Embed Size (px)
Citation preview

Investigating Spelling Impairment and Changes related to Intervention by means of Functional MRI and DTI
Doctoral Thesis
submitted by
Mag. rer. nat. Daniela Gebauer
Supervisor: Ass. Prof. PD. Dr. Mag. Andreas Fink
Supervisor: Assoc. Prof. PD. Dr. Christian Enzinger

2
Acknowledgement
First and foremost I offer my regards and blessings to all who supported me in any respect
during the completion of the thesis.
I offer my sincerest gratitude to my supervisor Ass.-Prof. Dr. Andreas Fink, who has
supported me throughout my thesis with his guidance and dedication whilst allowing me the
room to work in my own way. He provided me unflinching encouragement and support.
Many thanks go in particular to my supervisor Assoc. Prof. Dr. Christian Enzinger for his
valuable support, guidance and encouraging advices. I am much indebted to him for his
valuable knowledge and assistance.
I am heartily thankful to my colleagues of the Research Unit for “Neuronal Plasticity and
Repair” and my colleagues of the “Research Unit for NeuroImaging”: Marisa Loitfelder,
Margit Jehna, Patricia Linortner, Christian Langkammer, Michaela Söllinger und Stefan
Ropele for their constructive comments and enriching cooperation.
Moreover, I gratefully thank all professors, colleagues and staff members, who supported the
work on this thesis: Prof. Franz Fazekas and Karin Brodtrager (Department of Neurology);
Prof. Franz Ebner (Department of Neuroradiology); Mag. Karl Koschutnig and Dr. Gernot
Reishofer (assistance in the development of the functional paradigm); Dr. Reinhard Kargl and
Christian Purgstaller (creators of the spelling intervention); Heidi Johansen-Berg and Nicola
Filippini (University of Oxford, supporting DTI analyses) and Nadja Kozel, Bernd
Schneeberger, Hanna Vogel and Stephanie Rohrer (behavioural testing and training of
spelling impaired children).
Furthermore, I most sincerely thank the Division of Science and Research of the Styrian
government and the Jubilee Fund of the Austrian National Bank for the financial support of
this doctoral thesis.
Most especially I thank my parents for unconditionally supporting me for my whole life and
Oliver Pinter for his continuous support and love.

3
TABLE OF CONTENTS
ABSTRACT............................................................................................................................... 5
ZUSAMMENFASSUNG........................................................................................................... 6
I INTRODUCTION ................................................................................................................... 7
1. SPELLING AND READING IMPAIRMENT (SRI) ........................................................ 7
1.1. Etiology ....................................................................................................................... 7
2. NEUROPHYSIOLOGY OF SRI ....................................................................................... 8
2.1. Functional MRI and SRI ............................................................................................. 9
2.2. Diffusion Tensor Imaging and SRI ........................................................................... 10
3. ISOLATED SPELLING IMPAIRMENT ........................................................................ 12
3.1. Neurophysiology of Spelling Impairment................................................................. 12
4. INTERVENTION ............................................................................................................ 13
4.1. Spelling Intervention ................................................................................................. 14
4.1.1. Morpheme-based intervention............................................................................ 14
4.1.2. Computer-based intervention ............................................................................. 15
4.1.3. Morpheus-intervention....................................................................................... 15
4.2. Neurophysiologic changes related to intervention .................................................... 17
II OBJECTIVES AND METHODS......................................................................................... 21
III STUDIES ............................................................................................................................ 25
STUDY I – Isolated spelling impairment ............................................................................ 25
Abstract ............................................................................................................................ 26
1. Introduction .................................................................................................................. 27
2. Method ......................................................................................................................... 30
3. Results .......................................................................................................................... 35
4. Discussion .................................................................................................................... 40
5. Conclusion.................................................................................................................... 42
STUDY II – Functional changes related to intervention...................................................... 43
Abstract ............................................................................................................................ 44
1. Introduction .................................................................................................................. 45
2. Materials and Methods ................................................................................................. 46
3. Results .......................................................................................................................... 51
4. Discussion .................................................................................................................... 57

4
STUDY III – Structural Changes related to intervention..................................................... 60
Abstract ............................................................................................................................ 61
1. Introduction .................................................................................................................. 62
2. Methods........................................................................................................................ 64
3. Results .......................................................................................................................... 68
4. Discussion .................................................................................................................... 74
5. Conclusion.................................................................................................................... 78
IV GENERAL DISCUSSION AND CONCLUSIONS........................................................... 79
V REFERENCES..................................................................................................................... 86
VI APPENDIX......................................................................................................................... 98
Appendix Study I ................................................................................................................. 98
Appendix Study II .............................................................................................................. 103
VII LIST OF ABBREVIATIONS.......................................................................................... 106

ABSTRACT
5
ABSTRACT Approximately 3-15% of all children show reading and spelling difficulties. Research into
this topic mainly focused on the neural correlates of reading impairments, whereas spelling
impairments have been largely neglected so far. Hence, the aim of this doctoral project was to
investigate brain structure and function in children with spelling difficulties and whether a
specific spelling intervention would also be associated with changes in functional patterns of
brain activity and structural parameters. Specifically, this doctoral thesis is composed of three
different studies:
In our first study, we investigated structural and functional characteristics of the brain in
children with isolated spelling impairment, compared to children with spelling and reading
impairment and non-impaired controls, by means of functional magnetic resonance imaging
(MRI) and diffusion tensor imaging (DTI). We provided evidence that children with isolated
spelling impairment exhibited increased right hemispheric activation in the absence of
structural differences compared to controls.
In our second study, we investigated the effects of a morpheme-based spelling intervention
on functional patterns of brain activity in 20 German-speaking spelling impaired children
(divided into a training- and a waiting group) using repeated functional MRI. We found that
relative to 10 matched controls, children with poor spelling abilities showed increased
activation in frontal medial and right hemispheric regions and decreased activation in left
occipito-temporal regions prior to the intervention. After five weeks of intervention, spelling
and reading comprehension significantly improved in the training group, along with increased
activation in left temporal and (para)hippocampal regions.
In our third study , we investigated the effects of a spelling intervention on white matter
integrity in 20 spelling impaired children using repeated DTI. Results generally suggested
that after five weeks of intervention, spelling ability improved in the training group, along
with right hemispheric increases in white matter integrity compared to controls.
The main findings of this doctoral project can thus be briefly summarized as follows:
First, children with spelling impairment exhibit a stronger right hemispheric activation
compared to non-impaired controls. Secondly, successful intervention is associated with
changes in brain structure and function.

ZUSAMMENFASSUNG
6
ZUSAMMENFASSUNG Schätzungsweise 3-15% aller Kinder weisen Schwierigkeiten beim Lesen und Rechtschreiben
(LRS) auf. Die bisherige neurowissenschaftliche Forschung konzentrierte sich primär auf die
Leseschwäche (LS), wohingegen nur wenige Studien über die spezifischen neuronalen
Charakteristika von Kindern mit Rechtschreibschwäche (RS) existieren. Deshalb war es das
Ziel der vorliegenden Doktorarbeit, die Struktur und Funktion des Gehirns bei Kindern mit
RS näher zu untersuchen und zu überprüfen, ob eine gezielte Intervention mit Veränderungen
in der Struktur und Funktion des Gehirnes assoziiert sein könnte.
Das vorliegende Dissertationsprojekt umfasste drei Studien:
In unserer ersten Studie untersuchten wir mittels funktioneller Magnetresonanztomographie
(fMRT) und diffusionsgewichteter Bildgebung (DTI), ob Unterschiede in der Struktur und
Funktion des Gehirns bei Kindern mit isolierter RS, verglichen zu Kindern mit LRS und
unbeeinträchtigten Kontrollen bestehen. Dabei fanden wir bei Kindern mit isolierter RS eine
stärkere rechtshemisphärische Aktivierung, wohingegen sich keine strukturellen Unterschiede
im Vergleich zur Kontrollgruppe nachweisen ließen. In unserer zweiten Studie untersuchten
wir die Auswirkungen eines morphembasierten Rechtschreibtrainings auf die Funktion
des Gehirns bei 20 deutschsprachigen, rechtschreibschwachen Kindern (unterteilt in eine
Trainings- und eine Wartegruppe) mittels wiederholter fMRT Messung. Kinder mit RS
wiesen vor dem Training (verglichen zu einer Kontrollgruppe) eine stärkere Aktivierung
frontal medial und in der rechten Hemisphäre, sowie eine geringere Aktivierung in links
okzipito-temporalen Regionen auf. Nach einem fünfwöchigen Training verbesserten sich die
Rechtschreibung und das Leseverständnis in der Trainingsgruppe, begleitet von einer
gesteigerten Aktivierung in links temporalen und (para)hippocampalen Regionen.
In unserer dritten Studie untersuchten wir die Auswirkungen eines Rechtschreibtrainings
auf die Struktur des Gehirnes bei 20 rechtschreibschwachen Kindern (unterteilt in eine
Trainings- und eine Wartegruppe) mittels wiederholter DTI Messung. Es zeigte sich, dass
eine Verbesserung der Rechtschreibleistung in der Trainingsgruppe mit einer verbesserten
Integrität der weißen Substanz in der rechten Hemisphäre einherging.
Die beiden Hauptergebnisse dieses Dissertationsprojektes können folgendermaßen
zusammengefasst werden:
Erstens weisen Kinder mit RS eine stärkere rechtshemisphärische Hirnaktivierung im
Vergleich zu nicht-beeinträchtigten Kontrollen auf. Zweitens scheint ein erfolgreiches
Training mit strukturellen und funktionellen Veränderungen des Gehirns einherzugehen.

INTRODUCTION
7
I INTRODUCTION
1. SPELLING AND READING IMPAIRMENT (SRI) Reading and spelling are essential skills in modern society. Reading is the most
important portal to knowledge acquisition in our information age (Gabrieli, 2009) and due to
increased use of communication technologies, such as the mobile phone (e.g. SMS) and
internet (e-mail, networks), spelling is essential for private and professional interaction.
Hence, impairments in these skills affect everyday life and are related to a greater risk of
school anxiety, unemployment and multiple emotional and behavioral difficulties (Ise &
Schulte-Körne, 2010). A severe and well-known form of spelling and reading difficulties is
dyslexia.
According to the ICD 10 definition (Dilling et al., 2005), dyslexia or spelling and
reading impairment (SRI; F 81.0) is diagnosed if reading and spelling skills are located two
standard deviations below the level that might be expected based on general intelligence, age
and education. However, it has to be noted that difficulties in spelling and reading occur in
various degrees of severity. Although cut points are placed to help define groups, they have
been criticized for being arbitrary and lack biological validity (Shaywitz et al., 2008).
Children scoring one standard deviation below the criteria may still require and profit from
intervention.
Spelling and reading impairments (SRI) have been described in every ethnic group,
language and geographic region (Shaywitz et al., 2008). Prevalence rates range from 3 – 15
%, depending on definition and stringency of criteria used (Eden & Zeffiro, 1998; Gabrieli,
2009; Habib, 2000; Shaywitz et al., 2003). SRI represents the most common learning
disability affecting over 80% of learning disabled children (Shaywitz et al., 2008).
Multiple problems are associated with SRI (Klicpera et al., 2007; Warnke et al.,
2004). An augmented rate of emotional problems, ranging from low self-esteem, loss of
motivation, depressive mood and anxiety to increased suicide rates, have been observed in
various studies (e.g. Maughan et al.; 2003; Riddick et al., 1999). Typical behavioral problems
associated with SRI are: attentional deficits, agitation, aggression and delinquent behaviour
(Arnold et al., 2005; Fluss et al., 2009; Morgan et al., 2008). Consequently, SRI seems to
correlate with a greater probability of school drop-out, lower educational achievement and
increased risk of unemployment (Daniel et al., 2006; Esser et al., 2002).
1.1. Etiology It is assumed that an interaction of multiple factors is responsible for precipitating
SRI, including biological (genetics), social and neurophysiological factors. Dyslexia, the most

INTRODUCTION
8
severe form of SRI, is considered as a neurodevelopmental disorder influenced by genetic
factors.
Family and twin studies show a moderate to high heritability of SRI (Schulte-Körne,
2001). Heritability estimates of 50-60% are reported for reading skills and 50-70% for
spelling skills (Schulte-Körne et al., 2006). Nine candidate risk genes, implicated in neural
migration and brain development, on chromosomes 1, 2, 6, 15, and 18, are associated with
SRI (e.g. Fisher & DeFries, 2002; Galaburda et al., 2006; Scerri & Schulte-Körne, 2010;
Shaywitz et al., 2008).
There are a number of social factors which have an impact on SRI including: low
socio-economic status, low maternal education, less reading outside school, number of
available books at home and number of siblings (Cunningham & Stanovich, 1998; Klicpera et
al., 2007; Morgan et al., 2008; Warnke et al., 2004). These factors influence family
interaction (vocabulary), as well as individual learning conditions and motivation.
“Neurophysiological” characteristics related to SRI are investigated by neuroimaging
methods. The most consistent finding of functional MRI studies is decreased brain activation
in parieto-temporal and occipito-temporal regions of the left hemisphere, along with increased
activation in frontal and right hemispheric language-related regions in individuals with SRI
(for an overview see Bartl-Pokorny et al., 2011; Shaywitz et al., 2006).
2. NEUROPHYSIOLOGY OF SRI Neuroimaging studies revealed differences in brain function and structure between
individuals with spelling and reading impairments compared to non-impaired controls.
In general, reduced gray matter volume in individuals with SRI has been found in the
temporal lobe bilaterally (occipito-temporal and parieto-temporal), in the bilateral inferior
frontal gyrus (IFG) and in the cerebellum bilaterally (Brambati et al., 2004; Brown et al.,
2001; Casanova et al., 2004; Eckert et al., 2003; Kronbichler et al., 2008; Silani et al., 2005;
Steinbrink et al., 2008; Vinckenbosch et al., 2005). Remarkably, gray matter alterations in
these regions are already observed in pre-reading children with a family-history of dyslexia
(Raschle et al., 2010), suggesting that these differences might be present at birth rather than
being experience-dependent.
Accordingly, DTI studies report decreased white matter integrity in individuals with
SRI in left occipito-temporal, parieto-temporal and left frontal white matter (cf. Beaulieu et
al., 2005; Carter et al., 2009; Deutsch et al., 2005; Klingberg et al., 2000; Niogi &
McCandliss, 2006; Rimrodt et al., 2010; Steinbrink et al., 2008).

INTRODUCTION
9
Heterogeneous patterns of brain activation differences in cortical and subcortical
regions between children and adults with SRI and non-impaired controls were found across
studies. Frequently, a decreased activation in two posterior left hemispheric regions (parieto-
temporal and occipito-temporal) along with increased activation in frontal and right
hemispheric language-related regions is thought to be related to SRI (Maisog et al., 2008;
Richlan et al., 2009). A more detailed description of fMRI findings will be presented in the
next section.
2.1. Functional MRI and SRI The majority of studies investigating “neurophysiological” correlates of SRI are
using functional MRI. Meanwhile, a multitude of fMRI studies applying different tasks (e.g.
orthographic decision, sentence comprehension, letter processing) in different samples (e.g.
children, adults in different languages) exist. However, findings with respect to patterns of
brain activation differences between individuals with SRI and non-impaired controls have
been heterogeneous so far. In many studies, decreased activation in two posterior left
hemispheric regions, the parieto-temporal and occipito-temporal region (word-form area), is
mentioned to be associated with SRI. The left parieto-temporal region is related to phoneme-
grapheme-conversion and the occipito-temporal (fusiform) region is critical for skilled, fluent
reading (see Figure 1; Richlan et al., 2009, 2011). Decreased activation in these areas may
represent a specific neurophysiological characteristic of dyslexia. Hoeft et al. (2007)
examined differences of brain activation patterns in a dyslexic group compared to an age-
matched control group, and a (younger) reading-matched control group. Relative to both
control groups, the dyslexic group exhibited decreased activation in left parietal and occipito-
temporal regions. Increased activation in frontal and right hemispheric language-related
regions in individuals with SRI has often been observed, and has been related to (inefficient)
compensatory mechanisms such as internal articulation (Maisog et al., 2008; Richlan et al.,
2009; Shaywitz et al., 2006).
To identify consistent neural activity across different functional neuroimaging
studies and to overcome the limited generalizability of single experiments, quantitative
coordinate-based meta-analyses such as activation likelihood estimation (ALE) are applied
(Eickhoff et al., 2009). Richlan et al. (2009) used activation likelihood estimation (ALE)
including 17 studies to identify the typical patterns of increased and decreased activation in
individuals with dyslexia. They found maxima of decreased activation primarily in the left
hemisphere (inferior parietal, superior temporal, middle and inferior temporal and fusiform
regions, inferior frontal gyrus). Furthermore, increased activation in the primary motor cortex
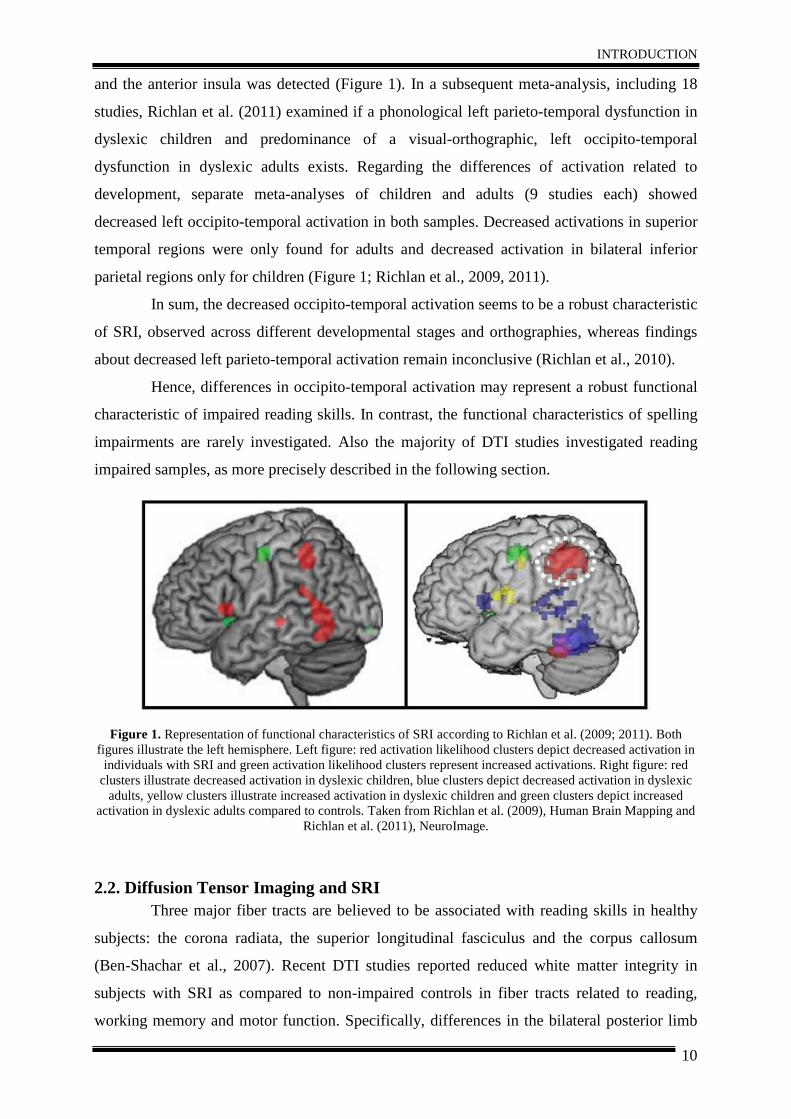
INTRODUCTION
10
and the anterior insula was detected (Figure 1). In a subsequent meta-analysis, including 18
studies, Richlan et al. (2011) examined if a phonological left parieto-temporal dysfunction in
dyslexic children and predominance of a visual-orthographic, left occipito-temporal
dysfunction in dyslexic adults exists. Regarding the differences of activation related to
development, separate meta-analyses of children and adults (9 studies each) showed
decreased left occipito-temporal activation in both samples. Decreased activations in superior
temporal regions were only found for adults and decreased activation in bilateral inferior
parietal regions only for children (Figure 1; Richlan et al., 2009, 2011).
In sum, the decreased occipito-temporal activation seems to be a robust characteristic
of SRI, observed across different developmental stages and orthographies, whereas findings
about decreased left parieto-temporal activation remain inconclusive (Richlan et al., 2010).
Hence, differences in occipito-temporal activation may represent a robust functional
characteristic of impaired reading skills. In contrast, the functional characteristics of spelling
impairments are rarely investigated. Also the majority of DTI studies investigated reading
impaired samples, as more precisely described in the following section.
Figure 1. Representation of functional characteristics of SRI according to Richlan et al. (2009; 2011). Both
figures illustrate the left hemisphere. Left figure: red activation likelihood clusters depict decreased activation in individuals with SRI and green activation likelihood clusters represent increased activations. Right figure: red
clusters illustrate decreased activation in dyslexic children, blue clusters depict decreased activation in dyslexic adults, yellow clusters illustrate increased activation in dyslexic children and green clusters depict increased
activation in dyslexic adults compared to controls. Taken from Richlan et al. (2009), Human Brain Mapping and Richlan et al. (2011), NeuroImage.
2.2. Diffusion Tensor Imaging and SRI Three major fiber tracts are believed to be associated with reading skills in healthy
subjects: the corona radiata, the superior longitudinal fasciculus and the corpus callosum
(Ben-Shachar et al., 2007). Recent DTI studies reported reduced white matter integrity in
subjects with SRI as compared to non-impaired controls in fiber tracts related to reading,
working memory and motor function. Specifically, differences in the bilateral posterior limb

INTRODUCTION
11
of the internal capsule (PLIC; e.g. Beaulieu et al., 2005; Klingberg et al., 2000), superior
corona radiata (SCR; e.g. Deutsch et al., 2005), the superior longitudinal fasciculus (SLF; e.g.
Carter et al. 2009; Hoeft et al., 2011; Rimrodt et al. 2010; Steinbrink et al. 2008), inferior
longitudinal fasciculus (ILF; e.g. Rollins et al., 2009; Steinbrink et al. 2008), corpus callosum
(CC; Ben-Shachar et al. 2007; Dougherty et al., 2007) and anterior corona radiata (ACR; e.g.
Beaulieu et al. 2005; Niogi & McCandliss, 2006) have been observed.
The most common imaging parameter used to assess white matter integrity in-vivo is
fractional anisotropy (FA). Furthermore, information about mean diffusivity (MD), axonal
and radial diffusion can be derived from DTI data, which allow a more precise interpretation
of differences in white matter integrity. In line with functional findings, several DTI studies
reported positive correlations between white matter integrity (mostly assessed by FA) and
reading ability in left parieto-temporal areas (Beaulieu et al. 2005; Deutsch et al. 2005;
Klingberg et al. 2000; Niogi & McCandliss 2006). Furthermore, a positive correlation
between white matter integrity of frontal pathways and working memory capacity were found
(Niogi & McCandliss, 2006). Steinbrink et al. (2008) observed decreased FA in bilateral
fronto-temporal and left temporo-parietal white matter regions (ILF and SLF) in German-
speaking dyslexics, suggesting a less efficient communication of regions associated with
reading and working memory.
Figure 2. Fiber tractography of the left superior corona radiate (SCR; left figure) and anterior corona radiata
(ACR). The white matter integrity of the fiber tracts of the left SCR (superior–inferior) positively correlated with reading performance. White matter integrity of the fiber tracts of the ACR (anterior–posterior) bilaterally positively correlated with working memory. Taken from Niogi & McCandliss (2006), Neuropsychologia.
As abovementioned, the majority of neuroimaging studies investigating SRI had a
strong focus on reading impairments, whereas neurophysiological correlates of spelling
ability and spelling impairment have been rarely investigated. Reading and spelling abilities
are significantly related to each other, but not completely identical regarding underlying
cognitive and brain mechanisms. Hence, the next section will focus on isolated spelling
impairment and neurophysiology of spelling ability.

INTRODUCTION
12
3. ISOLATED SPELLING IMPAIRMENT Currently, research focussing on (isolated) spelling impairment is rare, although
different diagnoses for children with difficulties in reading and spelling (F 81.0), as opposed
to children with isolated difficulties in spelling ability, exist in German-speaking countries.
According to the ICD 10 (WHO, 2003), the main feature of isolated spelling disorder (F
81.1.) is a specific and significant impairment in the development of spelling skills in the
absence of a history of specific reading disorder, which is not accounted for by age, visual
acuity problems or inadequate education. The ability to spell orally and to write down words
correctly is affected.
In German-speaking samples about 3-6% show isolated spelling difficulties (Moll &
Landerl, 2009; Wimmer & Mayringer, 2002). Due to the transparent orthography in German,
individuals often manage to read slowly but accurately in the course of their development,
whereas spelling mistakes rather persist into adulthood (Landerl & Klicpera, 2009). Spellings
are frequently phonologically adequate, but orthographically incorrect, possibly due to access
problems to the orthographic lexicon (Landerl & Wimmer, 2008; Wimmer & Schurz, 2010).
The course of an isolated spelling impairment is variable. Thirty-three percent of affected
children are able reach average spelling ability in the course of their development, in another
33% spelling impairments persist and in the remaining 33% additional reading difficulties
occur (Klicpera et al., 1993).
3.1. Neurophysiology of Spelling Impairment
So far only little is known about the neural substrates of spelling (Hillis et al., 2002)
and even less about the underlying brain mechanisms of spelling difficulties. For a long time,
the left angular gyrus and left supramarginal gyrus had been assumed to play a critical role in
spelling (Booth et al., 2002, 2004; Roeltgen, 1993; Rapcsak & Beeson, 2002). Other studies
rather suggested that the left inferior posterior temporal lobe (Beeson et al., 2003; Petrides et
al., 1995; Rapcsak & Beeson, 2004) or the left mid-fusiform region (Rapp & Lipka, 2011;
Tsapkini & Rapp, 2010) might be associated with spelling.
Further insights about brain regions associated with spelling impairment stem from
stroke studies. Lesions in the left posterior, inferior frontal and parietal cortex (Cloutman et
al., 2009) and left perisylvian regions (Henry et al., 2007; Hillis et al., 2002, 2004) have been
reported to be associated with spelling impairment. Lanzinger et al. (1999) observed that
lesions in the left medio-basal temporal lobe are involved in the emergence of spelling
impairment after stroke. Taken together, spelling ability in healthy subjects with unimpaired
development of spelling skills, is primarily related to left hemispheric brain activation.

INTRODUCTION
13
However, studies investigating the neural characteristics of developmental spelling
impairment are missing. As mentioned above, the term developmental spelling impairment
describes a specific and significant impairment in the development of spelling skills, which is
not accounted for by age, intelligence or inadequate education. Richards et al. (2009)
conducted a study with 11-year old poor spellers and found a stronger activation of the
precuneus, bilateral frontal regions, left angular gyrus and right temporal regions in poor
spellers, probably related to inefficient access to orthographic representations and increased
effort compared to good spellers. These findings provide first insights about
neurophysiological characteristics of spelling impairment. However, the study comprised only
seven poor spellers and did not examine any structural characteristics of spelling impairment.
The presented doctoral project thus aimed to investigate neurophysiological
characteristics specifically involved in spelling impairment and changes in brain structure and
function related to a spelling intervention.
4. INTERVENTION The development and provision of intervention programs for individuals with SRI is
a crucial objective, requiring fundamental knowledge of the involved cognitive and cortical
mechanisms in order to best tailor specific interventions according to individual impairments.
Evidence-based intervention could help to avoid that SRI and associated emotional and
behavioral difficulties persist over time or at least to alleviate them. The sooner SRI is
diagnosed and prevented, the better the outcome. Intervention programs for children beyond
second grade are effective but challenging (Shaywitz et al., 2008).
Basically, two kinds of intervention can be distinguished. The first kind focuses on
sensory deficits that are supposed to underlie SRI. The problem with this approach is that it
often ignores the complexity of SRI, as only about 30 to 40 % of individuals with SRI show
sensory deficits. In addition, improvement of sensory processing does not necessarily have a
positive impact on reading and/or spelling skills.
The second kind of intervention focuses on the symptom level of SRI. Recent
behavioral studies showed that interventions based on the symptom level (focusing on reading
and spelling skills) more efficiently improve literacy skills than interventions based on
sensory skills, in both English- (Alexander & Slinger-Constant, 2004) and German-speaking
samples (Suchodoletz, 2010).

INTRODUCTION
14
4.1. Spelling Intervention Compared to reading relatively few studies dealt with spelling intervention so far.
This is a severe limitation in German-speaking samples, as the impairments in spelling skills
are often more prominent in and relevant for children’s everyday lives and persist longer.
Furthermore, a considerable number of children experience serious deficits in spelling in spite
of intact reading skills (Moll & Landerl, 2009).
Most of the available interventions are based on phonological skills, as strong
evidence for the efficacy of such programs in English-speaking samples has been found (Eden
et al., 2004). However, in more transparent orthographies (e.g. German), phonological errors
are rather exceptional (Bergmann & Wimmer, 2008; Landerl & Wimmer, 2008) and provision
of morpheme-based and orthographic interventions for children in higher grades appears to be
particularly important.
Hence, in addition to phonologically-based interventions, methods targeting at the
morphematic and orthographic structure of words are essential to avoid incorrect spellings.
The evaluations of the effectiveness of orthographic spelling intervention revealed that
learning of explicit orthographic rules improve spelling ability as well as orthographic
knowledge (Ise & Schulte-Körne, 2010; Faber, 2010). Furthermore, several studies found that
morpheme-based interventions were able to significantly increase spelling skills (Kargl et al.,
2008, 2011; Schneeberger et al., 2011; Walter et al. 2001).
4.1.1. Morpheme-based intervention A morpheme is defined as the “smallest meaningful unit of language” (Bhatt, 1991).
Every word is built by different parts, which follow particular spellings (e.g. unforgetful =
prefix [un], suffix [ful], root [forget]). Therefore, the spelling of the German verb “verfahren”
can be derived by two rules: the prefix [ver] is always written with [v], the root [fahr] always
with an “h”. Children do not need to remember the spelling of every single word, but only to
memorize the spelling of their component parts. Furthermore, morphosemantic information
can support the development of a meaning-oriented decoding strategy, e.g. the correct
spelling of the noun “Motor-rad” (motor-bike) can be derived by the meaning (May et al.,
2000). In addition, this strategy seems to be easy to apply as only “100 of the most frequent
morphemes cover 70% of all written material” (Scheerer-Neumann, 1979).
Indeed, behavioral studies showed that morpheme-based interventions significantly
enhanced reading and/or spelling ability (Arnbak & Elbro, 2000; Lyster, 2002; Nunes et al.,
2003). Specifically, Nunes et al. (2003) found reading and the use of morphological rules in
spelling to be improved after morpheme-based intervention (12 weekly sessions for 30

INTRODUCTION
15
minutes each). Arnbak and Elbro (2000) showed that a morphological awareness training (15
minutes, three times a week, for 12 weeks) enhanced reading comprehension and spelling of
morphologically complex words also in dyslexic children.
4.1.2. Computer-based intervention The number of computer-based or computer-assisted interventions is constantly
rising and offers multiple advantages compared to conventional spelling interventions
(Zimdars & Zink, 2006). Application of computer-based interventions allows automatically
assessing training progress and individual adaption of different levels of difficulty.
Furthermore, a multitude of exercises e.g. grapheme-phoneme allocation through integrated
speech programs or writing exercises supported by a keyboard can be provided. The use of a
keyboard ensures legible writing and spelling mistakes can be easily corrected on screen.
Interventions supported by computers are also related to enhanced motivation (by constant
and immediate feedback) and concentration, which may positively affect training success.
Nevertheless, technical support cannot replace personal interaction with qualified
teachers and instructors, but rather represent helpful support and augmentation of
conventional programs (Suchodoletz, 2010).
4.1.3. Morpheus-intervention The Morpheus-intervention (Kargl & Purgstaller, 2010), which we applied in this
doctoral project, is a morpheme-based, computer-assisted spelling intervention, specifically
developed for children of the 4th to the 8th grade. The intervention consists of the most
frequent morphemes of the German language and is based on the empirical-based basic
vocabulary of fourth graders (Augst, 1989). As mentioned before, morpheme-based
interventions are easy to apply and efficient. The correct spelling of words can be derived by
memorizing the rules for particular component parts of a word.
The Morpheus-intervention consists of computerized tasks, a book of exercises and
morpheme-based games to facilitate the consolidation of the strategy. The intervention
includes daily handwritten and computer homework along with instructor-guided courses (e.g.
once a week, lasting approximately two hours). The consolidation of the morpheme-based
spelling strategy occurs by different exersises dealing with morphemes (such as detecting pre-
and suffixes of a word, arranging component parts of a word, detecting the root of a word,
identifying and arranging word families (e.g. all word including the same root) and word
classes (noun, verb, adjective), identifying and counting component parts of a word, writing

INTRODUCTION
16
down component parts of a word and completing cloze tests; see Figure 3). The Morpheus-
intervention has been constructed on the basis of the following principles: simplicity, relief
due to morpheme segmentation, rule-governed repetition, avoidance of mistakes,
productivity, and practicing handwriting.
The exercises are presented at different levels of difficulty. During the computer
tasks the achieved scores are displayed on the computer screen. Participants can only reach
the next difficulty level of the same exercise when they solve at least 75% of the given
problems correctly. Furthermore, the scores allow assessing the progress of the training.
The Morpheus-intervention has been approved as an evidence-based intervention for
individuals with reading and spelling deficits by the federal ministry of Austria and has shown
to significantly improve spelling ability in children in a series of behavioral studies in our
study group (Kargl et al., 2008, 2011; Weiss et al., 2010).
A recent EEG study provided evidence for a neurophysiological effect of the
Morpheus-intervention (lasting three-weeks). Increased EEG activation at centroparietal sites
which are thought to be involved in the neural network subserving reading and spelling, along
with performance gains in children with SRI were found (Kozel et al., submitted).

