Embed Size (px)
Citation preview

The Prostate 11:23-31 (1987)
Interaction of Sex Steroids and Prolactin on Phosphatases, Transaminases, and Citric Acid in the Ventral Prostate of Male Albino Rats N. Srinivasan, M. Michael Aruldhas, and P. Govindarajulu
Department of Endocrinology, P. G. Institute of Basic Medical Sciences, University of Madras, Taramani, Madras, India
The interaction of androgens, testosterone propionate (TP), and dihydrotestosterone (DHT) with prolactin (PRL) and estradiol-17fl (E$) on phosphatases, transaminases, and citric acid were studied in the ventral prostate of prepubertal and adult intact and castrated rats. None of the secretory products studied were affected by androgens or their combinations with E$/ PRL in prepubertal rats except for the increase observed in organ weight and DNA content. However, in adults the hormonal combinations produced some significant changes. Among the secretory products, phosphatases, glutamate oxaloacetic transaminase (GOT), and citric acid increased significantly in adult rats after the administration of TP/DHT. Glutamate pyruvic transaminase (GF'T) required PRL along with TP/DHT to show an appreciable increment in intact adult rats. The synergistic effect of PRL with androgens was observed in the adult ventral prostatic secretory acid phosphatase and citric acid. Unlike PRL, in combination with androgens produced an antagonistic effect in intact adult rats, which was well pronounced in the case of prostatic weight, DNA content secretory acid phosphatase, and citric acid. None of the androgens or their combination with PRL produced any significant change in DNA content compared to intact controls.
Key words: ventral prostate, acid phosphatase, glutamate oxaloacetic transaminase, glutamate pyruvic transaminase, citric acid, testosterone propionate, dihydrotestosterone, estra- diol, prolactin
INTRODUCTION
Testosterone and its active metabolite, dihydrotestosterone, are the major regu- lators of the structure and function of rat prostate [1,2,3]. Apart from these andro- genic steroids, estrogens were also implicated as regulators of the male accessory sex organs [4]. The androgen-estrogen antagonism or synergism on male accessory sex organs is a matter of dispute [4-91. Unlike estrogens, the synergistic effect of prolactin with androgens in the execution of structural and functional activities of male accessory sex organs is well established [lo, 111.
Acid phosphatases and citric acid serve as the markers for androgen action and secretory function of the rat prostate [ 12-15]. Secretory acid phosphatase, a tartrate- resistant enzyme, is specific to the prostate.
It has been postulated that transamination of aspartate to oxaloacetate is andro- gen dependent [16] where oxaloacetate serves as a source for citrate formation in the rat ventral prostate [17]. To our knowledge, the studies so far available on the interaction of androgens, estradiol, and prolactin are confined to adult rats. Since the
Received April 8, 1986; accepted April 3, 1987. Address reprint requests to Dr. N. Srinivasan, Department of Endocrinology, P.G. Institute of Basic Medical Sciences, University of Madras, Taramani, Madras 600 113, India.
0 1987 Alan R. Liss, Inc.

24 Srinivasan, Aruldhas, and Govindarajulu
status of the prostatic tissue and age of the animals may interfere with hormone actions, we have considered both intact as well as castrated prepubertal and adult rats for the present investigation. To establish the positive or negative interaction of these hormones, we have studied a few androgen sensitive parameters such as deoxyribose nucleic acid (DNA), phosphatases, transaminases, and citric acid.
MATERIALS AND METHODS Experimental Animals
Healthy male albino rats of Wistar strain were housed in a well-ventilated and humidity controlled animal room at a temperature of 25 k 2°C with 14-hour light and 10-hour dark schedule. These rats were fed on a standard pelleted diet (Gold Mohur, Hindustan Lever, India) and provided with drinking water ad libitum.
The rats were divided into prepubertal (25 days old) and adult (120 days old) age groups consisting of both intact and castrated animals. The rats were anesthetized with light ether and castrated by the transcortal route. Hormone treatments in the castrated animals were started 7 days postsurgery.
