Embed Size (px)
Citation preview

ORIGINAL PAPER
Intervertebral and Epiphyseal Fusion in the Postnatal Ontogenyof Cetaceans and Terrestrial Mammals
Meghan M. Moran & Sunil Bajpai & J. Craig George &
Robert Suydam & Sharon Usip & J. G. M. Thewissen
# Springer Science+Business Media New York 2014
Abstract In this paper we studied three related aspects of theontogeny of the vertebral centrum of cetaceans and terrestrialmammals in an evolutionary context. We determined patternsof ontogenetic fusion of the vertebral epiphyses in bowheadwha le (Balaena mys t i ce tus ) and be luga wha le(Delphinapterus leucas), comparing those to terrestrial mam-mals and Eocene cetaceans. We found that epiphyseal fusionis initiated in the neck and the sacral region of terrestrialmammals, while in recent aquatic mammals epiphyseal fusionis initiated in the neck and caudal regions, suggesting loco-motor pattern and environment affect fusion pattern. We alsostudied bony fusion of the sacrum and evaluated criteria usedto homologize cetacean vertebrae with the fused sacrum ofterrestrial mammals. We found that the initial ossification ofthe vertebral pedicles in the fetus may be a reliable indicator ofsacral homology in modern cetaceans. Finally, we also studiedfusion of the centra of cervical vertebrae in B. mysticetus andfound that it is not completed until after sexual maturity, andafter 20 years of age.
Keywords Intervertebral . Epiphyseal . Fusion . Cetacea .
Vertebra
Introduction
The vertebral column provides support for the body andallows for flexibility and mobility (Gegenbaur and Bell1878; Hristova et al. 2011; Bruggeman et al. 2012). Toachieve this mobility, individual vertebrae articulate with eachother through cartilaginous intervertebral joints between thecentra and synovial joints between the pre- and post-zygapophyses. The mobility of each vertebral joint variesgreatly between species as well as along the vertebral columnwithin a single species. Vertebral column mobility greatlyimpacts locomotor style, whether the animal is terrestrial oraquatic. In aquatic Cetacea, buoyancy counteracts gravity, andthe tail is the main propulsive organ (Fish 1996; Fish et al.2000). As a result, vertebral column design in cetaceans isvery different from that of terrestrial mammals (Slijper 1936;Buchholtz 1998, 2010). Since cetaceans are derived fromterrestrial mammals, evolutionary changes occurred in theirvertebral column, and these changes are recorded in modernadult morphologies as well as ontogeny. In this paper wecompare aspects of vertebral centrum joint ontogeny betweencetaceans and terrestrial mammals to understand the similar-ities among mammalian morphologies in the vertebral col-umn, regardless of habitat; and follow vertebral column mor-phologies as they change during cetacean evolution.
Our first objective is to document ontogenetic epiphysealfusion patterns in cetaceans and compare them to those ofsome terrestrial mammals. Epiphyseal fusion of the vertebralcentrum is epiphyseal (or growth) plate closure, an ossifica-tion event that results in cessation of longitudinal growth.Vertebral epiphyses fuse over an extended postnatal period,and in some species, such as Balaena mysticetus, some epiph-yses fuse long after sexual maturity. Epiphyseal fusion pat-terns have been studied in terrestrial mammals (Dawson 1925;Purdue 1983; Roach et al. 2003; Munro et al. 2009), as well assome cetaceans (Moore 1968; Ito and Miyazaki 1990; Mead
M. M. Moran (*)Department of Anatomy and Cell Biology, Rush Medical College,600 S. Paulina Street, Suite 506Ac/Fac, Chicago, IL 60612, USAe-mail: [email protected]
S. BajpaiDepartment of Earth Sciences, Indian Institute of Technology,Roorkee, Uttarakhand 247667, India
J. C. George : R. SuydamDepartment of Wildlife Management, North Slope Borough,P.O. Box 69, Barrow, AK 99723, USA
S. Usip : J. G. M. ThewissenDepartment of Anatomy and Neurobiology, Northeast Ohio MedicalUniversity, 4209 State Route 44, Rootstown, OH 44272, USA
J Mammal EvolDOI 10.1007/s10914-014-9256-7

and Potter 1990; Yoshida et al. 1994; Galatius 2010; Wheeler1930; Kato 1988, Kemper and Leppard 1999; Best andLockyer 2002), but little comparative work has been done.
Our second objective is to study the fusion of the sacralvertebrae in ontogeny and evolution and the absence of thisprocess in modern cetaceans. The sacrum is the area of leastmobility in most mammalian vertebral columns since it is thesite where the weight-bearing hind limb is anchored to theaxial skeleton. In terrestrial mammals, the process of sacralfusion, i.e., the ossification of the intervertebral disc, occurswell after birth, affecting the stability of the pelvis in immatureanimals. In modern cetaceans and sirenians, there is no weightbearing hind limb and the sacral vertebrae remain unfused(Eschricht et al. 1866; Slijper 1936; Buchholtz 2001, 2007;Buchholtz et al. 2005). Because of this, it is difficult todistinguish cetacean sacral vertebrae from adjacent lumbarand anterior caudal vertebrae, causing different authors touse different criteria to homologize cetacean vertebrae.
There is no consensus on criteria to distinguish sacralvertebrae in cetaceans and we compare different methods ofdistinguishing and homologizing sacral vertebrae in terrestrialmammals and cetaceans, and evaluate them against observa-tions on cetacean fetuses. Many marine mammal anatomistsavoid the issue altogether and use the term lumbar vertebraefor all vertebrae between the last rib-bearing vertebra and thefirst hemal-arch-bearing vertebra (e.g., Buchholtz 2001,2010). Lumbar, in this definition, includes traditionally de-fined lumbar vertebrae plus unfused sacral, and even anteriorcaudal vertebrae since the first hemal arch is often not presenton the first caudal vertebra in animals that have a sacrum(Eschricht et al. 1866; Slijper 1936; Evans 1993; Buchholtz2010). Slijper (1936) proposed that the modern cetacean sa-crum could be recognized on the basis of the location of thepudendal nerve as it exits the intervertebral foramen; the firstroot of this nerve passes through the foramen between S1 andS2 (Slijper 1936).
Ontogenetic fusion of sacral vertebrae is of special interestin fossil cetaceans. Cetaceans originated from terrestrial mam-mals (Thewissen et al. 2007, 2009), and early cetaceanspossessed fused sacra (Gingerich et al. 2001; Thewissenet al. 2001) and were able to locomote on land (Thewissenet al. 1996, 2001; Madar et al. 2002). The origin of cetaceansis well documented by fossils and vertebral columns areknown for many of these (Fraas 1904; Kellogg 1936;Thewissen et al. 1996; Uhen 1999; Gingerich et al. 2001;Madar et al. 2002; Uhen 2004; Thewissen and Bajpai 2009;Bebej et al. 2012). For many Eocene fossil cetaceans, there isno ambiguity about the homology of the sacrum becausemultiple vertebrae are fused together and this vertebral regionarticulates with the illium. However, for some of these spec-imens, only a single fossil sacrum has been found, and theontogenetic age of these fossil cetaceans is unknown. Giventhe slow pace of sacral centrum fusion after birth, low sample
sizes complicate determination of the number of vertebrae thateventually make up the sacrum in fossils.
Our third objective is to document the fusion of cervicalvertebrae in B. mysticetus. Unlike most mammals,B. mysticetus and some other cetaceans undergo intervertebralfusion of their cervical vertebrae centra (Eschricht et al. 1866;Wheeler 1930; Slijper 1936; Haldiman and Tarpley 1993;Buchholtz 2001, 2007; Buchholtz et al. 2005, 2007). Thisseverely limits neck mobility in cetaceans, even though thisis the area of greatest vertebral mobility in most other mam-mals (Graf et al. 1995; Bebej 2011; Bebej et al. 2012). Just likesacral fusion in terrestrial mammals, cervical fusion is a pro-cess with a protracted ontogeny, but no age series of cetaceannecks with different levels of fusion have been described.
Materials and Methods
This study used an ontogenetic series of 20 mice (Musmusculus); skeletal samples of nine pigs (Sus scrofa); anatom-ical specimens of eight bowhead whales (Balaena mysticetus)and nine beluga whales (Delphinapterus leucas); two pan-tropical spotted dolphin (Stenella attenuata) fetuses; oneB. mysticetus fetus; and skeletons of the Eocene cetaceansAmbulocetus natans and Kutchicetus minimus. Techniquesused include computed tomography imaging, whole mountclearing and staining, anatomical dissections, and osteologicalstudy.
Epiphyseal Fusion
Epiphyseal fusion within a single vertebra and across thevertebral column was illustrated in a diagram that we refer toas a fusion map. Each vertebra is represented by a rectangleand the degree of epiphyseal fusion is indicated in shades ofgray. To determine the epiphyseal fusion in the fossils, spec-imens were scored based on the fusion scar along the epiph-yses. If an epiphysis was missing, the joint was scored asunfused (light gray). If a fusion scar was visible for part or theentire circumference of the centrum, it was scored as partiallyfused (dark gray). If no fusion scar was visible along any partof the epiphysis, the joint was scored as fused (black). Scoringis more difficult in fossil specimens than in our recent sam-ples. In some fossils the entire circumference of a vertebra wasnot visible because of damage to the specimen, or because itwas covered with sediment. In those cases, specimens werescored on the basis of the visible segment. If the vertebra wasnot recovered, it was scored as missing data (white). Thisprocedure is likely to have introduced some noise in our datathat is not present in the scores of the modern animals.Vertebrae were identified using the descriptions forA. natans (Thewissen et al. 1996; Madar et al. 2002) andK. minimus (Thewissen and Bajpai 2009).
J Mammal Evol

