Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Investigating a crow die-off in January–February 2011during the introduction of a new clade of highly pathogenic avianinfluenza virus H5N1 into Bangladesh
Salah Uddin Khan • LaShondra Berman • Najmul Haider • Nancy Gerloff • Md Z. Rahman •
Bo Shu • Mustafizur Rahman • Tapan Kumar Dey • Todd C. Davis • Bidhan Chandra Das •
Amanda Balish • Ausraful Islam • Jens P. Teifke • Nord Zeidner • Steven Lindstrom •
Alexander Klimov • Ruben O. Donis • Stephen P. Luby • H. L. Shivaprasad • Andrea B. Mikolon
Received: 15 April 2013 / Accepted: 28 July 2013 / Published online: 1 October 2013
� Springer-Verlag Wien 2013
Abstract We investigated unusual crow mortality in
Bangladesh during January-February 2011 at two sites.
Crows of two species, Corvus splendens and C. macro-
rhynchos, were found sick and dead during the outbreaks.
In selected crow roosts, morbidity was *1 % and mor-
tality was *4 % during the investigation. Highly patho-
genic avian influenza virus H5N1 clade 2.3.2.1 was
isolated from dead crows. All isolates were closely related
to A/duck/India/02CA10/2011 (H5N1) with 99.8 % and
A/crow/Bangladesh/11rs1984-15/2011 (H5N1) virus with
99 % nucleotide sequence identity in their HA genes. The
phylogenetic cluster of Bangladesh viruses suggested a
common ancestor with viruses found in poultry from India,
Myanmar and Nepal. Histopathological changes and
immunohistochemistry staining in brain, pancreas, liver,
heart, kidney, bursa of Fabricius, rectum, and cloaca were
consistent with influenza virus infection. Through our
limited investigation in domesticated birds near the crow
roosts, we did not identify any samples that tested positive
for influenza virus A/H5N1. However, environmental
samples collected from live-bird markets near an outbreak
site during the month of the outbreaks tested very weakly
positive for influenza virus A/H5N1 in clade 2.3.2.1-spe-
cific rRT-PCR. Continuation of surveillance in wild and
domestic birds may identify evolution of new avian influ-
enza virus and associated public-health risks.
Introduction
Bangladesh has reported highly pathogenic avian influenza
virus A (HPAI) H5N1 in domestic poultry since February
2007 [1]. From 2007 through 2011, 514 outbreaks were
reported in 51 out of 64 districts [2]. The hemagglutinin (HA)
of all H5N1 influenza A viruses isolated from the 2007-2009
outbreaks belonged to clade 2.2.2, with very limited (\5 %)
divergence in their genetic makeup [3]. HPAI H5N1 was not
reported in wild birds during this period.
S. U. Khan (&) � N. Haider � M. Z. Rahman � M. Rahman �A. Islam � N. Zeidner � S. P. Luby � A. B. Mikolon
CCD, International Centre for Diarrhoeal Disease Research,
Bangladesh (ICDDR, B), Dhaka, Bangladesh
e-mail: [email protected]; [email protected]
URL: www.icddrb.org
S. U. Khan
Department of Environmental and Global Health, College of
Public Health and Health Professions, University of Florida,
Gainesville, FL, USA
L. Berman � N. Gerloff � B. Shu � T. C. Davis � A. Balish �N. Zeidner � S. Lindstrom � A. Klimov �R. O. Donis � S. P. Luby � A. B. Mikolon
Centers for Disease Control and Prevention, (CDC), Atlanta,
GA, USA
T. K. Dey
Forest Department, Ministry of Environment and Forest, Dhaka,
Bangladesh
B. C. Das
Department of Livestock Services, Ministry of Fisheries and
Livestock, Dhaka, Bangladesh
J. P. Teifke
Friedrich-Loeffler-Institut (FLI), Federal Research Institute for
Animal Health, Greifswald-Insel Riems, Germany
H. L. Shivaprasad
California Animal Health and Food Safety Laboratory System,
Tulare branch, University of California, Davis,
Tulare, CA, USA
123
Arch Virol (2014) 159:509–518
DOI 10.1007/s00705-013-1842-0

In January-February 2011, Bangladesh reported wide-
spread outbreaks of HPAI A/H5N1 in domestic poultry [4].
During the same period, the Forest Department and the
Department of Livestock Services of Bangladesh received
multiple reports of crow die-offs from at least two
administrative divisions of the country (personal commu-
nication, Department of Livestock Services, Bangladesh)
[5]. As a part of outbreak response, members from the
Department of Livestock Services, Forest Department of
Bangladesh, International Centre for Diarrhoeal Disease
Research, Bangladesh (icddr,b), and Centers for Disease
Control and Prevention (CDC), Atlanta, conducted field
and laboratory investigations. We report here the results of
outbreak investigations conducted to determine the cause
of crow die-offs and subsequent studies to determine the
source of H5N1 infection.
Materials and methods
Outbreak settings
We investigated die-offs in two crow roosts from 17 to 23
January 2011 in Patuakhali (N22.35944 E90.34912), a
small town in southern Bangladesh, and from 13 to 15
February 2011 in the capital city of Dhaka (N23.73989
E90.39829). The sites are 153 kilometers apart. In Patu-
akhali, the crows roosted in orchards (approximately 200
trees) surrounded by dense human settlement, whereas in
Dhaka, a city with a population density of 35,000/square
kilometer, the crows roosted in a park (approximately
3000-5000 trees) [6].
