Anal Biochem 162:156
Dodds P, Lawrence G, Ellis J (2001) Contrasting modes of evolu-tion acting on the complex N locus for rust resistance in Xax.Plant J 27:439–453
Druka A, Kudrna D, Han F, Kilian A, SteVenson B, Frisch D,Tompkins J, Wing R, Kleinhofs A (2000) Physical mappingof the barley stem rust resistance gene rpg4. Mol Gen Genet264:283–290
Ellis J, Lawrence G, Finnegan E, Anderson P (1995) Contrastingcomplexity of two rust resistance loci in Xax. Proc Natl AcadSci USA 92:4185–4188
Hanks S, Quinn A, Hunter T (1988) The protein kinase family:conserved features and deduced phylogeny of the catalyticdomains. Science 241:42–52
Horvath H, Rostoks N, Brueggeman R, SteVenson B, von Wett-stein D, Kleinhofs A (2003) Genetically engineered stemrust resistance in barley using the Rpg1 gene. Proc Natl AcadSci USA 100:364–369
Kim Y, Lin N, Martin G (2002) Two distinct Pseudomonas eVec-tor proteins interact with the Pto kinase and activate plantimmunity. Cell 109:589–598
Kleinhofs A, Graner A (2001) An integrated map of the barleygenome. In: Phillips RL, Vasil I (eds) DNA-based markers inplants, 2nd edn. Kluwer, Boston, pp 187–199
Kleinhofs A, Kilian A, Saghai Maroof M, Biyashev R, Hayes P,Chen F, Lapitan N, Fenwick A, Blake T, Kanazin V, Ana-niev E, Dahleen L, Kudrna D, Bollinger J, Knapp S, Liu B,Sorrells M, Heun M, Franckowiak J, HoVman D, Skadsen R,SteVenson B (1993) A molecular, isozyme and morphologi-cal map of the barley (Hordeum vulgare) genome. TheorAppl Genet 86:705–712
Luo H, Roe P, Barber D, Hanratty W, Lee S, Roberts T, D’An-drea A, Dearolf C (1997) Mutation in the Jak kinase JH2 do-main hyperactivates Drosophila and mammalian Jak-Statpathways. Mol Cell Biol 17:1562–1571
Martin G, Brommonschenkel S, Chunwongse J, Frary A, GanalM, Spivey R, Wu T, Earle E, Tanksley S (1993) Map-basedcloning of a protein kinase gene conferring disease resistancein tomato. Science 262:1432–1436
Martin G, Frary A, Wu T, Brommonschenkel S, Chunwongse J(1994) Member of the Pto gene family confers sensitivity tofenthion resulting in rapid cell death. Plant Cell 6:1543–1552
Martin G, Bogdanove A, Sessa G (2003) Understanding the func-tion of plant disease resistance proteins. Annu Rev PlantBiol 54:23–61
Nirmala J, Brueggeman R, Maier C, Clay C, Rostoks N, Kann-angara G, Wettstein D, SteVenson B, Kleinhofs A (2006)Sub-cellular localization and functions of the barley stemrust resistance receptor-like serine/threonine-speciWc pro-tein kinase Rpg1. Proc Natl Acad Sci USA 103:7518–7523
Pellegrini S, Dusanter-Fourt I (1997) The structure, regulationand function of the Janus kinases (JAKs) and the signaltransducers and activators of transcription (STATs). EurJ Biochem 248:615–63
Richter T, Ronald P (2000) The evolution of disease resistancegenes. Plant Mol Biol 42:195–204
Richter T, Pryor T, Bennetzen J, Hulbert S (1995) New rust resis-tance speciWcities associated with recombination in the Rp1complex in maize. Genetics 141:373–381
Ronald P, Albano B, Tabien R, Abenes L, Wu K, McCouch S,Tanksley S (1992) Genetics and physical analysis of the ricebacterial blight disease resistance locus, Xa21. Mol GenGenet 236:113–120
Rosenberg E, Holmes M, Tenenholz T, Abd El Raouf Khalil A,Valerie K (2003) Mapping of transcription start sites by di-rect sequencing of SMARTTM RACE products. BioTech-niques 34:482–486
Rostoks N, SteVenson B, Kleinhofs A (2004) Structure andexpression of the barley stem rust resistance gene Rpg1 mes-senger RNA. Phys Mol Plant Path 64:91–101
Saitou N Nei M (1987) The Neighbor-joining method: A newmethod for reconstructing phylogenetic trees. Mol Biol Evol4:406–425
Salmeron J., Oldroyd G, Rommens C, ScoWeld S, Kim H (1996)Tomato Prf is a member of the leucine-rich repeat class ofplant disease resistance genes and lies embedded within thePto kinase gene cluster. Cell 86:123–133
Sambrook J, Fritsch E, Maniatis T (1989) Molecular cloning: alaboratory manual, 2nd edn. Cold Spring Harbor Labora-tory, Cold Spring Harbor
Shao F, Goldstein C, Ade J, Stoutemyer M, Dixon J, Innes R(2003) Cleavage of Arabidopsis PBS1 by a bacterial type IIIeVector. Science 301:1230–1233
Shiu S-H, Bleeker A (2003) Expansion of the receptor-like ki-nase/Pelle gene family and receptor-like proteins in Arabid-opsis. Plant Physiol 132:530–543
Shiu S-H, Karlowski W, Pan R, Tzeng Y-H, Mayer K, Li W-H(2004) Comparitive analysis of the receptor-like kinase fam-ily in Arabidopsis and rice. Plant Cell 15:1220–1234
Song W, Wang G, Chen L, Kim H, Pi L, Gardner J, Wang B, Hol-sten T, Zhai W, Zhu L, Fauquet C, Ronald P (1995) A recep-tor kinase-like protein encoded by the rice disease resistancegene Xa21. Science 270:1804–1806
Song W, Pi L, Wang G, Gardner J, Holsten T, Ronald P (1997)Evolution of the Rice Xa21 disease resistance gene family.Plant Cell 9:1279–1287
Stein J, Howlett B, Boyes D, Nasrallah M, Nasrallah J (1991)Molecular cloning of a putative receptor protein kinase geneencoded at the self-incompatibility locus of Brassica olera-cea. Proc Natl Acad Sci USA 88:8816–8820
Stein J, Dixit R, Nasrallah M, Nasrallah J (1996) SRK, the stigma-speciWc S locus receptor kinase of Brassica, is targeted to theplasma membrane in transgenic tobacco. Plant Cell 8:429–445
Swiderski M, Innes R (2001) The Arabidopsis PBS1 resistancegene encodes a member of a novel protein kinase subfamily.Plant J 26:101–112
Takasaki T, Hatakeyama K, Suzuki G, Watanabe M, Isogal A,Hinata K (2000) The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 403:913–916
Yu Y, Tomkins J, Waugh R, Frisch D, Kudrna D, Kleinhofs A,Brueggeman R, Muehlbauer G, Wise R, Wing R (2000) Abacterial artiWcial chromosome library for barley (Hordeumvulgare L.) and the identiWcation of clones containing puta-tive resistance genes. Theor Appl Genet 101:1093–1099
63

CHAPTER THREE
Allele sequencing of the barley stem rust resistance gene Rpg1 identifies
regions relevant to disease resistance
Aghafakhr Mirlohi1*, Robert Brueggeman2*, Tom Drader4, Jayaveeramuthu Nirmala2, Brian J.
Steffenson3, and Andris Kleinhofs2,4
1College of Agriculture, Isfahan University of Technology, Esfahan, Iran
2Department of Crop and Soil Sciences, Washington State University, Pullman, WA
3Department of Plant Pathology, University of Minnesota, Minneapolis, MN
4School of Molecular Biosciences, Washington State University, Pullman, WA
Corresponding author: Brian Steffenson; [email protected];
As Published In
Phytopathology (2008) 98:910-918
* A. Mirlohi and R. Brueggeman contributed equally to this work
64

910 PHYTOPATHOLOGY
Genetics and Resistance
Allele Sequencing of the Barley Stem Rust Resistance Gene Rpg1 Identifies Regions Relevant to Disease Resistance
A. Mirlohi, R. Brueggeman, T. Drader, J. Nirmala, B. J. Steffenson, and A. Kleinhofs
First author: College of Agriculture, Isfahan University of Technology, Esfahan, Iran; second, third, and fourth authors: Department of Crop and Soil Sciences, Washington State University, Pullman; fifth author: Department of Plant Pathology, University of Minnesota, St. Paul; and sixth author: School of Molecular Biosciences and Department of Crop and Soil Sciences, Washington State University, Pullman.
Accepted for publication 2 April 2008.
ABSTRACT
Mirlohi, A., Brueggeman, R., Drader, T., Nirmala, J., Steffenson, B. J., and Kleinhofs, A. 2008. Allele sequencing of the barley stem rust resistance gene Rpg1 identifies regions relevant to disease resistance. Phytopathology 98:910-918.
The stem rust resistance gene Rpg1 has protected North American barley cultivars from significant yield losses for over 65 years. The remarkable durability of this gene warrants further study as to its possible origin and allelic variation. Eight Swiss barley (Hordeum vulgare) land-races and eight wild barley (H. vulgare subsp. spontaneum) accessions from diverse geographic regions were analyzed to uncover new alleles of Rpg1 and learn about its possible origin. The two germplasm groups included accessions that were resistant and susceptible to Puccinia graminis f. sp. tritici pathotype MCCF. Allele-specific primers were utilized to amplify 1 kbp overlapping fragments spanning the Rpg1 gene and sequenced if a polymerase chain reaction (PCR) fragment was generated. Variation among the PCR products revealed significant polymorphisms among these Hordeum accessions. Landraces and wild barley accessions sus-
ceptible to pathotype MCCF exhibited the highest degree of Rpg1 polymorphism. One resistant landrace (Hv672) and one resistant wild barley accession (WBDC040) yielded all seven Rpg1-specific PCR frag-ments, but only landrace Hv672 coded for an apparently functional Rpg1 as determined by comparison to previously characterized resistant and susceptible alleles and also resistance to HKHJ, a stem rust pathotype that can specifically detect Rpg1 in the presence of other resistance genes. Accessions resistant to stem rust pathotype MCCF, but completely lack-ing Rpg1-specific PCR amplification and hybridization with an Rpg1-specific probe, suggested the presence of stem rust resistant gene(s) different from Rpg1 in the Hordeum germplasm pool. Some Rpg1 alleles that retained the ability to autophosphorylate did not confer resistance to Puccinia graminis f. sp. tritici pathotype MCCF, confirming our previous observations that autophosphorylation is essential, but not sufficient for disease resistance. Thus, the RPG1 protein plays a complex role in the stem rust disease resistance-signaling pathway.
Additional keywords: functional analysis, R genes.
Stem rust (caused by the wheat stem rust fungus Puccinia
graminis f. sp. tritici) has been one of the most important patho-gens of barley (Hordeum vulgare L.) in the Upper Midwest region of the United States and Prairie Provinces of Canada over the past century. A single dominant gene (Rpg1) introgressed into barley cultivars has effectively controlled the stem rust epidemics which devastated the crop in the Northern Great Plains prior to 1942 (18). Many qualitative rust resistance genes in other cereal crops have been overcome by virulent pathotypes in a relatively short period of time (12); however, Rpg1 has been durable, protecting barley from serious stem rust losses for more than 65 years. This remarkable durability warrants further characterization of Rpg1.
Rpg1 was mapped to the short arm of barley chromosome 1(7H) and cloned from cultivar Morex by construction of high-resolution genetic and physical maps (2). Highly resistant transgenic plants were obtained by transforming the susceptible barley cultivar Golden Promise, devoid of Rpg1, with the Morex Rpg1 gene, demonstrating that the gene isolated was functional in conferring stem rust resistance (9). Rpg1 contains 14 exons in 4,466 bp of genomic sequence encoding an 837 amino acid pro-tein with two tandem kinase domains, which is a unique resis-tance gene structure (2).
Sequence analysis of Rpg1 from resistant North American barley cultivars revealed no amino acid sequence polymorphism
(2). This suggests a singularity of the Rpg1 source that can be traced to an unimproved bulked seed lot (from which the sister selections of Chevron [PI 38061] and Peatland [PI 539109] were derived) obtained by the United States Department of Agriculture from Switzerland in 1914. Another potential source of stem rust resistance in barley was the cultivar Kindred (CIho 6969), a farmer selection of a single stem rust resistant plant from a field of susceptible Wisconsin 37 barley (18). The plant from which Kindred was derived was probably an admixture because it dif-fered from Wisconsin 37 for many more traits than just stem rust resistance. Moreover, sequence analysis revealed that the Rpg1 in silico translated amino acid sequence in Kindred was identical to that of Chevron and Peatland (2). The source of Rpg1 in Kindred was likely Chevron because this cultivar had been used in the Wisconsin barley breeding program for a number of years prior to the release of Wisconsin 37.
During the map-based cloning effort of Rpg1, allele sequencing revealed three groups of stem rust susceptible barley lines (2). The first group contained susceptible rpg1 genes with 3 base pairs (GTT) inserted in the sixth exon at nucleotide position 1,546 relative to the Rpg1 sequence of Morex. The second group con-tained an Rpg1-like gene with a highly diverged 5′ region (2), and the third group had no detectable Rpg1 gene sequences. The second group, including barley line Sm89010, was recently cha-racterized as containing a recombinant Rpg1 gene with ABC1037, an Rpg1 gene family member located ≈50 kbp proximal to Rpg1 (1). Lines with this recombinant Rpg1 gene have a 5′ end highly homologous to ABC1037 and a 3′ end with high homology to Rpg1 (1). Reverse transcription (RT)-PCR and sequence analysis indicated that the Sm89010 recombinant Rpg1 gene is tran-
Corresponding author: B. Steffenson; E-mail address: [email protected]
doi:10.1094 / PHYTO-98-8-0910 © 2008 The American Phytopathological Society
65

Vol. 98, No. 8, 2008 911
scribed, but does not function in conferring stem rust resistance (to pathotype MCCF) because it and two other barley cultivars (Dicktoo and Gobernadora) containing the Sm89010-like Rpg1 gene were susceptible (2). It is possible that the Sm89010 Rpg1 gene expresses novel specificities to other stem rust pathotypes, as unequal exchange events have been suggested as a means for the evolution of plant disease resistance genes (7,15). In flax, 13 alleles at the L rust resistance locus have been reported to express 10 different rust resistance specificities. These resistance speci-ficities are presumed to have evolved through various mecha-nisms, including single base substitutions, insertion/deletions (indels), unequal recombination, and reassortment of variable re-gions within the alleles by multiple intra-genic sequence ex-change events (7).
Naturally occurring allelic diversity in plants has been sug-gested to be an important genetic component for phenotypic variation and crop improvement. It is therefore important to characterize this diversity in order to identify economically im-portant alleles and polymorphisms (3). In addition to landraces of domesticated barley (H. vulgare), the wild progenitor H. vulgare subsp. spontaneum is known to be a valuable source for alterna-tive forms of disease resistance genes for barley improvement (8,22). Stem rust resistance has been reported in wild barley, but the frequency of resistance is often low (<10%) (8,20). Given the recent threat of new pathotypes such as P. graminis f. sp. tritici TTKS (i.e., isolate Ug99) in the world, wild barley may become an important new source of stem rust resistance genes (20).
The lack of Rpg1 polymorphism among previously-tested stem rust resistant barley cultivars suggests either a highly conserved or single source for this gene. High conservation of a protein kinase disease resistance gene has been shown with the tomato Pto gene, which confers resistance to Pseudomonas syringae pv. tomato. The wild tomato species Lycopersicon pimpinellifolium is the source of the Pto gene in cultivated tomato (L. esculentum). A Pto ortholog LhirPto, isolated from another wild tomato relative (L. hirsutum), shares 97% amino acid identity with the Pto from L. pimpinellifolium (16). Even more surprising, the domesticated tomato has a highly conserved, transcriptionally active pto allele that fails to confer disease resistance (10). Since the Lycopersicon species separated presumably millions of years ago, these find-ings suggest that the PTO kinase has been highly conserved, even when it does not retain disease resistance function.
The objectives of this study were to use molecular techniques to uncover new alleles of Rpg1 and learn about its possible origin
in eight Swiss barley (H. vulgare) landraces and eight wild barley (H. vulgare subsp. spontaneum) accessions from diverse geo-graphic regions. The presence/absence of Rpg1, its primary structure, and expression were analyzed by PCR of genomic DNA and mRNA (RT-PCR), Southern blotting, sequencing, western blotting, and autophosphorylation analyses.
MATERIALS AND METHODS
Plant materials and disease phenotyping. From a germplasm collection of 74 barley landraces from Switzerland and 318 ecogeographically diverse wild barley accessions (20), 16 (eight from each group) were selected and used in this study (Table 1). Landraces from Switzerland were chosen for analysis because they are the donors of Rpg1 (i.e., Chevron and Peatland) in North American barley cultivars and may possess additional allelic diversity for the gene. Wild barley accessions also were included because of their great allelic diversity and potential for identifying the origin of Rpg1 in cultivated barley. Within each germplasm group, four resistant and four susceptible accessions to stem rust pathotype MCCF were chosen from ecogeographically diverse locations to identify functional Rpg1 variants as well as spon-taneous mutations that result in loss of function. Pathotype MCCF has been used for a number of years to select breeding lines with Rpg1, develop high resolution maps for map-based cloning of the resistance gene, and conduct functional analyses with the cloned gene (2,13,19). Recently, a comprehensive evaluation was made on the reaction of known stem rust resistance genes in barley to diverse pathotypes of P. graminis f. sp. tritici and isolates of P. graminis f. sp. secalis (the rye stem rust fungus) (21). This study revealed that P. graminis f. sp. tritici pathotype HKHJ was more effective than MCCF for detecting Rpg1 in the presence of several other known stem rust resistance genes (21). Thus, the germplasm initially selected for this study based on reaction to pathotype MCCF was subsequently evaluated to pathotype HKHJ to further resolve the functionality of Rpg1. Cultivars Morex (with Rpg1) and Steptoe (without any known resistance genes) also were included as the resistant and susceptible controls, respectively. Plants were grown and assayed for their reaction to stem rust as previously described (19).
DNA and RNA were isolated from a single plant of each of the 16 selected accessions based initially on their stem rust pheno-type. The plants were grown in plastic pots containing potting mix in a growth chamber under cool fluorescent lights with a 16 h
TABLE 1. Reaction of Swiss landrace and wild barley accessions to Puccinia graminis f. sp. tritici pathotypes MCCF and HKHJ at the seedling stage
Pathotype MCCF Pathotype HKHJ
Line Origin IT rangea General reaction IT rangea General reaction
Swiss Hv584 Fuormis, Switzerland 0;1 to 10; Resistant 3– to 3 Susceptible Swiss Hv587 Platta, Switzerland 0;1 to 21 Resistant to moderately resistant 3– to 3 Susceptible Swiss Hv619 Fetan, Switzerland 10; Resistant 3– to 33+ Susceptible Swiss Hv672 Surava, Switzerland 10; to 12 Resistant 10; to 21 Resistant to moderately resistant Swiss Hv489 Trans, Switzerland 3 Susceptible 3 to 33+ Susceptible Swiss Hv492 Scheid, Switzerland 3 to 33+ Susceptible 3– to 3 Susceptible Swiss Hv611 Plan, Switzerland 3– to 3 Susceptible 3– to 3 Susceptible Swiss Hv645 Scuol, Switzerland 3– to 3 Susceptible 3– to 3 Susceptible WBDC040 Haifa, Israel 0;1 to 21 Resistant to moderately resistant 3– to 3 Susceptible WBDC214 Samarkand, Uzbekistan 0;1 to 213– Resistant to moderately resistant 3 Susceptible WBDC220 Chimkent, Kazakhstan 10;2 to 213– Resistant to moderately resistant 3 to 33+ Susceptible WBDC225 Dushanbe, Tajikistan 0;1 to 21 Resistant to moderately resistant 3 to 3+ Susceptible WBDC019 West Azerbaijan, Iran 3 to 33+ Susceptible 33+ Susceptible WBDC160 Sweida, Syria 3 to 3+3 Susceptible 3 to 3+ Susceptible WBDC269 Lebanon 3-2 to 33+ Moderately susceptible to susceptible 3– to 3 Susceptible WBDC323 Ahalskii, Turkmenistan 3 to 33+ Susceptible 3 to 3+ Susceptible Morexb Minnesota, USA 0;1 to 213– Resistant to moderately resistant 12 to 21 Resistant to moderately resistant Steptoeb Washington, USA 3 to 33+ Susceptible 3 to 33+ Susceptible
a IT = infection types are based on a 0 to 4 scale, + and – denote more or less sporulation of uredinia, respectively. ITs of 0; 1, 2, and combinations thereof are indicative of resistance, whereas ITs of 3, 4 and combinations thereof are indicative of susceptibility (21).
b Morex (with Rpg1) and Steptoe (without any known stem rust resistance gene) are the resistant and susceptible controls, respectively.
66