INTRODUCTION
17
Figure 3. Illustrating six different tasks of the computer-based Morpheus-intervention. Examples for the 1st level of difficulty: (1) identifying the correct suffix of a word, (2) arranging component parts of a word, (3, 4)
identifying and arranging word families and classes. Examples for the 3rd level of difficulty: (5, 6) identifying and counting component parts of a word. For each task the number of trials is illustrated at the left upper corner (e.g. word 1 of 18) and the number of correctly solved trials is depicted on the right upper corner (e.g. 74 of 74 points). The instruction is presented at the bottom of the screen (e.g. Which suffix is matching the wort root?).
Screenshots taken from the computer tutorial of the Morpheus-intervention.
4.2. Neurophysiologic changes related to intervention Intervention studies using functional MRI revealed changes in brain activation
patterns along with successful intervention (Aylward et al., 2003; Eden et al., 2004; Gaab et
al., 2007; Meyler et al., 2008; Richards et al., 2006; Shaywitz et al., 2004; Simos et al., 2002;
2006; Temple et al., 2003). However, the majority of these findings stems from reading
impaired, English-speaking samples and existing studies typically focus on phonology-based
intervention programs (e.g. Eden et al., 2004; Shaywitz et al., 2004), while only few studies
were performed involving subjects with spelling impairment (Richards et al., 2009).
In the following, an overview of current findings about neurophysiological changes
related to successful intervention in subjects with SRI will be presented. In the first
paragraphs, the neurophysiological effects of phonologically-based intervention will be

INTRODUCTION
18
discussed. In the next paragraph we will focus on changes in functional patterns of brain
activation related to reading interventions and the last paragraph will outline functional
changes associated with morpheme-based and orthographic intervention.
Several studies showed that phonologically targeted interventions result in
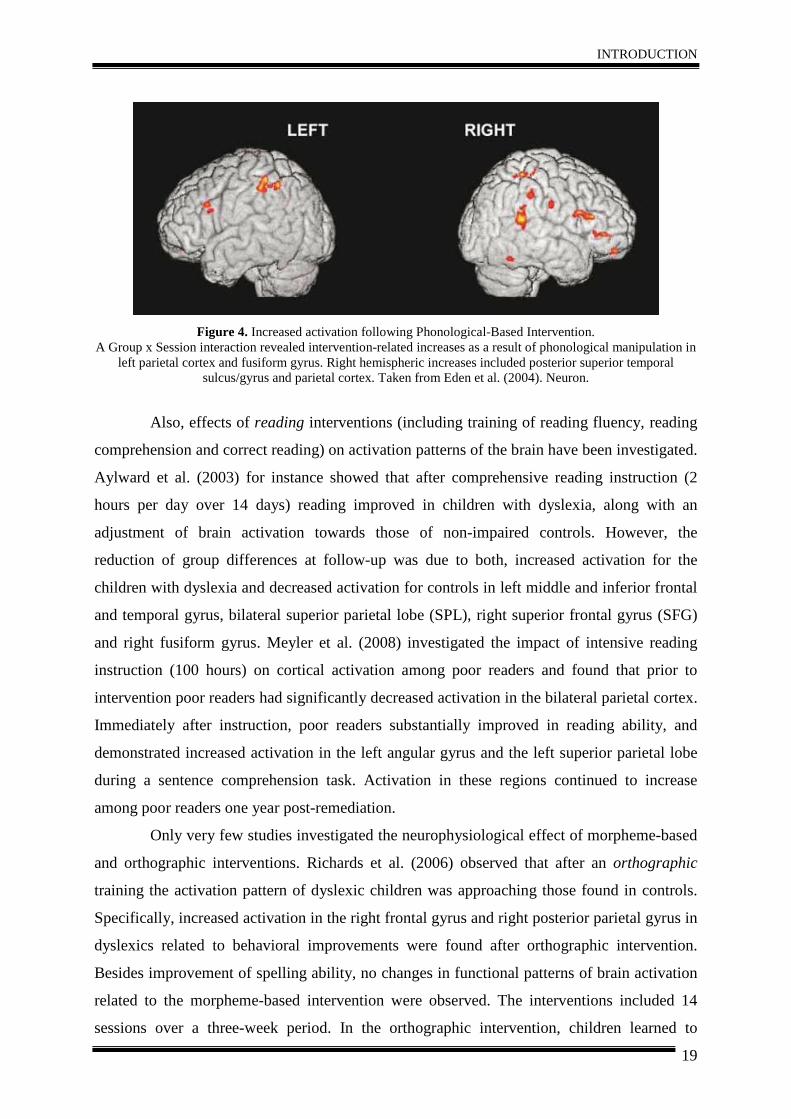
improvements in reading ability associated with neurophysiological changes. Eden et al.
(2004) reported performance improvements in adults with developmental dyslexia after eight
weeks of intervention, associated with signal increases in left parietal cortex and right
hemispheric regions (inferior frontal, parietal and parieto-temporal; Figure 4). It was
concluded that a combination of right hemispheric compensatory activation and increased
activation in left parietal regions, which are typically involved in phonological processing, are
associated with effective intervention. Similarly, Temple et al. (2003) observed behavioral
improvements in children with dyslexia, along with increased activation in the left parieto-
temporal cortex and in a right hemispheric compensatory network (frontal and temporal
regions) after intervention. In line with these findings, a year-long phonological intervention
was associated with significant performance gains in reading fluency, along with increased
activity in left hemispheric reading network (e.g. inferior frontal gyrus, superior and middle
temporal gyrus, occipitotemporal gyrus) in children with reading disability (Shaywitz et al.,
2004).
Evolvement of deviant into similar activation patterns in children with reading
impairment compared to non-impaired controls due to phonologically based intervention was
also supported by magnetic source imaging studies (magnetoencephalography; Simos et al.,
2002, 2006). Before intervention, children with reading impairment showed decreased
activation in the posterior superior temporal gyrus (STG) and increased activation in the
corresponding right hemispheric region. Activation in the left STG increased, along with
improvements of reading skills (Simos et al., 2002). Furthermore, Gaab et al. (2007) found
that children with dyslexia developed similar activation patterns compared to typical-reading
children in the left precentral gyrus after eight weeks of remediation. The intervention
included phoneme discrimination and sentence comprehension tasks. It has to be noted,
however, that the mentioned studies included phonological processing tasks (Eden et al.,
2004; Shaywitz et al., 2004; Simos et al., 2002; 2006; Temple et al., 2003) during the
functional MRI assessment, whereas Gaab et al. (2007) investigated brain activation during
rapid auditory processing, examining differences in activation during slow and fast transitions
of acoustic stimuli. In sum, it seems that phonological-based intervention results in activation
increases of the left parieto-temporal region and a right hemispheric compensatory reading
network.
INTRODUCTION
19
Figure 4. Increased activation following Phonological-Based Intervention.
A Group x Session interaction revealed intervention-related increases as a result of phonological manipulation in left parietal cortex and fusiform gyrus. Right hemispheric increases included posterior superior temporal
sulcus/gyrus and parietal cortex. Taken from Eden et al. (2004). Neuron.
Also, effects of reading interventions (including training of reading fluency, reading
comprehension and correct reading) on activation patterns of the brain have been investigated.
Aylward et al. (2003) for instance showed that after comprehensive reading instruction (2
hours per day over 14 days) reading improved in children with dyslexia, along with an
adjustment of brain activation towards those of non-impaired controls. However, the
reduction of group differences at follow-up was due to both, increased activation for the
children with dyslexia and decreased activation for controls in left middle and inferior frontal
and temporal gyrus, bilateral superior parietal lobe (SPL), right superior frontal gyrus (SFG)
and right fusiform gyrus. Meyler et al. (2008) investigated the impact of intensive reading
instruction (100 hours) on cortical activation among poor readers and found that prior to
intervention poor readers had significantly decreased activation in the bilateral parietal cortex.
Immediately after instruction, poor readers substantially improved in reading ability, and
demonstrated increased activation in the left angular gyrus and the left superior parietal lobe
during a sentence comprehension task. Activation in these regions continued to increase
among poor readers one year post-remediation.
Only very few studies investigated the neurophysiological effect of morpheme-based
and orthographic interventions. Richards et al. (2006) observed that after an orthographic
training the activation pattern of dyslexic children was approaching those found in controls.
Specifically, increased activation in the right frontal gyrus and right posterior parietal gyrus in
dyslexics related to behavioral improvements were found after orthographic intervention.
Besides improvement of spelling ability, no changes in functional patterns of brain activation
related to the morpheme-based intervention were observed. The interventions included 14
sessions over a three-week period. In the orthographic intervention, children learned to

INTRODUCTION
20
strengthen the precise representation of a written word in the working memory. In the
morpheme-based intervention, children learned to divide morphologically complex words into
their meaning parts. Unlike this, behavioral improvements and changes in brain activation
patterns associated with a three weeks morpheme-based intervention were reported in a recent
study using EEG (Kozel et al., submitted; see also Weiss et al., 2010).
In this doctoral project we aimed to assess the effects of an intensive morpheme-
based intervention (i.e. Morpheus-intervention, which lasts approximately five weeks) on
brain structure and function.
To the best of our knowledge, only two studies investigated structural changes
related to intervention in individuals with SRI so far. Increases in gray matter volume in the
left anterior fusiform/hippocampus, left precuneus, right hippocampus and right anterior
cerebellum have been associated with gains in reading skills following eight weeks of
intervention (Krafnick et al., 2011). Using DTI among poor readers, changes in white matter
integrity (increase of FA and decrease of radial diffusivity) were observed after 100 hours of
reading instruction in the left anterior centrum semiovale (Keller & Just, 2009).
In conclusion, evidence supporting the malleability of neural systems in individuals
with SRI has been found. On the functional level, neurophysiologic changes in terms of
increased activation of the “common” reading networks or right hemispheric “compensatory”
networks were observed. Also, first indications of structural plasticity related to successful
intervention have been reported. These promising findings provide some hope that targeted
intervention is associated not only with improvements in spelling and reading ability, but also
with persisting neurophysiological changes.

OBJECTIVES AND METHODS
21
II OBJECTIVES AND METHODS Previous studies indicated that characteristic differences of brain structure and
function in children with SRI compared to non-impaired controls exist. Based on existing
evidence in this field, we aimed to address two central research questions: 1) We investigated
whether differences in brain structure and function of children with spelling impairment
compared to non-impaired controls exist, using multimodal imaging. 2) This doctoral project
should also deal with the question how an intensive morpheme-based intervention would
result in improvement in spelling skills and associated changes in brain structure and function.
For this reason we apply the Morpheus-intervention (Kargl & Purgstaller, 2010), which has
repeatedly been observed to be associated with reliable increases in spelling performance
(Kargl et al., 2008, 2011; Kozel et al., submitted; Schneeberger et al., 2011; Weiss et al.,
2010).
The studies presented in this thesis were based on a large pre-experimental screening
from which participants for the subsequent functional MRI measurement were drawn. In this
screening, we assessed reading and spelling abilities and socio-demographic data in a sample
of 107 subjects. Standardized tests for the assessment of reading and spelling skills, non-
verbal intelligence and personality were administered. In addition, relevant socio-
demographic data such as age, sex, handedness, suitability of MRI assessment and native
language were obtained. The duration of the screening was about one hour and thirty minutes.
Non-verbal intelligence was measured by the Standard Progressive Matrices (SPM) by Raven
(1960) and personality was assessed by the Five-Factor-Questionnaire for children by
Asendorpf (1998) to control for potential influences of non-verbal intelligence and personality
traits on task performance. To assess spelling skills, we used a standardized spelling test
(Hamburger-Schreibprobe, HSP) by May et al. (2000). Additionally, we administered the
“Salzburger-Lese-Sreening” (SLS; Mayringer & Wimmer, 2003, 2005) which measures
reading speed and basic reading ability (automaticity, accuracy). Furthermore, we also
assessed reading comprehension (i.e. comprehension of words, sentences and text) by means
of a standardized German-speaking test (ELFE 1-6; Lenhard & Schneider, 2006).
According to the demographic data (e.g. handedness, suitability of MRI assessment)
and reading and spelling ability, subjects were chosen to participate in one out of three
different research projects: In the studies of this doctoral project we included only right-
handed, spelling impaired subjects with mild reading impairment who were suitable for MRI

OBJECTIVES AND METHODS
22
assessment (e.g. no braces, no claustrophobia). In another doctoral project (Kozel et al.,
submitted) children with poor to average reading and/or spelling abilities were included. In
this project, the behavioral and neurophysiological effects of reading and spelling
interventions (lasting five weeks each) were assessed by means of EEG. The remaining
subjects participated in a longitudinal behavioral study, investigating more long lasting effects
of a reading intervention and a morpheme-based spelling intervention (Schneeberger et al.,
2011). In the Schneeberger et al. (2011) study, the behavioral effects of the interventions were
assessed directly after the training (which took about five weeks to complete) and one month
after the intervention.
For our project we investigated a subgroup of 45 subjects behaviorally and by
repeated structural and functional neuroimaging, before and after the intervention.
Imaging was performed on a 3.0 Tesla Trio Tim scanner (Siemens Medical Systems,
Erlangen, Germany) using a 12-channel head coil. A high-resolution isotropic (1x1x1 mm)
structural scan (TR = 1900 ms, TE = 2.2 ms) was acquired to allow precise registration of
functional data to individual anatomy. Furthermore, a single shot EPI DTI data including four
averages (TR = 6700 ms, TE = 95 ms, 12 directions) was obtained. Functional images were
acquired with a single-shot gradient echo EPI sequence (TR = 2190 ms, TE = 30 ms).
As described in more detail in the studies reported below, three different
orthographic decision conditions were presented during event-related fMRI (1: correctly
spelled words, 2: misspelled words, 3: pseudowords). Similarly to the spelling judgment task
of Richards et al. (2009), children had to decide whether a presented word was spelled
correctly (e.g. Bäume; trees), incorrectly (e.g. fergesslich instead of vergesslich; forgetfull
instead of furgetful), or if it is a pseudoword (e.g. Ostablast). The correct decision for
misspelled versus correctly spelled words requires orthographic processing, as the misspelled
words are phonologically correct, resembling the pseudohomophones (see Kronbichler et al.,
2007; van der Mark et al., 2009). Visual stimuli were synchronized with the MR-scanner
using the software “Presentation” (Neurobehavioral Systems, Albany, CA). Answers were
given via a button response box as described above.
All subjects were right-handed and suitable for MRI assessment. Due to the
functional MRI paradigm and the applied Morpheus-intervention, we included children with
spelling impairment and mild reading impairment. Based on methodological and ethical
reasons, children with severe reading impairment were allocated to projects including reading
intervention. Furthermore, the applied functional MRI paradigm requested the correct

OBJECTIVES AND METHODS
23
perception of a word (or pseudoword) within three seconds. Spelling skills ranged from one to
two standard deviations below average (HSP percentile ranking score one to 39).
The applied intervention is a computer-aided morpheme-based spelling training
(Morpheus; Kargl & Purgstaller, 2010) which was realized over a time period of five weeks.
The training material of Morpheus consists of the most frequent morphemes (component parts
of a word e.g. prefix, root, suffix) of the German language and contains different levels of
difficulty. The intervention comprised a weekly instructor-guided course (lasting
approximately two hours) along with daily handwritten and computer homework. Children do
not need to remember the spelling of every single word, but only to memorize the spelling of
their component parts (e.g. unfriendly = prefix [un], suffix [ly], root [friend]). This strategy
seems to be easy to apply as only “100 of the most frequent morphemes cover 70% of all
written material” (Scheerer-Neumann, 1979, p. 125). The effectiveness of the Morpheus-
intervention has already been proven in behavioural studies (e.g. Kargl et al., 2008, 2011;
Weiss et al., 2010). Furthermore, first indications of neurophysiological changes (as measured
by EEG) due to the Morpheus-intervention have been reported (Kozel et al., submitted).
Regarding more long lasting behavioral effects of the Morpheus-intervention, improvements
of spelling ability are less prominent one month after the intervention, but still significantly
above the baseline value (Schneeberger et al., 2011).
In the studies of this doctoral project, each participant was tested twice, at baseline
and after five weeks of intervention (or, after a five week waiting period). This doctoral
project addresses two central research questions, 1) the investigation of isolated spelling
impairment and 2) the investigation of intervention-related changes of functional and
structural brain characteristics. These two research questions were investigated in three
different studies:
To our very best knowledge, no study investigating the neural characteristics of
isolated spelling impairment appears to exist so far. Therefore, in our first presented study,
we investigated structural and functional brain characteristics in children with isolated
spelling impairment. More specifically, we compared a group of children with isolated
spelling impairment, a group of children with SRI and a group of non-impaired controls.
Moreover, studies investigating the neurophysiological effects of a morpheme-based
spelling intervention are rare. Hence, in our second study, we examined the effects of the
Morpheus-intervention on functional patterns of brain activity. For this reason we divided the
spelling impaired sample into a training group (receiving the intervention between the pre-

OBJECTIVES AND METHODS
24
and the post-test) and into a waiting group (which completed the intervention after the post-
test).
In addition, studies examining structural changes related to spelling intervention are
completely missing. Therefore, in our third study, we assessed the potential effects of the
applied Morpheus-intervention on brain structure by specifically investigating changes in
white matter integrity in the training- and the waiting group, using repeated DTI.
In the following sections the three studies are described in more detail.

STUDY I
25
III STUDIES
STUDY I – Isolated spelling impairment
Distinct patterns of brain function in children wit h
Isolated Spelling Impairment: New Insights.
D. Gebauer 1,2, C. Enzinger 1,4, M. Kronbichler 3, M. Schurz 3, G. Reishofer 4 , K. Koschutnig 2,4, R. Kargl 5, C. Purgstaller 5, F. Fazekas 1, A. Fink 2
1 Department of Neurology, Medical University of Graz, Austria 2 Department of Psychology, Karl-Franzens-University Graz
3 Department of Psychology & Center for Neurocognitive Research, University of Salzburg 4 Section of Neuroradiology, Department of Radiology, Medical University of Graz
5 Institute for Reading and Spelling in Graz, Austria (Submitted to Neuropsychologia on 28 June 2011; submitted in revised form on 14 November 2011)

STUDY I
26
Abstract Studies investigating reading and spelling difficulties heavily focused on the neural correlates
of reading impairments, whereas spelling impairments have been largely neglected so far.
Hence, the aim of the present study was to investigate brain structure and function of children
with isolated spelling difficulties. Therefore, 31 children, aged ten to 15 years, were
investigated by means of functional MRI and DTI. This study revealed that children with
isolated spelling impairment exhibit a stronger right hemispheric activation compared to
children with reading and spelling difficulties and controls, when engaged in an orthographic
decision task, presumably reflecting a highly efficient serial grapheme-phoneme decoding
compensation strategy. In addition, children with spelling impairment activated bilateral
inferior and middle frontal gyri during processing correctly spelled words and misspelled
words, whereas the other two groups showed bilateral activation only in the misspelled
condition, also suggesting that additional right frontal engagement could be related to
generally higher task demand and effort. DTI analyses revealed stronger frontal white matter
integrity (fractional anisotropy) in controls (compared to spelling and reading impaired
children), whereas no structural differences between controls and spelling impaired children
were observed.
KEYWORDS: isolated spelling impairment, fMRI, right hemisphere, dyslexia, DTI

STUDY I
27
1. Introduction
Depending on definition and stringency of criteria used, approximately 3-15% of children
show difficulties in reading and spelling (Eden & Zeffiro, 1998; Gabrieli, 2009; Habib, 2000;
Shaywitz et al., 2003) that may be associated with a greater risk of school anxiety,
unemployment and multiple emotional and behavioural difficulties (Arnold et al., 2005;
Daniel et al., 2006; Fluss et al., 2009; Ise & Schulte-Körne, 2010; Klicpera et al., 2007;
Maughan et al., 2003; Morgan et al., 2008). To date, numerous studies investigated
behavioural and brain processes involved in reading impaired individuals (e.g. Eden et al.,
2000; Meyler et al., 2007; Richlan et al., 2010; Shaywitz et al., 2007) and frequently imply
that spelling impairment is only a secondary phenomenon. Only few studies focused on
spelling impairment itself (e.g. Richards et al., 2009). Nevertheless, deficits frequently go
beyond reading and it is important to achieve a better understanding of common
neuropsychological deficits besides reading ability. Although spelling difficulties are
associated with dyslexia they have been largely neglected by the majority of studies in this
field (Angelelli et al., 2010).
Especially the English-speaking community focuses strongly on reading impairment. For
instance, the DSM IV (APA, 1994) contains a diagnosis of isolated “reading disorder”
(315.00), but no diagnosis for spelling impairment, instead containing the “disorder of written
expression” (315.2). In more transparent orthographies such as Spanish, Finnish, Italian,
Greek, reading accuracy is hardly affected, whereas impairments of reading speed and severe
and persistent impairments in spelling are observed (Wimmer et al., 1998). Accordingly, in
the ICD 10 (Dilling et al., 2005), which is the major diagnostic manual used in German
speaking countries, an isolated impairment is only assumed for spelling (F 81.1), whereas for
reading disorder (F 81.0) associated spelling problems are supposed to be frequent and mainly
secondary to reading difficulties. It was also found that children with isolated spelling
difficulties show different patterns of cognitive deficits compared to children with both
reading and spelling difficulties (Moll & Landerl, 2009, Wimmer & Schurz, 2010). Isolated
spelling difficulties in German-speaking samples were found in about 3-6% (Moll & Landerl,
2009; Wimmer & Mayringer, 2002). Due to the transparent orthography in German, dyslexics
manage to read slow but accurate in the course of their development, whereas spelling
mistakes rather persist into adulthood (Landerl & Klicpera, 2009). Interestingly, Moll and
Landerl (2009) found that children with isolated spelling deficits named pseudohomophones
as quickly as their corresponding words, and that their phonological awareness skills were
adequate. The authors suggested that the reading in children with isolated spelling deficits

STUDY I
28
may be based on highly efficient grapheme-phoneme decoding procedures. Due to the
asymmetry in German language (grapheme-phoneme correspondence is high, but phoneme-
grapheme correspondence is low) this strategy is not helpful for spelling difficulties, as
different spellings for words with the same pronunciation (e.g. Wal / whale – Wahl / election)
exist.
Another explanation could be that deficits in vowel length perception might be responsible for
spelling disorders. In German orthography vowel length is not marked by the vowel letter
itself, but by the letters following the vowel. Short vowels are, often marked by two following
consonants (e.g., Stall, /$tal/, [barn]), whereas for a long vowel frequently a ‘‘silent h’’ (e.g.,
Stahl, /$ta:l/, [steel]) is added (Groth et al. 2011). Difficulties in perceiving these differences
in vowel length might impair spelling ability.
Based on these considerations we speculated that the functional patterns of brain activity
should dissociate between children with isolated spelling impairment and children with
deficits in reading and spelling. Several theories about deficits underlying spelling and
reading impairment emerged from behavioural and neuroimaging studies (e.g. phonological
deficit theory, see further Ramus et al., 2003; magnocellular deficit theory, Stein, 2001 ;
double-deficit theory, Wolf & Bowers, 1999; cerebellar deficit theory, Nicolson et al., 1999;
temporal processing theory, Steinbrink et al., 2009). Heterogeneous patterns of brain
activation differences in cortical and subcortical regions between children and adults with
difficulties in reading and spelling and non-impaired controls were found across studies.
Frequently, a lower activation in parieto-temporal and occipito-temporal brain regions of the
left hemisphere, associated with deficits in reading-related skills, such as grapheme-phoneme-
conversion and automatic and fluent reading (Kronbichler et al., 2008; Shaywitz et al., 2004)
is mentioned to be related with dyslexia. In addition, dyslexics often show increased frontal
and right hemispheric activation, which is thought to be related to compensatory activity, e.g.
internal articulation (Maisog et al., 2008; Richlan et al., 2009; Shaywitz et al., 2006). Recent
research on reading difficulties found substantial support that left occipito-temporal reading
circuits (comprising the visual word form area; see further Dehaene & Cohen, 2011) are the
origin of persistent impairments of fast fluent reading (e.g. Cohen et al., 2000; Kronbichler et
al., 2006), but so far only little is known about the neural substrate of spelling (Hillis et al.,
2002) and even less about underlying brain mechanisms of spelling difficulties.
For a long time, the left angular gyrus (and left supramarginal gyrus) was assumed to play a
critical role in spelling (Booth et al., 2002, 2004; Roeltgen, 1993; Rapcsak & Beeson, 2002).
Other studies rather suggest that the left posterior temporal lobe (BA 37; Petrides et al., 1995),

STUDY I
29
especially the inferior region (Beeson et al., 2003; Rapcsak & Beeson, 2004) is associated
with spelling. Rapp and colleagues (Rapp & Lipka, 2011; Tsapkini & Rapp, 2010) propose
the left mid-fusiform region to be related to spelling. Purcell et al. (2011) state that a left
hemispheric region just lateral and superior to the VWFA plays a significant role in typed
spelling (see also Katanoda et al., 2001; Sugihara et al., 2006). Richards et al. (2009)
conducted a study with poor spellers and found a stronger activation of the bilateral
precuneus, posterior cingulum and frontal regions, probably reflecting inefficient neural
mechanisms and increased effort compared to controls.
Several studies using diffusion tensor imaging (DTI) found a correlation between white
matter integrity (mostly measured by fractional anisotropy, FA) and reading ability in the left
parieto-temporal area (e.g. Beaulieu et al., 2005; Deutsch et al., 2005; Klingberg et al., 2000;
Niogi & McCandliss, 2006). To our knowledge only two DTI studies included behavioural
measures of spelling. Deutsch et al. (2005) found positive correlations between measurements
of reading (r = .62) and spelling (r = .66) with left parieto-temporal connectivity in children
with reading and spelling impairment. Suggesting that better white matter integrity in this
region is related to a general improved processing efficiency. Steinbrink et al. (2008)
investigated German dyslexic adults and found a decreased FA in bilateral fronto-temporal
and left temporo-parietal white matter regions, probably indicating less efficient
communication in dyslexics. No significant correlations with spelling were reported, but a
significant correlation between FA and reading speed.
In a previous study (Gebauer et al., submitted) we examined the effects of spelling
intervention on children with spelling and reading impairment, investigating a training group
and a waiting group (each group included children with isolated spelling impairment and
reading and spelling impairment) by means of repeated behavioral assessment, functional
MRI and DTI. We here addressed the central research question whether functional and
structural patterns of brain activity in children with isolated spelling impairments differ from
those observed in children with impairments in reading and spelling and in healthy controls.
We hypothesized that (a) children with poor spelling and reading abilities might show a
reduced activation of the left occipito-temporal region compared to controls. We further
presumed that (b) children with isolated spelling difficulties might possibly show reduced
brain activation in the left angular gyrus or posterior inferior temporal cortex, accompanied
with an increased activation in the homologue right hemispheric areas. We further expected

STUDY I
30
(c) decreased white matter integrity (FA) in children with spelling and reading impairment in
the left parieto-temporal cortex.
2. Method 2.1. Participants
Out of 107 subjects, 42 German-speaking children aged between nine and 15 years were
recruited based on a behavioural pre-screening. Standardized tests for the assessment of
reading and spelling abilities were administered, and information on relevant socio-
demographic data such as age, sex, native language and school year was recorded.
To assess spelling skills, we used the Hamburger-Schreibprobe (HSP; May et al., 2000), a
standardized spelling test. In the HSP, words and sentences are dictated by the experimenter
and have to be written next to the corresponding pictures that illustrate the respective words
or sentences. This test takes about 15 minutes and within this study the version for 4th/5th
graders and 5th to 9th graders were applied. The HSP provides measures for the number of
correctly spelled words and the number of grapheme-related mistakes. The latter was used in
this study as it provides a more precise measure of spelling ability.
Additionally, we measured reading speed and basic reading ability (automaticity, accuracy),
by means of the Salzburger-Lese-Sreening (SLS; Mayringer & Wimmer, 2005). The SLS 1-4
was used for children up to the 4th grade, and the SLS-5-8 was applied for older children,
parallel versions exist for both. In the SLS, children have to decide whether the content of a
presented sentence is correct or not. Testing time is limited to three minutes.
In addition we also measured reading comprehension (i.e. comprehension of words, sentences
and text) by means of a standardized “German-speaking test” (ELFE 1-6; Lenhard &
Schneider, 2006). Furthermore, non-verbal intelligence was measured by means of the
Standard Progressive Matrices (SPM) by Raven (1960). We also assessed personality by
means of the Five-Factor-Questionnaire for children by Asendorpf (1998) to control for
potential influences of specific traits on performance. However, no differences between the
groups were observed. Hence, we did not further include personality factors.
Only participants with less than 3 mm motion and less than 1 mm motion between sequential
functional volumes were included in the analysis. Eleven children had to be excluded due to
movement artefacts (n=7), poor behavioural performance inside the scanner (n=2, Mean
Accuracy < 50%) or because they decided to terminate the fMRI session (n=2), rendering a

STUDY I
31
final sample of 31 children1 (15 males) in the age between ten and 15 years (M = 11.81; SD =
1.56). All participants were healthy, right-handed and had normal or corrected-to normal
vision. Structural brain scans were reported as normal in all children. The study was approved
by the ethics committee of the Medical University of Graz, Austria. All participants and their
parents gave written informed consent.
In a previous study (Gebauer et al., submitted) we investigated the effects of spelling
intervention on children with spelling and reading impairment. We here examined whether
functional and structural patterns of brain activity in children with isolated spelling
impairments differ from those observed in children with impairments in reading and spelling
and in healthy controls. Therefore, we divided the children into three groups: (1) Eleven
children with isolated spelling impairment (SI) and (2) nine spelling and reading impaired
children (SRI), as it was determined by means of standardized psychometric tests for the
assessment of reading and spelling abilities. Furthermore (3), a control group consisting of 11
children (CG) was investigated. We analyzed the functional MRI and DTI data in order to
address the central research question whether functional and structural patterns of brain
activity in children with isolated spelling impairments differ from those observed in children
with impairments in reading and spelling and in healthy controls.
Children in the spelling impaired (SI) group showed poor spelling skills (spelling scores one
to two standard deviations below average) along with average reading skills, whereas children
in the SRI group showed impairments in spelling and reading (spelling and reading scores one
to two standard deviations below average). Controls had significantly higher spelling scores
than both impaired groups (p< .05; see Table 1), while the comparison between the SI and the
SRI were not significant2. Furthermore, controls had significantly higher reading speed scores
than both impaired groups. The spelling impaired (SI) group had significantly higher reading
speed scores compared to the SRI group (p< .05; see Table 1). The same pattern was found
for reading comprehension (p< .05; see Table 1). The groups did not differ with respect to age
and non-verbal intelligence (p>.05; see Table 1).
1 Children participating in this study were investigated in another research context; see Gebauer et al. (submitted). 2 Revealed by specific post-hoc comparison by means of the Tukey HSD test.
STUDY I
32
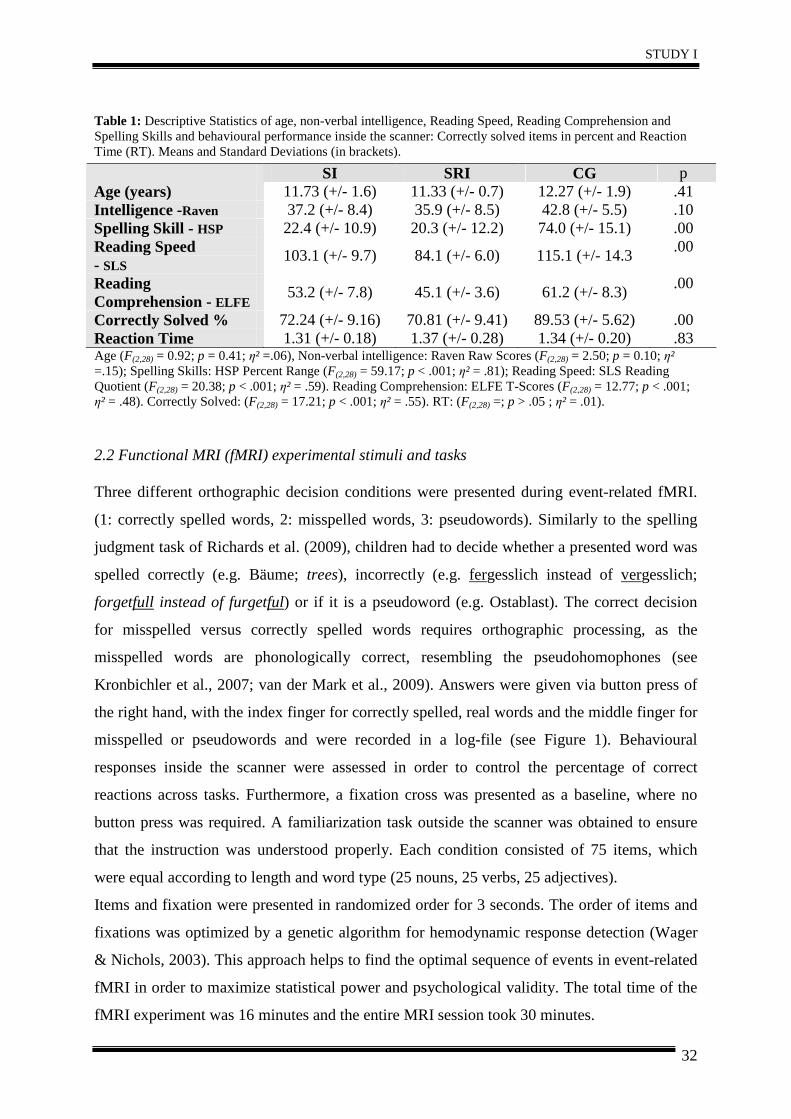
Table 1: Descriptive Statistics of age, non-verbal intelligence, Reading Speed, Reading Comprehension and Spelling Skills and behavioural performance inside the scanner: Correctly solved items in percent and Reaction Time (RT). Means and Standard Deviations (in brackets).
SI SRI CG p Age (years) 11.73 (+/- 1.6) 11.33 (+/- 0.7) 12.27 (+/- 1.9) .41 Intelligence -Raven 37.2 (+/- 8.4) 35.9 (+/- 8.5) 42.8 (+/- 5.5) .10 Spelling Skill - HSP 22.4 (+/- 10.9) 20.3 (+/- 12.2) 74.0 (+/- 15.1) .00 Reading Speed - SLS
103.1 (+/- 9.7) 84.1 (+/- 6.0) 115.1 (+/- 14.3 .00
Reading Comprehension - ELFE
53.2 (+/- 7.8) 45.1 (+/- 3.6) 61.2 (+/- 8.3) .00
Correctly Solved % 72.24 (+/- 9.16) 70.81 (+/- 9.41) 89.53 (+/- 5.62) .00 Reaction Time 1.31 (+/- 0.18) 1.37 (+/- 0.28) 1.34 (+/- 0.20) .83 Age (F(2,28) = 0.92; p = 0.41; η² =.06), Non-verbal intelligence: Raven Raw Scores (F(2,28) = 2.50; p = 0.10; η² =.15); Spelling Skills: HSP Percent Range (F(2,28) = 59.17; p < .001; η² = .81); Reading Speed: SLS Reading Quotient (F(2,28) = 20.38; p < .001; η² = .59). Reading Comprehension: ELFE T-Scores (F(2,28) = 12.77; p < .001; η² = .48). Correctly Solved: (F(2,28) = 17.21; p < .001; η² = .55). RT: (F(2,28) =; p > .05 ; η² = .01).
2.2 Functional MRI (fMRI) experimental stimuli and tasks
Three different orthographic decision conditions were presented during event-related fMRI.
(1: correctly spelled words, 2: misspelled words, 3: pseudowords). Similarly to the spelling
judgment task of Richards et al. (2009), children had to decide whether a presented word was
spelled correctly (e.g. Bäume; trees), incorrectly (e.g. fergesslich instead of vergesslich;
forgetfull instead of furgetful) or if it is a pseudoword (e.g. Ostablast). The correct decision
for misspelled versus correctly spelled words requires orthographic processing, as the
misspelled words are phonologically correct, resembling the pseudohomophones (see
Kronbichler et al., 2007; van der Mark et al., 2009). Answers were given via button press of
the right hand, with the index finger for correctly spelled, real words and the middle finger for
misspelled or pseudowords and were recorded in a log-file (see Figure 1). Behavioural
responses inside the scanner were assessed in order to control the percentage of correct
reactions across tasks. Furthermore, a fixation cross was presented as a baseline, where no
button press was required. A familiarization task outside the scanner was obtained to ensure
that the instruction was understood properly. Each condition consisted of 75 items, which
were equal according to length and word type (25 nouns, 25 verbs, 25 adjectives).
Items and fixation were presented in randomized order for 3 seconds. The order of items and
fixations was optimized by a genetic algorithm for hemodynamic response detection (Wager
& Nichols, 2003). This approach helps to find the optimal sequence of events in event-related
fMRI in order to maximize statistical power and psychological validity. The total time of the
fMRI experiment was 16 minutes and the entire MRI session took 30 minutes.