Data and sample collection procedures
The field team consisted of veterinarians and veterinary
technicians from the Department of Livestock Services,
Forest Department, and icddr,b. We used the total count
method to estimate the number of healthy, sick, and dead
crows in both roosts at two time points [7]. We recorded
detailed clinical presentations of a subset of sick crows
found at the Patuakhali outbreak site three times a day for
two consecutive days. All of the sick crows that we
included in the clinical observation had fallen from their
roosts and were unable to fly. These crows were a subset of
the ‘‘sick crows’’ that were identified during the investi-
gation. The field investigation team kept the sick crows in
an enclosed place under the crow roost, providing adequate
food and water. However, the sick crows were free to roam
within the orchard and none of them were sacrificed. The
orchard where the crows roosted was in the backyard of a
household fenced by a brick wall that made the place
inaccessible to the neighbors. The household members
rarely entered their backyard. We also interviewed the
people whose dwellings were under the crow roost in Pa-
tuakhali to estimate frequencies of crow deaths and history
of poultry die-offs in their backyards during the outbreak.
We collected oropharyngeal and cloacal swab samples
from dead and sick crows found under the roosts and
pooled them in viral transport medium, one pool per crow.
We kept two carcasses of crows that had recently died for
necropsy and histopathology. We also collected cloacal
swab samples from three domestic ducks raised by
household members living under the crow roost in Patu-
akhali. At the same time, we collected environmental swab
samples from live-bird markets located nearest to the out-
break sites to investigate the spread of the virus to domestic
poultry: one market in Patuakhali in January 2011 and two
in Dhaka during February 2011. These markets were within
three kilometers of the outbreak sites and sold about 200 to
2000 birds per day, including native chickens, commer-
cially raised chickens, ducks, geese, and pigeons. Sampling
at each of the markets included swabs (n=10) from poultry
cages, feed and water trays, and fecal droppings from
poultry stalls. Swabs were pooled into one environmental
sample per market.
We conducted a follow-up investigation in wild birds
around the Patuakhali crow roost on 7-8 February 2011,
two weeks after the primary investigation. The team col-
lected oropharyngeal and cloacal swabs from five crows
lying on the ground under the roost and unable to fly. We
caught wild birds around the crow roost using mist nets in
their flyways [8], banded them, and released them at the
same location where they were captured, after collecting
oropharyngeal and cloacal swabs.
During clinical observation and sampling, the team used
appropriate personal protective equipment, including
gloves, goggles, and face masks, and they made regular use
of disinfection.
Virus isolation, sequencing, and phylogeny
We performed real-time reverse transcription polymerase
chain reaction (rRT-PCR) to identify the influenza A virus
targeting the matrix (M) gene and the HA for subtyping and
clade-specific rRT-PCR as described previously [9]. Virus
culture and isolation in eggs, RNA extraction and sequencing
procedures were performed as described previously [10]. We
performed phylogenetic analysis as described elsewhere [11].
Briefly, complete HA sequences were aligned to a subset of
reference sequences representing currently circulating South
East Asian H5 clades using BioEdit [12], and a neighbor-
joining phylogenetic tree with 1000 bootstrap replicates was
calculated with MEGA version 4 [13]. HA sequences were
submitted to GISAID (http://platform.gisaid.org) with
accession numbers: EPI448167, EPI448159, EPI448151,
510 S. U. Khan et al.
123

EPI448063, EPI448055, EPI448047, EPI465423,
EPI448031, EPI353381, and EPI353364.
Histopathology
We performed histopathology and immunohistochemistry
on two bird samples collected during the outbreak at the
Patuakhali site. Tissue samples were collected and fixed in
10 % buffered neutral formalin. The tissues were trimmed,
processed, embedded in paraffin, sectioned at 4 microme-
ters, stained with haematoxylin and eosin (H&E), and
examined by bright field microscopy.
Immunohistochemistry (IHC)
To study the distribution of nucleoprotein (NP) of AI virus
in various organs and cells, IHC was performed on selected
tissues according to a method described elsewhere [14].
Briefly, paraffin sections were mounted on positively
charged Super Frost Plus microscopic slides, dewaxed, and
rehydrated. The sections were pretreated with 10 mM citric
acid, pH 6.0, for 10 min at 110 �C in a decloaking chamber
(Biocare Medical, Concord, CA) and subsequently incu-
bated with the rabbit anti-NP serum, diluted 1:750 in Tris-
buffered saline. Tissue sections were stained using the
avidin-biotin-peroxidase complex method, with 3-amino-9-
ethylcarbazole (red), and with hematoxylin (blue) accord-
ing to the method by Klopfleisch et al. [14].
Ethical considerations
This investigation was part of an outbreak response to a
possible public-health threat and was not considered
human subject research. However, before initiating field
investigation and sampling, the team obtained informed
consent from the property owners where the crows roosted,
owners of the individual birds that were sampled, and the
live-bird market authorities where we performed environ-
mental sampling. We collected all of the dead crows found
under the roost in biohazard bags during the investigation
and incinerated them at icddr,b or at the local livestock
hospital facility. During the investigation, we requested the
inhabitants close to the crow roost not to come into contact
with dead or sick crows. The local livestock and forest
department officials regularly disinfected the surface
beneath the crow roost at both sites. The crow samples
were handled at biosafety level (BSL) II for initial
screening by rRT-PCT, and they were later transferred to
BSL III laboratory facilities for virus culture. The protocols
for sampling domestic and wild birds and collecting envi-
ronmental samples were approved by the Institutional
Review Boards of the International Centre for Diarrhoeal
Disease Research, Bangladesh.