Depending on the treatment schedule, the animals were divided into 6 subgroups:
Subgroup I:
Subgroup 11:
Subgroup 111:
Received daily injections of testosterone propionate (TP) alone for 21 days. Dosage: 1 mg TP/prepubertal rat, 2 mg TP/adult rat. Received daily injections of TP + & for 21 days. Dosage: 1 mg TP + 10 pg E2/prepubertal rat, 2 mg TP + 20 pg E,/adult rat. Received daily injections of TP + ovine prolactin (PRL) for 21 days. Dosage: 1 mg TP + 250 pg PRL/prepubertal rat, 2 mg TP + 500 pg PRL/adult rat. Received daily injection of dihydrotestosterone (DHT) alone for 21 days. Dosage: 1 mg DHT/prepubertal, 2 mg DHT/adult rat. Received daily injections of DHT + E2 for 21 days. Dosage: 1 mg DHT + 10 pg E2/prepubertal rat, 2 mg DHT + 20 pg E2/ adult rat. Received daily injections of DHT + PRL for 21 days. Dosage: 1 mg DHT + 250 pg PRL/prepubertal rat, 2 mg DHT + 500 pg PRL/adult rat.
Subgroup IV:
Subgroup V:
Subgroup VI:
Each subgroup consisted of 60 intact (30 prepubertals and 30 adults) and 80 castrated (40 prepubertals and 40 adults) rats. The specified hormone treatment was given to 30 rats (15 intact and 15 castrates) from each age group, and an equal number of age-matched controls received the vehicle alone. Steroid hormones were dissolved in propane 1,2-diol, and PRL was dissolved in 0.02 M sodium hydroxide. Each hormone was injected i.p. separately at 0900 hours. Twenty-four hours after the last injection, the rats were sacrificed by decapitation. Ventral prostates were resected out and freed of external fascia. The tissues were weighed and stored at -20°C until used.
Extraction of the Enzyme From the Prostatic Tissue The tissues were homogenized in cold 0.25 M sucrose using a Potter-Elvejhem
homogenizer. The homogenates were frozen, thawed twice, and centrifuged at 1 ,OOO

Sex Steroids and Prolactin on Phosphatases 25
x g for 10 minutes at 4°C. The supernatant was filtered and the filtrate was used as the source of enzyme and was frozen at -75°C until use. Analytical Methods
DNA. Quantification of DNA was performed according to the method of Burton [ 181.
Acid phosphatase (EC 3.1.3.2). Total acid phosphatase activity was estimated following the procedure of Tenniswood et al [ 151, a modified method of Bessey et a1 [ 191, using p-nitrophenyl phosphate as the substrate. In the quantification of secretory acid phosphatase, tartrate buffer (0.2 M pH 4.8) was used instead of citrate buffer.
Transaminases. Glutamate oxaloacetic transaminase (GOT) (EC 2.6.1.1) and glutamate pyruvic transaminase (GPT) (EC 2.6.1.2) were assayed following the method of Reitman and Frankal[20].
Citric acid. The method adopted was essentially that of Beutler and Yeh [21]. All chemicals and reagents used were of analytical grade (Sigma Chemical Company, St. Louis, MO).
Statistical analysis. The data were statistically analyzed using one-way analy- sis of variance adopted by Sokal and Rohlf [22]. When the ‘F’ test was significant, multiple comparisons were made using Student Newmans-Keul’s test (SNK) accord- ing to the method of Zar [23]. The significance was calculated at the 0.05 level.
