Embed Size (px)
Citation preview

Introduction to Viral ImmunologyPart I
Dr. David J. Topham
Portions adapted from Dr. Colin R.A. Hewitt

References/Texts
• Principles of Virology: Molecular Biology, Pathogenesis, and Control Author(s)/Editor(s): S.J. Flint, L.W. Enquist, R.M. Krug, V.R. Racaniello, and A.M. Skalka ASM Press, December 2003
– Chapter 14
• Immunobiology:The Immune System in Health and Disease (6th Edition) Authors/Editors: C.A. Janeway, Jr., P. Travers, M. Walport, M.J. Schlomchik Garland Science Publishing, 2005.
– Chapters 2, 5, 10
• Viruses and Human DiseasesAuthors: James H. Strauss and Ellen G. Strauss. Elsevier.

Outline
• Innate mechanisms of antiviral immunity– Toll like receptors– Type I interferons– Natural Killer cells
• Antigen-specific recognition of virus and virus infected cells– Antigen processing and presentation– Dendritic cells– Cytotoxic CD8 T cells and Helper CD4 T cells

Preventing infection
• Physical barriers– Skin, mucosa
• Neutralization – Antibodies
• requires immunization
• Limit replication– Interferons (Type I and Type 2)– Defensins
• MULTIPLE ROLES OF ANTIMICROBIAL DEFENSINS, CATHELICIDINS, AND EOSINOPHIL-DERIVED NEUROTOXIN IN HOST
DEFENSE De Yang, Arya Biragyn,DavidM. Hoover,Jacek Lubkowski, and Joost J. Oppenheim Annu. Rev. Immunol. 2004. 22:181–215
• Mostly restricted to bacterial and fungal pathogens

Recognition of a virus infection• Toll like receptors (TLR)
– TLR 2, 3, 7, 9– MyD88, MAL/TIRAP,TRIF, TRAM