Results
Morbidity and mortality in crows
The Patuakhali outbreak occurred at a crow roost with an
estimated population of 1,500 birds. Seventeen (1 %) of
the birds showed clinical signs of illness, and 57 (4 %)
were found dead during the one-week investigation. The
Patuakhali residents dwelling under tree canopies with
roosting crows first noticed unusually high crow mortality
in the first week of January 2011. The crow deaths con-
tinued for approximately one month. The Dhaka city out-
break involved about 900 crows that roosted in the trees of
a city park. Of these, five (0.6 %) appeared sick, and 39
(4 %) were found dead during the investigation. In the
Dhaka city outbreak site, crow deaths were first reported
during the first week of February and continued until mid-
March 2011. We identified two species of crows, house
crows (Corvus splendens) and large-billed crows (Corvus
macrorhynchos); once the crows became ill, they were
found dead within 24 hours.
Clinical features in crows
In Patuakhali, the seven sick crows we observed in cap-
tivity for two days were unable to fly, appeared drowsy,
had dilated pupils, and had whitish fecal material attached
to the plumage surrounding the cloaca. The birds showed
neurologic signs, including uncoordinated gait, tremors,
pronounced opisthotonos, and torticollis at the terminal
stage (Fig. 1). We observed two (29 %) crows with dis-
charges from the nares during the later stage of clinical
illness. The field team observed cannibalism and carcass
scavenging among the crows (Fig. 1). The average cloacal
temperature of the sick or moribund crows was 100.3 �F
(range: 94 �F to 107 �F) (37.94 �C; range: 34.4–41.7 �C).
Phylogeny of H5N1viruses from crows
We collected 31 pooled oropharyngeal-cloacal swabs: 11
from C. splendens and 20 from C. macrorhynchos from the
two outbreak sites. All tested positive for avian influenza
virus A/H5 by rRT-PCR. Virus was successfully isolated
from 8 out of 31 samples (C. splendens. n=4, and C.
macrorhynchos, n=4), including samples from Patuakhali
(n=4) and from Dhaka (n=4). Sequence analysis of the HA
and NA genes indicated that the eight isolates were influ-
enza virus A/H5N1, and the HA contained the multibasic
cleavage site motif (PQRERRRKR*G) characteristic of
HPAI. Preliminary phylogenetic analysis showed that these
viruses were almost identical to each other in their HA
gene sequences and were most closely related to viruses
with clade 2.3.2.1 HA genes (Fig. 2). All isolates were
Highly pathogenic avian H5N1 influenza virus in Bangladesh 511
123

closely related to A/duck/India/02CA10/2011 (H5N1) and
A/crow/Bangladesh/11rs1984-15/2011(H5N1) virus, with
99.8 % and 99 % nucleotide sequence identity, respec-
tively, in their HA genes [15, 16]. Although virus culture
for the remaining samples was not successful, HA and NA
genes were sequenced using RNA extracted from the
specimen, and phylogenetic analysis revealed that these
viruses also belonged to clade 2.3.2.1.
Dispersion of H5N1 to the domestic poultry and poultry
marketing chain
We identified 12 households with 80 backyard poultry
living under the crow roost in Patuakhali. The household
members reported that 43 (54 %) of their poultry became
sick and 28 (35 %) died during 12 to 22 January 2011.
However, during our investigation, the backyard poultry in
those households did not show clinical signs of illness, and
none were found dead. Three ducks were available during
the investigation, and cloacal samples from all of them
tested negative for influenza virus A by rRT-PCR. The
epidemiologic investigation did not identify additional
domestic or wild birds with clinical signs of disease in
areas surrounding crow roosts in Patuakhali or Dhaka
during the outbreak investigations.
Environmental swab samples collected from one live-
bird market from Patuakhali city during the month of the
outbreaks tested very weakly positive for influenza virus
A/H5N1 by clade-2.3.2.1-specific rRT-PCR. Partial HA
gene sequence data from the two Dhaka market samples
also indicated closest relationship to clade 2.2.2; however,
we failed to isolate virus from any of the environmental
samples from the Patuakhali and Dhaka live-bird market
samples.
Follow-up investigation in wild birds in Patuakhali
During the follow-up investigation on 7-8 February 2011 in
Patuakhali, the team visited the outbreak site. They cap-
tured two large-billed crows (C. macrorhynchos) and three
house crows (C. splendens) under the crow roost. These
birds appeared sick and were unable to fly; we detected
influenza virus A/H5N1 clade 2.3.2.1 from oropharyngeal
and cloacal swabs by rRT-PCR. Additionally, the team
captured 30 wild birds of five different species near the
crow roost: house crows (C. splendens, n=2), pintail snipes
(Gallinago stenura, n=7), greater painted snipes (Rostra-
tula benghalensis, n=12), Asian pied starling (Sturnus
contra, n=8), and a barn owl (Tyto alba, n=1). Of birds,
only the barn owl was apparently sick and unable to fly.
Oropharyngeal and cloacal swabs samples from all of these
apparently healthy birds and the barn owl tested negative
for influenza virus A by rRT-PCR.