RESULTS Organ Weights and DNA Content (Table I)
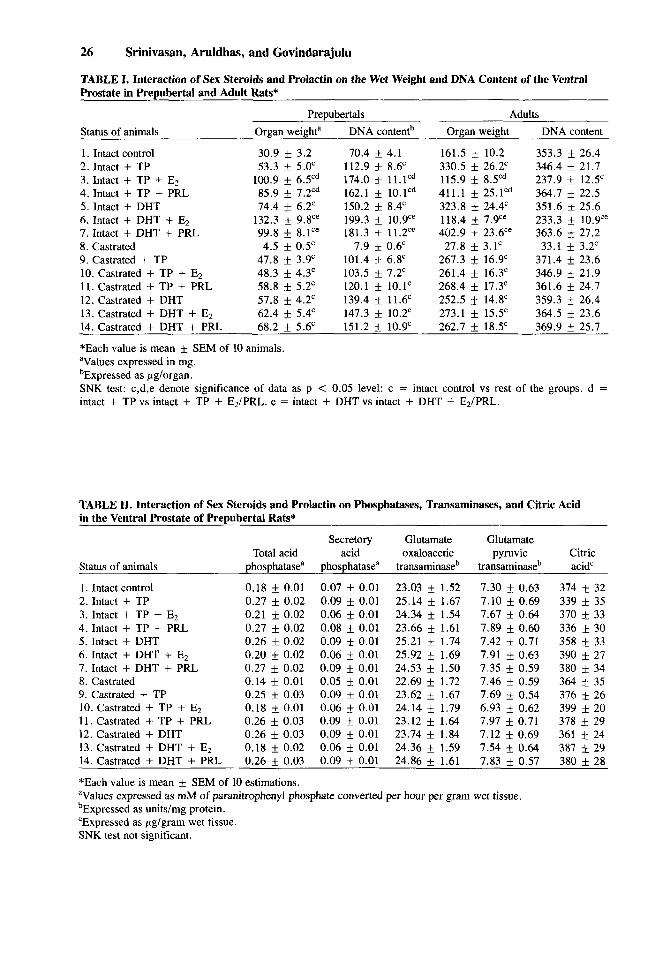
Prepubertals. Castration resulted in an obvious decrease in prostatic weight and DNA content. A significant increase in these parameters was observed in all the hormone treated groups when compared to intact controls. Administration of &/PRL along with TP/DHT registered a further increase in organ weight and DNA content in intact rats when compared to TP/DHT treated rats. However, no such changes were observed in castrated rats.
Adults. Both organ weight and DNA contents were decreased markedly after castration. TP/DHT given alone increased the organ weight in intact as well as castrated rats. E2 given along with TP/DHT markedly decreased the tissue weight and DNA content in intact adults. However, it did not produce any appreciable change in castrated adults. PRL given in combination with TP/DHT significantly enhanced the organ weight when compared to intact adults given TP/DHT. Similar to E2, PRL did not produce any significant change in castrated adults. None of the androgens or their combination with &/PRL produced any significant change in DNA content compared to intact controls.
Prepubertal (Table II) Total acid phosphatase and secretory acid phosphatase exhibited no significant
change between the treatment groups. None of the hormonal combinations produced any overall significant effect on transaminases or citric acid concentration. Adults (Table 111)
Whereas castration produced no significant change in the phosphatases, it decreased the transaminases and citric acid. TP/DHT given either alone or in combi- nation with PRL enhanced the phosphatases in both intact and castrated rats. How- ever, these combinations increased the activity of GOT only in intact rats. In fact, PRL further augmented the effect of androgens on citric acid. The response of GPT revealed the requirement of PRL along with TP/DHT for an increased activity in
26 Srinivasan, Aruldhas, and Govindarajulu
TABLE I. Interaction of Sex Steroids and Prolactin on the Wet Weight and DNA Content of the Ventral Prostate in Prepubertal and Adult Rats*
Prepubertals Adults Status of animals Organ weighta DNA contentb Organ weight DNA content
