Embed Size (px)
Citation preview

J. Cell Set. 54. 287-298 (1982) 287Printed in Great Britain © Company of Biologists Limited 1982
INTERACTION BETWEEN INTRACELLULAR
VACUOLES AND THE CELL SURFACE
ANALYSED BY FINITE APERTURE THEORY
INTERFERENCE REFLECTION MICROSCOPY
DAVID GINGELL, IAN TODD AND NORMAN OWENSDepartment of Biology as Applied to Medicine, The Middlesex Hospital Medical School,London WiP 6DB, U.K.
SUMMARY'
Using finite aperture theory we have shown that localized very dark areas in the interferencereflection images of Dictyostelium discoideum amoebae are due to the close intracellular approachof vesicles and tubular elements of the contractile vacuole system to the plasma membraneadjacent to the substratum. Vesicles interacting in this way become locally deformed to theplanar contour of the substratum and are separated from the cell surface membrane by aconstant < c i fim of cytoplasm. Lamellar processes formed by these cells on very adhesivesurfaces have identical dimensions. This minimal thickness may be a mechanical consequenceof a contractile mechanism which pulls membranes together.
INTRODUCTION
The contacts between cells and optically transparent substrata have been extensivelyobserved by interference reflection microscopy. Using this technique a careful analysisby Izzard & Lochner (1976) showed that mammalian cells in culture form adhesiveplaques, which appear as localized zero-order grey/black areas closer to the sub-stratum than the rest of the cell. These authors argued that at high illuminatingnumeri6al aperture (IN A) cell thickness has a negligible influence on the image and onlycell-substratum contacts are seen. They state (p. 137 op. cit.) that an exception to thisgeneralization can occur if the thickness of the cytoplasm is less than ~ 100 nm,but conclude that this will not influence image interpretation since cells do not thinto this extent. However, it has been argued theoretically (Gingell, 1981) that cyto-plasmic thicknesses even up to i-o fim can contribute significantly to the image atINA = i-18. Gingell & Vince (1982 a) have also demonstrated that cells can formultrathin lamellae, < o-i fim thick, which give rise to very dark interference imagesathighINA(i-i8).
