Embed Size (px)
Citation preview

Faculty of Bioscience Engineering
Academic year 2014–2015
Inter- and intra-species leaf trait variability in a planted
rainforest in Yangambi (D.R. Congo)
Mumbanza Mundondo Francis
Promotors: Prof. dr. ir. Hans Verbeeck and Prof. dr. ir. Pascal Boeckx
Tutor: Ir. Marijn Bauters
Master’s dissertation submitted in partial fulfillment of the requirements for the
degree of
Master of Science in Environmental Sanitation

i
DEDICATION
I dedicate this work to my parents, Jérôme-Émilien and Marie-José, and to all my brothers and
sisters.

ii
ACKNOWLEDGEMENTS
I’m greatly indebted to Prof. dr. ir. Hans Verbeeck, Prof dr. ir. Pascal Boeckx and Ir. Marijn
Bauters under whose leadership and supervision these investigations were conducted. Thank you
for giving me the opportunity to work on this exciting master research and for your untiring
guidance and criticisms throughout this work.
My sincere thanks go to Prof. dr. ir. Peter Goethals, the promoter of Master of Science in
Environmental Sanitation programme and to Veerle Lambert and Sylvie Bauwens, Centre of
Environmental Science and Technology coordinators for all their encouragement, guidance and
advices in the course of these studies. I am also deeply grateful to the University of Kinshasa, my
employer for granting me a study leave to undertake these studies.
I would also wish to extend my appreciation to all my family members for supporting me during
these studies. Finally, I would like to express my gratitude to all my fellow students and friends
who in one way or another contributed to the success of these studies especially, Noel
N’guessan, Colins Chi, Blaise Ntirumenyerwa, Yves-Daddy Botula, Velma Kimbi, Chantal
Nikuze and Sylvia Ambali.
Thank you for everything and may the Almighty God bless you all!

iii
ABSTRACT
One of the greatest challenges in plant community ecology is to elucidate the patterns of species
co-existence. Functional trait values have been often used to provide an insight into various
mechanisms that form the basis of plant species co-existence. Nowadays, there is a growing body
of evidence that not only inter-specific plant trait variability has a significant influence on the
dynamics and functioning of ecosystems, but also intra-specific plant trait variability.
In this study, the role of intra-specific trait variability in mediating inter-specific interactions was
examined among twelve co-existing species (target species) in a tropical plantation, consisting of
monoculture plots and two-species mixture plots. Nine functional traits were measured and both
single species trait and multiple trait analyses were used to quantify trait variation within and
between these co-existing tree species. The following specific questions were addressed: Is the
relative contribution of intra-specific trait variation to the overall trait variation more important
than that of inter-specific trait variation? Are the functional trade-offs and strategies adopted by
the target species at the intra-specific level similar to that at the inter-specific level? Are there
significant differences in trait values and/or in multivariate trait distributions between target
species in monocultures and in two-species mixtures?
The results obtained revealed a non-negligible contribution of the intra-specific trait variation to
the overall functional trait variability for the majority of the traits examined. In particular, the
intra-specific trait variability was higher than the inter-specific variability for the traits height
(H), diameter at breast height (DBH) and leaf phosphorus content (LPC). The results also
showed that the co-existing target species in this planted tropical forest deploy more or less the
same functional trade-offs and strategies at both the inter-specific level and the intra-specific
level. Finally, some significant differences in trait values and in multivariate trait distributions
were detected between target species in monocultures and in two-species mixtures for nine
species out of the twelve investigated. This latter finding seems to indicate a probable role played
by phenotypic plasticity in shaping species co-existence in this planted tropical forest.
The results of this study point to the fact that intra-specific plant trait variability may play a
determinant role in shaping species co-existence under certain circumstances. Hence, it should
not be systematically neglected in quantitative functional trait-based analyses. The decision on
whether or not to ignore the intra-specific trait variability should be made on a case-by-case basis
taking into account the trait, the species and the system under investigation.

iv
LIST OF FIGURES
Figure 2.1 Seven plant organs or whole-plant properties and their functional significance……. 16
Figure 2.2 Relationship between the number of traits and the ability to predict and explain
variation in community composition……………………………………………………….. 17
Figure 2.3 Hypothetical changes in the magnitude of inter-specific (INTER) and intra-specific
(INTRA) trait variability over geographical scales………………………………………… 22
Figure 3.1 Study site localization……………………………………………………………….23
Figure 3.2 Monthly average (from 2000-2008) for precipitation and temperature in the Yangambi
region……………………………………………………………………………………….. 24
Figure 3.3 Variance partitioning using multi-trait approach……………………………………. 29
Figure 4.1 Variance decomposition in inter-specific and intra-specific contributions for single-
trait and multi-trait patterns……………………………………………………………….... 32
Figure 4.2 Multidimensional structure within the trait space: Inter-specific and intra-specific
trade-offs…………………………………………………………………………………….34
Figure 4.3 Multidimensional structure within the trait space: Intra-specific trade-offs……….... 35
Figure 4.4 Dispersion of species and individuals of each species in the trait space…………….. 36

v
LIST OF TABLES
Table 2.1 Association of plant functional traits with 1) plant responses to four classes of
environmental change (i.e. “environmental filters”), 2) plant competitive strength and plant
“defense” against herbivores and pathogens (i.e. “biological filters”) and 3) plants effects on
biogeochemical cycles and disturbance regimes……………………………………………10
Table 2.2 Summary of nine studies that have explicitly measured inter-specific and intra-specific
variation in functional traits…………………………………………………………………19
Table 3.1 Experimental design of the Yangambi arboretum……………………………………. 25
Table 4.1 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species A. congolensis in the monoculture population and the two-
species mixture population…………………………………………………………………. 37
Table 4.2 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species L. trichilioides in the monoculture population and the two-
species mixture population…………………………………………………………………. 38
Table 4.3 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species M. africana in the monoculture population and the two-
species mixture population…………………………………………………………………. 39
Table 4.4 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species M.excelsa in the monoculture population and the two-
species mixture population…………………………………………………………………. 39
Table 4.5 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. oleosa in the monoculture population and the two-species
mixture population…………………………………………………………………………..40
Table 4.6 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. soyauxii in the monoculture population and the two-
species mixture population…………………………………………………………………. 41
Table 4.7 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. tessmannii in the monoculture population and the two-
species mixture population…………………………………………………………………. 42

vi
Table 4.8 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species S. tetrandra in the monoculture population and the two-
species mixture population…………………………………………………………………. 43
Table 4.9 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species E. cylindricum in the monoculture population and the two-
species mixture populations………………………………………………………………....43
Table 4.10 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species G.cedrata in the monoculture population and the two-
species mixture populations…………………………………………………………………45
Table 4.11 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species P. macrophylla in the monoculture population and the two-
species mixture populations…………………………………………………………………46
Table 4.12 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species P.elata in the monoculture populations and the two-species
mixture populations………………………………………………………………………… 48
Table 4.13 Between analysis tests for detecting the significance of grouping for the BPCAs
performed on each species…………………………………………………………………..50
Table 4.14 Between analysis tests for detecting the segregation of monoculture populations from
two-species mixture populations for the grouping of target species with more than two
population…………………………………………………………………………………... 51

