Embed Size (px)
Citation preview

Int. j. Adv. Lif. Sci., Available online on at www. 27
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 27
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Introduction
Microbes used to produce extra-cellular enzymes
that were able to degrade cellulose material into
their smaller components (Bedford and Partridge, 2001).
Natural cellulolysis occurs as combined action of fungi
and bacteria that shift their biomasses depending on the
substrate which is being metabolized (Hu and Van
Bruggen, 1997). Enzyme production is closely controlled
in microorganisms and therefore, to improve its
productivity these controls can be exploited and modified
(Gautam et al., 2011). Compared to fungi the bacterial
cellulase producers are less effective but they produce
enzymes quicker and can be genetically engineered
effectively (Ponnambalam et al., 2011).The production of
cellulase by microbe would bring benefits, such as
promoting rural economy, biomass utilization in an eco-
friendly manner (Himmel et al., 1999) and enhancing
national energy security (Lynd et al., 2008; Zhang
et al., 2008). They have attracted considerable attention
in recent years due to their great biotechnological
and industrial applications, for example, formulation
of washing powder, animal feed production (Han
and He, 2010), textile industry, pulp and paper
industry (Prasetyo et al., 2011), starch processing, grain
alcohol fermentation, malting and brewing, extraction of
fruits and vegetable juices (Bhat, 2000). At present most
Abstract
Cellulase biosynthesis has been established for a variety of bacteria. A novel
bacterium, Bacillus sphaericus JS1 showing production of carboxymethylcellulase
(CMCase) was isolated from the premises of cotton mill using Hirokoshi medium pH
9.5. CMC (1.5% w/v) and gelatin (0.5%, w/v) were recorded as the best carbon and
nitrogen sources for the production of enzyme. The production of CMCase was
maximum when the pH of the media was set at 7.0, inoculated with 12h inoculum of
6% (v/v), aerated at 150 rev/min and incubated at 40oC. Bacillus sphaericus was
found resistant to very few antibiotics (Clindamycin, Bacitracin, Fusidic acid,
Clindamycin, Amikacin, Novabiocin, Oxytetracyclin and Vancomycin) and sensitive
to many on the plates. The Km and Vmax were calculated to be 1.9 mg/ml and 10
µM/min/ml respectively. The outstanding stability of the CMCase from Bacillus
sphaericus JS1 to alkaline pH and high temperature makes this enzyme, a candidate
to be used as an effective additive to laundry detergents.
Keywords: Bacillus sphaericus JS1, carboxymethylcellulase, substrate specificity,
kinetic parameters, carbon source and nitrogen source.
Optimization of nutrient sources and process parameters for the production of carboxymethylcellulase from Bacillus sphaericus JS1
J. Singh Department of Biotechnology, Panjab University, Chandigarh, India
Email : [email protected]
Corresponding Author
J. Singh Department of Biotechnology,
Panjab University, Chandigarh, India

Int. j. Adv. Lif. Sci., Available online on at www. 28
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 28
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
of the advanced countries like United States and some
parts of Europe almost invariably use the detergents
incorporated with cellulases. Bacillus sp. continues to be
dominant bacterial workhorses due to the capacity of
some selected species to produce and secrete large
quantities of extracellular enzymes (Aa et al., 1994;
Ariffin et al., 2006; Mawadza et al., 1996; Rastogi et al.,
2010; Schallmey et al., 2004 and Singh et al., 2004).
Reports available on strains belonging to species such as
Bacillus sphaericus and Bacillus subtilis express high
cellulose degradation activities (Mawadza et al., 1996;
Rastogi et al., 2010; Schallmey et al., 2004; Singh et al.,
2004). To exploit the potentials of biodiversity of Indian
cellulolytic organisms, the current study was undertaken
to screen the potent strain of Bacillus sphaericus JS1 and
optimization of its process parameters for the production
of alkaline CMCase.
Materials and Methods
Dinitrosalicylic acid (DNS) and CMC (Ulta low
viscosity, SD=0.60-0.95) were purchased from Fluka Rdh
(Sigma-Aldrich, Cat. No. 21901). All media components
were of bacteriological grade and purchased from
HiMedia (Mumbai, India). All other chemicals were of
analytical grade and procured from Qualigens Fine
Chemicals (Mumbai, India) and SD Fine Chemicals
(Mumbai, India).
Screening, identification and checking the antibiotic
sensitivity of potent bacterial strain
Soil samples were collected from the premises of
different industries i.e. paper, cotton and wool industries
and screened for alkaliphilic bacteria using congo red
staining method (Beguin and Auber, 1994). The isolation
medium (g/l) contained (Carboxymethyl cellulose 10 g;
peptone 5g; yeast extract 5g; NaCl 5g, KH2PO4 1g
and the pH was adjusted to 9.5 with Na2CO3 (10%,
w/v). Cultures were grown on the same solid medium
(containing 2.0% agar (w/v)) at 40oC in Petri plates. The
isolate producing the maximum zone of hydrolysis was
selected, inoculated into the sterilized liquid medium and
incubated in a rotary shaker (40°C, 150 rev/min). The
samples were withdrawn at regular intervals of time,
centrifuged at 10,000 rev/min at 4°C for 10 min and
supernatant was assayed for enzyme activity. The potent
strain was sent to Microbial Type Culture Collection and
Gene Bank (MTCC-An Internationally depository
Authority), Institute of Microbial Technology,
Chandigarh (India) for identification and was used for
further experimentation.
Effect of antibiotics was studied by pouring an
optimized, sterilized media in the Petri plates and inoculating
them with ten microliter of liquid B. sphaericus culture
(grown over nightly) uniformly. Then, antibiotic discs
(OD 001, OD 002, OD 003, OD 0049) were placed on
them with different combinations and the plates were
incubated. After 24-48 hour of incubation the diameter
around the disc were measured and recorded.
Enzyme assay
The total reducing sugar was determined by
dinitrosalicylic acid reagent (Summer and Somers, 1954).
The yellow colored dinitrosalicylic acid curve of glucose
was made using pure glucose. The diluted enzyme (0.25)
was mixed with 0.25 ml CMC (4%, w/v, in Tris-HCl
buffer pH 8.0) and after incubation for 1h, 0.75 ml
solution of dinitrosalicylic acid was added. The mixture
was then heated in a boiling water bath for 10 minutes.
This was followed by the addition of 1.25 ml buffer and
absorbance was measured at 600nm (Beckman
spectrophotometer DU 640B). Glucose (0-2mg/ml) was
used for the preparation of standard curve. Glucose
concentration was measured in terms of color intensity. A
standard is required to catalyze the formation of 1.0
µM/min/ml of reducing sugar (expressed as glucose).