STUDY I
33
Figure 1: Overview of experimental design and measurement intervals. Correctly spelled words, misspelled
words, pseudowords and a fixation cross were presented in a randomized order for 3 seconds. In each orthographic decision condition participants were instructed to respond, by pressing either the “correct” button with the index finger [verfahren = to proceed] or the “misspelled/pseudoword” button with the middle finger on the response console [Techrot, Fortrag = lecture]. The responses were given with the right hand and recorded
and logged for further analyses.
2.3 Magnetic Resonance Imaging (MRI) data acquisition
Imaging was performed on a 3.0 Tesla Trio Tim scanner (Siemens Medical Systems,
Erlangen, Germany) using a 12-channel head coil. To minimize head movement, subjects’
heads were stabilized with foam cushions. A high resolution (1x1x1 mm) structural scan (TR
= 1900 ms, TE = 2.2 ms) was acquired to allow precise registration of functional data to
individual anatomy. Structural brain scans were reviewed by an expert and did not show
morphological abnormalities. Furthermore, diffusion tensor imaging (2x2x2.5 mm, TR =
6700 ms, TE = 95 ms, FOV = 250 mm, Flip Angle: 90°) data were obtained. Functional
images were acquired with a single-shot gradient echo EPI sequence (TR = 2190 ms, TE = 30
ms, FOV = 192 mm, Flip Angle = 90°, 36 three mm thick slices). Visual stimuli were
synchronized with the MR-scanner using the Software Presentation (Neurobehavioral
Systems, Albany, CA) and back-projected onto a translucent plastic screen which was
installed on the roof of the scanner bore. Participants watched the screen through a mirror
attached on the top of the head coil. Answers were given via button response box as described
above.
2.3.1. Behavioural data analyses
Task performance during fMRI recording (accuracy and reaction time of responding to the
experimental spelling tasks), was assessed by means of univariate ANOVA´s, using the
Tukey HSD test for post-hoc comparisons (PASW 18).

STUDY I
34
2.3.2. Functional Magnetic Resonance Imaging (fMRI) data analyses
Functional MRI data analysis was performed using FEAT (fMRI Expert Analysis Tool;
Version 4.1.5., part of FMRIB´s Software Library, www.fmrib.ox.ac.uk/fsl). The following
preprocessing steps were applied: motion correction using MCFLIRT; High-pass filter cut-off
100 s; non-brain removal using BET; interleaved slice time correction; spatial smoothing
using a Gaussian kernel of 6 mm FWHM; and high-pass temporal filtering. Time series
statistical analysis was carried out using FILM. The motion parameters were included in the
model as covariates of no interest. Nonlinear registration to high-resolution and standard
images (Montreal Neurological Institute (MNI) space) was carried out using FNIRT. Higher
level analysis was done using FLAME (FMRIB´s Local Analysis of Mixed Effects). If not
otherwise specified, Z statistic images were thresholded using clusters determined by Z >2.3
and a corrected cluster significance threshold of p = 0.05 (using Gaussian Random Field
Theory).
Analyses for the entire group were performed by computing linear t-contrasts between
selected experimental conditions for the orthographic decision task for each participant
individually, which were then entered into a random effects two sample t-test. For a detailed
overview of the FSL design matrix see Figure A.1 in the appendix.
According to our findings and theoretical background we computed five functional regions of
interest (ROI) in the right supramarginal gyrus (SMG), right superior parietal lobe (SPL),
right inferior frontal gyrus (triangularis), the medial frontal gyrus and the left occipito-
temporal (OT) region. ROI analyses were computed using FEATQUERY and PASW 18
(computing ANOVA´s along with Tukey HSD test for post-hoc comparisons; see Figure 4).
2.3.3. Diffusion tensor imaging (DTI) data analyses
Diffusion tensor imaging (DTI) analysis was performed using FDT (fMRIB´s Diffusion
Toolbox, Version v 2.0, part of fMRIB´s Software Library) and TBSS (Tract-Based Spatial
Statistics, Version v 1.2., part of fMRIB´s Software Library).
Raw images were pre-processed using eddy current correction. A brain mask was created
using BET (Brain Extraction Tool, Version 2.1). Maps for fractional anisotropy (FA), were
generated using DTIFit (FDT). Subsequently voxelwise statistical analysis of the FA data was
carried out using TBSS. Because standard templates do not appear appropriate for the smaller
head and brain size of children, a study-specific template was generated by registering the FA
images to the mean_FA of the group (created with the FMRIB58_FA image as the target).
The FA skeleton was thresholded at 0.20, to include major white matter pathways but avoid

STUDY I
35
peripheral tracts (vulnerable to intersubject variability; see Figure A.2 in the appendix). Each
subjects FA was then projected onto the mean skeleton. Voxel-wise cross-subject statistics
(TFCE thresholding, p<.05) using nonparametric testing as implemented in “randomize”
(5000 permutations), calculating group contrasts was applied. The anatomical location of the
significant clusters was determined by reference to the fibre tract-based atlas of human WM
(JHU ICBM-DTI-81 White-Matter Labels, JHU White-Matter Tractography Atlas, Juelich
Histological Atlas), implemented at FSL.
3. Results
3.1. Behavioral Performance inside the scanner
In order to investigate task performance during fMRI recording (as it was measured by means
of accuracy and reaction time of responding to the experimental spelling tasks), the ANOVA
yielded a significant GROUP effect. Post-hoc comparisons revealed that the controls reached
significantly higher percentage of correctly solved words than both impaired groups during
the fMRI session. The spelling impaired (SI) group and spelling and reading impaired (SRI)
group did not differ significantly (p< .001; see Table 1). No differences in reaction time were
found between groups (p > .05; see Table 1).
3.2. Functional MRI Results
3.2.1. Contrasts between Experimental Conditions:
In all three groups processing of the orthographic decision tasks (1: correctly spelled words;
2: misspelled words; 3: pseudowords) elicited activation in left pre- and postcentral gyrus
(button press) and large areas in the occipital and occipito-temporal cortex of both
hemispheres (visual input), relative to rest. The judgment of correctly spelled words evoked
additional activation in the left inferior frontal gyrus (IFG) in all three groups, and an
additional activation of the right IFG in the spelling impaired group (SI). The condition
misspelled words caused activation in the right inferior frontal gyrus in all three groups (see
Figure 2). For a more detailed overview of activation clusters and local maxima see Table A.1
in the appendix.
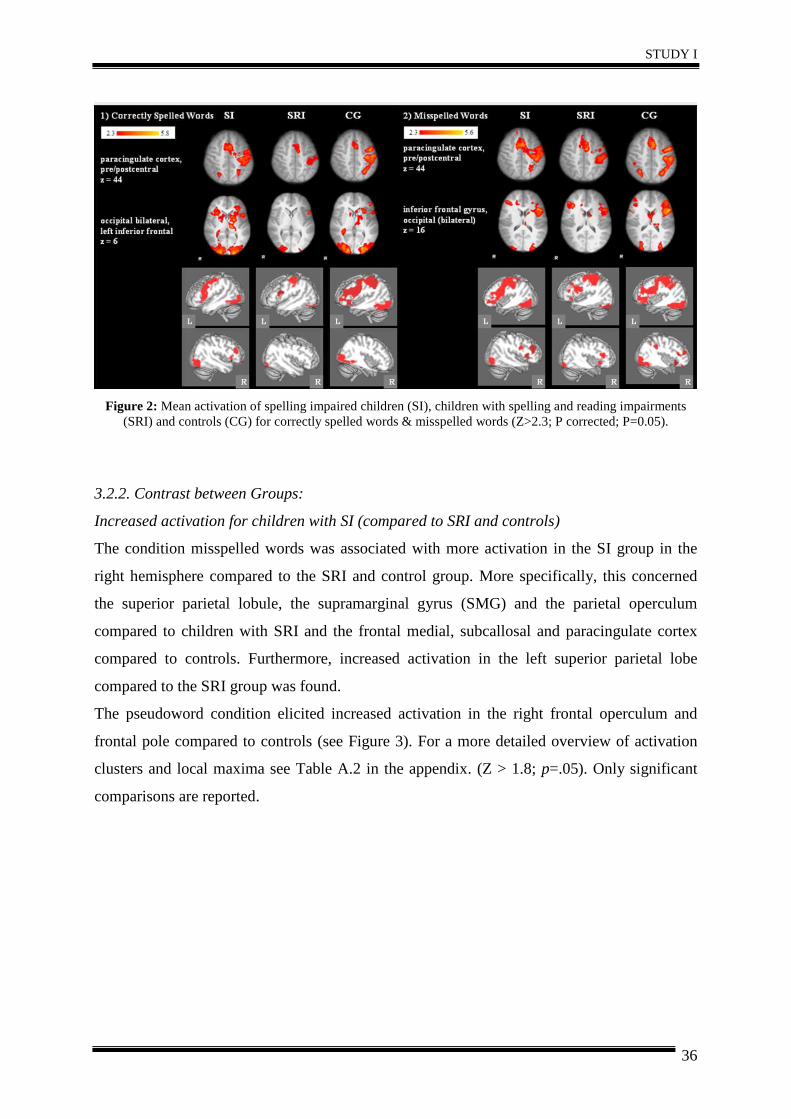
STUDY I
36
Figure 2: Mean activation of spelling impaired children (SI), children with spelling and reading impairments
(SRI) and controls (CG) for correctly spelled words & misspelled words (Z>2.3; P corrected; P=0.05).
3.2.2. Contrast between Groups:
Increased activation for children with SI (compared to SRI and controls)
The condition misspelled words was associated with more activation in the SI group in the
right hemisphere compared to the SRI and control group. More specifically, this concerned
the superior parietal lobule, the supramarginal gyrus (SMG) and the parietal operculum
compared to children with SRI and the frontal medial, subcallosal and paracingulate cortex
compared to controls. Furthermore, increased activation in the left superior parietal lobe
compared to the SRI group was found.
The pseudoword condition elicited increased activation in the right frontal operculum and
frontal pole compared to controls (see Figure 3). For a more detailed overview of activation
clusters and local maxima see Table A.2 in the appendix. (Z > 1.8; p=.05). Only significant
comparisons are reported.
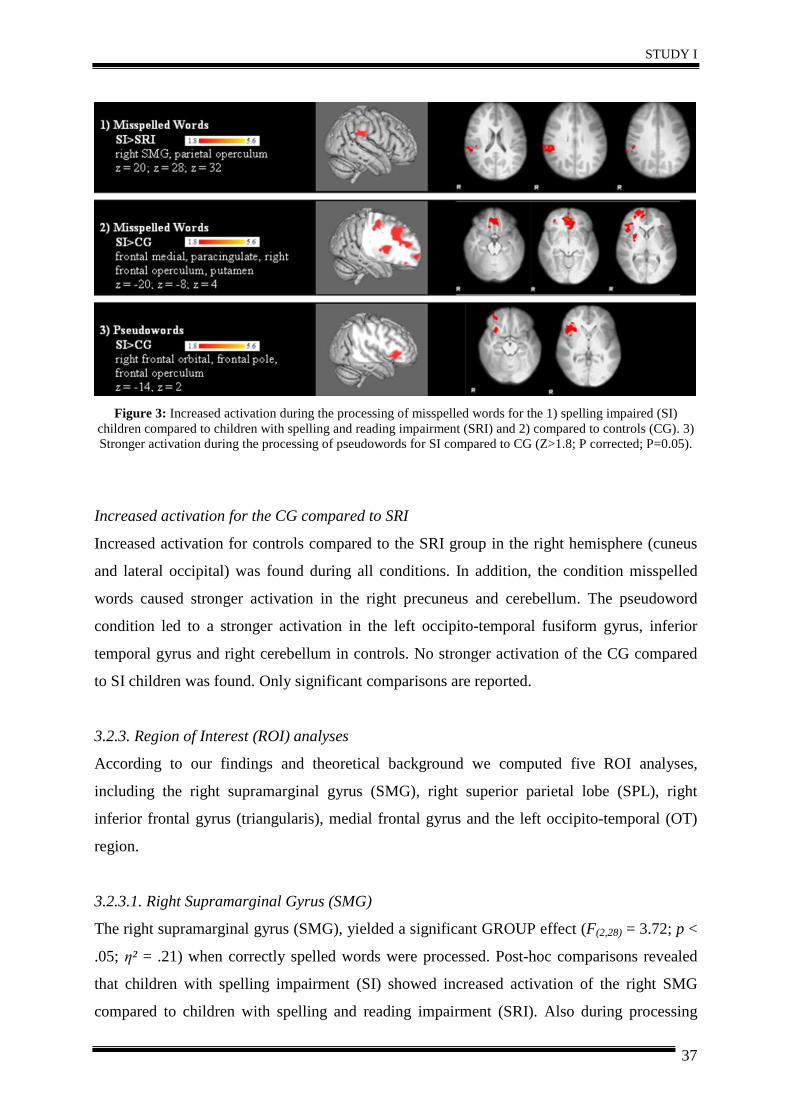
STUDY I
37
Figure 3: Increased activation during the processing of misspelled words for the 1) spelling impaired (SI)
children compared to children with spelling and reading impairment (SRI) and 2) compared to controls (CG). 3) Stronger activation during the processing of pseudowords for SI compared to CG (Z>1.8; P corrected; P=0.05).
Increased activation for the CG compared to SRI
Increased activation for controls compared to the SRI group in the right hemisphere (cuneus
and lateral occipital) was found during all conditions. In addition, the condition misspelled
words caused stronger activation in the right precuneus and cerebellum. The pseudoword
condition led to a stronger activation in the left occipito-temporal fusiform gyrus, inferior
temporal gyrus and right cerebellum in controls. No stronger activation of the CG compared
to SI children was found. Only significant comparisons are reported.
3.2.3. Region of Interest (ROI) analyses
According to our findings and theoretical background we computed five ROI analyses,
including the right supramarginal gyrus (SMG), right superior parietal lobe (SPL), right
inferior frontal gyrus (triangularis), medial frontal gyrus and the left occipito-temporal (OT)
region.
3.2.3.1. Right Supramarginal Gyrus (SMG)
The right supramarginal gyrus (SMG), yielded a significant GROUP effect (F(2,28) = 3.72; p <
.05; η² = .21) when correctly spelled words were processed. Post-hoc comparisons revealed
that children with spelling impairment (SI) showed increased activation of the right SMG
compared to children with spelling and reading impairment (SRI). Also during processing

STUDY I
38
misspelled words a significant GROUP effect (F(2,28) = 7.55; p < .05; η² = .35) emerged. Post-
hoc comparisons revealed increased activation for SI compared to controls and SRI (see Table
2, Figure 4). No significant group effect was found for the condition pseudowords.
3.2.3.2. Right Superior Parietal Lobe (SPL)
Significant GROUP effects were found during all orthographic decision tasks (Correct: F(2,28)
= 4.30; p < .05; η² = .24; Misspelled: F(2,28) = 4.39; p < .05; η² = .24; Pseudowords: F(2,28) =
4.83; p < .05; η² = .26). Post-hoc comparisons revealed increased activation for spelling
impaired children (SI) compared to SRI when correctly spelled words were presented and
increased activation for SI children and controls compared to SRI when misspelled words or
pseudowords were presented (see Table 2, Figure 4).
Figure 4: ROI with significant GROUP effect. Above = right superior parietal lobe and right supramarginal
gyrus. Below = medial frontal gyrus and left occipito-temporal region. Correctly Spelled Words (C), Misspelled Words (M) and Pseudowords (P).
3.2.3.3. Right Inferior Frontal Gyrus (IFG)
For the right IFG no significant GROUP effects were observed.
3.2.3.4. Medial Frontal Gyrus
A significant GROUP effect for the medial frontal gyrus was observed (F(2,28) = 5.38; p < .05;
η² = .28) when misspelled words were presented. Post-hoc comparisons revealed decreased
activation for controls compared to spelling impaired children (SI) and tentatively decreased
activation of controls compared to SRI (p = .056; see further Table 2, Figure 4).
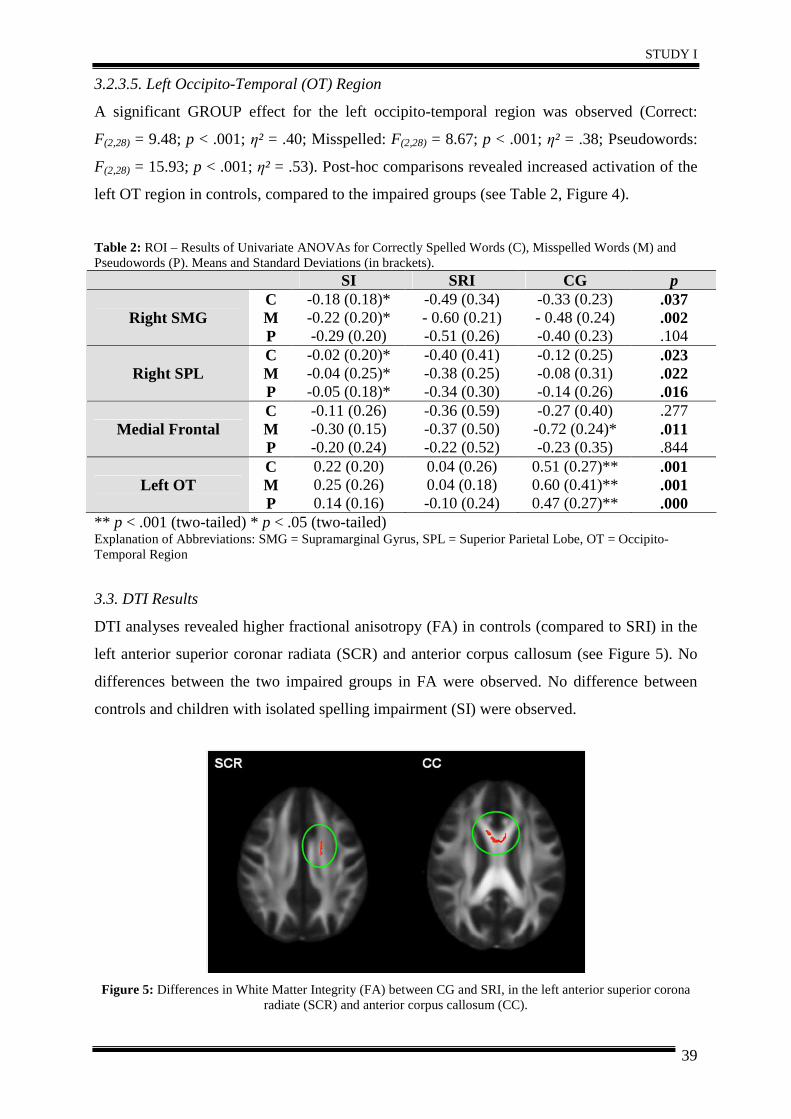
STUDY I
39
3.2.3.5. Left Occipito-Temporal (OT) Region
A significant GROUP effect for the left occipito-temporal region was observed (Correct:
F(2,28) = 9.48; p < .001; η² = .40; Misspelled: F(2,28) = 8.67; p < .001; η² = .38; Pseudowords:
F(2,28) = 15.93; p < .001; η² = .53). Post-hoc comparisons revealed increased activation of the
left OT region in controls, compared to the impaired groups (see Table 2, Figure 4).
Table 2: ROI – Results of Univariate ANOVAs for Correctly Spelled Words (C), Misspelled Words (M) and Pseudowords (P). Means and Standard Deviations (in brackets).
SI SRI CG p
Right SMG C M P
-0.18 (0.18)* -0.22 (0.20)* -0.29 (0.20)
-0.49 (0.34) - 0.60 (0.21) -0.51 (0.26)
-0.33 (0.23) - 0.48 (0.24) -0.40 (0.23)
.037
.002
.104
Right SPL C M P
-0.02 (0.20)* -0.04 (0.25)* -0.05 (0.18)*
-0.40 (0.41) -0.38 (0.25) -0.34 (0.30)
-0.12 (0.25) -0.08 (0.31) -0.14 (0.26)
.023
.022
.016
Medial Frontal C M P
-0.11 (0.26) -0.30 (0.15) -0.20 (0.24)
-0.36 (0.59) -0.37 (0.50) -0.22 (0.52)
-0.27 (0.40) -0.72 (0.24)* -0.23 (0.35)
.277
.011
.844
Left OT C M P
0.22 (0.20) 0.25 (0.26) 0.14 (0.16)
0.04 (0.26) 0.04 (0.18) -0.10 (0.24)
0.51 (0.27)** 0.60 (0.41)** 0.47 (0.27)**
.001
.001
.000 ** p < .001 (two-tailed) * p < .05 (two-tailed) Explanation of Abbreviations: SMG = Supramarginal Gyrus, SPL = Superior Parietal Lobe, OT = Occipito-Temporal Region
3.3. DTI Results
DTI analyses revealed higher fractional anisotropy (FA) in controls (compared to SRI) in the
left anterior superior coronar radiata (SCR) and anterior corpus callosum (see Figure 5). No
differences between the two impaired groups in FA were observed. No difference between
controls and children with isolated spelling impairment (SI) were observed.
Figure 5: Differences in White Matter Integrity (FA) between CG and SRI, in the left anterior superior corona
radiate (SCR) and anterior corpus callosum (CC).

STUDY I
40
4. Discussion
This study investigates for the first time patterns of brain activity in spelling impaired children
(SI) in comparison to children with spelling and reading impairment (SRI) and controls. As a
major finding, children with isolated spelling impairment showed a stronger right hemispheric
activation during processing orthographic decision tasks compared to children with SRI and
controls. This could reflect a compensatory mechanism, allowing German-speaking children
with isolated spelling deficits to avoid reading deficits by gaining word specific knowledge
through intact serial grapheme-phoneme decoding (Moll & Landerl, 2009). This
compensatory mechanism is successful for the compensation of reading deficits, whereas
spelling impairments remain, because of the asymmetry in German language (grapheme-
phoneme correspondence is high, but phoneme-grapheme correspondence is low). Equal word
spellings with different pronunciations (e.g. English the wind – to wind) do not exist in
German, whereas different spellings for words with the same pronunciation are common (e.g.
Wal / whale – Wahl / election). Hence, it is not possible to derive the correct spelling by this
serial phoneme-grapheme decoding strategy.
In line with this assumption, region of interest analyses revealed increased activation in right
posterior areas (SMG and SPL) related to grapheme-phoneme conversion (Booth et al., 2002,
2004), attention (Peyrin et al., 2010) and working memory (Berryhill & Olson, 2008; Koenigs
et al., 2009), in spelling impaired children. Hoeft et al. (2011) found that dyslexic children
who relied on right hemispheric pathways showed gains in reading. This further supports our
theory that the stronger right hemispheric engagement of spelling impaired children may
reflect a compensatory mechanism, related to adjustment of reading skills. The activation of
superior parietal regions could indicate increased attentional and working memory effort in
this group in order to overcome dysfunctions.
In addition (comparing mean activation maps of the groups), children with spelling
impairment activated bilateral inferior and middle frontal gyri during processing correctly
spelled words and misspelled words, whereas the other two groups showed left frontal
activation for the correctly spelled words and bilateral activation only in the misspelled
condition. Increased activation of the left and right inferior frontal gyri (IFG) has often been
interpreted as a compensatory process of dyslexics in order to overcome dysfunctions in left
posterior cortical areas subserving phonological and orthographic processing (Hoeft et al.,
2011). We found an increased right frontal engagement in spelling impaired children
compared to controls, although surprisingly not in the IFG (triangularis) but in adjacent
regions (frontal orbital region, frontal pole). The right IFG engagement occurred in all three
groups in the more difficult condition (misspelled words). Probably this additional bilateral

STUDY I
41
IFG engagement is reflecting a more general increase of activation because of higher task
demand, apparent in impaired children and non-impaired children. Nevertheless, we found
increased additional activation in the frontal medial region, related to more effortful and
attentionally guided reading strategy (Meyler et al., 2008) for both impaired groups.
According to current literature (Kronbichler et al., 2006; Richlan et al. 2009; Schurz et al.,
2010), children with difficulties in spelling and reading (SRI) exhibit decreased cortical
activation during orthographic decision tasks in left occipito-temporal (OT) regions compared
to controls, most prominently during the presentation of pseudowords. A region of interest
analyses revealed decreased activation of the left OT region for SI and SRI children, but with
a higher extent for the SRI. Also, decreased occipito-parietal activation (relative to controls)
in children with SRI was observed, probably indicating reduced serial guidance of visual
attention (Cohen et al., 2008; Schurz et al., 2010). Increased activation of controls (relative to
SRI) was observed in the precuneal and cuneal cortex, these regions are involved in attention,
semantic processing and most notably with the default-mode network (Binder et al., 2009;
Cavanna & Trimble, 2006; Graves et al., 2010). Higher activation in this region is related to
more self-referential thought and less task engagement. It seems that children with reading
and spelling impairments had to concentrate more on the task.
Activation in non-impaired controls was observed in expected regions of reading network
(e.g. occipital regions for visual processing, left occipito-temporal regions involved in visual
word and letter string recognition, left temporo-parietal regions involved in grapheme-
phoneme computation and phonological word processing, left inferior frontal regions; see
further Jobard et al., 2003) and spelling network (e.g. left supramarginal gyrus involved in
mapping between orthography and phonology, left posterior temporal lobe related to an
orthographic lexicon; see further Beeson et al., 2003; Booth et al., 2002, 2004; Rapcsak &
Beeson, 2004).
DTI analyses revealed higher frontal white matter integrity (assessed by means of FA) in
controls (compared to SRI) in the left anterior superior corona radiate (SCR) and anterior
corpus callosum, probably indicating less efficient communication in children with SRI.
Indications for a less efficient communication in bilateral fronto-temporal and left temporo-
parietal white matter regions were also found in German dyslexic adults (Steinbrink et al.,
2008), assuming that a disconnection syndrome of anterior and posterior regions involved in
reading and spelling might be underlying dyslexia. No difference of left parieto-temporal
white matter connectivity was found between our groups. Probably, differences in parieto-
temporal white matter integrity emerge through inefficient use and are therefore not yet
apparent in our young German-speaking sample. Nagy et al. (2004) also found correlations

STUDY I
42
between frontal fractional anisotropy (FA) in the anterior corpus callosum and working
memory capacity. Unfortunately, no working memory assessment was included in the present
study, but it is likely that working memory (important for the process of reading) is affected
in children with SRI. The SCR has been related to reading performance in children (e.g.
Beaulieu et al., 2005; Deutsch et al., 2005), possibly explaining that differences were only
observed for children with reading impairments. No evidence for differences in white matter
integrity between children with spelling impairment compared to controls were found,
supporting the theory of a functional compensatory mechanism, which probably induce also
structural changes, in spelling impaired children.
5. Conclusion
We investigated whether or to which extent dissociations between spelling impaired children
(SI) and children with spelling and reading impairments (SRI) exist in functional patterns of
brain activity. It was found that children with isolated spelling difficulties show different
patterns of brain activation compared to children with both reading and spelling difficulties.
Our study suggests that spelling impaired children compensate for probably initially existent
reading impairments by an increased recruitment of right posterior areas (SMG and SPL)
related to grapheme-phoneme conversion, attention and working memory. DTI analyses
revealed stronger frontal white matter integrity in controls, compared to spelling and reading
impaired children, whereas no structural differences between controls and children with
isolated spelling impairment were observed. This finding seems to support our theory of a
functional compensatory mechanism, which probably induce also structural changes.
Acknowledgments: The research presented in this paper was supported by grants from the Styrian government (Nr. A27214001062) and the Jubilee Fund of the Austrian National Bank (Nr. A26E16020013). The authors wish to express their large gratitude to Nadja Kozel, Bernd Schneeberger, Johanna Vogl and Stefanie Rohrer who greatly contributed to this research project.