• NK Receptors– Activation and Inhibitory receptors
• ITAM and ITIM intracellular signaling motifs
• Dendritic cells– Linking innate and adaptive immune responses
• Antigen processing and presentation of viral peptides– Class I and class II MHC

Toll Like ReceptorsInnate immunity to viruses
• Pattern recognition– Recognize conserved molecular structures
unique to pathogens
• Germline encoded
• Expressed on a variety of immune and non-immune cells
• Instruct Dendritic Cells (DC)
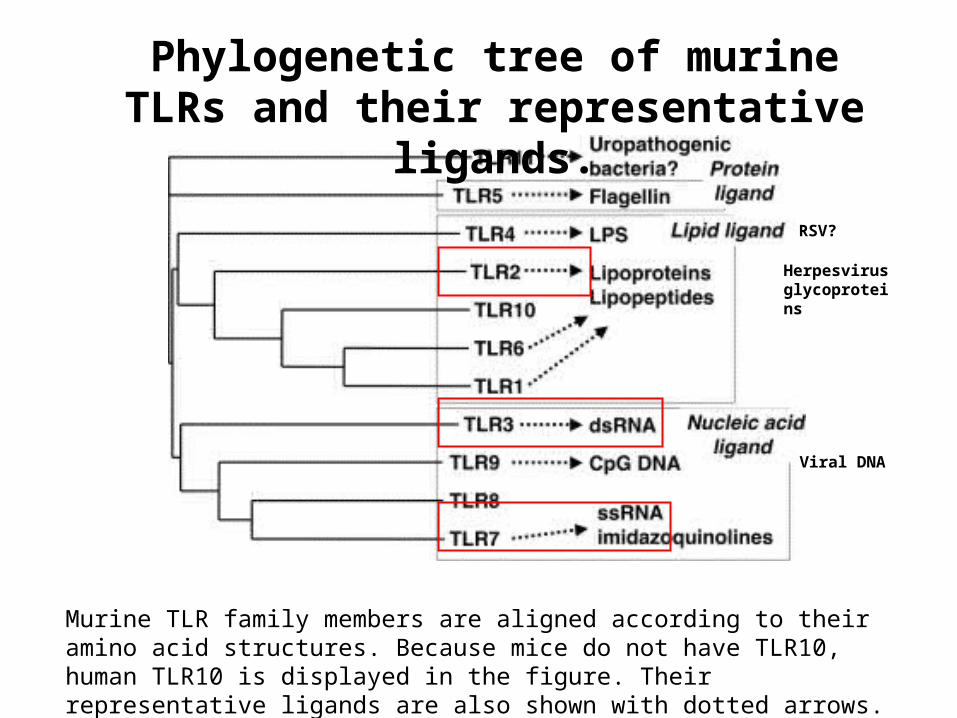
Murine TLR family members are aligned according to their amino acid structures. Because mice do not have TLR10, human TLR10 is displayed in the figure. Their representative ligands are also shown with dotted arrows.
Phylogenetic tree of murine TLRs and their representative ligands.
Herpesvirus glycoproteins
Viral DNA
RSV?
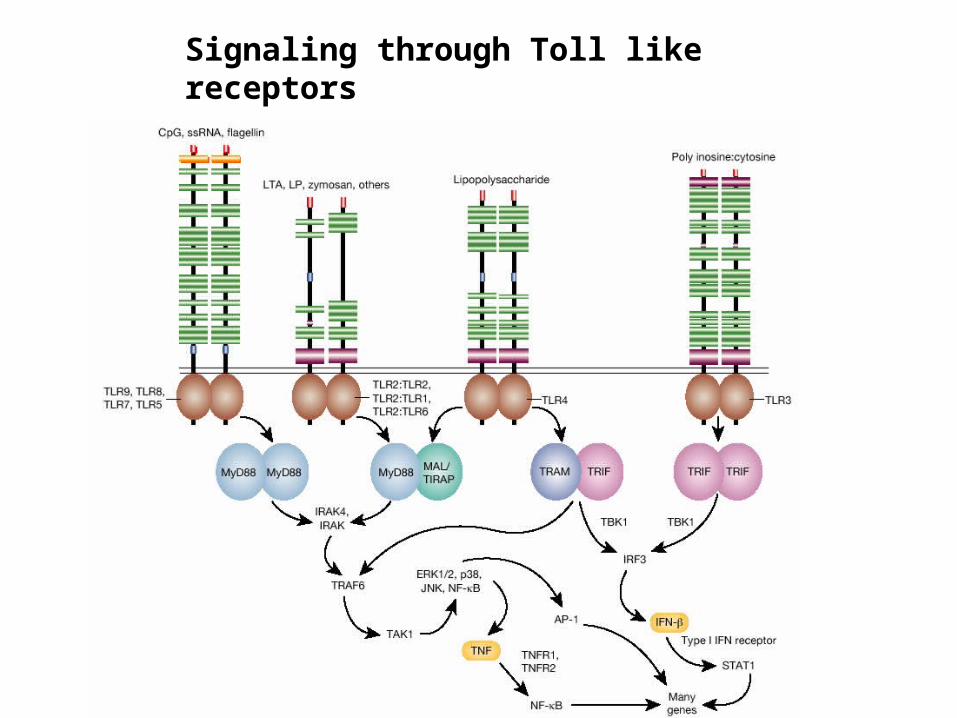
Signaling through Toll like receptors
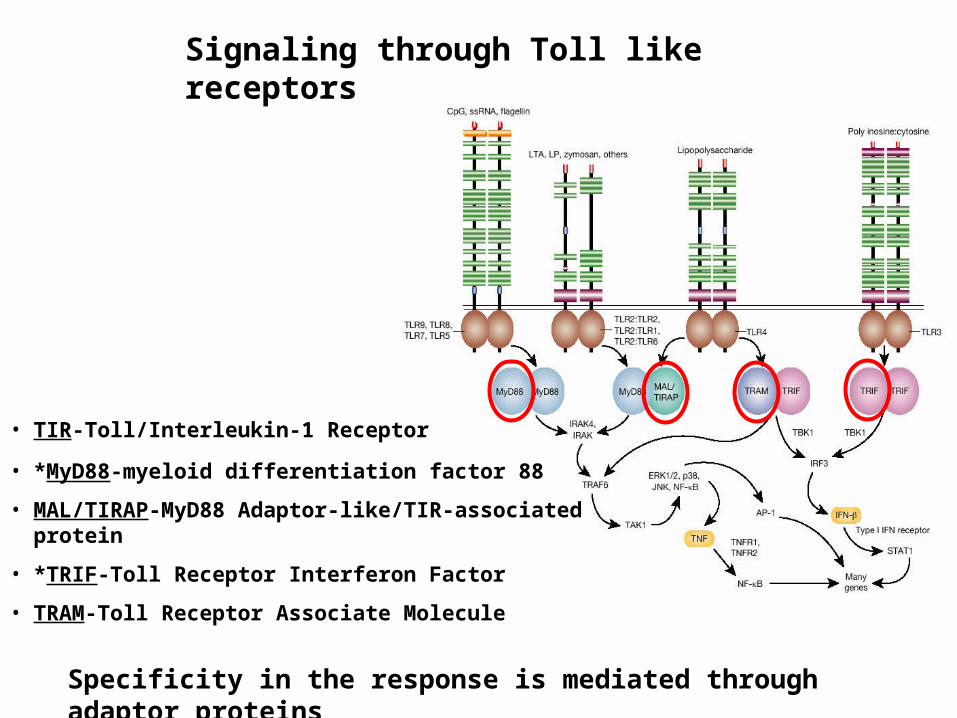
Signaling through Toll like receptors
• TIR-Toll/Interleukin-1 Receptor
• *MyD88-myeloid differentiation factor 88
• MAL/TIRAP-MyD88 Adaptor-like/TIR-associated protein
• *TRIF-Toll Receptor Interferon Factor
• TRAM-Toll Receptor Associate Molecule
Specificity in the response is mediated through adaptor proteins
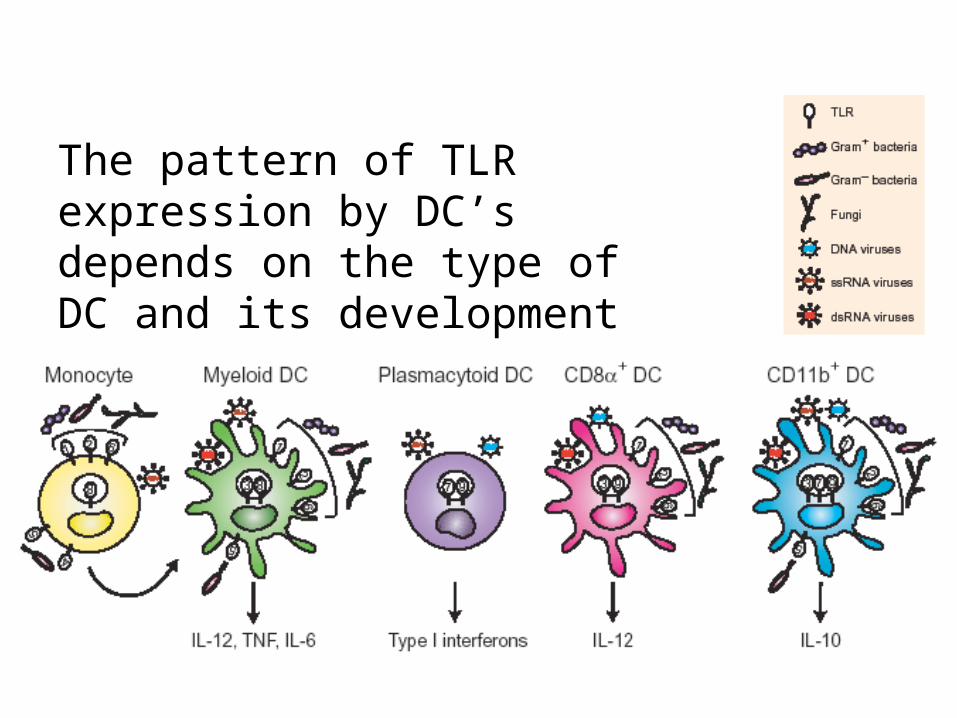
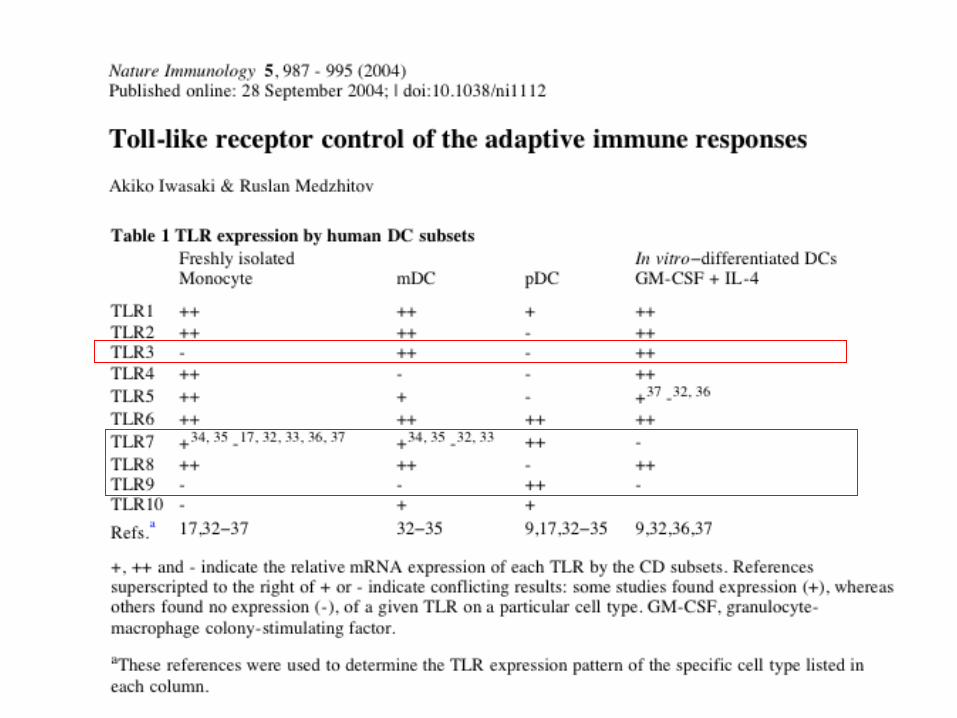
The pattern of TLR expression by DC’s depends on the type of DC and its development