912 PHYTOPATHOLOGY
photoperiod and day/night temperature of 21 ± 1°C/18 ± 1°C. For protein analysis by western blot and autophosphorylation, a single plant from the same seed source was grown under the above conditions.
DNA and RNA isolation. Genomic DNA for Southern hy-bridization was isolated from young barley leaves with a modified CTAB method (11). For PCR amplification, genomic DNA was isolated from young barley leaves according to Edwards et al. (6). Total RNA was isolated from young seedling leaves using Trizol reagent (4). Integrity and quantity of the RNA samples were checked by 1× TBE (0.089 M Tris-borate, 0.089 M boric acid, and 0.002 M EDTA) agarose gel electrophoresis.
PCR and RT-PCR. PCR primers were designed from cultivar Morex and line Sm89010 Rpg1 allele sequences (GenBank acces-sion numbers AF509748 and AF509762, respectively) to amplify approximately 1-kbp overlapping fragments covering the entire Rpg1 gene (Fig. 1 and Table 2). Primer set 228F6/228R6 ampli-fied a fragment from both cultivars Morex and Sm89010; how-ever, the Sm89010 fragment is of higher molecular weight (Table 2). Primer combination 37F2/228R6 amplified only the Sm89010 Rpg1 allele and resulted in a 1,678-bp band. DGK-F1 and DGSex2 primers are internal to 37F2/228R6 and were used only in the sequencing reaction in order to cover the gap between the forward 37F2 and reverse 228R6 primers. PCR reactions and analyses were as previously described (2).
Approximately 1µg of total RNA was used for oligo dT-primed first-strand cDNA synthesis using the Reverse Transcription Sys-tem (Promega, Madison, WI) according to the manufacturer’s recommended conditions followed by PCR reactions (2). Primers EX3cw2/228R6 and 37-F2/228R7 were used for RT-PCR to amplify Morex or Sm89010 Rpg1-specific cDNA, respectively.
Southern blot analysis. Southern blotting and hybridization were as previously described (11). Five micrograms of HindIII-digested barley genomic DNA was separated by agarose gel electrophoresis, blotted onto nylon membranes, and hybridized with the Rpg1-specific DNA probe RSB228 (2).
Sequencing. PCR fragments were eluted from the agarose gel, purified, concentrated by ethanol precipitation, and used as tem-plates for sequencing reactions. Sequencing reactions were per-formed using the Big-Dye Terminator kit (Applied Biosystems, Foster City, CA) and sequenced at the Bioanalytical Center, Washington State University, Pullman, WA. Sequences were ana-lyzed and contigs assembled using ContigExpress from the Vector NTI software package.
cDNA cloning and sequencing. Oligo dT-derived cDNA from Morex and WBDC040 was used as the template to amplify the ninth intron/exon junction of Rpg1. RT-PCR products were cloned in the pGEM-T vector (Promega) following the manufacturer’s protocol. Clones were confirmed by restriction digestion with EagI (New England Biolabs, Beverly, MA) and NcoI (Fermentas, Hanover, MD) and sequenced using T7 and SP6 universal primers.
Protein analysis. Polyclonal antibody was raised in New Zealand white rabbits at the Washington State University animal facility against a recombinant His-RPG1 protein purified from Pichia pastoris (13). Ribi adjuvant was used according to the manufacturer’s recommendation (Corixa, Hamilton, MO). West-ern blotting and autophosphorylation assays were performed as previously described (13).
Rapid amplification of cDNA 3′ ends (3′ RACE). 3′ RACE was carried out using the FirstChoice RLM-RACE Kit (Ambion, Austin, TX) and the 3′ RACE System (Invitrogen, Frederick,
Fig. 1. Schematic representation of the Rpg1 gene structure. A, The relative position of the seven cultivar Morex Rpg1-specific primer pairs and overlappingfragments generated in relation to the Rpg1 gene structure. Introns are shown in gray and exons in black shading. The 5′ and 3′ untranslated transcribed regions (UTRs) are indicated. B, Black bars indicate fragments amplified using the Morex-specific set of primer pairs, hatched bars indicate fragments amplified using the barley line Sm89010 allele-specific primers, and white bars indicates no amplification. The R or S designation within parentheses indicate resistance or suscep-tibility to Puccinia graminis f. sp. tritici pathotypes MCCF/HKHJ, respectively.
67

Vol. 98, No. 8, 2008 913
MD) under the manufacturer’s recommended standard conditions. The nested gene-specific primers used for both systems were 228-F3 and 228-F4 as previously described in Brueggeman et al. (2).
RESULTS
Stem rust reaction. The controls reacted as expected with Morex exhibiting resistant to moderately resistant infection types and Steptoe susceptible infection types to pathotypes MCCF and HKHJ (Table 1). Hv584, Hv587, Hv619, and Hv672 were among the most resistant accessions found to P. graminis f. sp. tritici pathotype MCCF in the Swiss landrace germplasm, giving infec-tion types ranging from 0;1 to 21 (Table 1). In contrast, Hv489, Hv492, Hv611, and Hv645 were among the most susceptible accessions found, giving infection types of 3– to 33+. Resistance to wheat stem rust in wild barley is fairly rare (8,20). The resistant wild barley accessions used in this study (WBDC040, WBDC214, WBDC220, and WBDC225) were selected from a previous pre-liminary study examining a large wild barley germplasm collec-tion (20) and were among the most resistant found to pathotype MCCF, giving infection types ranging from 0;1 to 213–. Acces-sions WBDC019, WBDC160, WBDC269, and WBDC323 were representative of the most susceptible accessions found and ex-
hibited infection types ranging from 3–2 to 3+3. All Swiss land-races and wild barley accessions also were evaluated to HKHJ, a pathotype that can effectively detect Rpg1 in the presence of other stem rust resistance genes (21). The Swiss landrace Hv672 was the only accession found with resistance (IT range of 10; to 21) to pathotype HKHJ. This suggests that it may be the only accession with a functional Rpg1 gene (Table 1).
Lines with an apparently intact Rpg1 gene. Seven primer pairs, designed to amplify the entire Rpg1 coding region, were used to amplify Rpg1 genes from all accessions. All expected DNA fragments were successfully amplified from Swiss Hv672, WBDC040, and WBDC323 (Fig. 1B), suggesting the presence of an intact Rpg1 gene in these genotypes. Southern blots of ge-nomic DNA digested with HindIII and hybridized with the Rpg1-specific probe RSB228 confirmed these results (Fig. 2). The hy-bridizing band was the same size (approximately 4.5 kbp) as observed in cultivar Morex and the wild barley accession OSU6 (2) containing an intact Rpg1 gene. These data were not in agree-ment with the stem rust phenotypes as only Hv672 was resistant to both pathotypes MCCF and HKHJ. OSU6 was previously characterized as resistant to stem rust pathotype MCCF, but multi-ple subsequent assays with both pathotypes MCCF and HKHJ on the selected plants used in the molecular analysis showed that
TABLE 2. Primers used to amplify Morex-like and Sm89010-like gene fragments for genotyping and sequencing Location and product sizeb
Fragment namea Primer name Primer sequence Sm89010 Morex
1 Sm 37F2/228R6 cttccactttcctctctctcagatc/cggaaggcgctcaaagcagtctc –52–1,626/1,678 bp None 1 228F11/228R11 tgcagcagtactctgcccacttgag/gggatgggtggctttgttgacaac None –135–736/870 bp 2 228F6/228R6 cggaaggcgctcaaagcagtctc/cggaaggcgctcaaagcagtctc 209–1,624/1,414 bp 130–1,090/959 bp 3 228F1/228R1 gccggggctggacgatgaggaattc/gaactcgaatgcaaactcccttgtc 1,420–2,506/1,085 bp 884–1,967/1,082 bp 4 228F2/228R2 tgtgtagacattccacccgaagag/catctgtcccaatcaagtttgcagga 2,151–3,247/1,095 bp 1,612–2,708/1,095 bp 5 228F3/228R3 catgcaacgaggaagccttgacgac/acccgatctactggtgatctct 2,893–3,980/1,086 bp 2,353–3,440/1,086 bp 6 228F4/228R4 ggactactccaactatgcacacac/gcatcgatgatatgatcggcagctgac 3,629–4,561/931 bp 3,090–4,020/929 bp 7 228F12/228R12 agatttcccaggctacttcttccag/gaccctccttatttgtctctgcttc 4,349–5,100/750 bp 3,810–4,561/750 bp RT-PCR1 EX3cw2/228R6 gccggtgtactatccctttc/cggaaggcgctcaaagcagtctc None 465–1,088/396 bp RT-PCR2 37-F2/228R7 cttccactttcctctctctcagatc/ttactgatgtaaagaccaccgtata 52–2,065/879 bp None Seq 1 Sm DGK-F1 cctcggatgctatcctcagcatgac 875 NAc Seq 1 Sm DGSex2 tggggtaatgagaaaggtgtgatag 953 NAc Ubiquitin Hvu_Ubi1_cw1/Hvu_Ubi1_ccw1 gccgcaccctcgccgactac/cggcgttggggcactccttc 219 bp 219 bp
a Fragments 1 to 7 primers were used to amplify the Morex Rpg1 coding regions as shown in Figure 1. Fragment 1 Sm primers amplified fragment 1 only from theSm89010-like Rpg1 allele. Fragment 2 primers amplified fragment 2 from both Morex and Sm type Rpg1 alleles, but a higher molecular weight fragment fromthe Sm89010-like allele. Reverse transcription-polymerase chain reaction (RT-PCR) 1 and 2 primers were used to amplify a specific Rpg1 mRNA fragment fromeither Morex or Sm89010, respectively. Seq1Sm primers DGK-F1 and DGSex2 are internal to the location of primers 37F2 and 228R6, and were used to coverthe sequencing gap in fragment 1Sm, which was approximately 1.7 kbp.
b Bases upstream of the start codon are designated with minus (–) values. The product size is given in base pairs and the location indicated by base pair position inMorex or Sm89010 with the first base in the ATG start codon designated as 1. Ubiquitin primers were used to amplify a 219 bp fragment of the ubiquitin 1 genefrom all cDNAs as shown in Figure 4.
c NA, not applicable.
Fig. 2. Southern blot analysis of Hordeum accessions. Genomic DNA of each line was digested with HindIII and hybridized with the Rpg1-specific probe RSB228. The Morex-specific Rpg1 band is approximately 4.5 kbp. The Sm89010-specific Rpg1 band is approximately 7.5 kbp (not shown) and of similar size to fragments hybridizing with DNA of Swiss landraces Hv619, Hv489, and Hv645. Harrington does not contain Rpg1. The R or S designation within parentheses indicate resistance or susceptibility to Puccinia graminis f. sp. tritici pathotypes MCCF/HKHJ, respectively.
68
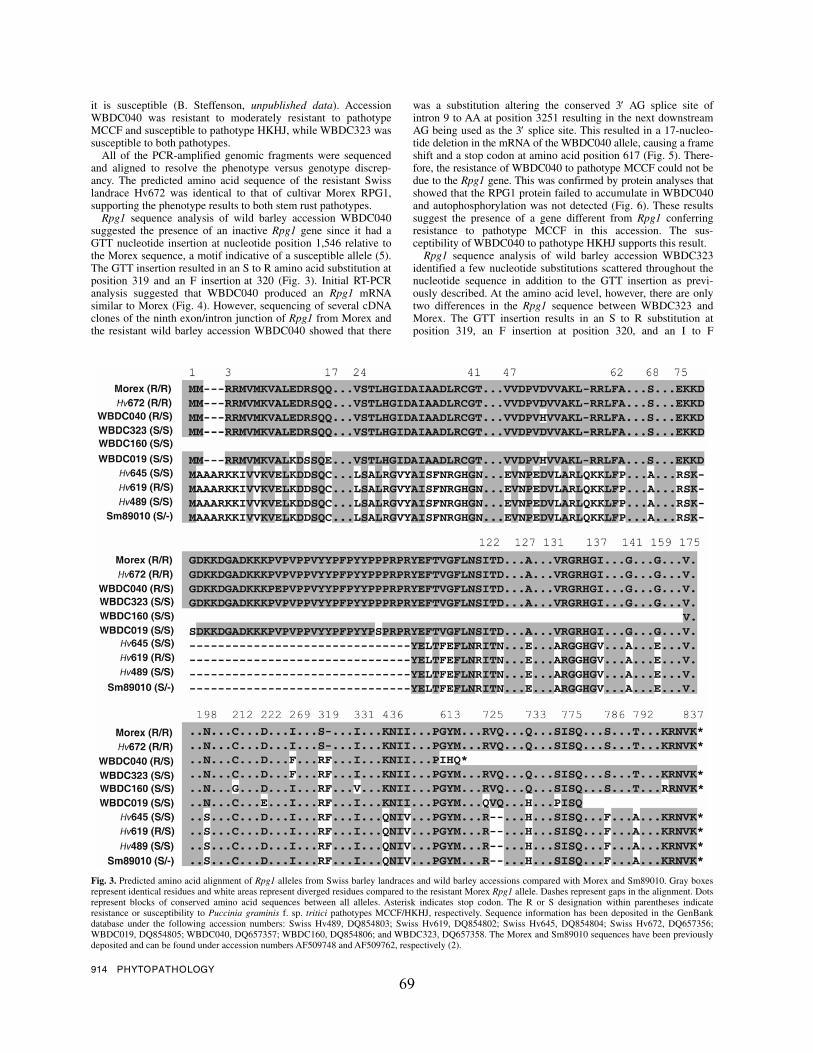
914 PHYTOPATHOLOGY
it is susceptible (B. Steffenson, unpublished data). Accession WBDC040 was resistant to moderately resistant to pathotype MCCF and susceptible to pathotype HKHJ, while WBDC323 was susceptible to both pathotypes.
All of the PCR-amplified genomic fragments were sequenced and aligned to resolve the phenotype versus genotype discrep-ancy. The predicted amino acid sequence of the resistant Swiss landrace Hv672 was identical to that of cultivar Morex RPG1, supporting the phenotype results to both stem rust pathotypes.
Rpg1 sequence analysis of wild barley accession WBDC040 suggested the presence of an inactive Rpg1 gene since it had a GTT nucleotide insertion at nucleotide position 1,546 relative to the Morex sequence, a motif indicative of a susceptible allele (5). The GTT insertion resulted in an S to R amino acid substitution at position 319 and an F insertion at 320 (Fig. 3). Initial RT-PCR analysis suggested that WBDC040 produced an Rpg1 mRNA similar to Morex (Fig. 4). However, sequencing of several cDNA clones of the ninth exon/intron junction of Rpg1 from Morex and the resistant wild barley accession WBDC040 showed that there
was a substitution altering the conserved 3′ AG splice site of intron 9 to AA at position 3251 resulting in the next downstream AG being used as the 3′ splice site. This resulted in a 17-nucleo-tide deletion in the mRNA of the WBDC040 allele, causing a frame shift and a stop codon at amino acid position 617 (Fig. 5). There-fore, the resistance of WBDC040 to pathotype MCCF could not be due to the Rpg1 gene. This was confirmed by protein analyses that showed that the RPG1 protein failed to accumulate in WBDC040 and autophosphorylation was not detected (Fig. 6). These results suggest the presence of a gene different from Rpg1 conferring resistance to pathotype MCCF in this accession. The sus-ceptibility of WBDC040 to pathotype HKHJ supports this result.
Rpg1 sequence analysis of wild barley accession WBDC323 identified a few nucleotide substitutions scattered throughout the nucleotide sequence in addition to the GTT insertion as previ-ously described. At the amino acid level, however, there are only two differences in the Rpg1 sequence between WBDC323 and Morex. The GTT insertion results in an S to R substitution at position 319, an F insertion at position 320, and an I to F
Fig. 3. Predicted amino acid alignment of Rpg1 alleles from Swiss barley landraces and wild barley accessions compared with Morex and Sm89010. Gray boxesrepresent identical residues and white areas represent diverged residues compared to the resistant Morex Rpg1 allele. Dashes represent gaps in the alignment. Dots represent blocks of conserved amino acid sequences between all alleles. Asterisk indicates stop codon. The R or S designation within parentheses indicate resistance or susceptibility to Puccinia graminis f. sp. tritici pathotypes MCCF/HKHJ, respectively. Sequence information has been deposited in the GenBankdatabase under the following accession numbers: Swiss Hv489, DQ854803; Swiss Hv619, DQ854802; Swiss Hv645, DQ854804; Swiss Hv672, DQ657356; WBDC019, DQ854805; WBDC040, DQ657357; WBDC160, DQ854806; and WBDC323, DQ657358. The Morex and Sm89010 sequences have been previouslydeposited and can be found under accession numbers AF509748 and AF509762, respectively (2).
69