1. Intact control 30.9 f 3.2 70.4 f 4.1 161.5 k 10.2 353.3 f 26.4 2. Intact + TP 53.3 f 5.0' 112.9 k 8.6' 330.5 f 26.2' 346.4 f 21.7 3. Intact + TP + Ez 100.9 f 6.5Cd 174.0 f 11.lCd 115.9 f 8.Yd 237.9 f 12.5' 4. Intact + TP + PRL 85.9 f 7.2Cd 162.1 f 10.lCd 411.1 f 25.1Cd 364.7 f 22.5 5 . Intact + DHT 74.4 f 6.2' 150.2 8.4' 323.8 f 24.4' 351.6 f 25.6 6. Intact + DHT + Ez 132.3 f 9.8'" 199.3 k 10.9'" 118.4 f 7.9'e 233.3 f 10.9'" 7. Intact + DHT + PRL 99.8 + 8.1'" 181.3 f 11.2" 402.9 f 23.6" 363.6 f 27.2 8. Castrated 4.5 f 0.5' 7.9 f 0.6' 27.8 f 3.1' 33.1 f 3.2' 9. Castrated + TP 47.8 f 3.9' 101.4 6.8' 267.3 f 16.9' 371.4 f 23.6 10. Castrated + TP + E2 48.3 f 4.3' 103.5 f 7.2' 261.4 f 16.3' 346.9 f 21.9 11. Castrated + TP + PRL 58.8 f 5.2' 120.1 f 10.1' 268.4 f 17.3' 361.6 f 24.7 12. Castrated + DHT 57.8 f 4.2' 139.4 f 11.6' 252.5 f 14.8' 359.3 f 26.4 13. Castrated + DHT + Ez 62.4 f 5.4' 147.3 k 10.2' 273.1 f 15.5' 364.5 f 23.6 14. Castrated + DHT + PRL 68.2 f 5.6' 151.2 k 10.9' 262.7 f 18.5' 369.9 f 25.7
*Each value is mean f SEM of 10 animals. aValues expressed in mg. bExpressed as pglorgan. SNK test: c,d,e denote significance of data as p < 0.05 level: c = intact control vs rest of the groups. d = intact + TP vs intact + TP + E,/PRL. e = intact + DHT vs intact + DHT + E2/PRL.
TABLE 11. Interaction of Sex Steroids and Prolactin on Phosphatases, Transaminases, and Citric Acid in the Ventral Prostate of Prepubertal Rats*
Secretory Glutamate Glutamate Total acid acid oxaloacetic pyruvic
Status of animals phosphatasea phosphatase" transaminase' transaminase'
1 . Intact control 0.18 f 0.01 0.07 f 0.01 23.03 f 1.52 7.30 L 0.63 2. Intact + TP 0.27 f 0.02 0.09 f 0.01 25.14 f 1.67 7.10 f 0.69 3. Intact + TP + Ez 0.21 f 0.02 0.06 f 0.01 24.34 f 1.54 7.67 f 0.64 4. Intact + TP + PRL 0.27 f 0.02 0.08 f 0.01 23.66 f 1.61 7.89 f 0.60 5 . Intact + DHT 0.26 f 0.02 0.09 f 0.01 25.21 f 1.74 7.42 f 0.71 6. Intact + DHT + E2 0.20 f 0.02 0.06 k 0.01 25.92 f 1.69 7.91 f 0.63 7. Intact + DHT + PRL 0.27 f 0.02 0.09 f 0.01 24.53 f 1.50 7.35 f 0.59 8. Castrated 0.14 f 0.01 0.05 f 0.01 22.69 f 1.72 7.46 f 0.59 9. Castrated + TP 0.25 f 0.03 0.09 f 0.01 23.62 f 1.67 7.69 f 0.54 10. Castrated + TP + E2 0.18 f 0.01 0.06 f 0.01 24.14 f 1.79 6.93 f 0.62 11. Castrated + TP + PRL 0.26 f 0.03 0.09 f 0.01 23.12 f 1.64 7.97 f 0.71 12. Castrated + DHT 0.26 f 0.03 0.09 f 0.01 23.74 f 1.84 7.12 f 0.69 13. Castrated + DHT + E2 0.18 f 0.02 0.06 f 0.01 24.36 f 1.59 7.54 f 0.64 14. Castrated + DHT + PRL 0.26 f 0.03 0.09 f 0.01 24.86 f 1.61 7.83 f 0.57
*Each value is mean f SEM of 10 estimations. aValues expressed as mM of paranitrophenyl phosphate converted per hour per gram wet tissue. 'Expressed as unitslmg protein. 'Expressed as pg/gram wet tissue. SNK test not significant.