vii
LIST OF ACRONYMS
Amax Photosynthetic rates
ATP Adenosine triphosphate
BPCA Between-group Principal Component Analysis
C Carbon
CO2 Carbon dioxide
DBH Diameter at Breast Height
DRC Democratic Republic of Congo
EA-IRMS Elemental Analyzer- Isotope Ratio Mass Spectrometer
FAO Food and Agriculture Organization
Ha Hectares
H Height
INERA Institut National pour l'Etude et la Recherche Agronomiques
IPCC Intergovernmental Panel on Climate Change
LDMC Leaf Dry Matter Content
LES Leaf Economics Spectrum
LMA Leaf Mass per unit Area
LNC Leaf Nitrogen Content
LPC Leaf Phosphorous Content
LWC Leaf Water Content
MECNT Ministère de l’Environnement, Conservation de la Nature et Tourisme
N Nitrogen
P Phosphorus
RAINFOR Rede Amazônica de Inventários Florestais, Red Amazónica de Inventarios
Forestales
Rd Leaf respiration rate
RDC République Démocratique du Congo
SLA Specific Leaf Area
SVP Spatial Variance Partitioning
UN-REDD United Nations Programme on Reducing Emissions from Deforestation and
forest Degradation
WPCA Within-group Principal Component Analysis
WUE Water Use Efficiency

viii
TABLE OF CONTENTS
DEDICATION ................................................................................................................................. i
ACKNOWLEDGEMENTS ............................................................................................................ ii
ABSTRACT ................................................................................................................................... iii
LIST OF FIGURES ....................................................................................................................... iv
LIST OF TABLES .......................................................................................................................... v
LIST OF ACRONYMS ................................................................................................................ vii
TABLE OF CONTENTS ............................................................................................................. viii
CHAPTER ONE: INTRODUCTION ............................................................................................. 1
1.1 Background ........................................................................................................................... 1
1.2 Problem statement ................................................................................................................. 3
1.3 Research objectives ............................................................................................................... 3
1.4 Research hypotheses ............................................................................................................. 4
CHAPTER TWO: LITERATURE REVIEW ................................................................................. 5
2.1 Importance of forest ecosystems ........................................................................................... 5
2.2 Structure and distribution of the world’s forests ................................................................... 5
2.2.1 Structure.......................................................................................................................... 6
2.2.2 Distribution ..................................................................................................................... 6
2.3 The Congolese forest ............................................................................................................. 7
2.4 Plant functional traits ............................................................................................................ 8
2.4.1 Key plant functional traits often measured in trait based ecology approach .................. 8
2.4.1.1 Whole plant traits ..................................................................................................... 9
2.4.1.2 Wood traits ............................................................................................................. 11
2.4.1.3 Leaf traits ............................................................................................................... 11
2.4.2 Dimensionality of plant functional traits ...................................................................... 15
2.4.3 Intra- specific variability of plant functional traits ....................................................... 18
2.4.3.1 Sources of intra-specific plant trait variability....................................................... 20
2.4.3.2 Structure of intra-specific plant trait variability ..................................................... 20
2.4.3.3 Magnitude of intra-specific plant trait variability .................................................. 21
CHAPTER THREE: MATERIALS AND METHODS ............................................................... 23

ix
3.1 Study location ...................................................................................................................... 23
3.2 Experimental set-up............................................................................................................. 24
3.3 Leaf sampling ...................................................................................................................... 26
3.4 Functional traits and measurement ...................................................................................... 26
3.5 Statistical analysis ............................................................................................................... 27
3.5.1 Variance partitioning .................................................................................................... 27
3.5.2 Main functional trade-offs and strategies ..................................................................... 29
3.5.3 Trait differences between target species in monocultures and two-species mixtures .. 30
3.5.4 Statistical packages ....................................................................................................... 30
CHAPTER FOUR: RESULTS ..................................................................................................... 31
4.1. Variance partitioning .......................................................................................................... 31
4.2 Main functional trade-offs and strategies ............................................................................ 33
4.3 Trait differences between target species in monocultures and two-species mixtures ......... 37
4.3.1 Single trait approach ..................................................................................................... 37
4.3.2 Multi-trait approach ...................................................................................................... 50
CHAPTER FIVE: DISCUSSION ................................................................................................. 53
5.1. Variance partitioning .......................................................................................................... 53
5.2 Main functional trade-offs and strategies ............................................................................ 54
5.3 Trait differences between target species in monocultures and two-species mixtures ......... 55
5.3.1 Single trait approach ..................................................................................................... 55
5.3.2 Multi-trait approach ...................................................................................................... 57
CHAPTER SIX: GENERAL CONCLUSION AND RECOMMENDATIONS .......................... 58
6.1 General conclusion .............................................................................................................. 58
6.2. Recommendations .............................................................................................................. 59
REFERENCES ............................................................................................................................. 60

1
CHAPTER ONE:
INTRODUCTION
1.1 Background
Forests, which are the dominant terrestrial ecosystem on Earth, play an essential role in the
planet’s life support system (Bonan, 2008). Besides providing basic ecological and
environmental services, they also supply humanity with valuable goods and services (Pearce,
2001). It is therefore necessary to constantly monitor forest ecosystems in order to comprehend
the various processes and factors that determine their ecology, function and patterns. This, in
turn, will allow devising proper management strategies to continue meeting the needs of a still
growing human population, especially in this era of global environmental change.
It is in this perspective that one of the major goals of forest ecology is the knowledge of the
mechanisms that drive life history variation among co-existing tree species (Westoby et al.,
2002). This is crucial in the understanding of forest successional dynamics following natural
and/or human induced disturbances. Qualitative life-history classifications, like shade tolerance
groupings, were commonly used to describe functional variation among tree species in the past
(Bazzaz and Pickett 1980; Swaine and Whitmore, 1988). However, these types of classifications
proved to be subjective and inefficient in describing variation observed among tree species in
very complex and diverse ecosystems such as tropical forests (Welden et al., 1991). This
situation prompted forest ecologists to switch towards the identification and quantification of
functional traits. These can constitute a good mechanistic basis for life-history strategies when
scaled to the whole-plant or species level (Ackerly et al., 2000; Westoby et al., 2002).
Plant functional traits are morphological, physiological, phenological, chemical features
measurable at individual level that represent ecological strategies. They determine how plants
respond to environmental factors, affect other trophic levels and influence ecosystem properties
(Perez-Harguindeguy et al., 2013). As such, their variability is an indication of the complex
ecosystem functional diversity, and they are often used to describe functional divergence among
various co-existing species within an ecosystem.
Over the past years, substantial functional trait datasets have been compiled, and their analysis
has made it possible to elucidate some of the leading dimensions of functional trait variation
among co-existing tree species in different forest ecosystems (Ackerly et al., 2000; Westoby et
al., 2002, Patiño et al., 2012). These dimensions include particularly tree size metrics,
reproductive traits, leaf-level physiological and morphological traits, and wood anatomical and
chemical traits (Thomas, 1996; Westoby et al., 2002; Wright et al. 2004; Chave et al., 2009).
Based on these functional trait dimensions, the relationships between different functional traits
on one hand and the link between traits and tree demography on the other hand can be built for
various forest ecosystems.