Vol. 98, No. 8, 2008 915
substitution at position 269 (Fig. 3). RT-PCR analysis showed that the Rpg1 gene was expressed in this accession at the mRNA level (Fig. 4), and western blot and autophosphorylation assays showed that the RPG1 protein is present and capable of autophos-phorylation (Fig. 6). Though the GTT insertion and/or the I to F
substitution did not affect the autophosphorylation activity, the protein was not functional in conferring resistance to either pathotype MCCF or HKHJ.
Lines with an apparent unequal recombination. We previ-ously described an apparent unequal recombination event in
Fig. 6. Western blot and autophosphorylation assays. A, RPG1 protein visualized with an RPG1-specific antibody. WBDC040 and GP (cultivar Golden Promise, which does not have the Rpg1 gene) failed to produce a detectable RPG1 protein. B, Autophosphorylation assay corresponding to RPG1 western blot. WBDC040and GP did not autophosphorylate. The K2 (kinase domain 2) mutant also failed to autophosphorylate, while the K1 (kinase domain 1) mutant autophosphorylatedas previously reported (13). C, Coomassie blue staining of total protein to demonstrate protein loading in each lane.
Fig. 5. Sequence comparison of Morex and WBDC040 Rpg1 intron 9/exon 10 junction and predicted amino acid sequence. The WBDC040 cDNA sequence isbased on three cloned reverse transcription-polymerase chain reaction generated fragments. Note the G to A substitution at the 3′ end of intron 9 (white A) in the WBDC040 genomic sequence and the subsequent use of the next AG pair (AG bold) for splicing of the mRNA. The resulting cDNA is missing 17 nucleotides from exon 10 causing a frame shift and a stop codon at position 617. The truncated translation product failed to accumulate a protein detectable by the RPG1-specific antibody.
Fig. 4. Reverse transcription-polymerase chain reaction showing Morex-like or Sm89010-like bands. A, Oligo dT-primed cDNA derived from total RNA was amplified with either Morex-like Rpg1-specific primers (right lane designated M) or Sm89010-like Rpg1-specific primers (left lane designated S). The accessions used for cDNA isolation are listed above. The Morex band is approximately 0.4 kbp and the Sm89010 band is approximately 0.9 kbp. Lines devoid of Rpg1 did not yield a band with either set of primers. B, Oligo dT-primed cDNA of each was amplified using ubiquitin-specific primers to demonstrate loading and RNA quality.
70

916 PHYTOPATHOLOGY
barley line Sm89010, resulting in a chimeric gene between ABC1037, a closely linked Rpg1 homologue, and Rpg1 (1). In the Swiss landraces Hv619, Hv489, and Hv645, amplification of fragments 3 to 7 with the Morex-specific primers was successful and of the expected size, but fragment 1 failed to amplify (Fig. 1). Fragment 2 amplified, but was larger than from Morex (1.6 kbp versus 1.0 kbp). This was similar to the situation previously observed with the Sm89010 allele. Successful amplification of fragment 1 using Sm89010 recombinant allele-specific primers confirmed the presence of an Sm89010-like recombinant allele in these lines (2). Southern blot hybridization of HindIII digested Hv619, Hv489, and Hv645 DNA yielded a band similar to that previously observed with Sm89010 (1) and with a higher mo-lecular weight than from the lines containing the Morex-like allele (Fig. 2). RT-PCR amplification using primers specific to Morex or Sm89010 alleles resulted only in a band similar to Sm89010 in these lines and no amplification with Morex-specific primers (Fig. 4). Nucleic acid sequence in silico translation predicted amino acid identity among the three Swiss landraces Hv619, Hv489, and Hv645 and with the Sm89010 Rpg1-like recombinant gene (Fig. 3). At the nucleotide level, there were only three differences between the entire Sm89010 Rpg1 gene sequence and the sequences of the three Swiss landraces (2). There was a G to A substitution at position 2,183 as well as single nucleotide deletions of A at 2,199 and C at 4,713, with numbering based on the Sm89010 Rpg1 sequence (data not shown). These single nucleotide polymorphisms (SNPs) did not change the amino acid sequence since the G 2,183 A substitution was in the third nucleotide in the codon and A 2,199 and C 4,713 deletions were in introns. The recombinant Rpg1-like gene is not expected to provide stem rust resistance as previously demonstrated with the Sm89010 line (1). Landraces Hv489 and Hv645 were indeed susceptible to stem rust pathotypes MCCF and HKHJ as ex-pected; however, Hv619 was resistant to pathotype MCCF, sug-gesting the presence of a gene different from Rpg1 conferring the resistance. The lack of a functional Rpg1 gene in Hv619 also was confirmed by susceptibility to pathotype HKHJ. All three lines with the recombinant type Rpg1 allele produced an RPG1 anti-body positive protein and autophosphorylated (Fig. 6).
Lines with apparent 3′ and 5′ deletions in the Rpg1 genes. The wild barley accessions WBDC019 and WBDC160 were susceptible to stem rust pathotypes MCCF and HKHJ and had apparent 3′ and 5′ deletions in the Rpg1-like genes. For WBDC019, fragment 7 (the most 3′ Rpg1 PCR fragment) failed to amplify and for WBDC160, fragments 1 and 2 (the most 5′ Rpg1 PCR fragments) failed to amplify with either the Morex- or Sm89010-specific primers (Fig. 1B). Southern blot analysis showed that both lines had Rpg1 probe-positive hybridizing fragments; however, accession WBDC019 showed a very high molecular weight fragment, while the WBDC160 fragment was about the same size as Morex (Fig. 2). Since WBDC019, the only accession lacking amplification for fragment 7, had a unique high molecular weight fragment hybridized, we assumed that there might be a large insertion in this region. Additional efforts using ampli-fication conditions for long range PCR (≈10 kbp fragments) and fragment 7 primers (228F12/228R12) did not result in amplifi-cation. Long range PCR also was applied for amplification of fragment 1 and 2 from WBDC160 using primers 228F11/228R11 and 228F6/228R6, respectively, but without success. RT-PCR produced a band similar to Morex (Fig. 4), indicating that the gene is expressed and suggesting the possibility of an mRNA similar to the Morex-like allele. The RT-PCR was carried out with primers that amplified the internal Morex-like region of the gene. Therefore, the bands are approximately the same size as the Morex Rpg1 mRNA.
The amplified genomic DNA fragments 1 to 6 from WBDC019 and fragments 3 to 7 from WBDC160 were sequenced and aligned with the other RPG1 derived amino acid sequences (Fig.
3). The susceptible Rpg1 allele from WBDC019 had diverged, including the GTT insertion, but still was very similar to the Morex RPG1 protein sequence until the C terminus where it could no longer be amplified and recognized as a Morex Rpg1 homologue. All attempts at extending the sequence with 3′ RACE failed.
The in silico translated sequence of the susceptible wild barley WBDC160 RPG1 starts at amino acid position 162 and is fairly similar to the Morex sequence from then on (Fig. 3). This gene also has the GTT insertion resulting in substitution of S 319 R and insertion of F 320. Both lines produced RPG1 antibody-positive protein and autophosphorylated (Fig. 6).
Rpg1 gene PCR amplification deficient lines. Of the 16 lines analyzed, 8 failed to produce any amplification products with either Morex-specific or Sm89010-specific Rpg1 primers (Fig. 1B). The eight lines were evenly divided between the germplasm groups with four Swiss landraces and four wild barley accessions failing to yield any amplification products. To confirm the absence of Rpg1 alleles in the eight genotypes, an additional 27 combinations of primers designed from the Morex and Sm89010 nucleotide sequences were tested on this group. No amplification products were obtained with these sets of primers, confirming the absence of a Morex-like or Sm89010-like Rpg1 in these lines (data not shown). In accordance with the PCR amplification re-sults, no hybridization to the RSB228 probe was observed with DNA from lines lacking PCR amplification (Fig. 2). To further confirm the lack of an Rpg1-like gene, two Swiss landraces resistant (Hv584 and Hv587) and one wild barley accession susceptible (WBDC269) to pathotype MCCF from this group were included in RT-PCR analysis. No amplification products were obtained using Morex or Sm89010 Rpg1 primers (Fig. 4). Five of these accessions (Hv584, Hv587, WBDC214, WBDC220, and WBDC225) were resistant to pathotype MCCF and three (Hv492, Hv611, and WBDC269) were susceptible. All of these accessions were susceptible to pathotype HKHJ. These results clearly indicate the presence of a stem rust resistance gene(s) different from Rpg1 in these Hordeum accessions.
DISCUSSION
To investigate the diversity and possible origin of the barley stem rust resistance gene Rpg1, we analyzed two subsets of H. vulgare and H. vulgare subsp. spontaneum germplasm: one com-prised of eight Swiss landraces and the second of eight wild bar-ley accessions from diverse geographic regions. Accessions resis-tant and susceptible to stem rust pathotype MCCF were chosen in order to identify functional Rpg1 variants as well as spontaneous mutations that result in loss of function (Table 1). Additionally, pathotype HKHJ also was used in this study because it is highly effective for detecting Rpg1 in the presence of other stem rust resistance genes.
The Swiss landraces used in this study were a potential source of stem rust resistant germplasm and possibly new Rpg1 alleles. However, only one accession (Hv672) with an intact Rpg1 gene was found to confer resistance to both P. graminis f. sp. tritici pathotypes MCCF and HKHJ (Fig. 1; Table 1). The gene in Hv672 was identical in predicted amino acid sequence to the cultivar Morex RPG1 protein (Fig. 3). Thus, Swiss landraces appear to be the sole source for Rpg1 as both cultivars Chevron and Peatland from Vorrenwald Eich in Canton Lucerne (18) and accession Hv672 from Surava in Canton Graubünden (B. Steffenson, unpublished data) have intact Rpg1 genes that encode a protein with amino acid sequences identical to Morex, and are all functional in conferring resistance to pathotype HKHJ. It is interesting to note that Hv672 (together with its duplicate selection Hv671) was the only accession among 74 Swiss landraces tested that exhibited resistance to pathotype HKHJ (B. Steffenson, unpublished data). Thus, the presence of Rpg1 also is
71

Vol. 98, No. 8, 2008 917
apparently rare in Swiss landraces. The Rpg1-derived amino acid sequence in Kindred was previously shown to be identical to that of Chevron and Peatland (2). As mentioned previously, the donor of Rpg1 in Kindred was likely Chevron.
Wild barley is known to be a rich source of different disease resistance alleles for cultivated barley improvement (8). However, to stem rust, the frequency of resistance is relatively low, even in a large (318 accessions) and diverse collection (20). To identify the possible source of Rpg1 in cultivated barley, eight ecogeo-graphically diverse wild barley accessions that were resistant or susceptible to stem rust pathotype MCCF were selected for this study. Analysis of the Rpg1 alleles from the eight wild barley accessions failed to identify any with an intact and functional Rpg1 gene as was found for the Swiss landrace accession Hv672 (Fig. 1 and Table 1). Surprisingly, most of the resistant wild barley accessions contained either a defective Rpg1 gene or did not contain a recognizable Rpg1 gene. This result indicates that other stem rust resistance gene(s) effective against P. graminis f. sp. tritici pathotype MCCF exist in wild barley. The fact that defective Rpg1 alleles were found among the wild barley accessions indicates that the gene exists or has existed in these populations.
A curious GTT indel occurs in the Rpg1 gene of several Swiss landraces and wild barley accessions. This indel results in an S 319 R substitution and insertion of F at position 320 (Fig. 3). In the wild barley accession WBDC323, the only other amino acid change relative to the Morex RPG1 sequence is an I 269 F sub-stitution resulting in a stem rust susceptible phenotype. We have previously observed the GTT indel in the wild barley accession OSU6 (PBI004-7-0-015 accession no. 8321) (2). OSU6 was previously characterized as resistant to stem rust pathotype MCCF, but multiple subsequent assays with both pathotypes MCCF and HKHJ on the selected plants used in the molecular analysis showed that it is susceptible (B. Steffenson, unpublished data). Except for the amino acid changes resulting from the GTT indel, OSU6 does not have any other differences in the RPG1 amino acid sequence compared to the Morex RPG1 sequence. The S to RF change due to the GTT indel appears in seven out of the eight accessions containing an Rpg1-like gene analyzed here and thus is very common (Fig. 3). These seven accessions (Hv619, Hv489, Hv645, WBDC040, WBDC019, WBDC160, and WBDC323) do not show a consistent pattern of resistance or susceptibility to the stem rust pathotype MCCF, but all were susceptible to HKHJ, the best pathotype for indicating the functionality of Rpg1. Thus, it seems that the GTT insertion affects Rpg1-mediated stem rust resistance as indicated by reaction to pathotype HKHJ, but may not affect resistance conferred by other genes, i.e., those con-ferring resistance to pathotype MCCF. The potential role of the GTT insertion is unknown. It occurs at position 319 in the RPG1 amino acid sequence, placing it near the end of the kinase 1 do-main. This domain is not essential for RPG1 autophosphorylation activity, but plays an important, yet undefined role in the overall stem rust resistance reaction (13). The fact that the RPG1 protein with the GTT insertion is active in autophosphorylation suggests that the kinase 2 domain, which is essential for autophosphoryla-tion and disease resistance, is intact and the gene is expressed and may retain some unknown function (Fig. 6).
A set of primers that can differentiate barleys with and without Rpg1 was developed based on the gene’s nucleotide sequence and the GTT insertion (5). The authors tested 42 different barley cultivars and showed that the primers correctly identified the phenotype in all cases, but one. The one exception was cultivar Maud, originally classified as resistant (with a question mark), but grouped with susceptible lines that do not contain an Rpg1 gene (5). We repeatedly tested Maud at both the seedling and adult plant stages and found it to be susceptible to stem rust pathotype MCCF (B. Steffenson, unpublished data). These results support our conclusion that the GTT indel is associated with a nonfunc-
tional Rpg1 gene, at least in terms of P. graminis f. sp. tritici resistance to pathotypes MCCF and HKHJ.
Another interesting group of Rpg1 alleles are those represented by an apparent unequal recombination between the Rpg1 gene and a closely linked paralog designated ABC1037 (1). This recombination was observed only in the Swiss landraces and not in any of the wild barley lines tested. The gene is expressed at the mRNA level (Fig. 4). Moreover, an RPG1 antibody positive protein is produced and is active in autophosphorylation (Fig. 6). The function, if any, of this recombinant allele is unknown, but we speculate that it may involve recognition of a putative AvrRpg1 effector of the pathogen. Recombination among homo-logous disease resistance genes has been observed and can result in the creation of novel resistance phenotypes (14,17).
The two wild barley accessions WBDC019 and WBDC160 with apparent 3′ and 5′, respectively, incomplete Rpg1 genes were susceptible to both stem rust pathotypes as expected (Fig. 1 and Table 1). Both genes, however, are expressed at the mRNA level (Fig. 4) and produce an RPG1 antibody positive protein that undergoes autophosphorylation (Fig. 6). The genomic nucleotide sequence of the Rpg1 gene in these lines is apparently similar enough to the Morex Rpg1 gene in the interior regions to produce mRNA and protein with Rpg1 properties and an intact kinase domain 2 that is active in autophosphorylation. The failure to amplify Rpg1 fragment 7 from WBDC019 and fragments 1 and 2 from WBDC160 suggest that those regions of the Rpg1 gene may be deleted, harbor a large insertion, or are highly diverged. A 19 nucleotide insertion and 11 nucleotide deletion in intron 12, plus a 17 nucleotide deletion at the 3′ end and 25 nucleotide deletion at the 5′ end of exon 13 of the WBDC019 allele, indicated high divergence in this region of the gene. However, these changes apparently are far enough downstream of the kinase 2 domain and do not affect the protein’s autophosphorylation activity (Fig. 6). The reason for the extensive divergence of the sequence at the 3′ end while the rest of the Rpg1 gene primary structure is mostly conserved is unknown.
The origin of the widely used and durable Rpg1 gene bred into North American barley cultivars remains a mystery since we have yet to discover a wild barley accession containing an intact and functional Rpg1 gene. Although the number of wild barley acces-sions examined by complete sequencing to date is small, they nevertheless represent some of the most stem rust resistant germ-plasm found to date in extensive evaluations of H. vulgare sp. spontaneum germplasm (8,20). In wild barley, we repeatedly found an Rpg1 gene with a very similar primary gene structure, but containing a GTT insertion that apparently renders the gene nonfunctional in providing stem rust resistance. Is this the progenitor resistance gene that later developed specificity for the stem rust pathotypes MCCF/HKHJ by deletion of the GTT triplet in the cultivated species? A similar, but inversed situation has been described with the Pto disease resistance gene (10). In that case, the wild tomato relative L. pimpinellifolium contributed the functional resistance gene, but the domesticated L. esculentum has maintained highly related and expressed pto and fen alleles. These alleles encode predicted proteins of 87 and 98% identity with the PTO and FEN kinases and are active in in vitro auto-phosphorylation, but not functional in disease resistance. The answer to the question of the Rpg1 origin will require a more extensive search of wild barley germplasm for a potential Rpg1 donor.
Eight of the 16 Hordeum accessions analyzed in this study did not have an Rpg1-like gene recognizable by multiple PCR primers or Southern blot analysis, yet five exhibited some resis-tance to pathotype MCCF (Fig. 1 and Table 1). These results clearly indicate the presence of a gene(s) different from Rpg1 conferring resistance to stem rust pathotype MCCF. The observed resistance could be due to the rpg4/Rpg5 complex as there is some evidence that one or both of these genes confers resistance to pathotype
72