We here describe and analyse the optical effects that arise when intracellularvesicles closely approach the cell surface. The entirely unexpected mechanicalrelations that emerge provide an intriguing parallel to the formation of ultrathin cyto-plasmic lamellae.
288 D. Gtngell, I. Todd and N. Owens
12
15

Images of vacuoles 289
MATERIALS AND METHODS
The optical equipment, method of quantitative analysis and cell cultures used in this studyhave been described previously (Gingell, Todd & Heavens, 1982; Gingell & Vince, 19820).Dictyostelium discoideum strain NC4 was employed for all quantitative measurements.
RESULTS
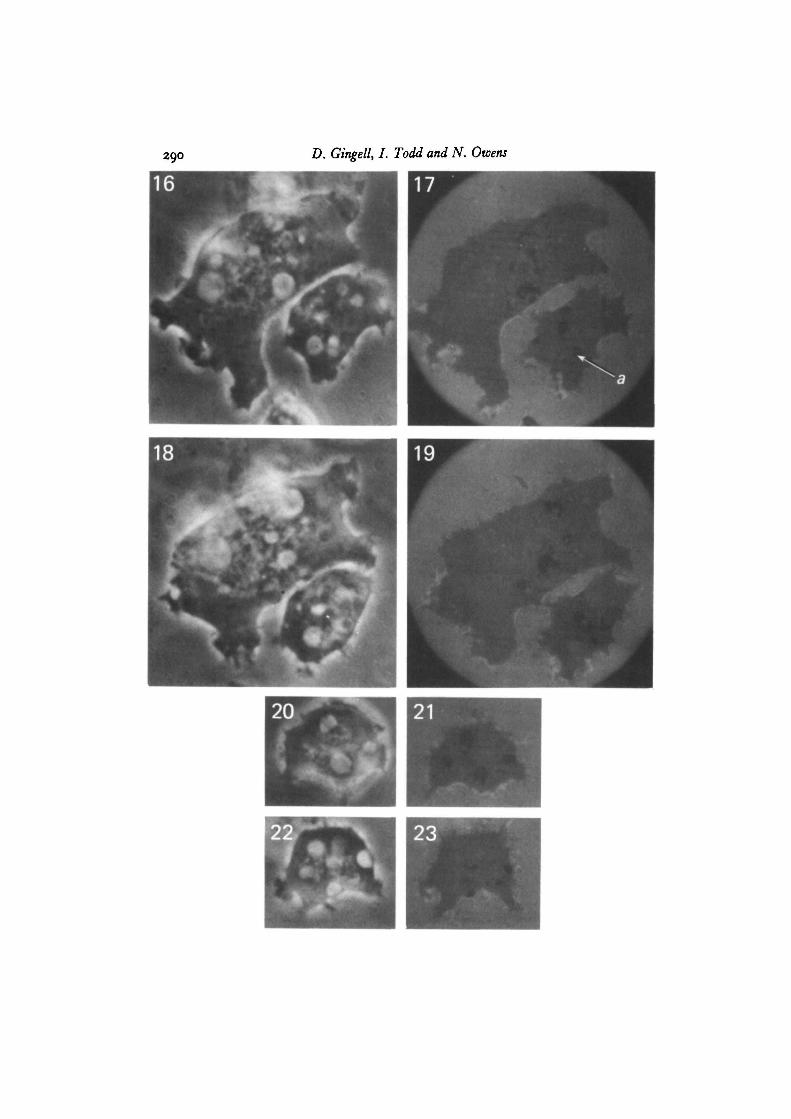
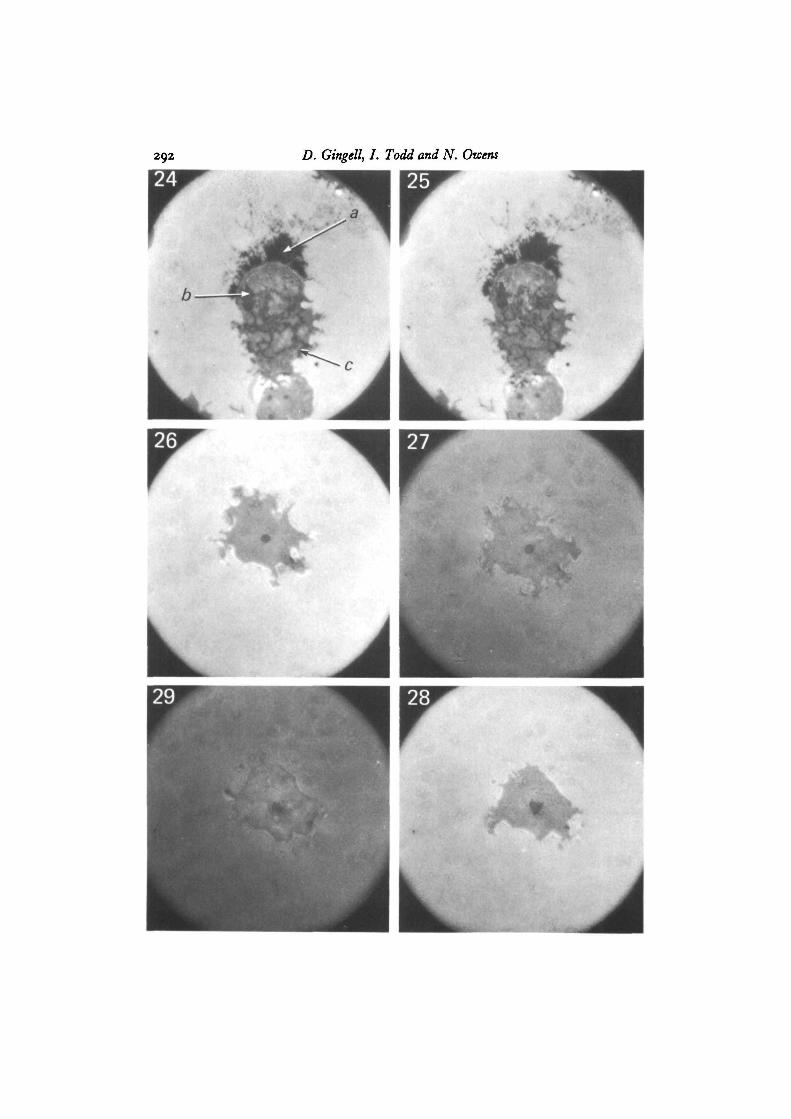
Observations at 546 run made with both NC4 and Ax2 strains, under the inter-ference microscope, showed that amoebae that have spread on clean glass, polylysine-treated glas9 and methylated glass, develop localized dark regions, which are zero-ordergrey/black in white light. These regions vary in shape with time, and are of two mainkinds. The larger of these have smooth contours and are often circular, ~ i-o/tmdiameter (arrow a in Fig. 2; arrow b in Fig. 24). The second are from thread-like struc-tures, which are able to change their form very rapidly and make reversible connectionswith the first kind (arrow b in Fig. 2; arrow c in Fig. 24). This reticulum of threadsand isolated elements exists in a state of continuous flux and interchange. The largerdark regions are relatively static but suddenly disappear, to be re-formed from therapidly moving threads over a period of a minute or so. The dark thread-like structuresseen in Figs. 24 and 25 have changed markedly in the few seconds that elapsed betweenthe two photographs. The black area at the upper end of this cell is due to a thincytoplasmic lamella (Gingell & Vince, 1982 a).
