Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Intravenous infusion of GDNF gene-modified human umbilicalcord blood CD34+ cells protects against cerebral ischemicinjury in spontaneously hypertensive rats
Yali Oua, SeongJin Yub, Yuji Kanekob, Naoki Tajirib, Eunkyung Cate Baeb,Sonia H. Chhedab, Christine E. Stahlc, Tianlun Yanga, Li Fanga, Kei Hua,Cesar V. Borlonganb,⁎, Guolong Yua,⁎aDepartment of Cardiology, Xiangya Hospital, Southern Central University, Changsha 410008, ChinabCenter of Excellence for Aging and Brain Repair, Department of Neurosurgery and Brain Repair,University of South Florida College of Medicine, Tampa, FL 33612, USAcDepartment of Flight Medicine, MacDill Air Force Base, Tampa, FL 33621, USA
A R T I C L E I N F O
⁎ Corresponding authors. G. Yu is to be contaE-mail addresses: [email protected]
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.09.098
A B S T R A C T
Article history:Accepted 24 September 2010Available online 1 October 2010
This study assessed the potential of intravenous transplantation of human umbilical cordblood (HUCB) CD34+ cells transfected with glial cell line-derived neurotrophic factor (GDNF)gene to exert therapeutic benefits in spontaneous hypertensive rats (SHR) exposed totransientmiddle cerebral artery occlusion (MCAO). SHRwithMCAOwere randomly assignedto receive intravenously transplantation of vehicle, the plasmid containing the enhancedgreen fluorescent protein (pEGFP)-CD34+ cells or pEGFP-GDNF-CD34+ cells at 6 h afterstroke. The CD34+ cells transfected with GDNF gene expressed higher levels of GDNF mRNAand protein than nontransfected HUCB CD34+ cells in vitro. At 28 days after transplantationof GDNF gene modified CD34+ cells, significantly more GFP positive cells, neurons, andastrocytes, likely derived from the grafted cells, populated the peri-infarct area compared tothose injected with pEGFP-CD34+ cells or vehicle. Furthermore, the stroke animalstransplanted with GDNF gene modified CD34+ cells showed a significant increase inGDNF level in the infarcted hemisphere, reduced brain infarction volume, and enhancedfunctional recovery compared with those that received pEGFP-CD34+ cells. This studysupports the use of a combined gene and stem cell therapy for treating stroke.
© 2010 Elsevier B.V. All rights reserved.
Keywords:StrokeNeurotrophic factorStem cellGene therapyTransplantation
1. Introduction
There is growing scientific evidence demonstrating thatsystemically transplanted human umbilical cord blood(HUCB) cells promote behavioral recovery in preclinical strokemodels (Boltze et al., 2005; Borlongan et al., 2004; Chen et al.,
cted at Fax: +86 731 432 73u (C.V. Borlongan), yuguo
er B.V. All rights reserved
2001; Makinen et al., 2006; Nystedt et al., 2006; Taguchi et al.,2004; Vendrame et al., 2004; Willing et al., 2003; Xiao et al.,2005). Previous studies also demonstrated that systemicadministration of HUCB cells increased the levels of trophicfactors including glial cell-derived neurotrophic factor (GDNF),nerve growth factor (NGF), brain-derived neurotrophic factor
32. C.V. Borlongan, Fax: +1 813 974 [email protected] (G. Yu).
.

218 B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
(BDNF), vascular endothelial growth factor (VEGF), and fibro-blast growth factor (FGF) in the ischemic brain (Borlonganet al., 2004; Lobel et al., 2003; Taguchi et al., 2004). In particular,our recent study showed that experimentally stroke animalsthat received HUCB cell grafts, which were pre-exposed to acocktail of antibodies against GDNF, BDNF, NGF, failed toexhibit any measurable neuroprotection of behavioral recov-ery (Borlongan et al., 2004). In addition, GDNF and BDNFdirectly administered by intracranial route promote neuro-protective effects in ischemic animals (Chen et al., 2000;Kobayashi et al., 2006). Taken together, these data suggest thattropic factors secreted by HUCB cells within the host brainmediate the reduction of ischemic damage and improvementof behavioral recovery following ischemia in recipientanimals.
Delivery of GDNF to the brain reduces cerebral infarctionand improved motor functions in stroke animals (Kobayashiet al., 2006). However, GDNF cannot directly reach to the brainparenchymal lesion after the systemic injection because of theblood–brain barrier. Recent studies have demonstrated thatintravenous administration of GDNF or BDNF gene-modifiedhuman mesenchymal stem cells affords functional recoverycoupled with reduction of infarct volume as revealed bymagnetic resonance imaging analysis in MCAO animals(Horita et al., 2006; Nomura et al., 2005).
That trophic factors critically participate in the therapeuticbenefits of cell transplantation prompted us to examinewhether the effects of HUCB cells in protecting againstischemic injury might be enhanced if the cells were engi-neered to over-express GDNF before transplantation. In thisstudy, we used a lipofection technique via non-viral deliveryof plasmid DNA encoding a rat pGDNF to CD34+ cells derivedfrom HUCB in order to generate cells with enhanced expres-sion of the GDNF gene and secretion of GDNF protein.Subsequently these transfected cells or the control non-transfected cells were intravenously delivered into spontane-ously hypertensive rats in order to reveal potentiation oftherapeutic efficacy by genetic modification of trophic factorsecretion in HUCB cells.
2. Results
2.1. Expression of GDNF mRNA and GDNF protein intransfected CD34+ cells in vitro
We were able to replicate our recent findings that CD34+ cellswere easily transfectable with GDNF gene and that GDNFmRNA and protein were stably expressed by these cells (Yuet al., 2010). About 85%CD-34 positive cells in total HUCBsweresuccessfully transfected with GDNF based on pEGF labeling.Here, we observed that the total RNA extracted from CD34+cells transfected by the plasmid pEGFP/GDNF or pEGFPresembled their amplified products, both demonstrating640 bp obtained by electrophoretic mobility shift assays(EMSA). Moreover, EMSA showed that the expression of GDNFmRNA from pEGFP/GDNF-transfected CD34+ cells was in-creased substantially, compared with pEGFP-transfectedCD34+ cells. Enzyme-linked immunosorbent assay (ELISA)
showed that compared with that of nontransfected or pEGFP-transfected CD34+ cells, the content of GDNF protein in theculture medium of pEGFP/GDNF-transfected CD34+ cells at 6,12, 24 and 48 h were increased significantly, and the peaklevel of GDNF protein was produced at 24 h post-transfection.These results validated our previous report (Yu et al., 2010) thatCD34+ cells transfected with recombinant plasmid pEGFP/GDNF via liposome enhanced the GDNF expression andsecretion. Of note, GDNF mRNA and protein of pEGFP-CD34+HUCBCs did not differ from nontransfected HUCBCs,indicating that GFP labeling did not alter GDNF expressionand secretion.