Int. j. Adv. Lif. Sci., Available online on at www. 29
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 29
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Optimization of nutrient sources and process parameters for production of CMCase
Different carbon sources (arabinose, CMC,
cellobiose, inositol, lactose, maltose, mannose, methyl
cellulose, salicine, starch, sucrose, xylose, glucose,
aesculin, dulcitol, fructose, galactose, glycerol, glycogen,
raffinose, sorbitol, and trehalose) were used at a
concentration of 0.5% (w/v). The pH of the medium was
adjusted to 8.5. The inoculum was prepared by incubating
the flasks in a rotary shaker at 40oC for overnight at 150
rpm. Different sets (in triplicate) of 250 ml Erlenmeyer
flasks each containing 50 ml medium was inoculated with
2.0 % (v/v) inoculum. Samples were withdrawn at regular
intervals of time and enzyme activity was measured.
Different concentrations (0-2%) of selected substrates
were further optimized by following the same steps.
Similarly different organic nitrogen sources (biopeptone,
beaf extract, casein, gelatin, luria broth, malt extract,
nutrient broth, peptone, soyabean casein, soyabean meal,
tryptone and yeast extract) at a concentration of 0.5 and
1.0% (w/v) were optimized. Different concentrations (0-
2%) of the selected nitrogen sources were further
optimized. The effects of pH and temperature on enzyme
activity were observed in a range of 6-10 and 30-55oC
respectively. Cultures of different ages (3-21 h) and
volume (1-10, % v/v) were used as inoculates at different
shaking speeds (0-200 rpm). Samples were withdrawn at
regular intervals of time and relative activity of CMCase
(%) was measured.
Determination of kinetic parameters with purified CMCase
Extracellular CMCase was precipitated with
ammonium sulphate (30–80% saturation) and dialyzed
against Tris-HCl buffer (50 mM, pH 8.0). The dialysate
was applied to a DEAE Sephadex A-50 column (3.6 x
30 cm) and absorbed material was eluted with a linear
gradient of NaCl in the range of 0.05–0.5M. The active
fractions (20–28) were concentrated using a Centrisart
filter (Sartorius, Gottingen,Germany), before loading
onto a Sephadex G-100 column (2.0 x 35 cm). Fractions
(2 ml; 12–18) corresponding to CMCase activity was
checked for purity on native PAGE and this purified
enzyme was subsequently used for characterization
work (Singh et al., 2004). The kinetic parameters like
reaction velocity (Vmax) and kinetic constant (Km) for
purified CMCase were determined using cellulose
modified substrates: carboxymethyl cellulose. For the
determination of Km and Vmax, the Lineweaver-Burk plots
were used under the conditions of 60oC and pH 8.0.
Results and discussion
Based on the morphological, physiological and
biochemical characteristics, the isolate JS1 was identified
as Bacillus sphaericus by the Microbial Type Culture
Collection and Gene Bank (MTCC-An Internationally
depository Authority), Institute of Microbial Technology,
Chandigarh (India) and designated an accession number
of 3243. The resistance to antibiotics was less with
Clindamycin (2 mcg), Bacitracin (10 mcg), Fusidic acid
(10 mcg), but it was much more with Clindamycin
(10 mcg), Amikacin (10 mcg), Novabiocin (30 mcg),
Oxytetracyclin (30 mcg) and Vancomycin (30 mcg).
The Bacillus sphaericus was sensitive to Amoxyclav
(10 mcg), Cephalexin (10 mcg), Ciprofloxacin 10 mcg),
Cloxacillin (1 mcg), Co-trimoxazole (25 mcg), Erythro-
mycin (15 mcg), Tetracycline (30 mcg), Ampicillin (10 mcg),
Carbenicillin (100 mcg), Cephotaxime (30 mcg),
Chloramphenicol (30 mcg), Co-trimazine (25 mcg),
Gentamicin (10 mcg), Norfloxacin (10 mcg), Oxacillin
(5 mcg), Amoxycillin (10 mcg), Cephalothin (30 mcg),
Erythromycin (15 mcg), Ampicillin (1unit), Erythromycin
(10 mcg), Gentamycin (2 mcg), Tetracycline (10 mcg),
Co-trimoxazole (25 mcg), Penicillin-G (25 mcg). Bacillus
sphaericus was found resistant to very few antibiotics and
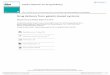
sensitive to many on the plates (Table - 1).
Int. j. Adv. Lif. Sci., Available online on at www. 30
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 30
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Table 1: Effect of antibiotics on the growth of Bacillus sphaericus JS1.
Name of antibiotic Conc. (mcg)
Growth radii (mm)
Name of antibiotic Conc. (mcg)
Growth radii (mm)
OD 001 OD 002
Amoxyclav (Ac) 10 - Ampicillin (A) 10 - Cephalexin (Cp) 10 - Carbenicillin (Cb) 100 - Ciprofloxacin (Cf) 10 - Chloramphenicol (C) 30 - Cloxacillin (Cx) 01 - Cephotexime (Ce) 30 - Co-Trimoprin (Co) 25 - Co-Trimazine (Cm) 25 - Erythromycin (E) 15 - Gentamycin (G) 10 - Tetracycline (T) 30 - Norfloxacin (Nx) 10 - Clindamycin (Cd) 02 13 Oxacillin (Ox) 05 -
OD 0049 - OD 003 Clindamycin (Cd) 10 6.0 Amikacin (Ak) 10 13 Ampicillin (A) 1 unit - Amoxycillin (Am) 10 - Erythromycin (E) 10 - Bacitracin (B) 10 6.0 Gentamycin (G) 02 - Cephalothin (Ch) 30 - Fusidic Acid (Fc) 10 6.0 Erythromycin (E) 15 - Tetracycline (T) 10 - Novobiocin (Nv) 30 13 Co-Trimoxazole (Co) 25 - Oxytetracycline (O) 30 15 Penicillin-G (P) 25 - Vancomycin (Va) 30 10
(-) No growth of Bacillus sphaericus was seen and measured.
Table - 2: Effect of carbon sources on the production of CMCase by Bacillus sphaericus JS1.