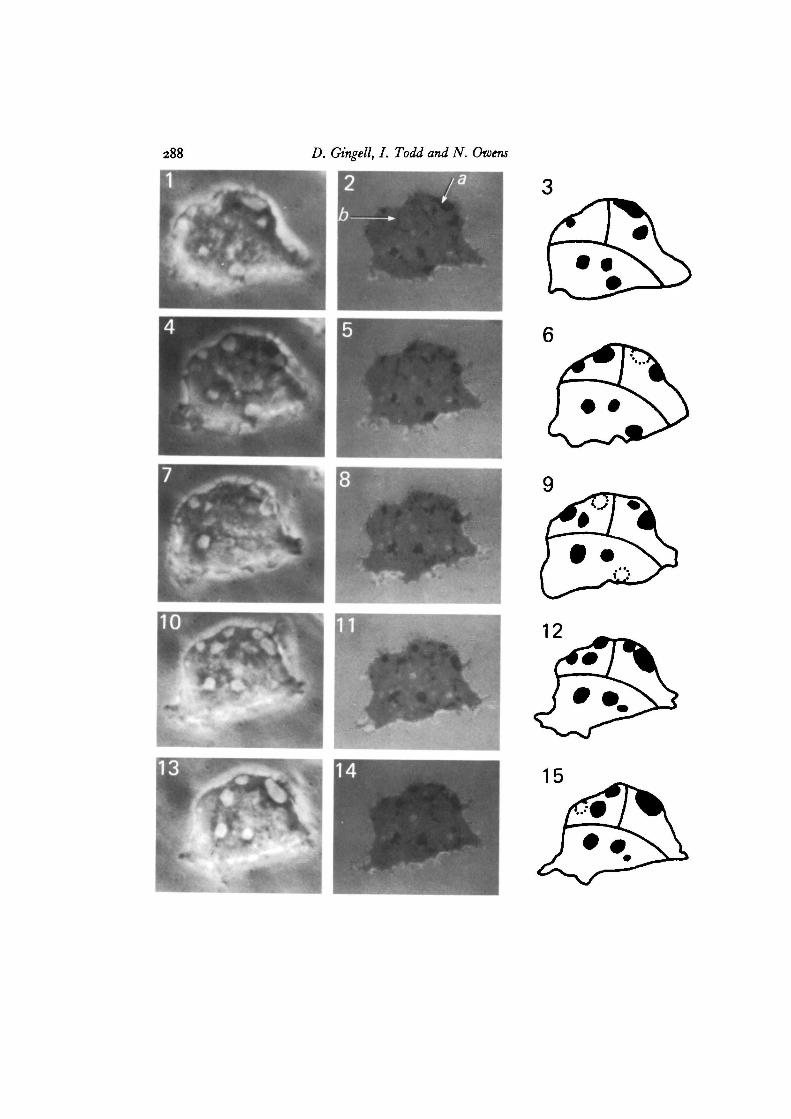
Alternate observations using interference and phase-contrast microscopy (seeFigs. 1-15 and 16-23) show that the larger dark interference images correspond withthe bright phase images of fluid-filled vacuoles. The fine threads could not be dis-tinguished by phase-contrast, against the background of rapidly moving refractileelements of the cytoplasm. Not all vacuoles seen under phase-contrast gave blackimages under interference, but all those giving interference images were found byphase-contrast to coincide with a vacuole near the glass/medium interface. Many freevacuoles were seen to follow the flow of cytoplasmic streams, whereas those generatinglocal dark interference images showed greatly constrained movement. Since the formsof the larger vacuoles change during the course of a minute or so, and since they alsorapidly disappear as they discharge their contents to the extracellular medium, it was
Figs. 1-15. One Axz strain amoeba attached to glass in 20 mM-NaCl. The left-handcolumn shows phase images, the centre column shows interference images and theright-hand column gives diagrams based on both sets of images, showing the positionsof contractile vacuoles grouped into 3 segments. Dotted circles represent the formerpositions of vacuoles. Contractile vacuoles appear bright in phase-contrast anddark in interference. The sequence of 5 phase-contrast images occupied about 5 min.Corresponding phase-contrast and interference images were taken practically simul-taneously, x 3500. Each segment of the diagrams 3, 6 etc. shows the sequence ofdischarge in a group of vacuoles. In each case a vacuole is re-formed at its previoussite of discharge. In addition, the right upper segment shows fusion between 2 con-tractile vacuoles. In Fig. 2 arrow a points to a contractile vacuole while b points to atubule. Figs. 24, 25 show tubules more clearly.