2
The temporal as well as the spatial structure and diversity in forest communities can be
characterized by the distribution of functional traits of the individuals they comprise. This is
because there are direct links between traits and the functioning of organisms. Trait distributions
constitute therefore an attractive means of looking into how forest communities are associated
and how they influence ecosystem processes (Cornwell and Ackerly, 2009). That is why there
has been a growing interest in describing the distribution of traits in plant communities and the
underlying processes responsible of these distributional patterns such as environmental filtering
or niche differentiation (Kraft et al., 2008; Baraloto et al., 2012; Violle et al., 2012).
The vast majority of research involving the measurement of trait distributions focuses on species
mean trait values. The emphasis is put on inter-specific trait differences between co-existing
species, while the intra-specific variability of plant traits is largely ignored. The assumption
behind this approach has been that intra-specific trait variation is negligible compared to
variation among species (Garnier et al., 2001; Baraloto et al., 2010). It is, however, known that
depending on plant genetic, developmental factors as well as changing environmental conditions,
plant functional traits can be quite variable within species (Violle et al., 2012), and that quite
important phenotypic variation for a range of traits can exist in plant population even within very
small areas (Linhart and Grant, 1996).
More and more studies are now pointing to the fact that intra-specific trait variation may account
for a quite large percentage of the total trait variation in some instances (Jung et al., 2010;
Messier et al., 2010, Albert et al., 2010). Similarly, intra-specific trait variation is believed to
play an important role in a broad range of ecological processes and properties including
resistance to disturbances, competition, co-existence or productivity (Hughes and Stachowicz,
2004; Fridley et al. 2007; Clark et al., 2010; Kotowska et al., 2010). Understanding to which
extent species co-existence and plant associations are mediated by intra-specific variability is
therefore necessary. This implies knowledge about trait variation distribution mechanisms over
ecological and spatial scales.
In this vein, Kang et al. (2014) noted that leaf and wood traits were varying differently in
relation to ecological scales in a subtropical forest of China. This brought them to suggest that
trait variability is tissue-specific. In a previous study, Messier et al. (2010) had reported that the
variation of leaf mass area and leaf dry matter content was more or less uniformly distributed
across six nested ecological scales in lowland rainforests of Panama. Earlier, Albert et al. (2010)
had concluded that intra-specific variability in the functional traits tree height, specific leaf area,
leaf dry matter, leaf carbon and leaf nitrogen contents of herbaceous species in the French Alps
was species-specific and trait-specific. These findings tend to suggest that the trait type (physical
or chemical traits), the ecological scale and the species identity influence the structure and the
extent of functional variability.

3
1.2 Problem statement
At regional scale, intra-specific trait variability is recognized to be mainly due to biogeographical
processes such as migration, climatic fluctuations and isolation. These processes are responsible
for the divergence in distant populations that enhances intra-specific variability (Moreira et al.,
2012). At local scales, trait variability in general is generated by local processes like
disturbances, heterogeneity in resource availability and species interaction (Moreira et al., 2012).
Species response to biotic and abiotic filters, which influence the population dynamics and the
community structure, is much dependent on this local trait variability (Jung et al., 2010; Bolnick
et al., 2011).
Studies seeking to evaluate the relative importance of intra-specific versus inter-specific trait
variability in shaping plant community are often conducted at large geographical scales or at
local scales but along very contrasting environmental gradients such as altitudinal or
precipitation gradients. Besides providing information on the patterns of intra-and inter-specific
variation, some of these studies have also tried to elucidate the main factors responsible for the
diversity of traits within a community. Despite the assumption that intra-specific trait variability
increases with decreasing spatial scale and environmental gradients (Albert et al., 2011), less
information has accumulated on the patterns of intra-specific trait variation and the factors
underlying trait variability within communities at small spatial scales with fairly homogeneous
environmental conditions.
The Yangambi arboretum in the tropical rainforest, in the North Eastern part of the Democratic
Republic of the Congo provides an excellent framework to study the patterns and distribution of
intra-specific plant trait variability at small spatial scale. This is because all plots in the
arboretum enjoy the same climatic conditions, and native tree species with different functional
types were planted there in both monocultures (target species) and two-species mixture plots
(target species and admixed species) in 1940. This can allow shedding more light on how intra-
specific trait variation impacts on the co-existence and diversity of species in this planted tropical
forest.
1.3 Research objectives
The main objective of this study was to explore how intra-specific variability can mediate small
spatial-scale inter-specific interactions in a planted tropical rainforest. Advantage was taken of
the presence of this unique experimental stand in Yangambi to quantify the variation between
and within twelve co-existing target tree species with respect to nine functional traits and explore
the following more specific objectives:
1) Determine whether the relative contribution of intra-specific trait variation to the overall trait
variation was more important than that of inter-specific trait variation;
2) Determine whether the functional trade-offs and strategies adopted by the target species at the
intra-specific level were similar to that at the inter-specific level;

4
3) Determine whether there were significant differences in trait values and/or in multivariate trait
distributions between target species in monocultures and in two-species mixtures.
1.4 Research hypotheses
The following hypotheses were made as part of this study:
1) The relative importance of inter-specific variation may be reduced for most traits due to the
small environmental variation in the arboretum and the biotic interaction between the individuals
of the target species at the neighborhood scale. Therefore, the intra-specific trait variation is
expected to account for a non-negligible fraction of the overall variation for most traits;
2) Individuals of the target species may adopt the same functional tradeoffs and strategies at the
intra-specific level as at inter-specific level to favor their co-existence. Therefore, the trait
variation structure within species should reflect the one that can be observed between species;
3) Resource competition between individuals of the target species and those of the admixed
species may favor niche partitioning among these competing individuals in two-species mixtures.
This may result in the occurrence of phenotypic plasticity, leading to a shift in the trait values
and/or in the multivariate trait distributions of target species in mixtures as compared to
monocultures.

