Embed Size (px)
Citation preview

ava i l ab l e a t www.sc i enced i r ec t . com
C l i n i ca l Immuno logy
www.e l sev i e r . com / l oca te / yc l im
Clinical Immunology (2012) 144, 98–108
Intranasal vaccination with an adjuvanted Norwalkvirus-like particle vaccine elicits antigen-specific Bmemory responses in human adult volunteersKarina Ramirez a, b, 1, Rezwanul Wahid a, b, 1, Charles Richardson d,Robert F. Bargatze d, Samer S. El-Kamary a, b, c,Marcelo B. Sztein a, b,⁎, Marcela F. Pasetti a, b,⁎
a Center for Vaccine Development, University of Maryland School of Medicine, 685 West Baltimore St. Room 480, Baltimore,MD 21201, USAb Department of Pediatrics, University ofMaryland School ofMedicine, 685West Baltimore St. Room480, Baltimore,MD 21201, USAc Department of Epidemiology and Public Health, University of Maryland School of Medicine,685 West Baltimore St. Room 480, Baltimore, MD 21201, USAd LigoCyte Pharmaceuticals, Inc. 2155 Analysis Drive, Bozeman, MT 59718, USA
Received 7 February 2012; accepted with revision 14 May 2012
⁎ Corresponding authors at: Center foBaltimore, MD 21201, USA. Fax: +1 41
E-mail addresses: msztein@medicin1 These authors contributed equally
1521-6616/$ - see front matter © 2012doi:10.1016/j.clim.2012.05.006
KEYWORDSNorwalk vaccines;Norwalk virus;Norwalk immuneresponses;Intranasal vaccinationand humans;B memory cells andvaccines
Abstract Noroviruses are the most frequent cause of acute gastroenteritis in humans of allages. No vaccines are currently available. An intranasally delivered Norwalk (NV) virus-likeparticle (VLP) vaccine was recently shown to be well tolerated, immunogenic and to protectagainst infection in Phase 1 studies. Here, we examined B memory (BM) responses in volunteerswho received the highest dosage levels of the NV-VLP vaccine (50 μg and 100 μg). We measuredthe frequency of NV-specific IgG and IgA-secreting BM cells in peripheral blood and the level ofantibodies produced by these cells in culture. All subjects immunized with 100 μg of the NV-VLPvaccine and 90% of those who received 50 μg had significant IgA or IgG BM responses. The BM cellfrequencies correlated with serum antibody levels and mucosally-primed antibody-secreting cellresponses. This is the first demonstration of dose-dependent, functional B responses in humans
Mimmunized intranasally with a NV-VLP vaccine.© 2012 Elsevier Inc. All rights reserved.
r Vaccine Development, Unive0 706 6205.e.umaryland.edu (M.B. Szteinto this work.
Elsevier Inc. All rights reserve
rsity of Maryland School of Medicine, 685 West Baltimore St. Room 480,
), [email protected] (M.F. Pasetti).
d.

99Intranasal vaccination with an adjuvanted Norwalk virus-like particle vaccine
1. Introduction
Noroviruses (NoV) are responsible for more than 95% ofepidemic viral gastroenteritis in humans. They persist in theenvironment and spread easily through contaminated foodand water; a very low infectious dose (10–1000 virions) issufficient to cause disease [1]. In the U.S. alone, NoV areestimated to cause more than 20 million cases yearly,representing 60% of all acute gastroenteritis and 50% offoodborne outbreaks [2,3]. Norwalk virus (NV), the prototypeof human NoV, is known for causing outbreaks in a widebreadth of community settings such as restaurants, hospices,hotels, military barracks or cruise ships [3–5]. The disease isusually self-limited, but can be severe and complicated invulnerable populations, such as young children, the elderlyand patients who are chronically ill or immunocompromised[5]. There is currently no vaccine available to prevent NoVinfection; treatment is generally limited to rehydration.Hence, appropriate infection control strategies and effectiveprevention tools are needed and their development andimplementation are considered a public health priority [3,6].
Although the virus cannot be cultured in vitro, NoVrecombinant virus-like particles (VLP) that are antigenicallyand morphologically similar to native viruses but lack thegenetic material that enables viral replication can beproduced using recombinant systems [7]. VLP could bedesigned to match individual viral strains and producedefficiently using simple technology. A handful of VLP-basedvaccines are commercially available for use in humans: thebest known probably being GlaxoSmithKline's Engerix®(hepatitis B virus) and Cervarix® (human papillomavirus),and Merck and Co., Inc.'s Recombivax HB® (hepatitis B virus)and Gardasil® (human papillomavirus) [8]. A VLP-based(seasonal and pandemic) influenza vaccine candidate wasrecently evaluated in a Phase 2 study and found to be welltolerated and to induce robust immune responses, meetinginternational and FDA licensure criteria [9]. These successessupport the development of a NoV VLP-vaccine [7].
We have recently completed a series of double-blind,placebo-controlled, dose-escalating Phase 1 studies in whichwe evaluated the safety and immunogenicity of an NV-VLPvaccine derived from NoV genogroup I, genotype 1 (GI.1).This vaccine was adjuvanted with monophosphoryl lipid A(MPL) (GlaxoSmithKline) and the mucoadhesin chitosan(ChiSys®) and was administered intranasally to healthyadult volunteers. The NV-VLP vaccine was found to be safeand highly immunogenic. NV-specific IgG and IgA antibodiesand antibody-secreting cells (ASC) expressing homing re-ceptors for gastrointestinal and peripheral lymphoid tissuewere elicited in a dose-dependent fashion [10].
During these clinical trials, an ancillary study wasperformed to further examine the breadth and quality ofthe humoral responses induced by the nasally administeredNV-VLP vaccine. Specifically, we investigated the inductionof NV-specific B memory (BM) cells to substantiate possiblelong-term protection under the premise that antibodies,particularly local IgA, represent an important host-defensemechanism against enteric viral infections. Upon re-exposureto microbial antigens vaccinated individuals mount a rapidanamnestic antibody response, which results from the
activation of BM cells primed during immunization. These BMcells, which reside mainly in niche compartments, willdifferentiate rapidly into plasma cells and heighten antibodyproduction. Increased levels of high-affinity antibodies arenecessary to block host cell binding of viral particles and toclear the antigenic overload typically associated with viralinfections, including NV [11]. The BM cells induced afterimmunization also replenish the pool of long-lived plasmacells, thus maintaining antibody levels long after immunizationand in the absence of a challenge antigen [12]. An ideal vaccineis expected to be immunogenic, but also capable of mounting arobust anamnestic response upon subsequent pathogen expo-sure, for which the induction and persistence of BM cells isessential.
Here we examined the frequencies of IgA and IgG BM cellscirculating in peripheral blood from volunteers who receivedthe NV VLP vaccine at the highest dosage levels (50 and100 μg) and from unvaccinated controls before and on days56 and 180 after immunization. We also investigated thefunctional capacity of these cells in terms of antibodyproduction. To assess whether there was an interdepen-dency of B-cell mediated immunity, the frequencies of NVIgA and IgG BM cells were compared with other immunolog-ical outcomes, i.e., mucosally primed NV-specific ASC andserum antibody titers. The most sensitive immunologicalreadouts and peak time points were defined and applied tothe overall evaluation of vaccine responders.
2. Materials and methods
2.1. Vaccine
The vaccine used in our studies consisted of NV-VLP fromNoV GI.1 genotype (Protein Sciences Corporation, Meriden,CT) adjuvanted with MPL® (GlaxoSmithKline) and chitosan(ChiSys®, Archimedes Development Ltd.), and sucrose andmannitol as excipients [10]. The vaccine was formulatedunder good manufacturing practices as a dry powder byArchimedes Development Ltd., Nottingham, UK, and it wasadministered intranasally using Bespak UniDose DP™ deliverydevices (Milton Keynes, UK). Each vaccine dose consisted oftwo loaded Bespak devices discharging 10 mg of dry powdervaccine formulation into each nostril for a total dose of20 mg.