Carbon source (0.5%, w/v)
Growth at 600 nm
Enzyme activity µµµµM/min/ml
Carbon source (0.5%, w/v)
Growth at 600 nm
Enzyme activity
µµµµM/min/ml Control 1.6 1.2 Control 1.6 1.2
Aesculin 0.8 0.5 Lactose 4.5 0.7
Arabinose 0.4 0.8 Maltose 0.8 0.2
CMC 3.4 3.3 4.1 4.1
2.8 2.9 3.8 3.9
Mannitol 2.5 0.5
Cellobiose 2.4 1.1 Mannose 0.6 0.2
Cellulose 2.7 2.5 2.8 2.6
2.5 2.7 2.6 2.5
Methyl cellulose 1.7 1.4
Int. j. Adv. Lif. Sci., Available online on at www. 31
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 31
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
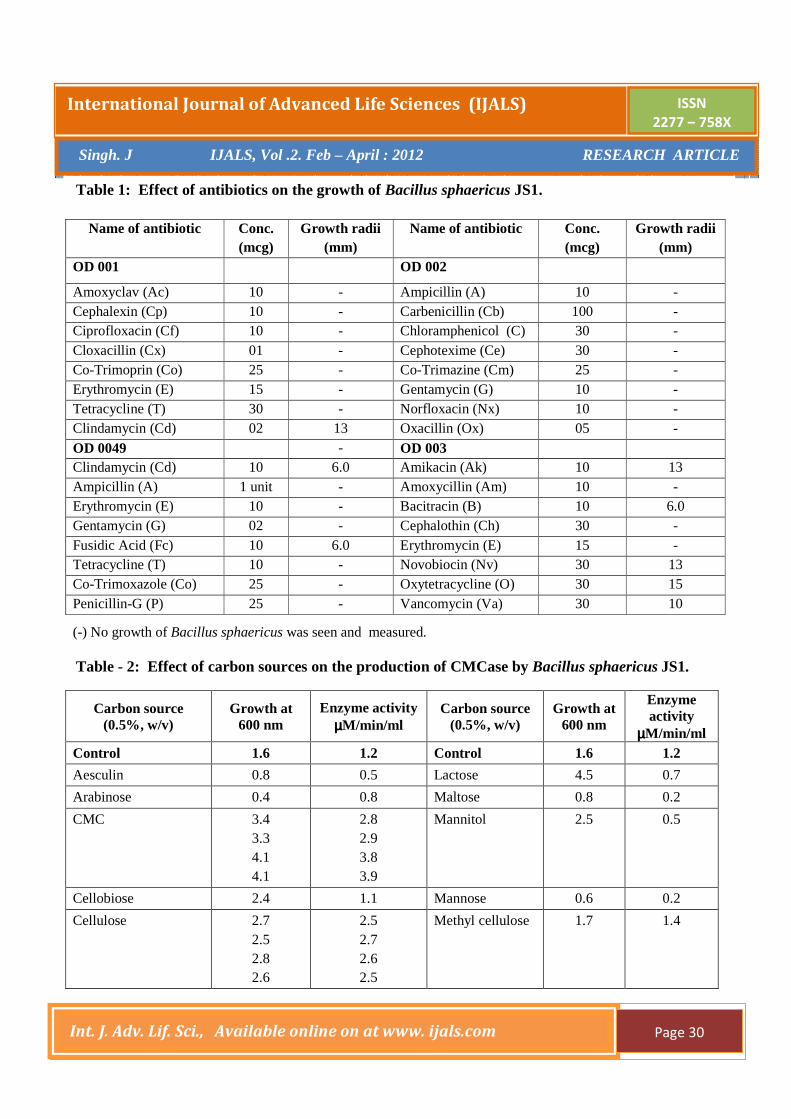
Dulcitol 0.8 1.6 Raffinose 2.7 1.2 Fructose 2.5 1.1 Salicine 2.2 0.2 Galactose 2.6 1.1 Sorbitol 0.7 0.6 Glucose 2.1 0.3 Starch 3.4 1.4 Glycerol 2.2 0.7 Sucrose 2.4 0.1 Glycogen 2.8 2.1
Trehalose 2.9 1.3
Inositol 3.6 0.7 Xylose 4.3 4.2 4.3 3.8
2.7 2.6 2.8 2.0
Table – 3. Effect of organic nitrogen sources on the production of CMCase by Bacillus sphaericus JS1.
Nitrogen source Conc. (%, w/v) Growth at 600 nm Enzyme activity (µµµµM/min/ml)
Control - Control -
Biopeptone 0.5 1.0
1.3 2.3
0.9 1.0
Beaf extract 0.5 1.0
1.2 2.6
1.3 1.8
Casein 0.5 1.0
4.1 4.2
1.2 1.1
Gelatin 0.5 1.0 1.5 2.0
2.0 2.7 2.3 2.8
2.4 2.2 2.3 2.3
Luria broth 0.5 1.0
1.5 1.4
1.4 0.9
Malt extract 0.5 1.0
1.3 2.8
1.3 1.1
Nutrient broth 0.5 1.0
1.3 1.3
1.6 1.2
Peptone 0.5 1.0
1.6 2.6
1.4 1.1
Soyabean casein 0.5 1.0
1.6 2.0
1.3 1.3
Soyabean meal 0.5 1.0
2.9 5.6
1.3 2.3
Trypton 0.5 1.0
1.6 2.9
1.5 1.2
Yeast extract 0.5 1.0
1.2 2.9
0.9 0.6

Int. j. Adv. Lif. Sci., Available online on at www. 32
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 32
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Optimization of carbon and organic nitrogen
sources for the production of CMCase
Effect of carbon source on the production of
CMCase by Bacillus sphaericus was shown in Table - 2.
The rate of production of enzyme was maximum with
CMC, cellulose and xylose than other carbon sources.