STUDY II
43
STUDY II – Functional changes related to intervention
Differences in Brain Function and Changes with
Intervention in Spelling Impaired Children
D. Gebauer 1,2, A. Fink 2*, R. Kargl 3, G. Reishofer 4, K. Koschutnig 2,4, C. Purgstaller 3, F. Fazekas 1, C. Enzinger 1,4
1 Department of Neurology, Medical University of Graz, Austria 2 Department of Psychology, Karl-Franzens-University Graz, Austria
3 Institute for Reading and Spelling in Graz, Austria 4 Division of Neuroradiology, Department of Radiology, Medical University of Graz, Austria
(Submitted to PLoS ONE on 14 December 2011)

STUDY II
44
Abstract
Previous fMRI studies in English-speaking samples suggested that specific interventions may
alter brain function in language-relevant networks in children with reading and spelling
difficulties, but this research strongly focused on reading impaired individuals. Only few
studies so far investigated characteristics of brain activation specifically associated with
spelling impairment and whether a specific spelling intervention may also be associated with
distinct changes in brain activity patterns. We here investigated such effects of a morpheme-
based spelling intervention on brain function in 20 spelling impaired children using repeated
fMRI. Relative to 10 matched controls, children with poor spelling abilities showed increased
activation in frontal medial and right hemispheric regions and decreased activation in left
occipito-temporal regions prior to the intervention, during processing of an orthographic
decision task. After five weeks of intervention, spelling and reading comprehension
significantly improved in the training group, along with increased activation in the left
temporal, parahippocampal and hippocampal regions. Conversely, the waiting group showed
increases in right posterior regions. Our findings could indicate an increased left temporal
activation associated with the recollection of the new learnt morpheme-based strategy related
to successful training.
KEYWORDS: fMRI; hippocampus; spelling impairment; neural plasticity

STUDY II
45
1. Introduction
Depending on definition and stringency of criteria used, approximately 3-16% of children
show difficulties in reading and spelling (Gabrieli, 2009; Habib, 2000; Klicpera et al., 2007)
that may be associated with a greater risk of school anxiety, unemployment and multiple
emotional and behavioural difficulties (Ise & Schulte-Körne, 2010). A severe and well-known
form of reading and spelling difficulty is dyslexia. Recent studies using functional magnetic
resonance imaging (fMRI) provided important insights into potential brain mechanisms
underlying dyslexia. Frequently, decreased brain activation in parieto-temporal and occipito-
temporal regions of the left hemisphere, along with increased activation in frontal and right
hemispheric language-related regions has been observed in individuals with dyslexia.
Decreased left parieto-temporal activation has been related to deficits in grapheme-phoneme-
conversion. The left angular gyrus (and left supramarginal gyrus) is assumed to play a critical
role in spelling (Booth et al., 2002, 2004). Occipito-temporal activation has been related to
automatic and fluent reading (Kronbichler et al., 2008; Shaywitz et al., 2006). The increased
frontal and right hemispheric activation has been interpreted to indicate inefficient
compensatory mechanisms such as internal articulation (Richlan et al., 2009; Maisog et al.,
2008).
Training studies using fMRI revealed changes in brain activation patterns along with
successful intervention (Aylward et al., 2003; Eden et al., 2004; Gaab et al., 2007; Meyler et
al., 2008; Richards et al., 2006; Shaywitz et al., 2004; Simos et al., 2002, 2006; Temple et al.,
2003). However, the majority of these findings comes from reading impaired individuals (e.g.
Eden et al., 2000; Meyler et al., 2007; Richlan et al., 2010; Shaywitz et al., 2007), while only
few studies focused on subjects with spelling impairment (e.g. Richards et al., 2009).
Isolated spelling difficulties in German-speaking samples were found in about 3-6% (Moll &
Landerl, 2009; Wimmer & Mayringer, 2002). Due to the transparent orthography in German,
dyslexics manage to read slow but accurate in the course of their development, whereas
spelling mistakes rather persist into adulthood (Landerl & Klicpera, 2009).
Here, we used a morpheme-based spelling intervention (Morpheus; Kargl & Purgstaller,
2010) which trains children to figure out the correct spelling of a word by separating it into
specific parts (morphemes). Behavioral studies in this field provided evidence that such
interventions significantly enhance reading and/or spelling ability (Arnbak & Elbro, 2000;
Lyster, 2002; Nunes et al., 2003; Weiss et al., 2010). Using EEG, we demonstrated a
neurophysiological training effect of this intervention, by increased EEG activation in left

STUDY II
46
hemispheric regions that are involved in the complex neural network subserving reading and
spelling (Weiss et al., 2010).
Using fMRI in a different sample, we sought to corroborate and extend these findings by
more detailed functional neuroanatomical insights. We specifically tested (a) if and how brain
activation patterns at baseline in spelling impaired children differed from controls. We
hypothesized that children with poor spelling abilities and controls would show different brain
activation prior to the applied spelling intervention, probably also in non-speech-relevant
regions. Secondly (b), we investigated whether spelling skills and brain activation can be
modulated by a specific spelling intervention, comparing two spelling impaired groups
(divided into a training group and into a waiting group).
2. Materials and Methods
2.1 Ethics Statement
The study was approved by the ethics committee of the Medical University of Graz, Austria.
All participants and their parents had given written informed consent.
2.2 Participants
Forty-two German-speaking children aged between nine and 15 years were recruited for this
study based on extensive behavioral pre-screening as described below (cf. 2.2). Three groups
(training group, waiting group and control group), each comprising 14 subjects, were
investigated. Children with overall motion > 3 mm or sudden movement > 1 mm during
scanning were excluded from further analyses. Based on this definition, twelve children had
to be excluded due to movement artifacts (n = 7). Furthermore, children had to be excluded
due to poor behavioral performance inside the scanner (n = 2; Mean Accuracy < 70%) or
because they did not attend all behavioral tests and fMRI sessions (n = 3), rendering a final
sample of 30 children (15 males), whose age ranged from 10 to 15 years (M = 11.80; SD =
1.58, see Table 1). All participants were right-handed. All participants were healthy, right-
handed and had normal or corrected-to normal vision.
We formed and investigated three experimental groups: (1) Ten children with below average
spelling abilities were assigned to the “training group” (TG), (2) another ten children with
poor spelling abilities were assigned to the so-called “waiting group” (WG, receiving the
training after the post-test) and (3) a control group (CG) of ten children (matched for age and
intelligence) with average spelling abilities, assessed at a single time-point, were investigated.
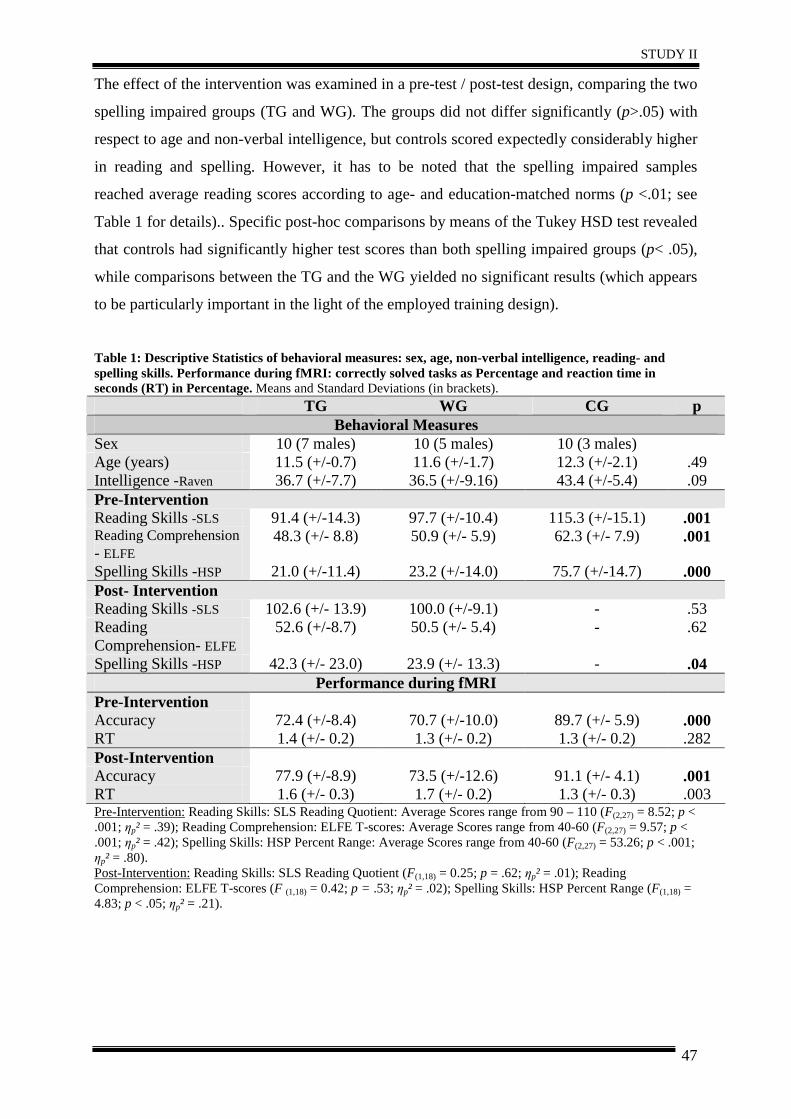
STUDY II
47
The effect of the intervention was examined in a pre-test / post-test design, comparing the two
spelling impaired groups (TG and WG). The groups did not differ significantly (p>.05) with
respect to age and non-verbal intelligence, but controls scored expectedly considerably higher
in reading and spelling. However, it has to be noted that the spelling impaired samples
reached average reading scores according to age- and education-matched norms (p <.01; see
Table 1 for details).. Specific post-hoc comparisons by means of the Tukey HSD test revealed
that controls had significantly higher test scores than both spelling impaired groups (p< .05),
while comparisons between the TG and the WG yielded no significant results (which appears
to be particularly important in the light of the employed training design).
Table 1: Descriptive Statistics of behavioral measures: sex, age, non-verbal intelligence, reading- and spelling skills. Performance during fMRI: correctly solved tasks as Percentage and reaction time in seconds (RT) in Percentage. Means and Standard Deviations (in brackets).
TG WG CG p Behavioral Measures
Sex 10 (7 males) 10 (5 males) 10 (3 males) Age (years) 11.5 (+/-0.7) 11.6 (+/-1.7) 12.3 (+/-2.1) .49 Intelligence -Raven 36.7 (+/-7.7) 36.5 (+/-9.16) 43.4 (+/-5.4) .09 Pre-Intervention Reading Skills -SLS 91.4 (+/-14.3) 97.7 (+/-10.4) 115.3 (+/-15.1) .001 Reading Comprehension - ELFE
48.3 (+/- 8.8) 50.9 (+/- 5.9) 62.3 (+/- 7.9) .001
Spelling Skills -HSP 21.0 (+/-11.4) 23.2 (+/-14.0) 75.7 (+/-14.7) .000 Post- Intervention Reading Skills -SLS 102.6 (+/- 13.9) 100.0 (+/-9.1) - .53 Reading Comprehension- ELFE
52.6 (+/-8.7) 50.5 (+/- 5.4) - .62
Spelling Skills -HSP 42.3 (+/- 23.0) 23.9 (+/- 13.3) - .04 Performance during fMRI
Pre-Intervention Accuracy 72.4 (+/-8.4) 70.7 (+/-10.0) 89.7 (+/- 5.9) .000 RT 1.4 (+/- 0.2) 1.3 (+/- 0.2) 1.3 (+/- 0.2) .282 Post-Intervention Accuracy 77.9 (+/-8.9) 73.5 (+/-12.6) 91.1 (+/- 4.1) .001 RT 1.6 (+/- 0.3) 1.7 (+/- 0.2) 1.3 (+/- 0.3) .003 Pre-Intervention: Reading Skills: SLS Reading Quotient: Average Scores range from 90 – 110 (F(2,27) = 8.52; p < .001; ηp² = .39); Reading Comprehension: ELFE T-scores: Average Scores range from 40-60 (F(2,27) = 9.57; p < .001; ηp² = .42); Spelling Skills: HSP Percent Range: Average Scores range from 40-60 (F(2,27) = 53.26; p < .001; ηp² = .80). Post-Intervention: Reading Skills: SLS Reading Quotient (F(1,18) = 0.25; p = .62; ηp² = .01); Reading Comprehension: ELFE T-scores (F (1,18) = 0.42; p = .53; ηp² = .02); Spelling Skills: HSP Percent Range (F(1,18) = 4.83; p < .05; ηp² = .21).

STUDY II
48
2.3 Psychometric Tests
In the pre-experimental screening, standardized tests for the assessment of reading and
spelling abilities were administered in 107 subjects, and we explored relevant socio-
demographic data such as age, sex and native language.
To assess spelling skills, we used a standardized spelling test (Hamburger-Schreibprobe,
HSP) by May et al. (2000). In the HSP, words and sentences are dictated by the experimenter
and have to be written next to the corresponding pictures that illustrate the respective words
or sentences. This test takes about 15 minutes. Within this study, versions for 4th/5th graders
and 5th to 9th graders were applied. The HSP provides measures for the number of correctly
spelled words and the number of grapheme-related mistakes. The latter measure was used in
this study as it provides a more precise measure of spelling ability.
Additionally, we administered the “Salzburger-Lese-Sreening” (SLS; Mayringer & Wimmer,
2003, 2005) that measures reading speed and basic reading ability (automaticity, accuracy).
The SLS 1-4 was used for children up to the 4th grade, and the SLS 5-8 was applied for older
children and parallel versions exist for both. In the SLS, children have to decide whether the
content of a presented sentence is correct or not. Testing time is limited to three minutes. In
addition, we also measured reading comprehension (i.e. comprehension of words, sentences
and text) by means of a standardized German-speaking test (ELFE 1-6; Lenhard & Schneider,
2006). Furthermore, non-verbal intelligence was measured by the Standard Progressive
Matrices (SPM) by Raven (1960).
2.4 Intervention
The applied intervention is a computer-aided morpheme-based spelling training (Morpheus;
Kargl & Purgstaller, 2010), which has been approved as an evidence-based intervention for
individuals with reading and spelling deficits by the federal ministry of Austria and has shown
to significantly improve spelling ability in children in a series of behavioral studies in our
laboratory (Kargl et al., 2008, 2011).
The Morpheus-intervention consists of computerized tutorials, a book of exercises and
morpheme-based games to facilitate the consolidation of the strategy. The intervention, which
includes daily handwritten and computer homework along with instructor-guided courses
(once a week, lasting approximately two hours), was realized within five weeks. These
tutorials on the computer include 12 different playful exercises dealing with morphemes (e.g.
recognizing and matching word families, morphological clozes, finding suffixes and
prefixes). During the tutorials achieved scores are displayed on the computer screen.
Participants can only reach the next difficulty level of the same exercise when they have

STUDY II
49
solved at least 75% of the given problems correctly. The automatically saved score of every
tutorial serves as basis for assessing training progress.
The training material of Morpheus consists of the most frequent morphemes of the German
language and contains different levels of difficulty. The words used for the training were
taken from an empirically-based collection of words (German basic vocabulary for 4th
graders; Augst, 1989). Morpheus has been constructed on the basis of the following
principles: simplicity, relief due to morpheme segmentation, rule-governed repetition,
playfulness, avoidance of mistakes, individuality, productivity, and practicing handwriting.
A morpheme is defined as the “smallest meaningful unit of language” (Bhatt, 1991). Every
word is built by different parts, which follow particular spellings (e.g. unforgetful = prefix
[un], suffix [ful], root [forget]). Therefore, the spelling of the German verb “verfahren” can be
derived by two rules: the prefix [ver] is always written with [v], the root [fahr] always with an
“h”. Children do not need to remember the spelling of every single word, but only to
memorize the spelling of their component parts. Furthermore, morphosemantic information
can support the development of a meaning-oriented decoding strategy, e.g. the correct
spelling of the noun “Motor-rad” (motor-bike) can be derived by the meaning (May et al.,
2000). In addition, this strategy seems to be easy to apply as only “100 of the most frequent
morphemes cover 70% of all written material” (Scheerer-Neumann, 1979).
2.5 Functional MRI (fMRI) experimental stimuli and tasks
Three different orthographic decision conditions were presented during event-related fMRI
(1: correctly spelled words, 2: misspelled words, 3: pseudowords). Each condition comprised
75 words (25 nouns, 25 verbs, 25 adjectives; mean word length: 7 letters).
Similarly to the spelling judgment task of Richards et al. (2009), children had to decide
whether a presented word was spelled correctly (e.g. Bäume; trees), incorrectly (e.g. Menner
instead of Männer; men) or if it was a pseudoword (e.g. Ostablast). The correct decision for
misspelled versus correctly spelled words requires orthographic processing, as the misspelled
words are phonologically correct, resembling the pseudohomophones (see Kronbichler et al.,
2007; van der Mark et al., 2009). Answers were given via button presses using the right
(dominant) hand, with the index finger for correctly spelled real words and the middle finger
for misspelled and pseudowords (see Figure 1). Behavioral responses inside the scanner were
assessed to obtain the percentage of correct responses and reaction time. Furthermore, a
fixation cross was presented as a baseline, with no button press required. Participants were
familiarized with the task outside the scanner to ensure the instruction had been understood
properly. Each condition consisted of 75 items, which were equal according to length and

STUDY II
50
word type (25 nouns, 25 verbs, 25 adjectives). Items and fixation were presented for three
seconds. Each condition was directly followed by the other. Participants had to respond
within the presentation interval of three seconds. The order of items and fixations was
optimized by a genetic algorithm for hemodynamic response detection (Wager & Nichols,
2003). The total time of the fMRI experiment was 16 minutes and the entire MRI session took
30 minutes.
Figure 1: fMRI Paradigm. Correctly spelled words, misspelled words, pseudowords and a fixation cross were presented in a randomized order for three seconds. In each orthographic decision condition, participants were
instructed to respond by either pressing the “correct” button with the index finger or the “misspelled/pseudoword” button with the middle finger on the response console. Responses were given with the
right hand and recorded and logged for further analyses. Subjects did not receive feedback to their responses.
2.6 Magnetic Resonance Imaging (MRI) data acquisition and analysis
Imaging was performed on a 3.0 Tesla Trio Tim scanner (Siemens Medical Systems,
Erlangen, Germany) using a 12-channel head coil. To minimize head movement, subjects’
heads were stabilized with foam cushions. A high-resolution isotropic (1x1x1 mm) structural
scan (TR = 1900 ms, TE = 2.2 ms) was acquired to allow precise registration of functional
data to individual anatomy. Structural brain scans were reviewed by an expert and did not
show morphological abnormalities. Functional images were acquired using a single-shot
gradient echo EPI sequence (TR = 2190 ms, TE = 30 ms, matrix 64 x 64 mm, FOV = 192,
Flip Angle 90°, 36 three mm thick slices). Visual stimuli were synchronized with the MR-
scanner using “Presentation” (Neurobehavioral Systems, Albany, CA) and back-projected
onto a translucent screen installed on the rear of the scanner bore. Participants watched the
screen through a mirror attached on the top of the head coil. Answers were given via a button
response box as described above.
Functional MRI data analysis was performed using FEAT (fMRI Expert Analysis Tool;
Version 4.1.5., part of FMRIB´s Software Library, www.fmrib.ox.ac.uk/fsl). The following
preprocessing steps were applied: motion correction using MCFLIRT; non-brain removal
using BET; interleaved slice time correction; spatial smoothing using a Gaussian kernel of 6

STUDY II
51
mm FWHM; and high-pass temporal filtering. Time series statistical analysis was carried out
using FILM. Motion parameters were included in the model as covariates of no interest.
Nonlinear registration to high-resolution and standard images (Montreal Neurological
Institute (MNI) space) was carried out using FNIRT. Higher level analysis was done using
FLAME (FMRIB´s Local Analysis of Mixed Effects). Z statistic images were thresholded
using clusters determined by Z >2.0 and a corrected cluster significance threshold of p = 0.05
(using Gaussian Random Field Theory).
Analyses for the entire group were performed by computing linear t-contrasts between
selected experimental conditions for the orthographic decision task for each participant, which
were then entered into a random effects two-sample t-test. To examine the correlation
between behavioral improvement and activation increase, as well as interaction effects
between increases in the TG and WG, we ran second-level (fixed effects) analyses for each
subject to calculate the differences between activation patterns (pre vs. post activation).
Subsequently, group level analyses (mixed effects) were run, including the number of
incorrect responses inside the scanner as covariate of no interest.
3. Results
3.1. Baseline differences in brain activation patterns (Pre-Intervention)
To test our first hypothesis, we looked for group differences (TG, WG and CG) prior to the
intervention (pre-test). Specifically, we analyzed whether the two spelling impaired groups
(TG, WG) displayed comparable brain activation patterns before the intervention. In addition,
we also investigated potential differences in brain activation between poor (TG, WG) and
good (CG) spellers. The number of incorrect responses inside the scanner was included as
covariate of no interest. There were only minor differences in brain activation between the TG
and the WG during processing of misspelled words before the intervention. For the TG,
increased activation in the left precuneus and left anterior cingulate gyrus was observed
(Table 2).
The pre-intervention comparison revealed increased activation in left occipito-temporal
regions and in the cerebellum for the CG compared to both spelling impaired samples (TG
and WG). Beyond that, the CG exhibited increased activation in the left lateral occipital
cortex, left inferior temporal gyrus, and left hippocampus relative to the TG, and increased
activation in the bilateral lateral occipital cortex and bilateral temporal regions compared to
the WG (Table 2).
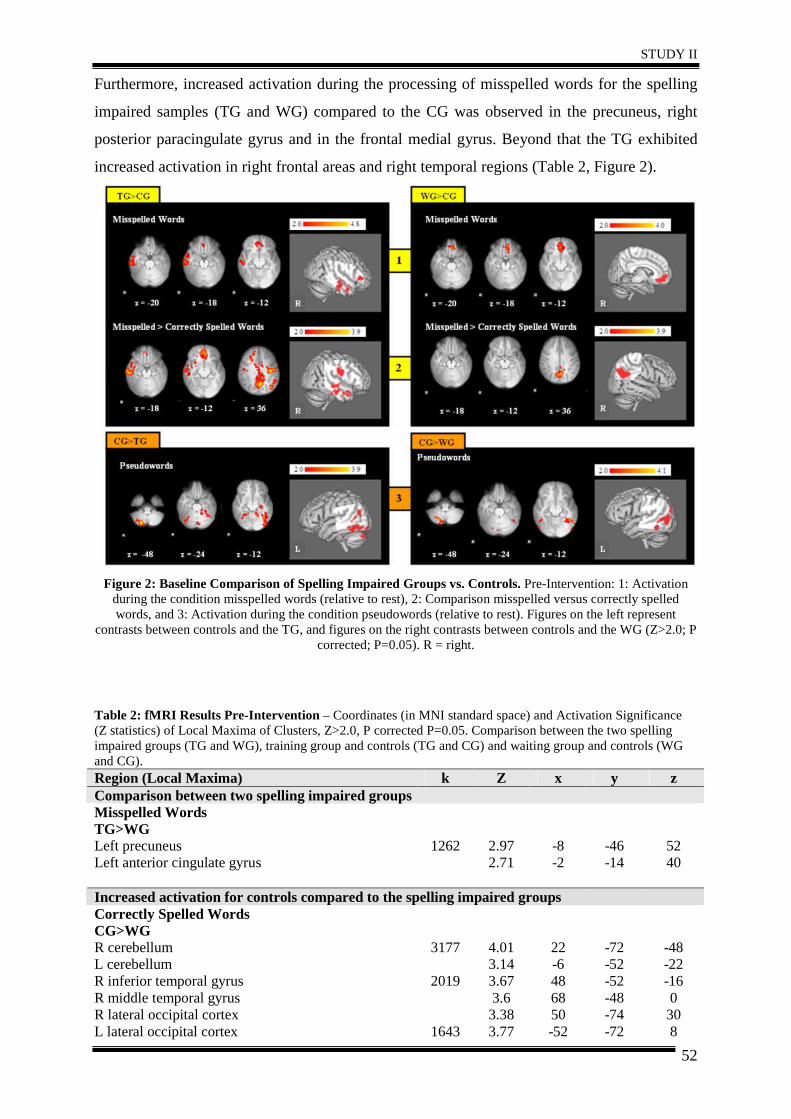
STUDY II
52
Furthermore, increased activation during the processing of misspelled words for the spelling
impaired samples (TG and WG) compared to the CG was observed in the precuneus, right
posterior paracingulate gyrus and in the frontal medial gyrus. Beyond that the TG exhibited
increased activation in right frontal areas and right temporal regions (Table 2, Figure 2).
Figure 2: Baseline Comparison of Spelling Impaired Groups vs. Controls. Pre-Intervention: 1: Activation
during the condition misspelled words (relative to rest), 2: Comparison misspelled versus correctly spelled words, and 3: Activation during the condition pseudowords (relative to rest). Figures on the left represent
contrasts between controls and the TG, and figures on the right contrasts between controls and the WG (Z>2.0; P corrected; P=0.05). R = right.
Table 2: fMRI Results Pre-Intervention – Coordinates (in MNI standard space) and Activation Significance (Z statistics) of Local Maxima of Clusters, Z>2.0, P corrected P=0.05. Comparison between the two spelling impaired groups (TG and WG), training group and controls (TG and CG) and waiting group and controls (WG and CG). Region (Local Maxima) k Z x y z Comparison between two spelling impaired groups Misspelled Words TG>WG Left precuneus 1262 2.97 -8 -46 52 Left anterior cingulate gyrus 2.71 -2 -14 40 Increased activation for controls compared to the spelling impaired groups Correctly Spelled Words CG>WG R cerebellum L cerebellum
3177 4.01 3.14
22 -6
-72 -52
-48 -22
R inferior temporal gyrus R middle temporal gyrus R lateral occipital cortex
2019 3.67 3.6 3.38
48 68 50
-52 -48 -74
-16 0 30
L lateral occipital cortex 1643 3.77 -52 -72 8
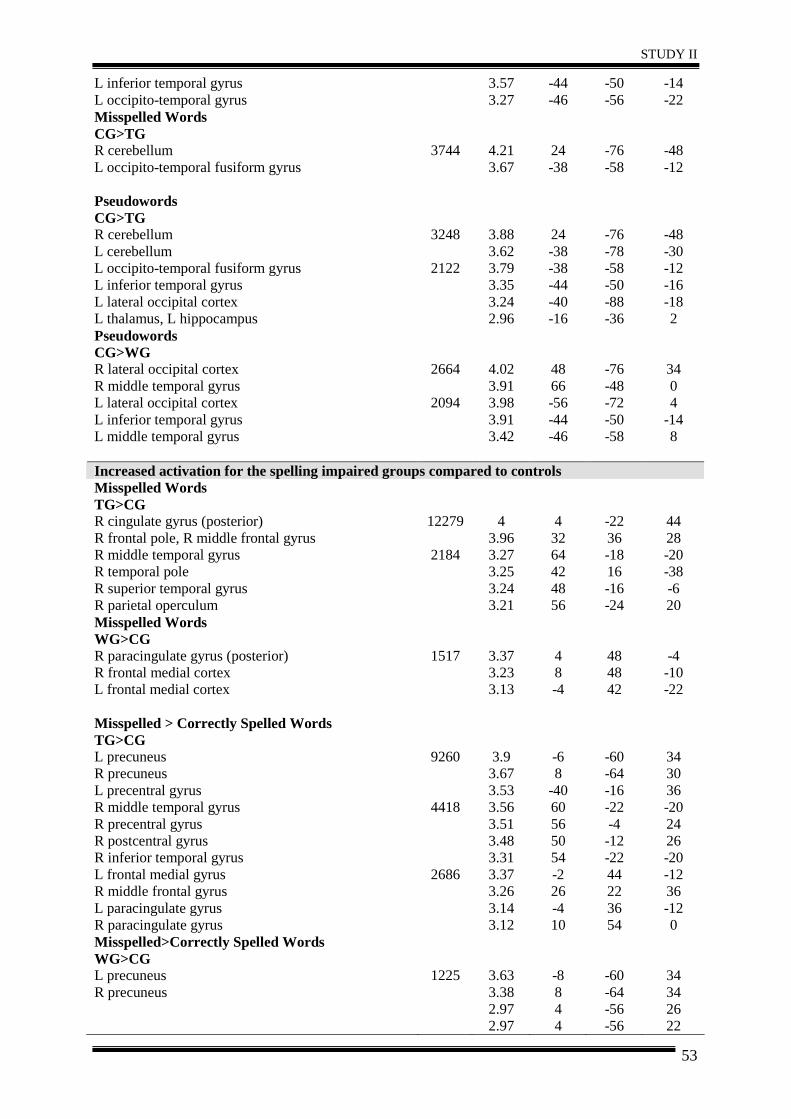
STUDY II
53
L inferior temporal gyrus L occipito-temporal gyrus
3.57 3.27
-44 -46
-50 -56
-14 -22
Misspelled Words CG>TG R cerebellum 3744 4.21 24 -76 -48 L occipito-temporal fusiform gyrus 3.67 -38 -58 -12 Pseudowords CG>TG R cerebellum L cerebellum
3248 3.88 3.62
24 -38
-76 -78
-48 -30
L occipito-temporal fusiform gyrus L inferior temporal gyrus L lateral occipital cortex L thalamus, L hippocampus
2122 3.79 3.35 3.24 2.96
-38 -44 -40 -16
-58 -50 -88 -36
-12 -16 -18 2
Pseudowords CG>WG R lateral occipital cortex 2664 4.02 48 -76 34 R middle temporal gyrus 3.91 66 -48 0 L lateral occipital cortex 2094 3.98 -56 -72 4 L inferior temporal gyrus L middle temporal gyrus
3.91 3.42
-44 -46
-50 -58
-14 8
Increased activation for the spelling impaired groups compared to controls Misspelled Words TG>CG R cingulate gyrus (posterior) R frontal pole, R middle frontal gyrus
12279 4 3.96
4 32
-22 36
44 28
R middle temporal gyrus R temporal pole R superior temporal gyrus R parietal operculum
2184 3.27 3.25 3.24 3.21
64 42 48 56
-18 16 -16 -24
-20 -38 -6 20
Misspelled Words WG>CG R paracingulate gyrus (posterior) 1517 3.37 4 48 -4 R frontal medial cortex 3.23 8 48 -10 L frontal medial cortex 3.13 -4 42 -22 Misspelled > Correctly Spelled Words TG>CG L precuneus 9260 3.9 -6 -60 34 R precuneus 3.67 8 -64 30 L precentral gyrus 3.53 -40 -16 36 R middle temporal gyrus 4418 3.56 60 -22 -20 R precentral gyrus R postcentral gyrus R inferior temporal gyrus L frontal medial gyrus R middle frontal gyrus L paracingulate gyrus R paracingulate gyrus
2686
3.51 3.48 3.31 3.37 3.26 3.14 3.12
56 50 54 -2 26 -4 10
-4 -12 -22 44 22 36 54
24 26 -20 -12 36 -12 0
Misspelled>Correctly Spelled Words WG>CG L precuneus 1225 3.63 -8 -60 34 R precuneus 3.38
2.97 2.97
8 4 4
-64 -56 -56
34 26 22
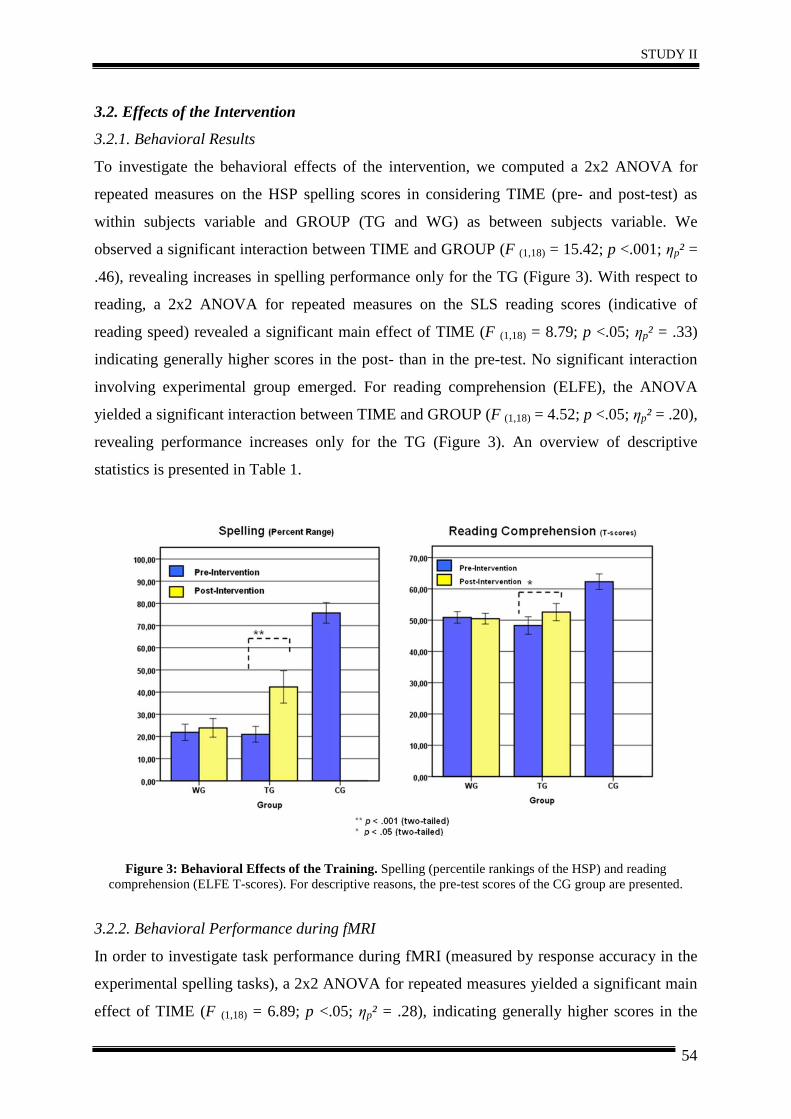
STUDY II
54
3.2. Effects of the Intervention
3.2.1. Behavioral Results
To investigate the behavioral effects of the intervention, we computed a 2x2 ANOVA for
repeated measures on the HSP spelling scores in considering TIME (pre- and post-test) as
within subjects variable and GROUP (TG and WG) as between subjects variable. We
observed a significant interaction between TIME and GROUP (F (1,18) = 15.42; p <.001; ηp² =
.46), revealing increases in spelling performance only for the TG (Figure 3). With respect to
reading, a 2x2 ANOVA for repeated measures on the SLS reading scores (indicative of
reading speed) revealed a significant main effect of TIME (F (1,18) = 8.79; p <.05; ηp² = .33)
indicating generally higher scores in the post- than in the pre-test. No significant interaction
involving experimental group emerged. For reading comprehension (ELFE), the ANOVA
yielded a significant interaction between TIME and GROUP (F (1,18) = 4.52; p <.05; ηp² = .20),
revealing performance increases only for the TG (Figure 3). An overview of descriptive
statistics is presented in Table 1.
Figure 3: Behavioral Effects of the Training. Spelling (percentile rankings of the HSP) and reading comprehension (ELFE T-scores). For descriptive reasons, the pre-test scores of the CG group are presented.
3.2.2. Behavioral Performance during fMRI
In order to investigate task performance during fMRI (measured by response accuracy in the
experimental spelling tasks), a 2x2 ANOVA for repeated measures yielded a significant main
effect of TIME (F (1,18) = 6.89; p <.05; ηp² = .28), indicating generally higher scores in the