5
CHAPTER TWO:
LITERATURE REVIEW
2.1 Importance of forest ecosystems
The importance of forest ecosystems to human well-being is undeniable. They provide a large
array of services to humankind including maintenance of biodiversity, nutrient cycling, climate
regulation, soil stabilization and erosion control, as well as economic, cultural and recreational
values (Krieger, 2001). In addition to that, many people across the world depend on forests for
products such as food, fiber, medicine, timber, fuel wood and charcoal, and for their income
(Pierce, 2001).
The role of forest ecosystems as both one of the major contributors to the global carbon (C) cycle
and an essential determinant of global climate patterns and processes is undeniably one of the
most critical scientific issues in these times of climatic change. In that respect, tropical forests
are particularly at the heart of the matter as they were shown to play an excessively large role in
the global C cycle. As a matter of fact, although tropical forests only represent 7-10 % of the
total Earth’s surface, they store more aboveground C than any other biome (IPCC, 2007).
Currently, the world’s forest C reserves are evaluated at 861 ± 66 petagrams of carbon (Pg C).
Of this total stock, 383 ± 30 Pg C (44 %) are present in soil (to 1-m depth), 363 ± 28 Pg C (42
%) in live biomass (above and below ground), 73 ± 6 Pg C (8 %) in deadwood, and 43 ± 3 Pg C
(5 %) in litter. As far as the geographical distribution is concerned, 471 ± 93 Pg C (55 %) is
stored in tropical forests, 272 ± 23 Pg C (32 %) in boreal forests, and 119 ± 6 Pg C (14 %) in
temperate forests (Pan et al., 2011).
Pan et al. (2011) also estimated the gross C emissions from tropical deforestation and
degradation at 2.9 ± 0.5 petagrams of carbon per year (Pg C/year) and the sinks due to tropical
re-growth at 1.6 ± 0.5 Pg C/year on a global scale. These figures clearly indicate that the rates of
tropical deforestation and degradation outpace tropical forest growth, making tropical forests a
net C source of 1.3 ± 0.7 Pg C/year. C fluxes from tropical forests are therefore considered as a
substantial contributor to increasing atmospheric carbon dioxide (CO2), representing
approximately 12-17 % of global anthropogenic C emissions (van der Werf et al. 2009).
Avoiding deforestation and forest degradation should consequently be considered as an
imperative duty.
2.2 Structure and distribution of the world’s forests
The main drivers controlling the structure and distribution of forests are environmental factors.
Because tree species adjust themselves to climate, resources gradients, disturbance regimes and
forest dynamics, complex geographical patterns in forest assemblage and structure are formed
(Pan et al., 2013).

6
2.2.1 Structure
Forest structure can be defined as the manner in which tree attributes are distributed within a
forest ecosystem (Gadow et al., 2012). Both the characteristics of individual structural elements
and the spatial (horizontal and vertical) patterns of structural elements are important in the
description of forest ecosystems (Franklin et al., 2002). Alongside with other biotic and abiotic
features, tree structural characteristics constitute the basis of forest ecosystem functioning and
processes (Pan et al., 2013).
Some individual structural elements including in particular leaf area, life form, branch
arrangement or soil depth can have a great influence on the basic functions of trees and on
competition for resources. Similarly, forest vegetation dynamics may be portrayed through
spatial pattern such as the size and distribution of trees, spatial distribution of biomass within a
stand, canopy layers, and gaps (Pan et al., 2013). The main drivers responsible for the alteration
of forest structure are disturbances. They produce landscape mosaics and establish the
prerequisites for successional dynamics and structural development (Swanson et al., 2011).
2.2.2 Distribution
The limits of forests are defined by several processes and factors. Forest vegetation primarily
follows the geographical distribution of climate. Depending on the seasonality of temperature
and rainfall, forests of different forms and growth adapt themselves to specific geographic
regions (Woodward et al., 2004). This global pattern can however be influenced at regional
and/or local scale by topography, soil types, and other local environmental factors through the
creation of microclimates (Pan et al., 2013). Due to this close correlation between the
geographical patterns of global forest and climate, global forest distributions have been quite
often explained by climate variables such as temperature and precipitation (Pan et al., 2013).
Global forests are also greatly impacted by a number of natural and human induced disturbances
which create complex mosaics of forest distribution and high landscape-scale diversity. Land use
change, particularly agriculture is to blame for most of the recent loss of forest, accounting for
nearly 80 % of deforestation across the world (Gibbs et al., 2007). Climatic change due to
anthropogenic green gas emission is another factor responsible for shaping forests globally. It
induces tree species migration resulting in new geographic distributions of forests (Pan et al.,
2013), and also disrupts disturbance regimes by increasing the frequency and/or the intensity of
wildfires, windstorms, or insect outbreaks (Dale et al., 2001).
The world’s forest covers an area estimated at 4.03 billion hectares (ha). This represents
approximately 30 % of Earth’s total land area (FAO, 2010). More widespread in the Northern
Hemisphere where larger land masses are found, 31 % of the world’s forested area is
encountered in Asia (including Asian Russia). South America with 21 % of the Earth’s total
forested area comes in the second place. Then, follow Africa and North and Central America
with 17 % of the Earth’s total forested area each. Europe and Oceania come last with 9 % and 5

7
% respectively of the world’s forest (FAO, 2010). It is also worth mentioning that 5 % of forests
in the world are commercial plantations (Pan et al., 2013).
The tropics account for 44 % of world’s forests, with tropical rain forests constituting the
dominant type. Tropical rain forests cover an area estimated at 600 million ha. The largest
portion of these rain forests is found in South America with 53 %. 27 % of these forests are
encountered in Africa and the remaining 20 % is shared between Asia and Oceania (Butler,
2014).
2.3 The Congolese forest
The Democratic Republic of Congo (DRC) has approximately 145 million hectares of forest
cover. This represents about 50 % of the humid tropical forests of the Congo Basin, the second
largest tropical forest bloc in the world after the Amazon. 62 % the DRC territory is made up of
forests distributed as follows: 37 % of rain forest cover accounting for almost half of the African
continent rain forests, 19 % of dry forests, 4% of swamp forests, and 2 % of mountain forests.
Much of these forests belongs to the domain of dense and humid forests of low and medium
altitude, and is part of the Guinean-Congolese massif (MECNT, RDC, 2009).
The Congolese forest sequesters a C stock estimated at about 27 Pg C. This represents 60 % of
the total C stock of the Congo basin countries (Gibbs et al., 2007). The presence of this forest
helps in the maintenance of major rain cycles sustaining rivers such the Congo River. This forest
also harbors many endemic species of wildlife and flora (MECNT, RDC, 2009).
The annual rate of deforestation in the country is relatively modest compared to other tropical
countries of Southeast Asia and Latin America. However, this rate has been increasing rapidly
over the past decades. Established at around 0.11 % between 1990 and 2000, this rate almost
doubled during the period 2000 and 2005 in which it was estimated at 0.22 % (Ernst, 2013). This
is equivalent to a loss of almost 406000 ha of forest each year, constituting the highest
deforestation annual rate among all the countries of the Congo Basin. During the same period
2000-2005, the DRC registered also the highest rate of forest degradation among all the Congo
Basin countries (Ernest, 2013). The principal direct causes of deforestation and forest
degradation are slash-and-burn agriculture and artisanal logging whereas demographic growth
and poor governance are the key underlying factors (MECNT, DRC/UN-REDD, 2012).
The swift degree of deforestation and forest degradation which the Congolese forest is
experiencing is expected to result in an important loss of biodiversity and serious C emissions.
Consequently, not only the timber production will be threatened but also the environment as a
whole. The development of mitigation measures at the local level has to be anticipated in order
to minimize the effects of these changes on the ecosystem functioning. This requires the
understanding of potential relationships between species diversity and ecosystem functioning. It
is within this context that studies like the present one fall.

