Embed Size (px)
Citation preview

Intestinal Alkaline Phosphatase Deficiency Leads toLipopolysaccharide Desensitization and Faster Weight Gain
Ye Yang,a* José Luis Millán,b Joan Mecsas,c Karen Guillemina
Institute of Molecular Biology, University of Oregon, Eugene, Oregon, USAa; Sanford Children’s Health Research Center, Sanford-Burnham Medical Research Institute, LaJolla, California, USAb; Department of Molecular Biology and Microbiology, Tufts University School of Medicine, and Graduate Program in Molecular Microbiology, SacklerSchool of Biomedical Sciences, Boston, Massachusetts, USAc
Animals develop in the presence of complex microbial communities, and early host responses to these microbes can influencekey aspects of development, such as maturation of the immune system, in ways that impact adult physiology. We previouslyshowed that the zebrafish intestinal alkaline phosphatase (ALPI) gene alpi.1 was induced by Gram-negative bacterium-derivedlipopolysaccharide (LPS), a process dependent on myeloid differentiation primary response gene 88 (MYD88), and functioned todetoxify LPS and prevent excessive host inflammatory responses to commensal microbiota in the newly colonized intestine. Inthe present study, we examined whether the regulation and function of ALPI were conserved in mammals. We found that amongthe mouse ALPI genes, Akp3 was specifically upregulated by the microbiota, but through a mechanism independent of LPS orMYD88. We showed that disruption of Akp3 did not significantly affect intestinal inflammatory responses to commensal micro-biota or animal susceptibility to Yersinia pseudotuberculosis infection. However, we found that Akp3�/� mice acquired LPS tol-erance during postweaning development, suggesting that Akp3 plays an important role in immune education. Finally, we dem-onstrated that inhibiting LPS sensing with a mutation in CD14 abrogated the accelerated weight gain in Akp3�/� mice receivinga high-fat diet, suggesting that the weight gain is caused by excessive LPS in Akp3�/� mice.
Mammals coexist with a consortium of microorganisms, theirmicrobiota. The most populous microbial community is
present in the gastrointestinal tract. A mutually beneficial rela-tionship has been forged between the host and its associated gutmicrobiota. While the host intestine provides a nutrient-rich en-vironment for the microbes, the gut microbiota modulate hostmetabolism (1), promote immune maturation (2), preserve gutepithelial barrier function (3), and prevent growth of pathogens(4). Despite these benefits, the intestinal microbiota are a contin-uous source of antigens and toxins, which can provoke host in-flammatory responses. Regulatory mechanisms therefore mustexist to prevent unlimited immune activation by microbial prod-ucts.
Alkaline phosphatases (ALPs) are a superfamily of metalloen-zymes that are widely found in organisms ranging from bacteria tohumans (5) and catalyze the hydrolytic removal of phosphatefrom a variety of molecules (6). Importantly, ALPs have beenshown to remove the lipid A phosphates of the endotoxin lipo-polysaccharide (LPS) (7–10). LPS is a constituent of the outermembrane of Gram-negative bacteria (11) which compose a sub-stantial proportion of the mammalian gut microbiota (12). Inmammals, LPS is transferred by LPS binding protein to CD14,which then presents the molecule to the TLR4 (Toll-like receptor4)/MD-2 receptor complex, resulting in the activation of innateimmune signaling. The two phosphate groups of the LPS lipid Amoiety support stable binding to the receptor complex, and de-phosphorylation of lipid A greatly reduces the inflammatory ac-tivity of LPS (13). Although this mechanism of LPS sensing is notconserved in nonmammalian species (e.g., fish lack orthologs ofCD14 and MD-2 and have functionally distinct TLR4 homologs[14, 15]), the dephosphorylation of LPS by ALPs ameliorates LPS-induced inflammation and reduces LPS endotoxic properties in awide range of organisms from cephalopod to tetrapods (16–20).
The mammalian ALP family consists of several isozymes which
can be classified as tissue-nonspecific (liver-bone-kidney type)and tissue-specific ALPs (intestinal, placental, and germ cell type)(6, 12). The intestinal ALP (ALPI) is abundantly present in theapical microvilli of the brush border of enterocytes (21) and ac-tively secreted into the intestinal lumen (22, 23). Recent studieshave shown that ALPI regulates neutral pH in the intestine (24),protects gut barrier function (18, 25–27), and preserves gut mi-crobial homeostasis (28, 29).
Our studies using zebrafish larvae have demonstrated an inte-gral role of ALPI in establishing a balanced host immune responseto the microbiota during gut colonization (16, 30). We showedthat intestinal colonization of Gram-negative bacteria upregu-lated zebrafish ALPI gene alpi.1 through an LPS-induced innateimmune signaling mechanism that required Myd88. Further-more, we showed that ALPI functioned to prevent excessiveinflammatory responses to the resident gut microbiota by detox-ifying LPS. The LPS-ALPI negative-feedback loop therefore pro-motes zebrafish immune tolerance to commensal Gram-negativebacteria. Analysis of the evolution of vertebrate ALPI genes re-
Received 18 August 2014 Returned for modification 16 September 2014Accepted 17 October 2014
Accepted manuscript posted online 27 October 2014
Citation Yang Y, Millán JL, Mecsas J, Guillemin K. 2015. Intestinal alkalinephosphatase deficiency leads to lipopolysaccharide desensitization and fasterweight gain. Infect Immun 83:247–258. doi:10.1128/IAI.02520-14.
Editor: A. J. Bäumler
Address correspondence to Karen Guillemin, [email protected].
* Present address: Ye Yang, Department of Medicine, University of Florida,Gainesville, Florida, USA.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.02520-14
January 2015 Volume 83 Number 1 iai.asm.org 247Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

vealed a dynamic history of gene losses and duplications and dem-onstrated that the extant zebrafish, mouse, and human genes arenot derived from a common descendant but instead arose inde-pendently in each lineage (12). This raises the question of whetherthe LPS-induced regulation and anti-inflammatory function ofALPI that we described in zebrafish are conserved in mammals. Toanswer this, we investigated the regulation and function of themouse ALPI gene Akp3 in host-microbe interactions.
The mouse genome contains four ALP genes: the tissue-non-specific type Alpl (MGI:87983), the embryonic Alppl2 (MGI:108009), the intestinal Alpi (MGI:1924018), and the duodenalAkp3 (MGI:87984) (6, 12). Alppl2, Alpi, and Akp3 all contribute toALP activity in the mouse intestine but exhibit distinct expressionpatterns (31). Alppl2 and Alpi are expressed after birth throughoutthe intestine but are enriched in the duodenum. In comparison,Akp3 expression starts around postnatal day 13 to 15 and is re-stricted to the duodenum. The concordant timing of Akp3 expres-sion with weaning and establishment of the adult gut microbiotaled us to test the possibility that Akp3 is induced by the microbiota.
Since the generation of Akp3�/� mice (32), several studies haveprobed the role of Akp3 in animal health and disease. Like otherALPs, AKP3 has LPS dephosphorylation activity, and Akp3�/�
mice had significantly lower ALP enzyme and LPS dephosphory-lating activities in stools and duodenal mucosa compared to wild-type littermates (18). Akp3�/� mice had increased gut permeabil-ity and hence increased LPS translocation from the intestinallumen into the blood (27). Consequently, Akp3�/� mice dis-played higher hepatic expression of major histocompatibilitycomplex class II molecules (20) and signs of metabolic endotox-emia (27). Akp3�/� mice were found to contain dramaticallyfewer and also different types of aerobic and anaerobic microbesin stools compared to wild-type mice (28, 29). Together, theseobservations from routinely maintained animals indicate thatAkp3 plays a role in maintaining gut barrier function and micro-bial homeostasis. The role of AKP3 as a gut mucosal defense factoris more evident when mice were challenged. Compared to wild-type mice, Akp3�/� mice suffered from more severe gut mucosaldisruption and higher bacterial translocation after ischemia/rep-erfusion injuries (18) and were more vulnerable to dextran sulfatesodium-induced colitis (33). Interestingly, Akp3�/� mice showedimmune tolerance to intraperitoneal LPS injection and resistanceto Salmonella enterica serovar Typhimurium infection (20).
All of the aforementioned results were obtained from full-grown mice, but little is known about the biological function ofAkp3 in developing animals. In this report, we compared intestinalinflammatory responses of wild-type and Akp3�/� mice at differ-ent ages to resident microbiota, orally administered LPS, and theGram-negative pathogen Yersinia pseudotuberculosis. Based onour previous observations in zebrafish (16), we speculated that thepartial reduction in ALPI activity due to disruption of one ALPIgene would result in subtle inflammatory reactions to the micro-biota in Akp3�/� mice during development, which might lead tolong-term changes in their immune sensitivity to future bacterialor LPS challenges. In addition, to test whether the reported resis-tance of Akp3�/� mice to colonic S. Typhimurium infection ispathogen or tissue specific, we investigated the requirement ofAkp3 for defense against infection by Y. pseudotuberculosis, whichinfects the mouse ileum, as well as the colon (34).
Finally, we explored a possible connection between the inflam-matory status of Akp3�/� mice and their accelerated weight gain
when maintained on a high-fat diet (HFD) (27, 32). Long-termHFD feeding has been shown to increase plasma LPS (35–38), andcontinuous infusion of LPS is sufficient to induce weight gain to asimilar extent as HFD feeding (36). Furthermore, loss-of-functionmutation of LPS receptors TLR4 or CD14 protects mice againstHFD-induced obesity (36, 39–41). We report that LPS sensing isrequired for the accelerated weight gain observed in Akp3�/� micereceiving HFD.
MATERIALS AND METHODSMice. Animals were maintained and all procedures were performed withapproval of the University of Oregon Institutional Animal Care and UseCommittee. Pairs of Akp3�/� C57BL/6 mice (32) were shipped from theMillán laboratory at the Sanford-Burnham Medical Research Institute, LaJolla, CA. Genotyping of Akp3�/� mice was performed as previously es-tablished (32). Pairs of Myd88�/� and CD14�/� C57BL/6 mice were ob-tained from the Jackson Laboratory, and genotyping was performed ac-cording to instructions in JAX mouse database. Heterozygous mice werebred at the University of Oregon animal facility to acquire homozygousmutants and wild-type littermates. For HFD feeding, a female Akp3�/�
mouse was crossed to a male CD14�/� mouse to generate Akp3�/�;CD14�/� mice, which were subsequently mated to produce progeny ofdesired genotypes. Mice undergoing the same experiment were mixedhoused. Intestinal tissue samples from age-matched Swiss Webster iso-genic germfree and “restricted flora” mice, originally from Taconic, wereprovided by Justin Sonnenburg (Stanford University, Stanford, CA).