2.2. Study design
Two Phase 1 clinical studies were performed as previouslydescribed [10]. Study 1 was a randomized, double-blind,placebo (adjuvant) controlled, dose-escalation evaluation ofthe adjuvanted NV-VLP vaccine conducted at the Universityof Maryland as single-site. A total of 28 healthy adults(18–49 years old, H type 1 secretors) were randomized toreceive two intranasal doses of: (1) 5 μg NV-VLP vaccine(n=5) or adjuvant control (n=2); (2) 15 μg NV-VLP vaccine(n=5) or adjuvant control (n=2); or (3) 50 μg NV-VLP vaccine(n=10) or adjuvant control (n=4). The two vaccine doseswere separated by 21 days. Study 2 was a dose-comparisonstudy of the two highest dosages previously tested (50 μg and100 μg) and was conducted as a multicenter, randomized,

100 K. Ramirez et al.
double-blind study conducted at the University of Maryland,Saint Louis University, the University of Cincinnati, and theUniversity of Rochester. The subjects were similar to those inStudy 1, except that those with B or AB blood-type wereadditionally excluded. A total of 61 healthy adults wereenrolled and randomized 2:2:1:1, respectively, to receiveeither two doses of: 50 μg NV-VLP vaccine (n=20); 100 μg NV-VLP vaccine (n=20); adjuvant control (n=10); or true placebo(n=11) consisting of a puff of air (no dry powder). Similarly,the two vaccine doses were separated by 21 days. All subjectsin both studies provided informed signed consent, and therespective Institutional Review boards approved the studies[10].
2.3. Study subjects and samples
The specimens used in the BM analysis were obtained fromvolunteers participating in the NV-VLP vaccine studiesmentioned above and included subjects from Study 1, whoreceived 50 μg of the NV-VLP vaccine (n=9) and adjuvantcontrols (n=3), and subjects from Study 2 (enrolled at theUniversity of Maryland), who received 50 μg (n=5) or 100 μg(n=5) of the NV-VLP vaccine, and controls who received theadjuvant only (n=3) or a puff of air (n=2). Altogether, thestudy population comprised 19 vaccine recipients (n=14 inthe 50 μg group and n=5 in the 100 μg group) and8 unvaccinated controls. These subjects were selectedbased on cell availability as the sole inclusion criteria.
2.4. BM ELISpot
NV-VLP-specific IgG and IgA BM cells were measured using amethod described by Crotty et al. with modifications[13,14]. Briefly, frozen Peripheral Blood Mononuclear Cells(PBMC) were thawed, rested overnight and expanded with Bcell expansion media containing 5 μM 2β-mercaptoethanol(Sigma-Aldrich, St. Louis, MO), 1:100,000 pokeweed mito-gen, 6 μg/mL CpG-2006 (Biopolymer/Genomics Core Facili-ty, University of Maryland, Baltimore, MD), and 1:10,000Staphylococcus aureus Cowan (Sigma‐Aldrich), in RPMI-1640(Invitrogen, Carlsbad, CA) supplemented with 100 U/mlpenicillin, 100 μg/ml streptomycin, 50 μg/ml gentamicin,2 mM L-glutamine, 2.5 mM sodium pyruvate, 10 mM HEPES(all from Invitrogen) and 10% heat-inactivated fetal bovineserum (Hyclone, Logan, UT) (complete RPMI). Cells wereexpanded for 6 days (1×106 cells/ml in 6-well plates) andfed with complete RPMI on day 3 or 4. The BM ELISpot wasperformed as follows: nitrocellulose plates (MAHAN, Milli-pore, Billerica, MA) were coated with NV-VLP (5 μg/ml),goat anti-human IgG (Jackson ImmunoResearch Laborato-ries, Inc., West Grove, PA; 5 μg/ml) or goat anti-human IgA(Jackson ImmunoResearch; 5 μg/ml) diluted in PBS. Plateswere incubated overnight at 4 °C, washed with PBS andblocked with 10% FBS in RPMI-1640 for 2 h at 37 °C.Expanded cells were seeded at a single concentration(1.5×105 cells/well) in triplicate wells for NV-VLP-specificand in 3 serial dilutions (1×104–2.5×103 cells/well) induplicate wells for total IgG and IgA BM ELISpot determina-tions. Plates were then incubated for 6 h at 37 °C, 5% CO2.After incubation, they were washed with PBS-T followed byPBS and biotinylated mouse anti-human Pan-IgG antibody
(Hybridoma Reagent Laboratory, Baltimore, MD) or anti-human Pan-IgA (Jackson ImmunoResearch) was added for18 h at 4 °C. Detection reagents included HRP-Avidin D(Vector Laboratories, Burlingame, CA) added for 1 h at roomtemperature, followed by 3-Amino-9 etheylcarbazole C (AECsubstrate; Calbiochem, La Jolla, CA) for 8–10 min in thedark. The reaction was stopped by washing wells withdistilled water. Plates were air-dried and spots counted withan ELISpot reader (Immunospot®, Cellular Technology Ltd.,Cleveland, OH) and analyzed with Immunospot® version 5.0software. The total or VLP specific spot-forming cells (SFC)/well were converted into SFC/106 based on the number ofcells seeded. VLP-specific BM frequency (also referred to asBM responses) was calculated as the percentage of total IgGor IgA SFC. The data were normalized to the number of totalIgG or IgA ASC in culture to account for cell yield variationsduring PBMC expansion in the different experiments. Forquality control purposes, a threshold level of detection wasdefined as the 5th percentile of the number of total IgG orIgA SFC measured in all volunteers at any point in time. Avaccine responder was defined as any subject who exhibitedan increase in the frequency of IgA or IgG BM cells of at least0.05% above the pre-vaccination level. The cut off level of0.05% was determined in previous studies and corresponds totwice the average response of multiple negative controls[14].
2.5. Flow cytometry
Peripheral blood mononuclear cells before and after BMexpansion were surface stained with Vivid dye to identifyliving (Vivid negative) cells (Invitrogen) and with fluorescentconjugated monoclonal antibodies specific for CD38 (BDPharmingen, San Diego, CA) a marker of human antibodysecreting and plasma cells, CD19 (Coulter) a marker of B-cells, CD3 (BD Biosciences, Franklin Lakes, NJ) a marker of Tcells, CD14 (Invitrogen) used to exclude macrophages fromanalysis and CD27 (Coulter) a marker of human memorycells, as previously described [15]. Samples were run in a MoFloflow cytometer/cell sorter system (Coulter-Cytomation, FortCollins, CO) and analyzed using WinList 6.0 3D software (VeritySoftware House, Topsham, ME).
2.6. Antibody measurements
NV-VLP-specific serum antibodies and antibodies in lympho-cyte supernatant (ALS) of BM cultures were measured byELISA as described earlier [10]. ALS culture supernatantswere collected from PBMC that had been expanded for6 days (as described above) and stored at −70 °C untilevaluation. ELISA end-point titers were calculated throughlinear regression parameters as the inverse of the serumdilution that produced an Absorbance value of 0.2 above theblank and expressed as EU/ml. NV hemagglutination inhibi-tion (HAI) titers were measured as described previously [10].Titers were calculated as the inverse of the highest serumdilution that inhibited agglutination of NV-VLP-coated O+red blood cells.
