Embed Size (px)
Citation preview

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 241, 570–573 (1997)ARTICLE NO. RC977859
Interaction of P1 RepA with Replication Originof Plasmid Rts1: Capability of an Initiator ProteinInducing Replication from a Foreign Origin
Yong Fang Li, Akira Tabuchi,1 and Yoshiro Terawaki2
Department of Bacteriology, Shinshu University School of Medicine, Asahi 3-1-1, Matsumoto 390, Japan
Received October 14, 1997
protein specifically inhibits the replication of its respec-Rts1 RepA and P1 RepA are trans-acting proteins tive plasmid when an excess amount of the protein is
essential for the initiation of replication of plasmid supplied in trans (4, 5). Thus, RepA proteins of Rts1Rts1 and prophage P1, respectively. In this study, we and P1 exhibit dual functions in the replication ma-found that, in vitro, P1 RepA bound to the Rts1 ori chinery. To investigate the functional domains of Rts1fragment and Rts1 incI fragment as strongly as Rts1 RepA, we recently constructed hybrid proteins of Rts1RepA. In addition P1 RepA, in trans, activated the Rts1
RepA with P1 RepA (6). In the course of the study, wereplication origin that was cloned in pBR322, thusnoticed that P1 RepA bound in vitro to the Rts1 oriallowing the ori plasmid to be maintained in a polAfragment, whereas Rts1 RepA did not bind to the P1E. coli host. Under these conditions, however, the oriori fragment. In this study, we examined interactionsplasmid was unstable as compared with that when ac-of P1 RepA with the Rts1 replication origin in vivo astivated by Rts1 RepA. In addition, we found that Rts1well as in vitro and found that P1 RepA could ineffi-RepA showed no interaction with the P1 replicationciently induce replication in vivo from the Rts1 ori.origin. q 1997 Academic Press
MATERIALS AND METHODS
Mini-Rts1, the minimal replication region of the plas- Bacterial strains and plasmids. E. coli JG112 (polA) (7) was usedmid Rts1, consists of 1,855 bp and contains three im- for the Rts1 ori and P1 ori activation assay, and HB101 (recA13)
(8) was for preparation of the RepA extract. Plasmids used wereportant components: the replication origin, ori, that en-pGB:LX01 and pALA619 (9), which are pGB2 recombinant plasmidscompasses a group of direct repeated sequences; a gene,with Rts1 repA (mini-Rts1 coordinates 1158 to 260) and P1 repArepA, encoding the replication initiator protein RepA(mini-P1 coordinates 609-1570), respectively. pGB:LX01 was con-
which binds to the direct repeated sequences (iterons) structed in this study. pGB2 (10) is a derivative of pSC101 and con-in ori, leading to initiation of Rts1 replication; and incI, fers spectinomycin (Sp) resistance. The repA expression from both
plasmids is under the control of a promoter derived from a pBR322containing five iterons, which is located downstream ofsubregion (coordinates 4168-29). pTW22:ori (6) and pBR322:P1orirepA and is involved in negative control of the plasmidare pBR322 recombinant plasmids containing Rts1 ori (mini-Rts1replication (1). In the ori region, tandem DnaA boxes coordinates 1441-1191) and P1 ori (mini-P1 coordinates 1-608), re-
and four GATC repeats are located upstream of the spectively, and which confer ampicillin (Ap) resistance.iterons. The replicon structure is quite similar to that Media and chemicals. Luria-Bertani (LB) broth (Difco Labora-of mini-P1, a minimal replicon of prophage P1, which tories) was used for bacterial cultivation. Penassay broth (Difco) agarconsists of ori iterons with DnaA boxes and GATC re- was used for selection of transformants, and 2YT medium (11) was
for isolation of plasmid DNA. Ap (30 mg/ml) and Sp (30 mg/ml) werepeats, a repA gene and incA iterons (2). The amino acidincluded in the medium when needed. [g-32P]ATP used for end label-sequence of Rts1 RepA consisting of 288 amino acidsing DNA fragments was purchased from Amersham.shares 60% homology with that of P1 RepA consisting
Preparation of cell extracts of RepA protein. RepA protein wasof 286 amino acids. Nevertheless, Rts1 and P1 belongprepared by ammonium sulfate fractionation of cell lysates fromto different incompatibility groups (2, 3). Each RepA HB101 harboring pGB: LX01 (or pALA619) by the method for prepa-ration of ‘‘fraction II’’ reported by Fuller et al. (12) with a slightmodification by Abeles (13).1 Present address: Department of Crop. & Animal Science and Bio-
science & Biotechnology, Shinshu University Faculty of Agriculture, Electrophoresis mobility shift assay for binding of RepA to ori andinc iterons. The binding was assayed as follows. The 10 ml binding8304 Minami-Minowa Village, Kamiina County 399-45, Japan.