At 5 days after the plasmid pEGFP/GDNF or pEGFP wastransfected into CD34+ cells, fluorescence microscopyrevealed that the GFP (as a reporter gene) positive cells was18.87±0.30% in CD34+ cells transfected by the plasmidpEGFP, and 19.52±0.81% in CD34+ cells transfected by theplasmid pEGFP/GDNF. There were no significant differencesbetween GFP positive cells transfected by the two differentplasmids.
2.2. Histological determination of infarction volume
As shown in Fig. 1, the infarct volume of the brain on day 7post-transplantation was significantly decreased in strokerats treated with pEGFP-GDNF modified CD34+ cells com-pared to stroke rats with pEGFP-CD34+cells (102.44±23.01 vs.138.35±37.54 mm3, P<0.05) or vehicle (102.44±23.01 vs.193.79±49.28 mm3, P<0.000). In addition, the infarct volumeof the brain was smaller in stroke rats treated with pEGFP-CD34+cells than those that received vehicle infusion(P<0.05).
2.3. GFP epifluorescence microscopy andimmunohistochemical assessment of grafted cells andneurogenesis in the stroke brain
On day 7 after transplantation, a few GFP-positive cells, andfew NeuN or GFAP-positive cells were detected, primarily inthe ischemic hemisphere of stroke animals that receivedpEGFP-GDNF or pEGFP modified CD34+ cells. However, thenumber of GFP-, NeuN- or GFAP-positive cells in the ischemichemisphere was increased on day 28 after transplantation.The number of these GFP-, NeuN- and GFAP-positive cells inthe brains was significantly increased in the stroke animalstreated with pEGFP-GDNF modified CD34+ cells compared tothose transplanted with pEGFP modified CD34+ cells (seeTable 1, Fig. 2). The ratio of NeuN+ to GFAP+ cells wasapproximately 1:4, with examination of more than fivesections per animal in both groups of stroke rats that receivedCD34+ cells. The GFP-, NeuN- and GFAP-positive cells weremainly distributed in peri-infarcted area in the ipsilateralstroke hemisphere, such as striatum, cortex, hippo-campus and subventricular zone. More than 75% of the totalGFAP single labeled cells were double-labeled with GFP-GFAP.Few of these positive cells were detected in the contralateralnon-infarcted hemisphere. No GFP, and very few NeuN- orGFAP-positive cells, if at all detected, were found in thecorresponding peri-infarct area of stroke rats treated withvehicle.

Fig. 1 – Histological examination the infarct volume of the brain (TTC staining) on day 7 after post-transplantation revealed asignificantly reduced cerebral ischemia in stroke rats treated with GDNF gene modified CD34+ cells (A) compared to stroke ratsthat received pEGFP-CD34+cells (B) or vehicle (C) (t tests; P<0.05).
219B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
2.4. ELISA for GDNF levels in brain tissue
The brain levels of GDNF, as revealed by ELISA, on 7 or 28 daysafter transplantation in the infarcted hemisphere of strokerats treated with GFP-GDNF-CD34+ cells were significantlyhigher than those treated with GFP-CD34+ cells (Fig. 3). Inaddition, compared with stroke rats treated with vehicle,GDNF levels in brains of stroke rats in both groups thatreceived CD34+ cells were significantly increased on day 7 or28 after transplantation. Finally, GDNF levels on day 7 in thethree groups were higher than that on day 28.
2.5. Behavioral tests
There were no significant differences in mNSS scores on day 1after transplantation among the three treatment groups (Fig. 4).Rats treatedwith pEGFP-GDNF-CD34+ cells displayed significant-ly better performance on mNSS scores compared with pEGFP-CD34+ cells or vehicle on day 7 and day 28 after transplantation.Compared to rats treatedwith vehicle,mNSS scores on day 7 andday 28 after transplantation were significantly decreased in ratstransplanted with pEGFP-CD34+. Moreover, mNSS scores weresignificantly decreased on day 28 compared to day 7 in ratstreated with pEGFP-GDNF-CD34+ cells.
3. Discussion
The present study demonstrated that intravenous injection ofGDNF gene transfected or untransfected HUCB CD34+ cells at
Table 1 – Comparison of phenotypic expression acrosstreatment groups mean cell counts±SEM).
pEGFP-GDNF-CD34+ cells
pEGFP-CD34+cells
Vehicle
GFP-positive cells 300.6±40.2** 210.9±36.1 0NeuN-positive cells 60.0±12.3* 30.6±11.5 <5GFAP-positive cells 200.1±20.4** 77.5±13.3 <10GFP-GFAP-positivecells
163.8±42.7** 27.8±8.1 0
*P<0.05 and **P<0.001 versus pEGFP-CD34+ cells or vehicle.
6 h after transient MCAO in SHR resulted in increased GDNFlevels in the infarcted cerebral hemisphere that accompaniedreduction in infarction volume and improvement in behav-ioral performance. These results are consistent with previousstudies showing beneficial effects of systemic delivery ofHUCB cells in stroke models (Boltze et al., 2005; Borlonganet al., 2004; Chen et al., 2001; Makinen et al., 2006; Nystedtet al., 2006; Taguchi et al., 2004; Vendrame et al., 2004; Willinget al., 2003; Xiao et al., 2005). This study also showed thatintravenously transplanted HUCB cells could enter the ratbrain (albeit a few cells), migrate to peri-infarct area, surviveand proliferate robustly at 1 week after transplantation.Furthermore, a handful of these grafted HUCB cells coulddifferentiate into astrocytes and neurons in ischemic brain. Ofnote, our data indicated that compared with nontransfectedCD34+ cells, the survival, proliferation and differentiation ofGDNF gene modified CD34+ cells into astrocytes and neuronswere enhanced significantly in ischemic brain, and promoteda much better recovery of functional outcome.