8
2.4 Plant functional traits
Traditionally, species were grouped based on their common evolutionary history using
phylogenetic methods. This approach has had some limitations to adequately answer ecological
questions at the ecosystem, landscape or biome scales (Cornelissen et al., 2003). In recent years,
there has been a shift towards functional classification of species that links population,
community and ecosystem processes to the key traits that influence the performance of
organisms in terms of growth, metabolism or reproduction. These traits are referred to as
functional traits.
A trait is defined as any morphological, physiological or phenological feature measurable at the
individual level irrespective of the environment and/or the level of organization (Violle et al.,
2007). A plant functional trait can therefore be considered as any relevant characteristic that
influences plant response to the environment and has an impact on the ecosystem functioning
(Diaz and Cabido, 2001). In simple words, it is a trait that influences plant function.
From an environmental point of view, a plant trait can either be a response trait or an effect trait.
A response trait varies in reaction to changes in environmental conditions whereas an effect trait
represents the influence of the plant on environmental conditions, communities or ecosystem
properties (Violle et al., 2007). Functional response traits in particular are very important
because they determine plant growth, survival and reproductive success. As such they are central
to understanding variability in plant distribution, form, function and diversity.
To help answer questions related to the reciprocal phenomena mentioned above, namely the
responses of vegetation to environmental variation or changes (climate, atmospheric chemistry,
land use, disturbance regimes) and the impacts of vegetation on large scale environmental
parameters, plant species are clustered as functional types. Cornelissen et al. (2003) define
functional types as groups of plant species sharing similar functioning at the organismic level,
similar responses to environmental factors and/or similar roles in (or effects on) ecosystems or
biomes. It is because these species have in common the same assortment of key functional traits
that they present similarities.
2.4.1 Key plant functional traits often measured in trait based ecology approach
In functional ecology, many traits that have an impact on the functioning and structure of plant
communities are measured. A list of some critical plant functional traits and their association
with plant response to environmental changes, plant competitive strength and plant “defense”
against herbivores and pathogens and plant effects on biogeochemical cycles and disturbance
regimes as described by Cornelissen et al. (2003) is provided in Table 2.1. Here, we give an
account of a few of them that are commonly used in trait based ecology to understand the
variability in plant form, function and diversity.

9
2.4.1.1 Whole plant traits
Whole plant traits are often the reflection of whole plant investment, allocation and growth.
These traits are believed to play a crucial role in the mechanisms behind the co-existence of
competing species.
Plant size
Plant size, which is measured as mass, height or diameter, can considerably vary over the
lifetime of different individuals. It has a strong influence on the form, function and life history of
plants (Westoby et al., 2002). Plant height (H) represents the distance between the uppermost
part of the photosynthetic tissue and the ground whereas the diameter at breast height (DBH)
refers to the diameter of plant stem measure at breast level. H and DBH are often used as
measures of plant growth (Sumida et al., 2013). H particulary is associated with competitive
vigour and has been shown to correlate well with the aboveground biomass (Cornelissen et al.,
2003).
The allometric relation DBH-H is said to strongly influence the safety factor against buckling of
tree species (van Gelder et al., 2006) and has been often used to describe tree species strategies.
It has been for instance observed that co-existing tree species could differ in H at the same
diameters (Poorter et al., 2003) and in diameters at a specific DBH. Two reasons have been
proposed that may justify a smaller DBH when different co-existing tree species are compared.
In the first place, it is argued that species with a small DBH deploy a fast-growth strategy and, at
the same time, they present a real risk of stem breakage. Secondly, it is suggested that species
with narrow DBH have strong, high density wood as a way of compensation for their limited
basal thickening (Kooyman and Westoby, 2009). Since dense wood has generally higher
modulus of rupture, trees are consequently less susceptible to breakage. This means that for tree
species with narrow DBH, a higher investment in wood density might result in comparable
biomass cost of H gain across different stem-widening strategies (Kooyman and Westoby, 2009).
The influence of other traits by plant size has been recognized in many studies. For example,
strong correlations across species were established between leaf size, specific leaf area (SLA)
and maximum H (Fonseca et al., 2000). Similarly, leaf size, wood density and seed size were
found to correlate with maximum H (Cornwell and Ackerly, 2009). Correlations between shade
tolerance as a reflection of successional status, wood density and maximum H have been also
reported (Falster and Westoby, 2005).

10
Table 2.1 Association of plant functional traits with 1) plant responses to four classes of environmental change (i.e. “environmental
filters”), 2) plant competitive strength and plant “defense” against herbivores and pathogens (i.e. “biological filters”) and 3) plants
effects on biogeochemical cycles and disturbance regimes (Reproduced from Cornelissen et al., 2003). *: Association established; ?:
Probable association.
Plant responses to environmental changes
(Environmental filters)
Plant competitive strength and
plant defence against herbivores
and pathogens (Biological filters)
Plant effects on biogeochemical
cycles and disturbance regimes
Climate
response
CO2
response
Response to
soil resources
Response to
disturbance
Competitive
strength
Plant defence/
protection
Effects on
biogeochemical
cycles
Effects on
disturbance
regimes
Whole-plant traits
1. Growth form * * * * * * * *
2. Life form * * * * * * *
3. Plant height * * * * * * * *
4. Clonality * ? * * * ?
5. Spinescence * ? * * ?
6. Flammability ? * ? * *
Leaf traits
1. Specific leaf area * * * * * *
2. Leaf size * ? * * * *
3. Leaf dry matter content * ? * * * *
4. Leaf N and P concentration * * * * * * *
5. Physical strength of leaves * ? * * * * *
6. Leaf life span * * * * * * * *
7. Leaf phenology * * * * *
8. Photosynthetic pathway * * *
9. Leaf frost resistance * * *
Stem and belowground traits
1. Stem specific density * ? ? * * * *
2. Twig dry matter content * ? ? ? * * *
3. Twig drying time * ? ? ? *
4. Bark thickness * * * ?
5. Specific root length * ? * * * ?
6. Diameter of fine root * ? *
7. Distribution of rooting depth * * * * * * *
8. 95% rooting depth * ? * * *
9. Nutrient uptake strategy * * * * * *
Regeneration traits
1. Dispersal mode *
2. Dispersule shape and size *
3. Seed mass * * * *
4. Resprouting capacity * * * *