Citric acid'
374 f 32 339 & 35 370 f 33 336 If: 30 358 * 33 390 f 27 380 If: 34 364 * 35 376 f 26 399 * 20 378 * 29 361 f 24 387 +_ 29 380 f 28

Sex Steroids and Prolactin on Phosphatases 27
TABLE III. Interaction of Sex Steroids and Prolactin on Phosphatases, Transaminases, and Citric Acid in the Ventral Prostate of Adult Rats*
Secretory Glutamate Glutamate Total acid acid oxaloacetic pyruvic
Status of animals phosphatasea phosphatasea transaminaseb transaminaseb
1. Intact control 0.32 f 0.02 0.17 f 0.01 38.33 k 2.14 11.73 f 0.86 2. Intact + TP 0.50 f O . M d 0.25 f 0.02d 52.19 f. 2.23d 14.89 f 0.93 3. Intact + TP + b 0.39 f 0.03 0.17 f O.Ole 44.12 f 2.17 12.00 f 0.84 4. Intact + TP + PRL 0.50 f O.Wd 0.33 f 0.03d 59.59 f 2.30d 17.76 f 0.96d 5. Intact + DHT 0.50 + 0.03d 0.25 f 0.02d 51.19 i 2.25d 15.13 f. 0.95 6. Intact + DHT + E2 0.39 f 0.03 0.16 f 0.02f 43.76 f 2.20 12.32 f 0.85 7. Intact + DHT + PRL 0.51 f. O.Md 0.33 f 0.03d 58.12 f 2.29d 18.05 f 0.93d 8. Castrated 0.24 k 0.01 0.10 f 0.01 26.53 f 2.Wd 7.63 k 0.56d 9. Castrated + TP 0.50 f O.Wd 0.24 f 0.02d 39.23 f 2.16 12.05 f. 0.89 10. Castrated + TP + Ez 0.50 f O.Wd 0.25 f 0.02d 46.37 f 2.20 14.85 f 0.91 11. Castrated + TP + PRL 0.51 f. O.Wd 0.30 k 0.02d 46.13 f 2.19 14.96 f 0.90 12. Castrated + DHT 0.51 f 0.Md 0.25 f 0.02d 39.12 k 2.15 12.19 f 0.91 13. Castrated + DHT + E2 0.51 f O . M d 0.26 f 0.02d 45.87 f 2.19 15.20 f. 0.94 14. Castrated + DHT + PRL 0.51 f O.Md 0.33 f 0.03d 46.20 + 2.18 15.11 f 0.93
*Each value is mean f SEM of 10 estimations. aValues expressed as mM of paranitrophenyl phosphate converted per hour per gram wet tissue. bExpressed as units/mg protein.
Citric acid'
857 f 79 1297 k 106d 890 f 80e
1620 f 114d3e 1289 i- 102d 876 f 84f
1645 f 119d3f 519 f 70d 840 f 82
1103 f 87 1118 f 89 859 f 80
1114 f 85 1104 f 89
'Expressed as pgigram wet tissue. SNK test: d,e,f, denote the significance of the data as p < 0.05 level: d = intact control vs rest of the groups. e = intact + TP vs intact + TP + b/PRL. f = intact + DHT vs intact + DHT + E2/PRL.
intact rats. Unlike PRL, E2 combined with TP/DHT produced a differential effect. E2 given along with TP/DHT retarded the effects of androgens on the secretory acid phosphatase and citric acid. However, E2 produced no significant changes on trans- aminases and total acid phosphatase in intact adult rats. On the other hand, E2 combination elevated the activity of phosphatases in castrated adults. Nevertheless, transaminases and citric acid showed no significant alterations to any of the hormonal combinations in castrated adult rats.
DISCUSSION
The data obtained in the present study suggests that the secretory function of the prostate and its response to androgens are age dependent. This is evident from our findings that androgen treatment did not bring about any significant change in the secretory products of prepubertal rats, but they markedly increased the same in adult rats. However, an opposite trend was observed in the case of DNA content. These variations are due to the functional differences of the prepubertal and adult prostates. Whereas the immature prostate is in a dynamic state of growth, the completely grown and well-differentiated adult prostate is committed to secretory function. Early reports on the ventral prostate of prepubertal rats also showed that the subcellular structures involved in secretion are poorly developed without any secretory granules [24,25]. Further, it is well known that the subcellular organelles involved in secretory function, such as golgi complex, endoplasmic reticulum, and lysosomes of the rat prostate, undergo morphological changes during sexual maturation [26,27,28]. Hence, the poor response of the prostatic secretory products to androgens in prepubertal rats may be attributed to the active growth and undifferentiated secretory apparatus. In the

28 Srinivasan, Aruldhas, and Govindarajulu
present study also, androgens enhanced the secretory activity alone in intact adult rats. In castrated adult rats, in addition to the stimulatory effect on secretory activities, the DNA content was also raised to the levels of controls. This is very much in correlation to the findings of Lesser and Bruchovsky [29]. According to them, the exogenous androgens will first initiate the proliferation of cells when they are in subnormal number. Once the cell number becomes normal, androgens will enhance the secretory activity.