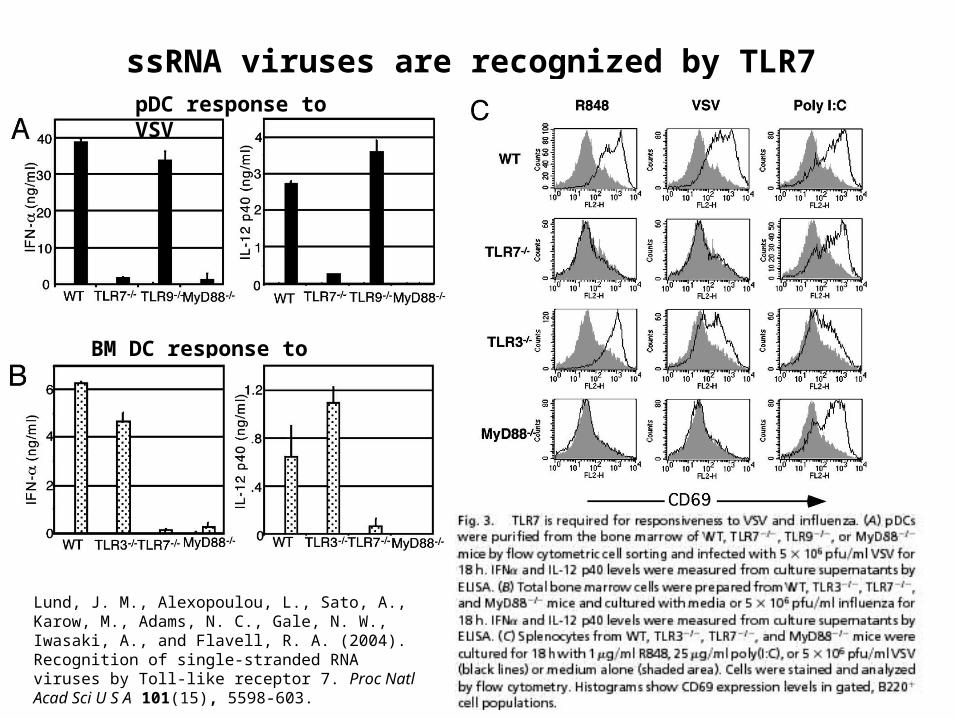
Lund, J. M., Alexopoulou, L., Sato, A., Karow, M., Adams, N. C., Gale, N. W., Iwasaki, A., and Flavell, R. A. (2004). Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc Natl Acad Sci U S A 101(15), 5598-603.
ssRNA viruses are recognized by TLR7pDC response to VSV
BM DC response to influenza
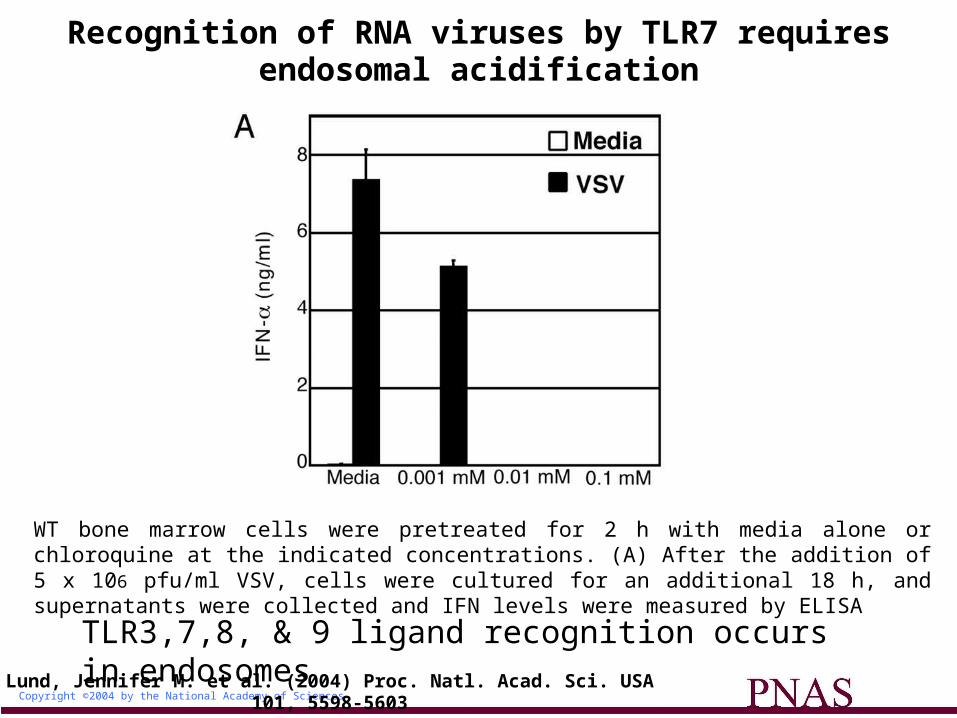
Copyright ©2004 by the National Academy of Sciences
Lund, Jennifer M. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 5598-5603
Recognition of RNA viruses by TLR7 requires endosomal acidification
WT bone marrow cells were pretreated for 2 h with media alone or chloroquine at the indicated concentrations. (A) After the addition of 5 x 106 pfu/ml VSV, cells were cultured for an additional 18 h, and supernatants were collected and IFN levels were measured by ELISA
TLR3,7,8, & 9 ligand recognition occurs in endosomes
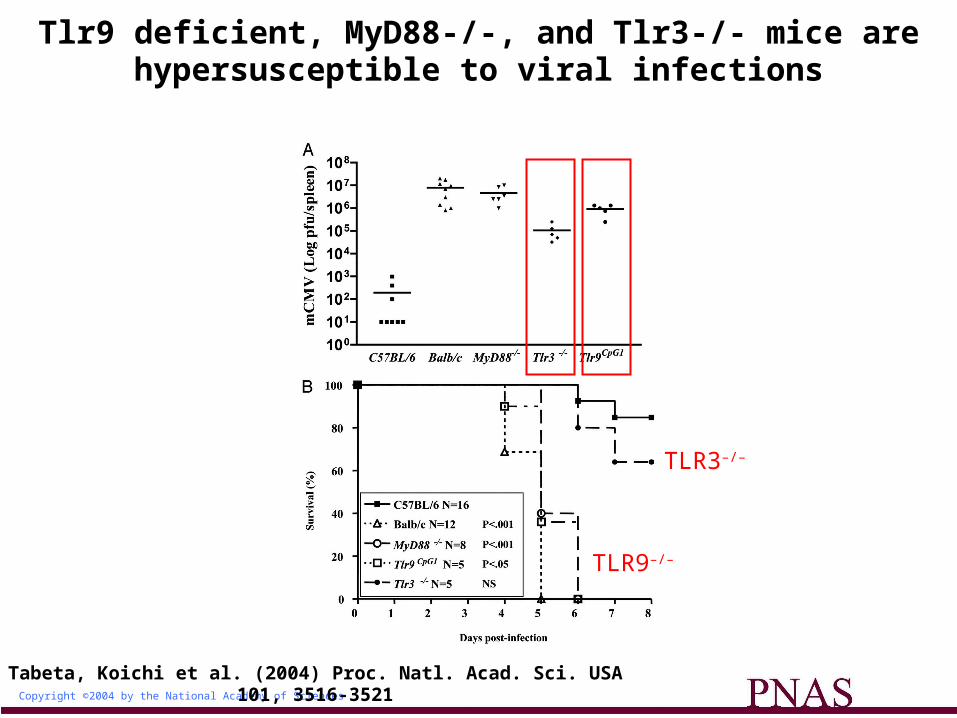
Copyright ©2004 by the National Academy of Sciences
Tabeta, Koichi et al. (2004) Proc. Natl. Acad. Sci. USA 101, 3516-3521
Tlr9 deficient, MyD88-/-, and Tlr3-/- mice are hypersusceptible to viral infections
TLR3–/–
TLR9–/–
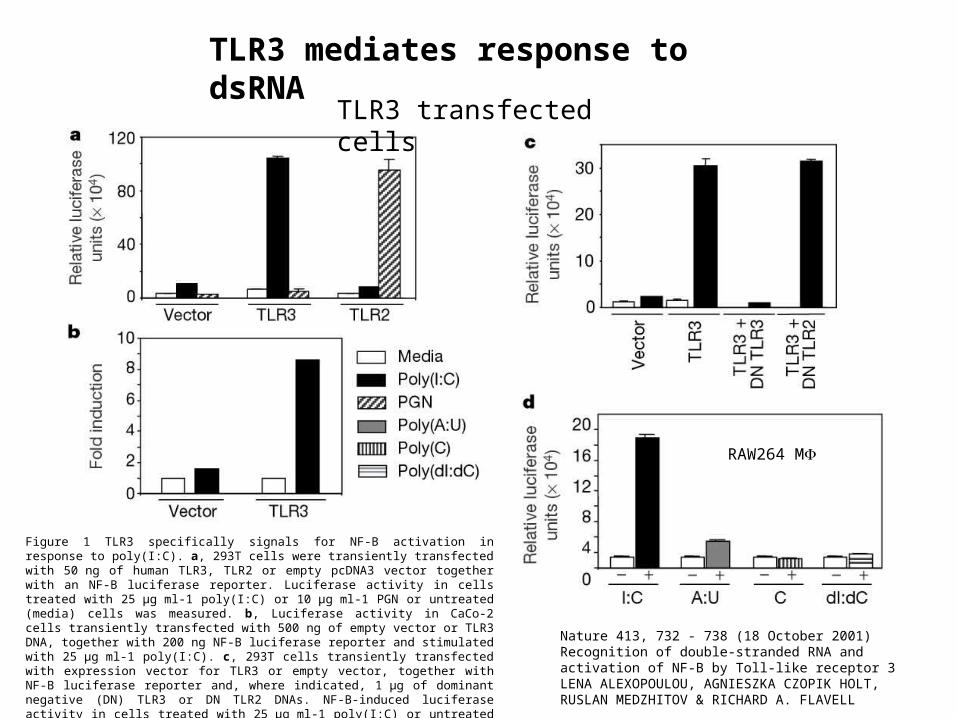
Figure 1 TLR3 specifically signals for NF-B activation in response to poly(I:C). a, 293T cells were transiently transfected with 50 ng of human TLR3, TLR2 or empty pcDNA3 vector together with an NF-B luciferase reporter. Luciferase activity in cells treated with 25 µg ml-1 poly(I:C) or 10 µg ml-1 PGN or untreated (media) cells was measured. b, Luciferase activity in CaCo-2 cells transiently transfected with 500 ng of empty vector or TLR3 DNA, together with 200 ng NF-B luciferase reporter and stimulated with 25 µg ml-1 poly(I:C). c, 293T cells transiently transfected with expression vector for TLR3 or empty vector, together with NF-B luciferase reporter and, where indicated, 1 µg of dominant negative (DN) TLR3 or DN TLR2 DNAs. NF-B-induced luciferase activity in cells treated with 25 µg ml-1 poly(I:C) or untreated cells was measured. d, Transfection of RAW 264.7 macrophages with a NF-B luciferase reporter. Luciferase activity in cells treated with 20 µg ml-1 poly(I:C), poly(A:U), poly(C) or poly(dI:dC), or untreated cells.