The exact mechanism of functional improvement by celltransplantation in the ischemic brain is unknown. Increasedneurogenesis, neuroprotection by neurotrophic and growthfactors, or new synaptic formation with CNS reorganization hasbeen implicated as potent cell repair processes (Yu et al., 2009a,b;Borlongan et al., 2004; Chen et al., 2001; Ding et al., 2006; Willinget al., 2003).Whether these same repairmechanismsmediate theobserved neuroprotective effects by GDNF gene modified CD34+cells in our study remain to be determined. HUCB cells have beendemonstrated to secrete a variety of neurotrophic factors such asGDNF, NGF, BDNF, VEGF, and FGF (Yu et al., 2009a,b; Borlonganet al., 2004; Chen et al., 2001; Ding et al., 2006;Willing et al., 2003).While numerous secreted trophic factors could contribute toneuroprotection, our study modified HUCB cells to over-expressonly their level of GDNF secretion. The GDNF gene wastransfected into HUCB cells using lipofectamine, and an elevatedsecretion of GDNF protein compared with nontransfected HUCBcellswas confirmed in vitro, and also in the ischemic hemisphereat 1 and 4 weeks after GDNF gene modified CD34+ celltransplantation. These findings suggest the GDNF genemodifiedCD34+ cells could maintain high levels of GDNF for atleast 4 weeks after transplantation and that the elevatedGDNF secretion might have facilitated the enhancedneuroprotection.
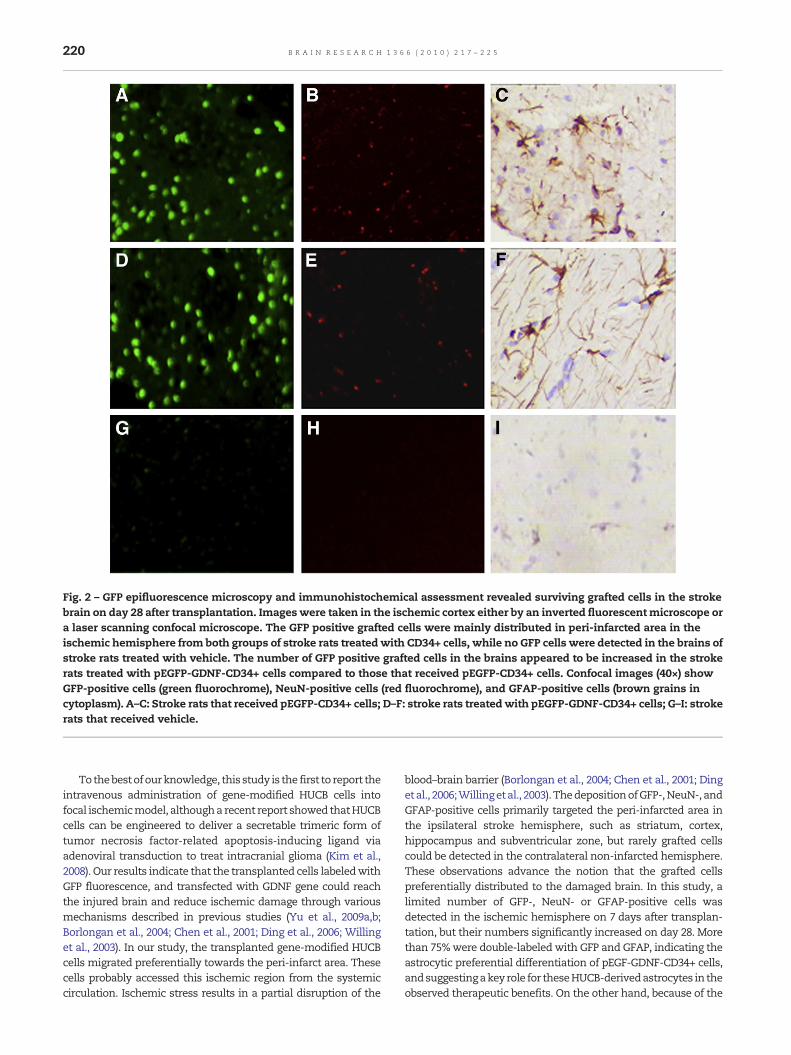
Fig. 2 – GFP epifluorescence microscopy and immunohistochemical assessment revealed surviving grafted cells in the strokebrain on day 28 after transplantation. Imageswere taken in the ischemic cortex either by an inverted fluorescentmicroscope ora laser scanning confocal microscope. The GFP positive grafted cells were mainly distributed in peri-infarcted area in theischemic hemisphere from both groups of stroke rats treatedwith CD34+ cells, while no GFP cells were detected in the brains ofstroke rats treated with vehicle. The number of GFP positive grafted cells in the brains appeared to be increased in the strokerats treated with pEGFP-GDNF-CD34+ cells compared to those that received pEGFP-CD34+ cells. Confocal images (40×) showGFP-positive cells (green fluorochrome), NeuN-positive cells (red fluorochrome), and GFAP-positive cells (brown grains incytoplasm). A–C: Stroke rats that received pEGFP-CD34+ cells; D–F: stroke rats treatedwith pEGFP-GDNF-CD34+ cells; G–I: strokerats that received vehicle.