2 Corresponding author. Fax: (81)-263-37-2616. E-mail: yter399@ mixture contained 20 mM Tris-acetate, pH 7.5, 50 mM magnesiumacetate, 1 mM dithiothreitol, 0.5 mM ATP, 5 mM EDTA, 0.2 mg polygipac.shinshu-u.ac.jp.
0006-291X/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
570
AID BBRC 7859 / 6942$$1321 12-08-97 16:37:56 bbrcg AP: BBRC
Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
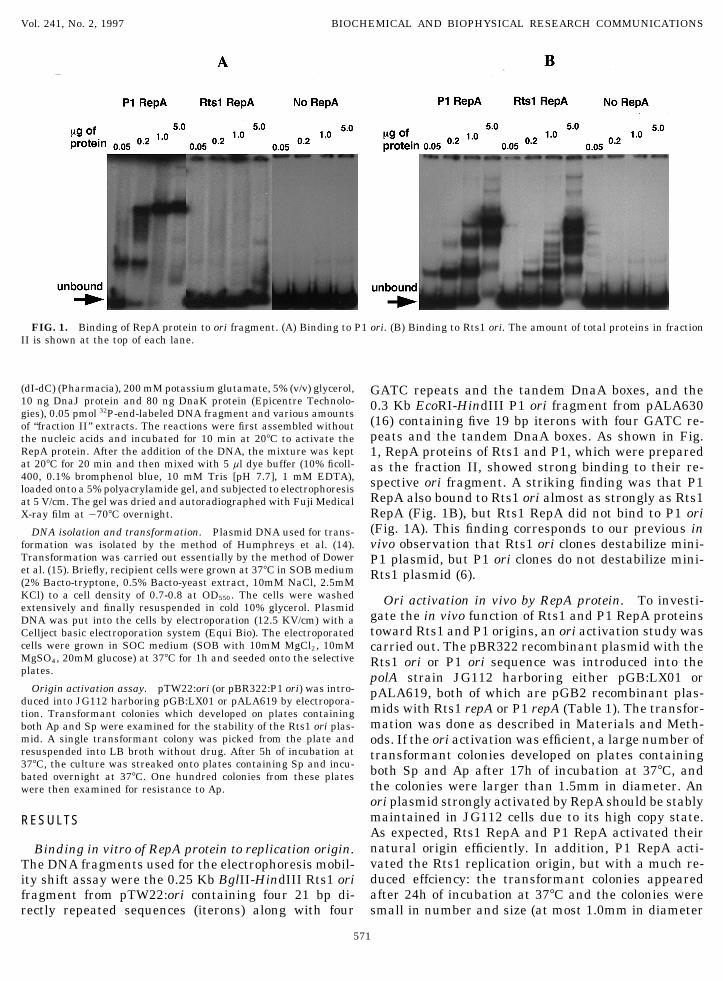
FIG. 1. Binding of RepA protein to ori fragment. (A) Binding to P1 ori. (B) Binding to Rts1 ori. The amount of total proteins in fractionII is shown at the top of each lane.