11
2.4.1.2 Wood traits
Over the past years, there has been a growing interest in anatomical and chemical wood traits in
plant ecological studies. This is because these traits have proven to be critical for studies related
to the estimation of forest aboveground biomass and carbon stock (Thomas and Malczewski,
2007).
Chave et al. (2009) clearly demonstrated the concept of wood economics spectrum along which
tree species differ one from the other. This concept encompasses a set of coordinated wood
chemical and anatomical traits that are presumed to be the mechanistic basis for inter-and intra-
specific variation in tree functional ecology. One of the key wood traits upon which lies this
concept is the wood density.
Wood density
Wood density, also referred to as wood specific gravity, is the ratio of wood dry mass to fresh
volume. It describes the fraction of stem that is tissue and cell walls and the space within cell
walls. Variation in wood density has been shown to be strongly linked to variation in other plant
traits such as the relative mechanical strength of a plant, the hydraulic capacity of the stem, the
timing of reproduction, the mortality rate, the diameter growth rate (Swenson and Enquist,
2007).
Based on the wood economics spectrum of Chave et al. (2009), it is believed that short-lived,
pioneer species will generally have low wood density as a way to achieve fast growth with
minimal structural investment. The structural investment referred to here is mainly in terms of
defensive compounds like lignin and secondary compounds. As for the long-lived, slow-growing
shade tolerant species, they will, on the contrary, have high wood density and incur heavy
investment in wood defenses. In so doing, these species are able to tolerate long periods in the
forest understorey. However, this strategy comes at a cost which is obviously a slow radial or
vertical growth.
In the context of tropical forests, traits involved in the wood economics spectrum were identified
as being among the principal determinants of carbon storage (Baker et al., 2004) and wood
decomposition rates (van Geffen et al., 2010). Wood anatomical traits in general and wood
density in particular have been also shown to be strong predictors of species-level growth and
mortality rates in tropical tree species (Poorter et al., 2008; Wright et al., 2010).
2.4.1.3 Leaf traits
Leaf traits are tightly linked to growth and survival of the plant. That is the main reason as to
why they are considered as good predictors of plant performance (Poorter and Bongers, 2006).
They are not only regarded as important for plants in terms of the acquisition and use of
resources and biomass production but also in relation to the ecosystem functioning as a whole
(Weiher et al., 1999; Vendramini et al., 2002). Their variations are quite often a result of the
adoption by plant species of different strategies (Westoby, 1998). On top of that, they offer the

12
advantage of being easy to quantify and convenient to compare among many plant species (Liu
et al., 2008).
The leaf economics spectrum (LES) with respect to forest ecosystems was introduced by Wright
et al. (2004) to demonstrate that species life history strategy could be simply explained by using
leaf functional traits as proxies. Traits involved in leaf economics spectrum tend to co-vary along
a spectrum of shade tolerance. The concept states that light-demanding, fast-growing pioneer
species express leaf traits that capture high rates of carbon in the short run. Slow-growing, shade
tolerant species, on the contrary, display a conservative resource investment strategy efficient for
long term carbon gain. These two opposing strategies are characterized by a set of traits
including photosynthetic rates (Amax), leaf nitrogen and leaf phosphorous content (LNC and
LPC), leaf respiration (Rd) rates, leaf mass per unit area (LMA) or specific leaf area (SLA) and
leaf life span (LL). Light-demanding, fast-growing pioneer species are characterized by high
Amax, LNC, and Rd rates, and low leaf LMA and short LL. The opposite is true for slow-
growing, shade-tolerant species.
Traits related to leaf structure, nutrient content and net photosynthetic rate all play a role in the
determination of the CO2 and water vapor fluxes between the vegetation and the atmosphere.
These traits are also associated to biogeochemical cycles that relate soil, climate and the
atmosphere (Reich et al., 2007). In addition to conditioning plant behavior and production by
their interaction, these traits provide an interesting conceptual connection between processes at
short-term leaf scales and long-term whole plant and stand-level scales (Meir et al., 2002).
Specific Leaf Area (SLA) and Leaf Mass per Area (LMA)
SLA characterizes the light-intercepting area of a leaf per unit dry mass (m2/g) in relation to the
net assimilation rate (Reich et al., 1992). This particular trait, which happens to be very easy to
measure, is a good correlate of both photosynthetic capacity and potential relative growth rate
(Westoby, 1998). It was also shown to be inversely related to the degree of physical defense of a
leaf (Cornellisen et al., 2003).
LMA is often used to predict leaf area expansion from leaf dry weight increase. It is actually a
measure of the investment of dry matter per unit of light-intercepting leaf area deployed (g/m2).
It can be computed as 1/SLA. Practically, a high LMA signifies a thicker leaf blade and/or a
denser tissue (Cornellisen et al., 2003).
Leaf carbon concentration (LCC)
LCC is the total quantity of carbon per unit of dry leaf mass (mg/g). The carbon content is
generally about 45-50 % of dry matter (Carvalho et al., 1998). It has been suggested that high
SLA was correlated to low LCC, and plants living in low-light environments have generally a
lower LCC (Ryser and Eek, 2000). LCC was also found to be leaf age dependent, with the
youngest tissues containing more carbon than the oldest ones (Alcoverro et al., 2000).

13
C has two stable isotopes, namely 13
C and 12
C. The relative abundance of these two isotopes of C
in plant leaves is also sometimes analyzed and reported as δ13
C. The isotope compostion δ13
C
has been shown to be negatively correlated to water used efficiency (WUE) in many C3 plant
species (Richards, 2005). WUE is defined as the quantity of the biomass produced per unit of
water used (Richards, 2005). The negative correlation between δ13
C and WUE is explained by
the fact that the δ13
C-value is dependent on the carbon isotope discrimination (∆) during carbon
fixation, and both the values of ∆ and WUE are closely linked to the ratio between the
concentration of CO2 in the leaf intercellular space and the concentration of CO2 in the ambient
air (Ci:Ca) (Ardnt and Wanek, 2002). In conditions where water is a limiting factor, it has been
shown that ∆ decreases as WUE increases. This leads to more positive values of δ13
C under
drought stress (Ardnt and Wanek, 2002).
Leaf Nitrogen Content (LNC) and Leaf Phosphorus Content (LPC)
LNC is expressed as the total amount of nitrogen per unit of dry leaf mass in mg/g. LNC is
integral to the proteins of photosynthetic machinery, in particular Rubisco, which constitutes the
basis for the drawdown of carbon dioxide within the leaf. Therefore, it is a reflection of the
concentration of proteins involved in photosynthesis, and it is linked to net photosynthesis rate
(Wright et al., 2004). Together with SLA, LNC provides the necessary information that is critical
for plant growth and development such as relative growth rate and leaf gas exchange (Garnier et
al., 1997).
At the whole plant level, LNC is said to effectively participate in the trade-off between fast
biomass production and effective nutrient use (Grime et al., 1997). At the ecosystem level, LNC
alongside with LSA are believed to significantly impact on primary productivity and nutrient
cycling (Aerts and Chapin, 2000). Finally, LNC is also reported to be useful as simple predictive
tool for litter decomposability that does not require prior knowledge of individual species
taxonomy and biology (Fortunel et al., 2009).
As it is the case of C, Nitrogen (N) also has two stable isotopes named 15
N and 14
N. The relative
abundance of these two isotopes (or isotope composition δ15
N ) in plant tissues is often used to
evaluate ecosystem N acquisition and cycling (Pardo et al., 2013). However, unlike C, the
influence of environmental variables on N isotope discrimination (∆15
N) is not fully elucidated.
Consequently, the interpretation of δ15
N in plant tissues is also less straightforward. δ15
N is said
to be dependent on several factors including soil N availability, land use history, climate,
mycorrhizal symbiosis but also species composition, especially N fixating species. (Pardo et al.,
2006). Despite this fact, it is however known that tropical forest ecosystems with high nitrogen
losses, thus an open N-cycle, exhibit a high δ15
N values both in plants and soil (Peri et al., 2012).
LPC represents the total amount of phosphorus per unit of dry leaf mass in mg/g (Cornelissen et
al., 2003). Leaf phosphorus is encountered in nucleic acids, lipid membranes and bio-energetic
molecules like ATP, and phosphorus mainly originates from soil mineral weathering (Wright et