STUDY II
55
post- than in the pre-test. The TIME by GROUP interaction failed to reach statistical
significance (p>.05), although the mean values (reported in Table 1) suggested somewhat
stronger increases in accuracy for the TG than for the WG. To investigate changes in reaction
time (RT in seconds) a 2x2 ANOVA for repeated measures was computed. The TIME by
GROUP interaction reached statistical significance (F (1,18) = 6.4; p <.05; ηp² = .26), indicating
a stronger increase in RT for the WG (see Table 1).
3.2.3. Functional MRI Results
To test for changes in brain activation patterns post- compared to pre-intervention, we
computed within group analyses for each group separately. Subsequently, to assess the
training effects more specifically, we compared increases in activation (post>pre) for the TG
and WG. The within group comparison revealed increased activation in the precuneus for all
three groups. Beyond that, for the TG increased activation in the right posterior cingulate, left
inferior and middle temporal gyrus and left hippocampus and parahippocampal region related
intervention was found. For the WG, increases in the right lateral occipital cortex and right
middle temporal cortex were observed (Table 3, Figure 4). In the CG additional increases in
activation in bilateral reading related regions at the second scan were found.
To investigate the effects of the intervention with respect to potential change of brain
activation patterns, we compared increases in activation (post>pre) for the TG and WG. We
observed a significant interaction effect, revealing increases in activation for the TG in the
bilateral parahippocampal area and in the cerebellum (extending into the brain stem), and
increased activation for the WG in the precuneus, cerebellum, left frontal pole and right
lateral occipital cortex and right parieto-temporal region (Table 3, Figure 4).
To assess the relation between improvement of spelling ability and increases in brain
activation patterns, we computed whole-brain correlation analyses. We found negative
correlations between improvement of spelling ability and activation increase in the
cerebellum and right lateral occipital cortex, right lingual gyrus and right middle temporal
gyrus in the TG (Table 3).
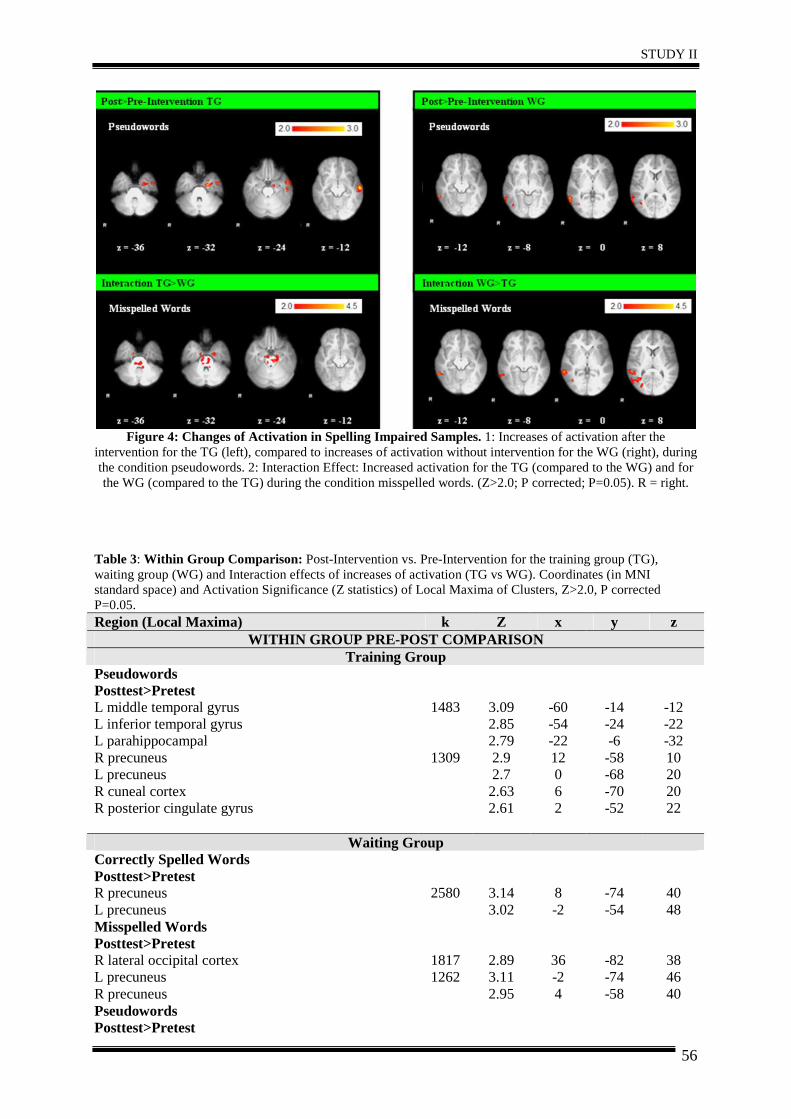
STUDY II
56
Figure 4: Changes of Activation in Spelling Impaired Samples. 1: Increases of activation after the
intervention for the TG (left), compared to increases of activation without intervention for the WG (right), during the condition pseudowords. 2: Interaction Effect: Increased activation for the TG (compared to the WG) and for the WG (compared to the TG) during the condition misspelled words. (Z>2.0; P corrected; P=0.05). R = right.
Table 3: Within Group Comparison: Post-Intervention vs. Pre-Intervention for the training group (TG), waiting group (WG) and Interaction effects of increases of activation (TG vs WG). Coordinates (in MNI standard space) and Activation Significance (Z statistics) of Local Maxima of Clusters, Z>2.0, P corrected P=0.05. Region (Local Maxima) k Z x y z
WITHIN GROUP PRE-POST COMPARISON Training Group
Pseudowords Posttest>Pretest L middle temporal gyrus L inferior temporal gyrus L parahippocampal R precuneus L precuneus R cuneal cortex R posterior cingulate gyrus
1483
1309
3.09 2.85 2.79 2.9 2.7 2.63 2.61
-60 -54 -22 12 0 6 2
-14 -24 -6 -58 -68 -70 -52
-12 -22 -32 10 20 20 22
Waiting Group Correctly Spelled Words Posttest>Pretest R precuneus L precuneus
2580
3.14 3.02
8 -2
-74 -54
40 48
Misspelled Words Posttest>Pretest R lateral occipital cortex
1817
2.89
36
-82
38
L precuneus R precuneus
1262 3.11 2.95
-2 4
-74 -58
46 40
Pseudowords Posttest>Pretest
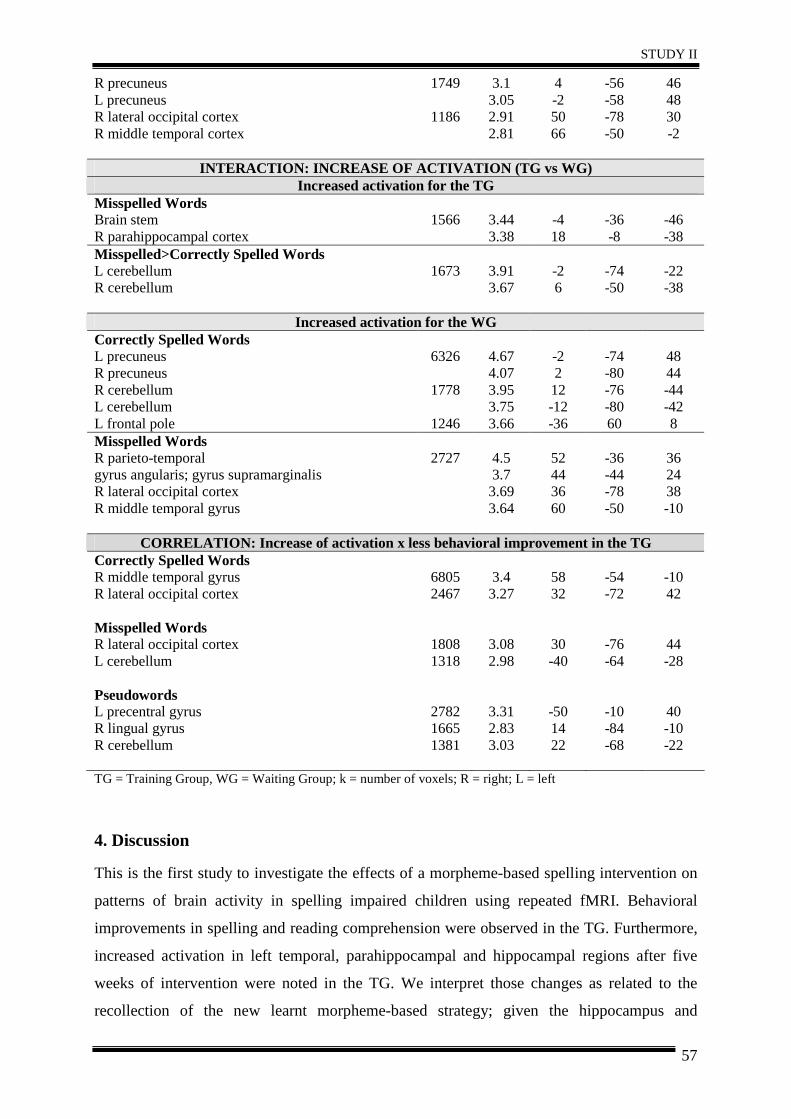
STUDY II
57
R precuneus L precuneus
1749 3.1 3.05
4 -2
-56 -58
46 48
R lateral occipital cortex R middle temporal cortex
1186 2.91 2.81
50 66
-78 -50
30 -2
INTERACTION: INCREASE OF ACTIVATION (TG vs WG) Increased activation for the TG
Misspelled Words Brain stem R parahippocampal cortex
1566 3.44 3.38
-4 18
-36 -8
-46 -38
Misspelled>Correctly Spelled Words L cerebellum R cerebellum
1673 3.91 3.67
-2 6
-74 -50
-22 -38
Increased activation for the WG Correctly Spelled Words L precuneus R precuneus R cerebellum L cerebellum L frontal pole
6326
1778
1246
4.67 4.07 3.95 3.75 3.66
-2 2 12 -12 -36
-74 -80 -76 -80 60
48 44 -44 -42 8
Misspelled Words R parieto-temporal gyrus angularis; gyrus supramarginalis R lateral occipital cortex R middle temporal gyrus
2727 4.5 3.7 3.69 3.64
52 44 36 60
-36 -44 -78 -50
36 24 38 -10
CORRELATION: Increase of activation x less behavioral improvement in the TG Correctly Spelled Words R middle temporal gyrus 6805 3.4 58 -54 -10 R lateral occipital cortex
2467 3.27 32 -72 42
Misspelled Words R lateral occipital cortex 1808 3.08 30 -76 44 L cerebellum
1318 2.98 -40 -64 -28
Pseudowords L precentral gyrus 2782 3.31 -50 -10 40 R lingual gyrus 1665 2.83 14 -84 -10 R cerebellum
1381 3.03 22 -68 -22
TG = Training Group, WG = Waiting Group; k = number of voxels; R = right; L = left
4. Discussion
This is the first study to investigate the effects of a morpheme-based spelling intervention on
patterns of brain activity in spelling impaired children using repeated fMRI. Behavioral
improvements in spelling and reading comprehension were observed in the TG. Furthermore,
increased activation in left temporal, parahippocampal and hippocampal regions after five
weeks of intervention were noted in the TG. We interpret those changes as related to the
recollection of the new learnt morpheme-based strategy; given the hippocampus and

STUDY II
58
parahippocampal gyrus have relevance for memory recollection (Diana et al., 2007; Gimbel et
al., 2011). In line with this notion, Krafnick et al. (2011) recently reported increases in gray
matter volume in the hippocampus in dyslexic children after an eight week reading
intervention. The activation increases in the left inferior and middle temporal gyri could be
indicative of enhanced reliance on concept retrieval, semantic processing and integration
processes in the TG (Binder et al., 2009).
Conversely to left hemispheric increases in the TG, the WG showed increases in right
posterior regions (i.e. lateral occipital cortex, gyrus angularis, gyrus supramarginalis). Several
studies observed a stronger engagement of the right hemisphere in dyslexic individuals,
suggestive of (probably inefficient) compensatory cognitive mechanisms (e.g. Maisog et al.,
2008; Shaywitz et al., 2006). These findings would be also in line with the negative
correlations between behavioral improvement and activation increase in the cerebellum and
right occipital and temporal regions in the TG. It seems that increased activation in the right
posterior hemisphere correlates with less improvement of spelling ability due to intervention,
which would further support the notion that reliance on the right posterior regions is probably
related to inefficient compensation.
Prior to the intervention, both spelling impaired groups showed increased activation in the
precuneus and frontal medial cortex and relatively decreased activation of left occipito-
temporal and cerebellar regions during an orthographic decision task (relative to controls).
Increased activation in the frontal medial region and paracingulate gyrus might be explained
by a more effortful and attentionally guided reading strategy (Fornito et al., 2004; Meyler et
al., 2008) used by the spelling impaired children. Increased activation of the precuneus in
children with spelling and reading impairments compared to non-impaired controls has also
been found by others (Kronbichler et al., 2008; Maisog et al., 2008; Richlan et al., 2009;
Shaywitz et al., 2004, 2006). The precuneus has been associated with attention, semantic
processing and most notably with the default-mode network (Binder et al., 2009; Cavanna et
al., 2006; Graves et al., 2010). This region is active during conscious rest and deactivated
during attentive task engagement. The general increase of activation in the precuneus found in
all groups at the second scan thus suggests a general decrease in attention or excitement
(Binder et al., 2009; Cavanna et al., 2006; Graves et al., 2010).
We also observed decreased activation of left occipito-temporal and cerebellar regions in
spelling impaired samples relative to controls prior to the intervention. The left occipito-
temporal region has been related to automatic and fluent reading (e.g. Kronbichler et al.,
2006; Shaywitz et al., 2006) and decreased activation has been found in multiple studies
investigating dyslexia or reading impaired individuals. Several structural and functional

STUDY II
59
imaging studies suggest cerebellar disruptions in individuals with dyslexia (e.g. Brown et al.,
2001; Brunswick et al., 1999; Eckert et al., 2003; Kronbichler et al., 2008; Richlan et al.,
2010). These have been related to semantic and phonological processing (Fulbright et al.,
1999), skill automatization and learning (Nicolson et al., 1999, 2001; Poldrack & Gabrieli,
2001) and linguistic performance (Riva & Giorgi, 2000; Scott et al., 2001).
Some limitations of this study also have to be considered, when interpreting our results. First,
the interpretation of the interaction effect (comparing within group changes of TG and WG)
has to be done carefully, as the two spelling impaired groups showed differences in activation
patterns prior to the intervention. However, the comparison of activation patterns post- vs.
pre-intervention for each group separately revealed increased activation in parahippocampal
regions for the TG, which were also observed by the interaction analyses. Second, a sample
size of ten children per group might be regarded as rather small. While this may certainly
compromise statistical power, it needs to be recognized that the employed study design
(requiring children to participate in the training and to take part in several behavioral and
fMRI testing sessions) imposed great efforts both on participants and the resources involved,
together making studies of this kind difficult and rare. Third, a follow-up assessment
including fMRI several months after the intervention would have been desirable to assess
potential long-term effects of training outcome.
Nonetheless, our study provides insights into the functional correlates of spelling impairment
and preliminary evidence for training-induced changes in brain function. We hope this work
encourages future investigations into this area that also seek to overcome some of these
shortcomings.
Acknowledgments: The authors wish to express their gratitude to Nadja Kozel, Bernd Schneeberger, Johanna Vogl and Stefanie Rohrer who greatly contributed to this project and to Franz Ebner, MD, for continued infrastructural support.

STUDY III
60
STUDY III – Structural Changes related to intervention
Differences in integrity of white matter and changes
with training in spelling impaired children – a
diffusion tensor imaging study
D. Gebauer 1,2, A. Fink 2, N. Filippini 3,6, H. Johansen-Berg 3, G. Reishofer 4, K. Koschutnig 4, R. Kargl 5, C. Purgstaller 5, F. Fazekas 1, C. Enzinger 1,4
(1) Department of Neurology, Medical University of Graz (2) Department of Psychology, Karl-Franzens-University Graz
(3) fMRIB centre, Nuffield Department of Clinical Neuroscience, University of Oxford (4) Division of Neuroradiology, Department of Radiology, Medical University of Graz
(5) Institute of Reading and Spelling, Graz (6) Department of Psychiatry, University of Oxford
(Submitted to Brain, Structure and Function on 15 September 2011; submitted in revised form on 24 October 2011; accepted on 4 December 2011; available online since 25 December 2011)

STUDY III
61
Abstract
While the functional correlates of spelling impairment have been rarely investigated, to our
knowledge no study exists regarding the structural characteristics of spelling impairment and
potential changes with interventions. Using diffusion tensor imaging at 3.0 T, we here
therefore sought to investigate (a) differences between children with poor spelling abilities
(Training Group and Waiting Group) and controls and (b) the effects of a morpheme-based
spelling intervention in children with poor spelling abilities on DTI parameters. A baseline
comparison of white matter indices revealed significant differences between controls and
spelling impaired children, mainly located in the right hemisphere (superior corona radiata
(SCR), posterior limb of internal capsule (PLIC), superior longitudinal fasciculus (SLF)).
After five weeks of training, spelling ability improved in the training group, along with
increases in fractional anisotropy and decreases of radial diffusivity in the right hemisphere
compared to controls. In addition, significantly higher decreases of mean diffusivity in the
right SCR for the spelling impaired training group compared to the waiting group were
observed. Our results suggest that spelling impairment is associated with differences in white
matter integrity in the right hemisphere. We also provide first indications that white matter
changes occur during successful training, but this needs to be more specifically addressed in
future research.
KEYWORDS: spelling impairment, DTI, intervention, white matter

STUDY III
62
1. Introduction Up to 16% of all children and adolescents may be affected by reading and spelling
difficulties, associated with greater risk of school anxiety, unemployment and multiple
emotional and behavioral difficulties (Klicpera et al. 2007; Schulte-Körne & Remschmidt
2003). A severe and well-known form of reading difficulties is dyslexia. Current
neuroimaging studies revealed that dyslexia is associated with lower activation in parieto-
temporal and occipito-temporal brain regions of the left hemisphere (Kronbichler et al. 2008;
Shaywitz et al. 2004), along with increased activation in frontal or right hemispheric regions
(Richlan et al. 2009; Maisog et al. 2008; Shaywitz et al. 2006). Intervention studies suggested
that these activation patterns may be “normalized” in regions associated with reading and
spelling in individuals with dyslexia or reading impairment (Aylward et al. 2003; Eden et al.
2004; Meyler et al. 2008; Richards et al., 2006; Shaywitz et al. 2004; Simos et al. 2002, 2006;
Temple et al., 2003). Previously, we observed a tendency of functional normalization of brain
activation patterns after an intense spelling intervention in children with spelling impairment.
As the neural signature of reading and spelling impairment may not only manifest
functionally but also at a microstructural level, we used diffusion tensor imaging (DTI) to
augment our understanding of neural plasticity related to intervention in this context.
Recent DTI studies reported regional white matter differences between individuals with
spelling and reading impairment and non-impaired controls in tracts related to reading,
specifically bilateral in the posterior limb of the internal capsule (e.g. Beaulieu et al. 2005;
Klingberg et al. 2000), the superior longitudinal fasciculus (e.g. Carter et al. 2009; Hoeft et
al., 2011 Rimrodt et al. 2010; Steinbrink et al. 2008), superior corona radiata (e.g. Deutsch et
al. 2005), inferior longitudinal fasciculus (e.g. Rollins et al. 2009; Steinbrink et al. 2008),
corpus callosum (Ben-Shachar et al. 2007; Dougherty et al. 2007) and anterior corona radiata
(e.g. Beaulieu et al. 2005; Niogi & McCandliss 2006).
Several studies found a correlation between white matter integrity (mostly assessed by
fractional anisotropy, FA) and reading ability in left parieto-temporal areas. Correlations
between reading ability and fibers with inferior-superior orientation in the PLIC (Beaulieu et
al. 2005) and the superior corona radiata (Deutsch et al. 2005; Niogi & McCandliss 2006)
were also found. Furthermore, correlations with fibers with anterior-posterior orientation in
left parieto-temporal regions (related to the external capsule and arcuate fasciculus; Klingberg
et al. 2000) were observed. Hoeft et al. (2011) reported that single-word reading improvement
in dyslexics over 2.5 years correlated positively with increased white matter integrity in the
right SLF and greater activation in the right IFG.

STUDY III
63
To our knowledge, so far only two DTI studies included behavioral measures of spelling.
Deutsch et al. (2005) found positive correlations between measurements of reading (r = .62)
and spelling (r = .66) with left parieto-temporal white matter in a pre-defined volume of
interest (VOI), suggesting more preserved white matter integrity in this region to be related to
improved processing efficiency. Steinbrink et al. (2008) investigated German dyslexic adults
and found decreased FA in bilateral fronto-temporal and left temporo-parietal white matter
regions (ILF and SLF), probably indicating less efficient communication in dyslexics. No
significant correlations with spelling were reported, but a significant correlation was found
between FA in a pre-defined region of interest (ROI) and speed of pseudoword reading for
dyslexic children (r = .85/.87; fronto-temporal bilateral, left parieto-temporal) and normal
controls (r = .82; left frontal). No significant correlations with mean diffusivity were
observed.
Learning new skills, be it motor or cognitive in nature, causes changes of brain function (e.g.
Poldrack et al. 1998; Raichle et al. 1994; Westerberg & Klingberg 2007) and also entails
structural changes in white and gray matter (Bengtsson et al., 2005; Draganski et al. 2004;
Scholz et al., 2009). Neuroimaging techniques provide new possibilities to investigate effects
of intervention and therapies of motor or cognitive impairments. The concept of neural
plasticity is crucial in various fields ranging from developmental language acquisition to
stroke recovery, providing new insight and hope for affected people.
Only few studies investigated changes in white matter connectivity due to instruction (e.g.
Scholz et al. 2009). To date only one study probed white matter changes due to intervention in
poor readers (Keller et al. 2009). The authors found that 100 hours of reading instruction over
six months resulted in increased FA and decreased radial diffusivity in the left anterior
centrum semiovale in poor readers, a pattern that could be indicative of increased myelination.
Furthermore, FA increase correlated with improvement in phonological decoding ability
(partial r = .23).
Functional correlates of spelling impairment have rarely been investigated (e.g. Richards et al.
2009), and to our knowledge no study about the structural characteristics of spelling
impairment and related interventions exist so far. Therefore, we here aimed to investigate the
effects of a morpheme-based spelling intervention on brain structure in children with poor
spelling abilities. We hypothesized that (a) children with poor spelling abilities show
differences in brain structure prior to the intervention compared to controls, consistent with
observations in reading impaired individuals (e.g. Beaulieu et al. 2005; Klingberg et al. 2000
Rimrodt et al. 2010; Rollins et al. 2009; Steinbrink et al. 2008). Furthermore, we investigated

STUDY III
64
whether (b) a spelling intervention would alter microstructural white matter changes as
measured by white matter indices obtained from DTI.
2. Methods 2.1. Participants
As part of a functional MRI study, which was designed to investigate the functional effects of
a spelling intervention on spelling impaired children, we also repetitively obtained DTI data
that allow addressing the central research question on microstructural changes in such a
setting. DTI data of 34 German-speaking children aged between nine and 16 years were
acquired. From this cohort, data of 6 children had to be excluded due to movement or scanner
artifacts, rendering a final sample of 28 children (16 males), whose age ranged from 10 to 16
years (M = 11.96; SD= 1.77; see Table 1). All participants were healthy, right-handed and had
normal or corrected to normal vision. The ethics committee of the Medical University Graz,
Austria approved the study. All participants and their parents gave written informed consent.
The effect of the intervention was investigated in a pre- / post-test design. We formed and
investigated three groups: (1) Ten children with below average spelling abilities were
assigned to the “Training-Group” (TG), (2) another nine children with poor spelling abilities
were assigned to the so-called “Waiting-Group” (WG, receiving the training after the post-
test), and (3) a control group (CG) of nine children with average spelling abilities, assessed at
a single time-point. Spelling ability was determined by standardized psychometric tests during
a pre-experimental screening, as specified below.
The groups did not differ significantly (p>.05) with respect to age and non-verbal intelligence,
but controls scored considerably higher in reading and spelling (p <.05; Table 1). Specific
post-hoc comparisons by means of the Tukey HSD test revealed that controls had
significantly higher test scores than both spelling impaired groups (p< .05), while the TG and
WG were not significantly different (Table 1).
STUDY III
65
Table 1: Descriptive Statistics of age, non-verbal intelligence, Reading- and Spelling Skills for Training Group (TG), Waiting Group (WG) and Control Group (CG); Means and Standard Deviations (in brackets).
TG WG CG p Sex 10 (7 males) 9 (6 males) 9 (3 males) Age (years) 11.5 (0.71) 12.1 (2.03) 12.3 (2.35) .58 Intelligence - Raven 36.7 (7.7) 38.0 (9.82) 44.3 (5.29) .10 Pre-Intervention Reading Speed Reading Comprehension
91.4 (14.27) 41.52 (24.57)
99.0 (10.77) 54.19 (21.98)
117.89 (12.71) 60.86 (7.98)
.000
.005 Spelling Skills - HSP 20.98 (11.37) 21.64 (12.34) 78.1 (15.98) .000 Post-Intervention Reading Speed Reading Comprehension
102.6 (13.88) 57.77 (27.91)
103.22 (10.51) 53.68 (20.80)
Spelling Skills - HSP 42.33 (23.03) 23.73 (14.11) Age (F(2,25) = 0.55; p = 0.58; ηp² =.04), Non-verbal intelligence: Raven Raw Scores (F(2,25) = 2.52; p = 0.10; ηp² =.17); Pre-Intervention: Reading Skills: SLS Reading Quotient (F(2,25) = 10.71; p < .001; ηp² = .46); Reading Comprehension: 6.62; p < .05; ηp² = .35). Spelling Skills: (F(2,25) = 55.71; p < .001; ηp² = .82).
2.2. Psychometric Tests
In the pre-experimental screening, participants underwent standardized tests for the
assessment of reading and spelling abilities, along with relevant socio-demographic data such
as age, sex, native language, and school year. In the “Hamburger-Schreibprobe” (HSP; May
et al. 2000), a standardized spelling test for children from the first to the ninth grade, words
and sentences are dictated by the experimenter and have to be written next to corresponding
pictures that illustrate them. This test takes about 15 minutes and we applied the version for
4th/5th graders and 5th to 9th graders. The HSP provides measures for the number of correctly
spelled words and the number of grapheme-related mistakes. The latter was used in this study
as it is more precisely. The “Salzburger-Lese-Screening” (SLS; Mayringer and Wimmer
2005) measures reading speed and basic reading ability (automaticity, accuracy). We used the
SLS 1-4 for children up to the 4th grade, and the SLS-5-8 was applied for older children,
parallel versions exist for both. In the SLS, children have to decide whether the content of a
presented sentence is correct or not. Testing time is limited to three minutes. In addition we
also measured reading comprehension (i.e. comprehension of words, sentences and text) by
means of a standardized German-speaking test (ELFE 1-6; Lenhard and Schneider 2006).
Furthermore, non-verbal intelligence was measured using the Standard Progressive Matrices
(SPM) by Raven (1960).

STUDY III
66
2.3. Intervention
The intervention was a computer-aided morpheme-based spelling training (Morpheus; Kargl
& Purgstaller 2010), which has been approved as an evidence-based intervention for
individuals with reading and spelling deficits by the federal ministry of Austria
(http://www.schulpsychologie.at/lernen-leistung/lese-rechtschreibschwaeche/).
Morpheus consists of computerized tutorials, a book of exercises and morpheme-based games
to facilitate the consolidation of the strategy. The intervention included daily handwritten and
computer homework along with instructor-guided courses (once a week, lasting
approximately two hours) over a period of five weeks. These tutorials on the computer
included 12 different playful exercises dealing with morphemes (e.g. recognizing and
matching word families, morphological clozes, finding suffixes and prefixes). During the
tutorials achieved scores were displayed on the computer screen. Participants could only
reach the next difficulty level of the same exercise when they had solved at least 75% of the
given problems correctly. The automatically saved score of every tutorial served as basis for
assessing training progress. Displaying the achieved performance increased the training
motivation of the participants.
In general, the training material of Morpheus consists of the most frequent morphemes of the
German language and contains different levels of difficulty. The words used for the training
were taken from an empirically based collection of words (German basic vocabulary for 4th
graders; Augst 1989). Morpheus has been constructed on the basis of the following principles:
simplicity, relief due to morpheme segmentation, rule-governed repetition, playfulness,
avoidance of mistakes, individuality, productivity, and practicing handwriting.
Every word is built by different parts, which follow particular spellings (e.g. unfriendly =
prefix [un], suffix [ly], root [friend]). Therefore, the spelling of the German verb “verfahren”
can be derived by two rules: the prefix [ver] is always written with [v], the root [fahr] always
with an “h”. Children do not need to remember the spelling of every single word, but only to
memorize the spelling of their component parts. Furthermore, morphosemantic information
can support the development of a meaning-oriented decoding strategy, e.g. the correct
spelling of the noun “Motor-rad” (motor-bike) can be derived by the meaning (May et al.
2000). In addition, this strategy seems to be easy to apply as only “100 of the most frequent
morphemes cover 70% of all written material” (Scheerer-Neumann, 1979).

STUDY III
67
2.4. Magnetic Resonance Imaging (MRI) data acquisition
Imaging was performed on a 3.0 Tesla Trio Tim scanner (Siemens Medical Systems,
Erlangen, Germany) using a 12-channel head coil. To minimize head movement, subjects’
heads were stabilized with foam cushions. Beside high-resolution 3D-T1 MPRAGE (1 mm
isotropic) structural scans (TR = 1900 ms, TE = 2.2 ms), single shot EPI DTI data including
four averages (1.9 x 1.9 x 2.5 mm acquisition voxel size, TR = 6700 ms, TE = 95 ms, matrix
128 x 128 mm; FOV = 250 mm, flip angle: 90°; b-value = 1000 s mm -2, 4 x B = 0 images, 12
directions) were obtained. Scan time was ten minutes for the T1-MPRAGE scan and seven
minutes for the DTI acquisition. Structural brain scans were reviewed by an MRI expert with
more than ten years of experience in neuroimaging and did not show morphological
abnormalities.
2.5. Diffusion tensor imaging (DTI) analysis
Diffusion tensor imaging (DTI) analysis was performed using FDT (fMRIB´s Diffusion
Toolbox, Version v 2.0, part of fMRIB´s Software Library) and TBSS (Tract-Based Spatial
Statistics, Version v 1.2 part of fMRIB´s Software Library).
Raw images were pre-processed using eddy current correction. A brain mask was created
using BET (Brain Extraction Tool, Version 2.1). Maps for fractional anisotropy (FA), Mean
Diffusivity (MD), axial diffusivity (λ 1, along the axis of the fiber) and radial diffusivity [(λ2
+ λ3) / 2] were generated, to increase interpretability of our findings.
Subsequently voxelwise statistical analysis of FA data was carried out using TBSS. Because
standard templates were not appropriate for the smaller head and brain size of children, we
generated a study-specific template by registering the FA images to the mean_FA of the
group (created with the FMRIB58_FA image as the target). The FA skeleton was thresholded
at 0.20 to include major white matter pathways but avoid peripheral tracts (vulnerable to
inter-subject variability). Each subject´s FA map was then projected onto the mean skeleton.
Voxel-wise cross-subject statistics (p<.05) by threshold-free cluster enhancement (TFCE),
avoiding use of an arbitrary threshold for the initial cluster-formation was applied. TFCE
represents a recently-proposed method to enhance cluster-like structures in an image (e.g. a z-
statistic activation image from an FMRI analysis) without having to define an initial cluster-
forming threshold or carry out a large amount of data smoothing (Smith & Nichols, 2009).
We used nonparametric testing as implemented in “randomise” (5000 permutations), for
calculating group contrasts. “Randomise” is a permutation method, which is used for