The castration-induced decrease in the activity of GOT, GPT, and citric acid concentration may be due to the increased degradation of these enzymes and the low rate of citric acid synthesis. The increase in the activity of GOT, GPT after the administration of TP/DHT, or their combination with PRL in adult rats suggests the increased provision of substrates like oxaloacetic acid and pyruvic acid for the synthesis of citric acid. The absence of any significant change in the activity of phosphatases after castration may be attributed to the decreased rate of degradation and/or the poor response of these enzymes to withdrawal of androgens. However, the significant increase in these parameters after the administration of androgens in castrated and intact adult rats clearly demonstrates their androgen dependency as reported previously [ 15,301.
Regarding the interaction of TP/DHT with PRL, the synergistic effect is well pronounced in citric acid concentration in intact adult rats. However, such a synergis- tic effect of PRL is not obviously seen in castrated rats. This is evident from the absence of any appreciable change in the activity of transaminases and citric acid concentration in these rats. It appears that PRL might have stimulated some testicular factor(s) other than androgens in intact rats, which may be responsible for its synergistic effects with androgens in intact animals. However, it requires further experimental evidence to confirm this hypothesis.
The interaction between E2 and androgens exhibits some interesting features. Whereas E2 is synergistic with androgen actions in prepubertals and castrated adult rats, it is antagonistic in intact adult rats. The antagonism is profoundly seen in parameters like DNA content secretory acid phosphatase activity and citric acid concentration. We have already demonstrated the antagonistic effect of E2 on prostatic weight when given along with TP/DHT in intact adult rats [31]. The androgen- estrogen antagonism in intact adults implies the possible involvement of some testic- ular factors for this unique feature. In this regard it is noteworthy to consider LH- RH-like peptides as the possible testicular factors. LH-RH-like peptides are reported to be produced by testes [32] and are adduced to have an inhibitory effect in the prostate [33,34]. It appears that E2 might have increased the synthesis and secretion of LH-RH-like peptides from testes, which in turn might have inhibited the effects of androgens. However, it warrants further study to unravel the exact mechanism that is involved in the estradiol-androgen antagonism in intact adult rats.
Estrogens are well known to modulate PRL secretion. Administration of estra- diol in animals and humans was shown to increase the pituitary content [35] blood levels and the release of PRL [36,37]. However, coadministration of E2 with increas- ing doses of testosterone was found to reduce the levels of PRL in castrated adult rats [38]. Further, DHT was reported not only to be inactive in stimulating PRL secretion [39], but also to partly reverse the stimulatory effect of E2 on PRL secretion in pituitary cell culture [40]. These investigations clearly disprove the chances for the increment in the levels of PRL when Q is given along with TP/DHT. The results

Sex Steroids and Prolactin on Phosphatases 29
obtained in the present study also suggest that at the dosage of E2 (20 pg/rat/day) and TP/DHT (2 mg/rat/day) used, there may not be any significant increase in the levels of PRL. Thus, from our studies it is reasonable to presume that either the synergistic or antagonistic effect of E2 with TP/DHT is a specific one and independent of PRL.
CONCLUSIONS
Our study has shown the potential usefulness of the secretory products for the elucidation of hormonal interaction in the ventral prostate. Further studies should be extended on some of the key metabolic enzymes that determine the accumulation of citric acid in the ventral prostate and the testicular factor@) responsible for the differential effects of PRL and E2 in adult rats to unravel the exact mode of interac- tions of these hormones with androgens.