Fusion data for M. musculus were collected from thecleared and stained ontogenetic mouse series. This techniqueprovided clear visualization of the epiphyseal fusion pattern.Fusion data in fossils were determined from inspection of theexternal surface of each vertebra. The inability to inspect theinternal morphology of these vertebrae introduces a level ofvariation that is not present in the other methods and is a likelysource of noise in our fossil data.
Modern Cetacean Osteological Samples
We recorded vertebral counts, intervertebral and epiphysealfusion patterns in B. mysticetus and D. leucas, during nativeAlaskan subsistence hunts in Barrow and Point Lay, Alaska.These samples were collected under permit NOAA-NMFS814-1899-01 with permission from the Alaska EskimoWhaling Commission and local subsistence hunters. Sagittalsamples of vertebral centra, intervertebral discs, growth plates,and epiphyses were cut using manual and power saws. Somesamples were skeletonized by cold water maceration. Balenamysticetuswere aged using criteria based on baleen length andbody length (Lubetkin et al. 2012), but D. leucas could onlybe relatively aged by comparing degrees of fusion and assum-ing that more epiphyseal fusion is present in older individuals.For D. leucas, a large number of partial vertebral columnswere available. Balena mysticetus and D. leucas specimensprovided data on epiphyseal fusion. Cervical fusion was stud-ied in B. mysticetus only, because D. leucas do not fuse theircervical vertebrae.
Ontogenetic Series of Mus Musculus
We collected an ontogenetic series ofM. musculus from birthto 60 days postnatal every ten days, plus individuals overone year old. Specimens were cleared and stained to showonly bone (Alizarin Red) and cartilage (Alcian Blue) struc-tures, and all other tissues were chemically cleared, using aprotocol adapted from Wassersug (1976). This collection of-fered details on the timing of bony epiphyseal and sacralfusion in a terrestrial mammal.
Artiodactyl Osteological Collection
We used S. scrofa as a second modern comparative sample toinvestigate epiphyseal and sacral fusion, because terrestrialartiodactyls are the closest mammalian relative of cetaceans(Thewissen and Madar 1999; Geisler et al. 2007; Thewissenet al. 2007; Spaulding et al. 2009). Osteological specimenswere studied at the Field Museum of Natural History inChicago, IL, and the Carnegie Museum of Natural Historyin Pittsburgh, PA. The S. scrofa studied were wild-caught ordomesticated individuals for which no exact ages were avail-able. Instead, ages of the S. scrofa were estimated from tooth
eruption sequences and wear pattern (Getty 1975; Hillson1986).
Modern Cetacean Fetuses
Stenella attenuata specimens were acquired from the LosAngeles Natural History Museum (LACM), and pertain toCarnegie Stage 21–23 (Thewissen and Heyning 2007). Thepelvic regions of S. attenuata LACM 94310 and B. mysticetus2000B3F fetuses were dissected to identify the location of thepudendal nerve before clearing-and-staining. LACM 94310 is185 mm in total length (TL) and is from C21-22. TheB. mysticetus fetus 2000B3F is 403 mm in TL and fromC23. These specimens, along with LACM 94382 (TL=225 mm), were cleared and stained to investigate the presenceof morphological traces of sacrum development.
Eocene Cetaceans
Two of the most basal cetaceans (Geisler and Uhen 2005;Uhen et al. 2011) were chosen for study because nearlycomplete vertebral columns are known for both A. natans(Ambulocetidae, HGSP 18507; Thewissen et al. 1996;Madar et al. 2002) and K. minimus (Remingtonocetidae,IITR-SB 2647; Bajpai and Thewissen 2000; Thewissen andBajpai 2009).
Computed Tomography
Computed tomography (CT) scans of modern and fossil ceta-cean specimens were completed at Loyola UniversityMedicalCenter in Maywood, IL, using a Siemens Somatom SensationCT scanner that produced sequential slices at 0.6 mm thick-ness and at NEOMED, Rootstown, OH, on a Norland StratecXCTBoneDensitometerμCTscanner yielding 0.25mm thickslices. Sequential CT images were studied using Image J(Rasband 2011).
Institutional Abbreviations
CM, Carnegie Museum of Natural History; FMNH, FieldMuseum of Natural History; HGSP, Howard University-Geological Survey of Pakistan, Quetta, Pakistan; IITR-SB,Indian Institute of Technology, Roorkee, India- Sunil Bajpaicollection; LACM, Los Angeles Natural History Museum,specimens deposited at Northeast Ohio Medical University(NEOMED); LUMC, Loyola University Medical Center;NOAA-NMFS, National Oceanic and AtmosphericAdministration- National Marine Fisheries Service.
J Mammal Evol