220 B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
Tothebestofourknowledge, this study is the first to report theintravenous administration of gene-modified HUCB cells intofocal ischemicmodel, althougha recent report showed thatHUCBcells can be engineered to deliver a secretable trimeric form oftumor necrosis factor-related apoptosis-inducing ligand viaadenoviral transduction to treat intracranial glioma (Kim et al.,2008). Our results indicate that the transplanted cells labeledwithGFP fluorescence, and transfected with GDNF gene could reachthe injured brain and reduce ischemic damage through variousmechanisms described in previous studies (Yu et al., 2009a,b;Borlongan et al., 2004; Chen et al., 2001; Ding et al., 2006; Willinget al., 2003). In our study, the transplanted gene-modified HUCBcells migrated preferentially towards the peri-infarct area. Thesecells probably accessed this ischemic region from the systemiccirculation. Ischemic stress results in a partial disruption of the
blood–brain barrier (Borlongan et al., 2004; Chen et al., 2001; Dingetal., 2006;Willingetal., 2003). ThedepositionofGFP-,NeuN-, andGFAP-positive cells primarily targeted the peri-infarcted area inthe ipsilateral stroke hemisphere, such as striatum, cortex,hippocampus and subventricular zone, but rarely grafted cellscould be detected in the contralateral non-infarcted hemisphere.These observations advance the notion that the grafted cellspreferentially distributed to the damaged brain. In this study, alimited number of GFP-, NeuN- or GFAP-positive cells wasdetected in the ischemic hemisphere on 7 days after transplan-tation, but their numbers significantly increased on day 28. Morethan 75%were double-labeled with GFP and GFAP, indicating theastrocytic preferential differentiation of pEGF-GDNF-CD34+ cells,andsuggestingakeyrole for theseHUCB-derivedastrocytes in theobserved therapeutic benefits. On the other hand, because of the
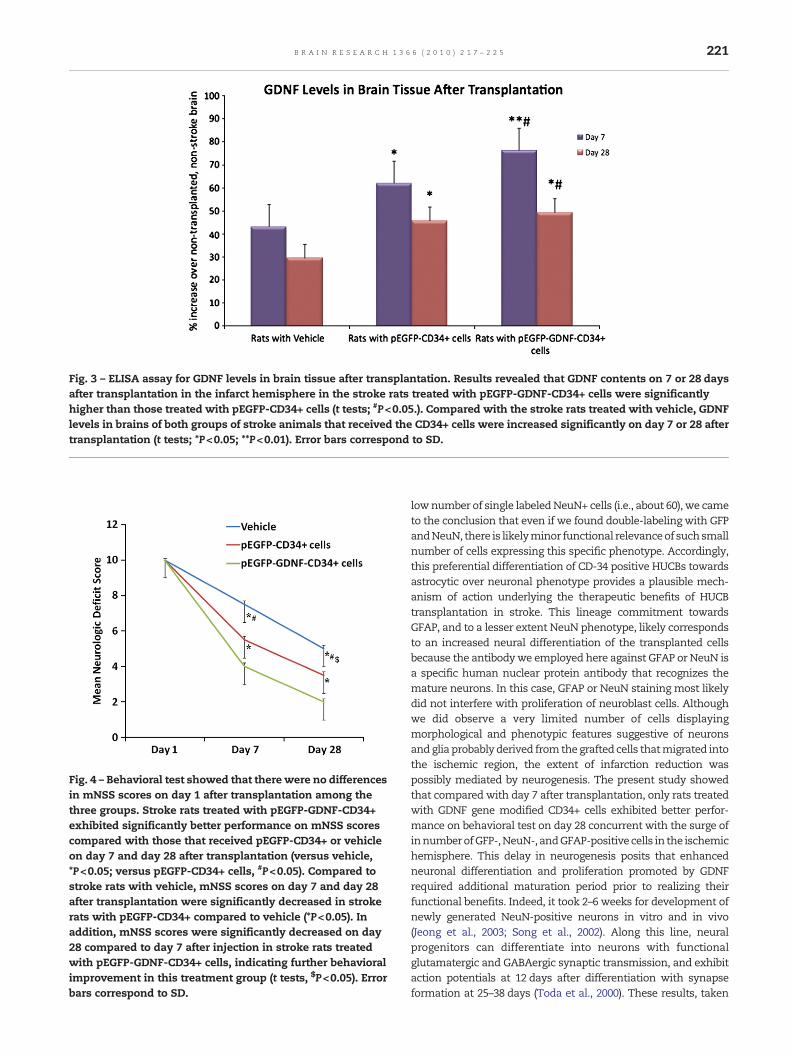
Fig. 3 – ELISA assay for GDNF levels in brain tissue after transplantation. Results revealed that GDNF contents on 7 or 28 daysafter transplantation in the infarct hemisphere in the stroke rats treated with pEGFP-GDNF-CD34+ cells were significantlyhigher than those treated with pEGFP-CD34+ cells (t tests; #P<0.05.). Compared with the stroke rats treated with vehicle, GDNFlevels in brains of both groups of stroke animals that received the CD34+ cells were increased significantly on day 7 or 28 aftertransplantation (t tests; *P<0.05; **P<0.01). Error bars correspond to SD.
Fig. 4 – Behavioral test showed that there were no differencesin mNSS scores on day 1 after transplantation among thethree groups. Stroke rats treated with pEGFP-GDNF-CD34+exhibited significantly better performance on mNSS scorescompared with those that received pEGFP-CD34+ or vehicleon day 7 and day 28 after transplantation (versus vehicle,*P<0.05; versus pEGFP-CD34+ cells, #P<0.05). Compared tostroke rats with vehicle, mNSS scores on day 7 and day 28after transplantation were significantly decreased in strokerats with pEGFP-CD34+ compared to vehicle (*P<0.05). Inaddition, mNSS scores were significantly decreased on day28 compared to day 7 after injection in stroke rats treatedwith pEGFP-GDNF-CD34+ cells, indicating further behavioralimprovement in this treatment group (t tests, $P<0.05). Errorbars correspond to SD.
221B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
lownumber of single labeledNeuN+ cells (i.e., about 60), we cameto the conclusion that even if we found double-labeling with GFPandNeuN, there is likelyminor functional relevanceof suchsmallnumber of cells expressing this specific phenotype. Accordingly,this preferential differentiation of CD-34 positive HUCBs towardsastrocytic over neuronal phenotype provides a plausible mech-anism of action underlying the therapeutic benefits of HUCBtransplantation in stroke. This lineage commitment towardsGFAP, and to a lesser extent NeuN phenotype, likely correspondsto an increased neural differentiation of the transplanted cellsbecause the antibodywe employed here against GFAP or NeuN isa specific human nuclear protein antibody that recognizes themature neurons. In this case, GFAP or NeuN staining most likelydid not interfere with proliferation of neuroblast cells. Althoughwe did observe a very limited number of cells displayingmorphological and phenotypic features suggestive of neuronsandglia probablyderived fromthegrafted cells thatmigrated intothe ischemic region, the extent of infarction reduction waspossibly mediated by neurogenesis. The present study showedthat compared with day 7 after transplantation, only rats treatedwith GDNF gene modified CD34+ cells exhibited better perfor-mance on behavioral test on day 28 concurrent with the surge ofinnumberofGFP-,NeuN-, andGFAP-positivecells in the ischemichemisphere. This delay in neurogenesis posits that enhancedneuronal differentiation and proliferation promoted by GDNFrequired additional maturation period prior to realizing theirfunctional benefits. Indeed, it took 2–6 weeks for development ofnewly generated NeuN-positive neurons in vitro and in vivo(Jeong et al., 2003; Song et al., 2002). Along this line, neuralprogenitors can differentiate into neurons with functionalglutamatergic and GABAergic synaptic transmission, and exhibitaction potentials at 12 days after differentiation with synapseformation at 25–38 days (Toda et al., 2000). These results, taken

222 B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
together with our data, suggest that cell replacement for the lostcells in ischemic brain might be a key cell repair mechanism forachieving stable, long-term functional recovery in stroke.