TLR3 transfected cells
RAW264 M
TLR3 mediates response to dsRNA
Nature 413, 732 - 738 (18 October 2001) Recognition of double-stranded RNA and activation of NF-B by Toll-like receptor 3 LENA ALEXOPOULOU, AGNIESZKA CZOPIK HOLT, RUSLAN MEDZHITOV & RICHARD A. FLAVELL
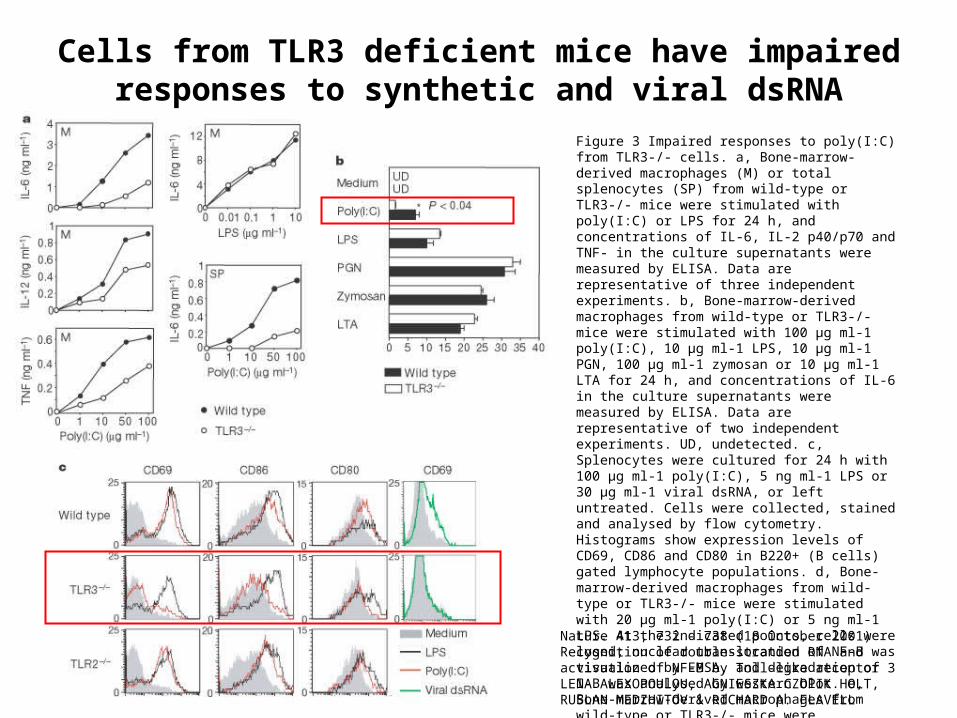
Cells from TLR3 deficient mice have impaired responses to synthetic and viral dsRNA
Figure 3 Impaired responses to poly(I:C) from TLR3-/- cells. a, Bone-marrow-derived macrophages (M) or total splenocytes (SP) from wild-type or TLR3-/- mice were stimulated with poly(I:C) or LPS for 24 h, and concentrations of IL-6, IL-2 p40/p70 and TNF- in the culture supernatants were measured by ELISA. Data are representative of three independent experiments. b, Bone-marrow-derived macrophages from wild-type or TLR3-/- mice were stimulated with 100 µg ml-1 poly(I:C), 10 µg ml-1 LPS, 10 µg ml-1 PGN, 100 µg ml-1 zymosan or 10 µg ml-1 LTA for 24 h, and concentrations of IL-6 in the culture supernatants were measured by ELISA. Data are representative of two independent experiments. UD, undetected. c, Splenocytes were cultured for 24 h with 100 µg ml-1 poly(I:C), 5 ng ml-1 LPS or 30 µg ml-1 viral dsRNA, or left untreated. Cells were collected, stained and analysed by flow cytometry. Histograms show expression levels of CD69, CD86 and CD80 in B220+ (B cells) gated lymphocyte populations. d, Bone-marrow-derived macrophages from wild-type or TLR3-/- mice were stimulated with 20 µg ml-1 poly(I:C) or 5 ng ml-1 LPS. At the indicated points, cells were lysed; nuclear translocation of NF-B was visualized by EMSA, and degradation of I-B was analysed by western blot. e, Bone-marrow-derived macrophages from wild-type or TLR3-/- mice were stimulated with 100 µg ml-1 poly(I:C) or 1 µg ml-1 LPS, or left untreated. After 4 h, total RNA was isolated and expression of IFN-, IFN- and HPRT was determined by RT-PCR
Nature 413, 732 - 738 (18 October 2001) Recognition of double-stranded RNA and activation of NF-B by Toll-like receptor 3 LENA ALEXOPOULOU, AGNIESZKA CZOPIK HOLT, RUSLAN MEDZHITOV & RICHARD A. FLAVELL
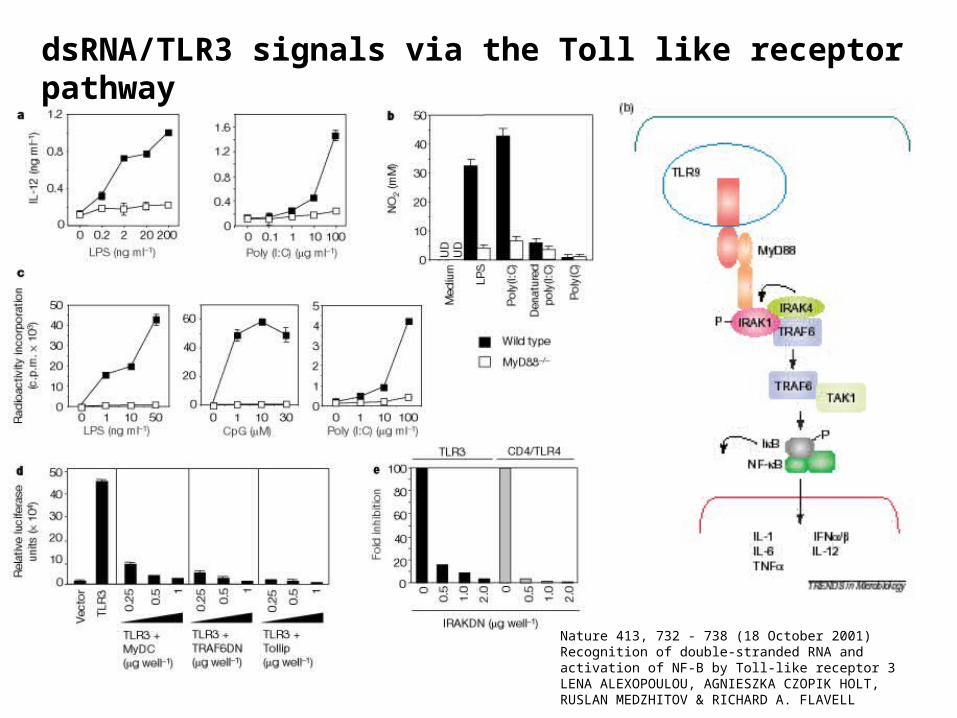
