Embed Size (px)
Citation preview

VIROLOGY 183, 44-51 (1991)
Inverted Repeats Direct Repair of Adenovirus Minichromosome Ends
KAI WANG,’ FU-YUN XU,‘KEVlN G. AHERN, AND GEORGE D. PEARSON3
Department of Biochemistry and Biophysics, Oregon State University, Corvallis, Oregon 9733 1
Received September 11, 1990; accepted March 25, 199 1
Adenovirus DNA initiates strand-displacement replication from origins located in identical inverted terminal repeti- tions (ITRs). Panhandle structures, formed by base pairing between ITRs on the displaced strands, have been proposed as replication intermediates for complementary strand synthesis. We have used a model system, which separates adenovirus replication origin sequences from those involved in panhandle formation, to study the length and sequence integrity of panhandles. By making a series of unidirectional deletions in the panhandle sequence, we show that 31 bp are necessary for panhandle formation. Removal of long stretches of 3’unpaired nucleotides distal to the panhandle is extremely efficient. Our results argue for the formation of panhandles during adenovirus DNA replication and provide a mechanism for maintaining sequence identity between distantly located inverted repetitions. The size constraint may explain why the adenovirus ITRs are larger than the viral DNA replication origins. o tSS1 Academic PRSS, I~C.
INTRODUCTION
All human adenovirus DNAs examined to date are linear, have a terminal protein covalently attached to the 5’-end of each strand, and contain inverted terminal repetitions (ITRs)~ spanning 102 to 166 bp (Shinagawa and Padmanabhan, 1980; Stillman et a/., 1982; Sus- senbach, 1984). Only the terminal 45 bp of the ITR are needed for optimal initiation of DNA replication in vitro (Tamanoi and Stillman, 1983; Enns et al,, 1983; Gug- genheimer et a/., 1984; Challberg and Rawlins, 1984; Rawlins et al., 1984) and in viva (Wang and Pearson, 1985; Hay, 1985a; Bernstein et a/., 1986; Hay and McDougall, 1986). Replication occurs in two phases: strand-displacement replication and complementary strand synthesis (Lechner and Kelly, 1977). Extensive evidence has established that initiation of strand-dis- placement replication is primed by terminal protein (Challberg and Kelly, 1979; Challberg eta/., 1980; Eno- moto et a/., 1981; Challberg et a/., 1982; Tamanoi and Stillman, 1982; for review, see Challberg and Kelly, 1989) and leads to the formation of Type I replication intermediates, linear molecules with one or more sin- gle-stranded branches (Lechner and Kelly, 1977). Elec- tron microscopic mapping has confirmed that dis- placed strands originate from either or both molecular ends (Lechner and Kelly, 1977). Completion of a round
’ Present address: Department of Biology 147-75, California Insti- tute of Technology, Pasadena, CA 91 125.
’ Present address: Department of Biochemistry, University of Southern California, Los Angeles, CA 90033.
3 To whom requests for reprints should be addressed, 4 Abbreviations used: ITR, inverted terminal repetition; E. co/i,
fscherichia co/i; bp, base pairs; kb, kilobase pairs,
of replication on Type I molecules produces displaced single strands. Subsequent conversion of displaced strands to double-stranded molecules (complemen- tary strand synthesis) constitutes the second phase of adenovirus replication. The precise mechanism(s) of complementary strand synthesis is not yet known, but an attractive hypothesis is that panhandle structures, created by base pairing between ITRs in the displaced strands, participate in the process by reestablishing double-stranded adenovirus origins (Daniell, 1976; Lechner and Kelly, 1977). Initiation of DNA synthesis on panhandle origins is also primed by terminal protein, and replication through the panhandles produces primed linear intermediates, Such intermediates have been identified by electron microscopy as Type II structures, linear molecules which are double- stranded at one end and single-stranded at the other (Lechner and Kelly, 1977). Although panhandle struc- tures have not been directly detected in infected cells (Revet and Benichou, 198 l), circumstantial evidence supports their formation. Terminal deletions removing one origin, but not the entire ITR, have no effect on adenovirus viability (Stow, 1982). The deleted origin is presumably restored by DNA repair of a residual pan- handle structure formed by base pairing between the remaining ITR sequences. The 3’-end of the panhandle structure serves as the primer for DNA synthesis using 5’-origin sequences as the template. This provides a mechanism to maintain sequence identity at both ends of the adenovirus genome. However, removal of one entire copy of the ITR, which precludes panhandle for- mation and regeneration of the deleted origin, is lethal (Stow, 1982).
We have used an adenovirus minichromosome sys-
0042-6822/91 $3.00 44 CopyrIght 0 1991 by Academtc Press, Inc. All rights of reproduction in any form reserved.

REPAIR OF ADENOVIRUS ENDS 45
tern (Hay el al., 1984; Wang and Pearson, 1985) to study panhandle structures during adenovirus DNA replication. This approach separates origin sequences from those involved in panhandle formation. Linear plasmids with a single adenovirus origin, transfected with helper adenovirus DNA, engage in strand-dis- placement replication (Hay, 1985b). If the plasmids also contain inverted repetitious sequences, new mole- cules with adenovirus origins at both ends are de- tected by Southern blot hybridization (Hay et al., 1984; Wang and Pearson, 1985). The creation of the new origin is consistent with DNA repair of a panhandle structure. Wang and Pearson (1985) have used this system to study origin sequences required for adenovi- rus DNA replication in viva. In this paper, we show that 31 bp are necessary for panhandle formation and that 3’-unpaired nucleotides distal to the panhandle are effi- ciently removed. Our results suggest a mechanism for maintaining sequence identity between distantly lo- cated inverted repetitions. In addition, these data place constraints on the mechanism(s) of producing defec- tive adenovirus molecules, and may explain why the adenovirus ITRs are larger than the viral DNA replica- tion origins.