Histopathology
Lesions in various organs of both birds were similar, with
minor variation between the two. In the brain, significant
lesions were confined to the cerebellum, brain stem and
medulla oblongata. Multifocal mild to severe enlargement,
degeneration and necrosis (characterized by red neurons)
and central chromatolysis of neurons including Purkinje
cells was observed. Some of the nuclei within the neurons
appeared enlarged and red, with homogenous composition
(Fig. 3a). Both cerebrum and cerebellum had mild multi-
focal increased glial cells, satellitosis, and neuronophagia
in the grey matter. Occasional perivascular cuffing and
mild hemorrhages scattered within the neuropil were
observed. The meninges were occasionally thickened due
to infiltration of lymphocytes.
Fig. 1 Sick and dead crows found under the roost in Patuakhali and
Dhaka city during January-February 2011. a A sick crow with ataxic
limbs found in the Dhaka city park, February 2011. b An apparently
healthy crow pecking on a sick crow under the roost in the Dhaka city
park, February 2011
512 S. U. Khan et al.
123

In the pancreas, severe multifocal foci of acute coagu-
lative necrosis of acinar cells were noted, with minimal
inflammation (Fig. 3b). The liver had acute mild multifocal
coagulative necrosis of hepatocytes accompanied by mild
infiltration of lymphocytes and macrophages (Fig. 3c). The
spleen had an increased number of mononuclear cells,
including plasma cells, as well as mild hemosiderosis.
Kidneys had mild multifocal acute necrosis of tubular
epithelial cells with no associated inflammation. The heart
had mild multifocal degeneration of myofibers with no
inflammation. An occasional segment of intestine demon-
strated mild necrosis of lymphocytes within Peyer’s
patches.
In one bird, the rectum/cloaca had increased cellularity
of the lamina propria, and the bursa of Fabricius had
changes ranging from cystic follicles to severe lymphoid
depletion.
Other organs, including the eyes, lungs, trachea, phar-
ynx, proventriculus, gizzard, intestine, gall bladder, testes
and skeletal muscles, did not have any significant lesions.
Immunohistochemistry (IHC)
IHC directed against the nucleoprotein (NP) of avian
influenza virus stained both the nucleus and cytoplasm in
various tests, and the results are summarized in Table 1.
Fig. 2 Phylogeny of HA gene
sequences of H5N1 (HPAI)
viruses, with the Bangladesh
viruses indicated in boldface.
Viruses that circulate in other
regions of South East Asia are
included and shown as
collapsed branches (triangles)
labeled with clade names (e.g.,
2.2.2). Clade 2.3.2.1 and the
lineages are indicated behind
the bracket. The tree was
calculated using the neighbor-
joining method with the Kimura
2-parameter distance model.
Numbers above nodes indicate
bootstrap values ([70 %) and
the tree was rooted at the
midpoint
Highly pathogenic avian H5N1 influenza virus in Bangladesh 513
123
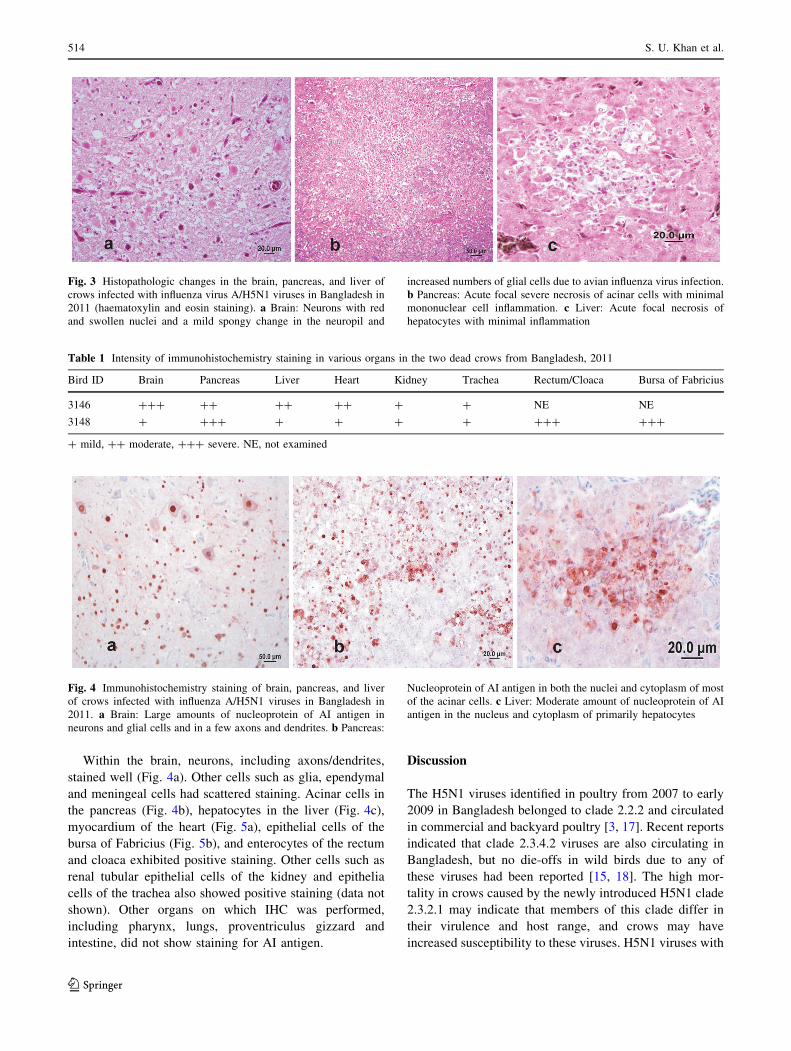
Within the brain, neurons, including axons/dendrites,
stained well (Fig. 4a). Other cells such as glia, ependymal
and meningeal cells had scattered staining. Acinar cells in
the pancreas (Fig. 4b), hepatocytes in the liver (Fig. 4c),
myocardium of the heart (Fig. 5a), epithelial cells of the
bursa of Fabricius (Fig. 5b), and enterocytes of the rectum
and cloaca exhibited positive staining. Other cells such as
renal tubular epithelial cells of the kidney and epithelia
cells of the trachea also showed positive staining (data not
shown). Other organs on which IHC was performed,
including pharynx, lungs, proventriculus gizzard and
intestine, did not show staining for AI antigen.