There have also beenmany reports on theuse of gene therapyfor neurological diseases (Chen et al., 2009; Horita et al., 2006;Kurozumi et al., 2004; Nomura et al., 2005). Recombinantadenoviruses have been the main vector for delivering genes toappropriate lesion sites (Horita et al., 2006; Nomura et al., 2005;Yasuhara et al., 2006). Amajor limitation in using viral vectors forgene delivery is the resulting unstable expression of specificproteins (Yasuhara et al., 2006). Conversely, stable transfection ofa foreign gene, i.e., rat GDNF gene to HUCB CD34+ cells, usinglipofectamine was found to maintain the specific trophicexpression as observed here and in our recent study (Yu et al.,2010). In this respect, GDNF gene-modified HUCB cells have a farreaching translational potential for improving neuroprotection incell transplantation.
Previous transplantation regimens in ischemic modelshave injected cells directly into the brain parenchyma,which might result in tissue damage along the needlepassages, as well as limitedmigration of grafted cells (Darsaliaet al., 2007; Jin et al., 2005; Lee et al., 2007; Willing et al., 2003).Although direct intracerebral injection of gene-modified stemcells in the rat revealed therapeutic efficacy, this approachmay be limited in clinical applications because of the surgicalrequirement and the difficulty in widespread distribution ofcells to ischemic areas. In this study, we transplanted gene-modified HUCB cells intravenously, and observed not onlytherapeutic efficacy, but an excellent site-directed depositionof grafted cells within the peri-infarct area. From a practicalstandpoint, systemic delivery of stem cells is less invasive,and can be more readily carried out in the clinic comparedwith intracerebral injection.
In the present study, SHRwasused as subjects for creating thestroke model. Previous studies (Coyle, 1984; Sokolova et al., 1985)have shown that the cerebral vascular anatomy in SHR closelyresembles that of thehuman, and the pathophysiological processof cerebral infarction in SHR also closely mimics that ofhypertensive patients with analogous cerebral vascular altera-tions in these animals. However, Sprague–Dawley or Wistar ratswith normal blood pressure have been used routinely as theanimal subject inmany past studies for evaluating the efficacy ofstem cell transplantation to treat cerebral infarction (Yu et al.,2009a,b). Unfortunately, Sprague–Dawley and Wistar rats lackhypertensive cerebral vascular pathological changes. Therefore,the use of SHR appears more clinically relevant than other ratstrains with normal blood pressure. Our recent study alsodemonstrated that the volume of cerebral infarction wassignificantly increased in SHR with MCAO compared with SDrats with MCAO (Yu et al., 2009a,b).
In conclusion, the present data provided initial evidencethat intravenously transplanted gene-modified HUCB cellsentered the ischemic rat brain, and survived, differentiated,and proliferated primarily within the peri-infarct area, andcoincided with a dramatic reduction of infarction volume anda remarkable improvement in neurological performance.These results further lent support to the utility of intravenoustransplantation of genetically engineered stem cells as aneffective and minimally invasive approach in affording CNSrepair for stroke therapy.
4. Experimental procedures
4.1. Main reagents
TM Total RNA Kit I and Trizol, a synthetic kit for singlestranded cDNA, were purchased from Shanghai bioengineer-ing companies, China. A DNA Purification kit was provided byJianCheng Biotechnology Company (Nanjing, China). Plasmidscontaining the enhanced green fluorescent protein-N1(pEGFP-N1) and bacteria E. coli DH5a were kindly provided byDr. Li Fang (Xiangya Hospital, Changsha, China). MMLVreverse transcriptase, T4DNA ligases, T4DNA polymerases,Taq DNA polymerases, nucleotides, endonuclease (XhoI, KpnI),DNA marker, Lipofectamine™ 2000 kit and ELISA kit for GDNFwere purchased from Promega (Madison, WI, USA). Theupstream and downstream primers, designed by Primer 5.0,were synthesized by Boshang Biotechnology Corporation(Shanghai, China). DMEM culture medium, fetal calf serumand G418 were obtained from GIBCO Company. The use ofanimals was approved by the Animal Care and Use Committeeof Central South University (Changsha, China). All procedureswere carried out in accordance with institutional guidelines.
4.2. Preparation of CD34+ cells
Human umbilical blood samples were collected from healthy,full-term neonates, immediately after delivery in the obstetricsection of Xiangya Hospital. The umbilical cord blood (UCB)sample [mean volume (64.6±13.6)ml] was collected into a250ml blood donor set containing 28ml citrate-phosphate-dextrose-adenosine anticoagulant. Nucleated cells were sepa-rated by sedimentation with 1.2% hydroxyethyl starch in 0.9%sodium chloride. Low-density mononuclear cells were thencollected after standard separation on Ficoll-Paque. For sortingCD34+ cell population, UCB cells were stained with anti-CD34+antibody and sorted by a flowmeter. The collected cellsat 2×106 cells/μl were resuspended in expansion mediumcontaining low-glucose Dulbecco's modified Eagle medium,supplemented with 20% fetal bovine serum (FBS). Interleukin-3(IL-3, 50 ng/ml), IL-6 (100 ng/ml) and SCF (100 ng/ml)were addedto maintain these cells in cycle. Quantitative analyses wereperformed using a flow cytometry (Becton Dickenson, USA).