MATERIALS AND METHODS
Reagents and enzymes
32P-labeled deoxyribonucleotides (3000 Ci/mmol) were from New England Nuclear. Restriction endonu- cleases, Escherichia co/i (E. co//) DNA polymerase I (Klenow fragment), T4 DNA ligase, Sl endonuclease, and E. co/i exonuclease Ill were purchased from New England Biolabs, Bethesda Research Laboratories, Boehringer-Mannheim Biochemicals, or Pharmacia. Pronase was from Calbiochem. Ribonuclease Tl and lysozyme were from Sigma. All of the enzymes were used as recommended by the suppliers.
Plasmids and bacteria
Plasmids pBR322 (Bolivar et al., 1977) pUCl9 (Messing, 1979), pMDCl0 (Enns et a/., 1983) and plR10 (Wang and Pearson, 1985; Fig. 1) have been described previously. Plasmid DNA was isolated by al- kaline extraction (Birnboim and Doly, 1979) and puri- fied by ethidium bromide-CsCI density gradient centrif- ugation. E. co/i RR1 in the form of competent cells was purchased from Bethesda Research Laboratories and used as recommended by the supplier.
Construction of deletion mutants
plRl0 was linearized with EcoRl and digested with exonuclease Ill at a concentration of 15-20 units/pm01
- plRSma - - pIREam
- plRXba - plRHnc - pwlsd - plRPst - plRHm
2.5 kb
ii
EcoRV Sphl
FIG. 1. Maps of plasmids plRl0, plRT6, plRT7, and the plasmids with polylinker inverted repeats (plRSma, etc.). Plasmid vector se- quences are indicated by the thin line. Tc (cross-hatched line) indi- cates the gene for tetracycline resrstance. The thick black line repre- sents adenovirus DNA. An adenovirus replication origin is located on each of the plasmids immediately adjacent to the EcoRl site (the direction of adenovirus-specific replication is clockwise on these maps). Short arrows show the orientation of repeated sequences (open boxes). Repeats are 89 bp long in plR10. plRT6. and plRT7. The repeats in the polylinker plasmids range from 51 bp (plRHin) down to 14 bp (plRSma). Sites in the polylinker region: E, EcoRI; Sm, Smal; B, BarnHI; X. Xbal; SI. Sall, P. Pstl, and H, HindIll.
of DNA. At this concentration of enzyme, 10 to 15 nu- cleotides/min were removed (Wu er a/., 1976). At various times, samples were removed and treated with Sl endonuclease at a concentration of 40 units/pm01 of DNA for 15 min at 20” to remove single-strand tails. The DNA was then incubated with the Klenow frag- ment of DNA polymerase I to blunt the ends. DNAfrom different time points were ligated to an EcoRl linker and cloned in E. co/i RR1 . Plasmid DNA from each ampicil- lin-resistant colony was screened for deletions after cleavage with EcoRl and Pvull. Since the deletions ex- tended bidirectionally from the original EcoRl site and removed adenovirus terminal sequences, the small EcoRI-BarnHI fragment from plRl0 was cloned back into the EcoRI-BarnHI backbone of each deletion mu- tant. One reconstructed mutant, called plRD1, con- tained inverted repeats of 54 bp as determined by DNA sequence analysis.

46 WANG ET AL.
Construction of inverted repetitious polylinker plasmids
The following scheme was used for constructing plasmids carrying inverted repeat sequences derived from all or part of the pUCl9 polylinker region. The EcoRI-Sspl fragment of pMDC10, containing adenovi- rus terminal sequences, was ligated to the EcoRI-Sspl backbone of pUC19. An intermediate plasmid, called T4, with 333 bp of the left end of adenovirus DNA and a pUC19 polylinker region (57 bp) was isolated and char- acterized from an ampicillin-resistant E. co/i RR1 col- ony. T4 was then cut with Sspl and any one of the restriction endonucleases (Smal, BarnHI, Xbal, Hincll, Sa/l, Pstl, or HindIll) cutting within the polylinker region. The small fragment was isolated in each case and blunted with DNA polymerase I. The purified frag- ments, which contained adenovirus terminal se- quences and all or part of the pUCl9 polylinker region, were ligated to the blunted pUCl9 EcoRI-Sspl back- bone. Plasmids in ampicillin-resistant E. co/i RR1 colo- nies were screened initially for the presence of a 390- bp EcoRI-BamHI fragment. Depending on which re- striction endonuclease was used, plasmids had inverted repetitions ranging from 14 bp (plRSma) to 51 bp (plRHin) as further determined by DNA sequence analysis (see Fig. 1).
Construction of plasmids with multiple inverted repeats
The following scheme was used to construct plas- mids carrying multiple copies of 89-bp inverted repeat sequences. A blunted 2.1 -kb EcoRI-Pvull fragment from pBR322 containing the tetracycline-resistance (Tc) gene was cloned into the blunted EcoRl site of pUCl9 under tetracycline selection. A 4.8-kb interme- diate clone, called pRT19, was isolated. The regener- ated EcoRl site in pRTl9 was removed to yield plasmid pT3. A 2.2-kb Smal-Pvull fragment from ~7-3 was then cloned into the Xmnl site of plR10 under tetracycline selection. Two 5.4-kb plasmids, called plRT6 and plRT7 (Fig. l), differing only in the orientation of the inserted fragment were recovered and characterized. Both plasmids contain three copies of the 89-bp repeat found in plR10.
DNA sequence analysis
Dideoxy sequencing on double-stranded plasmid DNA was done according to Zagursky et a/., (1985).