STUDY III
68
inference (thresholding) on statistic maps when the null distribution is not known (Nichols &
Holmes, 2002).
The anatomical location of significant clusters was determined by reference to the fibre tract-
based atlas of human WM (JHU ICBM-DTI-81 White-Matter Labels, JHU White-Matter
Tractography Atlas, Juelich Histological Atlas), implemented in FSL. MD, radial and axial
diffusivity were compared using TBSS in an analogous fashion.
Based on theoretical background we defined ten regions of interest (ROI´s), including the
right and left posterior limb of internal capsule (e.g. Beaulieu et al. 2005; Klingberg et al.
2000), superior corona radiata (e.g. Beaulieu et al. 2005; Klingberg et al. 2000), superior
longitudinal fasciculus (e.g. Carter et al. 2009; Hoeft et al. 2011; Steinbrink et al. 2008),
inferior longitudinal and occipito-frontal fasciculus (Rollins et al. 2009; Steinbrink et al.
2008) and anterior corona radiata (e.g. Beaulieu et al. 2005; Niogi & McCandliss 2006).
3. Results The two spelling impaired groups (training group – TG, waiting group – WG) did not
significantly differ with respect to any behavioral measure. Participants of the control group
(CG) scored significantly higher than both spelling impaired groups in reading and spelling
abilities (see further section 2.1. and Table 1). Figure 1 represents a schematic overview of the
statistical comparisons done regarding the significance of results.
Figure 1: Schematic overview of statistical comparisons and significance of results. Black arrows indicate significant differences and dashed lines indicate a tendency of difference. ROI = Results of Region of Interest
Analyses. TG = Training Group; WG = Waiting Group and CG = Controls.
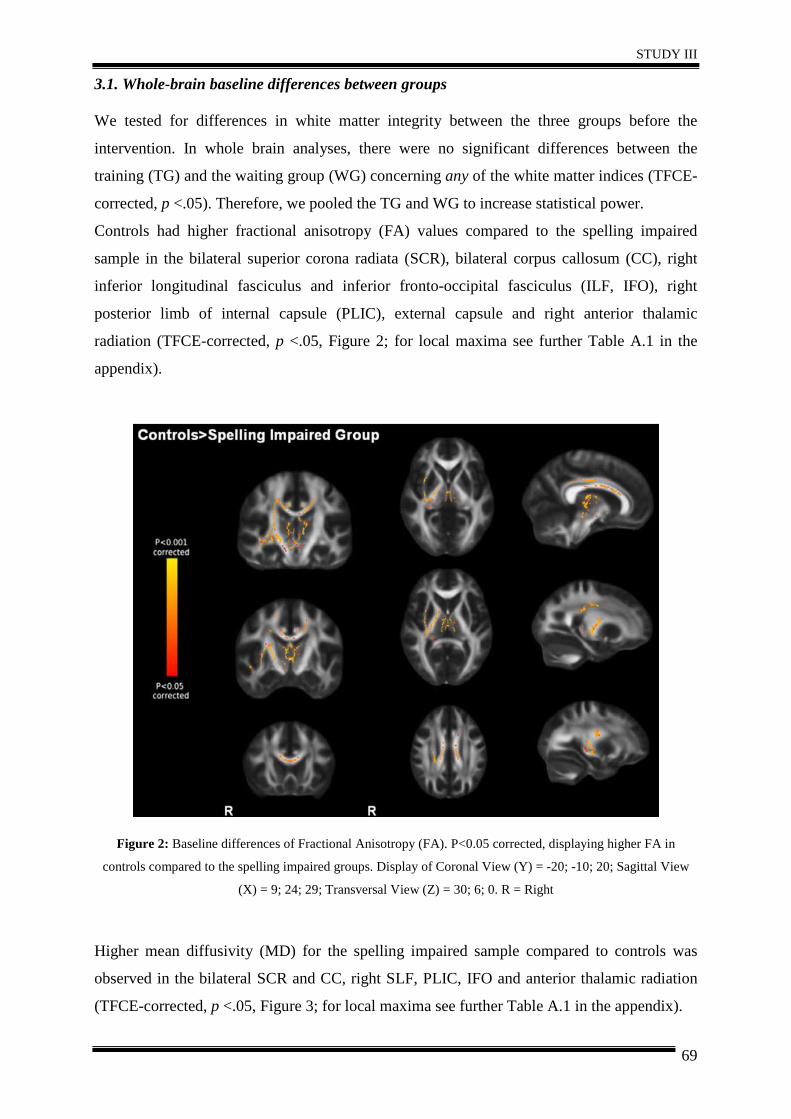
STUDY III
69
3.1. Whole-brain baseline differences between groups
We tested for differences in white matter integrity between the three groups before the
intervention. In whole brain analyses, there were no significant differences between the
training (TG) and the waiting group (WG) concerning any of the white matter indices (TFCE-
corrected, p <.05). Therefore, we pooled the TG and WG to increase statistical power.
Controls had higher fractional anisotropy (FA) values compared to the spelling impaired
sample in the bilateral superior corona radiata (SCR), bilateral corpus callosum (CC), right
inferior longitudinal fasciculus and inferior fronto-occipital fasciculus (ILF, IFO), right
posterior limb of internal capsule (PLIC), external capsule and right anterior thalamic
radiation (TFCE-corrected, p <.05, Figure 2; for local maxima see further Table A.1 in the
appendix).
Figure 2: Baseline differences of Fractional Anisotropy (FA). P<0.05 corrected, displaying higher FA in
controls compared to the spelling impaired groups. Display of Coronal View (Y) = -20; -10; 20; Sagittal View
(X) = 9; 24; 29; Transversal View (Z) = 30; 6; 0. R = Right
Higher mean diffusivity (MD) for the spelling impaired sample compared to controls was
observed in the bilateral SCR and CC, right SLF, PLIC, IFO and anterior thalamic radiation
(TFCE-corrected, p <.05, Figure 3; for local maxima see further Table A.1 in the appendix).
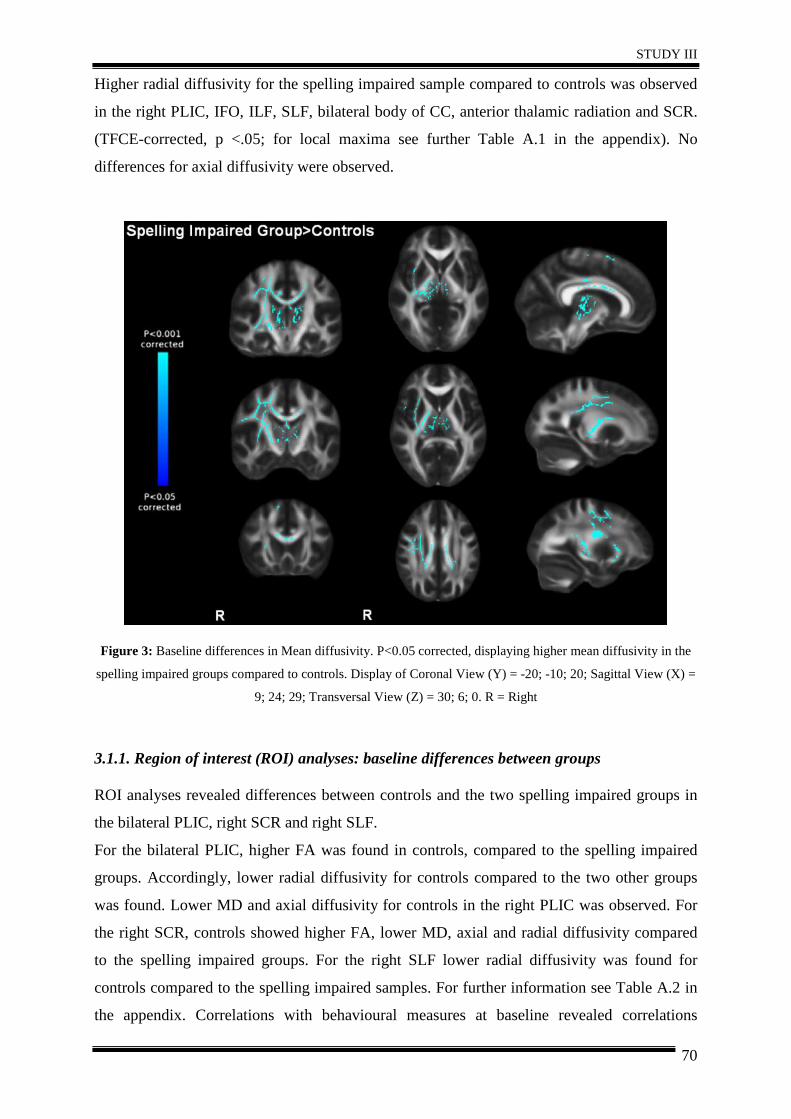
STUDY III
70
Higher radial diffusivity for the spelling impaired sample compared to controls was observed
in the right PLIC, IFO, ILF, SLF, bilateral body of CC, anterior thalamic radiation and SCR.
(TFCE-corrected, p <.05; for local maxima see further Table A.1 in the appendix). No
differences for axial diffusivity were observed.
Figure 3: Baseline differences in Mean diffusivity. P<0.05 corrected, displaying higher mean diffusivity in the
spelling impaired groups compared to controls. Display of Coronal View (Y) = -20; -10; 20; Sagittal View (X) =
9; 24; 29; Transversal View (Z) = 30; 6; 0. R = Right
3.1.1. Region of interest (ROI) analyses: baseline differences between groups
ROI analyses revealed differences between controls and the two spelling impaired groups in
the bilateral PLIC, right SCR and right SLF.
For the bilateral PLIC, higher FA was found in controls, compared to the spelling impaired
groups. Accordingly, lower radial diffusivity for controls compared to the two other groups
was found. Lower MD and axial diffusivity for controls in the right PLIC was observed. For
the right SCR, controls showed higher FA, lower MD, axial and radial diffusivity compared
to the spelling impaired groups. For the right SLF lower radial diffusivity was found for
controls compared to the spelling impaired samples. For further information see Table A.2 in
the appendix. Correlations with behavioural measures at baseline revealed correlations
STUDY III
71
between spelling skills and white matter indices in the right PLIC, SCR and left ILF/IFO for
the entire sample (see Table 2).
Table 2: Baseline Correlations: Significant Correlations between white matter indices and Spelling Skills for the Entire Sample. Pearson correlation coefficients are presented, ranging from -1 to +1 (positive values indicate positive correlations). FA MD Axial diffusivity Radial
diffusivity Right PLIC .38 -.58 -.45 -.50 Right SCR -.40 -.40 Left IFO/ILF .43 Explanation of Abbreviations: PLIC = posterior limb of internal capsule; SCR = superior corona radiata; IFO = Inferior occipito-frontal fasciculus; ILF = inferior longitudinal fasciculus; SLF = superior longitudinal fasciculus; ACR = anterior corona radiata
3.2. Training associated changes 3.2.1. Behavioral Results – Comparison of training group (TG) and waiting group (WG)
In order to investigate behavioral effects of the intervention, we computed a 2x2 ANOVA for
repeated measures on the HSP spelling scores considering TIME (pre- and post-test) as within
subjects’ variable and GROUP (TG and WG) as between subjects’ variable. We observed a
significant main effect of TIME (F (1,17) = 17.78; p <.05; ηp² = .50), indicating higher scores in
the post- than in the pre-test, but as it was evident by a significant interaction between TIME
and GROUP (F (1,17) = 11.33; p <.05; ηp² = .40), only the TG showed performance increases in
spelling (see Figure 4). The mean spelling score of the TG (42.33) after intervention reached
the range of average spelling ability (between 40 and 60; see Table 1).
With respect to reading, a 2x2 ANOVA for repeated measures on the SLS reading scores
(indicative of reading speed) revealed a significant main effect of TIME (F (1,17) = 11.31; p
<.05; ηp² = .40) indicating generally higher scores in the post- than in the pre-test. No
significant interaction with experimental group emerged. Also, for reading comprehension
(ELFE PR), the ANOVA yielded a significant main effect of TIME (F (1,17) = 4.56; p <.05; ηp²
= .21), indicating generally higher scores in the post- than in the pre-test (see Table 1).
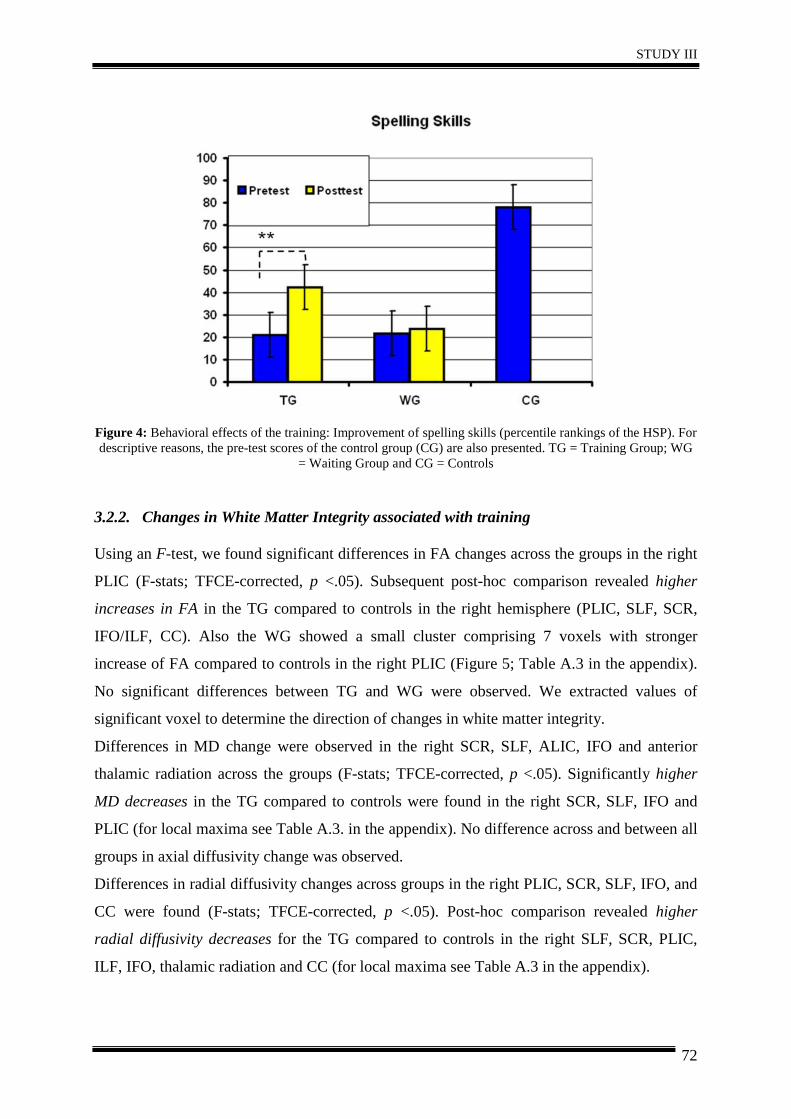
STUDY III
72
Figure 4: Behavioral effects of the training: Improvement of spelling skills (percentile rankings of the HSP). For descriptive reasons, the pre-test scores of the control group (CG) are also presented. TG = Training Group; WG
= Waiting Group and CG = Controls
3.2.2. Changes in White Matter Integrity associated with training
Using an F-test, we found significant differences in FA changes across the groups in the right
PLIC (F-stats; TFCE-corrected, p <.05). Subsequent post-hoc comparison revealed higher
increases in FA in the TG compared to controls in the right hemisphere (PLIC, SLF, SCR,
IFO/ILF, CC). Also the WG showed a small cluster comprising 7 voxels with stronger
increase of FA compared to controls in the right PLIC (Figure 5; Table A.3 in the appendix).
No significant differences between TG and WG were observed. We extracted values of
significant voxel to determine the direction of changes in white matter integrity.
Differences in MD change were observed in the right SCR, SLF, ALIC, IFO and anterior
thalamic radiation across the groups (F-stats; TFCE-corrected, p <.05). Significantly higher
MD decreases in the TG compared to controls were found in the right SCR, SLF, IFO and
PLIC (for local maxima see Table A.3. in the appendix). No difference across and between all
groups in axial diffusivity change was observed.
Differences in radial diffusivity changes across groups in the right PLIC, SCR, SLF, IFO, and
CC were found (F-stats; TFCE-corrected, p <.05). Post-hoc comparison revealed higher
radial diffusivity decreases for the TG compared to controls in the right SLF, SCR, PLIC,
ILF, IFO, thalamic radiation and CC (for local maxima see Table A.3 in the appendix).
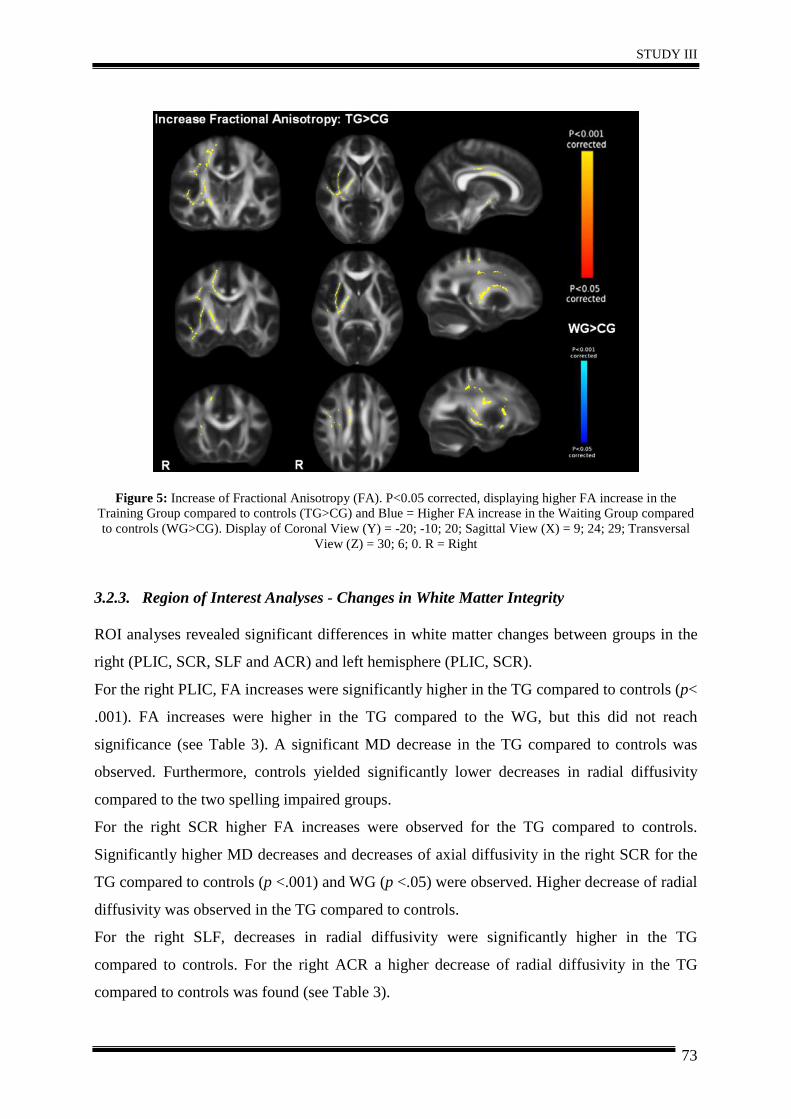
STUDY III
73
Figure 5: Increase of Fractional Anisotropy (FA). P<0.05 corrected, displaying higher FA increase in the Training Group compared to controls (TG>CG) and Blue = Higher FA increase in the Waiting Group compared to controls (WG>CG). Display of Coronal View (Y) = -20; -10; 20; Sagittal View (X) = 9; 24; 29; Transversal
View (Z) = 30; 6; 0. R = Right
3.2.3. Region of Interest Analyses - Changes in White Matter Integrity
ROI analyses revealed significant differences in white matter changes between groups in the
right (PLIC, SCR, SLF and ACR) and left hemisphere (PLIC, SCR).
For the right PLIC, FA increases were significantly higher in the TG compared to controls (p<
.001). FA increases were higher in the TG compared to the WG, but this did not reach
significance (see Table 3). A significant MD decrease in the TG compared to controls was
observed. Furthermore, controls yielded significantly lower decreases in radial diffusivity
compared to the two spelling impaired groups.
For the right SCR higher FA increases were observed for the TG compared to controls.
Significantly higher MD decreases and decreases of axial diffusivity in the right SCR for the
TG compared to controls (p <.001) and WG (p <.05) were observed. Higher decrease of radial
diffusivity was observed in the TG compared to controls.
For the right SLF, decreases in radial diffusivity were significantly higher in the TG
compared to controls. For the right ACR a higher decrease of radial diffusivity in the TG
compared to controls was found (see Table 3).
STUDY III
74
FA changes in the left PLIC were greater in the TG compared to the two other groups.
Furthermore, significantly higher decrease of radial diffusivity in the TG compared to
controls was found. Radial diffusivity in the left SCR was significantly decreased in the TG
compared to the two other groups (see Table 3).
Table 3: Region of interest Analyses. Mean changes of FA, MD, axial and radial diffusivity indices for the Training Group (TG), Waiting Group (WG) and controls (CG). TG WG CG p Right Hemisphere Right PLIC FA MD Radial D. (mm2.s-1 )
.0299 (.0171)
-.00004 (.00001) -.00005 (.00002)
.0265 (.0136)
-.00003 (.00002) -.00003 (.00002)
.0030 (.0095)
-.00001 (.00002) -.00001 (.00002)
.001 .006 .000
Right SCR FA MD Axial D. (mm2.s-1 ) Radial D. (mm2.s-1 )
.0120 (.0123)
-.000024 (.000012) -.000025 (.000012) -.000024 (.000013)
.0095 (.0117)
-.000011 (.000011) -.000010 (.000015) -.000012 (.000012)
-.0024 (.0050)
-.000002 (.000007) -.000007 (.000017) -.000001 (.000006)
.01 .001 .01 .001
Right SLF Radial D. (mm2.s-1 )
-.00003 (.00001)
-.00001 (.00002)
-.00001 (.00001)
.03
Right ACR FA
.0021 (.0072)
.0007 (.0112)
-.0075 (.0022)
.03
Left Hemisphere Left PLIC FA Radial D. (mm2.s-1 )
.0189 (.0104)
-.000022 (.00002)
.0027 (.0142)
-.000003 (.00002)
-.0030 (.0124)
-.000002 (.00001)
.002 .02
Left SCR Radial D. (mm2.s-1 )
-.000006 (.000002)
-.000002 (.000009)
.0000003 (.000005)
.01
R Plic (FA: F(2,25) = 10.23; p < .001; η² =.45; MD: F(2,25) = 6.39; p < .05; η² =.34; Radial D.: F(2,25) = 12.04; p < 001; η² =.49) R SCR (FA: F(2,25) = 5.20; p < .05; η² =.30; MD: F(2,25) = 9.6; p < .001; η² =.43; Axial D.: F(2,25) = 5.62; p < .05; η² =.31; Radial D.: F(2,25) = 10.27; p < .001; η² =.45) R SLF (Radial D.: F(2,25) = 3.71; p < .05; η² =.23) RACR (FA: F(2,25) = 4.16; p < .05; η² =.25) L Plic (FA: F(2,25) = 8.08; p < .05; η² =.39; Radial D.: F(2,25) = 4.38; p < .05; η² =.26) L SCR (Radial D.: F(2,25) = 5.11; p < .05; η² =.29) Explanation of Abbreviations: D. = diffusivity; PLIC = posterior limb of internal capsule; SCR = superior corona radiata; SLF = superior longidutinal fasciculus; ACR = anterior corona radiata Axial diffusivity and radial diffusivity values are expressed in mm2.s-1
4. Discussion Studies investigating neural correlates of spelling impairment are sparse (Richards et al.
2009). We here first report findings of differences in white matter integrity in spelling
impaired children, as assessed by DTI. Remarkably, differences were primarily observed in

STUDY III
75
the right hemisphere (superior corona radiata (SCR), posterior limb of internal capsule
(PLIC), superior longitudinal fasciculus(SLF)), as opposed to the DTI studies investigating
reading impaired samples that rather reported left hemispheric differences. On the functional
level, increased right hemispheric activation in subjects with reading and spelling impairment
have been interpreted as (probably inefficient) compensatory cognitive mechanisms, e.g.
internal articulation leading to phonological correct but orthographic incorrect spellings (e.g.
Hoeft et al. 2011; Maisog et al. 2008; Shaywitz et al. 2006).
We found increased fractional anisotropy (FA) in controls compared to both spelling impaired
groups in the superior corona radiata (SCR). FA reflects the degree of diffusion anisotropy
within a voxel, determined by fiber diameter and density, degree of myelination (Basser
1995), extracellular diffusion, inter-axonal spacing (Sen & Basser, 2005) and intravoxel fiber-
tract coherence (Basser & Pierpaoli 1996). One out of many possible explanations could
therefore be that increased FA might reflect more efficient axonal signal conduction (e.g.
Basser 1995; Ben-Shachar et al., 2007).
Differences in white matter integrity in inferior-superior oriented fibers in the corona radiata
and the PLIC have been found by prior studies investigating reading impairment (e.g.
Beaulieu et al. 2005; Deutsch et al. 2005; Klingberg et al. 2000; Nagy et al., 2004; Niogi &
McCandliss 2006). The corona radiata contains pathways devoted primarily to motor and
somatosensory function. Therefore, a relation between white matter integrity in this region
and reading and spelling skills is rather surprising and difficult to interpret. Cerebellar
deficits, reflections of differences in the corpus callosum (CC) or interdigitating pathways,
such as the superior longitudinal fasciculus (SLF) have been suggested as possible
explanations (Ben-Shachar et al. 2007).
The SLF connects the parieto-temporal cortex with the lateral frontal cortex and has been
related to articulatory and phonological processing in language (Makris et al., 2005;
Maldonado et al., 2011). The parieto-temporal cortex is known to be strongly related to the
phoneme-grapheme conversion (Booth et al., 2002, 2004; Eden et al. 2004; Shaywitz et al.
2006). The lateral frontal cortex (e.g. inferior frontal gyrus) has been related to sublexical
phonology-related processing (internal articulation) and lexico-semantic control and retrieval
processes (Heim et al., 2009; Richlan et al. 2009; Maisog et al. 2008; Shaywitz et al. 2006).
We observed higher mean diffusivity (MD) and radial diffusivity in the right hemisphere
(SLF, PLIC, SCR, IFO, inferior longitudinal fasciculus (ILF) and bilateral CC) for both
spelling impaired samples. Mean diffusivity (MD) is the mean of the eigenvalues of the
diffusion tensor, invariant with respect to orientation of the diffusion tensor (Ben-Shachar et

STUDY III
76
al. 2007). This suggests less efficient structural connectivity of right hemispheric pathways in
spelling impaired children.
Correlations between spelling ability and white matter indices were found in the right PLIC,
SCR and left inferior fronto-occipital fasciculus. A positive correlation between FA and
spelling skills in the right PLIC, along with negative correlations to MD, axial and radial
diffusivity and negative correlations between MD and axial diffusivity in the right SCR could
possibly indicate more efficient white matter connectivity in the right hemisphere to be
associated with better spelling skills. Axial diffusivity (λ1, along the axis of the fiber) has
been shown to change with changes in fiber coherence and radial diffusivity [(λ2 + λ3)/ 2]
and therefore has also sometimes been suggested to be related to factors such as fiber integrity
and myelination.
It is important to note that the observed differences in white matter indices at baseline in
children with spelling impairment may be one possible cause of spelling difficulties. Also,
poorer spelling skills could result in less efficient cerebral connectivity. Clearly, further
studies are needed to understand the relationship between white matter integrity and spelling
skills.
We further investigated if five weeks of morpheme-based spelling intervention in children
with poor spelling abilities would alter white matter microstructure. We indeed found first
indications of changes in white matter related to successful intervention. Spelling ability
improved in the TG, along with increases of fractional anisotropy and decreases of mean
diffusivity and radial diffusivity in the right hemisphere compared to controls, alluding to
increased connectivity after the intervention. The comparison between controls and WG
revealed just one significant area of greater FA change, including only 7 voxels (see Figure
5). The comparison of the spelling impaired TG and WG demonstrated significantly greater
decrease of mean diffusivity in the right SCR in the TG, consistent with increasing structural
connectivity in the group receiving the training. Descriptive data (see Table 3) suggested
more widespread structural changes in the TG compared to the WG. However, probably due
to the small sample size of nine to ten children per group and therefore reduced statistical
power, these differences often did not reach significance. Further studies investigating larger
samples and/or longer interventions would be needed to further clarify effects of spelling
intervention on white matter changes.
Increased FA and decreased radial diffusivity in the left anterior centrum semiovale in poor
readers after intervention over six months have been observed (Keller et al. 2009). As
mentioned earlier, differences between spelling impaired samples and controls were primarily

STUDY III
77
observed in the right hemisphere. We found increases of FA and decreases of MD and radial
diffusivity in right hemispheric pathways due to spelling intervention. Our results suggest that
spelling impairment and improvement are related to white matter integrity in the right
hemisphere. Positive correlations between stronger white matter integrity in the right SLF and
greater activation in the right IFG and single-word reading improvement have also been found
in a recent study that investigated children with dyslexia (Hoeft et al. 2011).
Some limitations of our study have to be considered, when interpreting our results. First, the
sample size of ten children per group might be regarded as rather small, resulting in reduced
statistical power. However, the employed study design (requiring children to participate in the
training and to take part in several behavioral testing sessions and repeated MRI) imposed
great efforts both on participants and the resources involved. We albeit hope our work might
encourage future investigators to specifically investigate the effects of a spelling intervention
on white matter integrity in larger spelling impaired samples. Further, biological
interpretation of DTI parameters is challenging. We found evidence that spelling impairment
was associated with lower FA and higher MD and radial diffusivity at baseline and that a
spelling intervention resulted in increasing FA and reducing MD and radial diffusivity. Our
results are therefore broadly consistent with the straight-forward but probably somewhat
simplistic interpretation that higher FA (lower MD, radial diffusivity) is generally indicative
of more preserved (“better”) white matter microstructure, as such values reflect greater fiber
density and coherence and greater myelination which together could support improved
neuronal efficiency. However, other microstructural changes, such as increasing axon
diameter, or changes in crossing fiber populations, could alternatively affect DTI parameters
in the opposite direction. Given this complexity, biological interpretations of DTI parameters
have to remain speculative. In addition, our acquisition scheme was simple (including only 12
diffusion encoding directions) and so only a simple tensor model could be fitted to the data.
Inclusion of more encoding directions could allow for greater accuracy of model parameter
estimation and greater potential for complex model fitting. It is hoped that improvements in
diffusion acquisition and modeling, as well as collection of multi-modal datasets, may allow
for more accurate biological conclusions to be made in future studies.

STUDY III
78
5. Conclusion
In our sample, children with spelling impairment exhibited differences in white matter
integrity mainly in the right hemisphere (SCR, PLIC, SLF). After five weeks of spelling
intervention, spelling ability improved in the training group, along with evidence for altered
integrity of white matter tracts in the right hemisphere compared to controls. We here thus
provide first indications of changes in white matter due to successful intervention in such a
cohort, which might stimulate future investigations into the effects of a spelling intervention
in larger samples.
Acknowledgments: The research presented in this paper was supported by grants from the Styrian government (Nr. A27214001062) and the Jubilee Fund of the Austrian National Bank (Nr. A26E16020013). The authors wish to express their large gratitude to Gwen Douaud, Nadja Kozel, Bernd Schneeberger, Johanna Vogl and Stefanie Rohrer who greatly contributed to this research project. We also thank Karin Brodtrager for technical assistance in the acquisition of the scans and Franz Ebner, MD, for continued infrastructural support.

GENERAL DISCUSSION AND CONCLUSIONS
79
IV GENERAL DISCUSSION AND CONCLUSIONS
This doctoral project comprised three different studies. In the first presented study
we specifically investigated isolated spelling impairment by means of functional MRI and
DTI. In our second study, we examined changes related to successful intervention with
respect to functional patterns of brain activity. In our third study, we explored whether
successful intervention would be also associated with changes in white matter integrity.
Our main findings can be summarized as follows:
1) Structural and functional differences between spelling impaired children and controls
rather occured in the right than in the left hemisphere.
2) In ROI analyses decreased activations in the left occipito-temporal region were
observed in children with SRI and children with isolated spelling impairment.
3) Successful intervention correlated with increased activation in the left temporal and
(para)hippocampal regions.
4) Successful intervention was associated with right-hemispheric changes in white matter
integrity.
In the following we will discuss each of the main findings in the context of recent research
findings in the field:
1) Functional MRI studies suggest increased right hemispheric brain activation to be
associated with spelling impairment in children. We observed increased activation in right
posterior (SMG and SPL) and bilateral frontal regions, probably reflecting a highly efficient
serial grapheme-phoneme decoding compensation strategy and increased demands on
attention and working memory. Due to the asymmetry of German language (grapheme-
phoneme correspondence is high, but phoneme-grapheme correspondence is low), this
strategy could successfully compensate reading difficulties, but not spelling difficulties. For
instance, the german words “Wal” (whale) and “Wahl” (election) are pronounced equally.
Therefore, reading aloud the words correctly will be possible following the rules of high
grapheme-phoneme correspondence (pronouncing one letter after the other). In contrast, the
correct spelling of the words cannot be derived following the phoneme-grapheme
correspondence. As mentioned in the introduction, in German-speaking samples spellings are
frequently phonologically adequate, but orthographically incorrect due to access problems to
the orthographic lexicon (Landerl & Wimmer, 2008; Wimmer & Schurz, 2010). Probably,

GENERAL DISCUSSION AND CONCLUSIONS
80
subjects with SI manage the spelling of a word through mentally pronouncing a word
(associated with increased demands on working memory and attention), leading to
orthographic spelling mistakes. Clearly, further studies are needed to clarify the cognitive
sub-processes and relation to specific brain regions affected in spelling impaired children.
Although, the left hemisphere is crucial for language, reading and spelling ability,
studies highlighting the contribution of right hemispheric regions in this context increasingly
emerge. Cognitive and linguistic functions are likely to be represented in distributed neural
networks often encompassing more than one lobe (Miceli, 2001). Enhanced right hemispheric
activation in subjects with reading and spelling disorder has been related to compensatory
mechanisms (Shaywitz et al., 2006). It is assumed that due to impairment in the common left
hemispheric reading-related network, homologue regions in the right hemisphere are used to
compensate for these deficits. A recent study reports positive correlations between right
hemispheric activation in the IFG, dorsolateral prefontral cortex and medial temporal gyrus
and performance in reading comprehension and word generation tasks (Van Ettinger-Veenstra
et al., 2011) in healthy subjects. The authors noted that the right hemisphere contributes to
modulation of language ability, as the right IFG is related to integration of semantic
knowledge and context interpretation (Caplan & Dapretto, 2001) and mediation of retrieval
processes (Fletcher et al., 1998), and activation of the right medial temporal cortex is
associated with word generation (Brown et al., 2005). These studies further support our
observation that spelling impairment is associated with increased activation in a right
hemispheric compensatory network, related to enhanced attentional demands, word
generation processes and increased involvement of working-memory processes.
Several left hemispheric regions (e.g. left angular gyrus, left posterior inferior
temporal lobe, left IFG; Cloutman et al., 2009; Hillis et al., 2002, 2004; Rapcsak & Beeson,
2004) are associated with spelling ability in normally developing individuals. However, in
line with our findings, Richards et al. (2009) found an increased activation of the precuneus,
bilateral frontal regions, left angular gyrus and right temporal regions in children with
spelling impairment, probably related to inefficient access to orthographic representations and
increased mental effort compared to good spellers. In line with this, we assume that the
enhanced right hemispheric engagement in children with SI is probably reflecting a
compensation of impairments in the left-hemispheric language-related network.
2) In accordance with recent research findings, we observed decreased activation in
the left occipito-temporal region in children with spelling and reading impairment. As
expected, we did not observe decreased left occipito-temporal activation in children with

GENERAL DISCUSSION AND CONCLUSIONS
81
isolated spelling impairment in our whole-brain analyses, but surprisingly compared to non-
impaired controls also children with isolated SI showed a decreased left occipito-temporal
activation in ROI analyses. The differences were less pronounced between controls (mean
activation left occipito-temporal = 0.53) and children with isolated SI (0.20), as opposed to
children with SRI (0.01; see further study I, Figure 4). However, an interpretation of this
finding remains challenging, as the decreased activation in the left occipito-temporal region is
the most robust characteristic of reading impairment and would not be expected in children
with isolated spelling impairment. The left occipito-temporal region (comprising the visual
word form area) is crucial for reading processes (Dehaene & Cohen, 2011). Decreased
activation in this region is associated with impairments of automatic, fluent reading (e.g.
Kronbichler et al., 2006; Richlan et al., 2009, 2011). Our study suggests that children with
spelling impairment probably compensate for initially existent reading impairment via an
increased recruitment of right posterior areas, which might indicate an efficient grapheme-
phoneme decoding strategy. In line with this suggestion, Hoeft et al. (2011) found that
dyslexic children who relied on right hemispheric pathways showed gains in reading. This
further supports our theory that the stronger right hemispheric engagement of spelling
impaired children may reflect a compensatory mechanism, related to adjustment of reading
skills.
An alternative explanation could be that the left occipito-temporal region is also
important for spelling. Purcell et al. (2011) found that a left hemispheric region just lateral
and superior to the VWFA may play a significant role in typed spelling. Also, Rapp and
colleagues (Rapp & Lipka, 2011; Tsapkini & Rapp, 2010) suggest that the left mid-fusiform
region may be related to spelling ability. Hence the decreased occipito-temporal activation
found in our children with isolated spelling impairment may probably reflect impairments of
spelling ability. Further studies could provide additional information, using a more precise
investigation of different substructures of the occipito-temporal lobe involved in reading and
spelling.
In the next section we will focus on the intervention-related functional and structural changes
of brain characteristics, which we have observed in our studies:
3) Behavioral improvements in the training group after five weeks of spelling
intervention were associated with increased activation in left temporal, parahippocampal and
hippocampal regions. We interpret these changes as being related to the recollection of the
new learnt morpheme-based strategy. Conversely, negative correlations between behavioral
improvement and right occipital and temporal brain activation were observed. These findings

GENERAL DISCUSSION AND CONCLUSIONS
82
support the notion that reliance on right posterior regions is probably related to inefficient
compensation of spelling difficulties.
In support with our functional findings, abnormalities in gray matter volume in the
hippocampus and parahippocampal area have been found in structural studies (Casanova et
al., 2005) in dyslexic subjects. In addition, increases in gray matter volume in the
hippocampus related to successful intervention in dyslexic children were observed (Krafnick
et al., 2011). The hippocampus is involved in retrieval processes (Gimbel & Brewer, 2011).
Probably increased activation (increased gray matter volume) is associated with improved
access to the orthographic lexicon.
Relating our results to prior studies investigating the neurophysiological effects of
spelling intervention is challenging, as the majority of findings investigating changes in brain
activation patterns as a result of spelling interventions come from English-speaking, reading
impaired samples (e.g. Eden et al., 2004; Meyler et al., 2008; Shaywitz et al., 2004). Only few
studies focused on subjects with spelling impairment (Richards et al., 2006; 2009).
Furthermore, some of these studies suffer from methodological limitations, as they, for
instance, did not examine changes in brain activation within the training group (only
presenting the contrast controls versus training group before and after intervention) or
interaction effects (e.g. Aylward et al., 2003; Meyler et al., 2008; Richards et al., 2006).
Hence, a reduction of group differences after the intervention could be due to increased
activation in children with SRI or decreased activation in controls. Richards et al. (2006)
found increased activation due to an orthographic intervention in the right frontal gyrus and
right posterior parietal gyrus which was accompanied by behavioral improvements,
approaching activation levels of a non-impaired control group. Despite behavioral
improvements, no changes in functional patterns of brain activity were observed after a three-
week morpheme-based intervention (Richards et al., 2006). In general, neurophysiologic
changes in terms of increased activation of the left hemispheric reading networks or right
hemispheric “compensatory” networks were observed in reading intervention studies (e.g.
Eden et al., 2004; Shaywitz et al., 2004; Temple et al., 2003). In our study, increases in left
temporal and parahippocampal regions were associated with spelling improvement.
4) We aimed to achieve a better understanding of the neural basis of spelling
impairment, by investigating not only functional, but also structural brain characteristics of
children with spelling impairment. In our first study, controls showed increased FA in the left
SCR and anterior CC compared to children with SRI, whereas no differences between
controls and children with isolated SI were observed.