(dI-dC) (Pharmacia), 200 mM potassium glutamate, 5% (v/v) glycerol, GATC repeats and the tandem DnaA boxes, and the10 ng DnaJ protein and 80 ng DnaK protein (Epicentre Technolo- 0.3 Kb EcoRI-HindIII P1 ori fragment from pALA630gies), 0.05 pmol 32P-end-labeled DNA fragment and various amounts
(16) containing five 19 bp iterons with four GATC re-of ‘‘fraction II’’ extracts. The reactions were first assembled withoutpeats and the tandem DnaA boxes. As shown in Fig.the nucleic acids and incubated for 10 min at 207C to activate the
RepA protein. After the addition of the DNA, the mixture was kept 1, RepA proteins of Rts1 and P1, which were preparedat 207C for 20 min and then mixed with 5 ml dye buffer (10% ficoll- as the fraction II, showed strong binding to their re-400, 0.1% bromphenol blue, 10 mM Tris [pH 7.7], 1 mM EDTA), spective ori fragment. A striking finding was that P1loaded onto a 5% polyacrylamide gel, and subjected to electrophoresis
RepA also bound to Rts1 ori almost as strongly as Rts1at 5 V/cm. The gel was dried and autoradiographed with Fuji MedicalRepA (Fig. 1B), but Rts1 RepA did not bind to P1 oriX-ray film at 0707C overnight.(Fig. 1A). This finding corresponds to our previous inDNA isolation and transformation. Plasmid DNA used for trans-
formation was isolated by the method of Humphreys et al. (14). vivo observation that Rts1 ori clones destabilize mini-Transformation was carried out essentially by the method of Dower P1 plasmid, but P1 ori clones do not destabilize mini-et al. (15). Briefly, recipient cells were grown at 377C in SOB medium Rts1 plasmid (6).(2% Bacto-tryptone, 0.5% Bacto-yeast extract, 10mM NaCl, 2.5mMKCl) to a cell density of 0.7-0.8 at OD550. The cells were washed Ori activation in vivo by RepA protein. To investi-extensively and finally resuspended in cold 10% glycerol. Plasmid
gate the in vivo function of Rts1 and P1 RepA proteinsDNA was put into the cells by electroporation (12.5 KV/cm) with atoward Rts1 and P1 origins, an ori activation study wasCellject basic electroporation system (Equi Bio). The electroporated
cells were grown in SOC medium (SOB with 10mM MgCl2, 10mM carried out. The pBR322 recombinant plasmid with theMgSO4, 20mM glucose) at 377C for 1h and seeded onto the selective Rts1 ori or P1 ori sequence was introduced into theplates. polA strain JG112 harboring either pGB:LX01 or
Origin activation assay. pTW22:ori (or pBR322:P1 ori) was intro- pALA619, both of which are pGB2 recombinant plas-duced into JG112 harboring pGB:LX01 or pALA619 by electropora-