Cells and virus
The 293 cell line (Graham et a/., 1977) was grown in Dulbecco’s modified Eagle’s medium (GIBCO) with 5%
fetal calf serum (HyClone). The HeLa cell line was grown in Joklik’s minimal essential medium (GIBCO) containing 5% fetal calf serum. Adenovirus type 5 was grown in HeLa cells; virus and viral DNA were isolated by the method of Pettersson and Sambrook (1973).
Transfection
The 293 cells were transfected according to the standard DNA-calcium phosphate coprecipitation method (Graham and van der Eb, 1973). Transfections utilized 5 pg of plasmid DNAand 5 pg of helper adenovi- rus type 5 DNA. Each 60-mm plate was inoculated with 4 X lo5 cells 24 hr before transfection. Cells were shocked for 1 min with 25% glycerol 4 hr after trans- fection.
Extraction and analysis of DNA
DNA was isolated from transfected 293 cells and analyzed by Southern blot hybridization using 32P-la- beled pUC8 or pUCl9 DNA (sp act > 1 X 1 O8 cpm1p.g) as a probe as described previously (Wang and Pear- son, 1985).
RESULTS
Adenovirus-specific replication of linear plasmids in viva
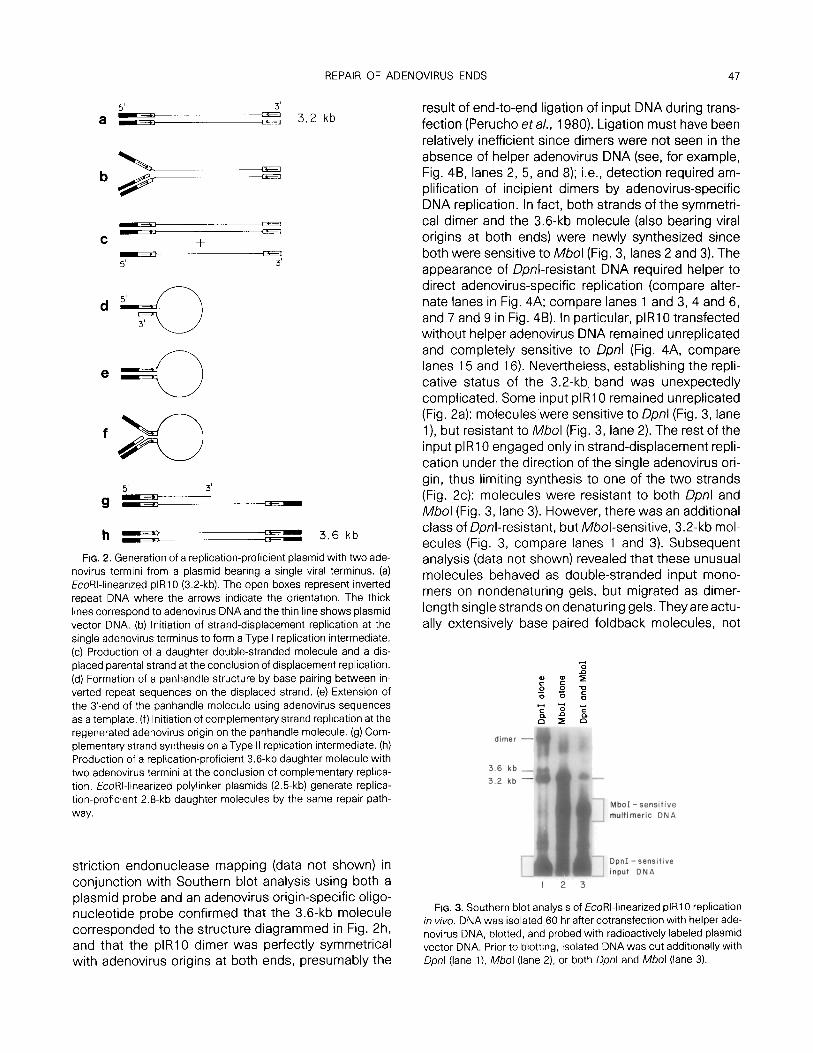
Figure 2 diagrams the postulated sequence of events required to produce replication-proficient mole- cules starting from EcoRI-linearized plasmid plRl0 (Wang and Pearson, 1985; Fig. 1). Central to this scheme is the formation of a panhandle structure (Fig. 2d), similar to a proposed intermediate in the replica- tion of adenovirus DNA (Daniell, 1976; Lechner and Kelly, 1977) by base pairing between the inverted repe- titious sequences on the displaced strand. Several predictions about this pathway can be tested using a strategy developed by Peden et a/. (1980). Plasmid DNA propagated in Dam+ E. co/i hosts is sensitive to Qonl but resistant to Mbol (both enzymes recognize the sequence GATC, but Dpnl only cuts when the site is methylated on both strands and Mb01 only cuts when the site is totally unmethylated). Since mamma- lian cells cannot methylate the sequence GATC, input plasmid DNA unable to replicate after transfection re- mains Dpnl-sensitive and Mbol-resistant, whereas DNA with one newly synthesized strand becomes re- sistant to both enzymes, and DNA with two newly repli- cated strands is Dpnl-resistant but A&ol-sensitive.