D. Gingell, I. Todd and N. Owens

Images of vacuoles 291
not possible to ascertain whether such vacuoles associated with the cell surfaceremained static with respect to the substratum during cell locomotion. Vacuolesshowing black interference images located near the edges of the grey cell-substratumcontact zone seemed particularly prone to release their contents and suddenly dis-appear. Rarely, one vacuole would release fluid into the region between the cell andthe substratum; when this happened the image at the discharge point became tran-siently diffuse and bright, then faded back to grey within a second or two. Occasionally,vacuoles could be seen generating oscillating patterns of Newton's rings, indicatingapproach towards the surface from deeper within the cytoplasm; this was invariablyfollowed by retreat rather than the formation of a stable uniform black contact. Thesequence in Figs. 1-15 shows paired images taken by phase-contrast and interferencealmost simultaneously. The corresponding diagrams show the positions of the largerinterferometrically dark vacuoles deduced from both sets of photographs. The mainfeature that becomes apparent is that the vacuoles disappear (dotted circles) and aresoon re-formed at the same sites; this can be seen in each demarcated group. In theupper right segment fusion between vacuoles is taking place. It is also apparent thatthe exocytosis of vacuolar contents has occurred only where vacuoles are adjacent to thefree lateral plasmalemma; the contents are therefore expelled into the surroundingmedium rather than into the contact zone. Exceptions to this were rare; the localdiffuse brightening of the image seen once when this happened can be attributed to atransient increase in cell-glass separation due to the volume of water expelled into thecontact zone.
We investigated the effects on the interference image of changing the refractiveindex of the medium, using bovine serum albumin (BSA) in 20 mM-NaCl. This had arefractive index of 1-368, which is the value determined by immersion refractometryfor Dictyostelium cytoplasm. The refractive index study was done because it iscapable of aiding distinction between the several different ways in which zero-ordergrey/black images can arise and also provides additional constraints for quantitativeanalysis (see Discussion).
By changing the medium around cells attached to either clean glass or polylysine-treated glass from 20 mM-NaCl to BSA in 20 mM-NaCl and then back again, weobtained striking changes in contrast (Figs. 26-29) that were fully reversible. Thechanges occurred as rapidly as the medium could be exchanged (~ 30 s). Althoughthe cells were virtually invisible in BSA, dark images of vacuoles and their relatedfilamentous connections remained visible. Disappearance of the cells was largelydue to the decreased reflectivity of the adjacent glass/medium interface.
Figs. 16-23. Pairs of phase-contrast (left) and interference (right) images of cells withcontractile vacuoles. Comparison of the images shows that, while all the major darkimages can be paired with a bright vacuole, the converse is not always true. Arrow ain Fig. 17 points to an image of a vacuole. Two adjacent vacuoles (Fig. 16) do notcontact the lower surface of the cell and do not, therefore, produce dark interferenceimages in Fig. 17. Ax2 cells in 20 mM-NaCl on glass, x 3500.
D. Gingell, I. Todd and N. Owens
Images of vacuoles 293
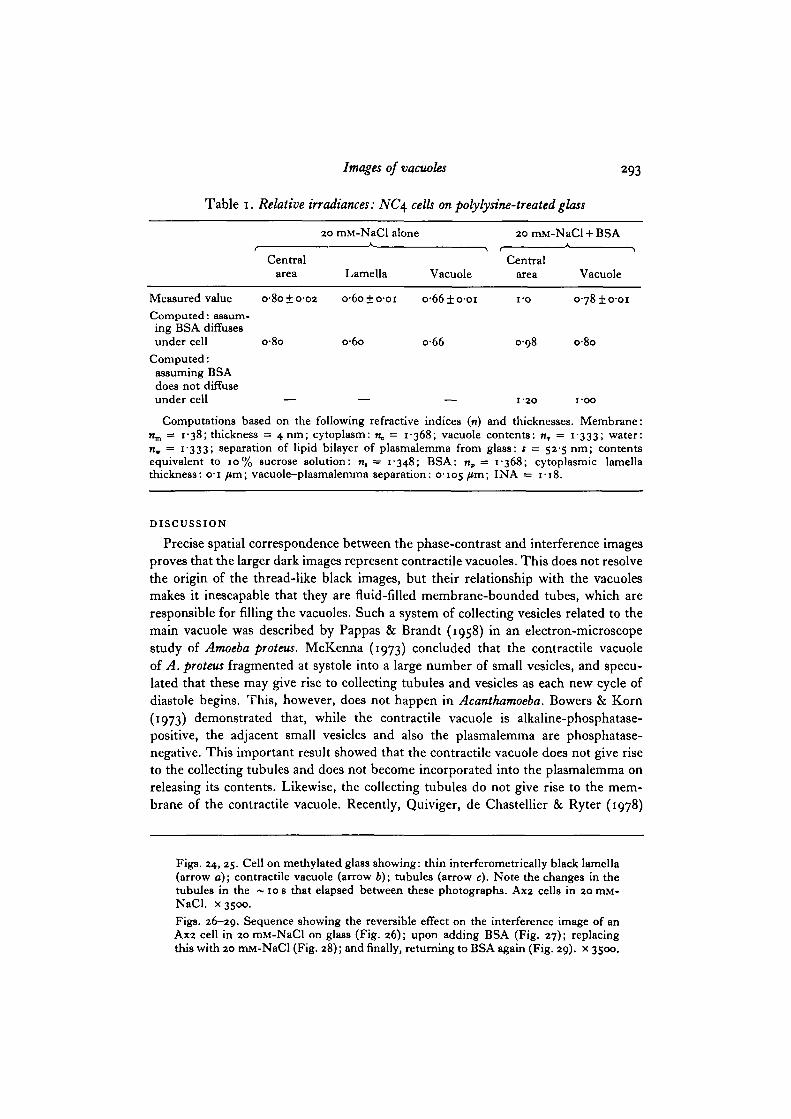
Table 1. Relative irradiances: NC\ cells on polylysine-treated glass
20 mM-NaCl alone 20 mM-NaCl + BSA
Central Centralarea Lamella Vacuole area Vacuole
Measured value o-8o±o-o2 o-6o±o-oi o-66±o-oi i-o o-78±o-oiComputed: assum-ing BSA diffusesunder cell o-8o o-6o o-66 0-98 o-8o
Computed:assuming BSAdoes not diffuseunder cell — — — 1-20 i-oo
Computations based on the following refractive indices (n) and thicknesses. Membrane:n ,̂ = 1-38; thickness = 4 nm; cytoplasm: n, = 1-368; vacuole contents: n, = 1-333; water:n, = 1-333; separation of lipid bilayer of plasmalemma from glass: s = 52-5 nm; contentsequivalent to 10% sucrose solution: n, = 1-348; BSA: np = 1-368; cytoplasmic lamellathickness: o-i /*m; vacuole-plasmalemma separation: 0-105 /*m> IN A = 1-18.