mids with Rts1 repA or P1 repA (Table 1). The transfor-tion. Transformant colonies which developed on plates containingmation was done as described in Materials and Meth-both Ap and Sp were examined for the stability of the Rts1 ori plas-
mid. A single transformant colony was picked from the plate and ods. If the ori activation was efficient, a large number ofresuspended into LB broth without drug. After 5h of incubation at transformant colonies developed on plates containing377C, the culture was streaked onto plates containing Sp and incu- both Sp and Ap after 17h of incubation at 377C, andbated overnight at 377C. One hundred colonies from these plates
the colonies were larger than 1.5mm in diameter. Anwere then examined for resistance to Ap.ori plasmid strongly activated by RepA should be stablymaintained in JG112 cells due to its high copy state.RESULTSAs expected, Rts1 RepA and P1 RepA activated theirnatural origin efficiently. In addition, P1 RepA acti-Binding in vitro of RepA protein to replication origin.vated the Rts1 replication origin, but with a much re-The DNA fragments used for the electrophoresis mobil-duced effciency: the transformant colonies appearedity shift assay were the 0.25 Kb BglII-HindIII Rts1 oriafter 24h of incubation at 377C and the colonies werefragment from pTW22:ori containing four 21 bp di-
rectly repeated sequences (iterons) along with four small in number and size (at most 1.0mm in diameter
571
AID BBRC 7859 / 6942$$1322 12-08-97 16:37:56 bbrcg AP: BBRC
Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
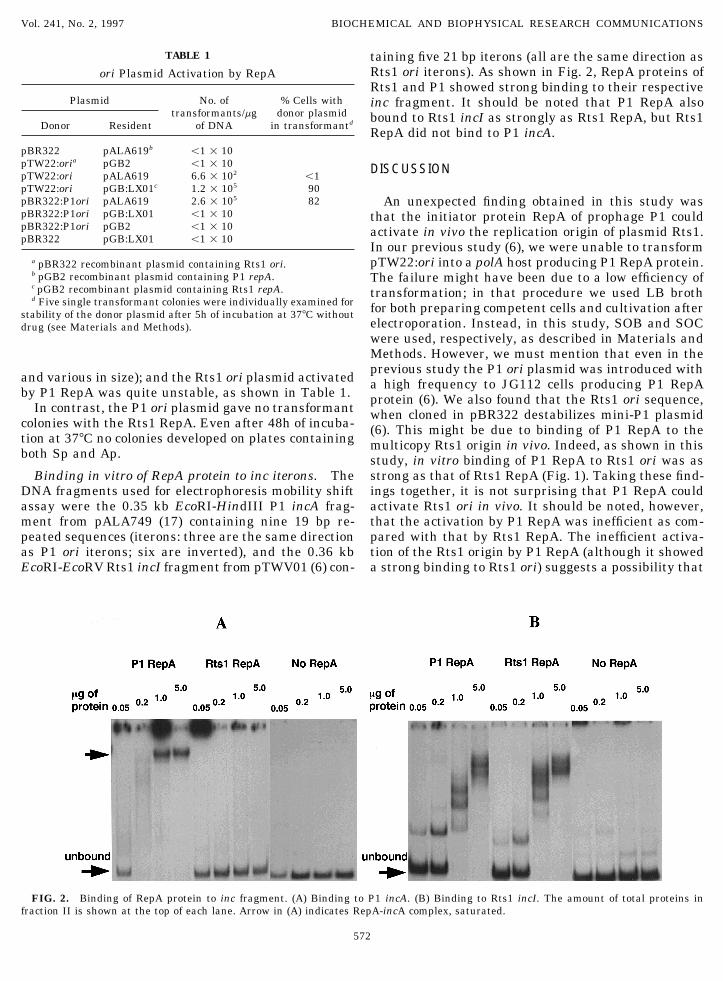
TABLE 1 taining five 21 bp iterons (all are the same direction asRts1 ori iterons). As shown in Fig. 2, RepA proteins ofori Plasmid Activation by RepARts1 and P1 showed strong binding to their respective
Plasmid No. of % Cells with inc fragment. It should be noted that P1 RepA alsotransformants/mg donor plasmid bound to Rts1 incI as strongly as Rts1 RepA, but Rts1
Donor Resident of DNA in transformantd
RepA did not bind to P1 incA.pBR322 pALA619b õ1 1 10pTW22:oria pGB2 õ1 1 10 DISCUSSIONpTW22:ori pALA619 6.6 1 102 õ1pTW22:ori pGB:LX01c 1.2 1 105 90pBR322:P1ori pALA619 2.6 1 105 82 An unexpected finding obtained in this study waspBR322:P1ori pGB:LX01 õ1 1 10 that the initiator protein RepA of prophage P1 couldpBR322:P1ori pGB2 õ1 1 10 activate in vivo the replication origin of plasmid Rts1.pBR322 pGB:LX01 õ1 1 10
In our previous study (6), we were unable to transformpTW22:ori into a polA host producing P1 RepA protein.a pBR322 recombinant plasmid containing Rts1 ori.