ALP activity assay. One-centimeter segments of the proximal duode-num from mice were collected and cut open longitudinally. The lumenside was gently washed with phosphate-buffered saline (PBS). Cells on thelumen side were harvested by using cell scrapers, homogenized in 1 ml oflysis buffer (50 mM Tris-HCl[pH 7.6], 150 mM NaCl, 1 mM EDTA, 1 mMEGTA, 1% Triton X-100), and centrifuged at 17,900 � g for 15 min at 4°C.The cell-free supernatant was diluted in double-distilled H2O (ddH2O),and the protein concentration was measured using a Bio-Rad proteinassay kit (Bio-Rad Laboratories, Inc.). Mouse stools were collected,weighed, homogenized in 1 ml of ddH2O, and centrifuged at 17,900 � gfor 15 min at 4°C to collect the supernatant. Diluted supernatants pro-cessed from duodenal samples and stools were assayed for ALP activitiesusing a phosphate substrate kit (Thermo Fisher Scientific, Inc.). The ALPactivities were normalized to the protein concentration for duodenal sam-ples and to the mass for stools.
LPS administration. LPS (purified from Salmonella enterica serotypeTyphimurium [Sigma-Aldrich]) was dissolved in ddH2O by a 2-min son-ication. Age-matched female wild-type and Akp3�/� mice were deprivedof food for 3 h, and then LPS was introduced by gavage at indicated doses.Oral gavage was performed using sterile 20 G 1.5-in. feeding needles(curved, ball tipped; Popper & Sons) in 8- and 10-week-old mice andsterilized 24 G 1-in. needles (straight, ball tipped; Roboz Surgical Instru-ment) in 19-day-old and 4-week-old mice. Water gavage was performedon the control group. Mice were euthanized 3 h after gavage. One-centi-meter segments of the proximal duodenum from mice were collected, andthe transcription levels of ALP genes were examined by quantitative PCR(qPCR). Then, 2-cm segments of the ileum were collected for qPCR ex-amination of inflammatory markers.
Yersinia pseudotuberculosis infection. The wild-type Y. pseudotuber-culosis strain (YPIII) was provided by Joan Mecsas (Tufts University,Medford, MA). The infection was performed according to the protocoldescribed previously (34). Briefly, female wild-type and Akp3�/� mice (10weeks old) were fasted overnight. Infection was performed by gavage at2 � 109 CFU/animal. Uninfected animals were used as the control. Threeto five mice of the infected or control group were weighed daily for 5 days.To quantify bacterial colonization, infected mice were euthanized 24 hafter the oral inoculation, and Peyer’s patches along the small intestinewere collected, homogenized in sterile PBS, and plated on TSA platescontaining 2 �g of Irgasan (Sigma-Aldrich)/ml to determine the CFU/g of
Yang et al.
248 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

tissue. Two-centimeter segments of the ileum tissue from mice were col-lected for qPCR examination of the inflammatory markers.
qPCR. The total RNA was isolated from collected mouse intestinal tissuesusing a PureLink RNA minikit (Life Technologies, Grand Island, NY), andcDNA was synthesized using a SuperScript III first-strand synthesis system(Life Technologies). qPCR was performed on the StepOnePlus real-timePCR system (Life Technologies) using the Kapa SYBR Fast qPCR kit(Kapa Biosystems). Endogenous reference genes were tested, and 36B4,Gapdh, and Rpl13a were selected for qPCR normalization based on theGeNorm and the Normfinder algorithms using the GenEx software(Multid Analyses AB). Primers used in qPCR are listed in Table 1. Primeramplification efficiency was determined using the LinRegPCR softwareand incorporated to normalize the relative levels of gene expression toendogenous references using ��CT analysis.
HFD feeding. Male mice of indicated genotypes (�10 of each) fromthe same generation were fed a 36.0% fat diet (Bio-Serv, F3282/S3282)after weaning. HFD feeding started when mice were 3 weeks old andcontinued for 8 weeks. Mouse body weights were recorded weekly.
Neutrophil staining and quantification. Mouse ileum tissue was col-lected, embedded in agar, and cryosectioned at a thickness of 16 �m.Slides were washed in PBDT (1% bovine serum albumin, 1% dimethylsulfoxide, and 0.1% Triton X-100 in PBS) for 30 min, blocked in the blockbuffer (2% normal goat serum in PBDT), and incubated with rat anti-mouse Ly-6G primary antibody (number 550291; BD Pharmingen, SanJose, CA) at 4°C overnight. The next day, the slides were washed withPBST (0.1% Triton X-100 in PBS) and then incubated with biotin-conju-gated mouse anti-rat IgG2b antibody (number 550327; BD Pharmingen)at 4°C overnight. The slides were washed with PBST, incubated with AlexaFluor 555-streptavidin (catalog no. S21381; Life Technologies), and fi-nally mounted with Vectashield mounting medium with DAPI (4=,6=-diamidino-2-phenylindole; Vector Laboratories, Burlingame, CA). Thetotal number of Ly6G-positive cells was counted from five ileal cross sec-tions from each individual mouse and was used as an indicator of neutro-phil infiltration in the mouse.
Statistics. All data were analyzed using Prism software (GraphPad).The statistical analyses used for different data sets are indicated in thefigure legends.
RESULTSAkp3 is upregulated by the microbiota. To test whether Akp3expression was induced by the microbiota, as is the case for ze-
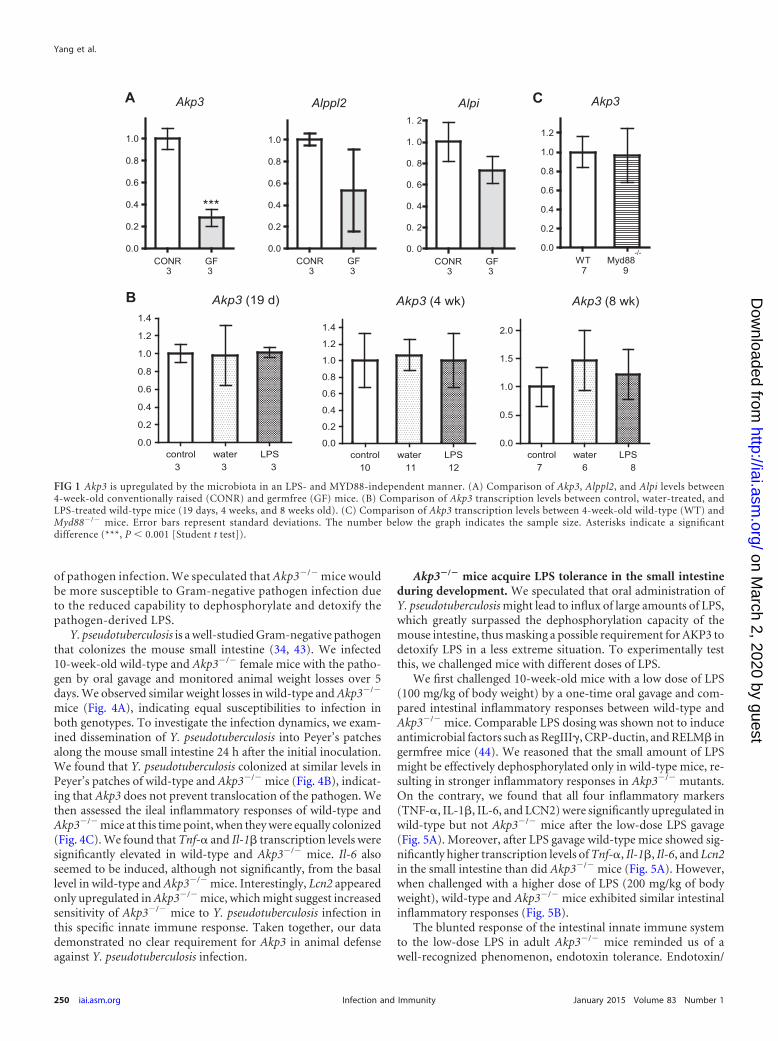
brafish alpi.1, we measured the transcription levels of ALP genes induodenal samples of age-matched, isogenic germfree and conven-tionally reared mice using qPCR. We found that germfree micehad reduced Akp3 transcription compared to conventional miceat 4 weeks of age (Fig. 1A). No significant differences in Alppl2 orAlpi transcription levels were observed between germfree andconventional mice. Consistent results were observed in two inde-pendent experiments (one shown in Fig. 1A), suggesting that themicrobiota specifically upregulate Akp3.
We next investigated whether MYD88-dependent LPS signal-ing induced Akp3, similar to the regulation of zebrafish alpi.1. Wetreated wild-type mice with water or LPS by oral gavage at varioustime points, concurrent with and after the normal onset of Akp3expression (at 19 days and at 4 and 8 weeks of age), and measuredAkp3 transcription levels. We observed no induction of Akp3 inthe mouse duodenum upon oral LPS administration at any ofthese time points (Fig. 1B). Furthermore, Myd88�/� mice andtheir wild-type littermates (4 weeks old) had comparable levels ofAkp3 transcription (Fig. 1C). We conclude that, whereas the mi-crobiota upregulate Akp3 in the mouse intestine, the mechanismis different from that in zebrafish.