Less than 50% of CMCase was observed in media
containing arabinose, cellobiose, inositol, lactose, starch,
aesculin, dulcitol, fructose, galactose, glycerol, raffinose,
sorbitol, trehalose and glycogen. Very low level (less
than 12%) of enzyme activity was seen with maltose,
mannose, glucose, sucrose and salicine. The different
concentrations (0.5, 1.0, 1.5 and 2.0%, w/v) of CMC,
cellulose or xylose was optimized. At both the
concentrations (0.5, 1.0% w/v), production of enzyme
was comparable among three substrates, but beyond this,
the CMC had shown the upper edge. It displayed 150%
of the enzyme activity as compared to xylose and
cellulose at 1.5% (w/v) concentration and reported to be
the best for the production of enzyme by JS1 isolate. All
other concentrations (1.0-2.0%, w/v) displayed the same
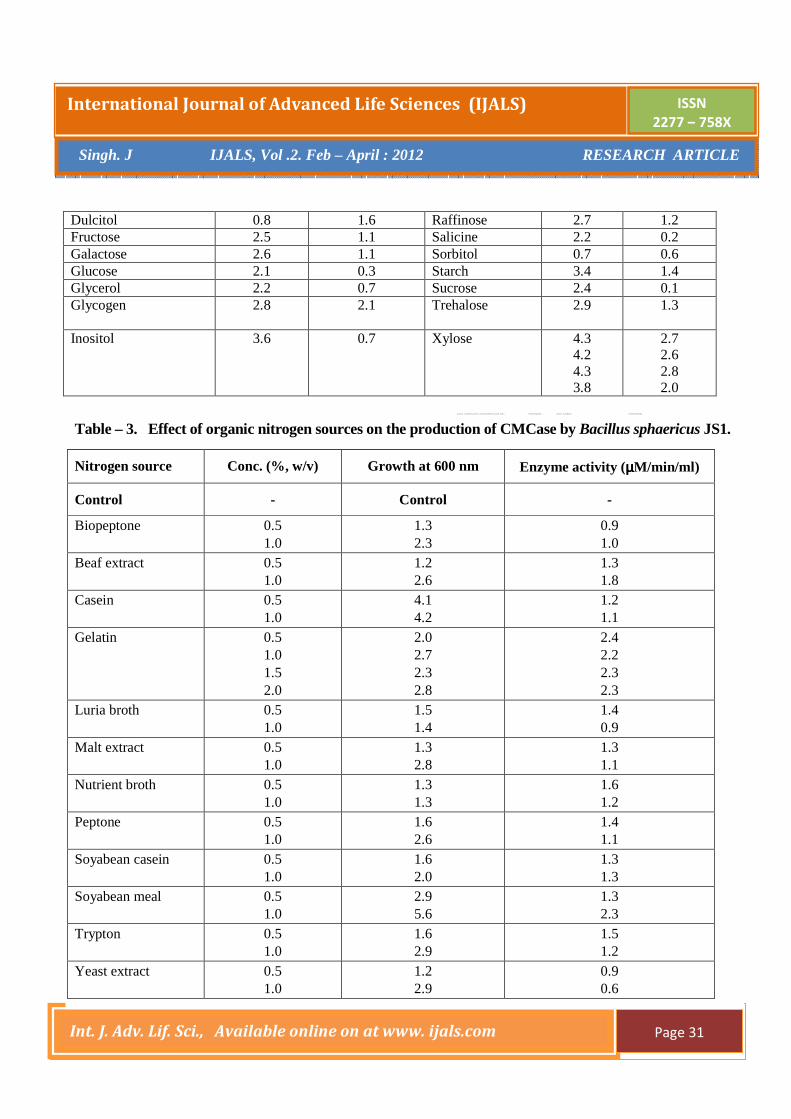
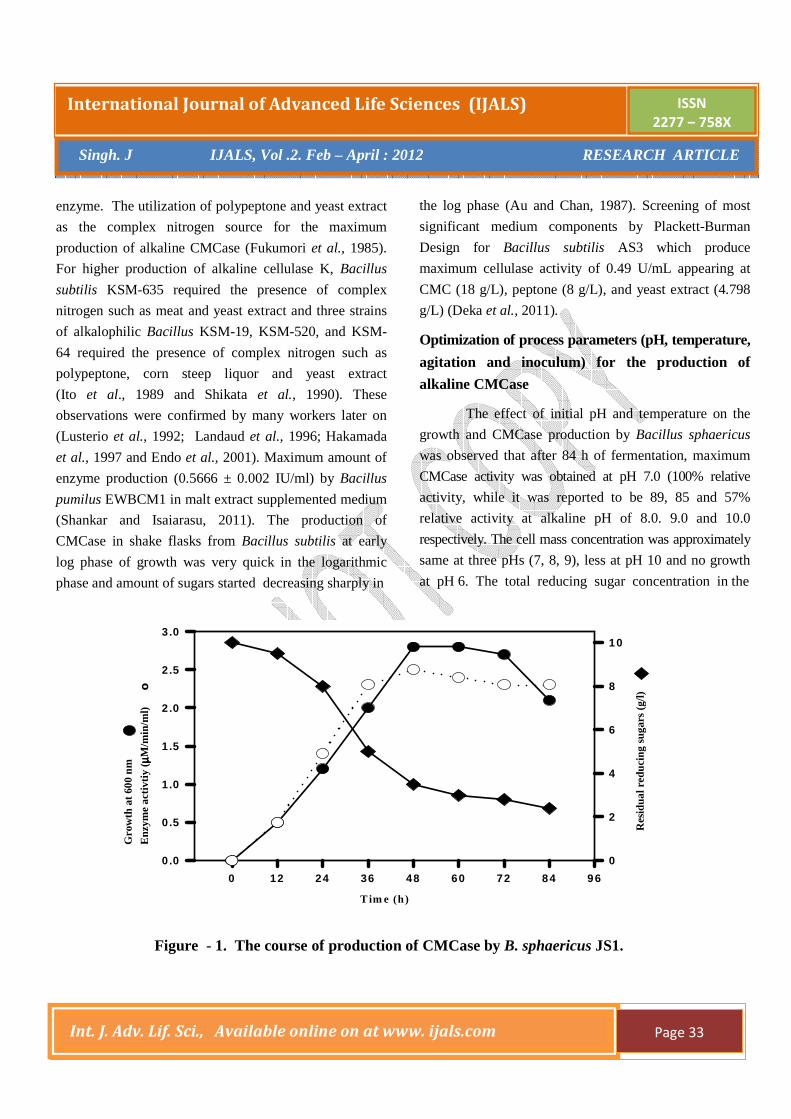
yield. Figure - 1 shows the course of cultivation of B.
sphaericus for the production of CMCase in shake flasks
in the optimized media. In the early hours of fermentation
(12-36h), the active cell growth took place and then
started decreasing. After 12 hours, CMCase activity
started increasing and continued to do so up to 48 h. After
the exponential phase, cell mass concentration started
deceasing and it continued up to 60h of fermentation.
Total residual sugar (as equivalent to glucose)
concentration started deceasing from the very beginning.
Rate of CMC utilization was highest during log phase of
growth. At the end of fermentation, total reducing sugar
utilization was 56% and 12% of initial sugar remained
unutilized, while the enzyme activity showed an
increasing trend. Many authors reported inducible
CMCases while optimizing carbon source. Cellobiose or
CMC (1% w/v) induced CMCase production in
alkalophilic Bacillus sp. No 1139 (Fukumori et al., 1985).
Production of alkaline cellulases required the presence of
CMC (1%) in the strains of alkalophilic Bacillus sp.
KSM-64 and KSM-520 (Shikata et al., 1990). CMCases
from Bacillus sp. KSM-S237 and KSM-N252 were
produced when the media was supplemented with CMC
(Hakamada et al., 1997 and Endo et al., 2001). Alkalophilic
Bacillus circulans synthesized neutral endoglucanases
when grown on N3 medium supplemented with glucose,
fluffy cellulose or wheat bran celluloses (Landaud et al.,
(1996). The highest yield was obtained on wheat bran,
cellobiose, lactose or polysaccharides such as crystalline
cellulose (Avicel, MN 300). Endoglucanase synthesis
was not completely repressed by glucose or cellobiose,
which was generally the case in cellulase producing
microorganisms. The effect of carbon sources on
cellulase production by the candidate species after 72
hours of incubation period at 37°C was observed. The
maximum cellulose production by Bacillus pumilus
EWBCM1 was recorded in galactose (0.5851 ± 0.006
IU/ml) supplemented medium (Shankar and Isaiarasu,
2011).