Results
Epiphyseal Fusion
The ontogenetic M. musculus series shows that, forindividual vertebrae, the cranial epiphysis always fusesto the centrum before the caudal epiphysis (Fig. 1).Epiphyseal fusion starts around ten days postnatal(Fig. 1ii) in two regions: the neck and sacrum. Fusionprogresses caudally from the cervical and sacral regionsas ontogeny progresses (Fig. 1ii–vi). Epiphyses of theseven cervical vertebrae and four sacral vertebrae arecompletely fused at 20 days (Fig. 1iii). At that age,cranial epiphyses of the thoracic vertebrae are complete-ly fused, but the caudal epiphyses are not, and bothepiphyses of the lumbar and anterior caudal vertebraeare partially fused. Epiphyseal fusion progresses to theanterior thoracic and caudal vertebrae by 30 dayspostnatal (Fig. 1iv). At 40 days postnatal, the cervical,anterior thoracic, sacral, and anterior caudal vertebraeare completely fused, as are the cranial epiphyses of thethoracics and most of the posterior caudal vertebrae
(Fig. 1v). The posterior lumbar epiphyses are the lastto fuse (Fig. 1vi). Complete epiphyseal fusion is presentby 60 days postnatal (Fig. 1vii, viii), well after sexualmaturity.
In S. scrofa, epiphyseal fusion starts after the erup-tion of the third molar (M3), when the animal is ap-proximately 18–20 months old (Getty 1975), and thepattern is similar to that of M. musculus in that thecranial epiphysis fuses to the centrum before the caudalepiphysis (Fig. 2). Similar to M. musculus, epiphysealfusion progresses caudally through the vertebral column,beginning in two different regions (Fig. 2i–v). In bothspecies, the anterior center of fusion is in the cervicalregion. However, the posterior ossification center inS. scrofa is located in the anterior caudal vertebrae,rather than in the sacrum as in M. musculus .Epiphyseal fusion progresses cranially and caudallyfrom this posterior ossification center (Fig. 2v, vi).The anterior thoracic vertebral epiphyses fuse last(Fig. 2viii, ix).
Epiphyseal fusion in B. mysticetus and D. leucas startsfrom the cervical and the caudal regions; fusion continues
Fig. 1 Mus musculus postnatal fusion maps. i, 1 day (M81); ii, 10 day(M54); iii, 20 day (M82); iv, 30 day (M78); v, 40 day (M143); vi, 50 day(M83); vii, 60 day (M84); viii, 230 day (M85). Blocks with two colorsshow different amounts of fusion within a single vertebra. Arrows show
direction of epiphyseal fusion. C2, second cervical vertebra; T1, firstthoracic vertebra; L1, first lumbar vertebra; S1, first sacral vertebra;Ca1, first caudal vertebra. Light gray, no epiphyseal fusion; dark gray,partial epiphyseal fusion; black, complete fusion
J Mammal Evol

towards mid-column from both these end regions (Fig. 3). Theanterior edge of the caudal ossification center in B. mysticetusis located at vertebra 36 (post-cervical 29 or post-thoracic 15)in one specimen (Fig. 3i) and four vertebral levels caudal tothat in another specimen (Fig. 3iv). Epiphyses of anteriorthoracic vertebrae are starting to fuse by five to ten years oldin B. mysticetus (Fig. 3ii). Cervical epiphyseal fusion is notcomplete in a one year old B. mysticetus (Fig. 3i) but iscomplete in the 40 year old B. mysticetus. The thoracic andlumbar vertebral epiphyses remain unfused through 20 yearsof age (Fig. 3iv).Delphinapterus leucas demonstrate the sameepiphyseal fusion pattern as B. mysticetus (Fig. 3v–xiii).Posterior thoracics are the last vertebrae to fuse in D. leucas.The posterior caudal vertebrae are not preserved in these wildcaught specimens and therefore fusion pattern is unknown.
As in modern mammals, including cetaceans, in gen-eral, cranial epiphyses fuse before caudal epiphyses, andin both A. natans and K. minimus there is an area near
the sacrum where fusion is completed before other areas(Fig. 4). A cervical center of fusion occurs inK. minimus, whereas the A. natans specimen appearsto be younger and lack this center (or possibly cervicalvertebrae are too incompletely preserved to detect it). Inboth fossil cetaceans, mid-caudal vertebrae have firmlyfused epiphyses and there is the suggestion that fusionhere does not start at the sacral center, but instead thatit commences at a separate, mid-caudal center (Fig. 4).
Sacral Centrum Fusion and Sacral Homology
Intervertebral sacral joints fuse in a cranial to caudal directionin M. musculus. Intervertebral fusion causes sacral discs todecrease in width and eventually be replaced by bone(Fig. 5i–iii). At 20 days postnatal all intervertebral discsare intact, and the nucleus pulposus is present in eachsacral joint (Fig. 5i). By 40 days postnatal, the S1/S2
Fig. 2 Sus scrofa postnatal fusion maps. i, FMNH 49844; ii, CM 59541;iii, FMNH 42440; iv, FMNH 42439; v, CM 16; vi, CM 30427; vii,FMNH 97884; viii, FMNH 92908; ix, FMNH 92907. Arrows show
direction of epiphyseal fusion. Light gray, no epiphyseal fusion; darkgray, partial epiphyseal fusion; black, complete fusion; white, missingvertebra
J Mammal Evol

and S2/S3 intervertebral discs are ossified (Fig. 5ii), andthe S3/S4 joint is maintained with the nucleus pulposusstill intact (Fig. 5ii). At 60 days all three sacral jointsare fused (Fig. 5iii). Residual growth plates are presentas thin strips of cartilage (Fig. 5ii, iii). This indicatesthat full sacral fusion in M. musculus occurs around orjust after sexual maturity.
In S. scrofa, the S1/S2 joint is the first to fuse and fusionprogresses caudally to the last sacral joint (S3/S4), similar toM. musculus (Fig. 5iv–vi). At six to eight months of age,sacral intervertebral fusion is present in the S1/S2 and S2/S3joints, but not in the S3/S4 joint (Fig. 5iv). By 18–20 months,the sacral centra are fusing (Fig. 5v), although at this age thegrowth plates are still present. This implies that sacral
Fig. 3 Balaena mysticetus and Delphinapterus leucas fusion maps. i–iv,B. mysticetus specimens: i, 1 year (09B11); ii, 5 years (11B8); iii, 5–10 years (09B5); iv, 20 years (11B9). The ages of these individuals aredetermined based on methods described by Lubetkin et al. (2012). v–xiii,D. leucas specimens. Individual belugas are ranked according to
epiphyseal fusion; ages are unknown as they were hunted, wild-caughtindividuals. Arrows show direction of epiphyseal fusion. Light gray, noepiphyseal fusion; dark gray, partial epiphyseal fusion; black, completefusion; white, missing data
J Mammal Evol

intervertebral fusion occurs before epiphyseal fusion inS. scrofa. In S. scrofa older than 20 months all sacral jointsare fused (Fig. 5vi).
Lacking intervertebral fusion in the sacrum of cetaceansmakes homologizing these vertebrae difficult. We evaluatedSlijper’s (1936) pudendal nerve location as a criterion for
sacral homology. Dissection of the pudendal nerve ofS. attenuata (Fig. 6i, ii) shows that it receives ventral ramifrom three spinal nerves. The first root of the pudendal nerveexits the intervertebral foramen between the 10th and 11thpost-thoracic vertebrae (Fig. 6i, ii). These nerves join anddescend through the abdominal wall to the genital region
Fig. 4 Ambulocetus natans and Kutchicetus minimus fusion maps. i, A. natans (HGSP 18507). ii, K. minimus (IITR-SB 2647). Specimen numbers arelisted for each vertebra. Light gray, no epiphyseal fusion; dark gray, partial epiphyseal fusion; black, complete fusion; white, missing data
Fig. 5 Terrestrial mammal sacra. i–iii, Cleared and stained mice(M. musculus) sacra, ventral view. i, 20 days; ii, 40 days; iii, 60 dayspostnatal. iv–vi, Skeletonized S. scrofa sacra. iv, 6–8 months (FMNH
49844); v, 18–20 months (FMNH 42439); vi, greater than18–20 months(CM 30427) postnatal. S1 S2 S3 S4, sacral vertebrae 1–4; gp, growthplate; np, nucleus pulposus; tp, transverse process; uf, unfused
J Mammal Evol