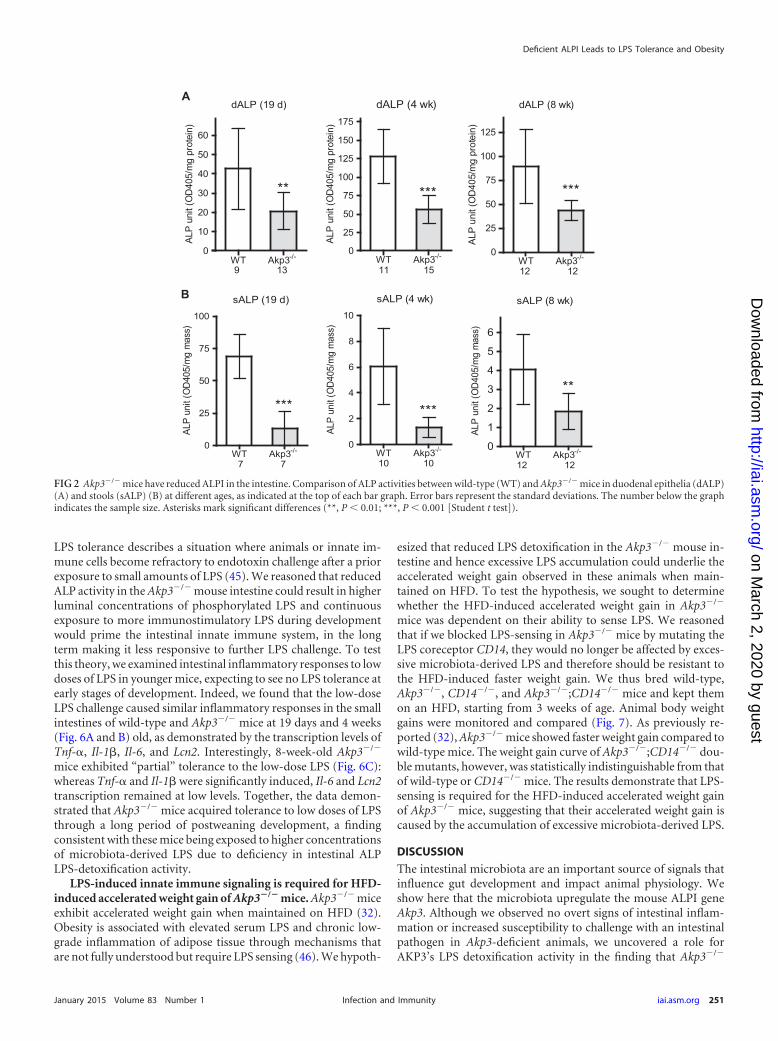
Basal inflammatory responses are normal in postweaningand adult Akp3�/� mice. We used the Akp3 loss-of-function miceto study the role of ALP in modulating intestinal inflammatoryresponses to resident microbiota during postweaning develop-ment. We first confirmed that ALP activities were significantlyreduced in the intestines of Akp3�/� mice versus wild-type miceafter the onset of Akp3 expression. We detected ALP deficiency inAkp3�/� mice, as measured by enzymatic assay on duodenal epi-thelial cells (Fig. 2A). ALP deficiency was also detected in stools ofAkp3�/� mice (Fig. 2B), indicating that the mutant mice havereduced ALP activity throughout the intestine.
We expected that ALP deficiency in the intestine of postwean-ing Akp3�/� mice would result in enhanced intestinal basal in-flammatory responses to excess microbiota-derived LPS. To assessinflammatory responses, we measured the transcription levels ofproinflammatory cytokines (tumor necrosis factor alpha [TNF-�], interleukin-1� [IL-1�], and IL-6) and the innate immune pro-tein LCN2 using qPCR and compared wild-type mice to Akp3�/�
mice. Although the duodenum is the site of Akp3 expression, it issparsely colonized by bacteria (42), and we detected only very lowlevels of cytokine transcripts in duodenal samples of both wild-type and Akp3�/� mice (data not shown). We hypothesized thatthe LPS-detoxifying function of AKP3 would be more importantin the distal intestine, which is more densely colonized and ex-posed to secreted ALP. We therefore examined basal inflamma-tory responses in the ileum. We detected comparable levels of thefour inflammatory markers in the ilea of postweaning wild-typeand Akp3�/� mice (19 days, 4 weeks, 8 weeks, or 10 weeks old)(Fig. 3). We also measured neutrophil infiltration in the ileum bycounting Ly6G-positive cells and observed no difference betweenwild-type and Akp3�/� mouse ileal tissues (data not shown). Wetherefore conclude that loss of AKP3 does not result in significantintestinal inflammation.
Akp3 is not required for defense against Yersinia pseudotu-berculosis infection. Although disruption of Akp3 has no obviouseffect on basal intestinal inflammatory responses in healthyanimals, we reasoned that the functional requirement for Akp3might be more pronounced when the equilibrium betweenhost and the gut microbiota is severely disturbed, as in the case
TABLE 1 Primers used in qPCR
Primer Sequence (5=–3=)36B4fwd TCCAGGCTTTGGGCATCA36B4rev CTTTATCAGCTGCACATCACTCAGAGapdhfwd GCACCACCAACTGCTTAGCGapdhrev GGCATGGACTGTGGTCATGAGRpl13afwd TTCTGGTATTGGATGGCCGAGRpl13arev ATGTTGATGCCTTCACAGCGAkp3fwd ACATTGCTACACAACTCATCTCCAkp3rev TCCTGCCATCCAATCTGGTTCAlppl2fwd AATGCCTGCCTCAGCGCTACAGGAAlppl2rev TGGTCTGGTGTCCCCTTGGGAAAlpifwd GGCTACACACTTAGGGGGACCTCCAAlpirev AGCTTCGGTGACATTGGGCCGGTTTnf�fwd AGGCTGCCCCGACTACGTTnf�rev GACTTTCTCCTGGTATGAGATAGCAAAIl1�fwd GAAATGCCACCTTTTGACAGTGIl1�rev TGGATGCTCTCATCAGGACAGIl6fwd TGGTACTCCAGAAGACCAGAGGIl6rev AACGATGATGCACTTGCAGALcn2fwd TGGCCCTGAGTGTCATGTGLcn2rev CTCTTGTAGCTCATAGATGGTGC
Deficient ALPI Leads to LPS Tolerance and Obesity
January 2015 Volume 83 Number 1 iai.asm.org 249Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
of pathogen infection. We speculated that Akp3�/� mice wouldbe more susceptible to Gram-negative pathogen infection dueto the reduced capability to dephosphorylate and detoxify thepathogen-derived LPS.
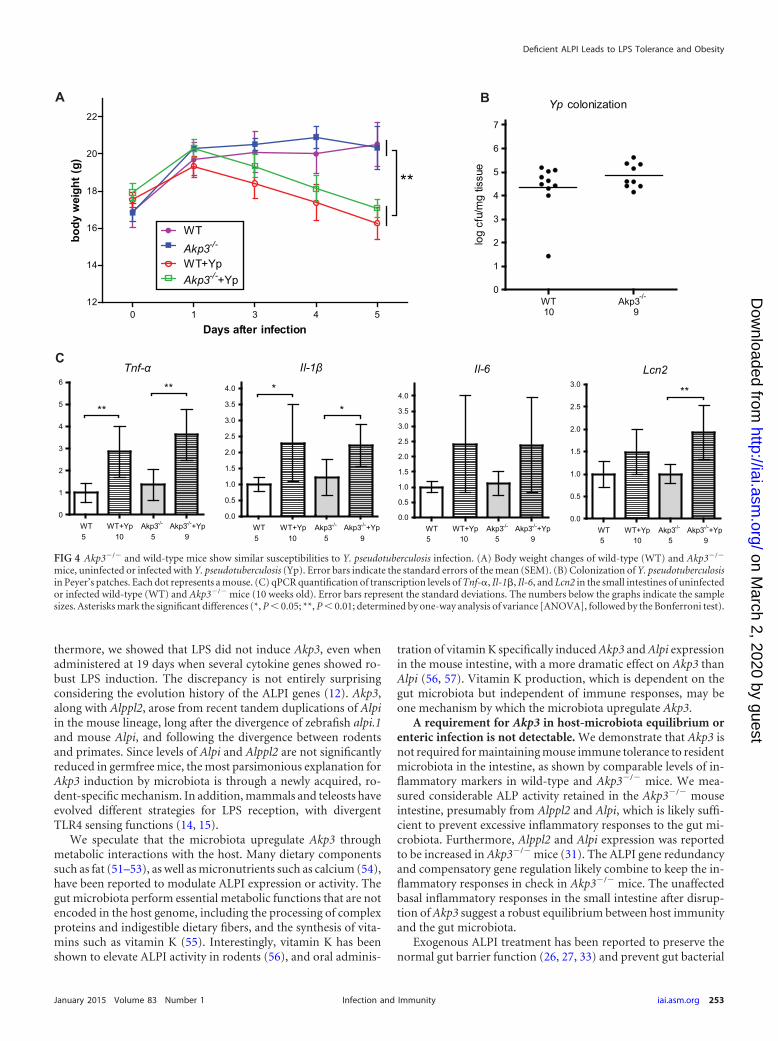
Y. pseudotuberculosis is a well-studied Gram-negative pathogenthat colonizes the mouse small intestine (34, 43). We infected10-week-old wild-type and Akp3�/� female mice with the patho-gen by oral gavage and monitored animal weight losses over 5days. We observed similar weight losses in wild-type and Akp3�/�
mice (Fig. 4A), indicating equal susceptibilities to infection inboth genotypes. To investigate the infection dynamics, we exam-ined dissemination of Y. pseudotuberculosis into Peyer’s patchesalong the mouse small intestine 24 h after the initial inoculation.We found that Y. pseudotuberculosis colonized at similar levels inPeyer’s patches of wild-type and Akp3�/� mice (Fig. 4B), indicat-ing that Akp3 does not prevent translocation of the pathogen. Wethen assessed the ileal inflammatory responses of wild-type andAkp3�/� mice at this time point, when they were equally colonized(Fig. 4C). We found that Tnf-� and Il-1� transcription levels weresignificantly elevated in wild-type and Akp3�/� mice. Il-6 alsoseemed to be induced, although not significantly, from the basallevel in wild-type and Akp3�/� mice. Interestingly, Lcn2 appearedonly upregulated in Akp3�/� mice, which might suggest increasedsensitivity of Akp3�/� mice to Y. pseudotuberculosis infection inthis specific innate immune response. Taken together, our datademonstrated no clear requirement for Akp3 in animal defenseagainst Y. pseudotuberculosis infection.
Akp3�/� mice acquire LPS tolerance in the small intestineduring development. We speculated that oral administration ofY. pseudotuberculosis might lead to influx of large amounts of LPS,which greatly surpassed the dephosphorylation capacity of themouse intestine, thus masking a possible requirement for AKP3 todetoxify LPS in a less extreme situation. To experimentally testthis, we challenged mice with different doses of LPS.