The production of alkaline CMCase by utilizing
gelatin as the nitrogen source has been reported for the
first time. It displayed 230% of enzyme activity and was
optimized as the best nitrogen source. Enzyme activity
was induced (150% approximately) by glycogen, beaf
extract, casein, luria broth, malt extract, nutrient broth,
peptone, soyabean meal and trypton, while the yeast
extract and biopeptone showed no effect (Table - 3).The
enzyme activity was same (2.3 µM/min/ml) at both the
concentrations (0.5, 1.0% w/v) of gelatin. By studying
enzyme synthesis on different concentrations (0.5, 1.0,
1.5, 2.0%, w/v), 0.5% (w/v) was reported to be the best
for the production of enzyme. Different workers had
reported different nitrogen sources for the production of
Int. j. Adv. Lif. Sci., Available online on at www. 33
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 33
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
enzyme. The utilization of polypeptone and yeast extract
as the complex nitrogen source for the maximum
production of alkaline CMCase (Fukumori et al., 1985).
For higher production of alkaline cellulase K, Bacillus
subtilis KSM-635 required the presence of complex
nitrogen such as meat and yeast extract and three strains
of alkalophilic Bacillus KSM-19, KSM-520, and KSM-
64 required the presence of complex nitrogen such as
polypeptone, corn steep liquor and yeast extract
(Ito et al., 1989 and Shikata et al., 1990). These
observations were confirmed by many workers later on
(Lusterio et al., 1992; Landaud et al., 1996; Hakamada
et al., 1997 and Endo et al., 2001). Maximum amount of
enzyme production (0.5666 ± 0.002 IU/ml) by Bacillus
pumilus EWBCM1 in malt extract supplemented medium
(Shankar and Isaiarasu, 2011). The production of
CMCase in shake flasks from Bacillus subtilis at early
log phase of growth was very quick in the logarithmic
phase and amount of sugars started decreasing sharply in
the log phase (Au and Chan, 1987). Screening of most
significant medium components by Plackett-Burman
Design for Bacillus subtilis AS3 which produce
maximum cellulase activity of 0.49 U/mL appearing at
CMC (18 g/L), peptone (8 g/L), and yeast extract (4.798
g/L) (Deka et al., 2011).
Optimization of process parameters (pH, temperature,
agitation and inoculum) for the production of
alkaline CMCase
The effect of initial pH and temperature on the
growth and CMCase production by Bacillus sphaericus
was observed that after 84 h of fermentation, maximum
CMCase activity was obtained at pH 7.0 (100% relative
activity, while it was reported to be 89, 85 and 57%
relative activity at alkaline pH of 8.0. 9.0 and 10.0
respectively. The cell mass concentration was approximately
same at three pHs (7, 8, 9), less at pH 10 and no growth
at pH 6. The total reducing sugar concentration in the
T im e (h)
0 12 24 36 48 60 72 84 96
Gro
wth
at 6
00 n
m
Enz
yme
activ
tiy ( µµ µµ
M/m
in/m
l)
o
0.0
0.5
1.0
1.5
2.0
2.5
3.0R
esid
ual r
educ
ing
suga
rs (
g/l)
0
2
4
6
8
10
Figure - 1. The course of production of CMCase by B. sphaericus JS1.
Int. j. Adv. Lif. Sci., Available online on at www. 34
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 34
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
culture broth reduced to same level at all pHs (Data not
shown). The maximum enzyme production took place at
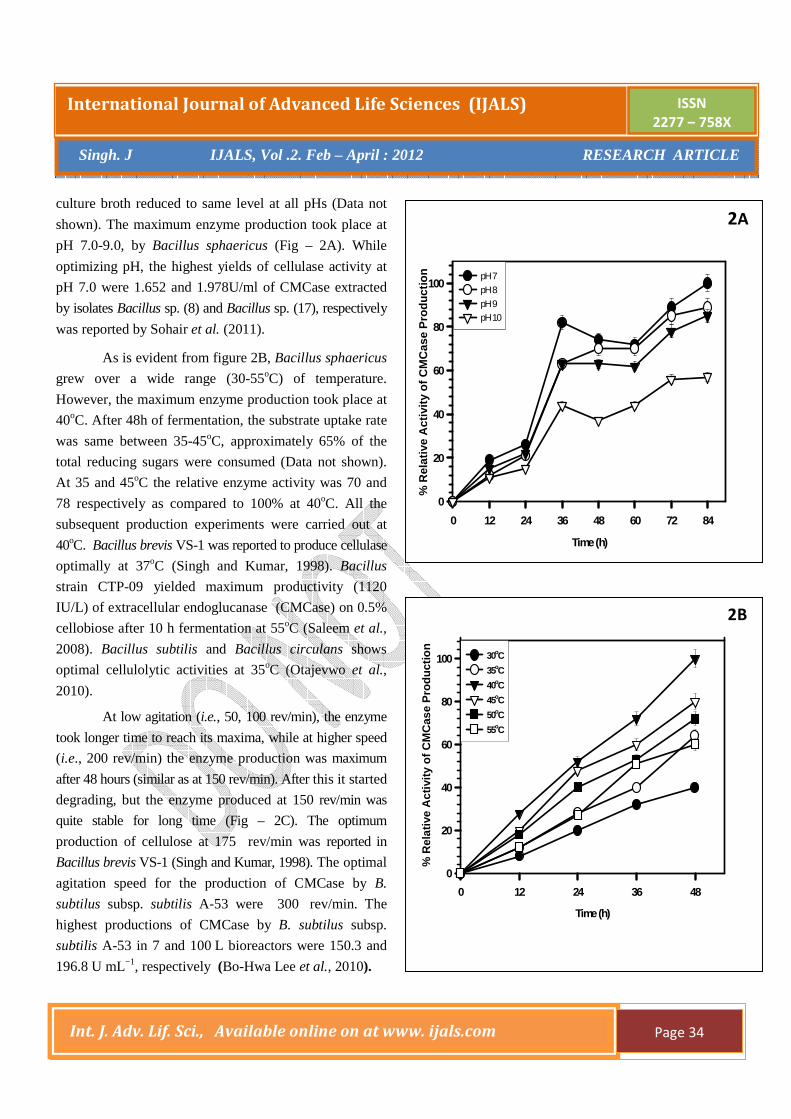
pH 7.0-9.0, by Bacillus sphaericus (Fig – 2A). While
optimizing pH, the highest yields of cellulase activity at
pH 7.0 were 1.652 and 1.978U/ml of CMCase extracted
by isolates Bacillus sp. (8) and Bacillus sp. (17), respectively
was reported by Sohair et al. (2011).
As is evident from figure 2B, Bacillus sphaericus
grew over a wide range (30-55oC) of temperature.
However, the maximum enzyme production took place at
40oC. After 48h of fermentation, the substrate uptake rate
was same between 35-45oC, approximately 65% of the
total reducing sugars were consumed (Data not shown).
