Embed Size (px)
Citation preview

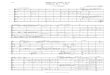
FE ring height above Phosphates
1.25
0.75
0.25
Radial distance from fengycin (Å)
Phosp
hate
heig
hts
rela
tive t
o F
E r
ing (Å
)
20 40 60
Radial distance from fengycin (Å)
Diff
ere
nce
in t
hic
kness
betw
een
upper
and low
erl
eaflets
(Å
)
1
0.5
0
DPPCPOPE/POPGPOPC
20 40 60
DPPCPOPE/POPGPOPC
-80 -40 0 40 80
Aaron Cravens, Joshua N. Horn, Alan GrossfieldUniversity of Rochester Medical School, Rochester, NY, USA
Fengycin
http: / / l oos. sourceforge. net
For reference, use the code to the left to access a digitalcopy of this poster. To learn more about this work or tocontact the authors, visit us at:
http: / / membrane. urmc. rochester. edu
Lightw
eight Object Oriented StructureAnalysis
G rossfield L ab
INTERACTIONS OF THE ANTIFUNGAL FENGYCIN WITH MODELBIOMEMBRANES CHARACTERIZED USING MOLECULAR SIMULATION
INTERACTIONS OF THE ANTIFUNGAL FENGYCIN WITH MODEL BIOMEMBRANES CHARACTERIZED USING MOLECULAR SIMULATION
Aaron Cravens, Joshua N. Horn, Alan GrossfieldUniversity of Rochester Medical School, Rochester, NY, USA
Simulation Details Clustering of Fengycin Varies Between Bilayer Types
Conclusions
Abstract
Coarse-Grained Modeling
•Potent anti-fungal agent - acts by disrupting outer membrane - little toxicity towards mammalian and bacterial cells - hypothesized to make pores at moderate concentrations and dissolve bilayers at high conc.•Structural characteristics - net charge equals -2 - amphipathic molecule with 10 amino acid cyclic peptide and 14 carbon tail - hydrophobic tail promotes insertion into lipid bilayers
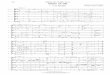
•Analysis of FE clusters - criteria for cluster was at least 2 contacts at a distance less than 7Å•Discreet data shown right - data was combined from 16 trajectories with 13 fengycin inserted each•Each bilayer had distinct ordering of fengycin•In POPC fengycin remains largerly clustered•In POPE/POPG fengycin tends to disperse
•Trade off between speed and detail•MARTINI forcefield used - parametized to reproduce thermodynamic properties - primarily used with lipids and proteins - maps approximately four heavy atoms to one CG (coarse-grained) 'bead' - simulation speeds 100x faster than AA (all-atom) due to fewer atoms - 20fs time step•CG fengycin - 29 'bead' model
FE Has Distinct Ring States
Model supports hypothesis that fengycin acts through bilayer disruption. Fengycin clusters in POPC and disperses in POPE/POPG and DPPC.•DPPC and POPC - neutral lipids - fengycin appears to has affinity for self-self interaction•POPE/POPG - preferential interactions, or at least not disfavorable, between fengycin and lipids results in the diffusion of fengycin into bilayer•Lack of distinct ring conformation of single fengycin in each bilayer type - perhaps ring-lipid interactions are not as important
•Test whether the fengycin clustering properties in POPC are a result of the FA tail length, fengycin ring structure (specifically charged beads) or a combination of the two - new coarse-grained fengycin lacking charged ring beads - new coarse-grained fengycin with additional/decreased carbons on the FA tail - hold other parameters constant and evaluate clustering behavior to see dependency on these factors
FE Binding Bends Membrane•Heat map of phosphate heights in the upper and lower bilayer leaflets - representative image of PO4 curvature when 1 FE is inserted in DPPC, POPC, or POPE/POPG bilayers - curvature occurs in the upper and lower leaflets where fengycin is inserted - no average curvature in neat bilayer
•Difference between upper and lower leaflet RDFs (radial distribution function) of phosphate heights - XY centered about 1 fengycin - Z of bilayer centered•All three bilayer types have a similar relative displacement of phosphate with one fengycin inserted
•Location of phosphates relative to height of fengycin ring - centered about 1 fengycin•Fengycin ring location differs in each bilayer - in POPC FE ring is inserted further - in POPE/POPG and DPPC FE ring sits atop lipid heads
POPC Lowers FE Ring Relativeto DPPC and POPE/POPG
0.009
0.006
0.003
0.009
0.006
0.003
0.009
0.006
0.003
Torsions Angle (degrees)
Average number of fengycin clusters
•Torsion Calculations - used Ala-6, Pro-7, Ile-10, and Tyr-3 ring beads (highlighted purple in fengycin introduction) - 13 FE calculation run after 400ns equilibration•Fengycin-fengycin contacts modulate ring structure in two possible ways - slowing of ring transition due to stabilization of the transition state - increased oscillation btw. the two main ring states•Free fengycin in each bilayer has similar torsion angle distribution. - other torsions remain the same between bilayers - ring-lipid interactions are similar between the three bilayer types? - fengycin ring might not determine if clustering or dispersal occurs
Analysis done using LOOS (Lightweight Object Oriented Structure analysis library), an open source C++ library designed and maintained by the Grossfield lab. LOOS provides a framework for designing analysistools that interface with file formats of most simulation packages.