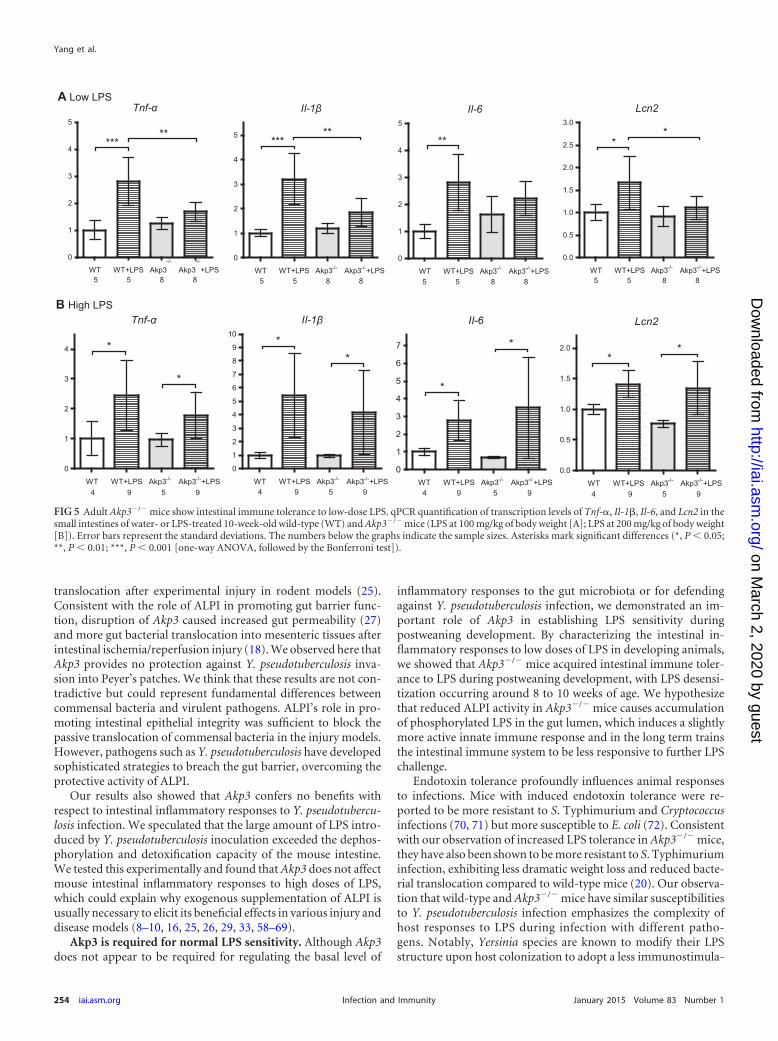
We first challenged 10-week-old mice with a low dose of LPS(100 mg/kg of body weight) by a one-time oral gavage and com-pared intestinal inflammatory responses between wild-type andAkp3�/� mice. Comparable LPS dosing was shown not to induceantimicrobial factors such as RegIII, CRP-ductin, and RELM� ingermfree mice (44). We reasoned that the small amount of LPSmight be effectively dephosphorylated only in wild-type mice, re-sulting in stronger inflammatory responses in Akp3�/� mutants.On the contrary, we found that all four inflammatory markers(TNF-�, IL-1�, IL-6, and LCN2) were significantly upregulated inwild-type but not Akp3�/� mice after the low-dose LPS gavage(Fig. 5A). Moreover, after LPS gavage wild-type mice showed sig-nificantly higher transcription levels of Tnf-�, Il-1�, Il-6, and Lcn2in the small intestine than did Akp3�/� mice (Fig. 5A). However,when challenged with a higher dose of LPS (200 mg/kg of bodyweight), wild-type and Akp3�/� mice exhibited similar intestinalinflammatory responses (Fig. 5B).
The blunted response of the intestinal innate immune systemto the low-dose LPS in adult Akp3�/� mice reminded us of awell-recognized phenomenon, endotoxin tolerance. Endotoxin/
A
B
CAkp3
CONR GF0.0
0.2
0.4
0.6
0.8
1.0
***
3 3
Alppl2
CONR GF0.0
0.2
0.4
0.6
0.8
1.0
3 3
Alpi
CONR GF0. 0
0. 2
0. 4
0. 6
0. 8
1. 0
1. 2
3 3
Akp3
WT Myd88-/- 0.0
0.2
0.4
0.6
0.8
1.0
1.2
7 9
Akp3 (4 wk)
control water LPS0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
10 11 12
Akp3 (8 wk)
control water LPS0.0
0.5
1.0
1.5
2.0
7 6 8
Akp3 (19 d)
control water LPS0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
3 3 3
FIG 1 Akp3 is upregulated by the microbiota in an LPS- and MYD88-independent manner. (A) Comparison of Akp3, Alppl2, and Alpi levels between4-week-old conventionally raised (CONR) and germfree (GF) mice. (B) Comparison of Akp3 transcription levels between control, water-treated, andLPS-treated wild-type mice (19 days, 4 weeks, and 8 weeks old). (C) Comparison of Akp3 transcription levels between 4-week-old wild-type (WT) andMyd88�/� mice. Error bars represent standard deviations. The number below the graph indicates the sample size. Asterisks indicate a significantdifference (***, P 0.001 [Student t test]).
Yang et al.
250 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
LPS tolerance describes a situation where animals or innate im-mune cells become refractory to endotoxin challenge after a priorexposure to small amounts of LPS (45). We reasoned that reducedALP activity in the Akp3�/� mouse intestine could result in higherluminal concentrations of phosphorylated LPS and continuousexposure to more immunostimulatory LPS during developmentwould prime the intestinal innate immune system, in the longterm making it less responsive to further LPS challenge. To testthis theory, we examined intestinal inflammatory responses to lowdoses of LPS in younger mice, expecting to see no LPS tolerance atearly stages of development. Indeed, we found that the low-doseLPS challenge caused similar inflammatory responses in the smallintestines of wild-type and Akp3�/� mice at 19 days and 4 weeks(Fig. 6A and B) old, as demonstrated by the transcription levels ofTnf-�, Il-1�, Il-6, and Lcn2. Interestingly, 8-week-old Akp3�/�
mice exhibited “partial” tolerance to the low-dose LPS (Fig. 6C):whereas Tnf-� and Il-1� were significantly induced, Il-6 and Lcn2transcription remained at low levels. Together, the data demon-strated that Akp3�/� mice acquired tolerance to low doses of LPSthrough a long period of postweaning development, a findingconsistent with these mice being exposed to higher concentrationsof microbiota-derived LPS due to deficiency in intestinal ALPLPS-detoxification activity.
LPS-induced innate immune signaling is required for HFD-induced accelerated weight gain of Akp3�/� mice. Akp3�/� miceexhibit accelerated weight gain when maintained on HFD (32).Obesity is associated with elevated serum LPS and chronic low-grade inflammation of adipose tissue through mechanisms thatare not fully understood but require LPS sensing (46). We hypoth-
esized that reduced LPS detoxification in the Akp3�/� mouse in-testine and hence excessive LPS accumulation could underlie theaccelerated weight gain observed in these animals when main-tained on HFD. To test the hypothesis, we sought to determinewhether the HFD-induced accelerated weight gain in Akp3�/�
mice was dependent on their ability to sense LPS. We reasonedthat if we blocked LPS-sensing in Akp3�/� mice by mutating theLPS coreceptor CD14, they would no longer be affected by exces-sive microbiota-derived LPS and therefore should be resistant tothe HFD-induced faster weight gain. We thus bred wild-type,Akp3�/�, CD14�/�, and Akp3�/�;CD14�/� mice and kept themon an HFD, starting from 3 weeks of age. Animal body weightgains were monitored and compared (Fig. 7). As previously re-ported (32), Akp3�/� mice showed faster weight gain compared towild-type mice. The weight gain curve of Akp3�/�;CD14�/� dou-ble mutants, however, was statistically indistinguishable from thatof wild-type or CD14�/� mice. The results demonstrate that LPS-sensing is required for the HFD-induced accelerated weight gainof Akp3�/� mice, suggesting that their accelerated weight gain iscaused by the accumulation of excessive microbiota-derived LPS.
DISCUSSION
The intestinal microbiota are an important source of signals thatinfluence gut development and impact animal physiology. Weshow here that the microbiota upregulate the mouse ALPI geneAkp3. Although we observed no overt signs of intestinal inflam-mation or increased susceptibility to challenge with an intestinalpathogen in Akp3-deficient animals, we uncovered a role forAKP3’s LPS detoxification activity in the finding that Akp3�/�
A
B
dALP (19 d)
WT Akp3-/-0
10
20
30
40
50
60
**
9 13
ALP
unit
(OD
405/
mg
prot
ein)
dALP (4 wk)
WT Akp3-/-0
25
50
75
100
125
150
175
***
11 15
ALP
unit
(OD
405/
mg
prot
ein)
dALP (8 wk)
WT Akp3-/-0
25
50
75
100
125
***
12 12
ALP
unit
(OD
405/
mg
prot
ein)
sALP (19 d)
WT Akp3-/-0
25
50
75
100
***
7 7
ALP
unit
(OD
405/
mg
mas
s)
sALP (4 wk)
WT Akp3-/-0
2
4
6
8
10
***
10 10
ALP
unit
(OD
405/
mg
mas
s)
sALP (8 wk)
WT Akp3-/-0
1
2
3
4
5
6
**
12 12AL
P un
it (O
D40
5/m
g m
ass)
FIG 2 Akp3�/� mice have reduced ALPI in the intestine. Comparison of ALP activities between wild-type (WT) and Akp3�/� mice in duodenal epithelia (dALP)(A) and stools (sALP) (B) at different ages, as indicated at the top of each bar graph. Error bars represent the standard deviations. The number below the graphindicates the sample size. Asterisks mark significant differences (**, P 0.01; ***, P 0.001 [Student t test]).
Deficient ALPI Leads to LPS Tolerance and Obesity
January 2015 Volume 83 Number 1 iai.asm.org 251Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

mice become desensitized to low doses of LPS during the first fewweeks after weaning. We further showed that this early subtle de-fect in LPS detoxification was related to the susceptibility of adultAkp3�/� mice to HFD-induced obesity, which was blocked with amutation in the LPS coreceptor CD14. These findings demon-strate how subtle alterations in host-microbiota interactions earlyin development can have long-lasting and systemic effects on an-imal physiology.