Nature 413, 732 - 738 (18 October 2001) Recognition of double-stranded RNA and activation of NF-B by Toll-like receptor 3 LENA ALEXOPOULOU, AGNIESZKA CZOPIK HOLT, RUSLAN MEDZHITOV & RICHARD A. FLAVELL
dsRNA/TLR3 signals via the Toll like receptor pathway
Requires:
MyD88
TRAF6
Tollip
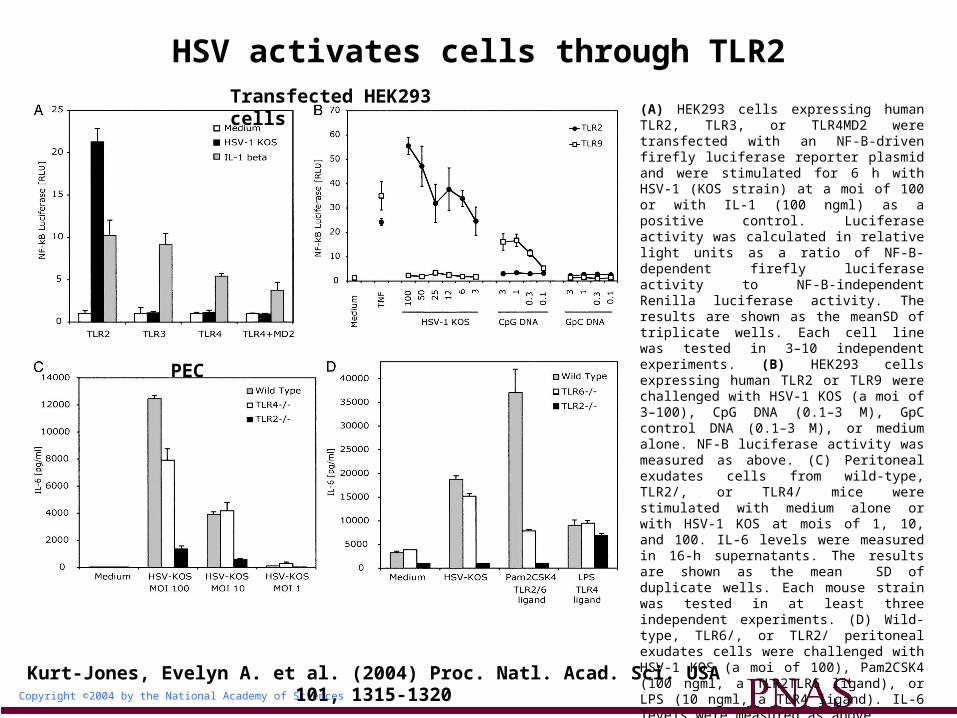
Copyright ©2004 by the National Academy of Sciences
Kurt-Jones, Evelyn A. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 1315-1320
HSV activates cells through TLR2
(A) HEK293 cells expressing human TLR2, TLR3, or TLR4MD2 were transfected with an NF-B-driven firefly luciferase reporter plasmid and were stimulated for 6 h with HSV-1 (KOS strain) at a moi of 100 or with IL-1 (100 ngml) as a positive control. Luciferase activity was calculated in relative light units as a ratio of NF-B-dependent firefly luciferase activity to NF-B-independent Renilla luciferase activity. The results are shown as the meanSD of triplicate wells. Each cell line was tested in 3–10 independent experiments. (B) HEK293 cells expressing human TLR2 or TLR9 were challenged with HSV-1 KOS (a moi of 3–100), CpG DNA (0.1–3 M), GpC control DNA (0.1–3 M), or medium alone. NF-B luciferase activity was measured as above. (C) Peritoneal exudates cells from wild-type, TLR2/, or TLR4/ mice were stimulated with medium alone or with HSV-1 KOS at mois of 1, 10, and 100. IL-6 levels were measured in 16-h supernatants. The results are shown as the mean SD of duplicate wells. Each mouse strain was tested in at least three independent experiments. (D) Wild-type, TLR6/, or TLR2/ peritoneal exudates cells were challenged with HSV-1 KOS (a moi of 100), Pam2CSK4 (100 ngml, a TLR2TLR6 ligand), or LPS (10 ngml, a TLR4 ligand). IL-6 levels were measured as above.
Transfected HEK293 cells
PEC
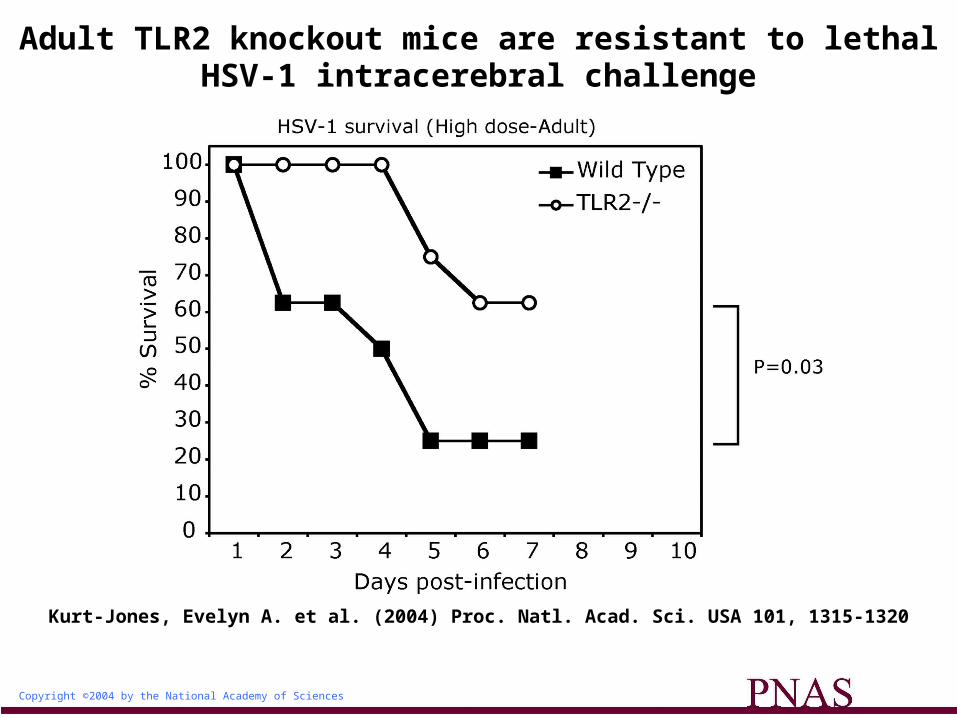
Copyright ©2004 by the National Academy of Sciences
Kurt-Jones, Evelyn A. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 1315-1320
Adult TLR2 knockout mice are resistant to lethal HSV-1 intracerebral challenge
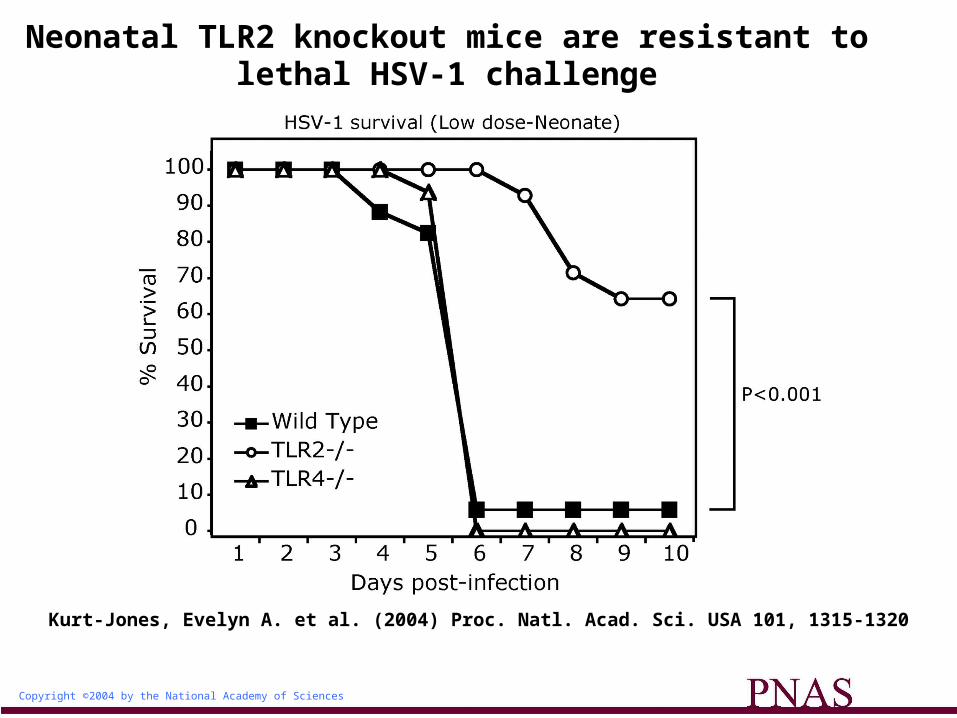
Copyright ©2004 by the National Academy of Sciences
Kurt-Jones, Evelyn A. et al. (2004) Proc. Natl. Acad. Sci. USA 101, 1315-1320
Neonatal TLR2 knockout mice are resistant to lethal HSV-1 challenge