918 PHYTOPATHOLOGY
MCCF in addition to other pathotypes (21)(B. Steffenson, unpub-lished data). Additional studies are underway to elucidate the genetics of stem rust resistance in these accessions.
In summary, Rpg1 alleles with a primary structure different from the one found in cultivar Morex, but apparently functional in conferring stem rust resistance to pathotypes MCCF and HKHJ were not discovered in this study. Moreover, we failed to find a progenitor Rpg1 gene among the wild barley accessions ex-amined. A common potential progenitor gene has a GTT indel that apparently results in failure to provide resistance to pathotype MCCF. Further, examples of an apparent unequal recombination were discovered, but only in the domesticated Hordeum group (i.e., Swiss landraces) suggesting that this may be a relatively recent event. We have previously shown that RPG1 autophos-phorylation is essential, but alone is not sufficient for conferring stem rust resistance (13). The observation that all the lines ex-amined in this study, except WBDC040, were active in autophos-phorylation reinforces the previous data that it is not sufficient for resistance. Thus, RPG1 plays a complex role in the stem rust disease resistance-signaling pathway.
ACKNOWLEDGMENTS
We thank G. M. Anderson and S. Dahl for technical assistance and J. Valkoun (International Center for Agricultural Research in the Dry Areas, Aleppo Syria) and G. Kleijer (Service de Genetique Swiss Federal Re-search Station for Plant Production, Nyon, Switzerland) for the gift of the wild barley and landrace barley germplasm, respectively. Research was supported by USDA-NRI grant No. 2004 35301-14635, the United States Barley Genome Project, and Lieberman-Okinow Endowment at the Uni-versity of Minnesota.
LITERATURE CITED
1. Brueggeman, R., Drader, T., and Kleinhofs, A. 2006. The barley serine/threonine kinase gene Rpg1 providing resistance to stem rust belongs to a gene family with five other members encoding kinase domains. Theor. Appl. Genet. 113:1147-1158.
2. Brueggeman, R., Rostoks, N., Kudrna, D., Kilian, A., Han, F., Chen, J., Druka, A., Steffenson, B., and Kleinhofs, A. 2002. The barley stem rust-resistance gene Rpg1 is a novel disease-resistance gene with homology to receptor kinases. Proc. Natl. Acad. Sci. USA 99:9328-9333.
3. Buckler, E. S., IV, and Thornsberry, J. M. 2002. Plant molecular diversity and applications to genomics. Curr. Opin. Plant Biol. 5:107-111.
4. Chomczynski, P., and Sacchi, N. 1987. Single-step method for RNA iso-lation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162:156.
5. Eckstein, P., Rossnagel, B. G., and Scoles, G. 2003. Allele-specific mark-ers within the barley stem rust resistance gene (Rpg1). Barley Genet. Newsl. 33:7-11.
6. Edwards, K., Johnstone, C., and Thompson, C. 1991. A simple and rapid method for the preparation of plant genomic DNA and PCR analysis. Nucleic Acids Res. 19:1351.
7. Ellis, J. G., Lawrence, G. J., Luck, J. E., and Dodds, P. N. 1999. Identification of regions in alleles of the flax rust resistance gene L that determine differences in gene-for-gene specificity. Plant Cell 11:495-506.
8. Fetch, T. G., Steffenson, B. J., and Nevo, E. 2003. Diversity and sources of multiple disease resistance in Hordeum spontaneum. Plant Dis. 87:1439-1448.
9. Horvath, H., Rostoks, N., Brueggeman, R., Steffenson, B., von Wettstein, D., and Kleinhofs, A. 2003. Genetically engineered stem rust resistance in barley using the Rpg1 gene. Proc. Natl. Acad. Sci. USA 100:364-369.
10. Jia, Y., Loh, Y.-T., Zhou, J., and Martin, G. B. 1997. Alleles of Pto and Fen occur in bacterial speck-susceptible and fenthion-insensitive tomato cultivars and encode active protein kinases. Plant Cell 9:61-73.
11. Kleinhofs, A., Kilian, A., Saghai Maroof, M. A., Biyashev, R. M., Hayes, P., Chen, F. Q., Lapitan, N., Fenwick, A., Blake, T. K., Kanazin, V., Ananiev, E., Dahleen, L., Kudrna, D., Bollinger, J., Knapp, S. J., Liu, B., Sorrells, M., Heun, M., Franckowiak, J. D., Hoffman, D., Skadsen, R., and Steffenson, B. J. 1993. A molecular, isozyme and morphological map of the barley (Hordeum vulgare) genome. Theor. Appl. Genet. 86:705-712.
12. Kolmer, J. A. 2001. Early research on the genetics of Puccinia graminis and stem rust resistance in wheat in Canada and the United States. Pages 51-82 in: Stem Rust of Wheat: From Ancient Enemy to Modern Foe. P. D. Peterson, ed. American Phytopathological Society, St. Paul, MN.
13. Nirmala, J., Brueggeman, R., Maier, C., Clay, C., Rostoks, N., Kannangara, C. G., von Wettstein, D., Steffenson, B., and Kleinhofs, A. 2006. Subcellular localization and functions of the barley stem rust resistance receptor-like serine/threonine-specific protein kinase Rpg1. Proc. Natl. Acad. Sci. USA 103:7518-7523.
14. Richter, T. E., Pryor, T. J., Bennetzen, J. L., and Hulbert, S. H. 1995. New rust resistance specificities associated with recombination in the Rp1 complex in maize. Genetics 141:373-381.
15. Richter, T. E., and Ronald, P. C. 2000. The evolution of disease resistance genes. Plant Mol. Biol. 42:195-204.
16. Riely, B. K., and Martin, G. B. 2001. Ancient origin of pathogen recognition specificity conferred by the tomato disease resistance gene Pto. Proc. Natl. Acad. Sci. USA 98:2059-2064.
17. Song, W.-Y., Pi, L.-Y., Wang, G.-L., Gardner, J., Holsten, T., and Ronald, P. C. 1997. Evolution of the rice Xa21 disease resistance gene family. Plant Cell 9:1279-1287.
18. Steffenson, B. J. 1992. Analysis of durable resistance to stem rust in barley. Euphytica 63:153-167.
19. Steffenson, B. J., Miller, J. D., and Yin, Y. 1993. Detection of the stem rust resistance gene Rpg1 in barley seedlings. Plant Dis. 77:626-629.
20. Steffenson, B. J., Olivera, P., Roy, J. A., Jin, Y., Smith, K. P., and Muehlbauer, G. J. 2007. A walk on the wild side: Mining wild wheat and barley collections for rust resistance genes. Aust. J. Agric. Res. 58:532-544.
21. Sun, Y., and Steffenson, B. J. 2005. Reaction of barley seedlings with different stem rust resistance genes to Puccinia graminis f. sp. tritici and P. graminis f. sp. secalis. Can. J. Plant Pathol. 27:80-89.
22. Williams, K. J. 2003. The molecular genetics of disease resistance in barley. Aust. J. Agric. Res. 54:1065-1079.
73

CHAPTER FOUR The stem rust resistance gene Rpg5 encodes a protein with nucleotide-binding-site, leucine-
rich, and protein kinase domains.
R. Brueggeman1*, A. Druka2, J. Nirmala1, T. Cavileer3, T. Drader4, N. Rostoks5, A. Mirlohi6, H.
Bennypaul7, U. Gill1, D. Kudrna8, C. Whitelaw9, A. Kilian10, F. Han11, Y. Sun12, K. Gill1, B.
Steffenson13, A. Kleinhofs1,4
1 Department of Crop and Soil Science, Washington State University, Pullman, WA 99164, USA 2 Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, Scotland, UK 3University of Idaho, Dept. Biological Sciences, Life Sciences Building, Moscow, ID 83844, USA 4 School of Molecular Biosciences, Washington State University, Pullman, WA 99164, USA 5 Faculty of Biology, University of Latvia, 4 Kronvalda Blvd., Riga, Latvia LV-1586 6 College of Agriculture, Isfahan University of Technology, Esfahan, Iran 7 CFIA Charlottetown Laboratory 93, Mount Edward Road, Charlottetown, Prince Edward Island C1A 5T1, Canada 8 Arizona Genomics Institute, Dept. Plant Sciences, University of Arizona, Tuscon AZ 85721-0036, USA 9 The Institute for Genomics Research, 9704 Medical Center Drive, Rockwille, MD 20850 10 Diversity Arrays Technology Pty Ltd, Building 1 CSIRO Forestry 1 Wilf Crane Crescent, Yarralumla PO Box 7141, Yarralumla, ACT 2600, Australia 11 Pioneer Hi-Bred International, Inc., 7300 NW 62nd Ave., Johnston, IA 50131, USA 12 Monsanto Company, 8350 Minnegan Road, Waterman, IL 60556 13 Department of Plant Pathology, University of Minnesota, St. Paul, MN 55108-6030, USA Corresponding author: Andris Kleinhofs, Dept. Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6420, tel.-509-335-4389, fax-509-335-8674, [email protected]
As Published In
Proceedings of the National Academy of Sciences (2008) 105(39):14970-14975
* R. Brueggeman is the main contributor
74

The stem rust resistance gene Rpg5 encodes aprotein with nucleotide-binding-site, leucine-rich,and protein kinase domainsR. Brueggemana, A. Drukab, J. Nirmalaa, T. Cavileerc, T. Draderd, N. Rostokse, A. Mirlohif, H. Bennypaulg, U. Gilla,D. Kudrnah, C. Whitelawi, A. Kilianj, F. Hank, Y. Sunl, K. Gilla, B. Steffensonm, and A. Kleinhofsa,d,n
aDepartment of Crop and Soil Science, Washington State University, Pullman, WA 99164; bScottish Crop Research Institute, Invergowrie, Dundee DD2 5DA,Scotland, United Kingdom; cDepartment of Biological Sciences, Life Sciences Building, University of Idaho, Moscow, ID 83844, dSchool of MolecularBiosciences, Washington State University, Pullman, WA 99164; eFaculty of Biology, University of Latvia, 4 Kronvalda Boulevard, Riga, Latvia LV-1586; fCollegeof Agriculture, Isfahan University of Technology, Esfahan, Iran; gCFIA Charlottetown Laboratory 93, Mount Edward Road, Charlottetown, Prince EdwardIsland C1A 5T1, Canada; hArizona Genomics Institute, Department of Plant Sciences, University of Arizona, Tucson AZ 85721-0036; iOffice of Research andGraduate Studies, National University of Ireland, Maynooth, Ireland; jDiversity Arrays Technology Pty. Ltd., Building 1, CSIRO Forestry 1 Wilf Crane Crescent,Yarralumla, P.O. Box 7141, Yarralumla, ACT 2600, Australia; kPioneer Hi-Bred International, Inc., 7300 NW 62nd Avenue, Johnston, IA 50131; lMonsantoCompany, 8350 Minnegan Road, Waterman, IL 60556; and mDepartment of Plant Pathology, University of Minnesota, St. Paul, MN 55108-6030
Communicated by Diter von Wettstein, Washington State University, Pullman, WA, August 1, 2008 (received for review March 1, 2008)
We isolated the barley stem rust resistance genes Rpg5 and rpg4by map-based cloning. These genes are colocalized on a 70-kbgenomic region that was delimited by recombination. The Rpg5gene consists of an unusual structure encoding three typical plantdisease resistance protein domains: nucleotide-binding site,leucine-rich repeat, and serine threonine protein kinase. The pre-dicted RPG5 protein has two putative transmembrane sites possi-bly involved in membrane binding. The gene is expressed at lowbut detectable levels. Posttranscriptional gene silencing using VIGSresulted in a compatible reaction with a normally incompatiblestem rust pathogen. Allele sequencing also validated the candidateRpg5 gene. Allele and recombinant sequencing suggested that theprobable rpg4 gene encoded an actin depolymerizing factor-likeprotein. Involvement of actin depolymerizing factor genes innonhost resistance has been documented, but discovery of theirrole in gene-for-gene interaction would be novel and needs to befurther substantiated.
actin depolymerizing factor � barley � disease resistancedomains � map-based cloning
S tem rust caused by the fungus Puccinia graminis (Pg) washistorically one of the most significant foliar diseases of
barley and wheat, with genetic resistance being the primarymeans of control. Durable resistance in barley has been achievedagainst many pathotypes for the past 60� years by the wide-spread use of the single resistance gene, Rpg1 (1).
Barley can be attacked by Pg f. sp. tritici (Pgt), the wheat stemrust pathogen and Pg f. sp. secalis (Pgs), the rye stem rustpathogen. In cultivated barley, five genes are known to conferresistance to Pgt and three to Pgs (2, 3). Only Rpg1 has beencloned and characterized (4, 5). A virulent Pgt pathotype,designated QCC, was isolated from Midwestern barley cvs.containing Rpg1 in 1989 (6). A resistance gene identified inbarley line Q21861 was designated rpg4. It acts in recessivemanner to pathotype QCC and is temperature-sensitive (3, 7, 8).Besides rpg4, Q21861 carries Rpg5 providing resistance to Pgsisolate 92-MN-90 (3, 8). The Rpg5 gene, previously RpgQ, isdominant or semidominant in action and was reported tocosegregate with rpg4, although three exceptions were found in769 F2 progeny (9). The rpg4 gene was mapped to the long armof barley chromosome 7(5H) (10). More detailed mapping andidentification of syntenic rice chromosome regions (11, 12)allowed the development of a physical map covering the pre-sumed rpg4 gene region (13).
Advances in molecular techniques and tools have facilitatedthe cloning of numerous plant disease resistance genes (R genes)in the past two decades. R genes are grouped into different
classes according to their protein domain structure (14). Thelargest group consists of the NBS-LRR family of R genes, whichis characterized by an N-terminal nucleotide-binding site (NBS)and C-terminal leucine rich repeats (LRRs). Another class, withrelatively few members, is the extracellular LRR and transmem-brane (TM) domain containing genes conferring resistance tothe fungus Cladosporium fulvum, the leaf mold pathogen oftomato. The rice genes Xa21 (15) and Xa26 (16) conferringresistance to the bacterial blight pathogen Xanthomonas oryzaeare the only examples of receptor-like kinase genes consisting ofan extracellular LRR, a TM, and a cytoplasmic serine/threonineprotein kinase (S/TPK). Last, there is a class of R genes thatconsists of S/TPK domains. S/TPK R genes include the previ-ously described barley Rpg1 gene (4); the tomato Pto gene, whichconfers resistance to the bacterial pathogen Pseudomonas syrin-gae pv. tomato (17); and the Arabidopsis PBS1 gene, whichconfers resistance to the bacterial pathogen Pseudomonas syrin-gae pv. phaseolicola (18).
The S/TPK group of R genes is unique in that two of themembers, Pto and PBS1, have been shown to require an NBS-LRR gene, Prf and RPS5, respectively, for resistance (19, 20).This demonstrates that NBS-LRR and protein kinases some-times work together to provide resistance to plant pathogenicorganisms.
Here, we report the cloning and preliminary characterizationof two unique barley stem rust resistance genes Rpg5 and rpg4.Rpg5 encodes an R gene protein containing the NBS, LRR, andS/TPK domains in a single transcript. We validated the candidateRpg5 gene by multiple allele sequencing and VIGS (21).
Available recombinants point to the rpg4 gene encoding anactin depolymerizing factor (Adf). Adfs play an important role
Author contributions: R.B., K.G., B.S., and A. Kleinhofs designed research; R.B., A.D., J.N.,T.C., T.D., N.R., A.M., H.B., U.G., D.K., C.W., A. Kilian, F.H., Y.S., and B.S. performed research;R.B., A.D., J.N., T.D., N.R., A.M., H.B., U.G., K.G., B.S., and A. Kleinhofs analyzed data; andR.B., A.D., and A. Kleinhofs wrote the paper.
The authors declare no conflict of interest.
Data deposition: The sequences reported in this paper have been deposited in the GenBankdatabase [accession nos. EU883792 (Rpg5 cDNA from barley line Q21861); EU878778(genomic sequence from barley line Q21861); EU812563 (genomic sequence from barleyline Morex); EU881932-EU881935 (HvAdf2 genomic sequence from barley lines GoldenPromise, Harrington, MD2, and Steptoe, respectively); EU883581-EU883583 (HvRGA1genomic sequence from barley lines Harrington, MD2 and Steptoe, respectively);EU883787-EU883790 (HvRGA2 genomic sequence from barley lines Golden Promise, Har-rington, MD2, and Steptoe, respectively); and EU883791 (HvAdf3 genomic sequence frombarley line Steptoe)].
nTo whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0807270105/DCSupplemental.
© 2008 by The National Academy of Sciences of the USA
14970–14975 � PNAS � September 30, 2008 � vol. 105 � no. 39 www.pnas.org�cgi�doi�10.1073�pnas.0807270105
75