where they innervate the external anal sphincter and genitals(Fig. 6i, ii). This fetus (LACM 94310) and severalothers were subsequently cleared and stained (Fig. 7i),and show that centers of ossification are present in mostof the cartilaginous vertebral centra and vertebral arches(Fig. 7).
LACM 94310 has a total of 79 vertebrae, sevencervical, 12 rib-bearing thoracics, and 56 post-thoracics(Fig. 7i). Sixty-nine of these vertebrae are partly ossi-fied, and the last ten caudal vertebrae are cartilaginous.As is normal among mammals, independent ossificationcores occur in the centrum, vertebral arch, and spinousprocess. Unusually though, the pedicle ossifications arelarger in post-thoracic vertebrae 13–17 (Fig. 7i), withthose of post-thoracic vertebra 13 and 17 smaller thanthose of 14–16. This pattern is similar to that in anolder S. attenuata fetus (LACM 94382, Fig. 7ii). Thisfetus also has 79 total vertebrae, but the larger sacralossifications are present in post-thoracic vertebrae 15–18
(Fig. 7ii). The larger spinous process ossifications cor-respond to the location of the sacrum as identified bythe pudendal nerve dissection in both individuals(Fig. 7).
Dissection of the B. mysticetus fetus (2000B3F) showedthat the first root for the pudendal nerve passes throughthe intervertebral foramen between the 7th and 8th post-thoracic vertebrae (Fig. 6iii, iv). This fetus has a total of55 vertebrae. Laminae and pedicles of the vertebrae areossified for most of the column, but larger ossificationsare present at post-thoracic vertebrae 3–7 (Fig. 7iii). Theselarger ossifications are more anteriorly located within thevertebral column than those in S. attenuata, and are ante-rior to the exit location of the pudendal nerve, which isalso unlike that in S. attenuata.
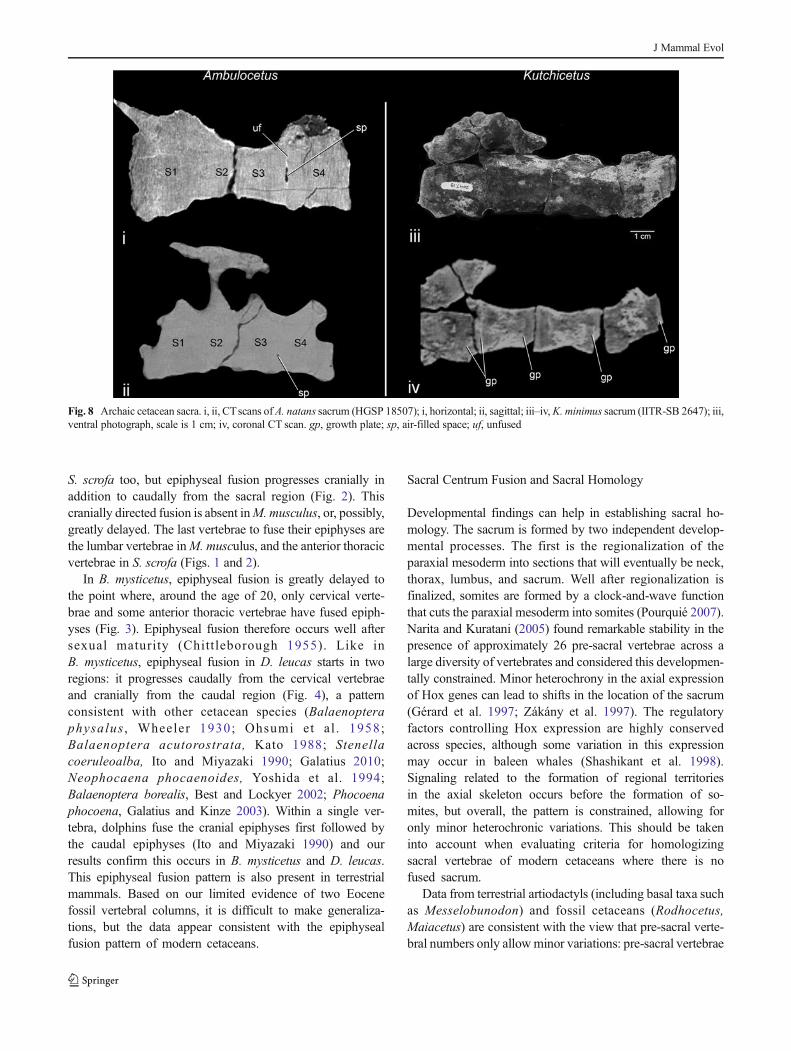
CT scans of the four vertebrae A. natans sacrum(HGSP 18507.834) indicate complete fusion of the S1/S2and S2/S3 joints (Fig. 8i, ii). The S3/S4 joint is fused atthe periphery of the joint, but an oval-shaped radiolucent
Fig. 6 Pudendal nerve of modern cetaceans. i, S. attenuata fetus,stage C21/22 (LACM 94268); ii, black and white line drawing ofS. attenuata pudendal nerve; iii, B. mysticetus fetus, stage C23(2000B3F); iv, black and white line drawing of B. mysticetuspudendal nerve. cc, crus of corpus cavernosum; dn, dorsal nerve
of the penis; ep, epididymus; pe, penis; pn, pudendal nerve; ri,right innominate; S1/Pt10, first sacral vertebra according to Slijperor post-thoracic vertebra 10; sp, spinous process; te, testis; ti,tibia; tp, transverse process; vr, ventral ramus
J Mammal Evol