DISCUSSION
Precise spatial correspondence between the phase-contrast and interference imagesproves that the larger dark images represent contractile vacuoles. This does not resolvethe origin of the thread-like black images, but their relationship with the vacuolesmakes it inescapable that they are fluid-filled membrane-bounded tubes, which areresponsible for filling the vacuoles. Such a system of collecting vesicles related to themain vacuole was described by Pappas & Brandt (1958) in an electron-microscopestudy of Amoeba proteus. McKenna (1973) concluded that the contractile vacuoleof A. proteus fragmented at systole into a large number of small vesicles, and specu-lated that these may give rise to collecting tubules and vesicles as each new cycle ofdiastole begins. This, however, does not happen in Acanthamoeba. Bowers & Korn(1973) demonstrated that, while the contractile vacuole is alkaline-phosphatase-positive, the adjacent small vesicles and also the plasmalemma are phosphatase-negative. This important result showed that the contractile vacuole does not give riseto the collecting tubules and does not become incorporated into the plasmalemma onreleasing its contents. Likewise, the collecting tubules do not give rise to the mem-brane of the contractile vacuole. Recently, Quiviger, de Chastellier & Ryter (1978)
Figs. 24, 25. Cell on methylated glass showing: thin interferometrically black lamella(arrow a); contractile vacuole (arrow b); tubules (arrow c). Note the changes in thetubules in the ~ i o s that elapsed between these photographs. Ax2 cells in 20 mM-NaCl. x 3500.
Figs. 26-29. Sequence showing the reversible effect on the interference image of anAx2 cell in 20 mM-NaCl on glass (Fig. 26); upon adding BSA (Fig. 27); replacingthis with 20 mM-NaCl (Fig. 28); and finally, returning to BSA again (Fig. 29). x 3500.
294 D. Gingell, I. Todd and N. Owens
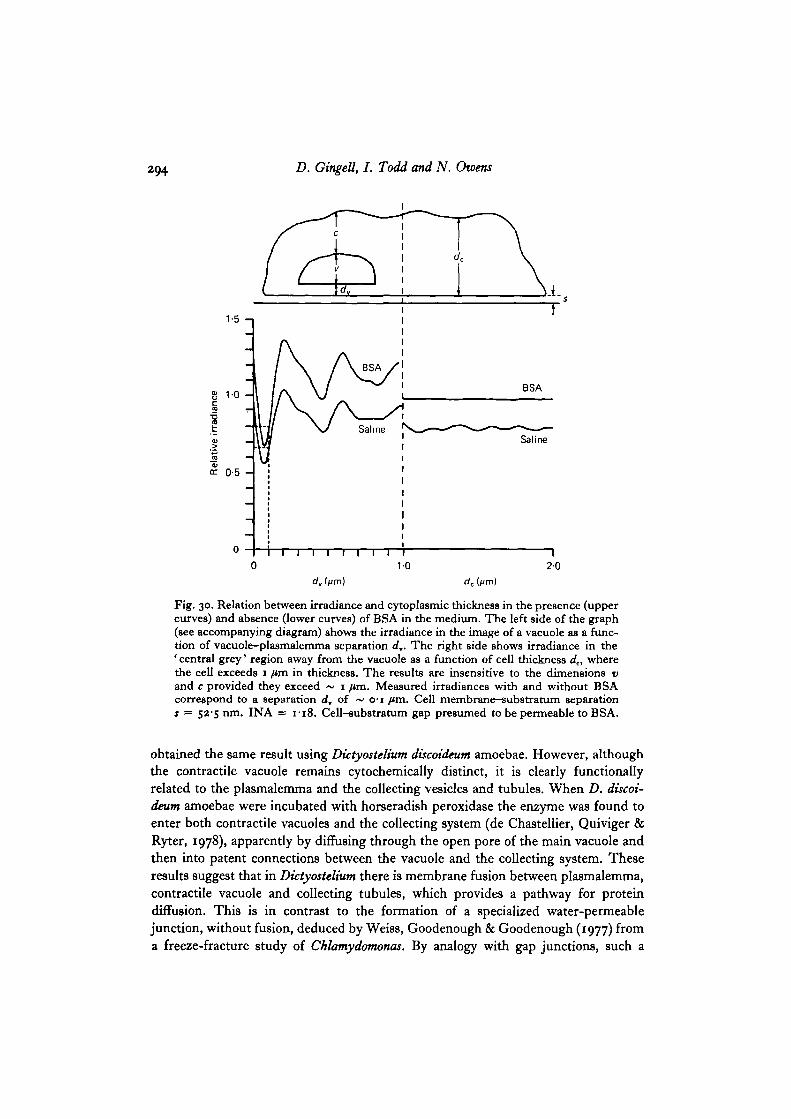
Fig. 30. Relation between irradiance and cytoplasmic thickness in the presence (uppercurves) and absence (lower curves) of BSA in the medium. The left side of the graph(see accompanying diagram) shows the irradiance in the image of a vacuole as a func-tion of vacuole-plasmalemma separation d,. The right side shows irradiance in the' central grey' region away from the vacuole as a function of cell thickness dc, wherethe cell exceeds 1 /un in thickness. The results are insensitive to the dimensions vand c provided they exceed ~ 1 fim. Measured irradiances with and without BSAcorrespond to a separation d, of ~ o-i ftm. Cell membrane-substratum separations = 52-5 run. INA = 1-18. Cell-substratum gap presumed to be permeable to BSA.
obtained the same result using Dictyostelium discoideum amoebae. However, althoughthe contractile vacuole remains cytochemically distinct, it is clearly functionallyrelated to the plasmalemma and the collecting vesicles and tubules. When D. discoi-deum amoebae were incubated with horseradish peroxidase the enzyme was found toenter both contractile vacuoles and the collecting system (de Chastellier, Quiviger &Ryter, 1978), apparently by diffusing through the open pore of the main vacuole andthen into patent connections between the vacuole and the collecting system. Theseresults suggest that in Dictyostelium there is membrane fusion between plasmalemma,contractile vacuole and collecting tubules, which provides a pathway for proteindiffusion. This is in contrast to the formation of a specialized water-permeablejunction, without fusion, deduced by Weiss, Goodenough & Goodenough (1977) froma freeze-fracture study of Chlamydomonas. By analogy with gap junctions, such a