Akp3 is upregulated by the microbiota through a noncon-served mechanism. The intestinal microbiota are an importantsource of signals that regulate gene transcription in animal intes-tines (47–50). Consistent with the upregulation of Akp3 at thetime of weaning and establishment of the adult gut microbiota, weshow that Akp3 expression is reduced in germfree mouse intes-tines. In contrast with zebrafish alpi.1, we found that mouse Akp3is upregulated by the microbiota independent of MYD88. Fur-
Tnf-α Il-1β Il-6 Lcn2
19 d
4 wk
8 wk
10 wk
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3 50. 0
0. 2
0. 4
0. 6
0. 8
1. 0
1. 2
1. 4
1. 6
3 50.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
3 50.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
3 5
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
7 70.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
7 70.0
0.5
1.0
1.5
7 70.0
0.5
1.0
1.5
2.0
2.5
7 7
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
7 70.00.20.40.60.81.01.21.4
7 70.0
0.2
0.4
0.6
0.8
1.0
1.2
7 70.0
0.2
0.4
0.6
0.8
1.0
1.2
7 7
0.0
0.5
1.0
1.5
2.0
5 50.0
0.5
1.0
1.5
2.0
5 50.0
0.5
1.0
1.5
5 50.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
5 5
FIG 3 Akp3�/� mice show normal basal intestinal inflammatory responses. Comparison of ileal transcriptional levels of Tnf-�, Il-1�, Il-6, and Lcn2 betweenwild-type (white columns) and Akp3�/� (gray columns) mice at different ages. Error bars represent the standard deviations. The number below the graphindicates the sample size.
Yang et al.
252 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
thermore, we showed that LPS did not induce Akp3, even whenadministered at 19 days when several cytokine genes showed ro-bust LPS induction. The discrepancy is not entirely surprisingconsidering the evolution history of the ALPI genes (12). Akp3,along with Alppl2, arose from recent tandem duplications of Alpiin the mouse lineage, long after the divergence of zebrafish alpi.1and mouse Alpi, and following the divergence between rodentsand primates. Since levels of Alpi and Alppl2 are not significantlyreduced in germfree mice, the most parsimonious explanation forAkp3 induction by microbiota is through a newly acquired, ro-dent-specific mechanism. In addition, mammals and teleosts haveevolved different strategies for LPS reception, with divergentTLR4 sensing functions (14, 15).
We speculate that the microbiota upregulate Akp3 throughmetabolic interactions with the host. Many dietary componentssuch as fat (51–53), as well as micronutrients such as calcium (54),have been reported to modulate ALPI expression or activity. Thegut microbiota perform essential metabolic functions that are notencoded in the host genome, including the processing of complexproteins and indigestible dietary fibers, and the synthesis of vita-mins such as vitamin K (55). Interestingly, vitamin K has beenshown to elevate ALPI activity in rodents (56), and oral adminis-
tration of vitamin K specifically induced Akp3 and Alpi expressionin the mouse intestine, with a more dramatic effect on Akp3 thanAlpi (56, 57). Vitamin K production, which is dependent on thegut microbiota but independent of immune responses, may beone mechanism by which the microbiota upregulate Akp3.
A requirement for Akp3 in host-microbiota equilibrium orenteric infection is not detectable. We demonstrate that Akp3 isnot required for maintaining mouse immune tolerance to residentmicrobiota in the intestine, as shown by comparable levels of in-flammatory markers in wild-type and Akp3�/� mice. We mea-sured considerable ALP activity retained in the Akp3�/� mouseintestine, presumably from Alppl2 and Alpi, which is likely suffi-cient to prevent excessive inflammatory responses to the gut mi-crobiota. Furthermore, Alppl2 and Alpi expression was reportedto be increased in Akp3�/� mice (31). The ALPI gene redundancyand compensatory gene regulation likely combine to keep the in-flammatory responses in check in Akp3�/� mice. The unaffectedbasal inflammatory responses in the small intestine after disrup-tion of Akp3 suggest a robust equilibrium between host immunityand the gut microbiota.
Exogenous ALPI treatment has been reported to preserve thenormal gut barrier function (26, 27, 33) and prevent gut bacterial
A
C
Yp colonization
WT Akp3-/-0
1
2
3
4
5
6
7
10 9
log
cfu/
mg
tissu
e
Tnf-α
WT WT+Yp Akp3-/- Akp3-/-+Yp0
1
2
3
4
5
6
5 10 5 9
**
**
Il-1β
WT WT+Yp Akp3-/- Akp3-/-+Yp0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0 *
*
5 10 5 9
Il-6
WT WT+Yp Akp3-/- Akp3-/-+Yp0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
5 10 5 9
Lcn2
WT WT+Yp Akp3-/- Akp3-/-+Yp0.0
0.5
1.0
1.5
2.0
2.5
3.0
5 10 5 9
**
0 1 3 4 512
14
16
18
20
22
WTAkp3-/-
WT+YpAkp3-/-+Yp
**
Days after infection
body
wei
ght (
g)
B
FIG 4 Akp3�/� and wild-type mice show similar susceptibilities to Y. pseudotuberculosis infection. (A) Body weight changes of wild-type (WT) and Akp3�/�
mice, uninfected or infected with Y. pseudotuberculosis (Yp). Error bars indicate the standard errors of the mean (SEM). (B) Colonization of Y. pseudotuberculosisin Peyer’s patches. Each dot represents a mouse. (C) qPCR quantification of transcription levels of Tnf-�, Il-1�, Il-6, and Lcn2 in the small intestines of uninfectedor infected wild-type (WT) and Akp3�/� mice (10 weeks old). Error bars represent the standard deviations. The numbers below the graphs indicate the samplesizes. Asterisks mark the significant differences (*, P 0.05; **, P 0.01; determined by one-way analysis of variance [ANOVA], followed by the Bonferroni test).
Deficient ALPI Leads to LPS Tolerance and Obesity
January 2015 Volume 83 Number 1 iai.asm.org 253Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
translocation after experimental injury in rodent models (25).Consistent with the role of ALPI in promoting gut barrier func-tion, disruption of Akp3 caused increased gut permeability (27)and more gut bacterial translocation into mesenteric tissues afterintestinal ischemia/reperfusion injury (18). We observed here thatAkp3 provides no protection against Y. pseudotuberculosis inva-sion into Peyer’s patches. We think that these results are not con-tradictive but could represent fundamental differences betweencommensal bacteria and virulent pathogens. ALPI’s role in pro-moting intestinal epithelial integrity was sufficient to block thepassive translocation of commensal bacteria in the injury models.However, pathogens such as Y. pseudotuberculosis have developedsophisticated strategies to breach the gut barrier, overcoming theprotective activity of ALPI.
Our results also showed that Akp3 confers no benefits withrespect to intestinal inflammatory responses to Y. pseudotubercu-losis infection. We speculated that the large amount of LPS intro-duced by Y. pseudotuberculosis inoculation exceeded the dephos-phorylation and detoxification capacity of the mouse intestine.We tested this experimentally and found that Akp3 does not affectmouse intestinal inflammatory responses to high doses of LPS,which could explain why exogenous supplementation of ALPI isusually necessary to elicit its beneficial effects in various injury anddisease models (8–10, 16, 25, 26, 29, 33, 58–69).
Akp3 is required for normal LPS sensitivity. Although Akp3does not appear to be required for regulating the basal level of
inflammatory responses to the gut microbiota or for defendingagainst Y. pseudotuberculosis infection, we demonstrated an im-portant role of Akp3 in establishing LPS sensitivity duringpostweaning development. By characterizing the intestinal in-flammatory responses to low doses of LPS in developing animals,we showed that Akp3�/� mice acquired intestinal immune toler-ance to LPS during postweaning development, with LPS desensi-tization occurring around 8 to 10 weeks of age. We hypothesizethat reduced ALPI activity in Akp3�/� mice causes accumulationof phosphorylated LPS in the gut lumen, which induces a slightlymore active innate immune response and in the long term trainsthe intestinal immune system to be less responsive to further LPSchallenge.
Endotoxin tolerance profoundly influences animal responsesto infections. Mice with induced endotoxin tolerance were re-ported to be more resistant to S. Typhimurium and Cryptococcusinfections (70, 71) but more susceptible to E. coli (72). Consistentwith our observation of increased LPS tolerance in Akp3�/� mice,they have also been shown to be more resistant to S. Typhimuriuminfection, exhibiting less dramatic weight loss and reduced bacte-rial translocation compared to wild-type mice (20). Our observa-tion that wild-type and Akp3�/� mice have similar susceptibilitiesto Y. pseudotuberculosis infection emphasizes the complexity ofhost responses to LPS during infection with different patho-gens. Notably, Yersinia species are known to modify their LPSstructure upon host colonization to adopt a less immunostimula-
A Low LPS
B High LPS
Tnf-α
WT WT+LPS Akp3-/-
Akp3-/-
+LPS
0
1
2
3
4
5
5 5 8 8
*****
Il-1β
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
5 5 8 8
*****
Il-6
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
5 5 8 8
**
Lcn2
WT WT+LPS Akp3-/- Akp3-/-+LPS
0.0
0.5
1.0
1.5
2.0
2.5
3.0
**
5 5 8 8
Tnf-α
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
4 9 5 9
*
*
Il-1β
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
6
7
8
9
10*
*
4 9 5 9
Il-6
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
6
7 *
*
4 9 5 9
Lcn2
WT WT+LPS Akp3-/- Akp3-/-+LPS
0.0
0.5
1.0
1.5
2.0*
*
4 9 5 9
FIG 5 Adult Akp3�/� mice show intestinal immune tolerance to low-dose LPS. qPCR quantification of transcription levels of Tnf-�, Il-1�, Il-6, and Lcn2 in thesmall intestines of water- or LPS-treated 10-week-old wild-type (WT) and Akp3�/� mice (LPS at 100 mg/kg of body weight [A]; LPS at 200 mg/kg of body weight[B]). Error bars represent the standard deviations. The numbers below the graphs indicate the sample sizes. Asterisks mark significant differences (*, P 0.05;**, P 0.01; ***, P 0.001 [one-way ANOVA, followed by the Bonferroni test]).
Yang et al.
254 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

tory form, suggesting that these bacteria’s pathogenesis strategymay be better adapted to an LPS-tolerant host than S. Typhimu-rium (73).