At 35 and 45oC the relative enzyme activity was 70 and
78 respectively as compared to 100% at 40oC. All the
subsequent production experiments were carried out at
40oC. Bacillus brevis VS-1 was reported to produce cellulase
optimally at 37oC (Singh and Kumar, 1998). Bacillus
strain CTP-09 yielded maximum productivity (1120
IU/L) of extracellular endoglucanase (CMCase) on 0.5%
cellobiose after 10 h fermentation at 55oC (Saleem et al.,
2008). Bacillus subtilis and Bacillus circulans shows
optimal cellulolytic activities at 35oC (Otajevwo et al.,
2010).
At low agitation (i.e., 50, 100 rev/min), the enzyme
took longer time to reach its maxima, while at higher speed
(i.e., 200 rev/min) the enzyme production was maximum
after 48 hours (similar as at 150 rev/min). After this it started
degrading, but the enzyme produced at 150 rev/min was
quite stable for long time (Fig – 2C). The optimum
production of cellulose at 175 rev/min was reported in
Bacillus brevis VS-1 (Singh and Kumar, 1998). The optimal
agitation speed for the production of CMCase by B.
subtilus subsp. subtilis A-53 were 300 rev/min. The
highest productions of CMCase by B. subtilus subsp.
subtilis A-53 in 7 and 100 L bioreactors were 150.3 and
196.8 U mL−1, respectively (Bo-Hwa Lee et al., 2010).
Time (h)
0 12 24 36 48
% R
elat
ive
Act
ivit
y o
f C
MC
ase
Pro
du
ctio
n
0
20
40
60
80
100 30oC
35oC
40oC
45oC
50oC
55oC
2B
Time (h)
0 12 24 36 48 60 72 84
% R
elat
ive
Act
ivit
y o
f C
MC
ase
Pro
du
ctio
n
0
20
40
60
80
100pH 7
pH 8
pH 9pH 10
2A
Int. j. Adv. Lif. Sci., Available online on at www. 35
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 35
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
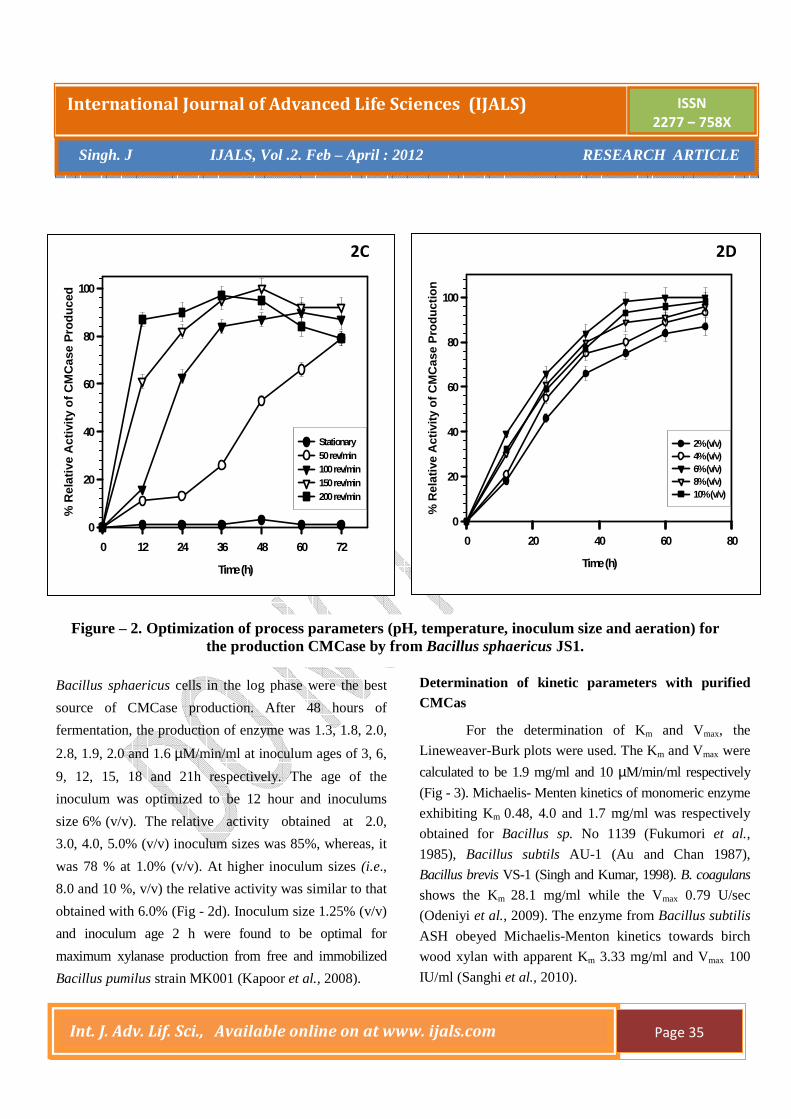
Bacillus sphaericus cells in the log phase were the best
source of CMCase production. After 48 hours of
fermentation, the production of enzyme was 1.3, 1.8, 2.0,
2.8, 1.9, 2.0 and 1.6 µM/min/ml at inoculum ages of 3, 6,
9, 12, 15, 18 and 21h respectively. The age of the
inoculum was optimized to be 12 hour and inoculums
size 6% (v/v). The relative activity obtained at 2.0,
3.0, 4.0, 5.0% (v/v) inoculum sizes was 85%, whereas, it
was 78 % at 1.0% (v/v). At higher inoculum sizes (i.e.,
8.0 and 10 %, v/v) the relative activity was similar to that
obtained with 6.0% (Fig - 2d). Inoculum size 1.25% (v/v)
and inoculum age 2 h were found to be optimal for
maximum xylanase production from free and immobilized
Bacillus pumilus strain MK001 (Kapoor et al., 2008).