Toll like receptors
• Innate recognition of pathogen-specific molecular motifs
• Initiation of inflammatory response– IL6, IFN-, IL8, IL12, chemokines, IL1, etc.
• Maturation of dendritic cells– Promote activation of naïve T cells– Promote Th1 like responses
• Initiate antiviral IFN- response

Limiting viral replication
• Type I interferons: IFN-, IFN-– Direct inhibition of viral gene expression
– Activate NK cells – Many viral evasion strategies target interferon
• Ex. Influenza NS1 protein (Influenza A Virus Lacking the NS1 Gene Replicates in Interferon-Deficient Systems Adolfo Garcı´a-Sastre,*,1 Andrej Egorov,†,2 Demetrius Matassov,* Sabine Brandt,†,‡ David E. Levy,§ Joan E. Durbin,¶ Peter Palese,*,1 and Thomas Muster†,‡ VIROLOGY 252, 324–330 (1998)
– Promotes Th1 response

IFN-
– Released in response to TLR ligation by viral products (ssRNS, dsRNA, glycoproteins)
– Acts in paracrine and autocrine fashion– Inhibit viral gene expression/replication
• Jak1/Stat-1 upregulation of 2’5’oligoadenylate synthetase which polymerizes ATP and activates an endoribonuclease
• P1 Kinase activation causes phosphorylation of eIF2• Induces MxA required for influenza resistance
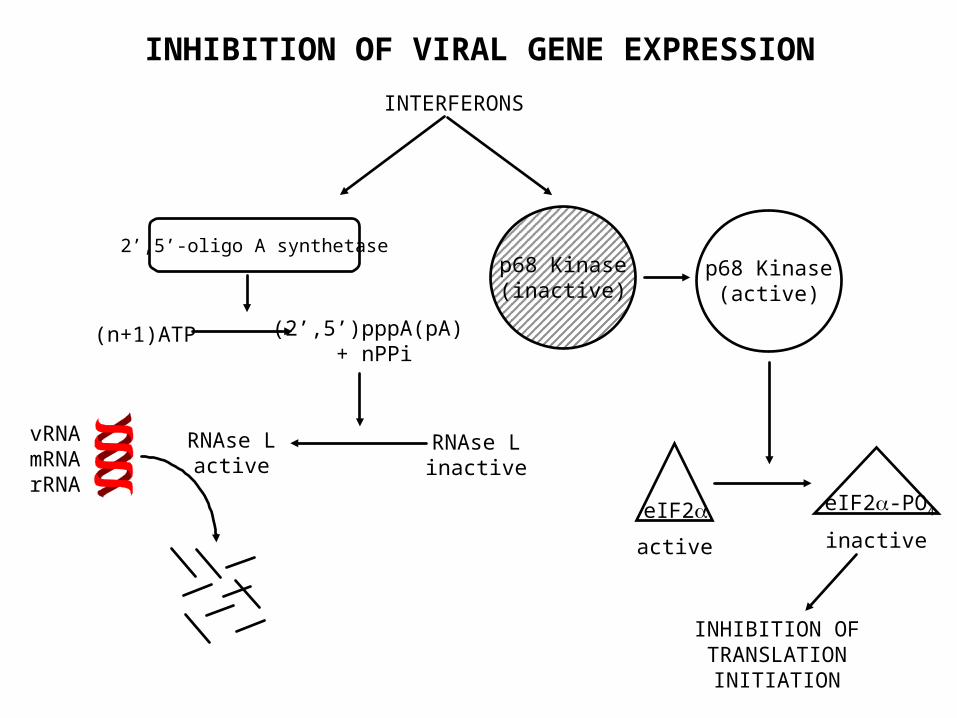
INTERFERONS
p68 Kinase(inactive)
p68 Kinase(active)
eIF2 eIF2-PO4
active inactive
INHIBITION OF TRANSLATION
INITIATION
RNAse Linactive
2’,5’-oligo A synthetase
(n+1)ATP (2’,5’)pppA(pA) + nPPi
RNAse Lactive
vRNAmRNArRNA
INHIBITION OF VIRAL GENE EXPRESSION
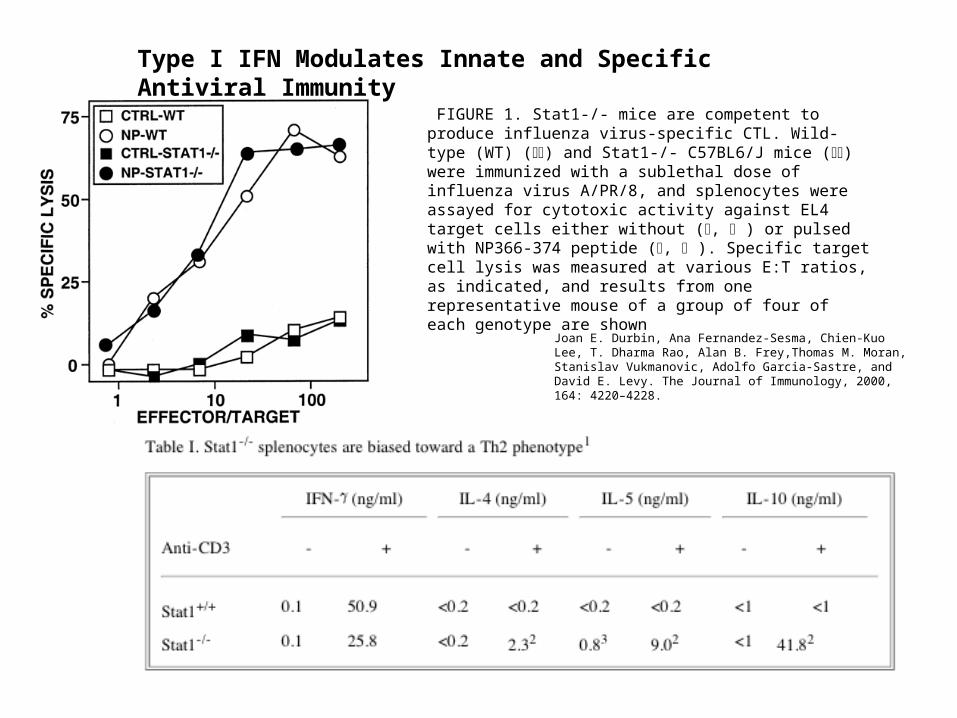
Type I IFN Modulates Innate and Specific Antiviral Immunity
FIGURE 1. Stat1-/- mice are competent to produce influenza virus-specific CTL. Wild-type (WT) () and Stat1-/- C57BL6/J mice () were immunized with a sublethal dose of influenza virus A/PR/8, and splenocytes were assayed for cytotoxic activity against EL4 target cells either without (, ) or pulsed with NP366-374 peptide (, ). Specific target cell lysis was measured at various E:T ratios, as indicated, and results from one representative mouse of a group of four of each genotype are shown
Joan E. Durbin, Ana Fernandez-Sesma, Chien-Kuo Lee, T. Dharma Rao, Alan B. Frey,Thomas M. Moran, Stanislav Vukmanovic, Adolfo Garcia-Sastre, and David E. Levy. The Journal of Immunology, 2000, 164: 4220–4228.
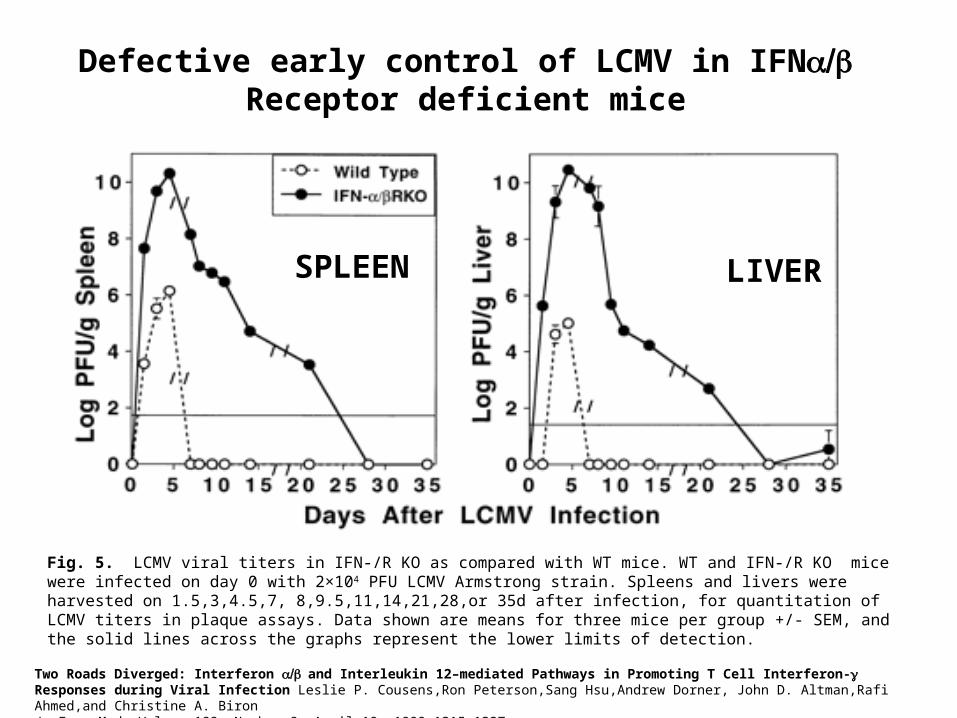
Two Roads Diverged: Interferon and Interleukin 12–mediated Pathways in Promoting T Cell Interferon- Responses during Viral Infection Leslie P. Cousens,Ron Peterson,Sang Hsu,Andrew Dorner, John D. Altman,Rafi Ahmed,and Christine A. BironJ. Exp. Med. Volume 189, Number 8, April 19, 1999 1315–1327
SPLEEN LIVER
Fig. 5. LCMV viral titers in IFN-/R KO as compared with WT mice. WT and IFN-/R KO mice were infected on day 0 with 2×104 PFU LCMV Armstrong strain. Spleens and livers were harvested on 1.5,3,4.5,7, 8,9.5,11,14,21,28,or 35d after infection, for quantitation of LCMV titers in plaque assays. Data shown are means for three mice per group +/- SEM, and the solid lines across the graphs represent the lower limits of detection.