ACKNOWLEDGMENTS
The authors are thankful to NIADDK, USA, for providing ovine PRL as a gift. N.S. is grateful to the Council of Scientific and Industrial Research, New Delhi, for financial support.
REFERENCES
1 . Williams-Ashman HG, Reddi AH: Androgen regulation of tissue growth and function. In Litwack G (ed): “Biochemical Actions of Hormones,” Vol 11. New York, Academic Press, 1972, pp 257- 294.
2. Coffey DS: The effects of androgens on DNA and RNA synthesis in sex accessory tissue. In Brandes D (ed): “Male Accessory Sex Organs: Structure and Function in Mammals.” New York, Academic Press, 1974, pp 307-328.
3. Bruchovsky N, Lesser B: Control of proliferative growth in androgen responsive organs and neoplasms. In Singhal RL, Thomas JA (4s) : “Cellular Mechanisms Modulating Gonadal Hormone Action, Advances in Sex Hormone Research” Vol2. Baltimore, University Park Press, 1976, pp 1- 56.
4. Mawhinney MG, Neubauer BL: Actions of estrogen in the male. Invest Urol 16:409-420, 1979. 5. Tasar C, Scott WW: A search for inhibitors of prostatic growth stimulators. Invest Urol 1:482-498,
1964. 6. Shimazaki J, Furuya N, Yamakana M, Shida K: Effects of estrogen administration on testosterone-
induced growth and increase in enzyme activities in the ventral prostate of castrated rats. Endocrinol Jap 16: 163-168, 1969.
7. Suffrin G, Coffey DS: A comparison of the hormone responsiveness of the prostate and seminal vesicle. Invest Urol 11:386-391, 1974.
8. Mawhinney MG, Belis JA: Androgens and estrogens in prostatic neoplasia. In Singhal RL, Thomas JA (eds): “Cellular Mechanisms Modulating Gonadal Hormone Action, Advances in Sex Hormone Research,” Vol2. Baltimore, University Park Press, 1976, pp 141-210.
9. Ehrlichman RJ, Isaacs JT, Coffey DS: Differences in the effects of estradiol on dihydrotestosterone induced prostatic growth of the castrate dog and rat. Invest Urol 18:466-470, 1981.
10. Nicoll CS: Physiological actions of prolactin. In Knobil E, Sawyer WH (eds): “The Pituitary Gland and Its Neuroendocrine Control,” Vol IV. Washington, DC, American Physiological Society, 1974, pp 253-292.

30 Srinivasan, Aruldhas, and Govindarajulu
11. Thomas JA, Keenan EJ: Prolactin influence upon androgen action in male accessory sex organs. In Singhal RL, Thomas JA (eds): “Cellular Mechanisms Modulating Gonadal Hormone Action,” Vol 2. Baltimore, University Park Press, 1976, pp 425-470.
12. Humphrey GF, Mann T: Studies on the metabolism of semen: Citric acid in semen. Biochem J
13. Lostroh AJ: Regulation by testosterone and insulin of citrate secretion and protein synthesis in explanted mouse prostates. Proc Natl Acad Sci USA 60:1312-1318, 1968.
14. Dondero F, Sciara F, Isidori A: Evaluation of relationship between plasma testosterone and human seminal citrate content. Fertil Steril23: 168-171, 1972.
15. Tenniswood M, Bird CE, Clark AF: Acid phosphatases: Androgen dependent markers of rat prostate. Can J Biochem 54:350-357, 1976.
16. Awapara J: The influence of sex hormones on the transaminases of the accessory sex organs of the male rat. Endocrinology 51:75-79, 1952.
17. Franklin RB, Brandly RL, Costello LC: Effects of inhibitors of RNA and protein synthesis on mitochondria1 aspartate aminotransferase response to testosterone in rat ventral prostate. The Prostate 3:637-642, 1982.
18. Burton D: A study of the conditions and mechanisms of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem J 62:3 15-323, 1956.
19. Bessey OA, Lowry OH, Brock MJ: A method for the rapid determination of alkaline phosphatase with five cubic millimeters of serum. J Biol Chem 164:321-325, 1946.
20. Reitman S, Frankel S: A colorimetic method for the determination of serum glutamic oxaloacetic and glutamic pyruvic transaminases. Amer J Clin Pathol28:56-63, 1957.