Discussion
The H5N1 viruses identified in poultry from 2007 to early
2009 in Bangladesh belonged to clade 2.2.2 and circulated
in commercial and backyard poultry [3, 17]. Recent reports
indicated that clade 2.3.4.2 viruses are also circulating in
Bangladesh, but no die-offs in wild birds due to any of
these viruses had been reported [15, 18]. The high mor-
tality in crows caused by the newly introduced H5N1 clade
2.3.2.1 may indicate that members of this clade differ in
their virulence and host range, and crows may have
increased susceptibility to these viruses. H5N1 viruses with
Fig. 3 Histopathologic changes in the brain, pancreas, and liver of
crows infected with influenza virus A/H5N1 viruses in Bangladesh in
2011 (haematoxylin and eosin staining). a Brain: Neurons with red
and swollen nuclei and a mild spongy change in the neuropil and
increased numbers of glial cells due to avian influenza virus infection.
b Pancreas: Acute focal severe necrosis of acinar cells with minimal
mononuclear cell inflammation. c Liver: Acute focal necrosis of
hepatocytes with minimal inflammation
Table 1 Intensity of immunohistochemistry staining in various organs in the two dead crows from Bangladesh, 2011
Bird ID Brain Pancreas Liver Heart Kidney Trachea Rectum/Cloaca Bursa of Fabricius
3146 ??? ?? ?? ?? ? ? NE NE
3148 ? ??? ? ? ? ? ??? ???
? mild, ?? moderate, ??? severe. NE, not examined
Fig. 4 Immunohistochemistry staining of brain, pancreas, and liver
of crows infected with influenza A/H5N1 viruses in Bangladesh in
2011. a Brain: Large amounts of nucleoprotein of AI antigen in
neurons and glial cells and in a few axons and dendrites. b Pancreas:
Nucleoprotein of AI antigen in both the nuclei and cytoplasm of most
of the acinar cells. c Liver: Moderate amount of nucleoprotein of AI
antigen in the nucleus and cytoplasm of primarily hepatocytes
514 S. U. Khan et al.
123

clade 2.3.2.1 HA were first detected in a common magpie
in Hong Kong SAR in 2007 [19, 20]. Since then, and
especially in the past 3 years, viruses from this clade have
spread to Asia and Europe, including China, Japan, Mon-
golia, Vietnam, the Republic of Korea, Myanmar, India,
Nepal, the Russian Federation, Bulgaria, and Romania [16,
19–26]. Although the majority of clade 2.3.2.1 viruses that
were reported were in domestic poultry, seven out of ele-
ven countries reported this clade in wild birds [3]. In 2010,
Nepal was the first country in the South Asian region to
identify this clade in domestic poultry [18]. Although the
H5N1 clade 2.3.2.1 viruses were found to cause outbreaks
in India’s poultry population about a month after the out-
break in Bangladesh, the viruses were almost identical
[16]. This indicates the continuing emergence of this par-
ticular clade of H5N1 virus, causing rapid spread from one
location to another, as seen in other parts of the world [23].
Particularly in Southeast Asia, national and international
poultry trade and migratory wild birds are commonly
implicated in cross-border transmission of the virus, which
reemphasizes the need for continuation of wild and
domestic bird surveillance for influenza viruses [27].
Although there are several reports of HPAI H5N1
pathology in poultry, there has been only one report on
crows (Corvus macrorhynchos) [28]. Our findings suggest
that the lesions in the brain, pancreas, liver and other
organs of the crows were caused by HPAI virus, as con-
firmed by immunohistochemistry. Moreover, there was
good correlation between histopathology and IHC. Lesions
in the brain and pancreas were quite severe, and mild to
moderate in other organs. These lesions would explain the
deaths of the two crows. Both the histopathologic lesions
Fig. 5 Immunohistochemistry
staining of heart and bursa of
Fabricius of crows infected with
influenza A/H5N1 viruses in
Bangladesh in 2011. a Heart:
Large amounts of nucleoprotein
of AI antigen, primarily in the
sarcoplasm and in some nuclei
of cardiac myocytes. b Bursa of
Fabricius: Abundant
nucleoprotein of AI antigen in
the nucleus and cytoplasm of
epithelial cells
Highly pathogenic avian H5N1 influenza virus in Bangladesh 515
123

and IHC seen in various organs of these crows in our study
are in good agreement with the report of Tanimura et al.
[28].