in cytoskeleton organization and have been reported to beinvolved with nonhost disease resistance (22). Their role ingene-for-gene interaction would be novel and needs to be furthersubstantiated.
ResultsHigh-Resolution Genetic and Physical Mapping. High-resolution ge-netic mapping identified the Rpg5/rpg4 containing region be-tween RFLP markers ARD5112 and ARD5016. This region wasdelimited by 10 cross-overs, 7 in the HvAdf2 region and 3between ARD5112 and HvAdf3 (Fig. 1 A and D). The recom-binants around the HvAdf2 region separated rpg4 (resistance toPgt pathotype QCC) from Rpg5 (resistance to Pgs isolate 92-MN-90) (Fig. 1D).
Physical mapping identified BAC clones 543P19 and 116G10
spanning the ARD5112 to ARD5016 region (Fig. 1B). Geneticand physical mapping confirmed that these BAC clones includedthe flanking markers and the Rpg5/rpg4 locus.
Identification and Analysis of Candidate Rpg5 and rpg4 Genes. Se-quence analysis of the cv. Morex BAC clones 543P19 and 116G10identified a 70-kb region f lanked by the RFLP markersARD5016 and ARD5112. This region was annotated, and fivecandidate genes were identified, two encoding predicted R-likeNBS-LRR proteins (HvRGA1 and HvRGA2), two actin depo-lymerizing factors (HvADF2 and HvADF3), and a proteinphosphatase 2C protein (HvPP2C) (Fig. 1C).
PCR primers designed to the cv. Morex candidate genesamplified four of the five genes from the resistant line Q21861.The HvPP2C gene could not be amplified with Morex-specificPCR primers, suggesting a diverged gene or an indel event. TheQ21861 noncolinearity region was amplified by using PCRchromosome walking (23) and sequenced revealing a S/TPKdomain associated with the Rpg5 candidate gene HvRGA2 (Fig.2). The S/TPK domain was absent from susceptible cvs. Morex,Steptoe, and Harrington, suggesting its involvement in Rpg5-mediated resistance.
To characterize the noncolinearity region further, a Q21861 �library was developed and clone RSB762 identified. The RSB762sequence contained the Rpg5 gene with 5 kb 5� region and 7.5 kbextending 3� into the noncolinearity region. Additional chromo-some walking resulted in 15,708-bp Q21861 sequence. This
rpg4HvAdf2
Rpg5
A genetic map Rpg5HvRGA1HvRGA2HvPP2CHvAdf3
Chromosome 7(5H)
CT
HvAdf2
116G10
ARD5112
xx
xxx
xx
xx
x
543p19HvRGA1ARD5016
ARD5112
HvAdf2
HvAdf2
HvRGA1
HvRGA1
HvRGA2
HvRGA2
HvPP2C
HvPP2C
HvAdf3
HvAdf3
ARD5016
HvAdf3
HQ1 (R), (R)
HQ18 (S), (R)
MQ28 (S), (R)
MQ29 (S), (R)
SQ31 (S), (R)
SQ41 (S), (R)
SQ47 (S), (R)
HQ9 (S), (S)
MQ27 (S), (S)
SQ36 (S), (S)
NBS-LRR-S/TPK
kb 5 10 15
70 kb
ARD5005
kb 5
B physical map
C sequence
D recombinants
HvAdf2 HvRGA1
Morex
Q21861
1 6 3
***
**
Fig. 1. Genetic, physical, sequence, and recombinant characterization of theRpg5/rpg4 locus. (A) High-resolution genetic map of the Rpg5 region based on5,232 recombinant gametes. The Rpg5 flanking markers (ARD5016 andARD5112) are designated above the white vertical bars. The candidate genescosegregating with isolate 92-MN-90 (Rpg5) resistance and pathotype QCC(rpg4) resistance are inside the boxes labeled Rpg5 and rpg4, respectively.Numbers below the bar indicate the number of cross-overs. C and T representcentromere and telomere. (B) BAC physical map spanning the Rpg5/rpg4locus. The two BAC clones (543P19 and 116G10) spanning the region areshown as horizontal gray bars. The white circles indicate the flanking geneticmarkers. Black circles indicate the position of Rpg5 and rpg4 candidate genesand the marker (ARD5005) used for chromosome walking. (C) Sequenceannotation of the Rpg5 region. White horizontal bars represent sequencedregions from Morex and Q21861. White circles show the position of flankingmarkers. The black arrow represents the Rpg5 NBS-LRR-S/TPK gene. Grayarrows indicate annotated genes. The darker gray background betweenHvRGA2 and HvAdf3 indicates the region of colinearity breakdown. The scaleis shown above in kilobases. (D) Recombinant sequence analysis. Horizontalbars represent sequence and/or genotyping from 10 lines with recombinationdefining the Rpg5/rpg4 region. Black indicates susceptible genotype, andwhite indicates the resistant Q21861 genotype. The ‘‘X’’ indicates the approx-imate point of recombination. The recombinant designations are labeled tothe right with resistant (R) or susceptible (S), indicating the response topathotype QCC (rpg4) and isolate 92-MN-90 (Rpg5), respectively. Gray arrowsindicate candidate genes and the white circles represent flanking markers.Recombinant lines indicated with an asterisk have complete sequence analysisat the region of recombination. The scale is shown below in kilobases.
217
1,378
*
*
*
1 2 3 4 5 6 7
A gene structure
B protein structure
ATG
ATG
TGA
TGA PASTSS
5’ UTR 3’ UTR
kb 1 2 3 4 5 6 7 8 9
AAA
X100 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Group 1 (Resistant)
Group 2 (susceptible)
Group 3 (susceptible)
Group 4 (susceptible)
161
NBS
NBS LRR
LRR
S/TPK
1,075
*
Fig. 2. Rpg5 predicted gene structure from resistant and susceptible culti-vars. (A) cDNA and genomic DNA structures with introns (gray), exons (black),and UTRs white shown to scale. Rpg5 predicted mRNA structure is shownabove the genomic sequence with exons numbered above. ATG represents thestart methionine codon and TGA represents the stop codon. AAA is the mRNApoly(A) tail, TSS is the transcription start site, and PAS is the polyadenylationsignal. The scale below is shown in kilobases. (B) Organization of the predictedprotein structure is shown to scale with predicted boundaries indicated byboxes. NBS denotes the nucleotide-binding site, LRR denotes 12 imperfectleucine-rich repeats, S/TPK denotes the serine threonine protein kinase do-main, and the black bars represent predicted transmembrane domains. Num-bers above indicate the last amino acid preceding the stop codon. Stop codonsare represented by an asterisk. Cultivars or lines were placed into four groups.Group 1 (resistant) consists of Q21861 with a predicted intact and functionalRpg5 gene. Group 2 (susceptible) consists of the cultivars Morex and Steptoe.They have a cytosine-to-adenine conversion that introduces a stop codonresulting in a predicted truncated protein. In addition, they are missing thekinase domain coding region. Group 3 (susceptible) contains Golden Promiseand MD2. They have a single cytosine insertion causing a frame-shift mutationthat results in a stop codon and a predicted truncated protein. Group 4(susceptible) consists of Harrington. Harrington is missing the kinase domaincoding region and the second transmembrane domain.
Brueggeman et al. PNAS � September 30, 2008 � vol. 105 � no. 39 � 14971
GEN
ETIC
S
76

completed the noncolinearity region except for a small retro-transposon block, estimated at �2 kb by restriction mapping,which we could not assemble because of its repetitive nature. TheS/TPK domain was the only additional gene found in the Q21861indel region.
An approximately 18-kb region between HvRGA1 and Rpg5contained a large retrotransposon block in Morex. We were notable to assemble Q21861 contiguous sequence from this region.The partial sequences obtained were all repetitive and/or retro-transposon-like, with no evidence of additional genes. The57.3-kb total Q21861 sequence identified the same candidategenes as in the susceptible Morex sequence, except that theHvPP2C gene was replaced by a protein kinase domain.
Comparison of Morex and Q21861 sequences suggested thatHvRGA2 was the Rpg5 gene (nucleotide and amino acid num-bers given are from the Q21861 Rpg5 sequence, starting with thefirst nucleotide in the translation start codon and the first aminoacid). The Morex HvRGA2 consisted of typical NBS-LRRdomains and a PP2C gene a short distance downstream (Fig. 1C).The Q21861 HvRGA2 gene consisted of NBS-LRR-S/TPKdomains (Figs. 1C and 2). Furthermore, the susceptible cv.Morex genomic and cDNA sequence revealed a nucleotidesubstitution at position �483, resulting in a stop codon at aminoacid position 161 and a predicted truncated protein (Fig. 2B,Group 2). This correlation of an apparently nonfunctionalprotein and a susceptible phenotype suggested that HvRGA2could be the Rpg5 gene. Sequence analysis of the HvRGA2alleles from the susceptible parents showed Steptoe to be similarto Morex, encoding a predicted truncated protein. MD2 andGolden Promise contained a single-nucleotide insertion (C337)causing a frame-shift mutation at amino acid position 114resulting in a stop codon at aa position 217 (Fig. 2B, Group 3).The susceptible cv. Harrington HvRGA2 allele contained anamino acid sequence very similar to Q21861, but it was lackingthe PK domain (Fig. 2B).
The remaining candidate genes were eliminated based onallele sequencing. The candidate gene HvRGA1 codes for an895-aa (98.5-kDa) predicted NBS-LRR protein with highesthomology to an Oryza sativa hypothetical NBS-LRR protein(GenBank accession no. EAY80410). Sequence analysis ofHvRGA1 alleles revealed that the resistant parent Q21861 andthe susceptible parent MD2 had identical HvRGA1 predictedamino acid sequences. The susceptible parent Harrington dif-fered from Q21861 by only a single amino acid (C324R). Thesusceptible cvs. Morex and Steptoe were identical at the aminoacid level and differed from Q21861 by five amino acids (S290A,K340N, A445G, N474D, and F586L). The single amino aciddifference between Q21861 and Harrington occurred within theNBS domain but was not within one of the highly conserved subdomains. Two of the amino acid differences between Q21861and Morex/Steptoe occurred within the NBS domain and threeoccurred within the predicted LRR region. The amino acidsequence identity between Q21861 and MD2 in addition to thevery limited polymorphism between Q21861 and Harringtonsuggested that HvRGA1 was not Rpg5.
The HvAdf3 gene codes for a protein with the highest aminoacid homology to a Lophopyrum elongatum actin depolymerizingfactor-like gene (GenBank accession no. AAG28460). Allelesequencing from the mapping population parents (Steptoe,MD2, and Q21861) and cv. Morex showed that HvADF3 had nopolymorphism at the amino acid level, suggesting that it is ahighly conserved protein and not the Rpg5 gene.
The HvPP2C gene sequence from cv. Morex codes for aprotein with the highest homology to an expressed O. sativaprotein phosphatase 2C family protein (GenBank accession no.ABF99721). Specific HvPP2C primers (from Morex) failed toamplify Q21861, MD2 or cv. Golden Promise (susceptible),suggesting that either the HvPP2C gene was not present or was
highly diverged. Southern blot analysis confirmed that HvPP2Cwas absent from Q21861 genome (data not shown). Sequenceanalysis of the � clone RSB762 confirmed the absence ofHvPP2C gene from this region of the resistant line Q21861.
Genetic mapping identified seven recombination events be-tween the markers ARD5016 and HvRGA1, a physical region of12 kb containing HvAdf2 and HvRGA1 (Fig. 1D). The sites ofrecombination were identified to within �200-bp intervals bysequencing and SNP analysis. Recombinants with the susceptiblecvs. HvAdf2 allele were resistant to the stem rust isolate 92-MN-90, eliminating HvAdf2 from consideration as the Rpg5 gene(Fig. 1D). However, the six recombinants differentiating rpg4(Pgt pathotype QCC resistance) from Rpg5 (Pgs isolate 92-MN-90 resistance) and an additional recombinant occurringdistal of HvAdf2 (HQ1) identified HvAdf2 as the probable rpg4gene.
HvAdf2 codes for a 147 aa (16.2 kDa) actin-depolymerizingfactor-like protein with highest homology to the O. sativaactin-depolymerizing factor 4 expressed gene (GenBank acces-sion no. ABF99587.1). Sequence analysis of the HvAdf2 allelesrevealed that the resistant parent Q21861 and susceptible parentSteptoe and cv. Morex differed by three amino acids (Q39H,A101T, and S135G). However, the pathotype QCC susceptibleparent Harrington had an HvAdf2 gene identical to the Q21861allele at the amino acid level.
Structure and Expression of the HvRGA2 (Rpg5) Gene. RT-PCRanalysis of HvRGA2 showed it was expressed at the mRNA levelin all parents tested, indicating that transcription did not corre-late with resistance. However, PCR primers designed to amplifythe junction between the LRR domain and the S/TPK domainproduced an RT-PCR product only from Q21861, MD2, andGolden Promise [supporting information (SI) Table S1]. Al-though MD2 and Golden Promise contain the intact NBS-LRR-S/TPK transcript, both MD2 and Golden Promise alleles wereshown to contain a frame-shift mutation within the N-terminalregion of the gene resulting in a stop codon at amino acidposition 217 and a putative truncated protein (Fig. 2B, Group 3).
To obtain HvRGA2 (Rpg5) transcription start site (TSS)primers were designed from the genomic sequence in 100 bp(Table S1) increments. They were used in RT-PCRs to delimitthe TSS to within 100 bp at position �408 to �346 bp.
HvRGA2 (Rpg5) encodes an apparently functional NBS-LRR-S/TPK gene containing seven exons transcribed into apredicted 4.4-kb mRNA coding for a 1,378-aa (151.6-kDa)predicted protein (Fig. 2). The transcript size was confirmed byNorthern blot analysis, which showed a single hybridizing bandat �4.8 kb (Fig. 3A).
The S/TPK domain contains all nine conserved amino acids(24), suggesting a functional kinase. The kinase domain had thehighest homology to an O. sativa putative S/TPK (GenBankaccession no. EAZ08788) but also showed significant similarityto the known R gene Pto (36% amino acid identity and 53%amino acid similarity). The Rpg5 S/TPK domain was veryhomologous to the unknown function Rpg1 gene family membersABC1041 and ABC1063 with 60% and 61% amino acid identity,respectively, and 76% amino acid similarity (25).
The NBS-LRR region contained the 4 NBS conserved sub-domains and 12 imperfect LR repeats. This region had thehighest homology to an O. sativa hypothetical protein (GenBankaccession no. EAY98635). The Rpg5 NBS-LRR is very similar tothe rice gene Pi-ta (40% amino acid identity and 54% amino acidsimilarity) conferring resistance to the rice blast fungus (26).
The TMPRED program (www.ch.embnet.org/software/TMPRED�form.html) predicted two transmembrane domains,one on the C-terminal side of the NBS domain and the other onthe C-terminal side of the LRR domain (Fig. 2B, Group 1).
14972 � www.pnas.org�cgi�doi�10.1073�pnas.0807270105 Brueggeman et al.
77