Figure 3 (lane 1) shows that several Qonl-resistant bands were detected when plRl0 was cotransfected with helper adenovirus DNA: 3.2-kb input DNA, a novel 3.6-kb molecule, and a dimer of plR10. Extensive re-
REPAIR OF ADENOVIRUS ENDS 47
a
b
d
e
9
h
5’ 3’ I- --.c 3.2 kb
3.6 kb
FIG. 2. Generation of a replication-proficient plasmid with two ade- novirus termini from a plasmid bearing a single viral terminus. (a) EcoRI-linearized plRl0 (3.2.kb). The open boxes represent inverted repeat DNA where the arrows indicate the orientation. The thick lines correspond to adenovirus DNA and the thin line shows plasmid vector DNA. (b) Initiation of strand-displacement replication at the single adenovirus terminus to form a Type I replication intermediate. (c) Production of a daughter double-stranded molecule and a dis- placed parental strand at the conclusion of displacement replication. (d) Formation of a panhandle structure by base pairing between in- verted repeat sequences on the displaced strand. (e) Extension of the 3’.end of the panhandle molecule using adenovirus sequences as a template. (f) Initiation of complementary strand replication at the regenerated adenovirus origin on the panhandle molecule. (g) Com- plementary strand synthesis on a Type II replication intermediate. (h) Production of a replication-proficient 3.6-kb daughter molecule with two adenovirus termini at the conclusion of complementary replica- tion. EcoRI-linearized polylinker plasmids (2.5.kb) generate replica- tion-proficient 2.8.kb daughter molecules by the same repair path- way.
striction endonuclease mapping (data not shown) in conjunction with Southern blot analysis using both a plasmid probe and an adenovirus origin-specific oligo- nucleotide probe confirmed that the 3.6-kb molecule corresponded to the structure diagrammed in Fig. 2h, and that the plRl0 dimer was perfectly symmetrical with adenovirus origins at both ends, presumably the
result of end-to-end ligation of input DNA during trans- fection (Perucho eta/., 1980). Ligation must have been relatively inefficient since dimers were not seen in the absence of helper adenovirus DNA (see, for example, Fig. 4B, lanes 2, 5, and 8); i.e., detection required am- plification of incipient dimers by adenovirus-specific DNA replication. In fact, both strands of the symmetri- cal dimer and the 3.6-kb molecule (also bearing viral origins at both ends) were newly synthesized since both were sensitive to Mb01 (Fig. 3, lanes 2 and 3). The appearance of Dpnl-resistant DNA required helper to direct adenovirus-specific replication (compare alter- nate lanes in Fig. 4A; compare lanes 1 and 3, 4 and 6, and 7 and 9 in Fig. 4B). In particular, plR10 transfected without helper adenovirus DNA remained unreplicated and completely sensitive to Dpnl (Fig. 4A, compare lanes 15 and 16). Nevertheless, establishing the repli- cative status of the 3.2-kb band was unexpectedly complicated. Some input plR10 remained unreplicated (Fig. 2a): molecules.were sensitive to Qonl (Fig. 3, lane l), but resistant to Mbol (Fig. 3, lane 2). The rest of the input plR10 engaged only in strand-displacement repli- cation under the direction of the single adenovirus ori- gin, thus limiting synthesis to one of the two strands (Fig. 2~): molecules were resistant to both Dpnl and Mb01 (Fig. 3, lane 3). However, there was an additional class of Dpnl-resistant, but Il/lbol-sensitive, 3.2-kb mol- ecules (Fig. 3, compare lanes 1 and 3). Subsequent analysis (data not shown) revealed that these unusual molecules behaved as double-stranded input mono- mers on nondenaturing gels, but migrated as dimer- length single strands on denaturing gels. They are actu- ally extensively base-paired foldback molecules, not
dimar -1
3.6 kb -
3.2 kb -
MboI - sensitive multimeric DNA
DpnI - sensitive input DNA
FIG. 3. Southern blot analyss of EcoRI-linearized plR10 replication in viva. DNA was isolated 60 hr after cotransfection with helper ade- novirus DNA, blotted, and probed with radioactively labeled plasmid vector DNA. Prior to blotting, isolated DNA was cut additionally with Dpnl (lane l), Mf~ol (lane 2) or both Dpnl and n/lb01 (lane 3).