space is visible in the joint on the CT scan (Fig. 8i, ii).This may be where the nucleus pulposus once resided.There is also a thin radio-opaque line visible on the CTscans between S3 and S4 extending from the edges of thisoval space (Fig. 8i). This white line is a narrow extensionof the air-filled space between S3 and S4 that was infil-trated by mineral.
The four vertebrae of the sacrum (IITR-SB 2647.13) ofK. minimus were incompletely fused and broken after death,resulting in individual sacral vertebrae that are still distinct onthe CT scans (Fig. 8iii). Thewissen and Bajpai (2009) pro-posed that this specimen of K. minimus was immature(Fig. 8iii, iv); sacral centrum fusion pattern remains unknown.
Cervical Intervertebral Fusion
InB. mysticetus, the seven cervical vertebrae fuse together andare remodeled to form a solid, immobile bony block (Fig. 9).
Our postnatal series of B. mysticetus skeletal specimens showsintervertebral fusion in a 1 year old individual (dark gray inFig. 9i). Bony intervertebral fusion (black) is present at thecenter of the cervical vertebrae and progresses peripherally(Fig. 9i–iv). By the age of 20, most cervical vertebrae havefused, with the exception of the most caudal cervical joints(Fig. 9v, vi). By 40 years of age, all seven cervical vertebraeand all growth plates are completely fused and the individualcentra cannot be recognized (Fig. 9vii).
Discussion
Epiphyseal Fusion
InM. musculus, epiphyseal fusion starts in cervical and sacralvertebrae and simultaneously progresses from those two re-gions caudally (Fig. 1). These two fusion centers are present in
Fig. 7 Cleared and stainedmodern fetal cetaceans. i,S. attenuata LACM 94310; ii,S. attenuata LACM 94382; iiiB. mysticetus 2000B3F. The blackovals surround the larger vertebralossifications that correspond tothe location of the sacrum asidentified by pudendal nervedissection in S. attenuata (i, ii). InB. mysticetus (iii) the largervertebral ossifications are locatedanterior to the sacrum asidentified through pudendal nervelocation
J Mammal Evol
S. scrofa too, but epiphyseal fusion progresses cranially inaddition to caudally from the sacral region (Fig. 2). Thiscranially directed fusion is absent inM.musculus, or, possibly,greatly delayed. The last vertebrae to fuse their epiphyses arethe lumbar vertebrae inM. musculus, and the anterior thoracicvertebrae in S. scrofa (Figs. 1 and 2).
In B. mysticetus, epiphyseal fusion is greatly delayed tothe point where, around the age of 20, only cervical verte-brae and some anterior thoracic vertebrae have fused epiph-yses (Fig. 3). Epiphyseal fusion therefore occurs well aftersexual maturity (Chitt leborough 1955). Like inB. mysticetus, epiphyseal fusion in D. leucas starts in tworegions: it progresses caudally from the cervical vertebraeand cranially from the caudal region (Fig. 4), a patternconsistent with other cetacean species (Balaenopteraphysalus , Wheeler 1930; Ohsumi et al . 1958;Balaenoptera acutorostrata, Kato 1988; Stenellacoeruleoalba, Ito and Miyazaki 1990; Galatius 2010;Neophocaena phocaenoides, Yoshida et al. 1994;Balaenoptera borealis, Best and Lockyer 2002; Phocoenaphocoena, Galatius and Kinze 2003). Within a single ver-tebra, dolphins fuse the cranial epiphyses first followed bythe caudal epiphyses (Ito and Miyazaki 1990) and ourresults confirm this occurs in B. mysticetus and D. leucas.This epiphyseal fusion pattern is also present in terrestrialmammals. Based on our limited evidence of two Eocenefossil vertebral columns, it is difficult to make generaliza-tions, but the data appear consistent with the epiphysealfusion pattern of modern cetaceans.
Sacral Centrum Fusion and Sacral Homology
Developmental findings can help in establishing sacral ho-mology. The sacrum is formed by two independent develop-mental processes. The first is the regionalization of theparaxial mesoderm into sections that will eventually be neck,thorax, lumbus, and sacrum. Well after regionalization isfinalized, somites are formed by a clock-and-wave functionthat cuts the paraxial mesoderm into somites (Pourquié 2007).Narita and Kuratani (2005) found remarkable stability in thepresence of approximately 26 pre-sacral vertebrae across alarge diversity of vertebrates and considered this developmen-tally constrained. Minor heterochrony in the axial expressionof Hox genes can lead to shifts in the location of the sacrum(Gérard et al. 1997; Zákány et al. 1997). The regulatoryfactors controlling Hox expression are highly conservedacross species, although some variation in this expressionmay occur in baleen whales (Shashikant et al. 1998).Signaling related to the formation of regional territoriesin the axial skeleton occurs before the formation of so-mites, but overall, the pattern is constrained, allowing foronly minor heterochronic variations. This should be takeninto account when evaluating criteria for homologizingsacral vertebrae of modern cetaceans where there is nofused sacrum.
Data from terrestrial artiodactyls (including basal taxa suchas Messelobunodon) and fossil cetaceans (Rodhocetus,Maiacetus) are consistent with the view that pre-sacral verte-bral numbers only allowminor variations: pre-sacral vertebrae
Fig. 8 Archaic cetacean sacra. i, ii, CTscans of A. natans sacrum (HGSP 18507); i, horizontal; ii, sagittal; iii–iv,K. minimus sacrum (IITR-SB 2647); iii,ventral photograph, scale is 1 cm; iv, coronal CT scan. gp, growth plate; sp, air-filled space; uf, unfused
J Mammal Evol

in these taxa vary between 26 and 28 (Table 1; Lessertisseurand Saban 1967). However, this developmental constraint wasreleased at least once early in the cetacean lineage, as even theEocene cetacean A. natans has 31 pre-sacral vertebrae, and itis likely that such constraints are released in modern
cetaceans, where most homology criteria show vastly differentnumbers of pre-sacral vertebrae (Table 1).
A common practice among cetologists is to use the locationof the first hemal arch to indicate the first caudal vertebrae inmodern cetaceans. In modern terrestrial artiodactyls and
Fig. 9 Balaena mysticetus cervical specimen photographs and cervicalfusion maps. On the left, photographs of skeletonized bowhead cervicalsamples, sagittal view. Scale bar: 1 cm for i–vi. Scale bar: 5 cm for vii. Onthe right, corresponding fusion maps for each specimen. i, 1 year old(09B11); ii, 2 years old (10B16); iii, 5 years old (11B8); iv, 5–10 years old
(09B9); v, 20 years old (10B15); vi, 20 years old (11B9); vii, 40 years old(11B3). The vertebrae are black boxes separated by the cartilaginousintervertebral disc (i–vi). Growth plates are depicted as zigzag lines.C1–C7, cervical vertebrae 1–7
J Mammal Evol