b pGB2 recombinant plasmid containing P1 repA. The failure might have been due to a low efficiency ofc pGB2 recombinant plasmid containing Rts1 repA. transformation; in that procedure we used LB brothd Five single transformant colonies were individually examined for for both preparing competent cells and cultivation afterstability of the donor plasmid after 5h of incubation at 377C without
electroporation. Instead, in this study, SOB and SOCdrug (see Materials and Methods).were used, respectively, as described in Materials andMethods. However, we must mention that even in theprevious study the P1 ori plasmid was introduced with
and various in size); and the Rts1 ori plasmid activated a high frequency to JG112 cells producing P1 RepAby P1 RepA was quite unstable, as shown in Table 1. protein (6). We also found that the Rts1 ori sequence,
In contrast, the P1 ori plasmid gave no transformant when cloned in pBR322 destabilizes mini-P1 plasmidcolonies with the Rts1 RepA. Even after 48h of incuba- (6). This might be due to binding of P1 RepA to thetion at 377C no colonies developed on plates containing multicopy Rts1 origin in vivo. Indeed, as shown in thisboth Sp and Ap. study, in vitro binding of P1 RepA to Rts1 ori was as
strong as that of Rts1 RepA (Fig. 1). Taking these find-Binding in vitro of RepA protein to inc iterons. TheDNA fragments used for electrophoresis mobility shift ings together, it is not surprising that P1 RepA could
activate Rts1 ori in vivo. It should be noted, however,assay were the 0.35 kb EcoRI-HindIII P1 incA frag-ment from pALA749 (17) containing nine 19 bp re- that the activation by P1 RepA was inefficient as com-
pared with that by Rts1 RepA. The inefficient activa-peated sequences (iterons: three are the same directionas P1 ori iterons; six are inverted), and the 0.36 kb tion of the Rts1 origin by P1 RepA (although it showed
a strong binding to Rts1 ori) suggests a possibility thatEcoRI-EcoRV Rts1 incI fragment from pTWV01 (6) con-
FIG. 2. Binding of RepA protein to inc fragment. (A) Binding to P1 incA. (B) Binding to Rts1 incI. The amount of total proteins infraction II is shown at the top of each lane. Arrow in (A) indicates RepA-incA complex, saturated.
572
AID BBRC 7859 / 6942$$1322 12-08-97 16:37:56 bbrcg AP: BBRC

Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Tetsuya Hayashi, Makoto Ohnishi, Takahiro Murata, and Qingbaosome subregion(s) other than the DNA binding domainTian for valuable discussions, and Kaori Sato for her technical assis-might exist on the Rts1 RepA molecule which is re-tance. This work was supported by a Grant-in Aid for Scientific Re-quired for stable ori activation. We are now searching search (07457072) from the Ministry of Education, Science and Cul-
for such a subregion on Rts1 RepA. ture of Japan, and by grants from the Yakult Foundation and theAiko Foundation.Contrary to our results with P1 RepA, Rts1 RepA
showed neither binding in vitro to P1 ori nor activationin vivo of the origin. We have recently made another REFERENCESobservation regarding the origin sequence for RepAbinding. The ori sequences of Rts1 and P1 contain four 1. Kamio, Y., Tabuchi, A., Itoh, Y., Katagiri, H., and Terawaki, Y.21 bp and five 19 bp iterons, respectively. A 19 bp con- (1984) J. Bacteriol. 158, 307–312.sensus sequence of the Rts1 iterons (5*-GGTGTGTGC- 2. Abeles, A. L., Snyder, K. M., and Chattoraj, D. K. (1984) J. Mol.TGAGGGGAAA-3 *) has four mismatches with the con- Biol. 173, 307–324.sensus sequence of P1 iterons (5*-GATGTGTGCTGG- 3. Coetzee, J. N., Datta, N., and Hedges, W. (1972) J. Gen. Micro-CGGGATA-3 *) (18). We synthesized these two 19 mer biol. 72, 542–552.oligonucleotides and tested them in the binding assay. 4. Muraiso, K., Mukhopadhyay, G., and Chattoraj, D. K. (1990) J.