Virus-Induced Gene Silencing of the Rpg5 Gene. The BSMV vector(21) was used to posttranscriptionally silence the Rpg5 gene withits antisense RNA. Approximately 30% (average of three inde-pendent experiments) of the Q21861 BSMV-asRpg5 treatedseedlings showed a conversion from incompatible reaction tocompatible, whereas all of the control BSMV-MCS-treatedplants remained resistant to isolate 92-MN-90 (Fig. 3B). TheBSMV-MCS control demonstrated that the BSMV-VIGS vectoritself did not induce susceptibility to fungal infection.
Quantitative analysis (qRT-PCR) of the Rpg5 mRNA wascarried out on samples taken at the time of fungal infection(designated TP0) and at 14 d after fungal infection (designatedTP14). At TP0, the BSMV-asRpg5 construct (Fig. 4, bar 1)showed significant mRNA reduction with reference to theBSMV uninoculated control (Fig. 4, bar 3) and BSMV-MCSinoculated control (Fig. 4, bar 2). At TP14, the uninoculateduninfected control (Fig. 4, bar 4) showed an Rpg5 mRNA levelcomparable to the TP0 control (Fig. 4, bar 3), but the TP14
BSMV uninoculated control (Fig. 4, bar 5) showed much highermRNA levels which are comparable to the BSMV-MCS inocu-lated control (Fig. 4, bar 8) and the BSMV-asRpg5 inoculatedplants that did not show resistance phenotype (Fig. 4, bar 7).Only the BSMV-asRpg5 inoculated seedlings showing a suscep-tible phenotype (Fig. 4, bar 6) had highly reduced Rpg5 mRNAlevels compared with the uninoculated and MCS inoculatedseedlings (Fig. 4).
The data suggested that the Rpg5 mRNA may be induced byinfection with isolate 92-MN-90. A time-course experimentshowed that there is a strong induction of the Rpg5 mRNA at day1 (12.7%, relative to a GAPDH control) and a second smallerpeak at days 8 (3.5%) through 11 (3.2%). This response,however, was also observed in mock and uninoculated controlsand therefore is not fungus infection specific (data not shown).We conclude that the Rpg5 mRNA levels f luctuate because ofenvironmental or developmental reasons, but the exact causeand pattern remain to be investigated. However, the Rpg5mRNA silencing by the BSMV-asRpg5 VIGS inoculation mustbe made with a control sample taken at the same time pointunder identical environmental conditions.
DiscussionHigh-resolution mapping identified a 70-kb region on chromo-some 7(5H) encompassing the Rpg5 and rpg4 genes. Within thislocus, we identified five candidate genes. Multiple allele se-quencing and recombinant characterization eliminated fourgenes, leaving only the HvRGA2 as the candidate Rpg5 gene(Fig. 1). Allele sequencing and VIGS confirmed the candidateRpg5 gene. This is the first report of an R-gene encoding adisease-resistance protein composed of NBS-LRR-S/TPK do-mains. A search of the Oryza and Arabidopsis genomes did notidentify any genes with this novel three-domain structure. Theprotein also contains two predicted transmembrane domains,suggesting a potential membrane-bound protein. The location ofthe transmembrane domains, if functional, suggests that theRPG5 LRR domain may reside outside the cell and act as thepathogen receptor, whereas the NBS and PK domains areintracellular and propagate the disease resistance signaling.
An NBS-LRR domain is present in a majority of plant diseaseresistance proteins identified to date (14). Another class ofresistance proteins is represented by S/TPKs such as Pto, PBS1,and RPG1 (14, 18). The tomato Pto and the Arabidopsis PBS1S/TPKs require function of the NBS-LRR genes Prf and RPS5,respectively, for disease resistance (18, 19). The Rpg5 gene,identified here, is the only reported disease resistance genewhere all three domains are present in a single protein. TheS/TPK of the Rpg5 gene is clearly required for disease resistance,as indicated by the susceptible cv. Harrington allele that appearsto have an intact and expressed NBS-LRR protein but lacks theprotein kinase domain. The presence of all three domains in asingle protein may facilitate studying their interactions.
The Pto and Prf proteins have been reported to act coinci-dentally with one another for detecting and eliciting a disease-resistance response to the pathogen (27). Given this closeinteraction between the PK and NBS-LRR proteins, it is notsurprising that a gene has been found that combines the twodomains in one protein.
The Rpg5/rpg4 locus also confers resistance to pathotype QCC.The HvAdf2 gene was identified as the probable rpg4 gene basedon analysis of the genetic recombinants. Interestingly, whereaswe found recombinants that expressed resistance to isolate92-MN-90 but susceptibility to pathotype QCC, the reciprocalrecombinants were not observed. In general, resistance topathotype QCC corresponded with resistance to isolate 92-MN-90 in studies of a large number of domesticated and wild
A B96543
2
1
.51. 2. Steptoe asRpg5(S) asRpg5(R) MCS Q21861
Fig. 3. Northern analysis of Rpg5 and virus-induced gene silencing pheno-types. (A) Northern blot analysis of Rpg5 transcript. Lane 1 is Q21861 poly(A)mRNA with the Rpg5 specific probe hybridizing to a single band at �4.8 kb,indicated by the arrow to the right. Lane 2 is a methylene blue stain of the RNAmarker after transfer to membrane. Numbers to the right indicate molecularmass of Millennium Marker (Ambion) bands. (B) Barley seedlings inoculatedwith antisense Rpg5 or MCS VIGS constructs and challenged with Pgs isolate92-MN-90. Approximately 30% of the inoculated seedlings showed a typicalsusceptible reaction to the Pgs 92-MN-90 fungal infection. These are markedasRpg5(S), whereas seedlings that failed to show a reaction to the fungalinfection are marked asRpg5(R). A representative seedling is shown for eachgroup. The MCS antisense (BSMV-MCS) treated plant shows typical resistantreaction to Pgs 92-MN-90. The seedling leaves marked Steptoe and Q21861 arethe rust-susceptible and resistant virus uninoculated controls, respectively.
5
4
3
2
1
% G
APD
H e
xpre
ssio
n
Time (DPI with 92-MN-90)
1 2 3 4 5 6 7 80 14
Fig. 4. Expression of the Rpg5 gene after BSMV-VIGS induced gene silencingand infection with isolate 92-MN-90 given as percentage of GAPDH geneexpression. Bars represent BSMV-VIGS data from plants sampled at either timepoint 0 or 14 d postfungal infection (DPI). Sample 1 is inoculated withBSMV-asRpg5, 2 is inoculated with BSMV-MCS, and 3 is the uninoculatedcontrol all sampled at TP0. Sample 4 is the untreated control, 5 is the virusuninoculated control, 6 is inoculated with BSMV-asRpg5 sample taken fromseedlings showing susceptible fungal reaction, 7 is inoculated with BSMV-asRpg5 sample taken from seedlings failing to show susceptible fungal reac-tions, and 8 is inoculated with BSMV-MCS, all sampled at TP14. The plants wereexamined for disease reaction at 14 DPI. The tissue analyzed by qRT-PCR wastaken at TP0 or TP14 d after inoculation with isolate 92-MN-90.
Brueggeman et al. PNAS � September 30, 2008 � vol. 105 � no. 39 � 14973
GEN
ETIC
S
78

barleys (B.S., unpublished data). The observation that theQCC-susceptible cv. Harrington has an expressed Adf2 geneidentical to the resistant Q21861 allele at the amino acid levelalso suggested that additional factors may be involved. Onehypothesis is that the recessive rpg4 gene functions as a pathotypeQCC-specific disease-resistance gene only in the presence ofanother functional disease resistance gene, perhaps Rpg5.
ADFs are critical in remodeling the actin cytoskeleton duringnormal plant development and under biotic and abiotic stress.They are typically small proteins that function in rapid recyclingof actin monomers (28). Actin functions in cytoskeleton orga-nization, which coordinates essentially all aspects of plant growth(29). Actin microfilament polymerization has been shown to beinvolved in nonhost disease resistance. For example, usingcytochalasin E, an inhibitor of actin microfilament polymeriza-tion and Arabidopsis defense-related mutants eds1, pad4, andnah4, it was shown that nonhost resistance to the wheat powderymildew fungus Blumeria graminis f. sp. tritici in Arabidopsislargely depends on actin cytoskeleton dynamics and function ofthe EDS1 gene (30). Similar observations have been documentedin other systems.
The posttranscriptional gene silencing of Rpg5 by VIGSshowed significant reduction of Rpg5 transcript at both samplingtime points 0 d (seedlings pooled) and 14 d (susceptible andresistant plants sampled separately). Approximately 30% of theBSMV-asRpg5 plants resulted in a susceptible reaction to therust fungus. At the 14-d time point, the level of silencingobserved corresponded to the phenotype observed with thesusceptible plants showing significant reduction in the Rpg5mRNA, whereas the resistant plants had mRNA level similar tothe controls (Fig. 4). The time-course experiments indicated thatRpg5 mRNA levels were variable over time. However, inductionby fungal infection could not be established because of similarobservations in mock and uninoculated controls. The variableRpg5 mRNA levels were surprising, because previous experi-ments with Rpg1 showed low but steady mRNA levels with orwithout fungal infection (31, 32).
In summary, we have identified two types of plant disease-resistance genes. The Rpg5 gene combines features of NBS-LRRtype disease-resistance gene with the S/TPK domain, suggestingthat this gene may function both in pathogen perception andsignal transduction. The candidate gene for rpg4 needs furtherverification, but, if confirmed, it would show involvement of actincytoskeleton in race-specific disease resistance.
Materials and MethodsPlant Materials. Progeny from the crosses Steptoe/Q21861, Harrington/Q21861, and MD2/Q21861 were used for genetic mapping. Q21861 is thesource of the stem rust-resistance genes Rpg5 and rpg4 (3, 9). Steptoe andHarrington are barley cultivars, and MD2 is a genetic stock with multipledominant mutations. All three are susceptible to stem rust. Plants were grownin the greenhouse with day/night temperatures of 18°C/14°C, respectively.Metal halide lights supplemented a 16-/8-h light/dark photoperiod.
Molecular Markers. RFLP markers were generated as described (13) or bydesigning PCR primers based on the cv. Morex or line Q21861 sequence (thisstudy). Primers are described in Table S1.
Genetic and Physical Mapping. High-resolution mapping at the Rpg5/rpg4locus used 50 recombinants selected from 5,232 gametes between the flank-ing markers Aga5 and ABG391. These 50 recombinants were reduced to 10between the flanking markers ARD5016 and ARD5112. BAC physical mapswere generated as described (13).
Sequencing and Sequence Analysis. The cv. Morex BAC clones (543P19 and116G10) forming a contig across the Rpg5/rpg4 region were sequenced at theInstitute for Genomics Research (TIGR). All other cultivar DNA was sequencedwith the BigDye terminator system on an ABI 373 DNA sequencer (AppliedBiosystems) at the Laboratory for Biotechnology and Bioanalysis, WashingtonState University, Pullman. The � clone RSB762 was subcloned into the NotI site
of pBluescript (Stratagene) and sequenced by using the EZ::TNTM�KAN-2�Insertion Kit (Epicentre). All PCR-generated fragments were either directlysequenced or cloned into pGEM-T Easy vector (Promega) and sequenced byusing the EZ::TNTM�KAN-2� Insertion Kit (Epicentre). For direct sequencing,gel slices were placed in plugged tips (Rainin Instrument) and frozen at �20°C.After complete thawing, the tip was centrifuged at 4,000 � g for 10 min. Theeluate was extracted two times with 24:1 chloroform:isoamyl alcohol andprecipitated with 0.3 M NaOAc and 2.5 volumes 95% EtOH. Primers and clonesare described in Table S1.
Contigs were assembled by using Vector NTI Advance 9.0 contig express(Invitrogen). The cultivar sequence comparisons were done by using VectorNTI alignX and the National Center for Biotechnology Information bl2seqfunction (www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi). All sequenceswere analyzed with the BLASTX and BLASTN algorithms.
Library Construction. High-molecular-mass Q21861 genomic DNA was isolatedby using a modified CTAB method (33). Genomic DNA (�20 �g) was partiallydigested with 0.1 units of Sau 3AI for 1 hour to yield a majority of 15- to 23-kbfragments. DNA was dephosphorylated with 0.1 units of calf intestine alkalinephosphatase for 30 min and separated in 1.2% low melting agarose. DNAfragments 9–23 kb were excised from the gel and recovered by �-agarasetreatment. Genomic fragments were ligated into the Lambda Dash II vector(Stratagene) predigested with BamHI and packaged using the Gigapack III XLpackaging extract (Stratagene).
RNA Isolation and Northern Blot. Isolation of RNA, Northern blot analysis, andhybridization were as described (25).
RT-PCR. Approximately 1 �g of total RNA was used for RT-PCRs by usingM-MLV Reverse Transcriptase (Promega) under the manufacturer’s recom-mended conditions. RT-PCR fragments were directly sequenced after elutionfrom agarose gels as described for direct sequencing.
3� and 5� RACE. 3� RACE was carried out by using the 3� RACE System(Invitrogen) following the manufacturer’s recommended conditions. 5� RACEwas carried out by using the FirstChoice RLM-RACE Kit (Ambion) following themanufacturer’s recommended conditions and SMART technology (34). Thegene specific primers used for RACE are described in Table S1.
VIGS and qRT-PCR. Barley plants for VIGS experiments were grown in thegrowth chamber in plastic pots with a day/night temperature of 20°C �/�1°Cand 18°C �/� 1°C, respectively. A 20-/4-h light/dark photoperiod was providedby cool fluorescent tubes (525 uE/m2s).
Rpg5 was silenced by using VIGS as described (21). A 314-bp Rpg5 cDNAfragment (�2,033 to 2,346 bp) was generated by PCR and ligated in antisense(as) orientation into BSMV-VIGS infectious clone (p�PDS4as) digested withPacI and NotI (21). The BSMV-asRpg5 construct was cotranscribed with the �
and � genomes of the tripartite BSMV virus by using the mMessage mMachineT7 kit (Ambion), and the T7 promoter. RNA was inoculated onto Q21861barley plants at the two-leaf stage. The seedlings were inoculated with isolate92-MN-90 urediniospores at 0.025 mg per plant mixed with a talc carrier�11–12 days after virus infection. After inoculation, the plants were mistedand placed in the dark under high humidity conditions for 22 h, then exposedto light and misted periodically. After 4 h, the misting was stopped, and theleaves were left to dry slowly. When the leaf surfaces were completely dry,plants were moved to the growth chambers at 20°C and 80% relative humid-ity. The plants were scored for compatibility or incompatibility at 14 dayspostfungal infection
The negative control contained a 121-bp antisense fragment of the multicloning site (MCS) from pBluescript K/S (Stratagene). The MCS sequence didnot hybridize to barley genomic DNA at low stringency conditions, indicatingno homologous regions in the barley genome. Q21861 and Steptoe were usedas the resistant and susceptible virus uninoculated controls. qRT-PCR wasperformed on Rotor-Gene 2000 thermocycler (Corbet Research) using theQuantiTect SYBR green PCR system (Qiagen). The Rpg5 primers used aredescribed in Table S1 and GAPDH primers were described (35). Tissue samplesfor qRT-PCR were collected at 0 or 14 d after fungal infection. Tissue samplesfor the time course experiment were collected at 0, 1, 3, 5, 8, 11, and 14 daysafter fungal inoculation.
ACKNOWLEDGMENTS. This research was supported by U.S. Department ofAgriculture National Research Initiative Grant No.2004-35301-14635. This isScientific Paper No. 0402-08 from the College of Agricultural, Human, andNatural Resource Sciences Research Center, Washington State University,Project 0196.
14974 � www.pnas.org�cgi�doi�10.1073�pnas.0807270105 Brueggeman et al.
79

1. Steffenson BJ (1992) Analysis of durable resistance to stem rust in barley. Euphytica63:153–167.
2. Sun Y, Steffenson BJ (2005)Reaction of barley seedlings with different stem rustresistance genes to Puccinia graminis f sp tritici and P g f. sp. secalis. Can J Plant Pathol27:80–89.
3. Jin Y, Steffenson BJ, Miller JD (1994) Inheritance of resistance to pathotypes QCC andMCC of Puccinia graminis f sp tritici in barley line Q21861 and temperature effects onthe expression of resistance. Phytopathology 84:452–455.
4. Brueggeman R, et al. (2002) The barley stem rust-resistance gene Rpg1 is a noveldisease-resistance gene with homology to receptor kinases. Proc Natl Acad Sci USA99:9328–9333.
5. Nirmala J, et al. (2006) Subcellular localization and functions of the barley stem rustresistance receptor-like serine/threonine-specific protein kinase Rpg1. Proc Natl AcadSci USA 103:7518–7523.
6. Roelfs AP, Casper DH, Long DL, Roberts JJ (1991) Races of Puccinia graminis in theUnited States in 1989. Plant Dis 75:1127–1130.
7. Jin Y, Steffenson BJ, Fetch TGJ (1994) Sources of resistance to pathotype QCC ofPuccinia graminis f sp tritici in barley. Crop Sci 34:285–288.
8. Steffenson BJ, Jin Y, Rossnagel BG, Rasmussen JB, Kao K (1995) Genetics of multipledisease resistance in a doubled-haploid population of barley. Plant Breeding 114:50–54.
9. Sun Y, Steffenson BJ, Jin Y (1996) Genetics of resistance to Puccinia graminis f sp secalisin barley line Q21861. Phytopathology 86:1299–1302.
10. Borovkova IG, et al. (1995) Identification of molecular markers linked to the stem rustresistance gene rpg4 in barley. Phytopathology 85:181–185.
11. Han F, et al. (1998) Synteny with rice: Analysis of barley malting quality QTL and rpg4chromosome regions. Genome 41:373–380.
12. Kilian A, Chen J, Han F, Steffenson B, Kleinhofs A (1997) Towards map-based cloningof the barley stem rust resistance genes Rpg1 and rpg4 using rice as an intergenomiccloning vehicle. Plant Mol Biol 35:187–195.
13. Druka A, et al. (2000) Physical mapping of barley stem rust resistance gene rpg4. MolGen Genet 264:283–290.
14. Martin GB, Bogdanove AJ, Sessa G (2003) Understanding the function of plant diseaseresistance proteins. Annu Rev Plant Biol 54:23–61.
15. Song W-Y, et al. (1995) A receptor kinase-like protein encoded by the rice diseaseresistance gene, Xa21. Science 270:1804–1806.
16. Sun X, et al. (2004) Xa26, a gene conferring resistance to Xanthomonas oryzae pvoryzae in rice, encodes an LRR receptor kinase-like protein. Plant J 37:517–527.
17. Martin GM, et al. (1993) Map-based cloning of a protein kinase gene conferring diseaseresistance in tomato. Science 262:1432–1436.
18. Swiderski MR, Innes RW (2001) The Arabidopsis PBS1 resistance gene encodes amember of a novel protein kinase subfamily. Plant J 26:101–112.
19. Salmeron JM, et al. (1996) Tomato Prf is a member of the leucine-rich repeat class ofplant disease resistance genes and lies embedded within the Pto kinase gene cluster.Cell 86:123–133.
20. Warren RF, Merritt RFPM, Holub E, Innes RW (1999) Identification of three putativesignal transduction genes involved in R-gene specified disease resistance in Arabidop-sis. Genetics 152:401–412.
21. Holzberg S, Brosio P, Gross C, Pogue GP (2002) Barley stripe mosaic virus-induced genesilencing in a monocot plant. Plant J 30:315–327.
22. Kobayashi Y, Yamada M, Kobayashi I, Kunoh H (1997) Actin microfilaments arerequired for the expression of nonhost resistance in higher plants. Plant Cell Physiol38:725–733.
23. Cottage A, Yang A, Maunders H, de Lacy RC, Ramsay NA (2001) Identification of DNAsequences flanking T-DNA insertions by PCR-walking. Plant Mol Biol Rep 19:321–327.
24. Hanks S, Quinn A, Hunter T (1988) The protein kinase family: Conserved features anddeduced phylogeny of the catalytic domains. Science 241:42–52.
25. Brueggeman R, Drader T, Kleinhofs A (2006) The barley serine/threonine kinase geneRpg1 providing resistance to stem rust belongs to a gene family with five othermembers encoding kinase domains. Theor Appl Genet 113:1147–1158.
26. Bryan GT, et al. (2000) A single amino acid difference distinguishes resistant andsusceptible alleles of the rice blast resistance gene Pi-ta. Plant Cell 12:2033–2045.
27. Mucyn TS, et al. (2006) The tomato NBARC-LRR protein Prf interacts with Pto kinase invivo to regulate specific plant immunity. Plant Cell 18:2792–2806.
28. Remedios DC, et al. (2003) Actin binding proteins: Regulation of cytoskeletal micro-filaments. Physiol Rev 83:433–479.
29. Wasteneys GO, Galway ME (2003) Remodeling the cytoskeleton for growth and form:An overview with some new views. Annu Rev Plant Biol 54:691–722.
30. Yun B-W, et al. (2003) Loss of actin cytoskeletal function and EDS1 activity, in combi-nation, severely compromises non-host resistance in Arabidopsis against wheat pow-dery mildew. Plant J 34:768–777.
31. Rostoks N, Steffenson BJ, Kleinhofs A (2004) Structure and expression of the barleystem rust resistance gene Rpg1. Physiol Mol Plant Pathol 64:91–101.
32. Zhang L, Castell-Miller C, Dahl S, Steffenson B, Kleinhofs A (2008) Parallel expressionprofiling of barley stem rust interactions. Funct Integr Genom DOI10.1007/s10142-007-0069-0. 8:187–198.
33. Kleinhofs A, et al. (1993) A molecular, isozyme and morphological map of the barley(Hordeum vulgare) genome. Theor Appl Genet 86:705–712.
34. Tabansky I, Nurminsky DI (2003) Mapping of transcription start sites by direct sequenc-ing of SMARTTM RACE products. Biotechnology 34:482–486.
35. Horvath H, et al. (2003) Genetically engineered stem rust resistance in barley using theRpg1 gene. Proc Natl Acad Sci USA 100:364–369.
Brueggeman et al. PNAS � September 30, 2008 � vol. 105 � no. 39 � 14975
GEN
ETIC
S
80