Subtle dysregulation of LPS-induced innate immune signal-ing due to Akp3 deficiency contributes to HFD-induced obesity.ALPI activities are inversely related to the obesity proneness in ratsreceiving HFD (74, 75). Analogously, mice lacking Akp3 exhibitedaccelerated weight gain when maintained on HFD compared towild-type mice (32). A direct role of the mouse gene Alpi in pro-moting fatty acid intake was proposed, and upregulation of Alpiin Akp3�/� mice could therefore contribute to the HFD-inducedfaster weight gain (31, 76, 77). Akp3�/� mice have also beenshown to have elevated serum LPS levels, presumably due to theirimpaired LPS dephosphorylation in the intestine and increased
gut permeability (27). We hypothesized that elevated basal levelsof LPS in Akp3�/� mice result in a physiological state in which theinnate immune system no longer responses acutely to low-levelLPS challenge but rather the tissues exist in a state of chronic,low-grade inflammation that promotes weight gain (46). Here, wetested the specific contribution of LPS-induced innate immunesignaling to HFD-induced accelerated weight gain in Akp3�/�
mice and found that by blocking LPS sensing with CD14 defi-ciency we abrogated the faster weight gain phenotype in thesemice. This provides evidence that LPS-mediated innate immunesignaling is required for the HFD-induced phenotype of Akp3�/�
mice. Elevated, LPS-dependent, chronic, low-grade inflammationsignaling might also underlie the inverse relationship betweenALPI activities and the obesity proneness of rats receiving HFD. It
A 19 d
B 4 wk
C 8 wk
Tnf-α
WT WT+LPS Akp3-/- Akp3-/-+LPS0
1
2
3
4
5
6
7
8
9
10
3 3 5 5
* *
Il-1β
WT WT+LPS Akp3-/- Akp3-/-+LPS0
5
10
15
20
25
3 3 5 5
Il-6
WT WT+LPS Akp3-/- Akp3-/-+LPS0
5
10
15
20
25
3 3 5 5
Lcn2
WT WT+LPS Akp3-/- Akp3-/-+LPS0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
3 3 5 5
Tnf-α
WT WT+LPS Akp3-/- Akp3-/-+LPS
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
7 7 7 7
**
Il-1β
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
7 7 7 7
*
*
Il-6
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
10
20
30
40
50
60
7 7 7 7
Lcn2
WT WT+LPS Akp3-/- Akp3-/-+LPS
0
1
2
3
4
5
6
7 7 7 7
*
Tnf-α
WT WT+LPS Akp3-/- Akp3-/-+LPS0.0
0.5
1.0
1.5
2.0
2.5
3.0
**
6 8 6 7
Il-1β
WT WT+LPS Akp3-/- Akp3-/-+LPS0
1
2
3
4
5
6
7 **
*
6 8 6 7
Il-6
WT WT+LPS Akp3-/- Akp3-/-+LPS02468
10121416182022
6 8 6 7
***
Lcn2
WT WT+LPS Akp3-/- Akp3-/-+LPS0
2
4
6
8
10
12
14
6 8 6 7
*
FIG 6 Akp3�/� mice acquire endotoxin tolerance during postweaning development. qPCR quantification of transcription levels of Tnf-�, Il-1�, Il-6, and Lcn2in the small intestines of wild-type (WT) and Akp3�/� mice, treated with water or LPS (100 mg/kg of body weight) at 19 days (A), 4 weeks (B), or 8 weeks (C)of age. Error bars represent the standard deviations. The numbers below the graphs indicate the sample sizes. Asterisks mark the significant difference (*, P 0.05;**, P 0.01 [one-way ANOVA, followed by the Bonferroni test]).
Deficient ALPI Leads to LPS Tolerance and Obesity
January 2015 Volume 83 Number 1 iai.asm.org 255Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

would be interesting to evaluate the contribution of LPS sensing inthese models.
In conclusion, we have discovered that the mouse ALPI geneAkp3 is specifically upregulated by the microbiota in an LPS- andMYD88-independent manner. We have showed that disruptionof Akp3 leads to immune desensitization to LPS during postwean-ing development, demonstrating a requirement for Akp3’s LPSdetoxifying activity in immune education. Finally, we have linkedthese earlier subtle defects in LPS detoxification to later onset ofHFD-induced obesity in Akp3�/� mice by showing that the LPS-sensing machinery is required for the accelerated weight gain inAkp3�/� mice maintained on HFD. Collectively, these studiesshow that subtle dysregulation of host-microbiota interactions inthe gut can have long-term systemic effects on host immunity andphysiology.
ACKNOWLEDGMENTS
We thank Jennifer Bates for her contributions to this project in its earlystages, Lora Hooper for valuable early assistance, and Justin Sonnenbergfor providing germfree mouse tissue samples. We especially thank the staffof the University of Oregon Mouse Facility for animal management andPoh Kheng Loi and the staff of the University of Oregon histology facilityfor histology services.
The project described was supported by awards R01DK075549 fromthe National Institute of Diabetes and Digestive and Kidney Diseases andaward P50GM098911 from the National Institute of General Medical Sci-ences of the National Institutes of Health.
The content is solely the responsibility of the authors and does notnecessarily represent the official views of the National Institutes of Health.
We have no conflicting financial interests.
REFERENCES1. Tremaroli V, Backhed F. 2012. Functional interactions between the gut
microbiota and host metabolism. Nature 489:242–249. http://dx.doi.org/10.1038/nature11552.
2. Belkaid Y, Hand TW. 2014. Role of the microbiota in immunity andinflammation. Cell 157:121–141. http://dx.doi.org/10.1016/j.cell.2014.03.011.
3. Goto Y, Kiyono H. 2012. Epithelial barrier: an interface for the cross-communication between gut flora and immune system. Immunol Rev245:147–163. http://dx.doi.org/10.1111/j.1600-065X.2011.01078.x.
4. Kamada N, Chen GY, Inohara N, Nunez G. 2013. Control of pathogensand pathobionts by the gut microbiota. Nat Immunol 14:685– 690. http://dx.doi.org/10.1038/ni.2608.
5. McComb RB, Bowers GN, Jr, Posen S. 1979. Alkaline phosphatase.Plenum Press, Inc, New York, NY.
6. Millán JL. 2006. Mammalian alkaline phosphatases: from biology to ap-plications in medicine and biotechnology. Wiley-VCH, Weinheim, Ger-many.
7. Bentala H, Verweij WR, Huizinga-Van der Vlag A, van Loenen-Weemaes AM, Meijer DK, Poelstra K. 2002. Removal of phosphate fromlipid A as a strategy to detoxify lipopolysaccharide. Shock 18:561–566.http://dx.doi.org/10.1097/00024382-200212000-00013.
8. Riggle KM, Rentea RM, Welak SR, Pritchard KA, Jr, Oldham KT,Gourlay DM. 2013. Intestinal alkaline phosphatase prevents the systemicinflammatory response associated with necrotizing enterocolitis. J SurgRes 180:21–26. http://dx.doi.org/10.1016/j.jss.2012.10.042.
9. Tuin A, Poelstra K, de Jager-Krikken A, Bok L, Raaben W, Velders MP,Dijkstra G. 2009. Role of alkaline phosphatase in colitis in man and rats.Gut 58:379 –387. http://dx.doi.org/10.1136/gut.2007.128868.
10. van Veen SQ, Dinant S, van Vliet AK, van Gulik TM. 2006. Alkalinephosphatase reduces hepatic and pulmonary injury in liver ischaemia:reperfusion combined with partial resection. Br J Surg 93:448 – 456. http://dx.doi.org/10.1002/bjs.5275.
11. Elin RJ, Wolff SM. 1976. Biology of endotoxin. Annu Rev Med 27:127–141. http://dx.doi.org/10.1146/annurev.me.27.020176.001015.
12. Yang Y, Wandler AM, Postlethwait JH, Guillemin K. 2012. Dynamicevolution of the LPS-detoxifying enzyme intestinal alkaline phosphatasein zebrafish and other vertebrates. Front Immunol 3:314. http://dx.doi.org/10.3389/fimmu.2012.00314.
13. Park BS, Lee JO. 2013. Recognition of lipopolysaccharide pattern byTLR4 complexes. Exp Mol Med 45:e66. http://dx.doi.org/10.1038/emm.2013.97.
14. Sullivan C, Charette J, Catchen J, Lage CR, Giasson G, Postlethwait JH,Millard PJ, Kim CH. 2009. The gene history of zebrafish tlr4a and tlr4b ispredictive of their divergent functions. J Immunol 183:5896 –5908. http://dx.doi.org/10.4049/jimmunol.0803285.
15. Sepulcre MP, Alcaraz-Perez F, Lopez-Munoz A, Roca FJ, Meseguer J,Cayuela ML, Mulero V. 2009. Evolution of lipopolysaccharide (LPS)recognition and signaling: fish TLR4 does not recognize LPS and nega-tively regulates NF-�B activation. J Immunol 182:1836 –1845. http://dx.doi.org/10.4049/jimmunol.0801755.
16. Bates JM, Akerlund J, Mittge E, Guillemin K. 2007. Intestinal alkalinephosphatase detoxifies lipopolysaccharide and prevents inflammation inzebrafish in response to the gut microbiota. Cell Host Microbe 2:371–382.http://dx.doi.org/10.1016/j.chom.2007.10.010.
17. Rader BA, Kremer N, Apicella MA, Goldman WE, McFall-Ngai MJ.2012. Modulation of symbiont lipid A signaling by host alkaline phospha-tases in the squid-vibrio symbiosis. mBio 3:e00093-12. http://dx.doi.org/10.1128/mBio.00093-12.
18. Goldberg RF, Austen WG, Jr, Zhang X, Munene G, Mostafa G,Biswas S, McCormack M, Eberlin KR, Nguyen JT, Tatlidede HS,Warren HS, Narisawa S, Millan JL, Hodin RA. 2008. Intestinalalkaline phosphatase is a gut mucosal defense factor maintained byenteral nutrition. Proc Natl Acad Sci U S A 105:3551–3556. http://dx.doi.org/10.1073/pnas.0712140105.
19. Chen KT, Malo MS, Moss AK, Zeller S, Johnson P, Ebrahimi F, MostafaG, Alam SN, Ramasamy S, Warren HS, Hohmann EL, Hodin RA. 2010.Identification of specific targets for the gut mucosal defense factor intes-tinal alkaline phosphatase. Am J Physiol Gastrointest Liver Physiol 299:G467–G475. http://dx.doi.org/10.1152/ajpgi.00364.2009.
20. Chen KT, Malo MS, Beasley-Topliffe LK, Poelstra K, Millan JL, MostafaG, Alam SN, Ramasamy S, Warren HS, Hohmann EL, Hodin RA. 2011.A role for intestinal alkaline phosphatase in the maintenance of local gutimmunity. Dig Dis Sci 56:1020 –1027. http://dx.doi.org/10.1007/s10620-010-1396-x.