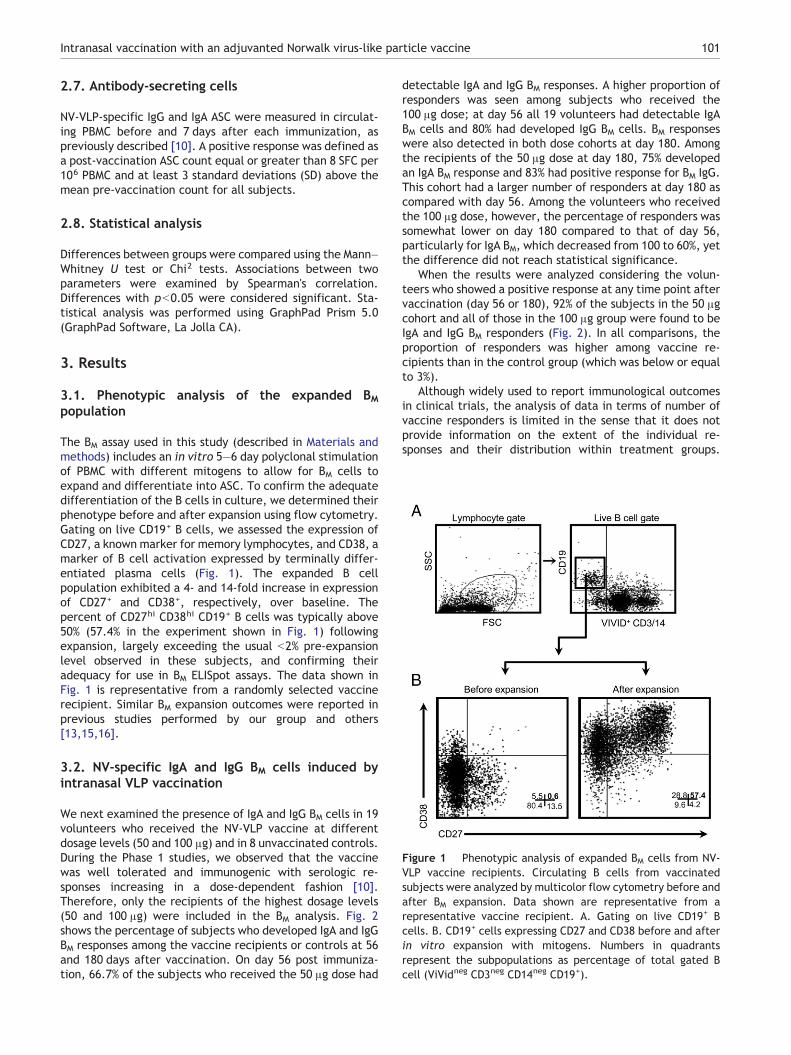
Figure 1 Phenotypic analysis of expanded BM cells from NV-VLP vaccine recipients. Circulating B cells from vaccinatedsubjects were analyzed by multicolor flow cytometry before andafter BM expansion. Data shown are representative from arepresentative vaccine recipient. A. Gating on live CD19+ Bcells. B. CD19+ cells expressing CD27 and CD38 before and afterin vitro expansion with mitogens. Numbers in quadrantsrepresent the subpopulations as percentage of total gated Bcell (ViVidneg CD3neg CD14neg CD19+).
101Intranasal vaccination with an adjuvanted Norwalk virus-like particle vaccine
2.7. Antibody-secreting cells
NV-VLP-specific IgG and IgA ASC were measured in circulat-ing PBMC before and 7 days after each immunization, aspreviously described [10]. A positive response was defined asa post-vaccination ASC count equal or greater than 8 SFC per106 PBMC and at least 3 standard deviations (SD) above themean pre-vaccination count for all subjects.
2.8. Statistical analysis
Differences between groups were compared using the Mann–Whitney U test or Chi2 tests. Associations between twoparameters were examined by Spearman's correlation.Differences with pb0.05 were considered significant. Sta-tistical analysis was performed using GraphPad Prism 5.0(GraphPad Software, La Jolla CA).
3. Results
3.1. Phenotypic analysis of the expanded BMpopulation
The BM assay used in this study (described in Materials andmethods) includes an in vitro 5–6 day polyclonal stimulationof PBMC with different mitogens to allow for BM cells toexpand and differentiate into ASC. To confirm the adequatedifferentiation of the B cells in culture, we determined theirphenotype before and after expansion using flow cytometry.Gating on live CD19+ B cells, we assessed the expression ofCD27, a known marker for memory lymphocytes, and CD38, amarker of B cell activation expressed by terminally differ-entiated plasma cells (Fig. 1). The expanded B cellpopulation exhibited a 4- and 14-fold increase in expressionof CD27+ and CD38+, respectively, over baseline. Thepercent of CD27hi CD38hi CD19+ B cells was typically above50% (57.4% in the experiment shown in Fig. 1) followingexpansion, largely exceeding the usual b2% pre-expansionlevel observed in these subjects, and confirming theiradequacy for use in BM ELISpot assays. The data shown inFig. 1 is representative from a randomly selected vaccinerecipient. Similar BM expansion outcomes were reported inprevious studies performed by our group and others[13,15,16].
3.2. NV-specific IgA and IgG BM cells induced byintranasal VLP vaccination
We next examined the presence of IgA and IgG BM cells in 19volunteers who received the NV-VLP vaccine at differentdosage levels (50 and 100 μg) and in 8 unvaccinated controls.During the Phase 1 studies, we observed that the vaccinewas well tolerated and immunogenic with serologic re-sponses increasing in a dose-dependent fashion [10].Therefore, only the recipients of the highest dosage levels(50 and 100 μg) were included in the BM analysis. Fig. 2shows the percentage of subjects who developed IgA and IgGBM responses among the vaccine recipients or controls at 56and 180 days after vaccination. On day 56 post immuniza-tion, 66.7% of the subjects who received the 50 μg dose had
detectable IgA and IgG BM responses. A higher proportion ofresponders was seen among subjects who received the100 μg dose; at day 56 all 19 volunteers had detectable IgABM cells and 80% had developed IgG BM cells. BM responseswere also detected in both dose cohorts at day 180. Amongthe recipients of the 50 μg dose at day 180, 75% developedan IgA BM response and 83% had positive response for BM IgG.This cohort had a larger number of responders at day 180 ascompared with day 56. Among the volunteers who receivedthe 100 μg dose, however, the percentage of responders wassomewhat lower on day 180 compared to that of day 56,particularly for IgA BM, which decreased from 100 to 60%, yetthe difference did not reach statistical significance.
When the results were analyzed considering the volun-teers who showed a positive response at any time point aftervaccination (day 56 or 180), 92% of the subjects in the 50 μgcohort and all of those in the 100 μg group were found to beIgA and IgG BM responders (Fig. 2). In all comparisons, theproportion of responders was higher among vaccine re-cipients than in the control group (which was below or equalto 3%).
Although widely used to report immunological outcomesin clinical trials, the analysis of data in terms of number ofvaccine responders is limited in the sense that it does notprovide information on the extent of the individual re-sponses and their distribution within treatment groups.