Viruses from clade 2.3.2.1 diverged into three distinct
groups, referred to as Hubei-like (A/Hubei/1/2010),
including strains circulating in South East Asian countries
such as Vietnam, Hong Kong/6841-like (A/Hong Kong/
6841/2010) strains detected in Japan, Korea, Russia and
primarily other Far Eastern countries, and barn swallow-
like (A/barn swallow/Hong Kong/1161/2010) strains,
including recent viruses from Hong Kong and Vietnam
(Fig. 2) [18, 29]. The clade 2.3.2.1 viruses identified in
crows during investigations in Bangladesh were most clo-
sely related to the Hubei-like clade 2.3.2.1 viruses found in
India in 2011 [16], Nepal in 2011, and Myanmar in 2010
and 2011 [18, 20, 30]. Bangladesh viruses from the Hubei-
like cluster were highly similar in their HA sequences and
were derived from both poultry and crow outbreaks [18]
(Fig. 2). The phylogenetic cluster of Bangladesh viruses
with viruses from neighboring countries suggested a
common ancestor with Hubei-like viruses found in poultry
from India, Myanmar, and Nepal and in wild birds (crows)
in Nepal [14, 18, 20, 30].
The rapid spread of HPAI H5N1 clade 2.3.2.1 has been
associated with illness in both domestic and wild birds
[24]. Our findings also suggest that once a crow popula-
tion in a roost becomes infected, the virus continues to
circulate among crows for a period of months. The most
likely way for virus to be transmitted between the crows
could be cannibalism and close contact with the ill crows.
The mortality rate in crows was very high, in agreement
with previous reports [31]. During January-April 2011,
Bangladesh reported 159 outbreaks of H5N1 in domestic
poultry, mostly in commercial farms [4]. Live-bird mar-
kets are considered high-risk sites for harboring influenza
viruses, and the poultry marketing chain may lead to
spread to a wider geographic area in the country [32].
Although we could not prove that birds from live-bird
markets near the crow roosts were infected with the same
clade of influenza virus, the possibility of horizontal
transmission of the virus between infected crows and
poultry in the live-bird market cannot be ruled out. Since
we collected only 10 environmental swab samples from a
live-bird market, we may have missed circulating influ-
enza viruses in the poultry. However, using an identical
sampling technique in the live bird markets of Bangladesh
has proven sufficient to identify circulating influenza
viruses, including A/H5N1 [32]. Additionally, it is also
possible that the infected crows may spread the virus
locally while scavenging near backyard poultry produc-
tion facilities and the commercial farms. Crows may
scavenge within eight kilometers from their roosting place
[33]. Our results indicate that crows infected with H5N1
exhibited clinical signs and were unable to fly, and the
clinical outcome was death. The combination of these
factors may limit the ability of infected crows to spread
the A/H5N1 clade 2.3.2.1 virus to a short geographical
range when compared to the marketing chain of live-bird
markets.
We systematically investigated crow outbreaks in only
two locations in Bangladesh during January-February
2011. However, there were reports of several crow out-
breaks at other locations during the same time period that
we did not have the resources to investigate. Additionally,
we counted the number of sick and dead crows only in the
area under the crow roost; this approach may have under-
estimated the morbidity and mortality in the crow
population.
Since the 2.2.2, 2.3.2.1 and 2.3.4 clades share common
hosts and reservoirs, surveillance is important to identify
whether they will reassort, co-circulate and/or replace one
another [34]. Bangladesh continues to be at risk for intro-
duction and evolution of additional new influenza viruses.
Informing and educating the public may help avoid direct
contact with sick and dead wild birds and promote sanitary
disposal to limit infection of humans and poultry.
Acknowledgments This research study was funded by the United
States Centers for Disease Control and Prevention (Cooperative
Agreement 5U01CI000628). icddr,b acknowledges with gratitude the
commitment of CDC to its research efforts. The contents of this report
are solely the responsibility of the authors and do not necessarily
represent the official views of the CDC or the icddr,b. We also
acknowledge and appreciate the diligent technical assistance of
Department of Livestock and Forest Department field officers of
Bangladesh, the icddr,b field team, and the immunohistochemistry
team of Gabriele Czerwinski at FLI.
References
1. World Organization for Animal health (2007) Highly pathogenic
avian influenza, Bangladesh (Immediate notification)
2. OIE (2011) Highly pathogenic avian influenza, Bangladesh.
Follow-up report No. 1–33. World Organization for Animal
health
3. Loth L, Gilbert M, Osmani MG, Kalam AM, Xiao X (2010) Risk
factors and clusters of Highly Pathogenic Avian Influenza H5N1
outbreaks in Bangladesh. Prev Vet Med 96(1–2):104–113. doi:10.
1016/j.prevetmed.2010.05.013
4. OIE (2011) Highly pathogenic avian influenza, Bangladesh.
Follow-up report No. 26–32. World Organization for Animal
health
5. Star TD (2011) Crows dying, poultry culled: Fear of fresh round
of bird flu grips Barisal, Kishoreganj. http://www.thedailystar.
net/newDesign/print_news.php?nid=172776. Accessed 24 Mar
2011, 6 Feb 2008
6. BBS (2012) Bangladesh Bureau of Statistics. Population density of
Dhaka city, 2008. http://www.bbs.gov.bd/WebTestApplication/user
files/Image/SubjectMatterDataIndex/pk_book_09.pdf. Accessed 23
Feb 2012
516 S. U. Khan et al.
123

7. Behrouzi-Rad B (2010) Population Estimation and Breeding
Biology of the House Crow Corvus splendens on Kharg Island,
Persian Gulf. Podoces 5(2):87–94
8. Ralph CJ, Dunn EH (eds) (2004) Monitoring bird populations
using mist nets, Studies in Avian Biology, vol 29. Cooper
Ornithological Society, Lawrence
9. Kis Z, Jones J, Creanga A, Ferdinand K, Inui K, Gerloff N, Davis
CT, Nguyen T, Donis RO (2013) Real-time RT-PCR assay to
differentiate clades of H5N1 avian influenza viruses circulating in
Vietnam. J Virol Methods 193:452–458. doi:10.1016/j.jviromet.