Eocene cetaceans, hemal arches indeed occur at the caudalend of the first caudal vertebra (Zhou et al. 1992; Buchholtz2007), although this is by no means a common feature ofmammals, e.g., dogs lack hemal arches on the anterior caudalvertebrae (Nickel et al. 1954; Evans 1993). If hemal archlocation is an indicator of sacral position (by way of identify-ing the first caudal vertebra) in cetaceans, the number of pre-sacral vertebrae varies much more than in other mammalorders (Table 1; Narita and Kuratani 2005; Buchholtz 2001,2007; Buchholtz et al. 2005). In late fetal S. scrofa, the firstand second caudal vertebra lack hemal arches, while they arepresent in more posterior vertebrae (personal observation).
Slijper (1936) suggested that the location of the firstspinal root of the pudendal nerve can be used as a markerfor the first sacral vertebra (S1). This feature cannot bechecked in fossils, but comparison of three modern ceta-cean species indicates that this criterion correlates verypoorly with the presence of the first hemal arch (Table 1).This holds true in M. musculus as well (Table 1), suggest-ing the location of the first spinal root of the pudendalnerve appears to be variable among mammals. The positionof the hind limb bud would appear to be a robust way todetermine the homology of sacral vertebrae in cetaceans,because the hind limb is initiated early in embryology byaxial signaling, and is initially intact in cetaceans(Thewissen et al. 2009). Interestingly, the position of thehind limb bud in S. attenuata matches that of the firsthemal arch, consistent with views by different authors, thatthese indicate sacrum position. However, there are no dataon the location of the hind limb bud with regard to thesomites in any other cetacean. In M. musculus embryos, thehind limb bud shifts position greatly during ontogeny(Table 1). More data on cetaceans are needed before
assumptions about the location of their hind limb bud canbe assumed to be stable.
The vertebral pedicle in some modern cetaceans (Table 1)shows an area of early ossification that could indicate thepresence of sacral vertebrae. Indeed, the first sacral vertebrain M. musculus fetuses at E18.5 also indicates stronger ossi-fication in the pedicle (McIntyre et al. 2007). Moreover, theidentical location of this area of ossification in two distantlyrelated modern cetaceans (B. mysticetus and S. attenuata,Table 1) could be taken to suggest that some developmentalsignaling of sacral formation has been retained, but that theirinitiation has been shifted to a more caudal somite position.Study of terrestrial artiodactyls fetuses could be used to testthe hypothesis that the first signs of ossification in the pedicleindicate the location of the sacral region.
Sacral Fusion
Figure 10 summarizes sacrum evolution in cetaceans based ona phylogenetic framework presented by Geisler et al. (2007),Geisler and Theodor (2009), and Spaulding et al. (2009).Archaic artiodactyls, including the sister group to cetaceans(Thewissen et al. 2007; Geisler et al. 2007), had three or fourvertebrae in their sacrum. Only a few fossil sacra are availablefor dichobunid artiodactyls (Rose 1985; Thewissen andHussain 1990), raoellid artiodactyls, and pakicetid cetaceans(Cooper et al. 2012); all have three fused sacral vertebrae.Since full sacral fusion occurs late in postnatal ontogeny, it ispossible that some of these are young animals and not allsacral vertebrae had fused yet. Usually, in Eocene cetaceans,there are four firmly fused vertebrae with a well-developedilio-sacral joint on S1. The only specimen of A. natans has afour-vertebra sacrum (Fig. 8), even though the number of
Table 1 Interspecies comparison of vertebral column morphological landmarks
# of presacral vertebrae, asbased on fused sacrum
1st hemal arch (Ca1) 1st root of pudendalnerve (S1)
1st vertebral pedicle withfetal sacral ossification
Fetal hind limb budlocation
Mus musculus (Mouse) 27 31 25 27 15–20 shifts to 25–30
Artiodactyla
Sus scrofa (Pig) 26–28 x 30–31 x 18
Messelobunodon 26 x x x x
Modern Cetacea
Balaena mysticetus(Bowhead)
x 33 27 33 x
Delphinapterus leucas(Beluga)
x 29 22 x x
Stenella attenuata(Dolphin)
x 46 30 33 45
Eocene Cetacea
Ambulocetus natans 31 x x x x
Rodhocetus kasrani 26 31 x x x
Maiacetus inuus 26 31 x x x
J Mammal Evol

unfused vertebrae suggests that is not skeletally mature(Figs. 4 and 8).
In cetaceans, the loss of fusion in the sacrum and the loss ofilio-sacral contact occur within the family Protocetidae(Fig. 10). The protocetid Rodhocetus was described as havingan ilio-sacral contact, but having unfused sacral vertebrae(Gingerich et al. 1994). It is possible that, with age, some ofthe sacral centra of Rodhocetus would fuse. Qaisracetus arifi
(Gingerich et al. 2001) and Takracetus simus (Gingerich et al.2001) retain a fused joint between S1 and S2, but the latter haslost any ilio-sacral joint contact (Fig. 10).
Cervical Intervertebral Fusion
In B. mysticetus, cervical intervertebral fusion starts in thecenter of each of the intervertebral discs and progresses
Fig. 10 Cetacean phylogeny of sacral fusion. Notice the evolutionary loss of sacral intervertebral fusion through cetacean evolution. The loss ofvertebral articulation with the pelvis can also be traced evolutionarily. ?- questionable articulation and fusion
J Mammal Evol

Fig. 11 Phylogeny of cervical fusion related as to neck length. 0 fusedcervical vertebrae, white, 2–4 fused cervical vertebrae, grey; 6–7 fusedcervical vertebrae, black. Phylogeny based on Messenger and McGuire(1998) strict consensus phylogeny, cervical fusion counts (Wheeler 1930;
Slijper 1936; Haldiman and Tarpley 1993; Buchholtz 2001, 2007, 2010;Buchholtz et al. 2005), Remingtonocetus (Bebej et al. 2012), Dorudon(Uhen 2004)
J Mammal Evol

peripherally: the first region to ossify is the nucleus pulposus(Fig. 9). Cervical intervertebral fusion in B. mysticetus isgradual with complete fusion occurring after 20 years of age(Fig. 9). This fusion pattern is different from the intervertebralfusion pattern in the sacrum of M. musculus and S. scrofa,which starts in the periphery of the disc (annulus fibrosus) andprogresses towards the nucleus pulposus (Fig. 5). This centralfusion in B. mysticetus cervical region causes ossification inthe intervertebral discs and also in the vertebral growth plates(Fig. 9). With larger sample sizes it could be possible to usethe ossification pattern of the neck to determine the age ofB. mysticetus, a subject of interest in managing these popula-tions (Lubetkin et al. 2012).
Fusion of cervical vertebrae limits neck mobility and fullfusion makes movement impossible. Neck length and cervicalvertebrae width also limit neck mobility (Buchholtz 1998;Bebej 2011; Bebej et al. 2012). Figure 11 summarizes necklength and cervical fusion in cetaceans in a phylogeneticcontext. This figure shows that cervical fusion was lost oracquired independently a number of times, and that there is abroad, but imperfect relation between neck shortening andcervical fusion.
Movements of the neck are partly determined by thevestibulocollic reflex (Sipla and Spoor 2008), and this reflexmust be lessened or absent in animals lacking a mobile neck(Spoor et al. 2002). Cetacean semicircular canals are unusu-ally small for mammals (Spoor et al. 2002; Spoor andThewissen 2008; Kandel and Hullar 2010). Among cetaceans,B. mysticetus have some of the largest semicircular canals(Spoor and Thewissen 2008). Given that cervical vertebraefuse later in life, it is possible that the vestibulocollic reflex isoperational, to some extent, in young B. mysticetus.
Conclusion
We studied three aspects of bony fusion in the vertebralcolumns of pre- and postnatal cetaceans in order to understandthe dramatic changes that the cetacean vertebral columnunderwent as cetaceans became an obligate aquatic taxa. Insome respects, cetaceans retained terrestrial mammal patterns.For instance, epiphyseal fusion patterns that start from twoareas, the neck and the sacro-caudal regions, are similar incetaceans and terrestrial mammals. The evolution of some ofthe vertebral fusion patterns was gradual, exemplified by theloss of fused vertebrae in the sacrum. In other aspects, highlyderived morphologies evolved multiple times in the cetaceanorder, as demonstrated by the pattern of cervical vertebraefusion in B. mysticetus. Taken as a whole, we view thecetacean vertebral column as more plastic than that of othermammals. Embryological study can elucidate developmentalconstraints that may have been released in the evolution of thecetacean vertebral column (Thewissen et al. 2012). This view
is consistent with previous authors who studied the cetaceanvertebral column (Slijper 1936; Buchholtz 2001, 2010).
Acknowledgments We would like to thank the Department of WildlifeManagement, North Slope Borough; the Alaska Eskimo Whaling Com-mission, local subsistence hunters; Dr. Terrence Demos, Department ofRadiology, LUMC, for his generosity with scanning fossil cetaceans; Dr.Sirpa Nummela, University of Helsinki; Dr. Lisa Noelle Cooper,NEOMED; Dr. John F. Moran, LUMC; Dr. Larry Heaney and JohnMead, FMNH and Sue McLaren, CM for their help and access tomuseum collections; and Dr. Emily Buchholtz for her correspondenceduring this project.
References
Bajpai S, Thewissen JGM (2000) A new, diminutive Eocene whale fromKachchh (Gujarat, India) and its implications for locomotor evolu-tion of cetaceans. Curr Sci India 79:1478–1482
Bebej RM (2011) Functional morphology of the vertebral column inRemingtonocetus (Mammalia, Cetacea) and the evolution of aquaticlocomotion in early archaeocetes. PhD Dissertation, University ofMichigan
Bebej RM, ul-HaqM, Zalmout IS, Gingerich PD (2012)Morphology andfunction of the vertebral column in Remingtonocetus domandaensis(Mammalia, Cetacea) from the middle Eocene Domanda Formationof Pakistan. J Mammal Evol 19:77–104
Best PB, Lockyer CH (2002) Reproduction, growth, andmigrations of seiwhales Balaenoptera borealis off the west coast of South Africa. SAfr J Marine Sci 24:111–133
Bruggeman BJ, Maier JA, Mohiuddin YS, Powers R, Lo Y, Guimarães-Camboa N, Evans SM, Arfe BD (2012) Avian intervertebral discarises from rostral sclerotome and lacks a nucleus pulposus: impli-cations for evolution of the vertebrate disc. Dev Dynam 241:567–683
Buchholtz EA (1998) Implications of vertebral morphology for locomo-tor evolution in early Cetacea. In: Thewissen JGM (ed) TheEmergence of Whales, Evolutionary Patterns in the Origin ofCetacea. Plenum Press, New York, pp 325–351
Buchholtz EA (2001) Vertebral osteology and swimming style in livingand fossil whales (Order: Cetacea). J Zool 253:175–190
Buchholtz EA (2007) Modular evolution of the cetacean vertebral col-umn. Evol Dev 9:278–289
Buchholtz EA (2010) Vertebral and rib anatomy in Caperea marginata:implications for evolutionary patterning of the mammalian vertebralcolumn. Mar Mammal Sci 27:382–397
Buchholtz EA, Booth AC, Webbnik KE (2007) Vertebral anatomy in theFlorida manatee, Trichechus manatus latirostris: a developmentaland evolutionary analysis. Anat Rec 290:624–637
Buchholtz EA, Wolkovich EM, Cleary RJ (2005) Vertebral osteologyand complexity in Lagenorhynchus acutus (Delphinidae) withcomparison to other delphinoid genera. Mar Mammal Sci 21:411–428
Chittleborough RG (1955) Puberty, physical maturity, and relativegrowth of the female humpback whale, Megaptera nodosa(Bonnaterre) on the Western Australian coast. Aust J Mar FreshRes 6:315–327
Cooper LN, Thewissen JGM, Bajpai S, Tiwari BN (2012) Postcranialmorphology and locomotion of the Eocene raoellid Indohyus(Artiodactyla: Mammalia). Historical Biol 24:279–310
Dawson AB (1925) The age order of epiphyseal union in the long bonesof the albino rat. Anat Rec 31:1–18
J Mammal Evol