CHAPTER FIVE
The rpg4/ Rpg5 stem rust resistance locus in barley; resistance genes and cytoskeleton
dynamics
Robert Brueggeman1*, Brian J. Steffenson2 and Andris Kleinhofs1
1 Department of Crop and Soil Science, Washington State University, Pullman, WA 99164, USA
2 Department of Plant Pathology, University of Minnesota, St. Paul, MN 55108-6030, USA
Corresponding author: Andris Kleinhofs, Dept. Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6420, tel.-509-335-4389, fax-509-335-8674, [email protected]
As Published In
Cell Cycle (2009) 8(7):977-981
* R. Brueggeman is the main contributor
81

1 Cell Cycle 2009; Vol. 8 Issue 7
Two closely linked resistance genes, rpg4 and Rpg5, conferring resistance to several races of Puccinia graminis, were cloned and characterized. The Rpg5 gene confers resistance to an isolate of Puccinia graminis f. sp. secalis (Pgs), while rpg4 confers resistance to Puccinia graminis f. sp. tritici (Pgt). Rpg5 is a novel gene containing nucleotide binding site-leucine rich repeat domains in combination with a serine threonine protein kinase domain. High-resolution mapping plus allele and recombinant sequencing identified the rpg4 gene, which encodes an actin depolymerizing factor-like protein (ADF2). Resistance against the Pgt races QCCJ, MCCF, TTKSK (aka Ug99) and RCRS requires both Rpg5 and rpg4, while Rpg5 alone confers resistance to Pgs isolate 92-MN-90. The depen-dency on the actin modifying protein ADF2 indicates cytoskeleton reorganization or redirection plays a role in pathogen-host inter-actions. Rpg5 may interact with ADF2 to activate or deactivate its function in the resistance response. Alternatively, Rpg5 could initiate signal transduction leading to resistance in response to detecting ADF2 protein modification. Pgt may redirect the actin cytoskeleton by inducing modifications of ADF2. The redirection of actin could possibly enable the pathogen to develop a haustoria-plant cell cytoskeleton interface for acquisition of nutrients.
The co-evolution of plants and pathogenic fungi has led to the development of an innate defense system in the former that acti-vates a series of responses to resist colonization by latter. Activation of defense mechanisms begins with a receptor recognizing the threat of a potential invader. Resisting introgression at the cell periphery is the first line of defense and is dependant upon a dynamic reorganization of the challenged cell’s cytoskeleton. If the initial defense strategy fails to stop penetration, a second line of defense is triggered by race specific resistance genes, resulting in a hypersensitive response (HR) and localized cell death.1,2
We recently reported the cloning and characterization of two closely linked stem rust resistance genes rpg4 and Rpg5.3 The Rpg5 gene is predicted to encode a protein with novel structure containing three R-gene domains: a nucleotide binding site (NBS), leucine rich repeat (LRR), and serine threonine protein kinase (STPK). The Rpg5 predicted protein structure suggests a typical R-gene function in recognition of the pathogen. Rpg5 confers resistance to an isolate of rye stem rust independent of rpg4, but resistance to several races of wheat stem rust requires both genes. The rpg4 gene is predicted to encode an actin depolymerizing factor-like (ADF) protein. The dependency on the actin modifying protein ADF2 indicates cytoskeleton reorganization or redirec-tion plays a role in the pathogen-host interaction. Rpg5 could potentially interact with rpg4 to activate or deactivate its function in the resistance response. Alternatively, Rpg5 may initiate signal transduction leading to resistance in response to detecting RPG4 (ADF2) protein modification. The pathogen may modify ADF2, via effector molecules, to interfere with the resistance reaction or redirect the actin cytoskeleton. The redirection of actin could possibly enable the pathogen to develop a haustoria-plant cell cytoskeleton interface for acquisition of nutrients.
Stem Rust Resistance Genes in Barley
Wheat stem rust, caused by the pathogen Puccinia graminis f. sp. tritici (Pgt), was a devastating disease of wheat and barley prior to the 1950’s.4 Additionally, the rye stem rust pathogen. P.g. f. sp. secalis (Pgs) also can attack and cause losses in barley. In the US and Canada, durable resistance in barley was achieved through the deployment of varieties with the single resistance gene Rpg1, while in wheat it was achieved by pyramiding several stem rust resis-tance genes in one cultivar. Since the deployment of this genetic resistance in barley and wheat in the early 1940s and mid-1950s, respectively, losses due to stem rust have been minimal, and wide-spread stem rust epidemics were considered by many to be a thing of the past until recent appearance of Pgt race TTKSK (aka isolate Ug99) in Uganda in 1999.5
The barley Rpg1 gene is remarkably durable and confers resis-tance to most races of Pgt; however, some races of Pgt and some isolates of Pgs are virulent on barley cultivars containing Rpg1. A
*Correspondence to: Andris Kleinhofs; Washington State University; Crop and Soil Sciences; Johnson Hall 259; Pullman, Washington 99164-6420 USA; Tel.: 509.335.4389; Email: [email protected]
Submitted: 02/02/09; Accepted: 02/04/09
Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/8079
Extra View
The rpg4/Rpg5 stem rust resistance locus in barleyResistance genes and cytoskeleton dynamics
Robert Brueggeman,1 Brian J. Steffenson2 and Andris Kleinhofs1,*
1Department of Crop and Soil Science; Washington State University; Pullman, Washington USA; 2Department of Plant Pathology; University of Minnesota; St. Paul, Minnesota USA
Key words: stem rust, barley, disease resistance genes, actin depolymerizing factor, cytoskeleton
Thi
s m
anus
crip
t ha
s be
en p
ublis
hed
onlin
e, p
rior
to
prin
ting
. Onc
e th
e is
sue
is c
ompl
ete
and
page
num
bers
hav
e be
en a
ssig
ned,
the
cit
atio
n w
ill c
hang
e ac
cord
ingl
y.
[Cell Cycle 8:7, 1-5; 1 April 2009]; ©2009 Landes Bioscience
82

www.landesbioscience.com Cell Cycle 2
Pgt race with virulence for Rpg1 (designated QCCJ) was identi-fied on barley in the Midwestern United States in 1989.6 Pgt QCCJ ultimately became one of the most common virulence types in the United States and caused local epidemics in both the US and Canada.7 The best source of resistance to race QCCJ was discovered in the unimproved barley line Q21861.8 Resistance in Q21861 is governed by a single gene rpg4, which is temperature sensitive and recessive in nature.8,9 Genetic mapping localized rpg4 to the long arm of barley chromosome 5H.10
Another resistance gene (designated Rpg5) conferring resistance to Pgs isolate 92-MN-90, also is present in line Q21861 and was mapped to the long arm of barley chromosome 5H, co-segregating with rpg4. Resistance to Pgs isolate 92-MN-90 was initially thought to be due to rpg4,11 but the recent cloning of the locus has shown that rpg4 and Rpg5 mediated resistance is distinct.3 The rpg4/Rpg5 locus also confers resistance to Pgt races MCCF and RCRS.12 Rpg1 is generally not effective against Pgs isolates like 92-MN-90.
The newly emerged Pgt race TTKSK (aka isolate Ug99) can attack nearly 70% of the world’s wheat varieties5 and is considered a threat to global food security.13 Moreover, race TTKSK also carries virulence for most barley varieties, including those carrying Rpg1.14 Fortunately, the rpg4/Rpg5 locus in line Q21861 was recently shown to confer resistance to race TTKSK (Steffenson B, unpublished).
Rpg5 has a Novel Gene Structure Containing all Three Major R-Gene Domains
Innate receptors or receptor complexes act as molecular switches that, once triggered by the pathogen, set into motion signaling events resulting in two different known defense mechanisms. The first line of defense, the major component of non-host resistance and a form of basal resistance, arrests the majority of invading pathogen units at the cell periphery.15 Pathogens that have adapted to circumvent the basal resistance encounter a second line of defense: the race specific, R-gene mediated resistance, associated with the HR and leading to localized cell death.
The barley R-gene, Rpg5, has a novel gene structure containing an N-terminal NBS, internal LRR, and C-terminal STPK with two predicted trans-membrane domains.3 Many studies investi-gating the molecular determinants of plant disease resistance have revealed that different combinations and polymorphisms of these three distinct protein domains (NBS, LRR and STPK) comprise the vast majority of pathogen recognition receptors or compo-nents of receptor complexes for R-gene mediated resistance.16 The Rpg5 gene structure appears to be novel when compared to the Arabidopsis, Brachypodium and Oryzae genomes. PCR reactions using several Rpg5 specific primers on genomic DNA and cDNA from several varieties of wheat and the wild wheat relative Triticum timopheevi identified NBS-LRR and STPK genes highly homologous to Rpg5, but did not detect genes or transcripts linking all three domains in a single gene (Brueggeman R, et al. unpublished). These data suggest that the Rpg5 structure combining the NBS, LRR and STPK domains probably evolved after wheat and barley split about 10 mya17 and may be unique to Hordeum.
The R-genes cloned to date have been grouped into different classes according to their protein domain structure.16 The major class of R-genes possess an N-terminal NBS and a C-terminal LRR and are designated NBS-LRR R-genes. In relatively few cases, NBS-LRR genes have been shown to specifically interact with pathogen avirulence (Avr) gene products.18-21 The guard hypoth-esis is an alternative to direct R-gene/Avr-gene interaction.22 In the guard hypothesis, the R-genes act as surveillance proteins detecting modification of host target proteins by pathogen effectors. The direct or indirect interaction between the host R-gene protein and pathogen AVR protein is believed to activate the signaling cascades resulting in HR and disease resistance. These signaling cascades may involve phosphorylation via the second major class of R-genes, the protein kinases.
Six R-genes containing at least one STPK domain make up the second major R-gene class. The STPK R-genes confer resistance to bacterial and fungal pathogens and have been cloned from a wide taxa of plant species.3,23-27 The STPK domain suggests that these proteins function in plant defense signal transduction pathways involving phosphorylation. However, STPK domain-containing genes are also known to interact directly with Avr genes, demonstrating a capacity to recognize non-self proteins through protein-protein interactions.28
Tomato Pto-mediated resistance against Pseudomonas syringae is one of the most extensively studied and understood mechanisms of disease resistance. Interestingly, Pto requires the presence of a second gene Prf (an NBS-LRR gene) for resistance.29 Functional analysis of the Pto/Prf system indicated that specific mutations in the activation loop of the Pto kinase induced spontaneous symptoms similar to HR observed when plants expressing Pto are infected with AvrPto containing strains of P. syringae. The sponta-neous HR response was AvrPto independent, but Prf dependent30 suggesting that Prf acts downstream or coincident with Pto. It was later demonstrated that Pto and Prf do act coincidently in the signaling pathway, and that the relationship is dependant on a physical interaction between the two proteins. In this resistance complex, Pto acts as a regulatory subunit of Prf, and Prf contrib-utes to the pathogen recognition specificity of Pto.31 Resistance against P. syringae strains that carry AvrPphB also requires two Arabidopsis genes, RPS5, a NBS-LRR gene, and PBS1, a serine threonine protein kinase gene.24 These systems indicate that resis-tance protein complexes require both an NBS-LRR and STPK protein for pathogen recognition and resistance.
The Rpg5 mechanism of pathogen recognition and signaling pathway may parallel Pto. The Pto-mediated resistance requires both NBS-LRR and STPK genes for resistance, and the RPG5 predicted protein structure contains all three NBS, LRR and STPK domains with significant amino acid homology to both Pto and Prf (kinase homology to Pto is 36% aa identity and 53% aa simi-larity and the NBS-LRR homology to PRF is 26% aa identity and 42% aa similarity). The RPG5 protein structure suggests that the three domains act coincident with one another in barley, similar to tomato, and are required for Rpg5 mediated resistance to Pgs isolate 92-MN-90. The RPG5 protein with all three domains in a single protein may facilitate the identification of other components
Stem rust resistance and cytoskeleton dynamics
83

Stem rust resistance and cytoskeleton dynamics
3 Cell Cycle 2009; Vol. 8 Issue 7
reaction to Pgt QCCJ. Since both positions involve amino acids that could be phosphorylated, i.e., either T added or S replaced, it suggests that phosphorylation may be involved in regulating ADF2 function. All Adf2 alleles tested were expressed at the transcription level, indicating that these two amino acid polymorphisms deter-mine susceptibility/resistance to Pgt QCCJ.
The Role of Actin Depolymerizing Factors in Resistance
During pathogen attack by both fungi and bacteria, the plant cytoskeleton dynamically rearranges.2 ADF proteins, in concert with other actin binding proteins, regulate actin filament dynamics.32 ADFs are believed to function by increasing the turn-over of filamentous F-actin by depolymerization of monomeric G-actin from the pointed ends of the filaments.33 In addition, ADFs may also nucleate the assembly of new actin filaments.32 Many interactions between the host and pathogen may be depen-dant on ADF proteins, including the cytoskeletal polarization in response to non-host pathogens.34 Cytoskeleton polariza-tion in response to pathogen challenge is well documented;1,2 however, very little is known concerning the signaling processes that mediate this response. Studies with the barley mlo non-race specific resistance system conferring resistance to Blumeria graminis f. sp. hordei using actin cytoskeleton pharmacological inhibitors and genetic interference by overexpressing a barley actin depolymerizing factor gene (HvAdf3), demonstrated that actin cytoskeleton function was required for basal defense against an appropriate mildew pathogen and for mlo-mediated non-race specific resistance at the cell wall. However, actin function was not required for several race specific immune responses. It was also demonstrated that actin cytoskeletal disruption led to increased fungal penetration by non-host pathogens.35
Chemical or genetic disruption of actin polymerization results in greater efficiency of non-host pathogen penetration; however, in tobacco cytoskeleton perturbation can also prime the cell for HR.36 It was proposed that the host detects disruption of the actin cytoskeleton similar to the guard hypothesis, thus triggering the HR response. Evidence is mounting that the actin cytoskeleton is the target of plant pathogen effectors to suppress non-race specific resistance, which may signal to induce a second line of defense mediated by the pathogen specific R-genes.37 There is also evidence suggesting non-host pathogens that evade the first line of defense and enter the cell are met by an HR mediated resistance that could be due to several simultaneously acting R-genes.38
ADF proteins possibly play an important role in response to pathogen challenge. It is plausible that ADF proteins are poten-tial effector targets of fungi to circumvent a defense response. Previous studies have shown that cytoskeleton disruption was only compromised in race non-specific and non-host resistance. This begs the question: is rpg4-mediated resistance a form of race non-specific resistance? Our data indicate that rpg4 functions to confer resistance to a number of different Pgt races, but not to all. Therefore it appears to have race specificity, but only in the pres-ence of the typical R-gene Rpg5. In the absence of Rpg5, the rpg4 gene may provide some non-host or non-race specific resistance to various pathogens.
of the resistance complex or the pathogen AVR protein by the yeast two-hybrid assay that otherwise may be missed when using an STPK or NBS-LRR gene as bait separately. It is plausible that very few identified AVR proteins interact with their corresponding NBS-LRR R-genes due to incomplete interaction because the resistance complex requires both STPK and NBS-LRR proteins for complete interaction. Many of the NBS-LRR R-genes identified may have an STPK counterpart that has not been identified due to lack of polymorphism.
The rpg4 Gene Encodes an Actin Depolymerizing Factor-Like Protein
Genetic mapping and allele analysis using two barley popula-tions containing HvAdf2 polymorphisms (Steptoe x Q21861 and MD2 x Q21861) identified recombinants that delimited the rpg4 region to 1kbp. This 1kbp region contains only the HvAdf2 gene, indicating that it is the rpg4 gene.3 However, the barley variety Harrington and line Sm89010, which are susceptible to Pgt QCCJ, contain an rpg4 allele identical at the amino acid level to the rpg4 allele in Q21861, which is resistant to Pgt QCCJ. This suggested that HvAdf2 was not the rpg4 gene. Further analysis of allele sequences of rpg4 and Rpg5 together with SNP analysis of the recombinants suggested that a functional Rpg5 may be required in addition to rpg4 for resistance to Pgt QCCJ. The Pgt QCCJ susceptible barleys Harrington and Sm89010 with rpg4 alleles identical to the resistant Q21861 rpg4 allele, do not contain a functional Rpg5 allele. The Harrington x Q21861 recombinant #18 combines the Harrington rpg4 allele with the Q21861 Rpg5 gene, and it is resistant to Pgt QCCJ, indicating that cultivar Harrington is susceptible to this race due to a lack of a functional Rpg5 (Brueggeman R, et al. unpublished). These data suggest that Pgt QCCJ resistance should behave as a single dominant gene in populations where only the Rpg5 gene is segregating in the presence of a non-polymorphic rpg4 gene. This hypothesis is being tested in the Harrington x Q21861 and Sm89010 x Q21861 F1 and F2 populations and by down-regulating the two alternative rpg4 alleles (Brueggeman R, et al. unpublished).
Phenotype analysis of three populations (Morex, Steptoe and Robust x Q21861) revealed 1:3 segregation for resistance : suscep-tibility, demonstrating the recessive nature of rpg4.8 The rpg4 and Rpg5 genes are tightly linked (~40 kbp physically) adding to the complexity of the two-gene requirement for resistance. The reces-sive behavior of rpg4 implies that it encodes a non-functional ADF2 protein, resulting in resistance or that the functional Rpg4 gene encoding a functional ADF2 protein acts as a susceptibility factor in the reaction with Pgt race QCCJ. The three QCCJ suscep-tible barley cultivars (Morex, Steptoe and Robust) have identical Adf2 alleles at the amino acid level and differ from the resistant Q21861 allele by only three amino acids (Q39H, A101T, S135G). Three Pgt QCCJ susceptible progeny from the Steptoe x Q21861 population have recombinations within the Adf2 gene and contain a functional Rpg5 allele. These recombinant Adf2 genes contain Q39 (Q21861-like) with T101 and G135 (Steptoe-like), showing that the amino acids at positions 101 and/or 135 are important for
84