48 WANG ET At
0 0 c - ‘, E
0 E
z a
E 2
0 z 2
0 z c =a .G
LL a: E 2 CK z is 2 5
A a a a a a a a a is (1 6
51 35 33 28 21 14 54 89 0 Panhandle Panhandle
+I- +I- +I- +I- +I- +[- +I- +I- +I- Helper Helper
Dpnl
dimer -
- 3.6
- 3.2
-D Dimer
,ena ‘.qe: &”
--c 2.8 Kb
i 1
* 2.5 hb Dpnl- cut
input DNA
I 2 3 4 5 6 7 8 9 IO II 12 13 I4 I5 16 I7 I8 123456789
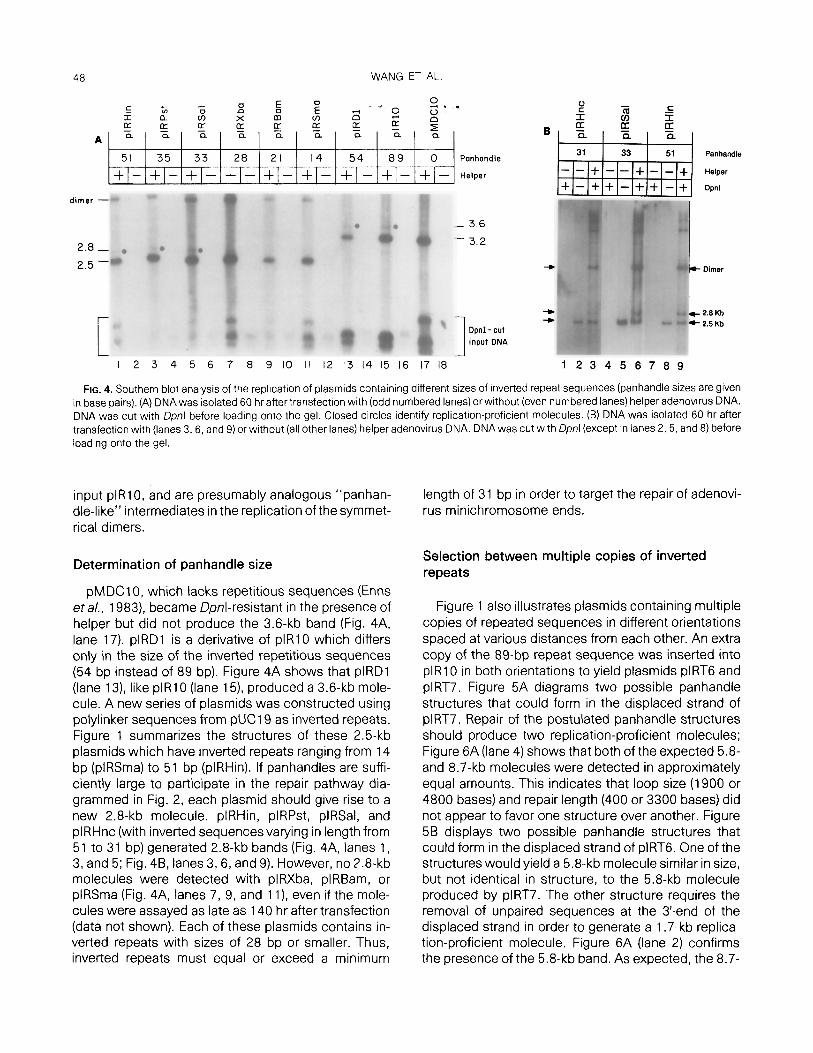
FIG. 4. Southern blot analysis of the replication of plasmids containing different sizes of inverted repeat sequences (panhandle sizes are given in base pairs). (A) DNA was isolated 60 hr after transfection with (odd numbered lanes) or without (even numbered lanes) helper adenovirus DNA. DNA was cut with Dpnl before loading onto the gel. Closed circles identify replication-proficient molecules. (B) DNA was isolated 60 hr after transfection with (lanes 3, 6, and 9) or without (all other lanes) helper adenovirus DNA. DNA was cut with /Ipnl (except in lanes 2, 5, and 8) before loading onto the gel
input plRl0, and are presumably analogous “panhan- dle-like” intermediates in the replication of the symmet- rical dimers.
Determination of panhandle size
pMDC10, which lacks repetitious sequences (Enns eta/., 1983) became Qonl-resistant in the presence of helper but did not produce the 3.6-kb band (Fig. 4A, lane 17). plRD1 is a derivative of plRl0 which differs only in the size of the inverted repetitious sequences (54 bp instead of 89 bp). Figure 4A shows that plRD1 (lane 13) like plR10 (lane 15) produced a 3.6-kb mole- cule. A new series of plasmids was constructed using polylinker sequences from pUCl9 as inverted repeats. Figure 1 summarizes the structures of these 2.5-kb plasmids which have inverted repeats ranging from 14 bp (plRSma) to 51 bp (plRHin). If panhandles are suffi- ciently large to participate in the repair pathway dia- grammed in Fig. 2, each plasmid should give rise to a new 2.8-kb molecule. plRHin, plRPst, plRSal, and plRHnc (with inverted sequencesvarying in length from 51 to 31 bp) generated 2.8-kb bands (Fig. 4A, lanes 1, 3, and 5; Fig. 4B, lanes 3, 6, and 9). However, no 2.8-kb molecules were detected with plRXba, plRBam, or plRSma (Fig. 4A, lanes 7, 9, and 1 l), even if the mole- cules were assayed as late as 140 hr after transfection (data not shown). Each of these plasmids contains in- verted repeats with sizes of 28 bp or smaller. Thus, inverted repeats must equal or exceed a minimum
length of 31 bp in order to target the repair of adenovi- rus minichromosome ends.
Selection between multiple copies of inverted repeats
Figure 1 also illustrates plasmids containing multiple copies of repeated sequences in different orientations spaced at various distances from each other. An extra copy of the 89-bp repeat sequence was inserted into plR10 in both orientations to yield plasmids pIRT6 and plRT7. Figure 5A diagrams two possible panhandle structures that could form in the displaced strand of plRT7. Repair of the postulated panhandle structures should produce two replication-proficient molecules; Figure 6A (lane 4) shows that both of the expected 5.8- and 8.7-kb molecules were detected in approximately equal amounts. This indicates that loop size (1900 or 4800 bases) and repair length (400 or 3300 bases) did not appear to favor one structure over another. Figure 58 displays two possible panhandle structures that could form in the displaced strand of plRT6. One of the structures would yield a 5.8-kb molecule similar in size, but not identical in structure, to the 5.8-kb molecule produced by plRT7. The other structure requires the removal of unpaired sequences at the 3’-end of the displaced strand in order to generate a 1.7-kb replica- tion-proficient molecule. Figure 6A (lane 2) confirms the presence of the 5.8-kb band. As expected, the 8.7-

REPAIR OF ADENOVIRUS ENDS
- plRT7 5.4Kb
49
DISCUSSION
P plRT6 5.4Kb
A
5.6 Kb 1.7 Kb
FIG. 5. Formation and fate of panhandle structures on displaced strands bearing multiple copies of inverted repeat sequences. See Fig. 2 for complete details of the replication pathway and an explana- tion of the symbols. (A) plRT7. (B) plRT6.