4.3. Construction and identification of eukaryotic expressionvector pEGFP/GDNF recombinant
Total RNA isolation, GDNF cDNA preparation with the cDNAsynthesis Kit, and PCR analysis were carried out according tostandard protocols and conditions as described by themanufacturers. Total RNA was isolated from fresh fetal braintissue of SD rats. The rat cDNA encoding GDNF was obtainedby amplification of a rat cortex cDNA library using specificprimers coding region of the rat prepro-GDNF gene andreverse transcription coupled with subsequent polymerasechain reaction (RT-PCR) to synthesize the entire codingsequence of rat GDNF. The specific primers of GDNF weredesigned by using the nucleotide sequence of rat GDNF(GenBank) to isolate rat GDNF cDNA. The upstream primer

223B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
contains a XhoI recognition sequence to facilitate subsequentcloning, the oligonucleotide sequences of upstream primerwere 5′-GGAAGCTTATGAAGTTATGGGATGTCG-3′. The down-stream primer contains KpnI recognition sequence, includingthe translation termination codon (5-GAGGATCCTCAGATA-CATCCACACC-3′). With rat GDNF cDNA as a template, theopen reading frame of GDNF was amplified by RT-PCR.
4.4. Transfection of GDNF gene into CD34+ cells
Transfection was performed according to the Nucleofector™manual. Recombinant plasmid pEGFP/GDNF or plasmid pEGFPas control was transfected into the CD34+ cells usingLipofectamineTM 2000 kit. At 6, 12, 24 and 48 h aftertransfection, supernatants were collected. The mRNA expres-sion of recombinant GDNF in the supernatants of culturesfrom pEGFP/GDNF or pEGFP transfected CD34+ cells main-tained in supplemented serum-free medium for 24 h wasperformed by reverse transcription-polymerase chain reaction(RT-PCR). Cultures were washed once in PBS before solubili-zation in Trizol (Life Technologies, Gaithersburg, MD) and thenstored at −80 °C. RNA extraction was performed according tothe instructions of the manufacturer (Life Technologies). Asuperscript kit (Life Technologies) was used for reversetranscription of 10 mg of RNA per condition. PCR conditionswere optimized by varying MgCl2 concentration and cyclenumber to determine the linear amplification range. Ampli-fication products were identified by size and confirmed byDNA sequencing. MgCl2 concentration was 1.5 mM. Primersequences were as follows: GAPDH, [forward (f)] 5′-CTCGTCTCA-TAGACAAGATGGTGAAG-3′; GAPDH, [reverse (r)] 5′-AGACTC-CACGACATACTCAGCACC-3′; GDNF, (f) 5′-GCCCTACTTTGTCAC-3′; GDNF, (r) 5′-TCACCAGCCTTCTATTTC-3′. For assay of thecontent of GDNF released by transfected CD34+ cells in cellcultures, the supernatants collected from 1×105 cells/μl for 6, 12,24 and 48 h after transfection were analyzed by two-sitesandwich ELISA (Duo-Kit ELISA for the rat GDNF protein). GFPexpression was examined using fluorescent microscopy todetermine transfection efficiency. Nontransfected culturedCD34+ cells were used as control.
4.5. Animal model and intravenous transplantation
Eight weeks old, male spontaneous hypertensive rats weigh-ing 220–240 g (provided by Shanghai Slack Laboratory AnimalCo., Ltd.) were anesthetized using 10% chloral hydrate(300 mg/kg, i.p.). Transient unilateral focal ischemia withreperfusion was produced using the intraluminal threadocclusion method for 120 min as described by our previousstudies and those of several other laboratories (Borlonganet al., 2004; Chen et al., 2001; Willing et al., 2003).
Following MCAO, animals were randomly divided intothree experimental groups: Group A (Control, stroke+medium as vehicle infused intravenously, n=20), Group B(pEGFP-CD34+ cells intravenously delivered at 6 h after stroke;n=20), and Group C (pEGFP-GDNF-CD34+ cells intravenouslydelivered at 6 h after stroke; n=20). At 6 h after MCAO, pEGFP-GDNF-CD34+ cell suspension (approximately 1×107 cells) in1 ml total fluid volume was injected slowly for 5 min in the rattail vein for Group C, pEGFP-CD34+ cell suspension (approx-
imately 1×107 cells) in the same volume for Group B, and thesame volume of medium as vehicle for Group A.
4.6. Behavioral tests
The modified Neurological Severity Scores (mNSS) wereperformed to confirm the functional consequences of theMCA occlusion and to quantify improvements in neurologicalfunction on days 1, 7, 14, 21 and 28 after transplantation by aninvestigatorwhowas blinded to the experimental designs. ThemNSS were calculated based on a series of motor, sensory,reflex, and balance tests (Lourbopoulos et al., 2008).
4.7. Histology
Following behavioral tests on day 7 post-transplantation, fiveanimals in each group were randomly anesthetized with lethaldose of chloral hydrate (500mg/kg, i.p.), perfused intracardiallywith 150ml of ice-cold saline, decapitated and their brainsharvested. Using a rat brain matrix (RBI), a tissue block (2mmwide) was obtained. Quantitative histological determinations ofinfarct volume were performed using standard TTCstaining and quantitative image analysis as previously described(Lourbopoulos et al., 2008; Nomura et al., 2005). Infarct volume(mm3) was determined using the following formula=2mm(thickness of the section)×[sum of the infarction area (mm2)].
4.8. Immunohistochemical assessment and GFPepifluorescence microscopy
On day 7 and 28 post-transplantation, five animals in eachgroup were randomly sacrificed and the brains were harvestedwith themethod as described above. For immunohistochemicalassessment, brains from three of five animals on day 7 and 28post-transplantation were fixed by 4% paraformaldehyde inphosphate-buffer, dehydrated by 25% sucrose in 0.1 M PBS forovernight, and then cut into coronal cryosections of 10-μmthickness with cryomicrotome. After the sections were dried,theywere fixedwith acetone and treatedwith 0.2%TritonX-100for 10min followed by 2 mol/l HCl for 1 h. Then the sectionswere blocked with normal serum, incubated with the mouseanti-human nuclei monoclonal antibody Neuronal Nuclei(NeuN) (1:100)/mouse anti-humanMonoclonal Anti-Glial Fibril-lary Acidic Protein (GFAP), (1:100) (Perprotech, USA) at 4 °Covernight, subsequently treated with peroxidase conjugatedrabbit anti-mouse IgG (Beijing Zhongshan Biotechnology Co.,China). Diaminobenzidine was then used as a chromogen. Thesections were further counterstained with hematoxylin formicroscopic examination by an inverted fluorescence micro-scope (Carl Zeiss, Germany) and a laser scanner confocalmicroscope (Olympus, Japan).