www.landesbioscience.com Cell Cycle 4
Stem rust resistance and cytoskeleton dynamics
alleles are expressed at the mRNA level, although we do not yet know if this is true at the protein level. The differential interaction between Rpg4 or rpg4 encoded proteins with Rpg5 could be modu-lated by the fungus or Rpg5 could modulate their interaction with the fungus. In order to satisfy the requirement that the rpg4 gene action is recessive, the presence of Rpg4 encoded ADF2 molecules must lead to susceptibility, while rpg4 encoded ADF2 must have a neutral or negative function.
It is well documented in animal pathogenic bacteria that pathogens produce proteins that disrupt actin cytoskeleton func-tion in order to become adapted to the host and thus become pathogenic.44,45 There is little evidence of plant pathogenic fungi producing effector molecules that disrupt actin dynamics; however, many genera of fungi produce cytochalasins the major chemical used for pharmacological inhibition of actin filament function.46 There is precedence of a bacterial plant pathogen secreting the effector, AvrPto, into the host cytoplasm, which disrupts the cytoskeleton polarization response. AvrPto protein has been shown to suppress callose deposition at the site of Pseudomonas syringae challenge, a cytoskeleton dependant response, leading to loss of basal resistance in Arabidopsis.47 Rpg5 could also be the guard protein monitoring the ADF2 protein for any modifications by the fungus effector; however, this would not explain why rpg4 is recessive. Rpg5 should still be able to guard ADF2 even when the rpg4 gene is present in a heterozygous state.
The rpg4/Rpg5 resistance system is the first link between an R-gene that putatively recognizes a race specific pathogen and an actin binding protein, which may be a link between the cytoskel-eton dynamics occurring post pathogen interaction.
The Next Step
The molecular basis of disease resistance appears very complex, and although progress has been made in identifying many components there are still key questions unanswered. A complete signal transduction pathway leading to fungal pathogen recogni-tion has yet to be identified and little is understood about how resistance genes activate defense signaling. It is apparent from the R-gene literature that different resistance pathways are acti-vated by pathogen recognition, and there is probably cross talk and common components involved among them. We are lacking answers to some of the key questions regarding the rpg4/Rpg5-mediated mechanism of resistance that may help answer questions regarding receptor activation and dynamic cytoskeleton rearrange-ment. Our future research will focus on addressing some of the questions regarding how and at what level rpg4 and Rpg5 interact to confer resistance to stem rust.
The rpg4/Rpg5 locus may answer some questions about disease resistance signaling. The novel gene structure of Rpg5 may provide answers to why NBS-LRR and STPK proteins interact and are required components of diverse R-gene receptor complexes. The interaction between rpg4 and Rpg5, whether direct or indirect, may also provide answers to how pathogen recognition signals the rapid rearrangement of the actin cytoskeleton and what role this rearrangement plays in the huastoria-plant cell interface. It may also be the first system where we have a pathogen effector target
Rpg5 and rpg4 Interact to Confer Stem Rust Resistance
In all known Pgt QCCJ resistant barley varieties and recombi-nants, it appears that the full resistance function of rpg4 requires the presence of a functional Rpg5 gene. We hypothesize that Rpg5 is the R-gene in this system that detects the Pgs and Pgt pathogens. Rpg5 mediated resistance to Pgs isolate 92-MN-90 is independent of rpg4. This leads to the speculation that Rpg5 may operate in two distinct resistance pathways, one that is similar to most race-specific R-gene mediated resistance mechanisms and is independent of actin reorganization events mediated by rpg4. The second pathway may be more like a non-race specific type of resistance mechanism dependant on rpg4-mediated actin reorganization. Like the Pto/Prf system, Rpg5 may be capable of recognizing more than one Puccina graminis derived AVR protein, leading to differential reactions and activation of different resistance pathways.
It is plausible that Rpg5 interacts with rpg4 directly or indirectly to activate or deactivate the actin depolymerizing factor through phosphorylation. As discussed earlier, the resistant and susceptible alleles differ by two amino acids that are both potential phosphory-lation sites. It has also been shown that ADFs can be deactivated or activated by phosphorylation or dephosphorylation, respectively, via protein kinases and phosphotases.39,40 This activation or deac-tivation may lead to an incompatible or compatible reaction with pathogens. Where the actin reorganization occurs and how R-gene mediated resistance is dependant on this reorganization still needs to be elucidated.
An attractive hypothesis is that the invading fungus actually captures the function of the cytoskeleton to feed itself. In this case, a non-functional adf gene that the fungus targets may prevent the fungus from becoming established within the plant. Miklis and co-workers have suggested that although actin disruption initially enhanced pathogen entry into the cell, it may have a long-term negative effect on the compatible interaction.1 It has been shown that in the compatible interaction between barley and powdery mildew, the haustoria (fungal feeding organs) are associated with actin filament rings, and actin filaments closely follow haustoria when invaginating the plasma membrane.41 This is similar to what occurs in the symbiotic interaction between a mycorrhizal fungus and tobacco root cells,42 suggesting similarity between how pathogenic and symbiotic fungi may establish the fungus-host interface for nutrient acquisition. The host cytoskeleton rearrangement machinery, initially utilized for resistance, may be redirected by the pathogen to establish a feeding mechanism that is actin filament-dependant.1,43 It is possible that functional Adf2 enables the fungus to produce or maintain this feeding structure, thus facilitating fungus growth or that non-functional or inap-propriately functional adf2 disrupts the actin filaments such that the fungus cannot maintain the feeding structure resulting in incompatibility.
A complete model for the function of rpg4 and Rpg5 in Pgt resistance must take into account the observations that these two genes work together and that the rpg4 gene function is recessive and temperature sensitive. It is, of course, possible that each gene functions independently, but weakly and both are required for a strong resistance phenotype. It should be noted that both Rpg4
85

Stem rust resistance and cytoskeleton dynamics
5 Cell Cycle 2009; Vol. 8 Issue 7
29. Salmeron JM, Oldroyd GED, Rommens CMT, Scofield SR, Kim H-S, Lavelle DT, et al. Tomato Prf is a member of the leucine-rich repeat class of plant disease resistance genes and lies embedded within the Pto kinase gene cluster. Cell 1996; 86:123-33.
30. Rathjen JP, Chang JH, Staskawicz BJ, Michelmore RW. Constitutively active Pto induces a Prf-dependant hypersensitive response in the absence of AvrPto. EMBO J 1999; 18.
31. Mucyn TS, Clemente A, Andriotis VME, Balmuth AL, Oldroyd GED, Staskawicz BJ, et al. The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate spe-cific plant immunity. Plant Cell 2006; 18:2792-806.
32. Dos Remedious CG, Chhabra D, Kekic M, Dedova IV, Tsubakihara M, Berry DA, et al. Actin binding proteins: regulation of cytoskeletal microfilamants. Physiol Rev 2003; 83:433-73.
33. Carlier MF, Laurent V, Santolini J, Melki R, Didry D, Xia GX, et al. Actin depolymeriza-tion factor (ADF/cofilin) enhances the rate of filament turnover: implication in actin-based motility. J Cell Biol 1997; 136:1307-22.
34. Skalamera D, Heath MC. Changes in the cytoskeleton accompanying infection-induced nuclear movements and hypersensitive response in plant cells invaded by rust fungi. Plant J 1998; 16:191-200.
35. Miklis M, Consonni C, Bhat RA, Lipka V, Schulze-Lefert P, Panstruga R. Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Phys 2007; 144:1132-43.
36. Kobayashi Y, Kobayashi I. Depolymerization of the actin cytoskeleton induces defense responses in tobacco plants. J Gen Plant Pathol 2007; 73:360-4.
37. Jones JDG, Dangl JL. The plant immune system. Nature 2006; 444:323-9. 38. Thordal-Christensen H. Fresh insights into processes of nonhost resistance. Curr Opin
Plant Biol 2003; 6:351-7. 39. Niwa R, Nagata-Ohashi K, Takeichi M, Mizuno K, Uemura T. Control of actin reorga-
nization by slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 2002; 108:233-46.
40. Nakano K, Kanai-Azuma M, Kanai Y, Moriyama K, Yakaki K, Hayashi Y, et al. Cofilin phosphorylation and actin polymerization by NRK/NESK, a member of the germinal center kinase family. Exp Cell Res 2002; 287:219-27.
41. Opalski KS, Schultheiss H, Kogel K-H, Huckelhoven R. The receptor-like MLO pro-tein and the RAC/ROP family G-protein RACB modulate actin reorganizatio in barley attacked by the biotrophic powdery mildew fungus Blumeria graminis f. sp. hordei. Plant J 2005; 41:291-303.
42. Genre A, Bonfante P. Actin versus tubulin configuration in arbuscule-containing cells from mycrorrizal tobacco roots. New Phytol 1998; 140:745-52.
43. Takemoto D, Hardham AR. The cytoskeleton as a regulator and target of biotic interac-tions in plants. Plant Phys 2004; 136:3864-76.
44. Cornelis GR. The Yersinia Ysc-Yop type III weaponry. Nat Rev Mol Cell Biol 2002; 3:742-52.
45. Buttner D, Bonas U. Common infection strategies of plant and animal pathogenic bacteria. Curr Opin Plant Biol 2003; 6:312-9.
46. Thomas DD. Cytochalasin effects in plants and eukaryotic microbial systems. In: Tanenbaum SW, ed. Cytochalisins: biochemical and cell biological aspects. New York: Elsevier, North Holland Biomedical Press 1978; 257-75.
47. Hauck P, Thilmony R, He SY. A Pseudomonas syringae type III effector suppresses cell wall-based extracellular defense in susceptible Arabidopsis plants. Proc Natl Acad Sci USA 2003; 100:8577-82.
that a fungus acts upon in order to perturb cytoskeleton depen-dant resistance.
References 1. Lipka V, Panstruga R. Dynamic cellular responses in plant-microbe interactions. Curr
Opin Plant Biol 2005; 8:625-31. 2. Schmelzer E. Cell polarization, a crucial process in fungal defense. Trends in Plant Sci
2002; 7:411-5. 3. Brueggeman R, Druka A, Nirmala J, Cavileer T, Drader T, Rostoks N, et al. The stem
rust resistance gene Rpg5 encodes a novel protein with nucleotide binding site, leucine-rich and protein kinase domains. Proc Natl Acad Sci USA 2008; 105:14970-5.
4. Steffenson BJ. Analysis of durable resistance to stem rust in barley. Euphytica 1992; 63:153-67.
5. Singh RP, Hodson DP, Huerta-Espino J, Jin Y, Njau P, Wanyera R, et al. Will stem rust destroy the world’s wheat crop? Adv Agron 2008; 98:271-309.
6. Roelfs AP, Casper DH, Long DL, Roberts JJ. Races of Puccinia graminis in the United States in 1989. Plant Dis 1991; 75:1127-30.
7. Roelfs AP, Long DL, Roberts JJ. Races of Puccinia graminis in the United States during 1991. Plant Dis 1993; 77:129-32.
8. Jin Y, Steffenson BJ, Fetch TGJ. Sources of resistance to pathotype QCC of Puccinia graminis f. sp. tritici in barley. Crop Sci 1994; 34:285-8.
9. Jin Y, Steffenson BJ, Miller JD. Inheritance of resistance to pathotypes QCC and MCC of Puccinia graminis f. sp. tritici in barley line Q21861 and temperature effects on the expression of resistance. Phytopathology 1994; 84:452-5.
10. Borovkova IG, Steffenson BJ, Jin Y, Rasmussen JB, Kilian A, Kleinhofs A, et al. Identification of molecular markers linked to the stem rust resistance gene rpg4 in barley. Phytopathology 1995; 85:181-5.
11. Sun Y, Steffenson BJ, Jin Y. Genetics of resistance to Puccinia graminis f. sp. secalis in barley line Q21861. Phytopathology 1996; 86:1299-302.
12. Sun Y, Steffenson BJ. Reaction of barley seedlings with different stem rust resistance genes to Puccinia graminis f. sp. tritici and P.g. f. sp. secalis. Can J Plant Pathol 2005; 27:80-9.
13. Stokstad E. Deadly wheat fungus threatens world’s breadbaskets. Science 2007; 315:1786-7.
14. Steffenson BJ, Jin Y. Resistance to race TTKS of Puccinia graminis f. sp. tritici in barley. Phytopathology 2006; 96:110.
15. Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Olu J-L, et al. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 2003; 425:973-7.
16. Martin GB, Bogdanove AJ, Sessa G. Understanding the function of plant disease resis-tance proteins. Annu Rev Plant Biol 2003; 54:23-61.
17. Wolfe KH, Gouy M, Yang YW, Sharp PM, Li WH. Date of the monocot-dicot diver-gence estimated from chloroplast DNA sequence data. Proc Natl Acad Sci USA 1989; 86:6201-5.
18. Dodds PN, Lawrence GJ, Catanzariti A-M, Teh T, Ching I, Wang A, et al. Direct protein interaction underlies gene-for-gene specificity and co-evolution of the flax resistance genes and flax rust avirulence genes. Proc Natl Acad Sci USA 2006; 103:8888-93.
19. Deslandes L, Olivier J, Peeters N, Feng DX, Khounlotham M, Boucher C, et al. Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc Natl Acad Sci USA 2003; 100:8024-9.
20. Jia J, McAdams SA, Bryan GT, Hershey HP, Valent B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J 2000; 19:4004-14.
21. Ueda H, Yamaguchi Y, Sano H. Direct interaction between the tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor during the hypersensi-tive response in tobacco plants. Plant Mol Biol 2006; 61:31-45.
22. Van Der Bizen EA, Jones JDG. Plant disease-resistance proteins and the gene-for-gene concept. Trends Biochem Sci 1998; 23:454-6.
23. Martin GM, Brommonschenkel SH, Chunwongse J, Frary A, Ganal MW, Spivey R, et al. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 1993; 262:1432-6.
24. Swiderski MR, Innes RW. The Arabidopsis PBS1 resistance gene encodes a member of a novel protein kinase subfamily. Plant J 2001; 26:101-12.
25. Song W-Y, Wang G-L, Chen L-L, Kim H-S, Pi L-Y, Gardner J, et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995; 270:1804-6.
26. Brueggeman R, Rostoks N, Kudrna D, Kilian A, Han F, Chen J, et al. The barley stem rust-resistance gene Rpg1 is a novel disease-resistance gene with homology to receptor kinases. Proc Natl Acad Sci USA 2002; 99:9328-33.
27. Sun X, Cao Y, Yang Z, Xu C, Li X, Wang S, et al. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J 2004; 37:517-27.
28. Tang X, Frederick RD, Zhou J, Halterman DA, Yia Y. Initiation of plant disease resis-tance by physical interaction of AvrPto and Pto kinase. Science 1996; 274:2060-3.
86

SUMMARY AND FUTURE DIRECTION
The focus of my doctoral research was to clone and characterize the genes underlying the
rpg4/Rpg5 stem rust resistance locus. This locus is very important because it is the best source of
resistance conferring resistance to races virulent on Rpg1, the only source of resistance deployed
in Midwestern barley cultivars. We cloned the Rpg1, rpg4 and Rpg5 genes by a map-based
approach and performed functional analysis of the RPG1 protein, gene family characterization
and elucidation of the resistance pathways. Rpg1 and Rpg5 both have novel R-gene structures
containing atleast one STPK domain with considerable homology to the Pto kinase. Therefore
what is known about both the Pto and Rpg1 mediated mechanisms will be used to drive the
functional analysis of the rpg4/Rpg5 mediated resistance mechanisms. The cytoskeleton
rearrangement aspect of the rpg4/Rpg5 mechanism appears to have far reaching scientific
importance beyond the average race specific resistance responses.
Functional analysis of the rpg4 and Rpg5 genes is in its infancy but with our Rpg1
research leading the way many techniques and tools needed are already in hand and
standardized. Although the rpg4/Rpg5 resistance mechanism is probably somewhat different
than Rpg1 it is currently the best model for gaining insight into the Rpg5 mechanism. We have
already discovered pieces of information about the rpg4/Rpg5 mechanism that will help in the
efforts to elucidate how these genes interact to confer resistance. We have interpreted rpg4 as a
disease resistance gene but recent data suggests that this designation may not fit. After
sequencing and analyzing the alleles from resistant and susceptible cultivars a more valid
interpretation maybe that rpg4 is a susceptibility factor present within some of the susceptible
cultivars. Analyzing the sequence between the different rpg4 and Rpg5 alleles along with the
interpretation of the recessive heritability and the populations used to determine this have shed
87

88
some light on the possible function of rpg4 and may explain why rpg4 inheritance is recessive.
Future research will be focused on how and at what level rpg4 and Rpg5 interact. I believe the
answers to these questions will provide great insight into how biotrophic fungal pathogens
maintain their life style within the plant host and will have great utility in future disease
management strategies.
Stem rust has reemerged as an important disease that has the potential to cause severe
barley and wheat yield losses and possibly could threaten world food security.
In order to keep the stem rust disease under control in an effective, sustainable and
environmentally friendly manner resistance genes must be deployed responsibly. Barley
production should not depend on the deployment of a limited number and sources of resistance
genes as is present situation with Rpg1 in the USA and Canada. Several sources of resistance
should be pyramided such that the genes will operate in an effective durable manner. This
depends on the identification and utilization of new sources of resistance. It also makes the
understanding of the mechanisms underlying these resistance genes important so this basic
scientific knowledge can be applied in the field to deploy R-genes for maximum effectiveness
and durability.





