Images of vacuoles 295
junction composed of intramembranous particles is unlikely to be permeable toprotein.
Our observations, while throwing no further light on these problems, providedirect evidence of a dynamic functional relationship between contractile vacuoles andthe collecting system. It shows the temporal sequence of changes that the electronmicroscope cannot show. It also shows that the collecting system in Dictyostelium isessentially tubular, a morphology easily misinterpreted in thin sections as beingvesicular.
Having demonstrated the spatial relationship between vacuoles and the largerdark images, the detailed origin of the interference images remains to be explained.We shall then consider what this implies about vacuole-plasmalemma interaction.Central to the image interpretation is the way in which irradiance is modified by BSA.This is because, in addition to providing constraints that the quantitative model mustexplain, the changes help to distinguish between several distinqt ways in which zero-order grey/black can arise. These are as follows: (1) from a thin cytoplasmic lamella,< o-i jim. Addition of BSA causes the image to brighten to near background as theupper cell surface/medium interface virtually disappears. (2) From the close approachof an intracellular vacuole to the cell surface, such that it remains separated from itby a uniform slab of cytoplasm not more than o-i /an thick. The image will be ex-tinguished in BSA if the vacuole approaches the upper surface,- but will remain if it isadjacent to the lower (substratum-apposed) surface. (3) From close-cell-glass contacts,which will brighten in BSA if the protein can penetrate beneath the cell but which willremain dark if it cannot.
The photometric results shown in Table 1 include data from Gingell & Vince(1982 a) on thin cytoplasmic lamellae. On the assumption that the NC4 cells areseparated from glass by a 52-5 nm region containing carbohydrate equivalent to10% sucrose, it can be seen that the measured and computed values are in excellentaccord with a model that allows diffusion of BSA underneath the cell. Since thicknessand refractive index have opposing effects on image brightness in this case, the thick-ness would be different if another value for the refractive index were chosen. Thecomputed values were arrived at in the following way. First, after arbitrarily assigningthe refractive index of the carbohydrate coat its thickness was adjusted empirically suchthat the computed and measured irradiances for thick cytoplasm (the central area inTable 1) were equal (Fig. 30). Then the irradiance for a thin lamella was computed,assuming that the carbohydrate-filled gap remained constant. The irradiance for avacuole making an extensive planar apposition with the cell membrane across a thinparallel slab of cytoplasm was calculated on the same basis, and finally calculationswere performed with BSA in the medium.