Eschricht DF, Reinhardt J, Lilljeborg W (1866) Recent Memoirs on theCetacea. Flower WH (ed), Published for the Ray Society by RobertHardwicke, London
Evans HE (1993) Miller’s Anatomy of the Dog. Third Ed. WB SaundersCompany, Philadelphia
Fish FE (1996) Transitions from drag-based to lift-based propulsion inmammalian aquatic swimming. Am Zool 36:628–641
Fish FE, Peacock JE, Rohr JJ (2000) Stabilization mechanism in swim-ming odontocetes cetaceans by phased movements. Mar MammalSci 19:515–528
Fraas E (1904) Neue Zeuglodonten aus dem unteren Mitteleocän vomMokattam bei Cairo. Geol Palaeontol Abh 6:199–220
Galatius A (2010) Paedomorphosis in two small species of toothedwhales (Odontoceti): how and why? Zool J Linn Soc 99:278–295
Galatius A, Kinze CC (2003) Ankylosis patterns in the postcranialskeleton and hyoid bones of the harbour porpoise (Phocoenaphocoena) in the Baltic and North Sea. Can J Zool 81:1851–1861
Gegenbaur C, Bell FJ (1878) Elements of Comparative Anatomy.Macmillan and Co, London
Geisler JH, Theodor JM (2009) Hippopotamus and whale phylogeny.Nature 458:E1–E4
Geisler JH, Theodor JM, Uhen MD, Foss SE (2007). Phylogeneticrelationships of cetaceans to terrestrial artiodactyls. In: ProtheroDR, Foss SE (eds) The Evolution of Artiodactyls. The JohnsHopkins University Press, Baltimore, pp 19–31
Geisler JH, Uhen MD (2005) Phylogenetic relationships of extinctcetartiodactyls: results of simultaneous analyses of molecular,morphological, and stratigraphic data. J Mammal Evol 12:145–160
Gérard M, Zákány J, Duboule D (1997) Interspecies exchange of a Hoxdenhancer in vivo induces premature transcription and anterior shift ofthe sacrum. Dev Biol 190:32–40
Getty R (1975) Sisson and Grossman’s The Anatomy of the DomesticAnimals. Fifth Ed. WB Saunders Company, Philadelphia
Gingerich PD, Raza SM, Arif M, Anwar M, Zhou X (1994) New whalefrom the Eocene of Pakistan and the origin of cetacean swimming.Nature 368:844–847
Gingerich PD, Ul-Haq, Khan I, Zalmount IS (2001) Eocene stratigraphyand archaeocete whales (Mammalia, Cetacea) of Drug Lahar in theEastern Sulaiman Range, Balochistan (Pakistan). Contrib MusPaleolotol Univ Mich 30:269–319
Graf W, de Waele C, Vidal PP (1995) Functional anatomy of the head-neck movement system of quadrupedal and bipedal mammals. JAnat 186:55–74
Haldiman JT, Tarpley RJ (1993) Anatomy and physiology. In: Burns JJ,Montague JJ, Cowles CJ (eds) The BowheadWhale. The Society ofMarine Mammology Press, Lawrence, Kansas, pp 71–156
Hillson S (1986) Cambridge Manuals in Archaeology- Teeth. CambridgeUniversity Press, London
Hristova GI, Jarzem P, Ouellet JA, Roughley PJ, Epure LM, Antoniou J,Mwale F (2011) Calcification in human intervertebral disc degener-ation and scoliosis. J Orthop Res 29:1888–1895
Ito H, Miyazaki N (1990) Skeletal development of the striped dolphin(Stenella coeruleoalba) in Japanese waters. Mammal Study 14:79–96
Kandel BM, Hullar TE (2010) The relationship of head movements tosemicircular canal size in cetaceans. J Exp Biol 213:1175–1181
Kato H (1988) Ossification pattern of the vertebral epiphyses in thesouthern minke whale. Sci Rep Whales Res Inst 39:11–19
Kellogg R (1936) A Review of the Archaeoceti. Carnegie Institution ofWashington, Washington
Kemper CM, Leppard P (1999) Estimating body length of pygmy rightwhales (Caperea marginata) frommeasurements of the skeleton andbaleen. Mar Mammal Sci 15:683–700
Lessertisseur J, Saban R (1967) Squelette axial. In: Grassé P-P (ed) Traité deZoologie, Anatomie, Systematique, Biologie, Vol. 16.1 Mammifères,Téguments, Squelette. Masson et Cie, Paris, pp 584–708
Lubetkin SC, Zeh JE, George JC (2012) Statistical modeling of baleenand body length at age in bowhead whales (Balaena mysticetus).Can J Zool 90:915–931
Madar SI, Thewissen JGM, Hussain ST (2002) Additional holotyperemains of Ambulocetus natans (Cetacea, Ambulocetidae) and theirimplications for locomotion in early whales. J Vertebr Paleontol 22:405–422
McIntyre DC, Rakshit S, Yallowitz AR, Loken L, Jeannotte L, CapecchiMR, Wellik DM (2007) Hox patterning of the vertebrate rib cage.Development 134:2981–2989
Mead JG, Potter CW (1990) Natural history of the bottlenose dolphinsalong the Central Atlantic Coast of the United States. In:Leatherwood S, Reeves RR (eds) The Bottlenose Dolphin.Academic Press, San Diego, pp 31–43
Messenger SL, McGuire JA (1998) Morphology, molecules, and thephylogenetics of cetaceans. Syst Biol 47:90–124
Moore JC (1968) Relationships among the living genera of beakedwhales with classifications, diagnoses, and keys. Fieldiana: Zool53:209–298
Munro ND, Bar-Oz G, Stutz AJ (2009) Aging mountain gazelle (Gazellagazelle): refining method of tooth eruption and wear and bonefusion. J Archaeol Sci 36:752–763
Narita Y, Kuratani S (2005) Evolution of the vertebral formulae inmammals: a perspective on developmental constraints. J Exp ZoolPart A 304B:91–106.
Nickel R, Schummer A, Seiferle E (1954) Lehrbuch der Anatomie derHaustiere, Vol. I. Bewegungsapparat. Parey P, Berlin, Hamburg,503 pp
Ohsumi SK, Nishiwaki M, Hibiya T (1958) Growth of fin whale in theNorthern Pacific. The Whales Research Institute, No. 13, Japan
Pourquié O (2007) Building the spine: the vertebrate segmentation clock.Cold Sh Q B 72:445–449
Purdue JR (1983) Epiphyseal closure in white-tailed deer. J WildlifeManage 47:1207–1213
Rasband WS (2011) Image J. U. S. National Institutes of Health,Bethesda, Maryland, http://imagej.nih.gov/ij/
Roach HI, Mehta G, Oreffo ROC, Clarke NMP, Cooper C (2003)Temporal analysis of rat growth plate: cessation of growth withage despite presence of a physis. J Histochem Cytochem 51:373–383
Rose KD (1985) Comparative osteology of North American dichobunidartiodactyls. J Paleontol 59:1203–1226
Shashikant CS, Kim CB, Borbély MA,Wang WCH (1998) Comparativestudies on mammalian Hoxc8 early enhancer sequence reveal abaleen whale-specific deletion of a cis acting element. Proc NatlAcad Sci USA 95:15446–15451
Sipla J, Spoor F (2008) The physics and physiology of balance. In:Thewissen JGM, Nummela S (eds) Sensory Evolution on theThreshold, Adaptations in Secondarily Aquatic Vertebrates.University of California Press, Berkeley, pp 227–232
Slijper EJ (1936) Die Cetaceen. A. Asher & Co, AmsterdamSpaulding M, O’Leary MA, Gatesy J (2009) Relationships of Cetacea
(Artiodactyla) among mammals: increased taxon sampling altersinterpretations of key fossils and character evolution. PLoS ONE4:1–14
Spoor F, Bajpai S, Hussain ST, Kumar K, Thewissen JGM (2002).Vestibular evidence for the evolution of aquatic behavior in earlycetaceans. Nature 417:163–166
Spoor F, Thewissen JGM (2008) Balance: comparative and functionalanatomy in aquatic mammals. In: Thewissen JGM, Nummela S(eds) Sensory Evolution on the Threshold, Adaptations inSecondarily Aquatic Vertebrates. University of California Press,Berkeley, pp 257–284
Thewissen JGM, Bajpai S (2009) New skeletal material of Andrewsiphiusand Kutchicetus, two Eocene cetaceans from India. J Paleontol 83:635–663
J Mammal Evol