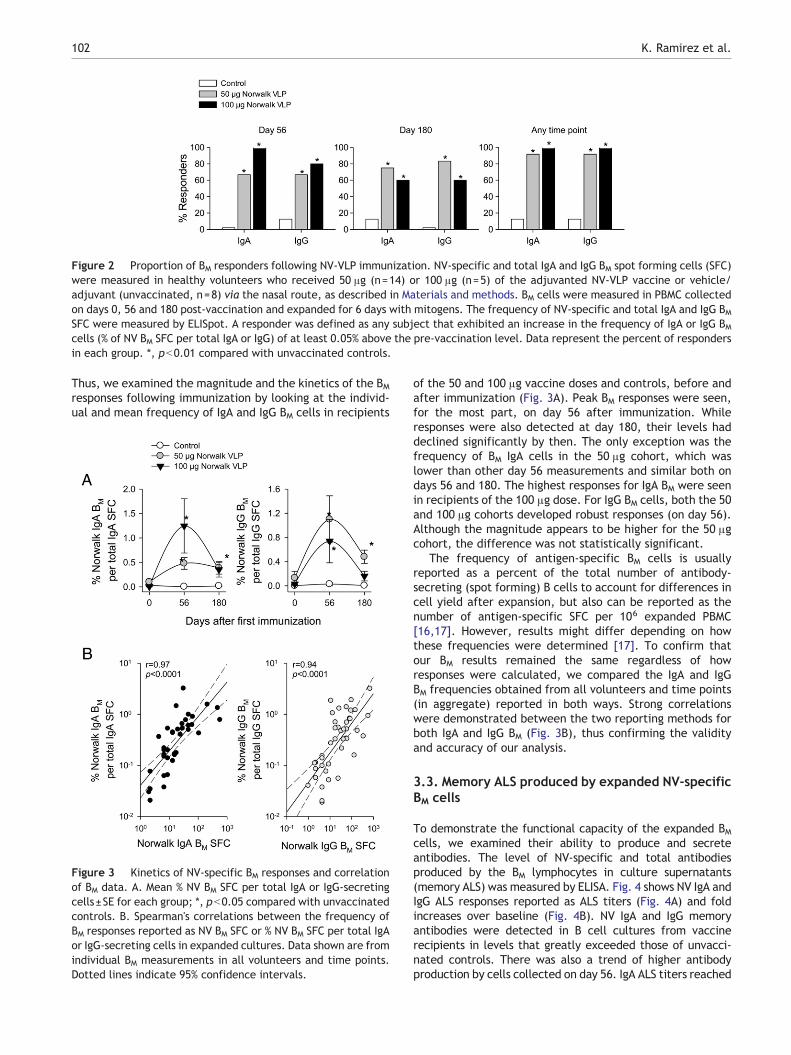
Figure 2 Proportion of BM responders following NV-VLP immunization. NV-specific and total IgA and IgG BM spot forming cells (SFC)were measured in healthy volunteers who received 50 μg (n=14) or 100 μg (n=5) of the adjuvanted NV-VLP vaccine or vehicle/adjuvant (unvaccinated, n=8) via the nasal route, as described in Materials and methods. BM cells were measured in PBMC collectedon days 0, 56 and 180 post-vaccination and expanded for 6 days with mitogens. The frequency of NV-specific and total IgA and IgG BMSFC were measured by ELISpot. A responder was defined as any subject that exhibited an increase in the frequency of IgA or IgG BMcells (% of NV BM SFC per total IgA or IgG) of at least 0.05% above the pre-vaccination level. Data represent the percent of respondersin each group. *, pb0.01 compared with unvaccinated controls.
102 K. Ramirez et al.
Thus, we examined the magnitude and the kinetics of the BMresponses following immunization by looking at the individ-ual and mean frequency of IgA and IgG BM cells in recipients
Figure 3 Kinetics of NV-specific BM responses and correlationof BM data. A. Mean % NV BM SFC per total IgA or IgG-secretingcells±SE for each group; *, pb0.05 compared with unvaccinatedcontrols. B. Spearman's correlations between the frequency ofBM responses reported as NV BM SFC or % NV BM SFC per total IgAor IgG-secreting cells in expanded cultures. Data shown are fromindividual BM measurements in all volunteers and time points.Dotted lines indicate 95% confidence intervals.
of the 50 and 100 μg vaccine doses and controls, before andafter immunization (Fig. 3A). Peak BM responses were seen,for the most part, on day 56 after immunization. Whileresponses were also detected at day 180, their levels haddeclined significantly by then. The only exception was thefrequency of BM IgA cells in the 50 μg cohort, which waslower than other day 56 measurements and similar both ondays 56 and 180. The highest responses for IgA BM were seenin recipients of the 100 μg dose. For IgG BM cells, both the 50and 100 μg cohorts developed robust responses (on day 56).Although the magnitude appears to be higher for the 50 μgcohort, the difference was not statistically significant.
The frequency of antigen-specific BM cells is usuallyreported as a percent of the total number of antibody-secreting (spot forming) B cells to account for differences incell yield after expansion, but also can be reported as thenumber of antigen-specific SFC per 106 expanded PBMC[16,17]. However, results might differ depending on howthese frequencies were determined [17]. To confirm thatour BM results remained the same regardless of howresponses were calculated, we compared the IgA and IgGBM frequencies obtained from all volunteers and time points(in aggregate) reported in both ways. Strong correlationswere demonstrated between the two reporting methods forboth IgA and IgG BM (Fig. 3B), thus confirming the validityand accuracy of our analysis.
3.3. Memory ALS produced by expanded NV-specificBM cells
To demonstrate the functional capacity of the expanded BMcells, we examined their ability to produce and secreteantibodies. The level of NV-specific and total antibodiesproduced by the BM lymphocytes in culture supernatants(memory ALS) was measured by ELISA. Fig. 4 shows NV IgA andIgG ALS responses reported as ALS titers (Fig. 4A) and foldincreases over baseline (Fig. 4B). NV IgA and IgG memoryantibodies were detected in B cell cultures from vaccinerecipients in levels that greatly exceeded those of unvacci-nated controls. There was also a trend of higher antibodyproduction by cells collected on day 56. IgA ALS titers reached

Figure 5 Spearman's correlations between BM responses andserum antibody titers. A. Correlation between frequency ofNV-specific BM cells and serum IgA and IgG titers measured byELISA B. Correlation between BM cell frequency and serum antibodytiters measured by hemagglutination inhibition (HAI). Data (A andB) represent individual BM antibody measurements from volunteersincluded in this ancillary study (as described in Materials andmethods) at days 0, 56 and 180. Dotted lines indicate 95%confidence intervals.
Figure 4 Production of IgG and IgA antibodies by NV-specificBM cells. A. NV-specific IgA and IgG produced by BM cells duringthe 6-day expansion and differentiation period. Culture super-natants were collected at the end of the incubation period andNV IgA and IgG produced by these cells were measured by ELISA(memory antibody in lymphocyte supernatants, ALS). Datarepresent mean ELISA Units end point titers±SE for vaccineand control groups *, pb0.05 as compared to the control group.B. Mean fold increases in NV-specific IgA and IgG ALS titers±SE.C. Spearman's correlation between the frequency of BM cellsreported as % NV SFC per total (IgA or IgG) and ALS titers. Datashown correspond to individual measurements from all volun-teers and time points. Dotted lines indicate 95% confidenceintervals.
103Intranasal vaccination with an adjuvanted Norwalk virus-like particle vaccine
similar peak levels both in the 50 and 100 μg-dose cohorts,whereas higher IgG ALS responses were seen in subjects thatreceived the 50 μg dose (Figs. 4A and B). When the antibodytiters were analyzed as fold-increases over baseline levels,peak ALS responses were seen on day 56, regardless of thedose of vaccine given (Fig. 5B). Interestingly, culturesupernatants contained approximately 10 times morevaccine-specific IgG, as compared with IgA, suggesting thatthe BM cells detected were mostly IgG-producing cells or that
more IgG was produced on an individual cell basis, than IgA(Fig. 4B).
The BM responses for all volunteers and time points alsowere examined in relation to their ALS responses. As shownin Fig. 4C, the frequencies of NV IgA and IgG SFC weresignificantly correlated with the levels of memory ALSantibodies. A closer association was seen between IgG BMand IgG ALS responses (r=0.75, pb0.0001). These resultssuggest that memory ALS could be a valid and simplermethod to assess vaccine-induced BM responses.