Bacteriol. 172, 4441–4447.Interestingly, Rts1 RepA and P1 RepA bound only to5. Terawaki, Y., Nozue, H., Zeng, H., Hayashi, T., Kamio, Y., andtheir respective 19 mer sequence (our unpublished
Itoh, Y. (1990) J. Bacteriol. 172, 786–792.data). In contrast to the inability of P1 RepA to bind6. Tabuchi, A., Ohnishi, M., Hayashi, T., and Terawaki, Y. (1995)to the Rts1 ori consensus oligonucleotides, the protein
J. Bacteriol. 177, 4028–4035.showed strong binding to the native Rts1 ori fragment,7. Miller, J., Manis, J., Kline, B., and Bishop, A. (1978) Plasmid 1,as demonstrated here (Fig. 1B), and induced replica-
273–283.tion from the Rts1 origin. This suggests that the origin8. Boyer, H. W., and Roulland-Dussoix, D. (1969) J. Mol. Biol. 41,structure, containing many iterons, facilitates binding
459–472.of a RepA molecule with a low affinity (to individual9. Abeles, A. L., Reaves, L. D., and Austin, S. J. (1990) J. Bacteriol.iterons), leading to the formation of a mature complex
172, 4386–4391.(18) for initiation of plasmid replication. We have also10. Churchward, G., Belin, D., and Nagamine, Y. (1984) Gene 31,found that P1 RepA binds to the Rts1 incI iterons (Fig. 165–171.
2B), but Rts1 RepA does not bind to the P1 incA iterons11. Miller, J. H. (1972) Experiments in molecular genetics, p. 352–
(Fig. 2A) which again suggests a subtleness of RepA 355. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,interaction with iteron structure. N.Y.
Another interesting subject raised by this study is 12. Fuller, R. S., Kaguni, J. M., and Kornberg, A. (1981) Proc. Natl.why the RepA functions of P1 and Rts1 are not recipro- Acad. Sci. USA 78, 7370–7374.cal. P1 RepA might be sufficiently multipotential to 13. Abeles, A. L. (1986) J. Biol. Chem. 261, 3548–3555.activate diverse replication origins. Alternatively, the 14. Humphreys, G. O., Willshaw, G. A., and Anderson, E. S. (1975)Rts1 ori sequence may be more flexible in binding for- Biochem. Biophys. Acta. 383, 457–463.eign replication proteins. It would be worthwhile to 15. Dower, W. J., Miller, J. F., and Ragsdale, C. W. (1988) Nucl.
Acids Res. 16, 6127-.search for the amino acid residue(s) that can convertP1 RepA protein to the Rts1 RepA type, or vice versa. 16. Brendler, T. G., Abeles, A. L., and Austin, S. J. (1991) J. Bacte-
riol. 173, 3935–3942.17. Abeles, A. L., Reaves, L. D., Youngren-Grimes, B., and Austin,ACKNOWLEDGMENTS
S. J. (1995) Mol. Microbiol. 18, 903–912.18. Brendler, T. G., Abeles, A. L., Reaves, L. D., and Austin, S. J.We thank Ann L. Abeles for critical reading of the manuscript
and for plasmids pALA619, pALA630, and pALA749. We also thank (1997) Mol. Microbiol. 23, 559–567.
573
AID BBRC 7859 / 6942$$1322 12-08-97 16:37:56 bbrcg AP: BBRC











![[PPT]Role of DNA - Elizabeth Rose · Web viewcopyright cmassengale DNA Replication As the 2 DNA strands open at the origin, Replication Bubbles form Prokaryotes (bacteria) have a single](https://img.pdfslide.us/doc/110x75/5aa6232f7f8b9a7c1a8e557b/pptrole-of-dna-elizabeth-rose-viewcopyright-cmassengale-dna-replication-as-the.jpg)

![DNA Replication Initiation Is Blocked by a Distant ... · replication and segregation of key chromosomal loci including the origin and terminus [4]. These steps also define two periods](https://img.pdfslide.us/doc/110x75/60020c885f9a0a1b3c145fd4/dna-replication-initiation-is-blocked-by-a-distant-replication-and-segregation.jpg)