From Fig. 30 it can be seen that an excellent fit is obtained at o-i fim of cytoplasm,but we cannot completely exclude the possibility that the smaller dimension of~ 0-04 fim is the correct one. Analysis of a wide range of models representing thecell surface in terms of plasmalemma and carbohydrate coat thicknesses, and refrac-tive indices, strongly suggests that the common constant thickness of both the cyto-plasmic lamella and the thin layer of cytoplasm separating vacuoles from the plasma

296 D. Gingell, I. Todd and N. Owens
membrane falls within these bounds. Insufficient knowledge of the surface propertiesof Dictyostelium amoebae makes greater precision unrealistic at present.
It wilt be noted that the photometric data were obtained from cells spread onpolylysine-treated glass. This was done in order to maximize cell adhesion whilechanging solutions. It in no way influences the results. The separation of the plasma-lemma from the glass in 20 mM-NaCl (but not low salt concentration) is the same forclean glass and polylysine-treated glass and the images observed on the two surfacesare indistinguishable.
The optical analysis is interesting in several respects. There is a very large differencebetween the computed irradiances in BSA according to whether or not the protein isassumed to have> diffused beneath the cell into the carbohydrate region. Comparisonwith measured values strongly supports the contention that BSA does in fact getbeneath the cell! Figs. 26-29 clearly show (1) that the vacuole is less bright than thebackground in BSA and (2) that the central area of the cell is not brighter than back-ground. (Use Table 1 in conjunction with Figs. 26-29.) The fact that there is anaqueous diffusion path beneath cells attached to glass is evident from the fact thatchanges in the salt concentration of the bathing medium around cells, which havealready spread, can cause large and virtually instantaneous changes in the plasma-lemma-glass separation (Gingell & Vince, 19826). Further observations (Gingell &Todd, unpublished) show that rapid ionic diffusion can also take place beneath cellsadherent to polylysine-treated glass. This region contains cell-surface carbohydrate,which might act as a sieve retarding the passage of large molecules, but from the opticalanalysis it seems that BSA is not appreciably restrained.
Since BSA appears to diffuse beneath the cell, we can exclude the possibility thatthe dark interference image of vacuoles is due, even in part, to localized close contactof the plasmalemma with the glass. The fact that the dark vacuole images do notdisappear rules out the possibility that they are due to vacuoles closely approaching thefar side of the cell, though this was never considered likely. Since the cell obviouslydoes not suddenly thin down to a lamella, the vacuole images must be generated by aslab of cytoplasm, < o-i fim thick, between the plasmalemma and the vacuolemembrane.
The origin of the filamentous black images is less easy to analyse, since our methodof computation is not reliable for structures with such a high curvature. Their blackappearance may be due to interference across tubes of water within the cytoplasm, orto separation from the lower plasmalemma of the cell by a < o-i /im region ofcytoplasm.
The tubules and vacuoles remained visible in BSA. Even when the outline of thecell was practically invisible, this highly dynamic system of emptying and refillingvacuoles and tubules was observed for long periods. This confirms that it is part of thecell's osmoregulatory apparatus rather than a vesicular system related to pinocytosisand it underlines the functional distinction between these processes. Had the latterbeen the case, vesicles containing endocytosed BSA would soon have replaced thosefilled with water and the images would have disappeared.
The fact that the image of the vacuolar contact is of uniform irradiance shows that

Images of vacuoles 297
the vacuole membrane is deformed and flattened against the cell surface. The factthat such images show restricted movement with respect to the cell surface indicatesthat the vacuoles make an extensive planar adhesion with the cell surface, separatedby a thin layer of submembranous cytoplasm.
The correctness of our interpretation is strengthened by an unexpected observationmade in the course of the experiments on the effect of changing the salt concentration(Gingell & Vince, 19826). The vacuoles and associated tubules become brighter thanbackground as soon as the medium bathing the cells is changed from 20 mM-NaCl to~ 1 mM-NaCl, but their activity continues unabated. From fig. 9 {op. cit.) it can beseen that the cell membrane-glass separation under this condition exceeds 100 run.On inserting this separation into the vacuole model and assuming that the surfacecarbohydrate is appropriately diluted (fig. 8, op. cit.), we find that a relative irradianceof 1 -4 is predicted, in satisfactory agreement with our qualitative observation.
Thus we deduce a situation analogous to that reported by Gingell & Vince (1982 a)where Dictyostelium amoebae produce a cytoplasmic lamella of the same thickness byan active contractile mechanism. A constant, though non-planar, vacuole-plasma-lemma separation can be seen in a published electron micrograph of Dictyostelium(Ryter & Hellio, 1980). Our observations leave open the question of whether there isa preferential interaction between vacuoles and the surface of the cell in contact withthe substratum, as opposed to free surfaces. The latter would be difficult to detectby interferometry and further study by transmission electron microscopy is indicated.