Defective early control of LCMV in IFN Receptor deficient mice
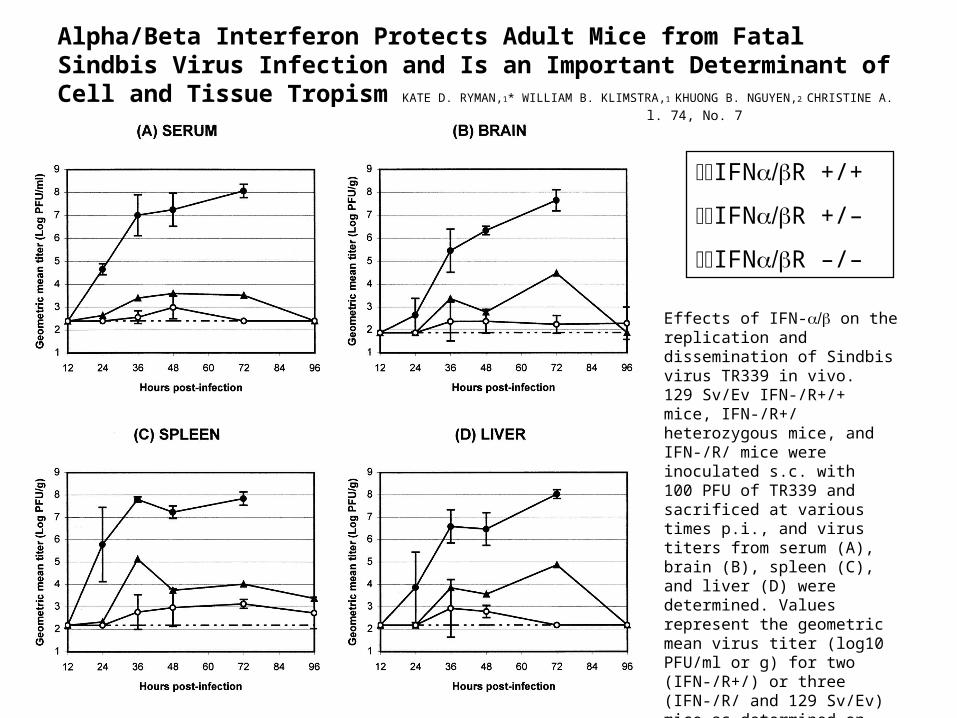
Alpha/Beta Interferon Protects Adult Mice from Fatal Sindbis Virus Infection and Is an Important Determinant of Cell and Tissue Tropism KATE D.
RYMAN,1* WILLIAM B. KLIMSTRA,1 KHUONG B. NGUYEN,2 CHRISTINE A. BIRON,2 AND ROBERT E. JOHNSTON1 JOURNAL OF VIROLOGY, Apr. 2000, p. 3366–3378 Vol. 74, No. 7
IFNR +/+
IFNR +/–
IFNR –/–
Effects of IFN- on the replication and dissemination of Sindbis virus TR339 in vivo. 129 Sv/Ev IFN-/R+/+ mice, IFN-/R+/ heterozygous mice, and IFN-/R/ mice were inoculated s.c. with 100 PFU of TR339 and sacrificed at various times p.i., and virus titers from serum (A), brain (B), spleen (C), and liver (D) were determined. Values represent the geometric mean virus titer (log10 PFU/ml or g) for two (IFN-/R+/) or three (IFN-/R/ and 129 Sv/Ev) mice as determined on BHK cells. Datum points are shown ± the standard deviation (SD), where n = 3. The lower limit of detection is indicated (broken line).
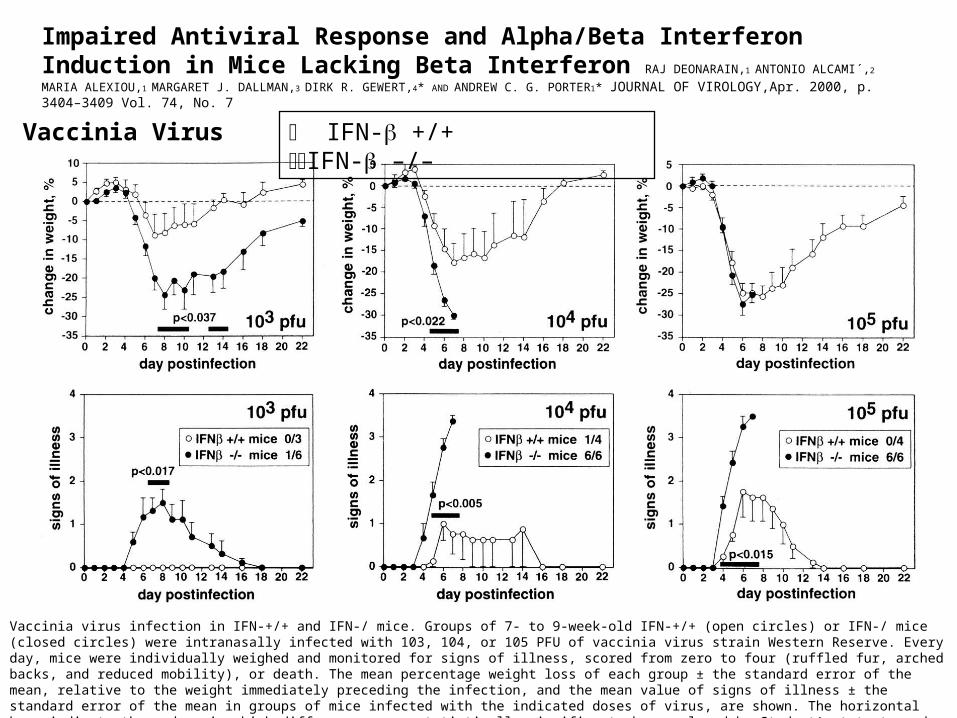
Impaired Antiviral Response and Alpha/Beta Interferon Induction in Mice Lacking Beta Interferon RAJ DEONARAIN,1 ANTONIO ALCAMI´,2 MARIA ALEXIOU,1 MARGARET J. DALLMAN,3 DIRK R. GEWERT,4*
AND ANDREW C. G. PORTER1* JOURNAL OF VIROLOGY,Apr. 2000, p. 3404–3409 Vol. 74, No. 7
IFN- +/+ IFN- –/–Vaccinia Virus
Vaccinia virus infection in IFN-+/+ and IFN-/ mice. Groups of 7- to 9-week-old IFN-+/+ (open circles) or IFN-/ mice (closed circles) were intranasally infected with 103, 104, or 105 PFU of vaccinia virus strain Western Reserve. Every day, mice were individually weighed and monitored for signs of illness, scored from zero to four (ruffled fur, arched backs, and reduced mobility), or death. The mean percentage weight loss of each group ± the standard error of the mean, relative to the weight immediately preceding the infection, and the mean value of signs of illness ± the standard error of the mean in groups of mice infected with the indicated doses of virus, are shown. The horizontal bars indicate those days in which differences were statistically significant when analyzed by Student's t test, and the P values are shown. The number of mice per group that either died or were sacrificed due to severe infection is shown in the insets.
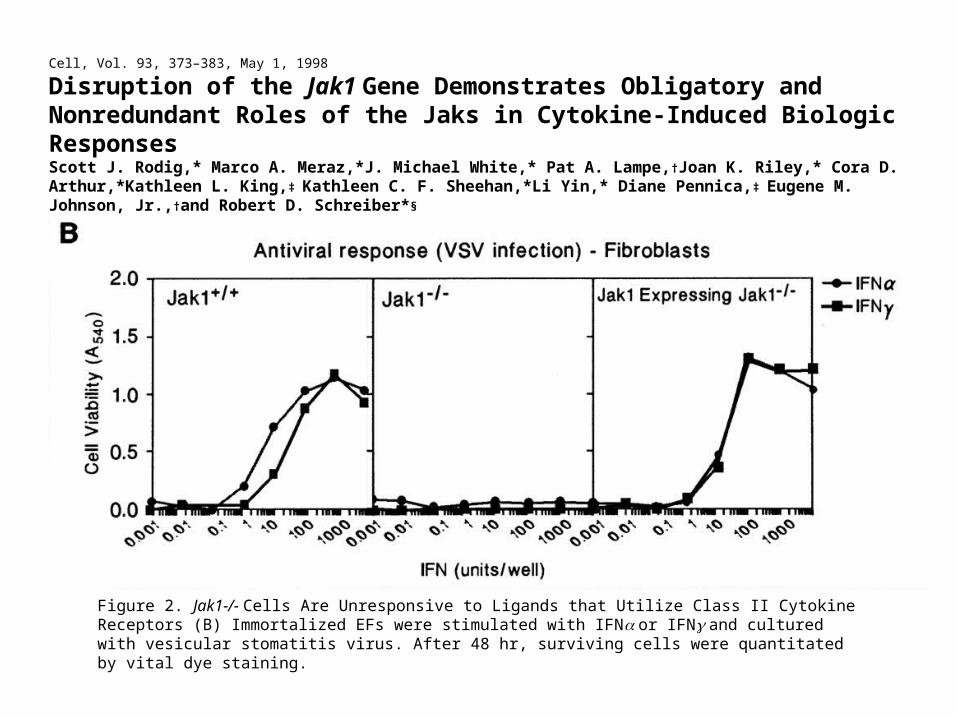
Cell, Vol. 93, 373–383, May 1, 1998
Disruption of the Jak1 Gene Demonstrates Obligatory and Nonredundant Roles of the Jaks in Cytokine-Induced Biologic ResponsesScott J. Rodig,* Marco A. Meraz,*J. Michael White,* Pat A. Lampe,†Joan K. Riley,* Cora D. Arthur,*Kathleen L. King,‡ Kathleen C. F. Sheehan,*Li Yin,* Diane Pennica,‡ Eugene M. Johnson, Jr.,†and Robert D. Schreiber*§
Figure 2. Jak1-/- Cells Are Unresponsive to Ligands that Utilize Class II Cytokine Receptors (B) Immortalized EFs were stimulated with IFN or IFN and cultured with vesicular stomatitis virus. After 48 hr, surviving cells were quantitated by vital dye staining.