3.4. Correlations between NV BM responses andserum antibodies
Because of the key role that BM cells play in maintaining thepool of plasma cells and the level of circulating antibodies,we also examined the NV IgA and IgG BM responses in theirassociation with serum antibody levels. A significant corre-lation was found between the frequency of IgA and IgG BMcells and serum antibody titers measured by ELISA (Fig. 5A,

Figure 6 Kinetics of mucosally primed NV antibody secretingcells (ASC) in NV-VLP vaccine recipient. NV specific IgA and IgGASC were measured in PBMC from vaccine recipients andcontrols before vaccination (day 0) and booster dose (day 21)and 7 days after each immunization (days 7 and 28). Datarepresents mean ASC×106 cells from all subjects included in thestudy±SE. *pb0.05 as compared to the control group.
104 K. Ramirez et al.
data from all subjects and time points were included in thecomparison). An association was also observed between BMresponses and serum HAI titers. Although less stringent thanfor ELISA antibodies, the correlations were nonethelesssignificant for both immunoglobulin types (Fig. 5B).
3.5. Mucosally primed ASC and their association toBM responses
We had previously shown that intranasal immunization withthe NV-VLP vaccine induced IgA ASC expressing gut and lymphnode-homing markers [10]. In this study we further examinedthe magnitude and kinetics of the ASC responses after eachimmunization and investigated a possible association with thefrequency of BM cells (Fig. 6 and Table 1). Both IgA and IgG ASCresponses had similar a time-course; a transient peak wasdetected 1 week after each immunization, preceded byundetectable pre-vaccination levels. Interestingly, the mag-nitude of the responses did not increase following the boosterdose given on day 21. In fact, the ASC counts were actuallylower on day 28, 7 days after the second vaccine dose, ascompared to 7 days following the first immunization. Vacci-nees who received the 100 μg dose had 4-fold higher IgA ASCresponses than those who received the 50 μg dose, althoughboth groups responded similarly, with high numbers of IgG ASCat peak time (day 7). No ASC responses were detected inunvaccinated subjects (Fig. 6). Significant correlations werefound between peak BM responses and the frequency of
Table 1 Spearman's correlation between the frequency of NV-sp
% NV-specific IgA/totalIgA SFC×106
Day 56
NV IgA ASC×106 cells R p
Day 7 0.68 0.0001Day 28 0.51 0.0097
mucosally primed ASC (Table 1), particularly for IgA on day 7,and for IgG on day 28 post- vaccination.
3.6. Compiled analysis of NV-VLP responders
Finally, we summarized the immunogenicity results from thePhase 1 NV-VLP vaccine studies based on the most sensitivereadouts, as we identified them in this ancillary study. Fig. 7shows vaccine-induced responders among recipients of the50 and 100 μg dose based on peak responses for serumantibodies (Fig. 7A), mucosally primed ASC (Fig. 7B) and ALS(Fig. 7C). The data analyzed in aggregate (for the 19volunteers included in this study) clearly show a higherproportion of responses being detected among subjects whoreceived the 100 μg dose of NV-VLP vaccine.
4. Discussion
One of the main benefits of active immunization is itscapacity to engender immunologic memory that would allowan individual to respond quickly and vigorously when re-exposed to microbial pathogens. Following vaccination,naïve B cells that have encountered foreign antigens willexpand and differentiate into antibody-secreting plasmacells, including long-lived plasma cells, and BM cells [18].Antibody-secreting plasma cells (or plasmablasts) are usuallydetected in circulation as antigen-specific ASC the first weekafter antigen exposure. These cells have a short life span andproduce a first wave of less refined antibodies [18]. Long-lived plasma cells, on the other hand, persist longer and helpto maintain a constant (and adequate) level of serumantibodies long after the antigen has been cleared [19].These cells have undergone isotype switch and affinitymaturation, and the antibodies produced are of enhancedquality (i.e., higher avidity and functional capacity). BM cellsrepresent the “central” reservoir of humoral defenses; theydo not secrete antibodies but will “reactivate” uponantigenic re-exposure, proliferating and differentiating intoplasma cells that will enter the circulation and produceantibodies. They also replenish the pool of long-lived plasmacells. BM cells and long-lived plasma cells are generated inthe germinal centers (GC) and from there they migrate toniche lymphoid tissues. Plasma cells migrate to the bonemarrow, and BM cells to bone marrow and secondarylymphoid organs [19]. The development of BM cells duringthe induction of adaptive immunity is critical to maintainantibody production and to ensure a rapid and robustanamnestic humoral response upon antigenic challenge.This is an essential requirement for a vaccine candidate toprovide long-term immunologic memory [20].
ecific BM responses and mucosally primed NV-specific ASC.
% NV specific IgG /totalIgG SFC×106
Day 56
NV IgG ASC×106 cells r P
Day 7 0.55 0.0039Day 28 0.67 0.0002
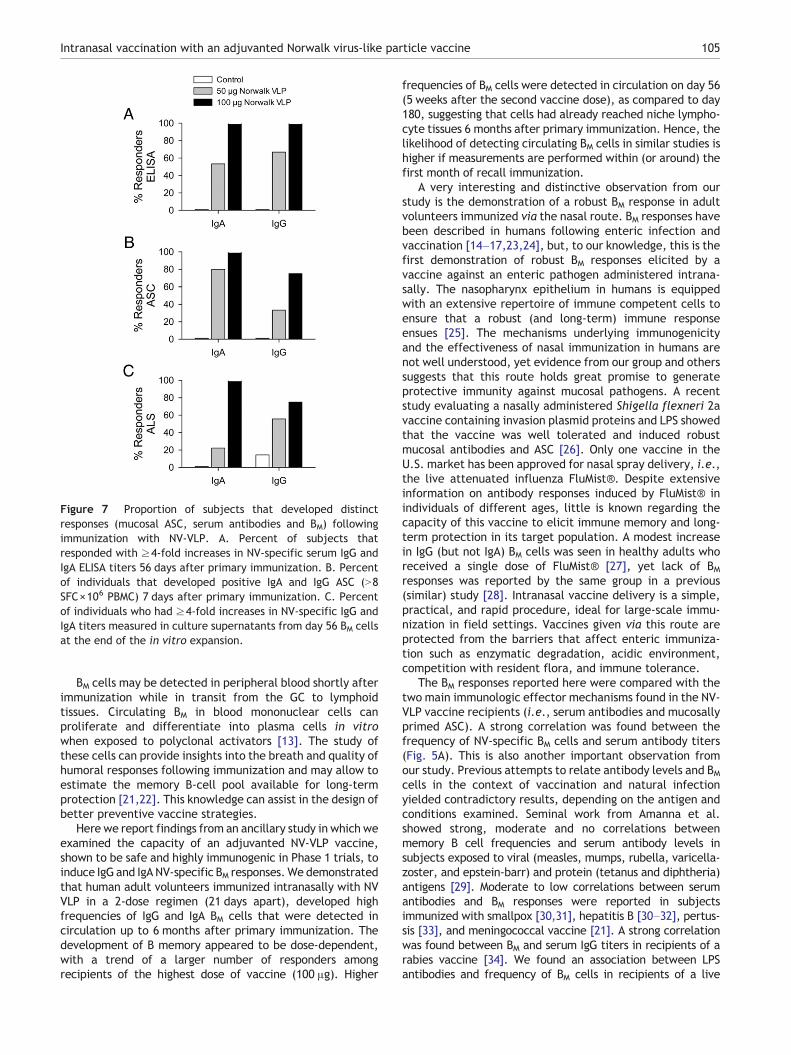
Figure 7 Proportion of subjects that developed distinctresponses (mucosal ASC, serum antibodies and BM) followingimmunization with NV-VLP. A. Percent of subjects thatresponded with ≥4-fold increases in NV-specific serum IgG andIgA ELISA titers 56 days after primary immunization. B. Percentof individuals that developed positive IgA and IgG ASC (N8SFC×106 PBMC) 7 days after primary immunization. C. Percentof individuals who had ≥4-fold increases in NV-specific IgG andIgA titers measured in culture supernatants from day 56 BM cellsat the end of the in vitro expansion.