GENERAL DISCUSSION AND CONCLUSIONS
83
In our second study, we found differences in white matter integrity (increased FA
and decreased radial diffusivity in controls) primarily in the right hemisphere (SCR, PLIC,
SLF), in line with our functional findings. Regarding the effects of spelling intervention, our
findings indicate a more efficient signal transduction (possibly associated with greater
myelination) in the right hemisphere to be related with improvements of spelling skills.
Differences in white matter integrity were found between children with SRI and
controls. Decreased frontal white matter integrity has been associated with less efficient
communication (Steinbrink et al., 2008) in frontal brain regions and decreased working
memory capacity (Nagy et al., 2004; Niogi & McCandliss, 2006).
In our first study, no structural differences between controls and children with
isolated SI have been observed. Clearly, the structural characteristics of isolated SI should be
further investigated.
Increased right hemispheric (SCR, PLIC, SLF) white matter integrity in controls was
observed prior to the intervention. Decreased white matter integrity in children with SRI
compared to non-impaired controls, in the SCR and PLIC, has been reported in previous
studies. The corona radiata is primarily related to motor and somatosensory function; hence
an involvement of the SCR in reading and spelling ability is rather difficult to interpret.
Cerebellar deficits related to automatization impairments or reflections of differences in
interdigitating pathways (e.g.CC, SLF) have been suggested as possible explanations (Ben-
Shachar et al., 2007). The SLF is related to language, grapheme-phoneme conversion,
articulatory processing and reading (Ben-Shachar et al., 2007).
We observed intervention-related increases of right hemispheric white matter
integrity in the training group, along with behavioral improvement. Similar findings are
reported by Keller and Just (2009), who reported increased white matter integrity after 100
hours of reading instruction in the left anterior centrum semiovale in poor readers.
In conclusion, we observed enhanced functional activation and decreased white
matter integrity in the right hemisphere before intervention in our training group. The
intervention resulted in an improvement of spelling ability along with increased activation of
the left temporal (parahippocampal and hippocampal) region and increased white matter
integrity in the right hemisphere. Further studies should address if and how changes in white
matter integrity in children with developmental learning disorders, are influencing BOLD
activity.

GENERAL DISCUSSION AND CONCLUSIONS
84
Some limitations have to be considered regarding our studies. First, the sample size
of ten children per group is rather small, resulting in reduced statistical power. However,
particularly in view of the complex longitudinal, multimodal study design, the presented
studies may nevertheless provide first indications of structural and functional characteristics
of spelling impairment and changes associated with successful intervention, and we hope that
this work will encourage future investigators to examine the neurobiology of spelling
impairment in larger samples.
Secondly, biological interpretation of DTI parameters is challenging. In general, FA
is related to fiber density, coherence and myelination. Therefore we speculate that the
observed increase in FA along with decreases of radial diffusivity could be indicative of
enhanced white matter integrity or greater myelination, related to successful training.
Acquisition of more encoding directions in DTI assessment along with associated
histopathological research in animal models may allow more accurate biological conclusions
in future studies.
Thirdly, interpretation of functional activation patterns and their relation to findings
of prior functional MRI studies remains difficult. Due to the scanner environment, the
performance and examination of spelling tasks is problematic, as the common method to
assess spelling ability is to read aloud words, which should be written by hand. Acoustic
representation of stimuli during event-related functional MRI is difficult because of the
scanner noise. In addition, the subject would have to write down words while lying inside the
scanner without moving the head (e.g. Beeson et al., 2003) or seeing the written word, which
further imposes difficulties. Therefore, most spelling tasks are presented visually and also
include reading processes. The orthographic decision task, which was applied in our studies,
required accurate perception of the presented word (or pseudoword) and a decision about the
correctness of its spelling. Therefore, we anticipated activation patterns associated with
reading, decision making and spelling ability. As expected, we observed activation in regions
of the reading and spelling network in the non-impaired control group during an orthographic
decision task. In line with current literature about reading impairment and dyslexia, we found
decreased occipito-temporal activation related to poor reading ability.
A major goal of cognitive neuroscience is to identify neural correlates of SRI and
risks for impairments at an early stage through combined behavioural and neuroscience
measures. In this way, the provision of preventive treatment and development of targeted,
evidence-based interventions specifically tailored for particular subtypes of SRI would be
possible.

GENERAL DISCUSSION AND CONCLUSIONS
85
Research focusing on spelling impairment is rare and therefore these studies provide
first indications of networks involved in spelling impairment. The information-processing
requirements of reading and spelling vary greatly across different orthographies and even
within a single orthography there are different subtypes of SRI (Hadzibeganovic et al., 2010).
Hence, our findings should be replicated in larger cohorts of spelling impaired individuals,
applying different tasks in diverse age groups and samples with varying levels of native
language ability, to achieve a better understanding of spelling impairment across development
and orthographies.
Combined behavioral and neuroimaging assessment possibly allows for the
identification of subtypes (e.g. presence of phonological, visual or cerebellar impairments)
and, consequently, the provision of appropriate treatment.
Studies of this kind already had implications for educational policy (e.g. providing
additional reading time on tests for children with SRI and adaption of reading and spelling
lessons due to recent research findings) and will also help to improve education in future.
We here highlighted the importance of the right hemisphere in spelling impairment
and we have shown that an intervention for five weeks already resulted in improvement of
spelling ability and changes in brain structure and functional activation patterns. We hope that
our study encourages further research on brain activation patterns in subjects with isolated
spelling impairment and associated structural brain characteristics. The importance of
multimodal longitudinal imaging, to examine mechanisms of neural plasticity on the
structural and functional level, is rising in research.
Taken together, this study provides first indications that distinct structural and
functional brain characteristics may underlie the closely related abilities of reading and
spelling. Furthermore, we hope that long-term effects of spelling interventions will be
investigated in the future. It would be also interesting to see how long changes in brain
activation and structure remain stable and how frequent and intense interventions should be,
in order to ensure permanent changes in spelling ability and brain structure and function.

REFERENCES
86
V REFERENCES
Alexander, A., & Slinger-Constant, A. (2004). Current Status of Treatments for Dyslexia: Critical Review. Journal of Child Neurology, 19, 744.
Angelelli, P., Notarnicola, A., Judica, A., Zoccolotti, P, & Luzzatti, C. (2010). Spelling impairments in Italian dyslexic children: phenomenological changes in primary school. Cortex, 46, 1299-1311.
APA (American Psychiatric Association). (1994). Diagnostic and Statistical Manual of Mental Disorders (DSM-IV). Washington, DC.
Arnbak, E., & Elbro, C. (2000). The Effects of Morphological Awareness Training on the Reading and Spelling Skills of Young Dyslexics. Scandinavian Journal of Educational Research, 44, 229-251.
Arnold, E. M., Goldston, D. B., Walsh, A. K., Reboussin, B. A., Daniel, S. S., Hickman, E., et al. (2005). Severity of emotional and behavioral problems among poor and typical readers. Journal of Abnormal Child Psychology, 33, 205–217.
Asendorpf, J.B. (1998). Fünf-Faktoren-Fragebogen für Kinder (FFFK). Berlin: Humboldt-Universität, Institut für Psychologie.
Augst, G. (1989) Schriftwortschatz: Untersuchungen und Wortlisten zum orthographischen Lexikon bei Schülern und Erwachsenen. Peter Lang, Frankfurt am Main.
Aylward, E. H., Richards, T. L., Berninger, V. W., Nagy, W. E., Field, K. M., Grimme, C., Richards, L., et al. (2003). Instructional treatment associated with changes in brain activation in children with dyslexia. Neurology, 61, 212-219.
Bartl-Pokorny, K.D., Landerl, K., Einspieler, C., Enzinger, C., Gebauer, D., Fink, A., Zhang, D., Kozel, N., Kargl, R., Seither-Preisler, A., Vollmann, R., & Marschik, P.B. (2011). Dyslexie und ihre neuronale Signatur. Klinische Neurophysiologie, 42, 166-171.
Basser, P.J. (1995). Inferring microstructural features and the physiological state of tissues from diffusion-weighted images. NMR Biomedicine, 8, 333-344.
Basser, P.J., & Pierpaoli, C. (1996). Microstructural and physiological features of tissues elucidated by quantitative-diffusion-tensor MRI. Journal of Magnetic Resonance B, 111, 209-219.
Beaulieu, C., Plewes, C., Paulson, L. A., Roy, D., Snook, L., Concha, L., & Phillips, L. (2005). Imaging brain connectivity in children with diverse reading ability. NeuroImage, 25, 1266-71.
Beeson, P.M., Rapcsak, S.Z., Plante, E., Chargualaf, J., Chung, A., Johnson, S., & Trouard, T. (2003). The neural substrates of writing: A functional magnetic resonance imaging study. Aphasiology, 17, 647-665.
Ben-Shachar, M., Dougherty, R. F., & Wandell, B. a. (2007). White matter pathways in reading. Current opinion in neurobiology, 17, 258-70.

REFERENCES
87
Bengtsson, S. L., Nagy, Z., Skare, S., Forsman, L., Forssberg, H., & Ullén, F. (2005). Extensive piano practicing has regionally specific effects on white matter development. Nature neuroscience, 8, 1148-1150.
Bergmann, J., & Wimmer, H. (2008). A dual-route perspective on poor reading in a regular orthography: evidence from phonological and orthographic lexical decisions. Cognitive neuropsychology, 25, 653-676.
Berryhill, M., & Olson, I. (2008). The right parietal lobe is critical for visual working memory. Neuropsychologia, 46, 1767-1774.
Bhatt, C. (1991) Einführung in die Morphologie. Gabel, Hürth-Efferen.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009). Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cerebral cortex, 19, 2767-2796.
Booth, J. R., Burman, D. D., Meyer, J. R., Gitelman, D. R., Parrish, T. B., & Mesulam, M. M. (2002). Functional Anatomy of Intra- and Cross-Modal Lexical Tasks. Neuroimage, 16, 7-22.
Booth, J. R., Burman, D. D., Meyer, J. R., Gitelman, D. R., Parrish, T. B., & Mesulam, M. M. (2004). Development of brain mechanisms for processing orthographic and phonologic representations. Journal of cognitive neuroscience, 16, 1234-49.
Brambati, S. M., Termine, C., Ruffino, M., Stella, G., Fazio, F., Cappa, S. F., & Perani, D. (2004). Regional reductions of gray matter volume in familial dyslexia. Neurology, 63, 742-745.
Brown, W. E., Eliez, S., Menon, V., Rumsey, J. M., White, C. D., & Reiss, a L. (2001). Preliminary evidence of widespread morphological variations of the brain in dyslexia. Neurology, 56, 781-3.
Brown, T.T., Lugar, H.M., Coalson, R.S., Miezin, F.M., Petersen, S.E., & Schlaggar, B.L. (2005). Developmental changes in human cerebral functional organization for word generation. Cerebral Cortex, 15, 275-290.
Brunswick, N. (1999). Explicit and implicit processing of words and pseudowords by adult developmental dyslexics: A search for Wernicke’s Wortschatz? Brain, 122, 1901-1917.
Bundesministerium für Unterricht und Kunst (BMUKK): Evidenzbasierte LRS-Förderung: http://www.schulpsychologie.at/lernen-leistung/lese-rechtschreibschwaeche/ (last accessed 06.04.2011)
Caplan, R. & Dapretto, M. (2001). Making sense during conversation: An fMRI study. NeuroReport, 12, 3625-3632.
Carter, J. C., Lanham, D. C., Cutting, L. E., Clements-Stephens, A. M., Chen, X., Hadzipasic, M., Kim, J., et al. (2009). A dual DTI approach to analyzing white matter in children with dyslexia. Psychiatry research, 172, 215-219.
Casanova, M. F., Araque, J., Giedd, J., & Rumsey, J. M. (2004). Reduced Brain Size and Gyrification in the Brains of Dyslexic Patients. Journal of Child Neurology, 19, 275-281.

REFERENCES
88
Casanova, M.F., Christensen, J.D., Giedd, J., Rumsey, J., Garver, D., & Postel, G. (2005). Magnetic Resonance Imaging study of brain asymmetries in dyslexic patients. Journal of Child Neurology, 20, 842-849.
Cavanna, A. E., & Trimble, M. R. (2006). The precuneus: a review of its functional anatomy and behavioural correlates. Brain: a journal of neurology, 129, 564-583.
Cloutman, L., Gingis, L., Newhart, M., Davis, C., Heidler-Gary, J., Crinion, J., & Hillis, A.E. (2009). A Neural Network Critical for Spelling. Annals of Neurology, 66, 249-253.
Cohen, L., Dehaene, S., Naccache, L., Lehéricy, S., Dehaene-Lambertz, G., Hénaff, M.A., & Michel, F. (2000). The visual word form area: spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain, 123, 291-307.
Cohen, L., Dehaene, S., Vinckier, F., Jobert, A., & Montavont, A. (2008). Reading normal and degraded words: Contribution of the dorsal and ventral visual pathways. NeuroImage, 40, 353-366.
Cunningham, A. & Stanovich, K. (1998). What reading does for the mind. American Educator, 22, 8-15.
Daniel, S. S., Walsh, A. K., Goldston, D. B., Arnold, E. M., Reboussin, B. A., & Wood, F. B. (2006). Suicidality, school dropout, and reading problems among adolescents. Journal of Learning Disabilities, 39, 507–514.
Dehaene, S., & Cohen, L. (2011). The unique role of the visual word form area in reading. Trends in Cognitive Sciences, 15, 254-262.
Deutsch, Gayle K., Dougherty, R. F., Bammer, R., Siok, W. T., Gabrieli, J. D. E., & Wandell, B. (2005). Children’s Reading Performance is Correlated with White Matter Structure Measured by Diffusion Tensor Imaging. Cortex, 41, 354-363.
Diana, R.A., Yonelinas, A.P., & Ranganath, C. (2007). Imaging recollection and familiarity in the medial temporal lobe: a three-component model. Trends in Cognitive Science, 11, 379-386.
Dilling, H., Mombour, W., & Schmidt, M.H. (2005). Internationale Klassifikation psychischer Störungen. ICD-10 V (F). Klinisch-diagnostische Leitlinien (5th ed.). Bern: Hans Huber.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., & May, A. (2004) Changes in gray matter induced by training. Nature, 427, 311-312.
Dougherty, R. F., Ben-Shachar, M., Deutsch, G. K., Hernandez, A., Fox, G. R., & Wandell, B. a. (2007). Temporal-callosal pathway diffusivity predicts phonological skills in children. Proceedings of the National Academy of Sciences, 104, 8556-61.
Eckert, M. (2003). Anatomical correlates of dyslexia: frontal and cerebellar findings. Brain, 126, 482-494.
Eden, G., & Zeffiro, T. (1998). Neural Systems Affected in Developmental Dyslexia Revealed by Functional Neuroimaging. Neuron, 21, 279-282.

REFERENCES
89
Eden, G., Brown, C., Jones, K., Given, B., & Zeffiro, T. (2000). Phonological and visual motion processing in reading impaired children. NeuroImage, 11, 183.
Eden, G., Jones, K. M., Cappell, K., Gareau, L., Wood, F. B., Zeffiro, T. A., Dietz, N., et al. (2004). Neural Changes following Remediation in Adult Developmental Dyslexia. Neuron, 44, 411-422.
Eickhoff, S.B., Laird, A., Grefkes, C., Wang, I., Zilles, K., & Fox, P. (2009). Coordinate-based activation likelihood estimation based on empirical estimates of spatial uncertainty. Hum Brain Mapping, 30, 2907-2926.
Esser, G., Wyschkon, A., & Schmidt, M. (2002). Was wird aus Achtjährigen mit einer Lese- und Rechtschreibstörung [Long-term outcome in 8-year-old children with specific reading retardation: results at age 25 years]. Zeitschrift für Klinische Psychologie und Psychotherapie, 31, 235–242.
Faber, G. (2010). Enhancing orthographic competencies and reducing domain-specific test anxiety: The systematic use of algorithmic and self-instructional task formats in remedial spelling training. International Journal of Special Education, 25, 78-88.
Fisher, S. E., & DeFries, J. C. (2002). Developmental dyslexia: genetic dissection of a complex cognitive trait. Nature reviews. Neuroscience, 3, 767-80.
Fletcher, P.C., Shallice, T., Frith, C.D., Frackowiak, R.S., & Dolan, R.J. (1998). The functional roles of prefrontal cortex in episodic memory. II Retrieval. Brain, 121, 1249-1256.
Fluss, J., Ziegler, J. C., Warszawski, J., Ducot, B., Richard, G., & Billard, C. (2009). Poor reading in French elementary school: the interplay of cognitive, behavioral and socioeconomic factors. Journal of Developmental & Behavioral Pediatrics, 30, 206–216.
Fornito, A., Yücel, M., Wood, S., Stuart, G.W., Buchanan, J.A., Proffitt, T., Anderson, V., Velakoulis, D. & Pantelis, C. (2004). Individual Differences in Anterior Cingulate/Paracingulate Morphology are related to Executive Functions in Healthy Males. Cereb. Cortex, 14, 424-431.
Frith, U. (1985). Beneath the surface of developmental dyslexia. In K.E. Patterson, J.C., Marshall, M., Coltheart (Eds.), Surface dyslexia: Neuropsychological and cognitive studies of phonological reading. London: Erlbaum.
Fulbright, R. K., Jenner, A. R., Mencl, W. E., Pugh, K. R., Shaywitz, B. A., & Shaywitz, S. E. (1999). The Cerebellum ’ s Role in Reading: A Functional MR Imaging Study. American Journal Of Neuroradiology, 20, 1925-1930.
Gaab, N, Gabrieli, J., Deutsch, G. K., Tallal, P., & Temple, E. (2007). Neural correlates of rapid auditory processing are disrupted in children with developmental dyslexia and ameliorated with training: an fMRI study. Restorative neurology and neuroscience, 25, 295-310.
Gabrieli, J. (2009). Dyslexia: a new synergy between education and cognitive neuroscience. Science, 325, 280-283.

REFERENCES
90
Galaburda, A. M., LoTurco, J., Ramus, F., Fitch, R. H., & Rosen, G. D. (2006). From genes to behavior in developmental dyslexia. Nature neuroscience, 9, 1213-7.
Gimbel, S.I., & Brewer, J.B. (2011). Reaction time, memory strength, and fMRI activity during memory retrieval: Hippocampus and default network are differentially responsive during recollection and familiarity judgments. Cognitive Neurosci., 2, 19-23.
Graves, W. W., Binder, J. R., Desai, R. H., Conant, L. L., & Seidenberg, M. S. (2010). Neural correlates of implicit and explicit combinatorial semantic processing. NeuroImage, 53, 638-646.
Groth, K., Lachmann, T., Riecker, A., Muthmann, I., & Steinbrink, C. (2011). Developmental dyslexics show deficits in the processing of temporal auditory information in German vowel length discrimination. Reading and Writing, 24, 285-303.
Habib, M. (2000). The neurological basis of developmental dyslexia: an overview and working hypothesis. Brain: a journal of neurology, 123, 2373-2399.
Hadzibeganovic, T., van den Noort, M., Bosch, P., Perc, M., van Kralingen, R., Mondt, K., & Coltheart, M. (2010). Cross-linguistic neuroimaging and dyslexia: A critical view. Cortex, 46, 1312-1316.
Heim, S., Eickhoff, S. B., Ischebeck, A. K., Friederici, A. D., Stephan, K. E., & Amunts, K. (2009). Effective connectivity of the left BA 44, BA 45, and inferior temporal gyrus during lexical and phonological decisions identified with DCM. Human brain mapping, 30, 392-402.
Henry, M.L., Beeson, P.M., Stark, A.J., & Rapcsak, S.Z. (2007). The role of left perisylvian cortical regions in spelling. Brain and Language, 100, 44-52.
Hillis, A.E., Kane, A., Tuffiash, E., Beauchamp, N., Norman, J., Barker, P., Jacobs, M., & Wityk, R. (2002). Neural substrates of the cognitive processes underlying spelling: Evidence from MR diffusion and perfusion imaging. Aphasiology, 16, 425-438.
Hillis, A.E., Chang, S., Breese, E., & Heidler, J. (2004). The crucial role of posterior frontal regions in modality specific components of spelling process. Neurocase, 10, 175-187.
Hillis, A.E., Newhart, M., Heidler, J., Barker, P., Herskovits, E., & Degaonkar, M. (2005). The roles of the “visual word form area” in reading. NeuroImage, 24, 548-559.
Hoeft, F., McCandliss, B. D., Black, J. M., Gantman, A., Zakerani, N., Hulme, C., Lyytinen, H., et al. (2011). Neural systems predicting long-term outcome in dyslexia. Proceedings of the National Academy of Sciences, 108, 361-366.
Hoeft, F., Meyler, A., Hernandez, A., Juel, C., Taylor-Hill, H., Martindale, J. L., McMillon, G., et al. (2007). Functional and morphometric brain dissociation between dyslexia and reading ability. Proceedings of the National Academy of Sciences, 104, 4234-4239.
Ise, E., & Schulte-Körne, G. (2010). Spelling deficits in dyslexia: evaluation of an orthographic spelling training. Annals of dyslexia, 60, 18-39.
Jobard, G., Crivello, F., & Tzourio-Mazoyer, N. (2003). Evaluation of the dual route theory of reading: a metanalysis of 35 neuroimaging studies. NeuroImage, 20, 693-712.

REFERENCES
91
Kargl, R., Purgstaller, C., Weiss, S. & Fink, A. (2008) Effektivitätsüberprüfung eines morphemorientierten Grundwortschatz-Segementierungstrainings (MORPHEUS) bei Kindern und Jugendlichen. Heilpädagogische Forschung, 31, 147-156.
Kargl, R. & Purgstaller, C. (2010) Morphem unterstütztes Segmentierungstraining MORPHEUS. Hogrefe, Göttingen.
Kargl, R., Purgstaller, C., Mrazek, C., Ertl, K. & Fink, A. (2011) Förderung der Lese- und Rechtschreibkompetenz auf Basis des morphematischen Prinzips. Zeitschrift für Heilpädagogik, 2, 61-68.
Katanoda, K., Yoshikawa, K., & Sugishita, M. (2001). A functional MRI study on the neural substrates for writing. Human Brain Mapping, 13, 34-42.
Keller, T., & Just, M. (2009). Altering Cortical Connectivity: Remediation-Induced Changes in the White Matter of Poor Readers. Neuron, 64, 624-631.
Klicpera C, Schabmann A, Gasteiger-Klicpera B. (2007). Legasthenie. München, Basel: Ernst Reinhardt Verlag.
Klingberg, T, Hedehus, M., Temple, E., Salz, T., Gabrieli, J. D., Moseley, M. E., & Poldrack, R. a. (2000). Microstructure of temporo-parietal white matter as a basis for reading ability: evidence from diffusion tensor magnetic resonance imaging. Neuron, 25, 493-500.
Koenigs, M., Barbey, A., Postle, B., & Grafman, J. (2009). Superior Parietal Cortex Is Critical for the Manipulation of Information in Working Memory. The Journal of Neuroscience, 29, 14980-14986.
Kozel, N., Kargl, R., Purgstaller, C., & Fink, A. (submitted). Uncovering the neurophysiological effects of a morpheme-based spelling training in children.
Krafnick, A. J., Flowers, D. L., Napoliello, E. M., & Eden, G. F. (2011). Gray matter volume changes following reading intervention in dyslexic children. NeuroImage, 57, 733-41.
Kronbichler, M., Bergmann, J., Hutzler, F., Staffen, W., Mair, A., Ladurner, G., & Wimmer, H. (2007). Taxi vs. taksi: on orthographic word recognition in the left ventral occipitotemporal cortex. Journal of cognitive neuroscience, 19, 1584-94.
Kronbichler, M., Hutzler, F., Staffen, W., Mair, A., Ladurner, G., & Wimmer, H. (2006). Evidence for a dysfunction of left posterior reading areas in German dyslexic readers. Neuropsychologia, 44, 1822-32.
Kronbichler, M., Wimmer, H., Staffen, W., Hutzler, F., Mair, A., & Ladurner, G. (2008). Developmental dyslexia: gray matter abnormalities in the occipitotemporal cortex. Human brain mapping, 29, 613-25.
Landerl, K. & Klicpera, C. (2009). Lese- und Rechtschreibstörung. In F. Petermann (Ed.), Fallbuch der Klinischen Kinderpsychologie (3rd ed.). (pp. 193-207). Göttingen: Hogrefe.
Landerl, K., & Wimmer, H. (2008). Development of word reading fluency and spelling in a consistent orthography: An 8-year follow-up. Journal of Educational Psychology, 100, 150-161.

REFERENCES
92
Lanzinger, S., Weder, B., Oettly, R., & Fretz, C. (1999). Neuroimaging findings in a patient recovering from global alexia to spelling dyslexia. Journal of Neuroimaging, 9, 48-51.
Lenhard, W. & Schneider, W. (2006) ELFE 1-6. Ein Leseverständnistest für Erst- bis Sechstklässler. Hogrefe, Göttingen.
Lyster, S. H. (2002). The effects of morphological versus phonological awareness training in kindergarten on reading development. Reading and Writing, 15, 261-294.
Maisog, J. M., Einbinder, E. R., Flowers, D. L., Turkeltaub, P. E., & Eden, G. F. (2008). A meta-analysis of functional neuroimaging studies of dyslexia. Annals of the New York Academy of Sciences, 1145, 237-59.
Makris, N., Kennedy, D. N., McInerney, S., Sorensen, G., Wang, R., Caviness, V. S., & Pandya, D. N. (2005). Segmentation of subcomponents within the superior longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study. Cerebral cortex, 15, 854-69.
Maldonado, I. L., Moritz-Gasser, S., & Duffau, H. (2011). Does the left superior longitudinal fascicle subserve language semantics? A brain electrostimulation study. Brain structure & function, 216, 263-74.
Maughan, B., Rowe, R., Loeber, R., & Stouthamer-Loeber, M. (2003). Reading problems and depressed mood. Journal of Abnormal Child Psychology, 31, 219–229.
May, P., Vieluf, U., & Malitzky, V. (2000). Hamburger Schreibprobe. Diagnose orthographischer Kompetenz. Hamburg: Verlag für pädagogische Medien.
Mayringer, H., & Wimmer, H. (2003) Salzburger Lese-Screening für die Klassenstufen 1 - 4. Hans Huber, Bern.
Mayringer, H., & Wimmer, H. (2005) Salzburger Lese-Screening für die Klassenstufen 5 - 8. Hans Huber, Bern.
Meyler, A., Keller, T. a, Cherkassky, V. L., Gabrieli, J. D. E., & Just, M. A. (2008). Modifying the brain activation of poor readers during sentence comprehension with extended remedial instruction: a longitudinal study of neuroplasticity. Neuropsychologia, 46, 2580-92.
Meyler, A., Keller, T. a, Cherkassky, V. L., Lee, D., Hoeft, F., Whitfield-Gabrieli, S., Gabrieli, J. D. E., et al. (2007). Brain activation during sentence comprehension among good and poor readers. Cerebral cortex, 17, 2780-7.
Miceli, G. (2001). Disorders of single word processing. Journal of Neurology, 248, 658-664.
Moll, K., & Landerl, K. (2009). Double Dissociation Between Reading and Spelling Deficits. Scientific Studies of Reading, 13, 359-382.
Morgan, P. L., Farkas, G., Tufis, P. A., & Sperling, R. A. (2008). Are reading and behavior problems risk factors for each other? Journal of Learning Disabilities, 41, 417–436.
Nagy, Zoltan, Westerberg, H., & Klingberg, T. (2004). Maturation of white matter is associated with the development of cognitive functions during childhood. Journal of cognitive neuroscience, 16, 1227-33.

REFERENCES
93
Nichols, T.E., & Holmes, A.P. (2002) Nonparametric Permutation Tests for Functional Neuroimaging: A Primer with Examples. Human Brain Mapping, 15, 1-25.
Nicolson, R. I., Fawcett, a J., Berry, E. L., Jenkins, I. H., Dean, P., & Brooks, D. J. (1999). Association of abnormal cerebellar activation with motor learning difficulties in dyslexic adults. Lancet, 353, 1662-1667.
Nicolson, R., Fawcett, A., & Dean, P. (2001). Dyslexia, development and the cerebellum. Trends in Neurosciences, 24, 515-516.
Niogi, S. N., & McCandliss, B. D. (2006). Left lateralized white matter microstructure accounts for individual differences in reading ability and disability. Neuropsychologia, 44, 2178-88.
Nunes, T., Bryant, P., & Olsson, J. (2003). Learning Morphological and Phonological Spelling Rules: An Intervention Study. Scientific Studies of Reading, 7, 289-307.
Petrides, M., Alivisatos, B., & Evans, A.C. (1995). Functional activation of the human ventrolateral frontal cortex during mnemonic retrieval of verbal information. PNAS USA, 92, 5803-5807.
Peyrin, C., Démonet, J., Guyen-Morel, M., Le Bas, J., & Valdois, S. (2010). Superior parietal lobule dysfunction in a homogenous group of dyselxic children with a visual attention span disorder. Brain & Language, 118, 128-138.
Plume, E., & Warnke, a. (2007). Definition, Symptomatik, Prävalenz und Diagnostik der Lese-Rechtschreib-Störung. Monatsschrift Kinderheilkunde, 155, 322-327.
Poldrack, R.A., & Gabrieli, J. D. (2001). Characterizing the neural mechanisms of skill learning and repetition priming: evidence from mirror reading. Brain: a journal of neurology, 124, 67-82.
Poldrack, R.A., Desmond, J.E., Glover, G.H., & Gabrieli, J.D. (1998). The neural basis of visual skill learning: an fMRI study of mirror reading. Cereb Cortex, 8, 1– 10.
Purcell, J.J., Napoliello, E.M., & Eden, G. (2011). A combined fMRI study of typed spelling and reading. NeuroImage, 55, 750-762.
Raichle, M.E., Fiez, J.A., Videen, T.O., MacLeod, A.M., Pardo, J.V., Fox, P.T., & Petersen, S.E. (1994). Practice-related changes in human brain functional anatomy during nonmotor learning. Cereb Cortex, 4, 8– 26.
Ramus, F., Rosen, S., Dakin, S. C., Day, B. L., Castellote, J. M., White, S., & Frith, U. (2003). Theories of developmental dyslexia: insights from a multiple case study of dyslexic adults. Brain: a journal of neurology, 126, 841-65.
Rapcsak, S.Z., & Beeson, P.M. (2002). Neuroanatomical correlates of spelling and writing. In A.E.Hillis (Ed.), Handbook of Adult Language Disorders: Integrating Cognitive Neuropsychology, Neurology and Rehabilitation. (pp. 71-99). Philadelphia: Psychology Press.
Rapcsak, S.Z., & Beeson, P.M. (2004). The role of left posterior inferior temporal cortex in spelling. Neurology, 62, 2221-2229.

REFERENCES
94
Rapp, B., & Lipka, K. (2011). The literate brain: the relationship between spelling and reading. Journal of Cognitive Neuroscience, 23, 1180-1897.
Raschle, N. M., Chang, M., & Gaab, N. (2010). NeuroImage Structural brain alterations associated with dyslexia predate reading onset. NeuroImage, 57, 742-749.
Raven, J.C. (1960) Guide to the Standard Progressive Matrices: Sets A, B, C, D and E. HK Lewis, London.
Riddick, B., Sterling, C., Farmer, M. & Morgan, S. (1999). Self-esteem and anxiety in the educational histories of adult dyslexic students. Dyslexia, 5, 227-248.
Richards, T., Aylward, E., Berninger, V., Field, K., Grimme, a, Richards, a, & Nagy, W. (2006). Individual fMRI activation in orthographic mapping and morpheme mapping after orthographic or morphological spelling treatment in child dyslexics. Journal of Neurolinguistics, 19, 56-86.
Richards, Todd L., Berninger, V. W., & Fayol, M. (2009). fMRI activation differences between 11-year-old good and poor spellers’ access in working memory to temporary and long-term orthographic representations. Journal of Neurolinguistics, 22, 327-353.
Richlan, F., Kronbichler, M., & Wimmer, H. (2009). Functional abnormalities in the dyslexic brain: a quantitative meta-analysis of neuroimaging studies. Human brain mapping, 30, 3299-308.
Richlan, F., Kronbichler, M., & Wimmer, H. (2011). Meta-analyzing brain dysfunctions in dyslexic children and adults. NeuroImage, 56, 1735-42.
Richlan, F., Sturm, D., Schurz, M., Kronbichler, M., Ladurner, G., & Wimmer, H. (2010). A common left occipito-temporal dysfunction in developmental dyslexia and acquired letter-by-letter reading? PloS one, 5, e12073.
Rimrodt, S.L., Peterson, D.J., Denckla, M.B., Kaufmann, W.E., & Cutting, L.E. (2010). White matter microstructural differences linke to left perisylvian language network in children with dyslexia. Cortex, 46, 739-749.
Riva, D., & Giorgi, C. (2000). The cerebellum contributes to higher functions during development: evidence from a series of children surgically treated for posterior fossa tumours. Brain: a journal of neurology, 123, 1051-1061.
Roeltgen, D.P. (1993). Agraphia. In: Heilman, K.M., Valenstein, E. (Eds.) Clinical Neuropsychology. (pp. 63-89). New York: Oxford University Press.
Rollins, N. K., Pickering, J., & Hughes, C. W. (2009). Simple Developmental Dyslexia in Children: Alterations in Diffusion- Tensor Metrics of White Matter Tracts at 3 T. Radiology, 251, 882-891.
Scerri, T. S., & Schulte-Körne, G. (2010). Genetics of developmental dyslexia. European child & adolescent psychiatry, 19, 179-97.
Scheerer-Neumann, G. (1979) Intervention bei Lese-Rechtschreibschwäche. Überblick über Theorien, Methoden und Ergebnisse. Kamp, Bochum.