The planar deformation of vacuoles near the glass interface implies that adhesiveforces pull the opposed membranes together, as argued in the case of the thin cyto-plasmic lamella. An electrostatic/electrodynamic balance of force in the intracellularmilieu at ~o-i /im is out of the question. The attraction is likely to be short-range,implicating macromolecular connections between the surfaces capable of shorteningor interdigitating to bring them into a constant separation. The zip model involvingactin filaments, proposed by Gingell & Vince (1982 a) to explain thin lamella forma-tion, could equally well apply to vacuole-membrane interaction.
An interesting possibility arises, that adhesive membrane-membrane appositionsinvolving intracellular organelles may follow a similar pattern. Despite the smallseparation we have described; the contractile vacuoles of Dictyostelium as well asAcanthamoeba (de Chastellier et al. 1978) ultimately fuse with the plasmalemma.Interference images, which may possibly represent a later stage in this process, are therarely seen areas of background irradiance coincident with vacuoles shown in fig. 24of Gingell & Vince (1982 a). Reference to Fig. 30 shows that this is the irradianceexpected as the intervening cytoplasm thins to zero. Membrane fusion cannot proceedbefore intercalated membrane proteins move laterally away from the contact zone(Lawson et al. 1977) and it is possible that lateral movement also carries membrane-associated actin aside, simultaneously on both apposed membranes, thereby creatinga local region wherein an hiatus in the thin cytoplasmic layer allows the bilayers tocome into contact as an essential prelude to fusion (Gingell & Ginsberg, 1979).
D.G. wishes to acknowledge support from the Science Research Council.

298 D. Gingell, I. Todd and N. Owens
REFERENCES
BOWERS, B. & KORN, E. D. (1973). Cytochemical identification of phosphatase activity in thecontractile vacuole of Acanthamoeba castellanii. J. Cell Biol. 59, 784-791.
DE CHASTELLIER, C. D., QUIVIGER, B. & RYTER, A. (1978). Observations on the functioning ofthe contractile vacuole of Dictyostelium discoideum with the electron microscope. J. Ultra-struct. Res. 6a, 220-227.
GINGELL, D. (1981). The interpretation of interference-reflection images of spread cells:significant contributions from thin peripheral cytoplasm. J. Cell Sci. 49, 237-247.
GINGELL, D. & GINSBERG, L. (1979). Problems in the physical interpretation of membraneinteraction and fusion. In Membrane Fusion, Cell Surface Reviews, vol. 5 (ed. G. Poste & G. L.Nicolson), pp. 791-883. Amsterdam: North-Holland.
GINGELL, D., TODD, I. & HEAVENS, O. S. (1982). Quantitive interference microscopy: effect ofaperture, Optica Acta (In Press).
GINGELL, D. & VINCE, S. (1982a). Substratum wettability and charge influence the spreadingof Dictyostelium amoebae and the formation of ultrathin cytoplasmic lamellae. J. Cell Sci.54, 255-285.
GINGELL, D. & VINCE, S. (19826). Cell-glass separation depends on salt concentration andvalency: measurements on Dictyostelium amoebae by finite aperture interferometry. J. CellSci. 54, 299-310.
IZZARD, C. A. & LOCHNER, L. R. (1976). Cell to substrate contact in living fibroblasts: aninterference reflexion study with an evaluation of the technique. J. Cell Sci. 21, 129-159.
LAWSON, D., RAFF, M. C, GOMPERTS, B., FEWTRELL, C. & GILULA, N. B. (1977). Molecularevents during membrane fusion. A study of exocytosis in rat peritoneal mast cells. J. Cell Biol.73, 242-259.
MCKENNA, J. A. (1973). Membrane recycling: vesiculation of the amoeba contractile vacuoleat systole. Science, N. Y. 179, 88-90.
PAPPAS, G. & BRANDT, P. W. (1958). The fine structure of the contractile vacuole in Ameba.J. biochem. biophys. Cytol. 4, 485-487.
QUIVIGER, B., DE CHASTELLIER, C. & RYTER, A. (1978). Cytochemical demonstration of alkalinephosphatase in the contractile vacuole of Dictyostelium discoideum. J. Ultrastruct. Res. 6a,228-236.
RYTER, A. & HELLIO, R. (1980). Electron-microscope study of Dictyostelium discoideum plasmamembrane and its modifications during and after phagocytosis. ,7. Cell Sci. 41, 75-88.
WEISS, R. L., GOODENOUGH, D. A. & GOODENOUGH, U. W. (1977). Membrane particle arraysassociated with the basal body and with contractile vacuole secretion in Chlamydomonas.J. Cell Biol. 72, 133-143.
{Received 20 July 1981)
























![Regulation of the intracellular Ca2+. Regulation of intracellular [H]:](https://img.pdfslide.us/doc/110x75/5a4d1b717f8b9ab0599b56a5/regulation-of-the-intracellular-ca2-regulation-of-intracellular-h.jpg)