kb molecule did not appear. The 1.7-kb molecule was not seen clearly on this blot due to obscuring Dpnl digestion products. However, it was unambiguously detected in a different blot (Fig. 66, lane 1). The 1.7- and 5.8-kb bands had roughly equal intensities, indi- cating that both panhandles were efficiently used even though more than 4000 unpaired bases had to be re- moved from the 3’-end of the structure postulated as an intermediate for the 1.7-kb molecule. Neither loop size (700 or 4800 bases) nor the presence (up to 4000 bases) or absence of 3’-unpaired tails appeared to bias the yield. Figure 6B also demonstrates that we could force the exclusive appearance of the 1.7-kb band by removing the inverted repeat sequence from 3’-end of plRT6 with various restriction endonucleases. EcoRI- cut plRT6 truncated additionally with C/al (lane 2) EcoRV (lane 3) Sphl (lane 4) or Aval (lane 5) gave rise only to the 1.7-kb molecule. The residual 3’-unpaired tails on the truncated plasmids ranged in length from 23 to 1425 bases, respectively. Although some bands in lanes 2 through 5 approximated the sizes of plRT6 (5.4 kb) or the 5.8-kb end-repaired molecule, they were clearly identified as replication-proficient oligomers of the truncated input plasmid in each transfection: a 5.2- kb tetramer (lane 2) a 6.0-kb tetramer (lane 3) and a 5.4-kb dimer (lane 5).
We have used an adenovirus minichromosome sys- tem (Hay et a/., 1984; Wang and Pearson, 1985) to provide new evidence that panhandle formation is an important part of adenovirus complementary strand replication. Linear plasmids with one adenovirus origin (pMDC10) only replicated a single strand to become Dpnl-resistant in the presence of helper adenovirus DNA. New, larger molecules with adenovirus origins at both ends were generated if the input plasmids also contained inverted repetitious sequences (plR10 or polylinker plasmids). Since these new molecules were sensitive to Mbol, both strands of the molecules were newly replicated. Only inverted repeats equaling or ex- ceeding a minimum length of 31 bp targeted the repair of adenovirus minichromosome ends. Furthermore, pairs of inverted repeats completely surrounded by
Dimer 8.7 Kb
5.8 Kb Monomer
Dpnl-sensitive inwt DNA
I--1+1-I+1 12 34
pIFiT BI, I
E- “1 $3 ZT SUwma 1++++ ----_
dimer +
5.6 kb input 3
.7 kb
FIG. 6. (A) Southern blot analysis of the replication of plasmids containing multiple copies of inverted repeat sequences. DNA was isolated 60 hr after transfection with (lanes 2 and 4) or without (lanes 1 and 3) helper adenovirus DNA. DNA was cut with Dpnl before loading onto the gel. Arrows show the positions of monomers, dimers, and 5.8- and 8.7.kb replication-proficient repaired mole- cules. Lanes 1 and 2, plRT6. Lanes 3 and 4, plRT7. (B) Southern blot analysis of plRT6 cut with EcoRl alone (lane 1) or EcoRl together with C/al (lane 2) EcoRV (lane 3) Sphl (lane 4) or Awl (lane 5). The 3’unpaired tails are respectively 4000, 23, 185, 562, and 1425 nt. Samples were digested with Dpnl prior to gel electrophoresis. Arrows show the positions of monomers, dimers, and 1.7- and 5.8- kb replication-proficient repaired molecules. Bands corresponding to the truncated input plasmids are indicated by the letter “i” and the truncated dimer bands are labeled with the letter “d” in lanes 2 through 5.

50 WANG ET AL.
large regions of heterology were equally as effective in promoting end-repair (e.g., the 1.7-kb product of plRT6) as pairs of inverted repeats where at least one of the repeat borders lacked adjacent sequences (e.g., the 5.8-kb product of plRT6). These results argue against homologous recombination as a mechanism for end-repair. First, homologous recombination has been detected between sequences ranging in size from 14 bp (Rubnitz and Subramani, 1984) to 25 bp (Ayares et a/., 1986). Second, strand invasion and dou- ble-strand-break-stimulated mechanisms of recombi- nation are inhibited by adjacent heterology as small as 40 bp in yeast (Struhl, 1987), Xenopus oocytes (Carroll et a/., 1986; Abastado et al,, 1987; Grzesiuk and Carroll, 1987), and mouse cells (Brenner et al,, 1986).
The structures of the new, end-repaired molecules are consistent with the pathway outlined in Fig. 2. This pathway exhibits several key features: (a) Only one strand, the displaced strand, from the input molecule participates in end-repair. (b) The intermediate, a pan- handle structure, arises from direct intramolecular hy- bridization of complementary sequences (inverted re- peats) within the displaced strand. Since interacting sequences are initially single-stranded and hybridiza- tion does not involve the progressive invasion of a du- plex structure, heterology surrounding the inverted re- peats does not block the formation of panhandles. (c) Initially only the inverted repeats are double-stranded in the panhandle intermediate. (d) Unpaired 3’-ends, but not 5’-ends, are excised from the intermediate (e.g., the 1.7-kb product of plRT6). We have also ob- served the efficient removal of unpaired 3’-ends from both strands of heteroduplex intermediates formed during adenovirus postreplicative overlap recombina- tion (Ahern et al., 1991). The mechanism for eliminat- ing unpaired 3’-tails is not yet known, but the potent 3’- to 5’-exonuclease activity of the adenovirus DNA poly- merase (Field et al., 1984) may be involved. (f) The adenovirus origin is regenerated by DNA repair synthe- sis from the base-paired S/-end of the panhandle in the intermediate using 5’-sequences as a template. Only panhandles 31 bp in size or longer can be repaired. The abrupt length dependence suggests a steric con- straint. In fact, recent work (unpublished data) sup- ports this conclusion since the repair of otherwise iso- genie mismatches located within the duplex panhan- dle region is position- and orientation-dependent. Only mismatches located at least 31 bp from the loop of the intermediate are corrected by DNA repair synthesis proceeding away from the loop, although mismatches as close as 6 bp can be rectified by synthesis toward the loop. The molecular basis of the steric obstruction remains to be determined. (g) Subsequent rounds of
adenovirus-specific DNA replication amplify the new, end-repaired molecules.