5
3
1
5
3
1
5
3
1
200 400 600 800 1000 Time (ns)
200 400 600 800 1000 Time (ns)
200 400 600 800 1000 Time (ns)
Time (ns)
Num
ber
of
fengyci
n c
lust
ers
Num
ber
of
fengyci
n c
lust
ers
With the advent of many immuno-compromising medicines as well as drug-resistant strains of fungi there is an increasing need for new classes of antifungal compounds. The lipopeptide fengycin (FE), produced by Bacillus subtilis, has demonstrated potent antifungal activity while having little inhibitory effect towards mammalian cells and bacteria. Experiment indicates fengycin acts as a toxic agent via cytoplasmic membrane disruption, resulting in permeabilization of the cell membrane. Here we explore this hypothesis using coarse-grained molecular dynamics simulations. While the insertion of multiple fengycins into our model bilayers causes notable thinning there is no significant difference in average thinning between the three bilayer types. This is likely due to fengycin clustering in the different types of membranes. Membranes with experimental susceptibility to fengycin demonstrated more clustering in our simulations. We hypothesize that fengycin-resistant membranes disperse fengycin about the membrane eliminating radical pockets of curvature.
AA CG
DPPCPOPE/POPGPOPC
DPPC POPE/POPG POPC
DPPC
POPE/POPG
POPC
Membrane plane (Å)
Dis
tance
fro
m m
em
bra
ne c
ente
r (Å
)
1 Fengycin in DPPC
Up
per
Leafl
et
Low
er
Leafl
et
Pure DPPC
-40 -20 0 20 40 -40 -20 0 20 40Membrane plane (Å)
-40
-20
0
20
40
-40
-20
0
20
40Mem
bra
ne p
lane (
Å)
Mem
bra
ne p
lane (
Å)
22
21
20
19
18-18
-19
-20
-21
-22
Probabili
tyPr
obabili
tyPr
obabili
ty
•Simulations carried out in physiologic salt (100mM NaCl) concentration plus neutralizing salt for fengycin/lipid charges - DPPC, POPC, POPE/POPG (2:1) membrane models were used - 256 lipids per leaflet •Pure bilayer, 1 fengycin, and 13 fengycin systems constructed•8,000-12,000 waters and 15,000-20,000 total 'atoms' per system•16 replicas per system type for a total of 144 trajectories - each trajectory at >1us
charge temperature saturated tails model DPPC 0 323K 2 NAPOPE 0 300K 1 bacterialPOPG -1 300K 1 POPC 0 300K 1 fungal
ON
H
OH
O
O
O
O
HNO
N
O
HN
O
OHN
O
HO
O
H
N
O
O
NH
N
H
OHN
NH3
O
O
O
NH
O
HO
D-allo-Thr-4
L-Glu-5
D-Ala-6
L-Pro-7
L-Glu-8
D-Tyr-9
L-Ile-10
L-Tyr-3
L-Glu-1
D-Orn-2
FA
DPPC
POPE/POPG
POPC
Future Directions
Phosphate-height RDF about 1 FE
![1-Aryl-3-[4-(thieno[3,2-d]pyrimidin-4-yloxy)phenyl]ureas ... · 2 BioMedResearchInternational N H O F HN N H N O H O N O HN O NH Cl F F F Sunitinib Sorafenib N N N N N NH O S O Pazopanib](https://img.pdfslide.us/doc/110x75/5f6429458f126977bb11bd13/1-aryl-3-4-thieno32-dpyrimidin-4-yloxyphenylureas-2-biomedresearchinternational.jpg)

















![2006 DOE Hydrogen Program Review Presentation · 2020. 11. 21. · HN O SO3 HOH O O [ ] HN AIBN SO OH + O O O O SO3H O HN Synthetic steps leading to a photo-crosslinkable sulfonic](https://img.pdfslide.us/doc/110x75/612f34bf1ecc515869434b3b/2006-doe-hydrogen-program-review-presentation-2020-11-21-hn-o-so3-hoh-o-o-.jpg)