The remaining brains were initially stored in 4% parafor-maldehyde with 25% sucrose for at least 48 h prior to cryostatsectioning. The entire tissue block was cut, and all 20-μmcryostat sections were examined to reveal grafted cellsurvival. Microscopy was conducted via GFP epifluorescence(fluorescein isothiocyanate fluorochrome) using a Zeiss Axio-phot 2 microscope (Carl Zeiss, Germany) at 20× and 40×magnifications. To determine the survival and differentiationof grafted cells semiquantitatively, the cells labeled with GFP,

224 B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
NeuN, and GFAP in the same focal plane were countedrespectively in six regions of interest situated within theperi-infarct striatum and cortex. The average of the positivecells in all slices for each field of vision was used for analysis.An investigator, blinded to the treatment conditions, per-formed the cell counts.
4.9. ELISA for GDNF in vivo
For measurement of GDNF protein content in brain, theremaining five animals in each group on day 28 aftertransplantation were anesthetized by 10% chloral hydrate(500 mg/kg, i.p.), and their brains were removed quickly. Thentheir ischemic hemispheres were dissected on ice and storedat −80 °C until use. Subsequently, each sample was homoge-nizedwith buffer (0.01 mol/l Tris–HCl buffer, pH 7.5, containing0.1 mol/l NaCl, 1 mmol/l ethylenediaminetetraacetic acid(EDTA), and 1 mmol/l phenylmethylsulfonyl fluoride) using aPolytron. The homogenate was centrifuged at 10,000×g for20 min at 4 °C, and then the supernatant was collected as asample for GDNF ELISA assay. A specific GDNF sandwichimmunoassay with a goat polyclonal antibody for humanGDNF was performed in the 96-well immunoassay plate(Peprotech USA), which was precoated with the anti-GDNFmonoclonal antibody, using the kit (GDNF Emax ImmunoassaySystems, Promega). After overnight incubation at 4 °C, reactionof a second antibody-HRP conjugate (kit component) for goatIgY was performed, followed by color development with TMBsolution (kit component). After the stop reaction with 1 mol/l phosphoric acid, OD450 was measured by a 96 well platereader, and GDNF protein content of each sample wascalculated. Using this system, immunoreactive GDNF in thesamples can be quantified in the range of 16 to 1000 pg/ml.
4.10. Statistical analysis
The data were presented as mean±standard deviation (SD).Behavioral scores, the number of cells, and infarct volumes wereanalyzed using ANOVA test followed by post hoc Bonferroni ttests when assays involved multiple time points, whereas onlypair-wise t testswere performed for single time point assays. Thestatistical software used was SPSS for Windows, version 13.0,from SPSS Inc. The level of significance was set at <0.05.
Acknowledgments
This work was supported by the grant (30572079) from theNational Natural Science Foundation of China (2006), fundingof the Chinese Ministry of Education for oversea students(2008), and James and Esther King Biomedical ResearchPrograms 09KB-01-23123 and 1KG01-33966.
R E F E R E N C E S
Boltze, J., Kowalski, I., Geiger, K., Reich, D., Gunther, A., Buhrle, C.,Egger, D., Kamprad, M., Emmrich, F., 2005. Experimentaltreatment of stroke in spontaneously hypertensive rats byCD34+ and CD34− cord blood cells. Ger. Med. Sci. 3 (9), 1–15.
Borlongan, C.V., Hadman, M., Sanberg, C.D., Sanberg, P.R., 2004.Central nervous system entry of peripherally injectedumbilical cord blood cells is not required for neuroprotection instroke. Stroke 35 (10), 2385–2389.
Chen, J., Li, Y., Chopp, M., 2000. Intracerebral transplantation ofbonemarrowwith BDNF after MCAo in rat. Neuropharmacology39 (5), 711–716.
Chen, J., Sanberg, P.R., Li, Y., Wang, L., Lu, M., Willing, A.E.,Sanchez-Ramos, J., Chopp, M., 2001. Intravenousadministration of human umbilical cord blood reducesbehavioral deficits after stroke in rats. Stroke 32 (11),2682–2688.
Chen, B., Gao, X.Q., Yang, C.X., Tan, S.K., Sun, Z.L., Yan, N.H., Pang, Y.G., Yuan, M., Chen, G.J., Xu, G.T., Zhang, K., Yuan, Q.L., 2009.Neuroprotective effect of grafting GDNF gene-modified neuralstem cells on cerebral ischemia in rats. Brain Res. 11 (1284), 1–11.
Coyle, P., 1984. Outcomes to middle cerebral artery occlusion inhypertensive and normotensive rats. Hypertension 6 (2 Pt 2),69–74.
Darsalia, V., Kallur, T., Kokaia, Z., 2007. Survival migration andneuronal differentiation of human fetal striatal and corticalneural stem cells grafted in stroke-damaged rat striatum. Eur. J.Neurosci. 26 (3), 605–614.
Ding, D.C., Shyu,W.C., Lin, S.Z., Li, H., 2006. Current concepts in adultstem cell therapy for stroke. Curr. Med. Chem. 13 (29), 3565–3574.
Horita, Y., Honmou, O., Harada, K., Houkin, K., Hamada, H., Kocsis,J.D., 2006. Intravenous administration of glial cell line-derivedneurotrophic factor gene-modified humanmesenchymal stemcells protects against injury in a cerebral ischemiamodel in theadult rat. J. Neurosci. Res. 84 (7), 1495–1504.
Jeong, S.W., Chu, K., Jung, K.H., Kim, S.U., Kim, M., Roh, J.K., 2003.Human neural stem cell transplantation promotes functionalrecovery in rats with experimental intracerebral hemorrhage.Stroke 34 (9), 2258–2263.
Jin, K., Sun, Y., Xie, L., Mao, X.O., Childs, J., Peel, A., Logvinova, A.,Banwait, S., Greenberg, D.A., 2005. Comparison ofischemia-directed migration of neural precursor cells afterintrastriatal, intraventricular, or intravenous transplantationin the rat. Neurobiol. Dis. 18 (2), 366–374.
Kim, S.M., Lim, J.Y., Park, S.I., Jeong, C.H., Oh, J.H., Jeong, M., Oh,W.,Park, S.H., Sung, Y.C., Jeun, S.S., 2008. Gene therapy usingTRAIL-secreting human umbilical cord blood-derivedmesenchymal stem cells against intracranial glioma. CancerRes. 68 (23), 9614–9623.