105Intranasal vaccination with an adjuvanted Norwalk virus-like particle vaccine
BM cells may be detected in peripheral blood shortly afterimmunization while in transit from the GC to lymphoidtissues. Circulating BM in blood mononuclear cells canproliferate and differentiate into plasma cells in vitrowhen exposed to polyclonal activators [13]. The study ofthese cells can provide insights into the breath and quality ofhumoral responses following immunization and may allow toestimate the memory B-cell pool available for long-termprotection [21,22]. This knowledge can assist in the design ofbetter preventive vaccine strategies.
Here we report findings from an ancillary study in which weexamined the capacity of an adjuvanted NV-VLP vaccine,shown to be safe and highly immunogenic in Phase 1 trials, toinduce IgG and IgA NV-specific BM responses.We demonstratedthat human adult volunteers immunized intranasally with NVVLP in a 2-dose regimen (21 days apart), developed highfrequencies of IgG and IgA BM cells that were detected incirculation up to 6 months after primary immunization. Thedevelopment of B memory appeared to be dose-dependent,with a trend of a larger number of responders amongrecipients of the highest dose of vaccine (100 μg). Higher
frequencies of BM cells were detected in circulation on day 56(5 weeks after the second vaccine dose), as compared to day180, suggesting that cells had already reached niche lympho-cyte tissues 6 months after primary immunization. Hence, thelikelihood of detecting circulating BM cells in similar studies ishigher if measurements are performed within (or around) thefirst month of recall immunization.
A very interesting and distinctive observation from ourstudy is the demonstration of a robust BM response in adultvolunteers immunized via the nasal route. BM responses havebeen described in humans following enteric infection andvaccination [14–17,23,24], but, to our knowledge, this is thefirst demonstration of robust BM responses elicited by avaccine against an enteric pathogen administered intrana-sally. The nasopharynx epithelium in humans is equippedwith an extensive repertoire of immune competent cells toensure that a robust (and long‐term) immune responseensues [25]. The mechanisms underlying immunogenicityand the effectiveness of nasal immunization in humans arenot well understood, yet evidence from our group and otherssuggests that this route holds great promise to generateprotective immunity against mucosal pathogens. A recentstudy evaluating a nasally administered Shigella flexneri 2avaccine containing invasion plasmid proteins and LPS showedthat the vaccine was well tolerated and induced robustmucosal antibodies and ASC [26]. Only one vaccine in theU.S. market has been approved for nasal spray delivery, i.e.,the live attenuated influenza FluMist®. Despite extensiveinformation on antibody responses induced by FluMist® inindividuals of different ages, little is known regarding thecapacity of this vaccine to elicit immune memory and long-term protection in its target population. A modest increasein IgG (but not IgA) BM cells was seen in healthy adults whoreceived a single dose of FluMist® [27], yet lack of BMresponses was reported by the same group in a previous(similar) study [28]. Intranasal vaccine delivery is a simple,practical, and rapid procedure, ideal for large-scale immu-nization in field settings. Vaccines given via this route areprotected from the barriers that affect enteric immuniza-tion such as enzymatic degradation, acidic environment,competition with resident flora, and immune tolerance.
The BM responses reported here were compared with thetwo main immunologic effector mechanisms found in the NV-VLP vaccine recipients (i.e., serum antibodies and mucosallyprimed ASC). A strong correlation was found between thefrequency of NV-specific BM cells and serum antibody titers(Fig. 5A). This is also another important observation fromour study. Previous attempts to relate antibody levels and BMcells in the context of vaccination and natural infectionyielded contradictory results, depending on the antigen andconditions examined. Seminal work from Amanna et al.showed strong, moderate and no correlations betweenmemory B cell frequencies and serum antibody levels insubjects exposed to viral (measles, mumps, rubella, varicella-zoster, and epstein-barr) and protein (tetanus and diphtheria)antigens [29]. Moderate to low correlations between serumantibodies and BM responses were reported in subjectsimmunized with smallpox [30,31], hepatitis B [30–32], pertus-sis [33], and meningococcal vaccine [21]. A strong correlationwas found between BM and serum IgG titers in recipients of arabies vaccine [34]. We found an association between LPSantibodies and frequency of BM cells in recipients of a live

106 K. Ramirez et al.
attenuated Shigella flexneri 2a vaccine [15]. Interestingly, thiswas true when BM frequencies were measured by ELISpot asopposed to flow cytometry, the latter being apparently lesssensitive [32]. A trend that emerges from these studies andours is that BM and serum antibody responses seem to be closelyassociated in individuals who had been immunized recently andreceived multiple doses of vaccine. The agreement betweenthe magnitude of serum antibody and BM responses in re-cipients of the NV-VLP vaccine suggest that the BM cellsdetected in circulation are contributing to the supply ofsystemic antibodies, possibly through maintenance of plasmacells. Thus, the BM cells detected in our system may representan indirect measurement of the existing pool of plasma cells.The association between the frequency of circulating BM cellsand mucosally derived ASC (detected in blood 7 days aftervaccination) suggest that naïve B cells primed in the mucosaare contributing to the pool of BM cells detected in the NV-VLPvaccinees. In fact, one would expect the frequency of BM cellsmeasured in circulation to be proportional to the number of Bcells primed in the mucosa. After differentiation, these cellsmay home back to effector sites as long-lived plasma cells [35].The correspondence between BM, serum antibody and ASCresponses also shows the consistency of the immunologicalreadouts following immunization and attests to the robustnessof the NV-VLP vaccine.
Another important observation in our study is theassociation between the frequency of NV IgG and IgA BMcells and the level of antibodies produced by these cellsduring expansion (memory ALS). This finding confirms thatthe BM cells measured were functionally active and that thememory ALS could be used as a practical alternative methodto demonstrate vaccine-induced B memory. An advantage ofthe ALS results is that they may reflect more accurately thenumber of BM cells originally present in blood. In contrast,when using ELISpot, BM frequencies are calculated based oncells that have been expanded, recounted and added to theplates. The usefulness of measuring BM-secreted antibodies(ALS) as indicative of long-term B cell-mediated protectionhas been reported by our group [14] and others [21].
The immunological effector mechanisms that mediateprotection against NoV infection remain unclear. Antibodiesthat block histoblood group antigens [11] and HAI antibodies[36] have been associated with protection against clinicalgastroenteritis. While antibodies are typically needed toprevent viral infection, the controversy remains as to thedegree of their contribution in overall protection againstNoV. Mucosal antibodies, as opposed to serum antibodies,are likely to play a more prominent role in host defenseagainst this intestinal viral pathogen. An early mucosal IgAresponse has been associated with protection from NVinfection in human challenge studies [37]. It is unclearwhether the NV-VLP-induced BM cells we detected incirculation contribute to the production of secretory IgA.Fecal IgA measurements in BM study participants whoreceived 50 μg of the VLP failed to detect vaccine-inducedresponses, except for a transient 2-fold rise over baselinefollowing the first immunization (data not shown). Theassociation between serum antibodies and BM cells argues infavor of these cells maintaining systemic rather thanmucosal antibodies. However, BM cells or plasma cellsoriginated through mucosal vaccination can presumablyhome back to mucosal tissues, thereby contributing to the
production of local antibodies. A correlation was seenbetween LPS-specific IgA BM cells, mucosal ASC andsecretory IgA in subjects immunized with live oral S. flexneri2a strains [16]. The analysis of mucosal (i.e., fecal)antibodies along with ASC and BM cells in future NV-VLPstudies will allow the evaluation of this hypothesis. Thecontribution of cell-mediated immunity in protection alsorequires elucidation. T cell proliferation and IFN-γ produc-tion were reported in volunteers immunized orally with NV-VLP [38]. Importantly, a predominant CD4-dependent IFN-γ-mediated Th1-type response was seen in a subset ofvolunteers who remained uninfected when challenged withGII NoV, even in the absence of IgG responses [39].