2013.06.023
10. Szretter KJ, Balish AL, Katz JM (2006) Influenza: propagation,
quantification, and storage. Curr Protoc Microbiol Chapter 15:
Unit 15G 11. doi:10.1002/0471729256.mc15g01s3
11. Rivailler P, Perry IA, Jang Y, Davis CT, Chen LM, Dubovi EJ,
Donis RO (2010) Evolution of canine and equine influenza
(H3N8) viruses co-circulating between 2005 and 2008. Virology
408(1):71–79. doi:10.1016/j.virol.2010.08.022
12. Hall TA (1999) BioEdit: a user-friendly biological sequence
alignment editor and analysis program for Windows 95/98/NT.
Nucl Acids Symp Ser 41:95–98
13. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molec-
ular Evolutionary Genetics Analysis (MEGA) software version
4.0. Mol Biol Evol 24(8):1596–1599. doi:10.1093/molbev/
msm092
14. Klopfleisch R, Werner O, Mundt E, Harder T, Teifke JP (2006)
Neurotropism of highly pathogenic avian influenza virus
A/chicken/Indonesia/2003 (H5N1) in experimentally infected
pigeons (Columbia livia f. domestica). Vet Pathol 43(4):463–470
15. Islam MR, Haque ME, Giasuddin M, Chowdhury EH, Samad
MA, Parvin R, Nooruzzaman M, Rahman MM, Monoura P
(2011) New Introduction of Clade 2.3.2.1 Avian Influenza Virus
(H5N1) into Bangladesh. Transbound Emerg Dis 59(5):460–463.
doi:10.1111/j.1865-1682.2011.01297.x
16. Nagarajan S, Tosh C, Smith DK, Peiris JS, Murugkar HV, Sridevi
R, Kumar M, Katare M, Jain R, Syed Z, Behera P, Cheung CL,
Khandia R, Tripathi S, Guan Y, Dubey SC (2012) Avian Influ-
enza (H5N1) Virus of Clade 2.3.2 in Domestic Poultry in India.
PLoS One 7(2):e31844. doi:10.1371/journal.pone.0031844
17. Khan SU, Gurley ES, Rahman M, Jahangir Hossain M, Uz
Zaman R, Nasreen S, Azim T, Mikolon A, Azziz-Baumgartner E,
Luby SP (2010) Live bird market surveillance for avian influenza
in Bangladesh, 2007–2009. In: Abstract in the International
Conference on Emerging Infectious Diseases. Atlanta
18. WHO (2012) Antigenic and genetic characteristics of zoonotic
influenza viruses and development of candidate vaccine viruses
for pandemic preparedness. September 2012. http://www.who.int/
influenza/vaccines/virus/201209_h5h7h9_vaccinevirusupdate.pdf.
Accessed 21 July 2013
19. Smith GJ, Vijaykrishna D, Ellis TM, Dyrting KC, Leung YH,
Bahl J, Wong CW, Kai H, Chow MK, Duan L, Chan AS, Zhang
LJ, Chen H, Luk GS, Peiris JS, Guan Y (2009) Characterization
of avian influenza viruses A (H5N1) from wild birds, Hong Kong,
2004–2008. Emerg Infect Dis 15(3):402–407
20. FAO-EMPRES (2012) H5N1 HPAI, GLobal Overview, April–
June 2012. http://www.fao.org/docrep/016/ap387e/ap387e.pdf.
Accessed 21 July 2013
21. Li Y, Liu L, Zhang Y, Duan Z, Tian G, Zeng X, Shi J, Zhang L,
Chen H (2011) New avian influenza virus (H5N1) in wild birds,
Qinghai, China. Emerg Infect Dis 17(2):265–267. doi:10.3201/
eid1702.100732
22. L’Vov DK, Shchelkanov M, Prilipov AG, Deriabin PG, Fediak-
ina IT, Galkina IV, Kireev DE, Frolov AV, Akanina DS, Usa-
cheva OV, Shliapnikova OV, Poglazov AB, Morozova TN,
Proshina ES, Grebennikova TV, Zaberezhnyi AD, Iakovlev SS,
Shcherbakova LO, Shapovalov AB, Zhalin MV, Rudenko VP,
Pichuev AE, Litvin KN, Varkentin AV, Steshenko VV, Khari-
tonov SP, Samokhvalov EI, Al’khovskii SV, Aliper TI, Mar-
tynovchenko VV, Lysenko SN, Vlasov NA, Nepoklonov EA
(2008) Interpretation of the epizootic outbreak among wild and
domestic birds in the south of the European part of Russia in
December 2007. Vopr Virusol 53(4):18–23
23. Reid SM, Shell WM, Barboi G, Onita I, Turcitu M, Cioranu R,
Marinova-Petkova A, Goujgoulova G, Webby RJ, Webster RG,
Russell C, Slomka MJ, Hanna A, Banks J, Alton B, Barrass L,
Irvine RM, Brown IH (2011) First reported incursion of highly
pathogenic notifiable avian influenza A H5N1 viruses from clade
2.3.2 into European poultry. Transbound Emerg Dis 58(1):76–78.