Kobayashi, T., Ahlenius, H., Thored, P., Kobayashi, R., Kokaia, Z.,Lindvall, O., 2006. Intracerebral infusion of glial cellline-derived neurotrophic factor promotes striatalneurogenesis after stroke in adult rats. Stroke 37 (9), 2361–2367.
Kurozumi, K., Nakamura, K., Tamiya, T., Kawano, Y., Kobune, M.,Hirai, S., Uchida, H., Sasaki, K., Ito, Y., Kato, K., Honmou, O.,Houkin, K., Date, I., Hamada, H., 2004. BDNF gene-modifiedmesenchymal stem cells promote functional recovery andreduce infarct size in the rat middle cerebral artery occlusionmodel. Mol. Ther. 9 (2), 189–197.
Lee, H.J., Kim, K.S., Kim, E.J., Choi, H.B., Lee, K.H., Park, I.H., Ko, Y.,Jeong, S.W., Kim, S.U., 2007. Brain transplantation ofimmortalized human neural stem cells promotes functionalrecovery in mouse intracerebral hemorrhage stroke model.Stem Cells 25 (5), 1204–1212.
Lobel, D.A., Hadman, P.R., Sanberg, P.R., Borlongan, C.V., 2003.Early intracerebral and intra-arterial transplantation of humanumbilical cord blood cells into an ischemic rat brain model.Exp. Neurol. 181 (1), 97.
Lourbopoulos, A., Karacostas, D., Artemis, N., Milonas, I., Grigoriadis,N., 2008. Effectiveness of a new modified intraluminal suture fortemporary middle cerebral artery occlusion in rats of variousweight. J. Neurosci. Methods 30 (173(2)), 225–234.
Makinen, S., Kekarainen, T., Nystedt, J., Liimatainen, T., Huhtala,T., Narvanen, A., Laine, J., Jolkkonen, J., 2006. Human umbilical

225B R A I N R E S E A R C H 1 3 6 6 ( 2 0 1 0 ) 2 1 7 – 2 2 5
cord blood cells do not improve sensorimotor or cognitiveoutcome following transient middle cerebral artery occlusionin rats. Brain Res. 1123 (1), 207–215.
Nomura, T., Honmou, O., Harada, K., Houkin, K., Hamada, H.,Kocsis, J.D., 2005. I.V. infusion of brain-derived neurotrophicfactor gene-modified humanmesenchymal stem cells protectsagainst injury in a cerebral ischemia model in adult rat.Neuroscience 136 (1), 161–169.
Nystedt, J., Makinen, S., Laine, J., Jolkkonen, J., 2006. Human cordblood CD34+ cells and behavioral recovery following focalcerebral ischemia in rats. Acta Neurobiol. Exp. (Wars) 66 (4),293–300.
Sokolova, I.A., Manukhin, E.B., Blinkov, S.M., Koshelev, V.B.,Pinelis, V.G., Rodionov, I.M., 1985. Rarefication of the arteriolesand capillary network in the brain of rats with different formsof hypertension. Microvasc. Res. 30 (1), 1–9.
Song, H.J., Stevens, C.F., Gage, F.H., 2002. Neural stem cells fromadult hippocampus develop essential properties of functionalCNS neurons. Nat. Neurosci. 5 (5), 438–445.
Taguchi, A., Soma, T., Tanaka, H., Kanda, T., Nishimura, H.,Yoshikawa, H., Tsukamoto, Y., Iso, H., Fujimori, Y., Stern, D.M.,Naritomi, H., Matsuyama, T., 2004. Administrationof CD34+ cells after stroke enhancesneurogenesis via angiogenesis in a mouse model. J. Clin. Invest.114 (3), 330–338.
Toda, H., Takahashi, J., Mizoguchi, A., Koyano, K., Hashimoto, N.,2000. Neurons generated from adult rat hippocampal stem cellsform functional glutamatergic and GABAergic synapses in vitro.Exp. Neurol. 165 (1), 66–76.
Vendrame, M., Cassady, J., Newcomb, J., Butler, T., Pennypacker, K.R., Zigova, T., Sanberg, C.D., Sanberg, P.R., Willing, A.E., 2004.Infusion of human umbilical cord blood cells in a rat model ofstroke dose-dependently rescues behavioral deficits andreduces infarct volume. Stroke 35 (10), 2390–2395.
Willing, A.E., Lixian, J., Milliken, M., Poulos, S., Zigova, T., Song, S.,Hart, C., Sanchez-Ramos, J., Sanberg, P.R., 2003. Intravenousversus intrastriatal cord blood administration in a rodentmodel of stroke. J. Neurosci. Res. 73 (3), 296–307.
Xiao, J., Nan, Z., Motooka, Y., Low, W.C., 2005. Transplantation of anovel cell line population of umbilical cord blood stem cellsameliorates neurological deficits associated with ischemicbrain injury. Stem Cells Dev. 14 (6), 722–733.
Yasuhara, T., Borlongan, C.V., Date, I., 2006. Ex vivo gene therapy:transplantation of neurotrophic factor-secreting cells forcerebral ischemia. Front. Biosci. 11 (1), 760–775.
Yu, G., Borlongan, C.V., Stahl, C.E., Hess, D.C., Ou, Y., Kaneko, Y.,Yu, S.J., Yang, T., Fang, L., Xie, X., 2009a. Systemic delivery ofumbilical cord blood cells for stroke therapy: a review. Restor.Neurol. Neurosci. 27 (1), 41–54.
Yu, G., Hu, K., O.Y.L., Yang, T.L., 2009b. The preparation forexperimental animal model of focal cerebral ischemicreperfusion injury in spontaneous hypertensive rats. Chin. J.Hypertens. 17 (7), 64–66.
Yu, G., Borlongan, C.V., Ou, Y., Stahl, C.E., Yu, S., Bae, E., Kaneko, Y.,Yang, T., Yuan, C., Fang, L., 2010. In vitro non-virallipofectamine delivery of the gene for glial cell line-derivedneurotrophic factor to human umbilical cord blood CD34+ cells.Brain Res. 1325, 147–154.





