21. Hanna SD, Mircheff AK, Wright EM. 1979. Alkaline phosphatase ofbasal lateral and brush border plasma membranes from intestinal ep-ithelium. J Supramol Struct 11:451– 466. http://dx.doi.org/10.1002/jss.400110404.
22. McConnell RE, Higginbotham JN, Shifrin DA, Jr, Tabb DL, Coffey RJ,Tyska MJ. 2009. The enterocyte microvillus is a vesicle-generating organ-elle. J Cell Biol 185:1285–1298. http://dx.doi.org/10.1083/jcb.200902147.
23. Shifrin DA, Jr, McConnell RE, Nambiar R, Higginbotham JN, CoffeyRJ, Tyska MJ. 2012. Enterocyte microvillus-derived vesicles detoxify bac-terial products and regulate epithelial-microbial interactions. Curr Biol22:627– 631. http://dx.doi.org/10.1016/j.cub.2012.02.022.
1 2 3 4 5 6 7 810
15
20
25
30
35
40
WT
Akp3-/-
CD14-/-
Akp3-/-;CD14-/-
***
weeks on high fat diet
body
wei
ght (
g)
FIG 7 LPS sensing is required for HFD-induced obesity in Akp3�/� mice.Body weight changes of wild-type (WT), Akp3�/�, CD14�/�, and Akp3�/�;CD14�/� mice fed an HFD were determined. Error bars represent the SEM.Asterisks indicate a significant difference (***, P 0.001 [two-way ANOVA]).
Yang et al.
256 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

24. Mizumori M, Ham M, Guth PH, Engel E, Kaunitz JD, Akiba Y. 2009.Intestinal alkaline phosphatase regulates protective surface microclimatepH in rat duodenum. J Physiol 587:3651–3663. http://dx.doi.org/10.1113/jphysiol.2009.172270.
25. Martinez-Moya P, Ortega-Gonzalez M, Gonzalez R, Anzola A, Ocon B,Hernandez-Chirlaque C, Lopez-Posadas R, Suarez MD, Zarzuelo A,Martinez-Augustin O, Sanchez de Medina F. 2012. Exogenous alkalinephosphatase treatment complements endogenous enzyme protection incolonic inflammation and reduces bacterial translocation in rats. Pharma-col Res 66:144 –153. http://dx.doi.org/10.1016/j.phrs.2012.04.006.
26. Rentea RM, Liedel JL, Welak SR, Cassidy LD, Mayer AN, Pritchard KA,Jr, Oldham KT, Gourlay DM. 2012. Intestinal alkaline phosphatase ad-ministration in newborns is protective of gut barrier function in a neona-tal necrotizing enterocolitis rat model. J Pediatr Surg 47:1135–1142. http://dx.doi.org/10.1016/j.jpedsurg.2012.03.018.
27. Kaliannan K, Hamarneh SR, Economopoulos KP, Nasrin Alam S,Moaven O, Patel P, Malo NS, Ray M, Abtahi SM, Muhammad N,Raychowdhury A, Teshager A, Mohamed MM, Moss AK, Ahmed R,Hakimian S, Narisawa S, Millan JL, Hohmann E, Warren HS, Bhan AK,Malo MS, Hodin RA. 2013. Intestinal alkaline phosphatase preventsmetabolic syndrome in mice. Proc Natl Acad Sci U S A 110:7003–7008.http://dx.doi.org/10.1073/pnas.1220180110.
28. Malo MS, Alam SN, Mostafa G, Zeller SJ, Johnson PV, Mohammad N,Chen KT, Moss AK, Ramasamy S, Faruqui A, Hodin S, Malo PS,Ebrahimi F, Biswas B, Narisawa S, Millan JL, Warren HS, Kaplan JB,Kitts CL, Hohmann EL, Hodin RA. 2010. Intestinal alkaline phosphatasepreserves the normal homeostasis of gut microbiota. Gut 59:1476 –1484.http://dx.doi.org/10.1136/gut.2010.211706.
29. Malo MS, Moaven O, Muhammad N, Biswas B, Alam SN, Economo-poulos KP, Gul SS, Hamarneh SR, Malo NS, Teshager A, MohamedMM, Tao Q, Narisawa S, Millan JL, Hohmann EL, Warren HS, RobsonSC, Hodin RA. 2014. Intestinal alkaline phosphatase promotes gut bac-terial growth by reducing the concentration of luminal nucleotide triphos-phates. Am J Physiol Gastrointest Liver Physiol 306:G826 –G838. http://dx.doi.org/10.1152/ajpgi.00357.2013.
30. Bates JM, Mittge E, Kuhlman J, Baden KN, Cheesman SE, Guillemin K.2006. Distinct signals from the microbiota promote different aspects ofzebrafish gut differentiation. Dev Biol 297:374 –386. http://dx.doi.org/10.1016/j.ydbio.2006.05.006.
31. Narisawa S, Hoylaerts MF, Doctor KS, Fukuda MN, Alpers DH, MillanJL. 2007. A novel phosphatase upregulated in Akp3 knockout mice. Am JPhysiol Gastrointest Liver Physiol 293:G1068 –G1077. http://dx.doi.org/10.1152/ajpgi.00073.2007.
32. Narisawa S, Huang L, Iwasaki A, Hasegawa H, Alpers DH, Millan JL.2003. Accelerated fat absorption in intestinal alkaline phosphatase knock-out mice. Mol Cell Biol 23:7525–7530. http://dx.doi.org/10.1128/MCB.23.21.7525-7530.2003.
33. Ramasamy S, Nguyen DD, Eston MA, Alam SN, Moss AK, Ebrahimi F,Biswas B, Mostafa G, Chen KT, Kaliannan K, Yammine H, Narisawa S,Millan JL, Warren HS, Hohmann EL, Mizoguchi E, Reinecker HC,Bhan AK, Snapper SB, Malo MS, Hodin RA. 2011. Intestinal alkalinephosphatase has beneficial effects in mouse models of chronic colitis. In-flamm Bowel Dis 17:532–542. http://dx.doi.org/10.1002/ibd.21377.
34. Logsdon LK, Mecsas J. 2003. Requirement of the Yersinia pseudotuber-culosis effectors YopH and YopE in colonization and persistence in intes-tinal and lymph tissues. Infect Immun 71:4595– 4607. http://dx.doi.org/10.1128/IAI.71.8.4595-4607.2003.
35. Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC,Chamontin B, Ferrieres J. 2008. Energy intake is associated with endo-toxemia in apparently healthy men. Am J Clin Nutr 87:1219 –1223.
36. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, NeyrinckAM, Fava F, Tuohy KM, Chabo C, Waget A, Delmee E, Cousin B,Sulpice T, Chamontin B, Ferrieres J, Tanti JF, Gibson GR, Casteilla L,Delzenne NM, Alessi MC, Burcelin R. 2007. Metabolic endotoxemiainitiates obesity and insulin resistance. Diabetes 56:1761–1772. http://dx.doi.org/10.2337/db06-1491.
37. Erridge C, Attina T, Spickett CM, Webb DJ. 2007. A high-fat mealinduces low-grade endotoxemia: evidence of a novel mechanism of post-prandial inflammation. Am J Clin Nutr 86:1286 –1292.
38. Ghanim H, Abuaysheh S, Sia CL, Korzeniewski K, Chaudhuri A,Fernandez-Real JM, Dandona P. 2009. Increase in plasma endotoxinconcentrations and the expression of Toll-like receptors and suppressor ofcytokine signaling-3 in mononuclear cells after a high-fat, high-
carbohydrate meal: implications for insulin resistance. Diabetes Care 32:2281–2287. http://dx.doi.org/10.2337/dc09-0979.
39. Davis JE, Gabler NK, Walker-Daniels J, Spurlock ME. 2008. Tlr-4deficiency selectively protects against obesity induced by diets high insaturated fat. Obesity 16:1248 –1255. http://dx.doi.org/10.1038/oby.2008.210.
40. Radin MS, Sinha S, Bhatt BA, Dedousis N, O’Doherty RM. 2008.Inhibition or deletion of the lipopolysaccharide receptor Toll-like recep-tor-4 confers partial protection against lipid-induced insulin resistance inrodent skeletal muscle. Diabetologia 51:336 –346. http://dx.doi.org/10.1007/s00125-007-0861-3.
41. Tsukumo DM, Carvalho-Filho MA, Carvalheira JB, Prada PO, Hi-rabara SM, Schenka AA, Araujo EP, Vassallo J, Curi R, Velloso LA, SaadMJ. 2007. Loss-of-function mutation in Toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Diabetes 56:1986 –1998. http://dx.doi.org/10.2337/db06-1595.
42. Uhlig HH, Powrie F. 2003. Dendritic cells and the intestinal bacterialflora: a role for localized mucosal immune responses. J Clin Invest 112:648 – 651. http://dx.doi.org/10.1172/JCI200319545.
43. Autenrieth IB, Firsching R. 1996. Penetration of M cells and destructionof Peyer’s patches by Yersinia enterocolitica: an ultrastructural and histo-logical study. J Med Microbiol 44:285–294. http://dx.doi.org/10.1099/00222615-44-4-285.
44. Vaishnava S, Behrendt CL, Ismail AS, Eckmann L, Hooper LV. 2008.Paneth cells directly sense gut commensals and maintain homeostasis atthe intestinal host-microbial interface. Proc Natl Acad Sci U S A 105:20858 –20863. http://dx.doi.org/10.1073/pnas.0808723105.
45. Fan H, Cook JA. 2004. Molecular mechanisms of endotoxin tolerance.J Endotoxin Res 10:71– 84. http://dx.doi.org/10.1177/09680519040100020301.
46. Cani PD, Osto M, Geurts L, Everard A. 2012. Involvement of gutmicrobiota in the development of low-grade inflammation and type 2diabetes associated with obesity. Gut Microbes 3:279 –288. http://dx.doi.org/10.4161/gmic.19625.
47. Larsson E, Tremaroli V, Lee YS, Koren O, Nookaew I, Fricker A,Nielsen J, Ley RE, Backhed F. 2012. Analysis of gut microbial regulationof host gene expression along the length of the gut and regulation of gutmicrobial ecology through MyD88. Gut 61:1124 –1131. http://dx.doi.org/10.1136/gutjnl-2011-301104.
48. Mutch DM, Simmering R, Donnicola D, Fotopoulos G, Holzwarth JA,Williamson G, Corthesy-Theulaz I. 2004. Impact of commensal micro-biota on murine gastrointestinal tract gene ontologies. Physiol Genomics19:22–31. http://dx.doi.org/10.1152/physiolgenomics.00105.2004.
49. Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI.2001. Molecular analysis of commensal host-microbial relationships inthe intestine. Science 291:881– 884. http://dx.doi.org/10.1126/science.291.5505.881.
50. Kanther M, Sun X, Muhlbauer M, Mackey LC, Flynn EJ, III, Bagnat M,Jobin C, Rawls JF. 2011. Microbial colonization induces dynamic tem-poral and spatial patterns of NF-�B activation in the zebrafish digestivetract. Gastroenterology 141:197–207. http://dx.doi.org/10.1053/j.gastro.2011.03.042.
51. Eliakim R, Mahmood A, Alpers DH. 1991. Rat intestinal alkalinephosphatase secretion into lumen and serum is coordinately regulated.Biochim Biophys Acta 1091:1– 8. http://dx.doi.org/10.1016/0167-4889(91)90213-H.
52. Kaur J, Madan S, Hamid A, Singla A, Mahmood A. 2007. Intestinalalkaline phosphatase secretion in oil-fed rats. Dig Dis Sci 52:665– 670.http://dx.doi.org/10.1007/s10620-006-9384-x.
53. Vazquez CM, Zanetti R, Santa-Maria C, Ruiz-Gutierrez V. 2000. Effectsof two highly monounsaturated oils on lipid composition and enzymeactivities in rat jejunum. Biosci Reports 20:355–368. http://dx.doi.org/10.1023/A:1010377900745.
54. Brun LR, Brance ML, Rigalli A. 2012. Luminal calcium concentrationcontrols intestinal calcium absorption by modification of intestinal alka-line phosphatase activity. Br J Nutr 108:229 –233. http://dx.doi.org/10.1017/S0007114511005617.
55. Resta SC. 2009. Effects of probiotics and commensals on intestinal epi-thelial physiology: implications for nutrient handling. J Physiol 587:4169 –4174. http://dx.doi.org/10.1113/jphysiol.2009.176370.
56. Sogabe N, Maruyama R, Hosori T, Goseki-Sone M. 2007. Enhancementeffects of vitamin K1 (phylloquinone) or vitamin K2 (menaquinone-4) on
Deficient ALPI Leads to LPS Tolerance and Obesity
January 2015 Volume 83 Number 1 iai.asm.org 257Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

intestinal alkaline phosphatase activity in rats. J Nutr Sci Vitaminol 53:219 –224. http://dx.doi.org/10.3177/jnsv.53.219.
57. Haraikawa M, Sogabe N, Tanabe R, Hosoi T, Goseki-Sone M. 2011.Vitamin K1 (phylloquinone) or vitamin K2 (menaquinone-4) inducesintestinal alkaline phosphatase gene expression. J Nutr Sci Vitaminol 57:274 –279. http://dx.doi.org/10.3177/jnsv.57.274.
58. Bol-Schoenmakers M, Fiechter D, Raaben W, Hassing I, Bleumink R,Kruijswijk D, Maijoor K, Tersteeg-Zijderveld M, Brands R, Pieters R.2010. Intestinal alkaline phosphatase contributes to the reduction of se-vere intestinal epithelial damage. Eur J Pharmacol 633:71–77. http://dx.doi.org/10.1016/j.ejphar.2010.01.023.
59. Heemskerk S, Masereeuw R, Moesker O, Bouw MP, van der Hoeven JG,Peters WH, Russel FG, Pickkers P. 2009. Alkaline phosphatase treatmentimproves renal function in severe sepsis or septic shock patients. Crit CareMed 37:417– 423. http://dx.doi.org/10.1097/CCM.0b013e31819598af.
60. Heinzerling NP, Liedel JL, Welak SR, Fredrich K, Biesterveld BE,Pritchard KA, Jr, Gourlay DM. 2014. Intestinal alkaline phosphatase isprotective to the preterm rat pup intestine. J Pediatr Surg 49:954 –960.http://dx.doi.org/10.1016/j.jpedsurg.2014.01.031.
61. Kats S, Brands R, Seinen W, de Jager W, Bekker MW, Hamad MA, TanME, Schonberger JP. 2009. Anti-inflammatory effects of alkaline phos-phatase in coronary artery bypass surgery with cardiopulmonary bypass.Recent Patents Inflamm Allergy Drug Discov 3:214 –220. http://dx.doi.org/10.2174/187221309789257388.
62. Lukas M, Drastich P, Konecny M, Gionchetti P, Urban O, Cantoni F,Bortlik M, Duricova D, Bulitta M. 2010. Exogenous alkaline phosphatasefor the treatment of patients with moderate to severe ulcerative colitis.Inflamm Bowel Dis 16:1180 –1186. http://dx.doi.org/10.1002/ibd.21161.
63. Pickkers P, Heemskerk S, Schouten J, Laterre PF, Vincent JL, Beishui-zen A, Jorens PG, Spapen H, Bulitta M, Peters WH, van der Hoeven JG.2012. Alkaline phosphatase for treatment of sepsis-induced acute kidneyinjury: a prospective randomized double-blind placebo-controlled trial.Crit Care 16:R14. http://dx.doi.org/10.1186/cc11159.
64. Rentea RM, Liedel JL, Fredrich K, Pritchard K, Jr, Oldham KT, Simp-son PM, Gourlay DM. 2013. Enteral intestinal alkaline phosphatase ad-ministration in newborns decreases iNOS expression in a neonatal necro-tizing enterocolitis rat model. J Pediatr Surg 48:124 –128. http://dx.doi.org/10.1016/j.jpedsurg.2012.10.026.
65. Rentea RM, Liedel JL, Fredrich K, Welak SR, Pritchard KA, Jr, OldhamKT, Simpson PM, Gourlay DM. 2012. Intestinal alkaline phosphataseadministration in newborns decreases systemic inflammatory cytokineexpression in a neonatal necrotizing enterocolitis rat model. J Surg Res177:228 –234. http://dx.doi.org/10.1016/j.jss.2012.05.039.
66. van Veen SQ, van Vliet AK, Wulferink M, Brands R, Boermeester MA,van Gulik TM. 2005. Bovine intestinal alkaline phosphatase attenuatesthe inflammatory response in secondary peritonitis in mice. Infect Immun73:4309 – 4314. http://dx.doi.org/10.1128/IAI.73.7.4309-4314.2005.
67. Verweij WR, Bentala H, Huizinga-van der Vlag A, Miek van Loenen-
Weemaes A, Kooi K, Meijer DK, Poelstra K. 2004. Protection against anEscherichia coli-induced sepsis by alkaline phosphatase in mice. Shock22:174 –179. http://dx.doi.org/10.1097/01.shk.0000132485.05049.8a.
68. Whitehouse JS, Riggle KM, Purpi DP, Mayer AN, Pritchard KA, Jr,Oldham KT, Gourlay DM. 2010. The protective role of intestinal alkalinephosphatase in necrotizing enterocolitis. J Surg Res 163:79 – 85. http://dx.doi.org/10.1016/j.jss.2010.04.048.
69. Alam SN, Yammine H, Moaven O, Ahmed R, Moss AK, Biswas B,Muhammad N, Biswas R, Raychowdhury A, Kaliannan K, Ghosh S, RayM, Hamarneh SR, Barua S, Malo NS, Bhan AK, Malo MS, Hodin RA.2014. Intestinal alkaline phosphatase prevents antibiotic-induced suscep-tibility to enteric pathogens. Ann Surg 259:715–722. http://dx.doi.org/10.1097/SLA.0b013e31828fae14.
70. Lehner MD, Ittner J, Bundschuh DS, van Rooijen N, Wendel A,Hartung T. 2001. Improved innate immunity of endotoxin-tolerant miceincreases resistance to Salmonella enterica serovar Typhimurium infectiondespite attenuated cytokine response. Infect Immun 69:463– 471. http://dx.doi.org/10.1128/IAI.69.1.463-471.2001.
71. Rayhane N, Fitting C, Lortholary O, Dromer F, Cavaillon JM. 2000.Administration of endotoxin associated with lipopolysaccharide toleranceprotects mice against fungal infection. Infect Immun 68:3748 –3753. http://dx.doi.org/10.1128/IAI.68.6.3748-3753.2000.
72. Lu M, Varley AW, Ohta S, Hardwick J, Munford RS. 2008. Hostinactivation of bacterial lipopolysaccharide prevents prolonged tolerancefollowing gram-negative bacterial infection. Cell Host Microbe 4:293–302. http://dx.doi.org/10.1016/j.chom.2008.06.009.
73. Rebeil R, Ernst RK, Gowen BB, Miller SI, Hinnebusch BJ. 2004. Vari-ation in lipid A structure in the pathogenic yersiniae. Mol Microbiol 52:1363–1373. http://dx.doi.org/10.1111/j.1365-2958.2004.04059.x.
74. de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, RaybouldHE. 2010. Propensity to high-fat diet-induced obesity in rats is associatedwith changes in the gut microbiota and gut inflammation. Am J PhysiolGastrointest Liver Physiol 299:G440 –G448. http://dx.doi.org/10.1152/ajpgi.00098.2010.
75. Sefcikova Z, Hajek T, Lenhardt L, Racek L, Mozes S. 2008. Differentfunctional responsibility of the small intestine to high-fat/high-energy dietdetermined the expression of obesity-prone and obesity-resistant pheno-types in rats. Physiol Res 57:467– 474.
76. Hansen GH, Niels-Christiansen LL, Immerdal L, Nystrom BT, Dan-ielsen EM. 2007. Intestinal alkaline phosphatase: selective endocytosisfrom the enterocyte brush border during fat absorption. Am J PhysiolGastrointest Liver Physiol 293:G1325–G1332. http://dx.doi.org/10.1152/ajpgi.00379.2007.
77. Lynes M, Narisawa S, Millan JL, Widmaier EP. 2011. Interactions betweenCD36 and global intestinal alkaline phosphatase in mouse small intestine andeffects of high-fat diet. Am J Physiol Regul Integr Comp Physiol 301:R1738–R1747. http://dx.doi.org/10.1152/ajpregu.00235.2011.
Yang et al.
258 iai.asm.org January 2015 Volume 83 Number 1Infection and Immunity
on March 2, 2020 by guest
http://iai.asm.org/
Dow
nloaded from





