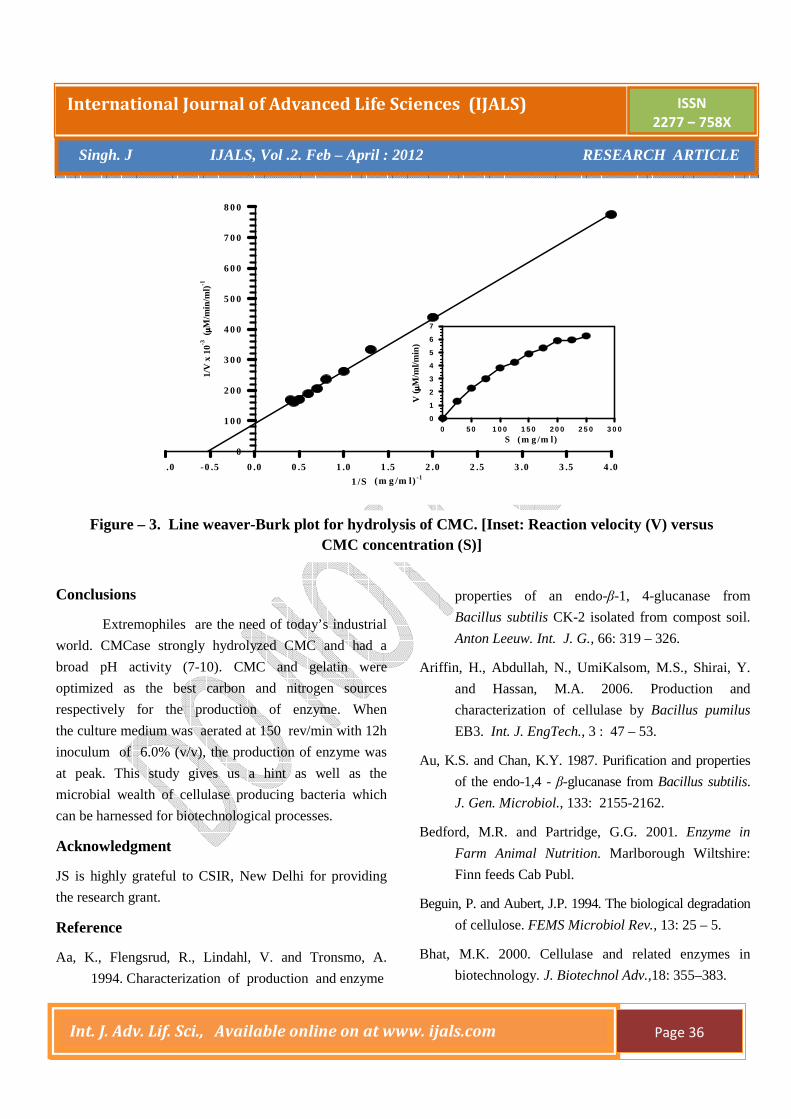
Determination of kinetic parameters with purified CMCas
For the determination of Km and Vmax, the
Lineweaver-Burk plots were used. The Km and Vmax were
calculated to be 1.9 mg/ml and 10 µM/min/ml respectively
(Fig - 3). Michaelis- Menten kinetics of monomeric enzyme
exhibiting Km 0.48, 4.0 and 1.7 mg/ml was respectively
obtained for Bacillus sp. No 1139 (Fukumori et al.,
1985), Bacillus subtils AU-1 (Au and Chan 1987),
Bacillus brevis VS-1 (Singh and Kumar, 1998). B. coagulans
shows the Km 28.1 mg/ml while the Vmax 0.79 U/sec
(Odeniyi et al., 2009). The enzyme from Bacillus subtilis
ASH obeyed Michaelis-Menton kinetics towards birch
wood xylan with apparent Km 3.33 mg/ml and Vmax 100
IU/ml (Sanghi et al., 2010).
Time (h)
0 12 24 36 48 60 72
% R
elat
ive
Act
ivit
y o
f C
MC
ase
Pro
du
ced
0
20
40
60
80
100
Stationary50 rev/min 100 rev/min 150 rev/min 200 rev/min
Time (h)
0 20 40 60 80
% R
elat
ive
Act
ivit
y o
f C
MC
ase
Pro
du
ctio
n
0
20
40
60
80
100
2% (v/v)4% (v/v)6% (v/v)8% (v/v)10% (v/v)
2C 2D
Figure – 2. Optimization of process parameters (pH, temperature, inoculum size and aeration) for the production CMCase by from Bacillus sphaericus JS1.
Int. j. Adv. Lif. Sci., Available online on at www. 36
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 36
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Conclusions
Extremophiles are the need of today’s industrial
world. CMCase strongly hydrolyzed CMC and had a
broad pH activity (7-10). CMC and gelatin were
optimized as the best carbon and nitrogen sources
respectively for the production of enzyme. When
the culture medium was aerated at 150 rev/min with 12h
inoculum of 6.0% (v/v), the production of enzyme was
at peak. This study gives us a hint as well as the
microbial wealth of cellulase producing bacteria which
can be harnessed for biotechnological processes.
Acknowledgment
JS is highly grateful to CSIR, New Delhi for providing
the research grant.
Reference
Aa, K., Flengsrud, R., Lindahl, V. and Tronsmo, A.
1994. Characterization of production and enzyme
properties of an endo-β-1, 4-glucanase from
Bacillus subtilis CK-2 isolated from compost soil.
Anton Leeuw. Int. J. G., 66: 319 – 326.
Ariffin, H., Abdullah, N., UmiKalsom, M.S., Shirai, Y.
and Hassan, M.A. 2006. Production and
characterization of cellulase by Bacillus pumilus
EB3. Int. J. EngTech., 3 : 47 – 53.
Au, K.S. and Chan, K.Y. 1987. Purification and properties
of the endo-1,4 - β-glucanase from Bacillus subtilis.
J. Gen. Microbiol., 133: 2155-2162.
Bedford, M.R. and Partridge, G.G. 2001. Enzyme in
Farm Animal Nutrition. Marlborough Wiltshire:
Finn feeds Cab Publ.
Beguin, P. and Aubert, J.P. 1994. The biological degradation
of cellulose. FEMS Microbiol Rev., 13: 25 – 5.
Bhat, M.K. 2000. Cellulase and related enzymes in
biotechnology. J. Biotechnol Adv.,18: 355–383.
1 /S (m g /m l) -1
-1 .0 -0 .5 0 .0 0 .5 1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0
1/V
x 1
0-3 ( µµ µµ
M/m
in/m
l)-1
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
6 0 0
7 0 0
8 0 0
S (m g /m l)0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 3 0 0
V (
µµ µµM/m
l/min
)0
1
2
3
4
5
6
7
Figure – 3. Line weaver-Burk plot for hydrolysis of CMC. [Inset: Reaction velocity (V) versus CMC concentration (S)]

Int. j. Adv. Lif. Sci., Available online on at www. 37
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 37
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Bo-Hwa, L., Bo-Kyung, K., You- Jung, L., Chung-Han,
C. and Jin-Woo, L. 2010. Industrial scale of
optimization for the production of carboxymethyl
cellulase from rice bran by a marine bacterium,
Bacillus subtilis subsp. subtilis A-53. Enzyme
Microbial Technol., 46 : 38-42.
Deka, D., Bhargavi, P., Sharma, A., Goyal, D., Jawed, M.
and Goyal, A. 2011. Enhancement of Cellulase
with Various Cellulosic Substrates Activity from a
New Strain of Bacillus subtilis by Medium Optimization
and Analysis. Enzyme Res., 8.
Endo, K., Hakamada, Y., Takizawa, S., Kubota, H.,
Sumitomo, N. and Kobayashi, T. 2001. A novel
alkaline endoglucanase from an alkalophilic Bacillus
isolate: Enzymatic properties, and nucleotide and
deduced amino acid sequences. App. Microb.
Biotech., 57: 109-116.
Fukumori, F., Kudo, T. and Horikoshi, K. 1985. Purification
and properties of a cellulose formal kalophilic
Bacillus sp. No. 1139. J Gen. Microbiol.,131 : 2239
- 3345.