A challenge study has recently been completed in 84adult human subjects who received 2 doses (100 μg each) ofthe NV-VLP vaccine or placebo. The vaccine was generallywell tolerated and had a 47% efficacy (or relative reduction)against any NoV illness, including mild illness, and a 26%efficacy against NoV infection [40]. In the 77 adults whocompleted the trial as per its original protocol, vaccinationdecreased the incidence of acute gastroenteritis due to NoVfrom 69% to 37% and the incidence of NoV infection from 82%to 61%. The severity of illness also was significantly reducedin the vaccinees [40]. This remarkable protection outcomeand our reported evidence of immunologic memory in NV-VLP recipients both confirm the effectiveness of this vaccineand support its potential usefulness.
Because of the high genetic variability among humanNoV, a vaccine that would confer broad protection againstmultiple virus strains is greatly desirable. This, in fact,represents one of the major challenges in the developmentof prophylactic tools to prevent this disease. The initialPhase 1 studies described herein were designed to provide aproof of concept for safety and immunogenicity of amucosally delivered NoV GI.1- derived VLP vaccine. Becausethere is only one (GI.1) NoV strain that has been approved bythe Food and Drug Administration for use in humanchallenges, efforts were focused on a monovalent GI.1vaccine as a suitable model for evaluating protectiveefficacy in vaccinated subjects. A clinical study is nowbeing conducted to evaluate a bivalent GI.1 and GII.4, MPL-adjuvanted, NV-VLP vaccine that would expand the protec-tion coverage to the most predominantly circulating NoVstrains (ClinicalTrials.gov NCT01168401).
In conclusion, our studies demonstrate that intranasalimmunization of human adult volunteers with an adjuvantedNV-VLP vaccine was safe and resulted in robust immuneresponses, which eventually lead to protection, and immu-nologic memory. The demonstration of high frequencies ofBM responses supports the notion that this vaccine mightprovide long-term protection against disease through per-sistence of antibodies. The strategy appears to be verypromising to prevent NoV infection in humans.
Conflict of interest statement
The authors declare that there are no conflicts of interest.

107Intranasal vaccination with an adjuvanted Norwalk virus-like particle vaccine
Acknowledgments
We thank Dr. S. Crotty for providing pokeweed mitogen andpersonnel from the Applied Immunology Section for performingserological measurements. This work was supported by grants fromLigoCyte Pharmaceuticals, Inc. The nasal vaccine incorporateschitosan. This application of chitosan (ChiSys®) has been licensedfrom Archimedes Development Ltd. ChiSys is a trademark ofArchimedes Development Ltd., and is registered as a CTM, as a USRegistered Trademark and in certain other jurisdictions.
References
[1] R.I. Glass, U.D. Parashar, M.K. Estes, Norovirus gastroenteritis,N. Engl. J. Med. 361 (2009) 1776–1785.
[2] P.S. Mead, L. Slutsker, V. Dietz, L.F. McCaig, J.S. Bresee, C.Shapiro, P.M. Griffin, R.V. Tauxe, Food-related illness anddeath in the United States, Emerg. Infect. Dis. 5 (1999)607–625.
[3] H.L. Koo, N. Ajami, R.L. Atmar, H.L. DuPont, Noroviruses: theleading cause of gastroenteritis worldwide, Discov. Med. 10(2010) 61–70.
[4] J.A. Marshall, L.D. Bruggink, The dynamics of norovirusoutbreak epidemics: recent insights, Int. J. Environ. Res.Public Health 8 (2011) 1141–1149.
[5] M.K. Estes, B.V. Prasad, R.L. Atmar, Noroviruses everywhere:has something changed? Curr. Opin. Infect. Dis. 19 (2006)467–474.
[6] Norovirus: Surveillance, Disease Burden, Center for DiseaseControl and Prevention. National Center for Immunization andRepiratory Disease, Division of Viral Diseases, 2011 , http://www.cdc.gov/ncidod/dvrd/revb/gastro/norovirus-surv-disease-burden.htm.
[7] M. Herbst-Kralovetz, H.S. Mason, Q. Chen, Norwalk virus-likeparticles as vaccines, Expert Rev. Vaccines 9 (2010) 299–307.
[8] A. Roldao, M.C. Mellado, L.R. Castilho, M.J. Carrondo, P.M.Alves, Virus-like particles in vaccine development, Expert Rev.Vaccines 9 (2010) 1149–1176.
[9] C. Lopez-Macias, E. Ferat-Osorio, A. Tenorio-Calvo, A. Isibasi,J. Talavera, O. rteaga-Ruiz, L. rriaga-Pizano, S.P. Hickman, M.Allende, K. Lenhard, S. Pincus, K. Connolly, R. Raghunandan,G. Smith, G. Glenn, Safety and immunogenicity of a virus-likeparticle pandemic influenza A (H1N1) 2009 vaccine in ablinded, randomized, placebo-controlled trial of adults inMexico, Vaccine 29 (2011) 7826–7834.
[10] S.S. el-Kamary, M.F. Pasetti, P.M. Mendelman, S.E. Frey, D.I.Bernstein, J.J. Treanor, J. Ferreira, W.H. Chen, R. Sublett, C.Richardson, R.F. Bargatze, M.B. Sztein, C.O. Tacket, Adju-vanted intranasal Norwalk virus-like particle vaccine elicitsantibodies and antibody-secreting cells that express homingreceptors for mucosal and peripheral lymphoid tissues, J. Infect.Dis. 202 (2010) 1649–1658.
[11] A. Reeck, O. Kavanagh, M.K. Estes, A.R. Opekun, M.A. Gilger,D.Y. Graham, R.L. Atmar, Serological correlate of protectionagainst norovirus-induced gastroenteritis, J. Infect. Dis. 202(2010) 1212–1218.
[12] I.J. Amanna, M.K. Slifka, Mechanisms that determine plasmacell lifespan and the duration of humoral immunity, Immunol.Rev. 236 (2010) 125–138.
[13] S. Crotty, R.D. Aubert, J. Glidewell, R. Ahmed, Trackinghuman antigen-specific memory B cells: a sensitive andgeneralized ELISPOT system, J. Immunol. Methods 286 (2004)111–122.
[14] R. Wahid, M.F. Pasetti, M. Maciel Jr., J.K. Simon, C.O. Tacket,M.M. Levine, M.B. Sztein, Oral priming with Salmonella typhivaccine strain CVD 909 followed by parenteral boost with the
S. typhi Vi capsular polysaccharide vaccine induces CD27+IgD-S. typhi-specific IgA and IgG B memory cells in humans, Clin.Immunol. 138 (2011) 187–200.
[15] J.K. Simon, R. Wahid, M. Maciel Jr., W.L. Picking, K.L. Kotloff,M.M. Levine, M.B. Sztein, Antigen-specific B memory cellresponses to lipopolysaccharide (LPS) and invasion plasmidantigen (Ipa) B elicited in volunteers vaccinated with live-attenuated Shigella flexneri 2a vaccine candidates, Vaccine 27(2009) 565–572.
[16] J.K. Simon, M. Maciel Jr., E.D. Weld, R. Wahid, M.F. Pasetti,W.L. Picking, K.L. Kotloff, M.M. Levine, M.B. Sztein, Antigen-specific IgA B memory cell responses to Shigella antigenselicited in volunteers immunized with live attenuated Shigellaflexneri 2a oral vaccine candidates, Clin. Immunol. 139 (2011)185–192.