Type I IFNIFN-
• Limits viral gene expression
• Makes uninfected cells resistant to infection
• Promotes Th1/IFN- response
• Causes attrition of memory T cells and then promotes memory T cell expansion

Figure 2-48

Innate recognition of infected cells by Natural Killer (NK)
cells

NK and NKT cells• NK cells: (Biron et al. Annual Review of Immunology Apr 1999, Vol. 17: 189-220)
– CD56+ (human) NK1.1+ (mouse)– DX5+ (CD49b/VLA-2) mouse (human?)
• NKT cells: (Bendelac et al. Annual Review of Immunology Apr 1997, Vol. 15: 535-562)
– Restricted TCR repertoire (V14, V2,7,8)– CD4+ (60%) or double negative (40%)– NK1.1+– Recognize CD1 non-classical MHC class-I-like
• Presents lipids (Porcelli et al Annu. Rev. Immunol. 1999. 17:297–329)
• Annu. Rev. Immunol. 2004. 22:405–29 THE DYNAMIC LIFE OF NATURAL KILLER CELLS Wayne M. Yokoyama, Sungjin Kim, and Anthony R. French

NK receptors
• Killer Ig-like receptors (KIR)
• Lectin-like receptors (CD94/NKG2, NKG2D, Ly49)
• ITIMs: immunoreceptor tyrosine-based inhibitory motifs
• ITAMs: immunoreceptor tyrosine-based activation motifs– Also present on adaptor proteins that bind to ITIM-less receptors,
ex. DAP12
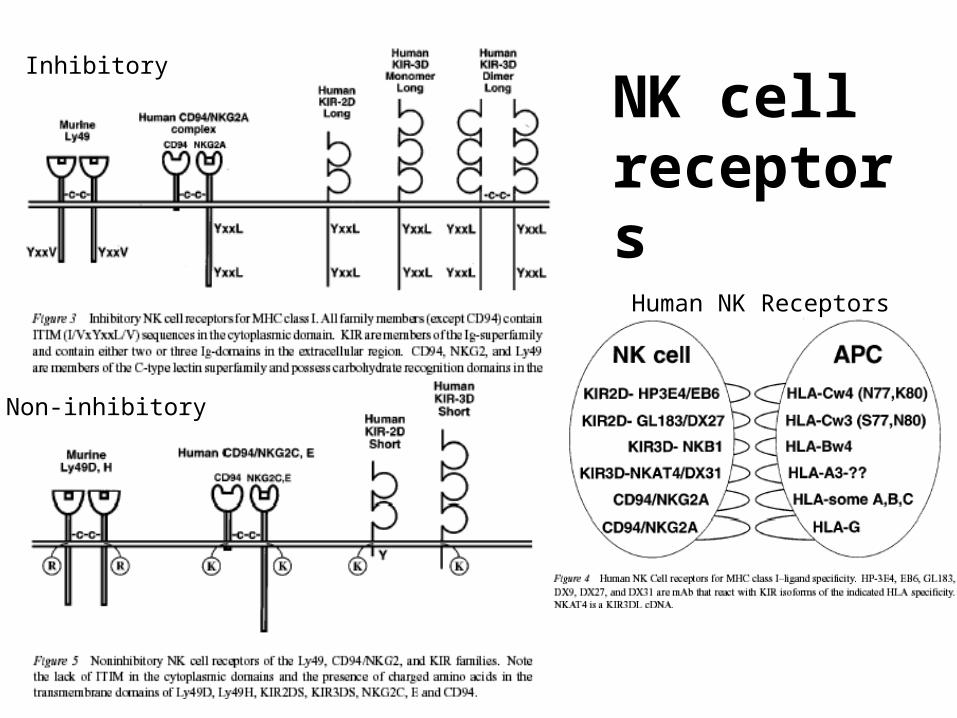
NK cell receptors
Inhibitory
Non-inhibitory
Human NK Receptors
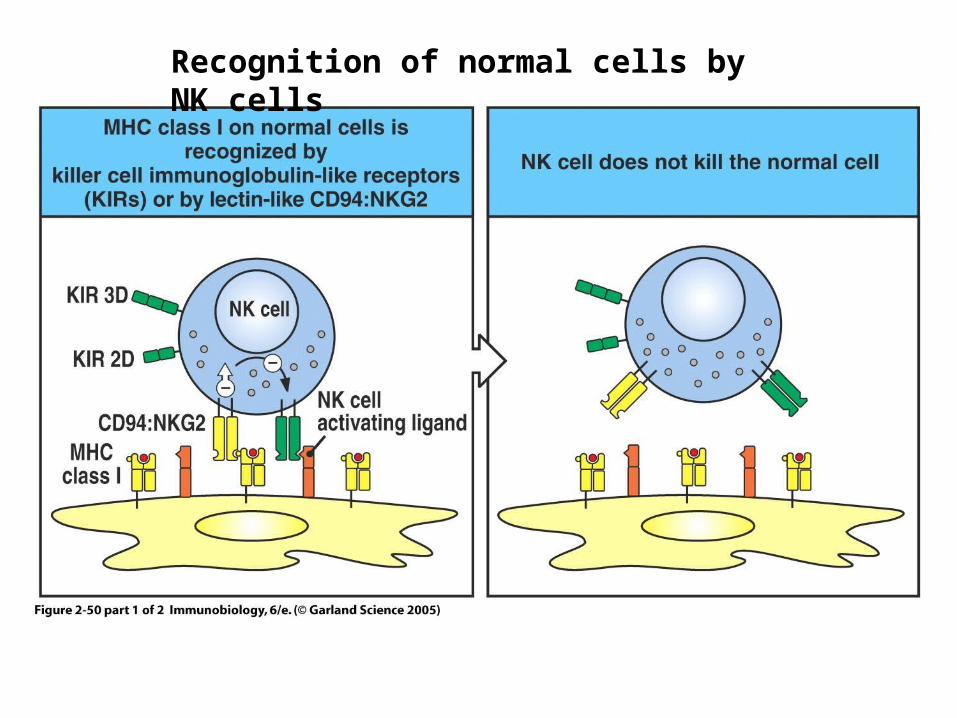
Figure 2-50 part 1 of 2Recognition of normal cells by NK cells
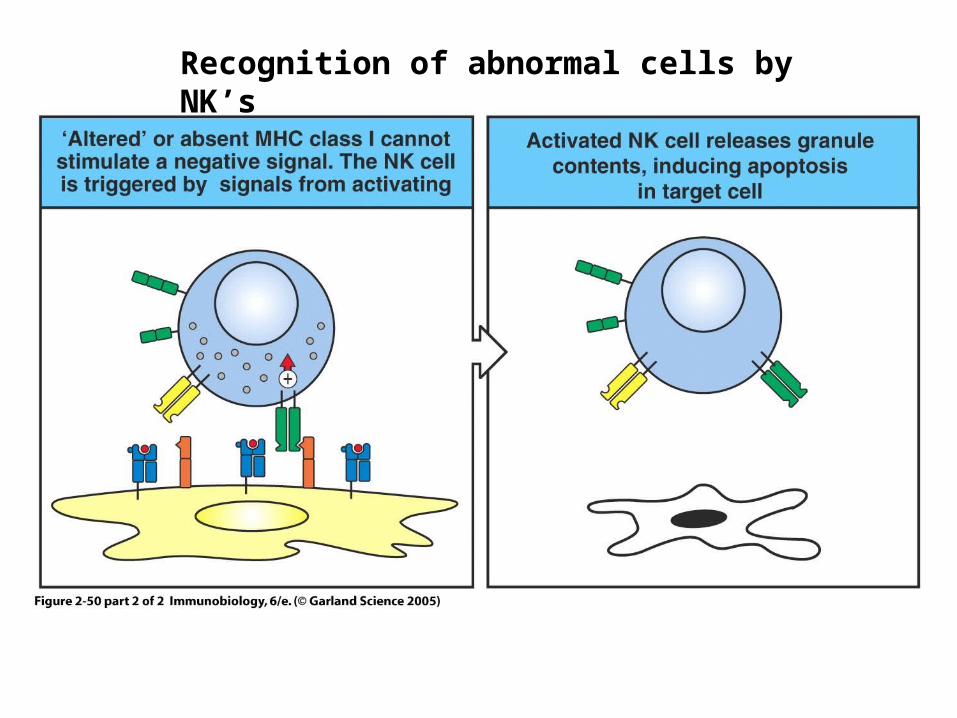
Figure 2-50 part 2 of 2Recognition of abnormal cells by NK’s

Missing-self hypothesis
• Downregulation of MHC molecules on infected cells triggers NK cell lysis
• Dependent on absence of MHC signaling through NK inhibitory receptors
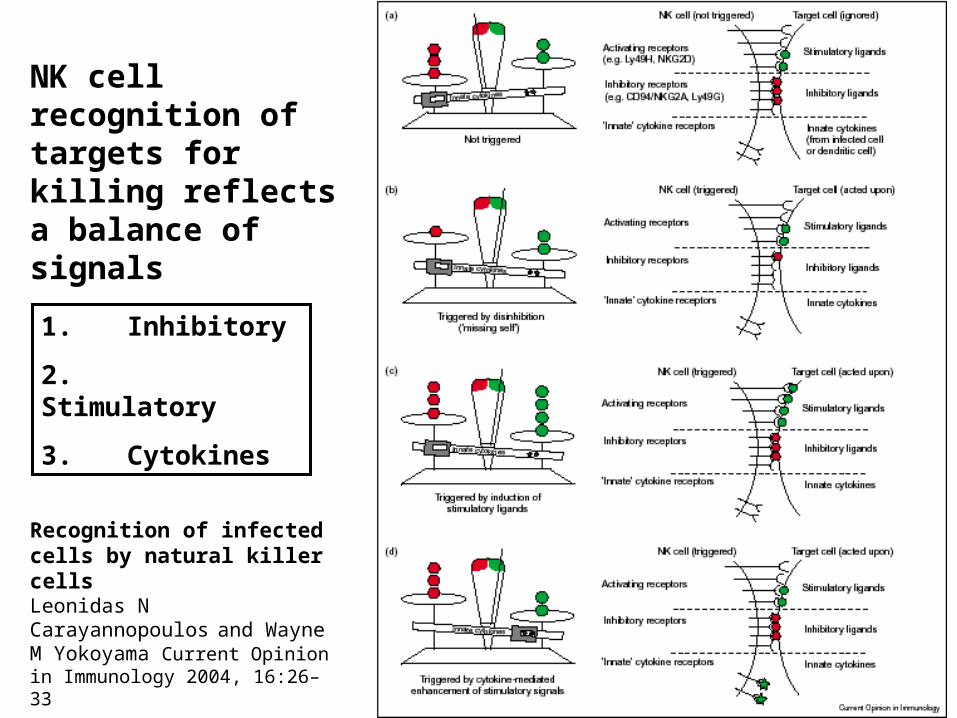
Recognition of infected cells by natural killer cellsLeonidas N Carayannopoulos
and Wayne M Yokoyama Current Opinion in Immunology 2004, 16:26–33
NK cell recognition of targets for killing reflects a balance of signals
1. Inhibitory
2. Stimulatory
3. Cytokines

(B) MCMV replication in F1 hybrid offspring from DBA/2 and BXD-8. Three days after infection [with MCMV Smith strain,2-3x104 plaque-forming units (PFUs)], organ viral titers were assessed in tissue homogenates collected from C57BL/6, DBA/2, BXD-8, and (DBA/2 3 BXD-8)F1 hybrid mice (five mice per group), as indicated. Spleen titers are shown here; liver titers are available online (8). Each point represents the average titer determined for an individual mouse. Mean viral titers for each group are depicted as horizontal bars. For mice with titers below the level of detection of the assay, the minimum number of detectable PFUs (102) was assumed to determine the mean.
Ly49H deficiencyLy49H NK1.1
MCMV
Ly49D
Anti-Ly49c
Anti-Ly49h
Anti-NK1.1
Science. 2001 May 4;292(5518):934-7.

NK cell recognition of CMV infected targets
• Ly49H activation receptor on NK cells
• M157 gene product from CMV– Class I MHC like molecule expressed on infected
cells– May have evolved to bind Ly49I, an inhibitory
receptor on NK cells
• Innate immune response encoded within the genome

Mechanisms to detect and eliminate virus infected cells
• Antibody dependent cellular cytotoxicity (ADCC)
• Activated macrophage, neutrophils, granulocytes
• NK/NKT cell mediated lysis
• Cytotoxic T cells (CTL)

ADCC
• Antibodies to pathogen or cell surface antigen bind to NK cells via FcRIII receptor
• IgG1, IgG3
• Antibodies bind FcR on macrophage to elicit TNF- secretion

Alternative modes of antibody mediated pathogen toxicity
• IgE/FcRI– Mast cells
• Release histamine, prostaglandins, leukotrienes, TNF-– Basophils
• Histamine and IL4
– Eosinophils• Toxic granule proteins and free radicals
• IgG1,IgG3/FcRIII• Fc
– Macrophage, neutrophils, mast cells, basophils, and eosinophils bind pathogen surface via Fc/Ig and deliver contents of lysosomes into membrane or on surface of pathogen

Antigen-specific recognition of viruses
Antigen processing and presentation through class I and
class II MHC molecules





