Thewissen JGM, Cooper LN, Behringer RR (2012) Developmental biol-ogy enriches paleontology. J Vertebr Paleontol 32:1223–1234
Thewissen JGM, Cooper LN, Clementz MT, Bajpai S, Tiwari BN (2007)Whales originated from aquatic artiodactyls in the Eocene epoch ofIndia. Nature 450:1190–1194
Thewissen JGM, Cooper LN, George JC, Bajpai S (2009) From land towater: the origin of whales, dolphins and porpoises. Evolution:Education and Outreach 2 272–288
Thewissen JGM, Heyning J (2007) Embryogenesis and development inStenella attenuata and other cetaceans. In: Jamieson BGM (ed)Reproductive Biology and Phylogeny of Cetacea: Whales,Dolphins, and Porpoises. Science Publishers, New Hampshire, pp307–329
Thewissen JGM, Hussain ST (1990) Postcranial osteology of the mostprimitive artiodactyls Diacodexis pakistanensis (Dichobunidae).Anat Histol Embryol19:37–48
Thewissen JGM, Madar SI (1999) Ankle morphology of the earliestcetaceans and its implications for the phylogenetic relations amongungulates. Syst Biol 48:21–30
Thewissen JGM, Madar SI, Hussain ST (1996) Ambulocetus natans, anEocene cetacean (Mammalia) from Pakistan. Vol. 191 CourierForschungsinstitut Senckenberg
Thewissen JGM, Williams EM, Roe LJ, Hussain ST (2001) Skeletons ofterrestrial cetaceans and the relationship of whales to artiodactyls.Nature 413:277–281
Uhen MD (1999) New protocetid (Mammalia, Cetacea) from the latemiddle Eocene Cook Mountain Formation of Louisiana. J VertebrPaleontol 18:664–668
Uhen MD (2004) Form, function, and anatomy of Dorudon atrox(Mammalia, Cetacea): an archaeocete from the middle to lateEocene of Egypt. Univ Michigan Pap Paleontol 34
Uhen MD, Pyenson ND, Devris TJ, Urbina M, Renne PR (2011) Newmiddle Eocene whales from the Pisco Basin of Peru. J VertebrPaleontol 85:955–969.
Wassersug RA (1976) A procedure for differential staining of cartilage andbone in whole formalin fixed vertebrates. Stain Technol 51:131–134
Wheeler JFG (1930) The Age of Fin Whales at Physical Maturity with aNote onMultiple Ovulatons. Cambridge University Press, Cambridge
Yoshida H, Shirakihara M, Takemura A, Shirokihara K (1994)Development, sexual dimorphism, and individual variation in theskeleton of the finless porpoise, Neophocaena phocaenoides, in thecoastal waters of western Kyushu, Japan. MarMammal Sci 10:266–282
Zákány J, Gérard M, Favier B, Duboule D (1997) Deletion of a HoxDenhancer induces transcriptional heterochrony leading to transposi-tion of the sacrum. EMBO J 16: 4393–4402
Zhou X, Sanders WJ, Gingerich PD (1992) Functional and behavorialimplications of vertebral structure in Pachyaena ossifraga(Mammalia, Mesonychia). Contrib Mus Paleolotol Univ Mich 28:289–319
J Mammal Evol





