14
al., 2004). LPC is said to be linked to photosynthetic capacity, and as such it also relates to
growth (Reich and Oleksyn, 2004).
To have an indication on which of the nitrogen or phosphorus is a more limiting factor for
carbon cycling processes in the ecosystem, LNC:LPC ratio is often used (Cornelissen et al.,
2003). LNC: LPC > 20 generally indicates LPC limitation on a vegetation level, while LNC:LPC
< 10 is indicative of LNC limitation. The average LNC:LPC ratio for most terrestrial plants is
estimated at 12-13 (Güsewel, 2004).
Sometimes nitrogen and phosphorus limitation are assessed by computing LCC:LNC and
LCC:LPC ratios. These two ratios can also be used as an indicative guide to estimate the
likelihood that there will be a net release of nitrogen or phosphorus during early stage of the
decomposition of plant leaves. Leaf residues of plants with LCC:LNC ratio < 20:1 and LCC:
LPC ratio < 200:1 favor a fast decomposition rate with net mineralization of nitrogen and
phosphorus happening right from the start (Giller, 2001; Cattanio et al., 2008). This is important
for the production of high quality litter and for nutrient cycling. McGroddy et al. (2004)
estimated the mean LCC:LNC and mean LCC:LPC ratios at 35.5 and 2457 across several
tropical forests.
Leaf Life span (LL)
Expressed in months, LL can be described as the mean duration of the revenue stream from each
leaf constructed. In other words, it is the period of time during which all or part of an individual
leaf is alive and physiologically active (Cornelissen et al., 2003). It is generally expressed in
months.
Because long LL necessitates vigorous construction, it normally corresponds to a high LMA.
Long LL has been often regarded as a form of adaptation to low nutrient availability. That is
because long LL allows for a much longer nutrient utilization period in the leaf biomass
(Mediavilla and Escudero, 2003).
Photosynthetic capacity (Amax)
The photosynthetic assimilation rate measured under high light, ample soil moisture and ambient
carbon dioxide represents the Amax (Field and Mooney, 1986). In other words, Amax is the
measure of the maximum rate at which leaves are capable of fixing C during photosynthesis. It is
often expressed in nmol/g/s. Amax gives a good indication on both chemical and physical
limitations of photosynthesis as set out by biotic and abiotic conditions over various time-scales
(Field and Mooney, 1986). Amax can be, for many plant species, predicted quite precisely by a
combination of SLA and LNC which also correlate with LL (Reich et al., 1999).

15
Rate of dark respiration (Rd)
Rd is the measure of the rate at which plants can release CO2 in the absence of light (nmol/g/s). It
is the reflection of metabolic cost of photosynthate, and more specifically protein turnover and
phloem-loading of photosynthates. In shaded environments, low dark respiration rate during
growth is often seen as a way of reducing carbon losses and maintaining a positive whole-plant
carbon balance (Reich et al., 2003).
Mass-based leaf traits versus area-based leaf traits
To assess the relationships among different leaf traits, these can be normalized either by mass or
by area. In the context of the LES, leaf traits are mostly expressed in terms mass than area. When
leaf traits are expressed in terms of mass, they show very tight relationships among them. This
presents the advantage of constraining the biodiversity of leaves to a single axis (Osnas et al.,
2013). It was however shown that the correlations among leaf traits involved in the LES,
principally Amax, Rd, LNC, and LCC were much weaker when these were expressed in terms of
area (Osnas et al., 2013). This raises the question of how best to interpret the contrasting strong
mass-based relationships and the weaker area-based relationships observed among traits involved
in the LES.
Osnas et al. (2013) argued that most traits involved LES are area-proportional traits that are
expressed in terms of mass. Mass normalization leads to strong correlations between area-
proportional traits owing to the fact that there is large variation between species in LMA. Osnas
et al. (2013) then found in their study that a LES that was independent of mass-or area-
normalization and LMA resulted in physiological relationships that were not in line with those
described in global vegetation model designed for climate change. In a similar way, Llyod et al.
(2013) suggested that the LES should be revisited because photosynthesis was in its essence an
area-based trait. Consequently, any photosynthesis-nutrient relationships or photosynthesis-
structure relationships should be strictly based on a leaf area analysis. For their part, Westboy et
al. (2013) insisted on the fact that mass-based expressions were best suited for studies related to
plant growth and economics of resource use.
2.4.2 Dimensionality of plant functional traits
As already stated, trait-based approaches to ecology are attractive because they require the
analysis of fewer traits compared to the number of species to understand the functioning and
predict the dynamics of plant communities. Despite this fact, it still remains true that many traits
are needed to fully understand the response of species or communities to biotic and abiotic
factors present in their environment.
It stands to reason that the number and type of traits to study will depend on the objectives of the
study at stake. The smallest number of parameters necessary to describe a multi-trait dataset is
referred to as intrinsic dimension (Lee and Verleyson, 2007). To put it in another way, the

16
intrinsic dimensionality of plant traits corresponds to the number of independent axes of
functional variation among plant species.
Plant trait dimensionality in plant ecology has been approached in various ways. Generally,
functional groups are recognized for plants with similar ecology (Grime and Pierce, 2012), and a
few important functional traits are identified that can be used as proxies to explain species life
history. Another approach that has been suggested is the leaf-height-seed strategy (Westoby,
1998). The latter has the advantage of capturing several dimensions by defining the main axes.
Still another approach that certain plant ecologists advocate for consists in the measurement of as
many traits as possible (Cornelissen et al., 2003).
In a recent study, Laughlin (2014) considers that to be defined as optimal, an approach to
dimensionality should take into consideration every organ of the plant (Figure 2.1). He argues
that each plant organ could produce potentially unique information regarding the functioning of a
given plant species within its environment on one hand, and how different plants are distributed
along environmental gradients on the other hand.
Figure 2.1 Seven plant organs or whole-plant properties and their functional significance. Known
statistical relationships among each circle are illustrated by black arrows, and weaker
relationships are shown as grey dashed arrows. The strength of all these relationships among a
set of plants determines the intrinsic dimensionality of plant traits (Reproduced from Laughlin,
2014).