21. Beutler E, Yeh MK: A simplified method for the determination of citric acid. J Lab Clin Med
22. Sokal RR, Rohlf FG: “Biometry-The Principle and Practice of Statistics in Biological Research.”
23. Zar JH: “Biostatistical Analysis.” Englewood Cliffs, NJ, Prentice-Hall, 1974, pp 151-155. 24. Moore CR, Price D, Gallagher TF: Rat-prostate cytology as a testis-hormone indicator and the
prevention of castration changes by testis-extract injections. Amer J Anat 45:71-107, 1930. 25. Brandes D: “Male Accessory Sex Organs, Structure and Function in Mammals. New York,
Academic Press, 1974, pp 18-113. 26. Harkin JC: An electron microscopic study of the castration changes in the rat prostate. Endocrinol-
27. Harkin JC: An electron microscopic study of the age changes in the rat prostate and a comparison with prostate hyperplasia in man. Am J Path 33502 (Abst), 1957.
28. Brandes D: The fine structure and histochemistry of prostatic glands in relation to sex hormones. In Bourne GH, Danielli JF (eds): “International Review of Cytology,” Vol 20. New York, Academic Press, 1966, pp 207-276.
29. Lesser B, Bruchovsky N: The effects of testosterone, 5~~-dihydrotestosterone and adenosine 3‘,5’- monophosphate on cell proliferation and differentiation in rat prostate. Biochim Biophys Acta
30. Tenniswood MP, Abrahams PP, Bird CE, Clark AF: Effects of castration and androgen replacement on acid phosphatase activity in the adult rat prostate gland. J Endocr 77:301-308, 1978.
31. Srinivasan N , Michael Aruldhas M, Govindarajulu P: Sex steroid-induced changes in collagen of the prostate and seminal vesicle of rats. J Androl 7:55-58, 1986.
32. Sharpe RM, Fraser HM, Cooper I, Rommerts FRG: Sertoli-Leydig cell communication via an LHRH like factor. Nature 290:785-787, 1981.
33. Sundaram K, Cao YQ, Wang NG, Bardin WC, Rivier J, Vale W: Inhibition of the action of sex steroids by gonadotropin releasing hormone (GnRH) agonist: A new biological effect. Life Sciences
34. Raju VS, Rukurnini V, Reddy PRK: Effect of gonadotropin releasing hormone on ventral prostate of rat. Horm Metabol Res 17:241-243, 1985.
35. Ajika K, Krulich L, Fawcett CP, McCann SM: Effects of estrogen on plasma and pituitary gonadotropins and prolactin and on hypothalamic releasing and inhibiting factors. Neuroendocrinol-
36. Frantz AG, Kleinberg DL, Noel GL: Studies on prolactin in man. Recent Prog Horm Res 28:527-
44:97-105, 1949.
54: 125-131, 1959.
San Francisco, W.H. Freeman, 1969.
ogy 601185-199, 1957.
3081426-437, 1973.
28:83-88, 1981.
Ogy 9:304-315, 1972.
590, 1972.

Sex Steroids and Prolactin on Phosphatases 31
37. Ojeda SR, Castro-Vazquez A, Jameson HE: Prolactin release in response to blockade of dopami- nergic receptors and to TRH injections in developing and adult rats: Role of estrogen in determining sex differences. Endocrinology 100:427-439, 1977.
38. Shi YF, Patterson AP, Sherins RJ: Increased plama and pituitary prolactin concentrations in adult male rats with selective elevation of FSH levels may be explained by reduced testosterone and increased estradiol production. J Androl 7: 105-111, 1986.
39. Nolin JM, Campbell GT, Nansel DD, Bogdanove EM: Does androgen influence prolactin secretion? Endocr Res Commun 4:61-70, 1977.
40. Labrie F, Ferland L, Denijeau F, Beaulieu M: Sex steroids interact with dopamine at the hypotha- lamic and pituitary levels to modulate prolactin secretion. J Steroid Biochem 12:323-330, 1980.





