Gautam, S.P., Bundela, P.S., Pandey, A.K., Khan, J.,
Awasthi, M.K. and Sarsaiya, S. 2011. Optimization
for the production of cellulaseenzyme from municipal
solid waste residue by two novel cellulolytic fungi.
Biotech. Res Int., 2011 : 8.
Hakamada, Y., Koike, K., Yoshimatsu, T., Mori, H.,
Kobayashi, T. and Ito, S. 1997. Thermostable alkaline
cellulases from an alkaliphilic isolate, Bacillus sp.
KSM - S237. Extremophil. ,1: 151-156.
Han, W. and He, M. 2010. The application of exogenous
cellulase to improve soil fertility and plant growth
due to acceleration of straw composition. Bioresour
Technol.,101: 3724–3731.
Himmel, M.E., Ruth, M.F. and Wyman, C.E. 1999.
Cellulase for commodity products from cellulosic
biomass. Curr Opin Biotechnol., 10: 358 – 364.
Hu, S. and Van Bruggen, A.H.C. 1997. Microbial
dynamics associated with multiphasic decomposition
of 14C-labelled cellulose in soil. Microb. Ecol., 33:
134 - 143.
Ito, S. 1997. Alkaline cellulase from alkaliphilic Bacillus:
Enzymatic properties, genetics, and application to
detergents. Extremophil.,1: 61-66.
Kapoor, M., Nair, L.M. and Kuhad, R.C. 2008. Cost-
effective xylanase production from free and
immobilized Bacillus pumilus strain MK001 and its
application in saccharification of Prosopisjuliflora.
J. Biochem. Eng., 38: 88–97.
Landaud, S., Davila, A.M. and Pourquie, J. 1996. Neutral
endoglucanases production by a newly isolated
alkalophilic Bacillus circulans. Biotech Let., 18:
741 - 746.
Lusterio, D.D., Suizo, F.G., Labunos, N.M., Valledor,
M.N., Ureda, S. and Kawai, S. 1992. Alkali-
resistant, alkaline endo-β-glucanase produced by
Bacillus sp. PKM-5430. Biosci Biotech Biochem.,
56: 1671-1672.
Lynd, L.R., Laser, M.S., Bransby, D., Dale, B.E.,
Davison, B., Hamilton, R., Himmel, M., Keller, M.,
McMillan, J.D., Sheehan, J. and Wyman, C.E.
2008. How biotech can transform biofuels. Nat
Biotechnol., 26 : 169 – 172.
Mawadza, C., Boogerd, F.C., Zvauya, R. and Van
Verseveld, H.W. 1996. Influence of environmental
factors on endo-β- 1,4-glucanase production by
Bacillus HR 68, isolated from a Zimbabwean hot
spring. Anton Leeuw. Int J. G., 69 : 363–369.

Int. j. Adv. Lif. Sci., Available online on at www. 38
Int. J. Adv. Lif. Sci., Available online on at www. ijals.com
Page 38
International Journal of Advanced Life Sciences (IJALS) ISSN
2277 – 758X
Singh. J IJALS, Vol .2. Feb – April : 2012 RESEARCH ARTICLE
Odeniyi, O.A., Onilude, A.A. and Ayodele, M.A. 2009.
Production characteristics and properties of
cellulose polygalacturonase by a Bacillus coagulans
strain from a fermenting palm-fruit industrial
residue. Afr. J. Microbiol. Res., 3: 407 - 417.
Otajevwo, F.D. and Aluyi, H.S. 2010. Cultural conditions
necessary for optimal cellulose yield by cellulolytic
bacterial organisms as they relate to residual sugars
released in broth medium. Nigerian J. Microbiol.,
24: 2168 – 2182.
Ponnambalam, A.S., Deepthi, R.S. and Ghosh, A.R.
2011. Qualitative display and measurement of
enzyme activity of isolated cellulolytic bacteria.
J. Biotech. Bioinf. Bioeng.,1:33-37.
Prasetyo, J., Zhu, J., Kato, T. and Park, E.Y. 2011.
Efficient production of cellulase in the culture of
Acremonium cellulolyticus using untreated waste
paper sludge. Biotechnol. Prog., 7: 104 – 110.
Rastogi, G., Bhalla, A., Adhikari, A., Bischoff, K.M.,
Hughes, S.R., Christopher, L.P., Sani, R.K.2010.
Characterization of thermo stable cellulases produced
by Bacillus and Geobacillus strains. Bioresource
Technol., 101: 8798 – 8806.
Saleem, M., Akhtar, M.S., Yasmin, R., Zahid, M., Malik,
N.N., Afzal, M. 2008. Production, purification and
characterization of beta-1,4 - endoglucanase from a
novel bacterial strain CTP-09 of a Bacillus sp.
Protein Pept. Lett., 15: 402 - 10.
Sanghi, A., Garg, N., Gupta, V.K., Mittalk, A., Kuhad,
R.C. 2010. One-Step Purification and Characterization
of Cellulase Free Xylanase Produced by Alkalophilic
Bacillus subtilis ASH. Braz J. Microbiol., 4: 467-476.
Schallmey, M., Singh, A. and Ward, O.P. 2004. Develop-
ments in the use of Bacillus species for industrial
production. Can. J. Microbiol., 50: 1 – 17.
Shikata, S., Saeki, K., Okoshi, H., Yoshimatsu, T., Ozaki,
K. and Kawai, S. 1990. Alkaline cellulase laundry
detergents: Production by alkalophilic strains of
Bacillus and some properties of the crude enzymes.
Agri. Biol. Chem., 54: 91 - 96.
Shankar, T. and Isaiarasu, L. 2011.Cellulase Production
by Bacillus pumilus EWBCM1 under Varying
Cultural Conditions. Middle East J. Sci. Res., 8:
40 - 45.
Singh, V.K. and Kumar, A. 1998. Production and
purification of an extracellular cellulose From
Bacillus brevis VS-1. Biochem. Mol. Biol. Int., 45:
443 - 452.
Singh, J., Batra, N. and Sobti, R.C. 2004. Purification and
characterization of alkaline cellulose produced by a
novel isolate Bacillus sphaericus JS1. J. Ind.
Microb. Biotech., 31: 51 - 56.
Sohair, A.N., Abozaid, A.A., Hussein, N.A. and Al-
salem, F.A. 2011. Cellulase production by local
bacteria isolated from Taif in Saudi Arabia.
J. Agric. Sci., 19: 163-170.
Summer, J.B. and Somers, G.F. 1954. Dinitrosalicylic
acid for glucose. In: Laboratory experiments in
Biological Chemistry. Academic Press, New York,
PP. 34–39.
Zhang, Y.H.P. 2008. Reviving the carbohydrate economy
via multiproduct biorefineries. Ind. J. Microb.
Biotech., 35 : 367–37.
Corresponding Author : J. Singh, Department of Biotechnology, Panjab University, Chandigarh, India, Email - [email protected].