[17] D.T. Leung, M.A. Rahman, M. Mohasin, M.A. Riyadh, S.M. Patel,M.M. Alam, F. Chowdhury, A.I. Khan, E.J. Kalivoda, A. Aktar,M.S. Bhuiyan, R.C. LaRocque, J.B. Harris, S.B. Calderwood, F.Qadri, E.T. Ryan, Comparison of memory B cell, antibody-secreting cell, and plasma antibody responses in youngchildren, older children, and adults with infection caused byVibrio cholerae O1 El Tor Ogawa in Bangladesh, Clin. VaccineImmunol. 18 (2011) 1317–1325.
[18] S. Crotty, R. Ahmed, Immunological memory in humans, Semin.Immunol. 16 (2004) 197–203.
[19] S.G. Tangye, Staying alive: regulation of plasma cell survival,Trends Immunol. 32 (2011) 595–602.
[20] M. Heyzer-Williams, S. Okitsu, N. Wang, L. Heyzer-Williams,Molecular programming of B cell memory, Nat. Rev. Immunol.12 (2011) 24–34.
[21] G. Blanchard-Rohner, G. Galli, E.A. Clutterbuck, A.J. Pollard,Comparison of a limiting dilution assay and ELISpot fordetection of memory B-cells before and after immunisationwith a protein–polysaccharide conjugate vaccine in children,J. Immunol. Methods 358 (2010) 46–55.
[22] E.A. Clutterbuck, P. Salt, S. Oh, A. Marchant, P. Beverley, A.J.Pollard, The kinetics and phenotype of the human B-cell responsefollowing immunization with a heptavalent pneumococcal-CRMconjugate vaccine, Immunology 119 (2006) 328–337.
[23] M.M. Alam, M.A. Riyadh, K. Fatema, M.A. Rahman, N. Akhtar,T. Ahmed, M.I. Chowdhury, F. Chowdhury, S.B. Calderwood,J.B. Harris, E.T. Ryan, F. Qadri, Antigen-specific memory B-cellresponses in Bangladeshi adults after one- or two-dose oral killedcholera vaccination and comparison with responses in patientswith naturally acquired cholera, Clin. Vaccine Immunol. 18(2011) 844–850.
[24] C.R. Jayasekera, J.B. Harris, S. Bhuiyan, F. Chowdhury, A.I.Khan, A.S. Faruque, R.C. LaRocque, E.T. Ryan, R. Ahmed, F.Qadri, S.B. Calderwood, Cholera toxin-specific memory B cellresponses are induced in patients with dehydrating diarrheacaused by Vibrio cholerae O1, J. Infect. Dis. 198 (2008)1055–1061.
[25] H. Kiyono, S. Fukuyama, NALT- versus Peyer's-patch-mediatedmucosal immunity, Nat. Rev. Immunol. 4 (2004) 699–710.
[26] M.S. Riddle, R.W. Kaminski, C. Williams, C. Porter, S. Baqar, A.Kordis, T. Gilliland, J. Lapa, M. Coughlin, C. Soltis, E. Jones, J.Saunders, P.B. Keiser, R.T. Ranallo, R. Gormley, M. Nelson,K.R. Turbyfill, D. Tribble, E.V. Oaks, Safety and immunogenic-ity of an intranasal Shigella flexneri 2a Invaplex 50 vaccine,Vaccine 29 (2011) 7009–7019.
[27] S. Sasaki, X.S. He, T.H. Holmes, C.L. Dekker, G.W. Kemble, A.M.Arvin, H.B. Greenberg, Influence of prior influenza vaccinationon antibody and B-cell responses, PLoS One 3 (2008) e2975.
[28] S. Sasaki, M.C. Jaimes, T.H. Holmes, C.L. Dekker, K. Mahmood,G.W. Kemble, A.M. Arvin, H.B. Greenberg, Comparison of theinfluenza virus-specific effector and memory B-cell responses toimmunization of children and adults with live attenuated orinactivated influenza virus vaccines, J. Virol. 81 (2007) 215–228.

108 K. Ramirez et al.
[29] I.J. Amanna, N.E. Carlson, M.K. Slifka, Duration of humoralimmunity to common viral and vaccine antigens, N. Engl. J.Med. 357 (2007) 1903–1915.
[30] S. Crotty, P. Felgner, H. Davies, J. Glidewell, L. Villarreal, R.Ahmed, Cutting edge: long-term B cell memory in humans aftersmallpox vaccination, J. Immunol. 171 (2003) 4969–4973.
[31] E. Tuaillon, Y.A. Tabaa, G. Petitjean, M.F. Huguet, G. Pajeaux,J.M. Fondere, B. Ponseille, J. Ducos, P. Blanc, J.P. Vendrell,Detection of memory B lymphocytes specific to hepatitis Bvirus (HBV) surface antigen (HBsAg) from HBsAg-vaccinated orHBV-immunized subjects by ELISPOT assay, J. Immunol.Methods 315 (2006) 144–152.
[32] S.M. Ward, P. Phalora, D. Bradshaw, H. Leyendeckers, P.Klenerman, Direct ex vivo evaluation of long-lived protectiveantiviral memory B cell responses against hepatitis B virus,J. Infect. Dis. 198 (2008) 813–817.
[33] A.M. Buisman, C.G. de Rond, K. Ozturk, H.I. Ten Hulscher, R.S.van Binnendijk, Long-term presence of memory B-cells specificfor different vaccine components, Vaccine 28 (2009) 179–186.
[34] G. Blanchard-Rohner, A.S. Pulickal, C.M. Jol-van der Zijde,M.D. Snape, A.J. Pollard, Appearance of peripheral bloodplasma cells and memory B cells in a primary and secondaryimmune response in humans, Blood 114 (2009) 4998–5002.
[35] D.T. Avery, J.I. Ellyard, F. Mackay, L.M. Corcoran, P.D. Hodgkin,S.G. Tangye, Increased expression of CD27 on activated human
memory B cells correlates with their commitment to the plasmacell lineage, J. Immunol. 174 (2005) 4034–4042.
[36] R. Czako, R.L. Atmar, A.R. Opekun, M.A. Gilger, D.Y.Graham, M.K. Estes, Serum hemagglutination inhibitionactivity correlates with protection from gastroenteritis inpersons infected with Norwalk virus, Clin. Vaccine Immunol.19 (2012) 284–287.
[37] L. Lindesmith, C. Moe, S. Marionneau, N. Ruvoen, X. Jiang, L.Lindblad, P. Stewart, J. LePendu, R. Baric, Human suscepti-bility and resistance to Norwalk virus infection, Nat. Med. 9(2003) 548–553.
[38] C.O. Tacket, M.B. Sztein, G.A. Losonsky, S.S. Wasserman, M.K.Estes, Humoral, mucosal, and cellular immune responses tooral Norwalk virus-like particles in volunteers, Clin. Immunol.108 (2003) 241–247.
[39] L. Lindesmith, C. Moe, J. LePendu, J.A. Frelinger, J. Treanor,R.S. Baric, Cellular and humoral immunity following SnowMountain virus challenge, J. Virol. 79 (2005) 2900–2909.
[40] R.L. Atmar, D.I. Bernstein, C.D. Harro, M.S. Al-Ibrahim, W.H.Chen, J. Ferreira, M.K. Estes, D.Y. Graham, A.R. Opekun, C.Richardson, P.M. Mendelman, Norovirus vaccine against exper-imental human Norwalk Virus illness, N. Engl. J. Med. 365(2011) 2178–2187.





