The limit placed on panhandle length is a fundamen- tal constraint and has important consequences for the biology of adenoviruses. Panhandle structures have been proposed as intermediates in the replication of complementary strands (Lechner and Kelly, 1977), in the repair of terminal deletions removing one origin (Stow, 1982), in the generation of subgenomic defec- tive molecules (Daniell, 1976), and in the rearrange- ment of nondefective mutant genomes with embedded inverted terminal repeats (Haj-Ahmad and Graham, 1986). The length of panhandles presumably must equal or exceed 3 1 bp in order to participate in each of these pathways. Thus, blocking DNA repair synthesis on all but sufficiently long panhandles may serve to minimize the fortuitous creation of subgenomic defec- tive molecules, while still allowing the viral DNA replica- tion origin to survive potentially lethal terminal dele- tions or substitutions damaging one entire origin. Long ITRs may have a selective advantage since the correc- tion of mismatched sequences anywhere within the adenovirus replication origin could occur if ITRs ex- ceeded 76 bp in length (at least 31 bp larger than the 45-bp viral origin). Interestingly, Lippb and Graham (1989) have recently shown that a hybrid virus with nonidentical terminal sequences is viable. Pure clones of the virus cannot be isolated since progeny virus with identical ends is rapidly generated, most likely by the same repair pathway illustrated in Fig. 5B. The viability of the hybrid virus, however, implies that the ends have some opportunity to elude mismatch repair. The signifi- cance of this escape remains to be established with regard to a mechanism(s) for adenovirus DNA replica- tion.
ACKNOWLEDGMENTS
We thank Catherine Z. Mathews, Margaret Nervik Ho, and Kelsie Thielen for excellent technical assistance. This work was supported by National Institutes of Health grant CA1 7699 (GDP) and American Cancer Society Grant PF-2938 (KGA).
REFERENCES
ABASTADO, J.-P., DARCHE, S., GODEAU, F., CAMI, B., and KOURILSKY, P. (1987). Intramolecular recombination between partially homolo- gous sequences in Escherichia cob and Xenopus laevis oocytes. Proc. Nat/. Acad. Sci. USA 84, 6496-6500.
AHERN, K. G., WANG, K.. Xu, F.-Y., MATHEWS, C. Z., and PEARSON, G. D. (1991). Strands hybridize in postreplicative adenovirus over- lap recombination. Proc. Nat/, Acad. Sci. USA 88, 105-l 09.
AYARES, D., CHEKURI, L., SONG, K.-Y., and KUCHERLAPATI, R. (1986). Sequence homology requirements for intermolecular recombina- tion in mammalian cells. Proc. Nat/, Acad. Sci, USA 83, 5199- 5203.
BERNSTEIN, J. A., PORTER. J. M., and CHALLBERG, M. D. (1986). Tem-

REPAIR OF ADENOVIRUS ENDS 51
plate requirements for in viva replication of adenovirus DNA. Mol. Cell. Biol. 6, 2115-2124.
BIRNBOIM, H. C., and DOLY, J. (1979). A rapid alkaline extraction pro- cedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7, 1513-l 523.
BOLIVAR, F., RODRIGUEZ, R. L., GREENE, P. J., BETLACH, M. V., HEY- NECKER, H. L., BOYER, H. W., CROSA, J. H., and FALKOW, S. (1977). Construction and characterization of new cloning vehicles. Il. A multipurpose cloning system. Gene 2, 95-1 13.
BRENNER, D. A., SMIGOCKI, A. C., and CAMERINI-OTERO, R. D. (1986). Double-strand gap repair results in homologous recombination in mouse L cells. Proc. Natl. Acad. Sci. USA 83, 1762-l 766.
CARROLL, D., WRIGHT, S. H., WOLFF, R. K., GRZESIUK, E., and MARYON. E. B. (1986). Efficient homologous recombination of linear DNA substrates after injection into Xenopus laevis oocytes. Mol. Cell. Biol. 6, 2053-2061.
CHALLBERG, M. D., and KELLY, T. J.. JR. (1979). Adenovirus DNA repli- cation in vitro. Proc. Natl. Acad. Sci. USA 76, 655-659.
CHALLBERG, M. D., and KELLY, T. J. (1989). Animal virus DNA replica- tion. Annu. Rev. Biochem. 58, 67 l-71 7.
CHALLBERG, M. D., DESIDERIO, S. V., and KELLY, T. J., JR. (1980). Ade- novirus DNA replication in vitro: Characterization of a protein co- valently linked to nascent DNA strands. Proc. Nat/. Acad. Sci. USA 77,5105-5109.
CHALLBERG, M. D., OSTROVE, J. M., and KELLY, T. J., JR. (1982). Initia- tion of adenovirus DNA replication: Detection of covalent com- plexes between nucleotide and the 80.kilodalton terminal protein. J. Viral. 41, 265-270.
CHALLBERG, M. D., and RAWLINS, D. R. (1984). Template require- ments for the initiation of adenovirus DNA replication. Proc. Nat/. Acad. Sci. USA 81, 100-104.
DANIELL, E. (1976). Genome structure of incomplete particles of ade- novirus. J. Viral. 19, 685-708.
ENNS, R. E., CHALLBERG, M. D., AHERN, K. G., CHOW, K-C., MATHEWS, K. Z., ASTELL, C. R., and PEARSON, G. D. (1983). Mutational map- ping of a cloned adenovirus origin. Gene 23, 307-3 13.
ENOMOTO, T., LICHY, J. H., IKEDA, J.-E., and HURWIT~, J. (1981). Adeno- virus DNA replication in vitro: Purification of the terminal protein in a functional form. Proc. Nat/. Acad. Sci. USA 78, 6779-6783.