doi:10.1111/j.1865-1682.2010.01175.x
24. Sakoda Y, Sugar S, Batchluun D, Erdene-Ochir TO, Okamatsu
M, Isoda N, Soda K, Takakuwa H, Tsuda Y, Yamamoto N,
Kishida N, Matsuno K, Nakayama E, Kajihara M, Yokoyama A,
Takada A, Sodnomdarjaa R, Kida H (2010) Characterization of
H5N1 highly pathogenic avian influenza virus strains isolated
from migratory waterfowl in Mongolia on the way back from the
southern Asia to their northern territory. Virology 406(1):88–94.
doi:10.1016/j.virol.2010.07.007
25. Uchida Y, Mase M, Yoneda K, Kimura A, Obara T, Kumagai S,
Saito T, Yamamoto Y, Nakamura K, Tsukamoto K, Yamaguchi S
(2008) Highly pathogenic avian influenza virus (H5N1) isolated
from whooper swans, Japan. Emerg Infect Dis 14(9):1427–1429
26. Wan XF, Nguyen T, Davis CT, Smith CB, Zhao ZM, Carrel M,
Inui K, Do HT, Mai DT, Jadhao S, Balish A, Shu B, Luo F, Emch
M, Matsuoka Y, Lindstrom SE, Cox NJ, Nguyen CV, Klimov A,
Donis RO (2008) Evolution of highly pathogenic H5N1 avian
influenza viruses in Vietnam between 2001 and 2007. PLoS One
3(10):e3462. doi:10.1371/journal.pone.0003462
27. Newman SH, Hill NJ, Spragens KA, Janies D, Voronkin IO,
Prosser DJ, Yan B, Lei F, Batbayar N, Natsagdorj T, Bishop CM,
Butler PJ, Wikelski M, Balachandran S, Mundkur T, Douglas
DC, Takekawa JY (2011) Eco-virological approach for assessing
the role of wild birds in the spread of avian influenza H5N1 along
the Central Asian Flyway. PLoS One 7(2):e30636
28. Tanimura N, Tsukamoto K, Okamatsu M, Mase M, Imada T,
Nakamura K, Kubo M, Yamaguchi S, Irishio W, Hayashi M,
Nakai T, Yamauchi A, Nishimura M, Imai K (2006) Pathology of
fatal highly pathogenic H5N1 avian influenza virus infection in
large-billed crows (Corvus macrorhynchos) during the 2004
outbreak in Japan. Vet Pathol 43(4):500–509. doi:10.1354/vp.43-
4-500
29. Creanga A, Thi Nguyen D, Gerloff N, Thi Do H, Balish A, Dang
Nguyen H, Jang Y, Thi Dam V, Thor S, Jones J, Simpson N, Shu
B, Emery S, Berman L, Nguyen HT, Bryant JE, Lindstrom S,
Klimov A, Donis RO, Davis CT, Nguyen T (2013) Emergence of
multiple clade 2.3.2.1 influenza A (H5N1) virus subgroups in
Vietnam and detection of novel reassortants. Virology
444:12–20. doi:10.1016/j.virol.2013.06.005
30. Saito T, Uchida Y, Myint WW, Thein WZ, Watanabe C, Take-
mae N, Mase M, Okamatsu M, Mar A, Mon CC, Gawng LT,
Sann K, Kyi TA, Yamaguchi S (2008) Characterisation of highly
pathogenic avian influenza viruses in Myanmar. Vet Rec 163
(24):722–723
31. Ellis TM, Dyrting KC, Wong CW, Chadwick B, Chan C, Chiang
M, Li C, Li P, Smith GJ, Guan Y, Malik Peiris JS (2009) Analysis
of H5N1 avian influenza infections from wild bird surveillance in
Hong Kong from January 2006 to October 2007. Avian Pathol
38(2):107–119. doi:10.1080/03079450902751855
32. Khan SU, Rahman M, Gurley ES, Mikolon AB, Rahman MZ, Uz
Zaman R, Jahangir Hossain M, Nasreen S, Widdowson M-A,
Azim T, Azziz-Baumgartner E, Luby SP (2011) Live bird market
environmental sampling: a tool for poultry influenza surveillance.
In: American Society for Tropical Medicine and Hygiene, 60th
Highly pathogenic avian H5N1 influenza virus in Bangladesh 517
123

annual meeting, December 4–8, 2011. Philadelphia Marriott
Downtown, Philadelphia
33. Sonerud Geir A, Hansen Henning, Smedshaug CA (2002) Indi-
viduarlo ostings trategies in a flock-living bird: movementa and
social cohesiono of hooded crows (Corvus corone comix) from
pre-roost gatheings to roost sites. Behav Ecol Sociobiol
51(4):309–318
34. WHO/OIE/FAO H5N1 Evolution Working Group (2012) Con-
tinued evolution of highly pathogenic avian influenza A (H5N1):
updated nomenclature. Influenza Other Respi Viruses 6(1):1–5.
doi:10.1111/j.1750-2659.2011.00298.x
518 S. U. Khan et al.
123





