REFERENCES
95
Schneeberger, B., Kargl, R., Purgstaller, C., Kozel, N., Gebauer, D., Vogl, J., Rohrer, S., & Fink A. (2011). Förderung von Kindern und Jugendlichen mit Problemen im Schriftspracherwerb. Zeitschrift für Heilpädagogik, 12, 476-483.
Scholz, J., Klein, M. C., Behrens, T. E. J., & Johansen-Berg, H. (2009). Training induces changes in white-matter architecture. Nature neuroscience, 12, 1370-1371.
Schulte-Körne, G., Warnke, A., & Remschmidt, H. (2006). [Genetics of dyslexia]. Zeitschrift für Kinder- und Jugendpsychiatrie und Psychotherapie, 34, 435-444.
Schulte-Körne, G., & Remschmidt, H. (2003). Legasthenie- Symptomatik, Diagnostik, Ursachen, Verlauf und Behandlung. Deutsches Ärzteblatt, 7, 396-406.
Schulte-Körne, G. (2001). Annotation: Genetics of reading and spelling disorder. Journal of child psychology and psychiatry, and allied disciplines, 42, 985-97.
Schurz, M., Sturm, D., Richlan, F., Kronbichler, M., Ladurner, G., & Wimmer, H. (2010). A dual-route perspective on brain activation in response to visual words: evidence for a length by lexicality interaction in the visual word form area (VWFA). NeuroImage, 49, 2649-2661.
Scott, R. B., Stoodley, C. J., Anslow, P., Paul, C., Stein, J. F., Sugden, E. M., & Mitchell, C. D. (2001). Lateralized cognitive deficits in children following cerebellar lesions. Developmental medicine and child neurology, 43, 685-91.
Sen, P.N., & Basser, P.J. (2005). A model for diffusion in white matter in the brain. Biophys J, 89, 2927-2938.
Shaywitz, S. E., Morris, R., & Shaywitz, B. (2008). The education of dyslexic children from childhood to young adulthood. Annual review of psychology, 59, 451-75.
Shaywitz, B., Skudlarski, P., Holahan, J. M., Marchione, K. E., Constable, R. T., Fulbright, R. K., Zelterman, D., et al. (2007). Age-related changes in reading systems of dyslexic children. Annals of neurology, 61, 363-70.
Shaywitz, B., Lyon, G. R., & Shaywitz, S. E. (2006). The role of functional magnetic resonance imaging in understanding reading and dyslexia. Developmental neuropsychology, 30, 613-32.
Shaywitz, B., Shaywitz, S. E., Blachman, B. a, Pugh, K. R., Fulbright, R. K., Skudlarski, P., Mencl, W. E., et al. (2004). Development of left occipitotemporal systems for skilled reading in children after a phonologically- based intervention. Biological psychiatry, 55, 926-33.
Shaywitz, S. E, Shaywitz, B., Fulbright, R. K., Skudlarski, P., Mencl, W. E., Constable, R. T., Pugh, K. R., et al. (2003). Neural Systems for Compensation and Persistence: Young Adult Outcome of Childhood Reading Disability. Biological Psychiatry, 54, 25-33.
Silani, G., Frith, U., Demonet, J.-F., Fazio, F., Perani, D., Price, C., Frith, C. D., et al. (2005). Brain abnormalities underlying altered activation in dyslexia: a voxel based morphometry study. Brain: a journal of neurology, 128, 2453-2461.

REFERENCES
96
Simos, P G, Fletcher, J. M., Bergman, E., Breier, J. I., Foorman, B. R., Castillo, E. M., Davis, R. N., et al. (2002). Dyslexia-specific brain activation profile becomes normal following successful remedial training. Neurology, 58, 1203-1213.
Simos, P. G., Fletcher, J. M., Denton, C., Sarkari, S., Billingsley-Marshall, R., & Papanicolaou, A. C. (2006). Magnetic source imaging studies of dyslexia interventions. Developmental neuropsychology, 30, 591-611.
Smith, S.M., & Nichols, T.E. (2009). Threshold-free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. NeuroImage, 44, 83-98.
Stein, J. (2001). The magnocellular theory of developmental dyslexia. Dyslexia, 7, 12-36.
Steinbrink, C., Vogt, K., Kastrup, a, Müller, H.-P., Juengling, F. D., Kassubek, J., & Riecker, A. (2008). The contribution of white and gray matter differences to developmental dyslexia: insights from DTI and VBM at 3.0 T. Neuropsychologia, 46, 3170-3178.
Steinbrink, C., Ackermann, H., Lachmann, T., & Riecker, A. (2009). Contribution of the anterior insula to temporal auditory processing deficits in developmental dyslexia. Human brain mapping, 30, 2401-11.
Suchodoletz, W. (2010). Konzepte in der LRS-Therapie. Zeitschrift für Kinder- und Jugendpsychiatrie und Psychotherapie, 38, 329-339.
Sugihara, G., Kaminaga, T., & Sugishita, M. (2006). Interindividual uniformity and variety of the “Writing center”: a functional MR study. NeuroImage, 32, 1837-1849.
Temple, E., Deutsch, G. K., Poldrack, R., Miller, S. L., Tallal, P., Merzenich, M. M., & Gabrieli, J. D. E. (2003). Neural deficits in children with dyslexia ameliorated by behavioral remediation: evidence from functional MRI. Proceedings of the National Academy of Sciences, 100, 2860-5.
Tsapkini, K. & Rapp, B. (2010). The orthography-specific functions of the left fusiform gyrus: evidence of modality and category specifity. Cortex, 46, 185-205.
Van der Mark, S., Bucher, K., Maurer, U., Schulz, E., Brem, S., Buckelmüller, J., Kronbichler, M., et al. (2009). Children with dyslexia lack multiple specializations along the visual word-form (VWF) system. NeuroImage, 47, 1940-1949.
Van Ettinger-Veenstra, H., Ragnehed, M., McAllister, A., Lundberg, P., & Engström, M. (2011). Right-hemispheric cortical contributions to language ability in healthy adults. Brain and Language. E-pub ahead of print.
Vinckenbosch, E., Robichon, F., & Eliez, S. (2005). Gray matter alteration in dyslexia: converging evidence from volumetric and voxel-by-voxel MRI analyses. Neuropsychologia, 43, 324-31.
Wager, T.D., & Nichols, T.E. (2003). Optimization of Experimental Design in fMRI: A General Framework Using a Genetic Algorithm. NeuroImage, 18, 293-309.
Walter, J. (2001). Förderung bei Lese- und Rechtschreibschwäche. Grundlagenforschung, methodische Konsequenzen, Praxisbeispiele und mediendidaktische Anregungen auf der

REFERENCES
97
Basis empirischer Forschungsmethoden. Hogrefe Verlag: Göttingen, Bern, Toronto, Seattle.
Warnke, A., Hemminger, U. & Plume, E. (2004): Lese-Rechtschreibstörungen- Leitfaden. Kinder- und Jugendpsychiatrie. Göttingen: Hogrefe.
Weiss, S., Grabner, R.H., Kargl, R., Purgstaller, C. & Fink, A. (2010) Behavioral and neurophysiological effects of morphological awareness training on spelling and reading. Reading and Writing: An Interdisciplinary Journal, 23, 645–671.
Westerberg, H., & Klingberg, T. (2007) Changes in cortical activity after training of working memory - A single subject analysis. Physiol Behavior, 92, 186–192.
Wimmer H, & Schurz, M. (2010): Dyslexia in Regular Orthographies: Manifestation and Causation. Dyslexia 16:283-299.
Wimmer, H., & Mayringer, H. (2002). Dysfluent Reading in the Absence of Spelling Difficulties: A Specific Disability in Regular Orthographies. Journal of Educational Psychology, 94, 272-277.
Wimmer, H., Mayringer, H., & Landerl, K. (1998). Poor Reading: A deficit in skill-automatization or a phonological deficit? Scientific Studies of Reading, 2, 321-340.
Wolf, M., & Bowers, P. (1999). The Double-Deficit Hypothesis for Developmental Dyslexias. Journal of Educational Psychology, 91, 415-438.
World Health Organization. (2003). International Classification of Disease (ICD-10). Second Edition.
Zimdars, K. & Zink, S. (2006). Computergestützte Trainingsverfahren. In W.v. Suchodoletz (Hrsg.), Therapie der Lese-Rechtschreibstörung (LRS). Traditionelle und alternative Behandlungsmethoden im Überblick. (S. 58-81). Stuttgart: Kohlhammer.
APPENDIX
98
VI APPENDIX
Appendix Study I Table A.1: Mean activation for the Conditions Correctly Spelled Words, Misspelled Words, Pseudowords (vs. Fixation) for all three groups (SI, SRI, CG)– Coordinates (in MNI standard space) and Activation Significance (Z statistics) of Local Maxima of Clusters, Z>2.3, P corrected P=0.05. Region (Local Maxima) k Z x y z Correctly spelled words SI L precentral gyrus L postcentral gyrus L inferior frontal gyrus L middle frontal gyrus R occipital pole R cerebellum L occipital pole R Hippocampus R lateral occipital Cortex SRI R occipital fusiform gyrus R lateral occipital Cortex R occipital pole L precentral gyrus L postcentral gyrus L occipital pole L occipital fusiform gyrus
L anterior cingulum L SMA R paracingulate gyrus L inferior frontal gyrus L precentral gyrus CG R occipital pole
16322
12648
900
2763
2344
2139
967
845
17910
5.19 4.91 4.9 4.87 4.72 4.64 5.32 4.97 4.81 4.8 4.79 3.71 3.35 3.19
4.56 4.38 3.85 4.04 4.25 3.35 3.29 3.27 3.27 3.24 4.03 4.02 3.93 3.72 3.81 3.69 3.6 3.52 3.5 3.28 4.07 4.01 3.82 3.33 3.85
5.48 5.74
-38 -42 -38 -48 -44 -52 16 20 28 -22 -12 22 28 26
30 46 38 18 -34 -36 -56 -52 -46 -44 -20 -16 -30 -44 -6 -10 -8 -4 8 -6 -54 -54 -56 -56 -56
26 16
-16 -14 -14 -16 18 8
-92 -102 -70 -102 -100 -28 -38 -64
-84 -86 -86 -98 -8 -4 -24 -28 -28 -26 -104 -92 -80 -82 16 20 16 6 22 18 14 14 14 18 10
-94 -96
64 58 56 58 16 42 -4 2
-24 6 0 -6 4 50
-8 -12 -18 -4 62 56 42 62 58 52 -8 -12 -16 -16 38 30 34 54 40 48 12 8 30 24 28
-6 -2
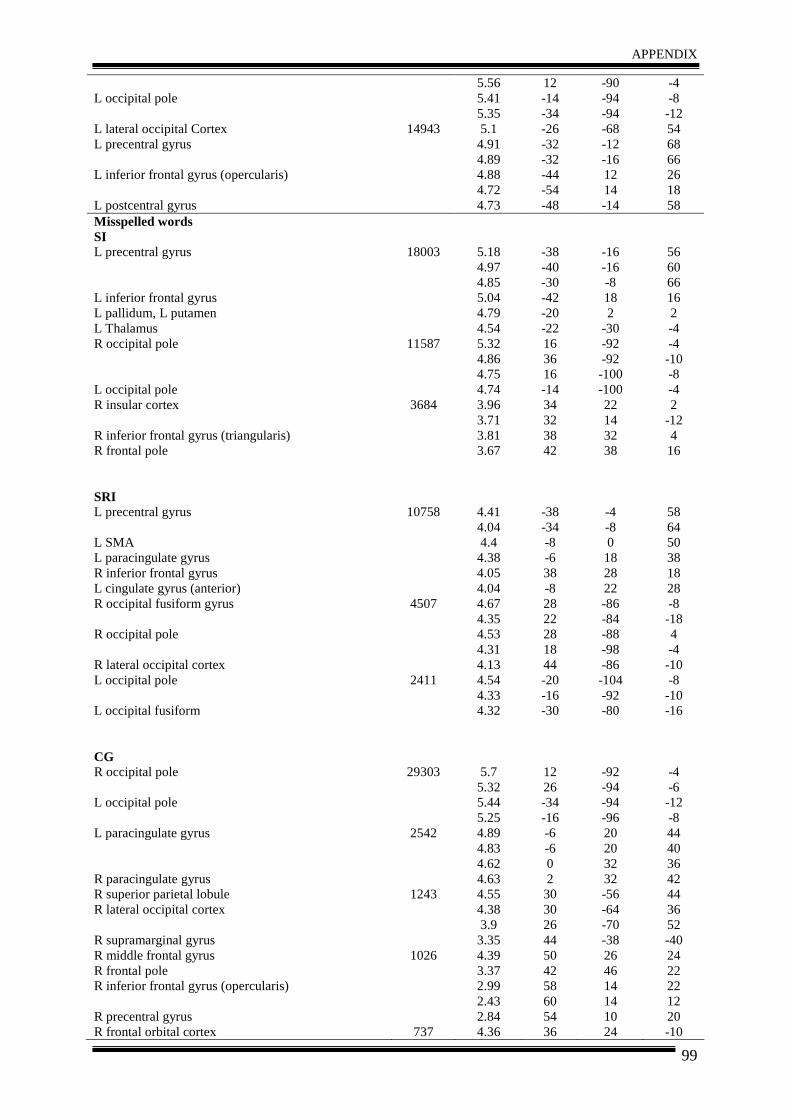
APPENDIX
99
L occipital pole L lateral occipital Cortex L precentral gyrus L inferior frontal gyrus (opercularis) L postcentral gyrus
14943
5.56 5.41 5.35 5.1 4.91 4.89 4.88 4.72 4.73
12 -14 -34 -26 -32 -32 -44 -54 -48
-90 -94 -94 -68 -12 -16 12 14 -14
-4 -8 -12 54 68 66 26 18 58
Misspelled words SI L precentral gyrus L inferior frontal gyrus L pallidum, L putamen L Thalamus R occipital pole L occipital pole R insular cortex R inferior frontal gyrus (triangularis) R frontal pole SRI L precentral gyrus L SMA L paracingulate gyrus R inferior frontal gyrus L cingulate gyrus (anterior) R occipital fusiform gyrus R occipital pole R lateral occipital cortex L occipital pole L occipital fusiform CG R occipital pole L occipital pole L paracingulate gyrus R paracingulate gyrus R superior parietal lobule R lateral occipital cortex R supramarginal gyrus R middle frontal gyrus R frontal pole R inferior frontal gyrus (opercularis) R precentral gyrus R frontal orbital cortex
18003
11587
3684
10758
4507
2411
29303
2542
1243
1026
737
5.18 4.97 4.85 5.04 4.79 4.54 5.32 4.86 4.75 4.74 3.96 3.71 3.81 3.67
4.41 4.04 4.4 4.38 4.05 4.04 4.67 4.35 4.53 4.31 4.13 4.54 4.33 4.32
5.7 5.32 5.44 5.25 4.89 4.83 4.62 4.63 4.55 4.38 3.9 3.35 4.39 3.37 2.99 2.43 2.84 4.36
-38 -40 -30 -42 -20 -22 16 36 16 -14 34 32 38 42
-38 -34 -8 -6 38 -8 28 22 28 18 44 -20 -16 -30
12 26 -34 -16 -6 -6 0 2 30 30 26 44 50 42 58 60 54 36
-16 -16 -8 18 2
-30 -92 -92 -100 -100 22 14 32 38
-4 -8 0 18 28 22 -86 -84 -88 -98 -86 -104 -92 -80
-92 -94 -94 -96 20 20 32 32 -56 -64 -70 -38 26 46 14 14 10 24
56 60 66 16 2 -4 -4 -10 -8 -4 2
-12 4 16
58 64 50 38 18 28 -8 -18 4 -4 -10 -8 -10 -16
-4 -6 -12 -8 44 40 36 42 44 36 52 -40 24 22 22 12 20 -10
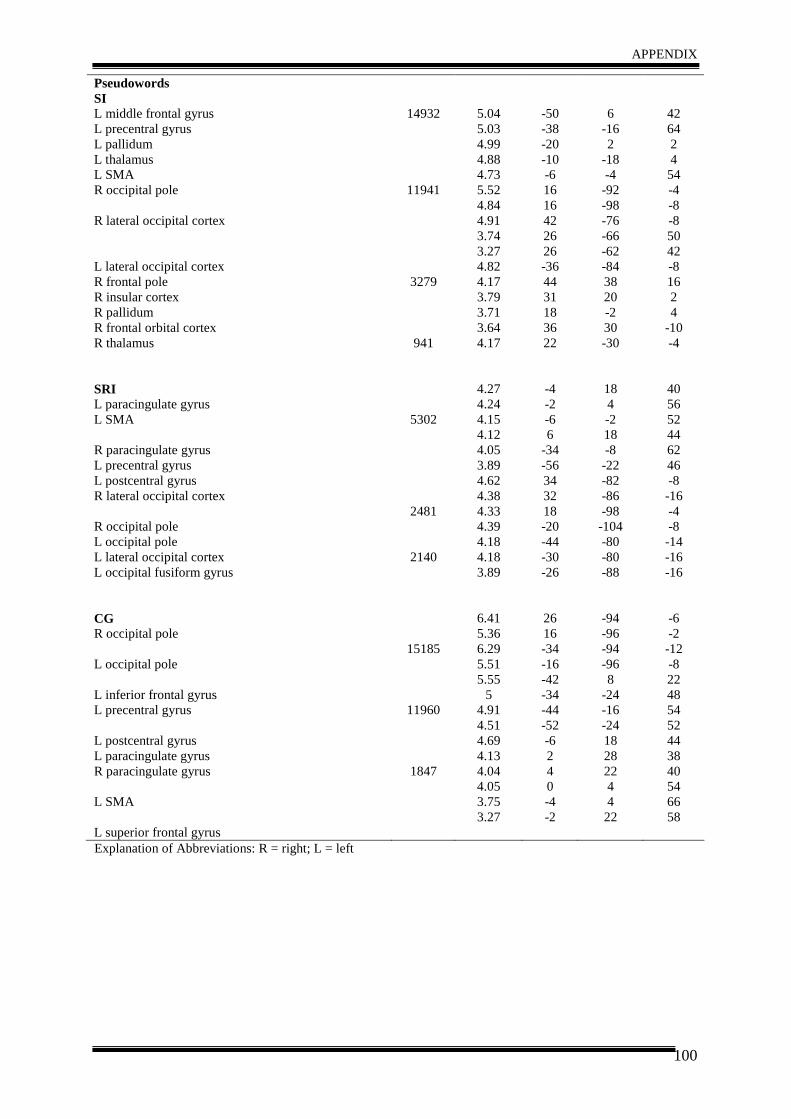
APPENDIX
100
Pseudowords SI L middle frontal gyrus L precentral gyrus L pallidum L thalamus L SMA R occipital pole R lateral occipital cortex L lateral occipital cortex R frontal pole R insular cortex R pallidum R frontal orbital cortex R thalamus SRI L paracingulate gyrus L SMA R paracingulate gyrus L precentral gyrus L postcentral gyrus R lateral occipital cortex R occipital pole L occipital pole L lateral occipital cortex L occipital fusiform gyrus CG R occipital pole L occipital pole L inferior frontal gyrus L precentral gyrus L postcentral gyrus L paracingulate gyrus R paracingulate gyrus L SMA L superior frontal gyrus
14932
11941
3279
941
5302
2481
2140
15185
11960
1847
5.04 5.03 4.99 4.88 4.73 5.52 4.84 4.91 3.74 3.27 4.82 4.17 3.79 3.71 3.64 4.17
4.27 4.24 4.15 4.12 4.05 3.89 4.62 4.38 4.33 4.39 4.18 4.18 3.89
6.41 5.36 6.29 5.51 5.55
5 4.91 4.51 4.69 4.13 4.04 4.05 3.75 3.27
-50 -38 -20 -10 -6 16 16 42 26 26 -36 44 31 18 36 22
-4 -2 -6 6
-34 -56 34 32 18 -20 -44 -30 -26
26 16 -34 -16 -42 -34 -44 -52 -6 2 4 0 -4 -2
6
-16 2
-18 -4 -92 -98 -76 -66 -62 -84 38 20 -2 30 -30
18 4 -2 18 -8 -22 -82 -86 -98 -104 -80 -80 -88
-94 -96 -94 -96 8
-24 -16 -24 18 28 22 4 4 22
42 64 2 4 54 -4 -8 -8 50 42 -8 16 2 4
-10 -4
40 56 52 44 62 46 -8 -16 -4 -8 -14 -16 -16
-6 -2 -12 -8 22 48 54 52 44 38 40 54 66 58
Explanation of Abbreviations: R = right; L = left
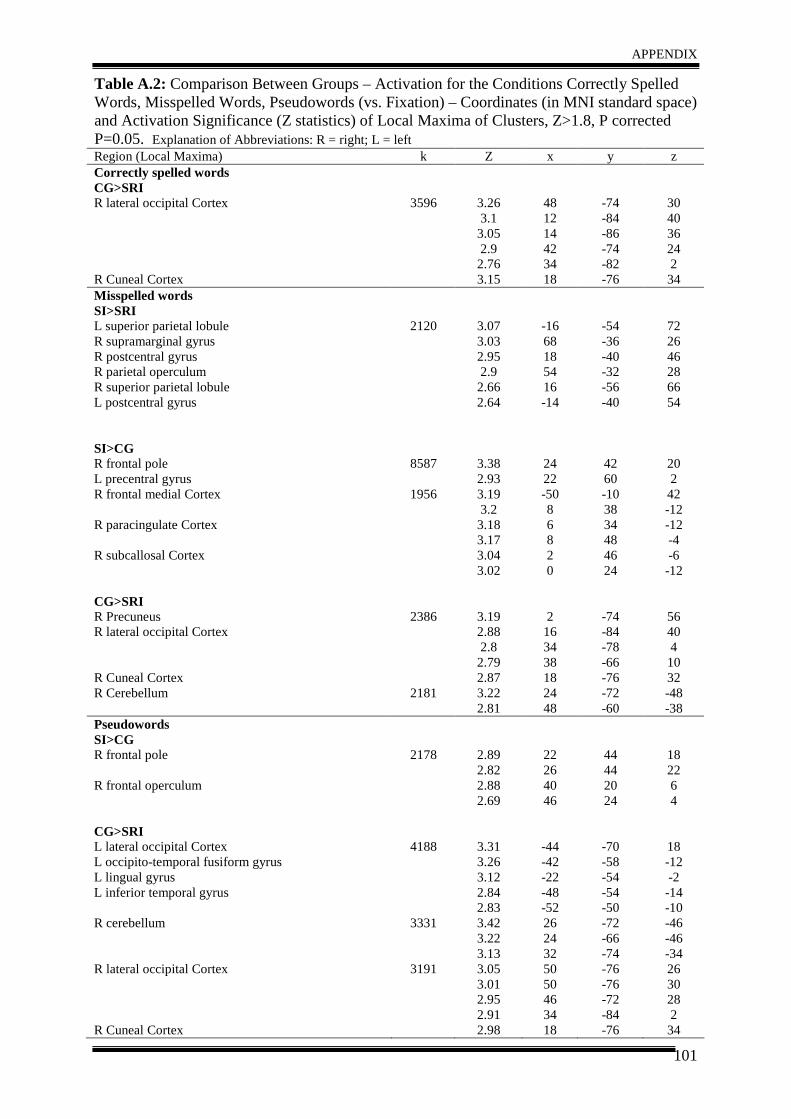
APPENDIX
101
Table A.2: Comparison Between Groups – Activation for the Conditions Correctly Spelled Words, Misspelled Words, Pseudowords (vs. Fixation) – Coordinates (in MNI standard space) and Activation Significance (Z statistics) of Local Maxima of Clusters, Z>1.8, P corrected P=0.05. Explanation of Abbreviations: R = right; L = left Region (Local Maxima) k Z x y z Correctly spelled words CG>SRI R lateral occipital Cortex R Cuneal Cortex
3596
3.26 3.1 3.05 2.9 2.76 3.15
48 12 14 42 34 18
-74 -84 -86 -74 -82 -76
30 40 36 24 2 34
Misspelled words SI>SRI L superior parietal lobule R supramarginal gyrus R postcentral gyrus R parietal operculum R superior parietal lobule L postcentral gyrus SI>CG R frontal pole L precentral gyrus R frontal medial Cortex R paracingulate Cortex R subcallosal Cortex CG>SRI R Precuneus R lateral occipital Cortex R Cuneal Cortex R Cerebellum
2120
8587
1956
2386
2181
3.07 3.03 2.95 2.9 2.66 2.64
3.38 2.93 3.19 3.2 3.18 3.17 3.04 3.02
3.19 2.88 2.8 2.79 2.87 3.22 2.81
-16 68 18 54 16 -14
24 22 -50 8 6 8 2 0 2 16 34 38 18 24 48
-54 -36 -40 -32 -56 -40
42 60 -10 38 34 48 46 24
-74 -84 -78 -66 -76 -72 -60
72 26 46 28 66 54
20 2 42 -12 -12 -4 -6 -12
56 40 4 10 32 -48 -38
Pseudowords SI>CG R frontal pole R frontal operculum CG>SRI L lateral occipital Cortex L occipito-temporal fusiform gyrus L lingual gyrus L inferior temporal gyrus R cerebellum R lateral occipital Cortex R Cuneal Cortex
2178
4188
3331
3191
2.89 2.82 2.88 2.69
3.31 3.26 3.12 2.84 2.83 3.42 3.22 3.13 3.05 3.01 2.95 2.91 2.98
22 26 40 46
-44 -42 -22 -48 -52 26 24 32 50 50 46 34 18
44 44 20 24
-70 -58 -54 -54 -50 -72 -66 -74 -76 -76 -72 -84 -76
18 22 6 4
18 -12 -2 -14 -10 -46 -46 -34 26 30 28 2 34
APPENDIX
102
Fig. A.1: FSL design matrix for the functional MRI analyses.
First-Level Analysis: Corr = correctly spelled words; miss = misspelled words; pseudo = pseudowords. Motion parameters were included as covariable of no interest.
Higher-Level Analysis: Glm for mean activation and contrast between groups. SI = Spelling impaired children; SRI = children with spelling and reading impairments; CG = control group.
Fig. A.2: Corona (y = -17.00) and transversal (z = 19.00) view of the FA skeleton (green) on top of the average standard FMRIB58_FA map (A) compared to the FA skeleton on top of the mean FA map of our sample (B). R
= right, L = left, S = superior, I = inferior, A = anterior, P = posterior.
APPENDIX
103
Appendix Study II Table A.1: Local maxima and cluster size at baseline (TFCE, p< 0.95). Region (Local Maxima) k x y z Fractional Anisotropy (FA) CG>Spelling Impaired Sample R Anterior thalamic radiation
7924
84
114
78
Mean Diffusity (MD) Spelling Impaired Sample>CG R PLIC
10873
67
112
76
Radial diffusity Spelling Impaired Sample>CG R PLIC
13771
67
110
74
Explanation of Abbreviations: TG = Training Group; CG = Control Group; WG = Waiting Group IFO = Inferior occipito-frontal fasciculus; SLF = superior longidutinal fasciculus; PLIC = posterior limb of internal capsule; SCR = superior corona radiata; ILF = inferior longitudinal fasciculus Table A.2: Significant ROI at Baseline. Comparison of mean FA, MD, axial and radial diffusivity indices across an a priori defined ROI between the spelling impaired sample and controls. Spelling Impaired Sample Control Group p Right PLIC FA MD Axial D. (mm2.s-1 ) Radial D. (mm2.s-1 )
.4838 .00054 .00104 .00029
.5079 .00051 .00101 .00026
.005 .000 .023 .000
Left PLIC FA Radial D. (mm2.s-1 )
.4989 .00030
.5176 .00028
.026 .022
Right SCR FA MD Axial D. (mm2.s-1 ) Radial D. (mm2.s-1 )
.2130 .00033 .00053 .00024
.2233 .00031 .00050 .00021
.032 .002 .004 .003
Right SLF Radial D. (mm2.s-1 )
.00037
.00034
.049
R PLIC (FA: F(1,26) = 9.25; p < .05; ηp² =.26; MD: F(1,26) = 21.03; p < .001; ηp² =.45; L1: F(1,26) = 5.83; p < .05; ηp² =.18; L23: F(2,25) = 17.30; p < .001; ηp² =.40) R SCR (FA: F(1,26) = 5.12; p < .05; ηp² =.16; MD: F(1,26) = 12.29; p < .05; ηp² =.32; L1: F(1,26) = 9.72; p < .05; ηp² =.27; L23: F(1,26) = 10.45; p < .05; ηp² =.29) R SLF (L23: F(1,26) = 4.27; p = .05; ηp² =.14) L PLIC (FA: F(1,26) = 5.59; p < .05; ηp² =.18; L23: F(1,26) = 5.98; p < .05; ηp² =.19) Explanation of Abbreviations: TG = Training Group; CG = Control Group; WG = Waiting Group FA = Fractional Anisotropy; MD = Mean diffusivity; PLIC = posterior limb of internal capsule; SCR = superior corona radiata; SLF = superior longitudinal fasciculus Axial diffusivity and radial diffusivity values are expressed in mm2.s-1
Table A.3: Local maxima and cluster size for training effect (TFCE, p< 0.95). Region (Local Maxima) k x y z
APPENDIX
104
FA F-test across the three groups R PLIC
18 9
70 67
118 113
78 81
TG>CG R IFO
5496
56
130
72
WG>CG R PLIC
7
71
118
77
MD F-test across the three groups R SCR
3619
63
125
94
CG>TG R PLIC
11993
66
116
84
Radial diffusity F-test across the three groups R SCR R IFO
4087
2
63 56
126 107
93 63
CG>TG R SCR
15416
66
124
91
Explanation of Abbreviations: TG = Training Group; CG = Control Group; WG = Waiting Group PLIC = posterior limb of internal capsule; IFO = Inferior occipito-frontal fasciculus; SCR = superior corona radiata
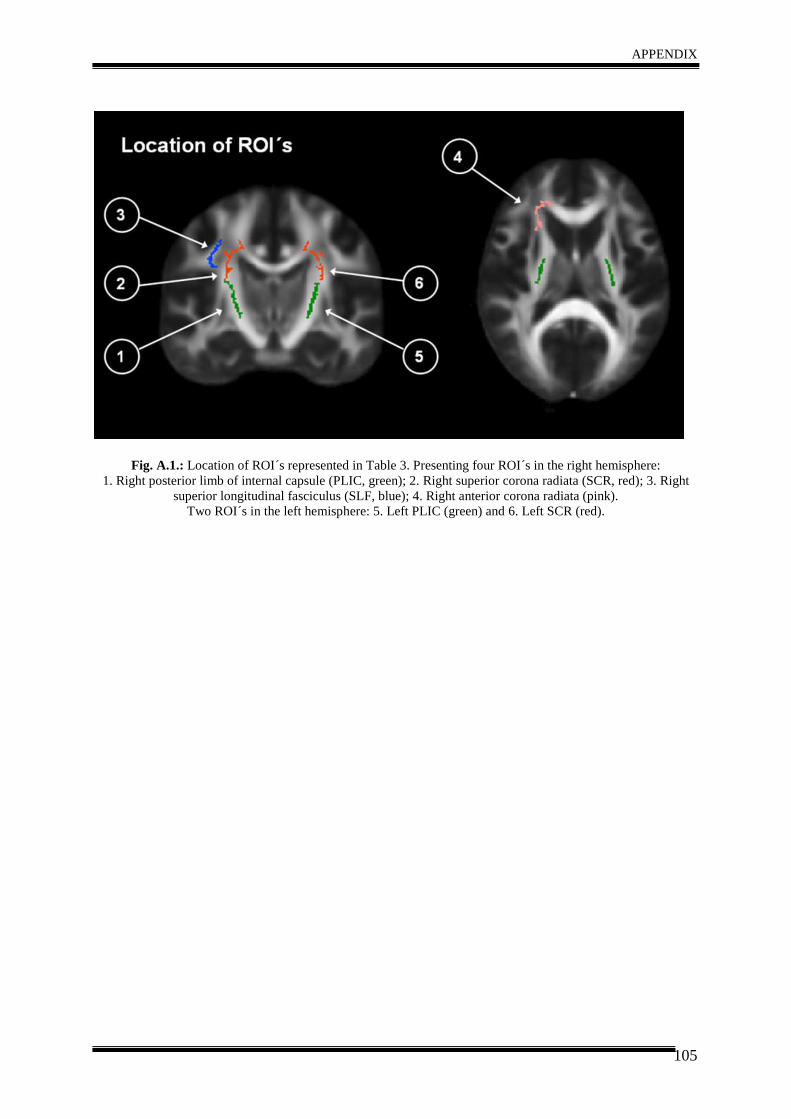
APPENDIX
105
Fig. A.1.: Location of ROI´s represented in Table 3. Presenting four ROI´s in the right hemisphere: 1. Right posterior limb of internal capsule (PLIC, green); 2. Right superior corona radiata (SCR, red); 3. Right
superior longitudinal fasciculus (SLF, blue); 4. Right anterior corona radiata (pink). Two ROI´s in the left hemisphere: 5. Left PLIC (green) and 6. Left SCR (red).

106
VII LIST OF ABBREVIATIONS (in alphabetical order) ACR – Anterior Corona Radiata
ALE – Activation Likelihood Estimation
ANOVA – Analyses of Variance
APA – American Psychological Association
BET – Brain Extraction Tool
C – Correctly Spelled Words
CC – Corpus Callosum
CG – Control Group
DSM IV – Diagnostic and Statistical Manual of Mental Disorders
DTI – Diffusion Tensor Imaging
EEG – Electroencephalogram
ELFE - Ein Leseverständnistest für Erst- bis Sechstklässler
EPI – Echo Planar Imaging
FA – Fractional Anisotropy
FDT – FMRIB´s Diffusion Toolbox
FEAT – FMRI Expert Analysis Tool
FILM – FMRIB´s Improved Linear Model
FLAME – FMRIB´s Local Analysis of Mixed Effects
FLIRT – FMRIB´s Linear Image Registration Tool
FMRIB – Oxford Centre of Functional MRI of the Brain
FNIRT – FMRIB´s Nonlinear Image Registration Tool
fMRI – functional Magnetic Resonance Imaging
FOV – Field of View
HSP – Hamburger Schreibprobe
ICD 10 – International Classification of Diseases 10
IFG – Inferior Frontal Gyrus
IFO – Inferior Fronto-Occipital Fasciculus
ILF – Inferior Longitudinal Fasciculus
K – Number of Voxels
L – Left
M – Misspelled Words
MD – Mean Diffusivity

107
MNI – Montreal Neurological Institute
MPRAGE – Magnetization Prepared Rapid Gradient Echo
MRI – Magnetic Resonance Imaging
OT – Occipito-Temporal
P – Pseudowords
PASW – Predictive Analytics Software
PLIC – Posterior Limb of Internal Capsule
PR – Percent Range
R – Right
ROI – Region of Interest
RT – Reaction Time
SCR – Superior Corona Radiata
SFG – Superior Frontal Gyrus
SI – Isolated Spelling Impairment
SLF – Superior Longitudinal Fasciculus
SLS – Salzburger Lese-Screening
SMG – Supramarginal Gyrus
SPL – Superior Parietal Lobe
SPM – Standard Progressive Matrices
SRI – Spelling and Reading Impairment
TBSS – Tract Based Spatial Statistics
TE – Time to Echo
TFCE – Threshold-Free Cluster Enhancement
TG – Training Group
TR – Time to Repeat
VWFA – Visual Word Form Area
WG – Waiting Group

108
STATEMENT OF AUTHORSHIP
I hereby certify that this dissertation has been composed by myself, and describes my own
work, unless stated otherwise. All references and verbatim have been quoted, and all sources
of information have been specifically acknowledged.
Date Signature





