FIELD, J., GRONOSTNSKI, R. M., and HURWITZ, J. (1984). Properties of the adenovirus DNA polymerase. /. Biol. Chem. 259, 9487-9495.
GRAHAM, F. L., SMILEY, J., RUSSELL, W. C., and NAIRN, R. (1977). Char- acteristics of a human cell line transformed by DNA from human adenovirus 5. J. Gen. Virol. 36, 59-72.
GRAHAM, F. L., and VAN DER EB, A. J. (1973). A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 52,456- 467.
GRZESIUK. E., and CARROLL, D. (1987). Recombination of DNAs in Xenopus oocytes based on short homologous overlaps. Nucleic Acids Res. 15, 971-985.
GUGGENHEIMER, R. A., STILLMAN, B. W., NAGATA, K.. TAMANOI, F., and HURWITZ, 1. (1984). DNA sequences required for the in vitro replica- tion of adenovirus DNA. Proc. Nat/. Acad. Sci. USA 81, 3069- 3073.
HN-AHMAD, Y., and GRAHAM, F. L. (1986). Characterization of an adenovirus type 5 mutant carrying embeded inverted terminal re- peats. Virology 153, 22-34.
HAY, R. T. (1985a). The origin of adenovirus DNA replication: Minimal DNA sequence requirement in vivo. EMBO /. 4, 42 l-426.
HAY, R. T. (198513). Origin of adenovirus DNA replication. Role of the nuclear factor I binding site in vivo. J. Mol. Biol. 186, 129-136.
HAY, R. T., and MCDOUGALL, I. M. (1986). Viable viruses with dele- tions in the left inverted terminal repeat define the adenovirus ori- gin of DNA replication. 1. Gen. Viral. 67, 321-332.
HAY, R. T.. STOW, N. D., and MCDOUGALL, I. M. (1984). Replication of adenovirus mini-chromosomes. f. Mol. Biol. 175, 493-5 10.
LECHNER, R. L., and KELLY, T. J. (1977). The structure of replicating adenovirus 2 DNA molecules. Cell 12, 1007-l 020.
LIPP~, R., and GRAHAM, F. L. (1989). Adenoviruses with nonidentical terminal sequences are viable. 1. Viral. 63, 5133-5141.
MESSING, J. (1979). “Recombinant DNATechnical Bulletin,” National Institutes of Health Publ. No. 79-99, Vol. 2, pp. 43-48. National Institutes of Health, Bethesda, MD.
PEDEN, K. W. C., PIPAS, J. M., PEARSON-WHITE. S., and NATHANS. D. (1980). Isolation of mutants of an animal virus in bacteria. Science 209, 1392-l 396.
PERUCHO, M., HANAHAN, D., LIPSICH, L., and WIGLER, M. (1980). Isola- tion of the chicken thymidine kinase gene by plasmid rescue. Na- ture 285, 207-210.
PE-TTERSSON, U., and SAMBROOK, J. (1973). Amount of viral DNA in the genome of cells transformed by adenovirus type 2. J. Mol. Biol. 73, 125-130.
RAWLINS, D. R., ROSENFELD, P. J., WIDE% R. J., CHALLBERG, M. D., and KELLY, T. J., JR. (1984). Structure and function of the adenovirus origin of replication. Cell 37, 309-319.
REVET, B., and BENICHOU, D. (1981). Electron microscopy of AD5 replicating molecules after in vivo photocrosslinking with trioxsa- len. Virology 114, 60-70.
RUBNITZ, J., and SUBRAMANI, S. (1984). The minimum amount of ho- mology required for homologous recombination in mammalian cells. Mol. Cell. Biol. 4, 2253-2258.
SHINAGAWA, M., and PADMANABHAN. R. (1980). Comparative se- quence analysis of the inverted terminal repetitions from different adenoviruses. Proc. Natl. Acad. Sci. USA 77, 3831-3835.
STILLMAN, B. W., TOPP, W. C., and ENGLER. J. A. (1982). Conserved sequences at the origin of adenovirus DNA replication. J. Viral. 44, 530-537.
STOW, N. D. (1982). The infectivity of adenovirus genomes lacking DNA sequences from their left-hand termini. Nucleic Acids Res. 10,5105-5119.
STRUHL, K. (1987). Effect of deletion and insertion on double-strand- break repair in Saccharomyces cerevisiae. Mol. Cell. Biol. 7, 1300-l 303.
SUSSENBACH, J. S. (1984). The structure of the genome. In “The Ade- noviruses” (H. S. Ginsberg, Ed.), pp. 271-308. Plenum, NewYork.
TAMANOI, F., and STILLMAN. B. W. (1982). Function of adenovirus terminal protein in the initiation of DNA replication. Proc. Nat/. Acad. Sci. USA 79,2221-2225.
TAMANOI. F., and STILLMAN, B. W. (1983). Initiation of adenovirus DNA replication in vitro requires a specific DNA sequence. Proc. Nat/ Acad. Sci. USA 80, 6446-6450.
WANG, K., and PEARSON, G. D. (1985). Adenovirus sequences re- quired for replication in vivo. Nucleic Acids Res. 13, 5173-5187.
Wu, R.. RUBEN, G., SIEGEL, B., JAY, E., SPIELMAN, P., and Tu, C. P. (1976). Synchronous digestion of SV40 DNA by exonuclease Ill. Biochemistry 15, 734-740.
ZAGURSKY, R., NICKLEN, N., and COULSON, A. R. (1985). Rapid and easy sequencing of large linear double-strand DNA and super- coiled plasmid DNA. Gene Anal. Tech. 2, 89-94.





