17
Analyzing different plant species-trait datasets for dimensionality reduction by a combination of
linear and non-linear methods, Laughlin (2014) came to the conclusion that the number of
dimensions required for capturing most of the variation in community structure does not exceed
6 for the most comprehensive dataset. He demonstrated also that although additional traits could
increase substantially the ability to predict the community composition, only 4 to 8 traits were
necessary to reach a plateau (Figure 2.2). To broaden the understanding of trait-based
community assembly, Laughlin (2014) recommends firstly that ecologists minimize the number
of traits measured while maximizing the number of dimensions. Secondly, whenever possible,
traits from multiple organs should be measured. These include particularly leaf, stem, root and
flowering traits as they have been shown to be consistent in explaining community assembly
across different ecosystems.
Figure 2.2 Relationship between the number of traits and the ability to predict and explain
variation in community composition (based on the R2 of the relationship between observed and
predicted relative abundances) using a trait-based model of community assembly in six published
studies. Vertical dotted lines indicate where predictive power begins to plateau (Reproduced
from Laughlin, 2014).

18
2.4.3 Intra- specific variability of plant functional traits
To be able to make predictions about community structure, describe plant species distribution
and understand various ecosystem processes such as nutrient cycling and plant productivity,
ecologists often resort to the study of variation in functional traits.
Variation in plant functional traits is caused by evolutionary (genetic) and environmental drivers
that act on phenotypes, and these usually operate at different scales (Reich et al., 2003).
According to Jung al. (2010), species or individuals of the species can establish themselves
under given environmental conditions if they present functional traits with values lying within a
specified range. This is in line with the filtering concept which argues that the environment
exerts significant influence over a given set of species by restricting the types and values of traits
that local communities will be composed of (Weiher and Keddy, 1995). This environmental
action on species leads to a convergence of traits. Competition on the contrary has the opposite
effect in the sense that it brings to trait divergence. This is expressed by a higher spread of trait
values at the community level (Cornwell and Ackerly, 2009). Both these processes may play a
role in shaping local community.
For years, much of the research involving plant functional traits had concentrated in capturing
variation only between species using the mean trait approach. It was assumed that the use of
robust traits will result in much lower intra-specific trait variation as compared to inter-specific
variation. Since intra-specific trait variation is negligible, it could be ignored. Albert et al. (2011)
mention 3 major reasons that promoted this shared understanding including 1) the search for
general patterns at the inter-specific level, 2) the establishment of standardized protocols aiming
at reducing intra-specific variation and 3) and the neglect of intra-specific variation in most trait
databases.
The use of inter-specific variation in plant functional traits has so far allowed the achievement of
important milestones as far as elucidating fundamental patterns and trade-offs in plant design and
functioning, understanding the effects of (changing) species composition on ecosystem functions
and classifying plant species into ‘functional types’ or strategies (Semenova and van der Maarel,
2000).
Nowadays, plant functional ecologists are in unanimous agreement that there can be substantial
variation within species and that intra-specific variability could be relatively important with
respect to inter-specific variation for some traits in some conditions (Table 2.2). In these
circumstances, intra-specific plant trait variation should not be ignored because it could play a
significant role in community processes and assembly mechanisms (Albert et al., 2011).

19
Table 2.2 Summary of nine studies that have explicitly measured inter-specific and intra-specific variation in functional traits
(modified from Auger and Shipley, 2013)
Number of species Environmental
gradient
Traits % Inter-specific
variation
% Intra-specific
variation
References
10 Herbaceous and tree
species North-South transect,
Southern France
Leaf mass, LDMC, Leaf
thickness, SLA
90-65 10-35 Roche et al., 2004
51 Herbaceous species Water depth in flood plain
(inundated to dry), single site,
France
SLA, LDMC, H 87-74 13-26 Jung et al., 2010
32 Tree species Water and soil nutrient
availability, 4 sites, Australia
Wood density, modulus of
elasticity modulus of rupture,
82-57 18-43 Onoda et al., 2010
13 Herbaceous and shrubs species
Altitudinal gradient, Alps, France
H, SLA, LMDC, LCC, LNC 80-60 20-40 Albert et al., 2010
10 Tree species Environmental variation not
specified, Single site dry
forest, Costa Rica
Leaf mass, Leaf area, SLA,
Leaf water content (LWC)
64-17 36-83 Hulshof and Swenson, 2010
119 Tree species Precipitation gradient, East-
West Panama
LMA, LDMC, 35-21* 48 Messier et al., 2010
39 Herbaceous species Soil fertility, mowing and
altitude, meadows in Czech
Republic and grass land in the
French Alps
LDMC, H 72-52 28-48 de Bello et al., 2011
422 Saplings Short environmental gradient
(Slope and amount of solar
radiation), deciduous forest,
Canada
15 functional traits 92-40 8-60 Auger and Shipley, 2013
96 woody species Gradient of anthropogenic
disturbance, 5 sites,
Subtropical forest, China
10 functional traits 74-2** 17-60 Kang et al., 2014
*Strictly inter-specific variation. Variation due to difference in plot : 0 %. Variation due to difference in site was estimated at 17-31%
** Strictly inter-specific variation. Variation due to difference in plot: 0-71%; variation due to difference in site: 2-10%

20
2.4.3.1 Sources of intra-specific plant trait variability
Intra-specific trait variability, which is sometimes referred to as to as intra-specific functional
variability or within species trait variability, can be defined as the total variability of trait values
and trait syndromes (sets of trait values including trait trade-offs) expressed by individuals
within a species (Albert et al., 2011).
Intra-specific variability actually reflects the aptitude of a given species to react to environmental
changes. This occurs via two main mechanisms, adaptation and acclimation and their interaction.
Adaptation is simply the phenotypic variability that occurs among individual genotypes, and is
the consequence of evolutionary processes such as genetic drift, mutation, selection, migration
and the raw material for species future evolution (Albert et al., 2011). As for acclimation, also
termed as phenotypic plasticity, it is the ability to generate several phenotypes by a single
genotype under different environmental conditions (Miner et al., 2005). In other words, it is the
trait variability that arises from environmental heterogeneity in space, time or during an
individual’s lifetime. Acclimation confers to a plant the capacity to alter its morphology and/or
physiology in order to cope with varying environmental conditions. It is therefore considered as
highly beneficial for the performance of the plant (Badyaev, 2009).
2.4.3.2 Structure of intra-specific plant trait variability
Intra-specific trait variability can take place at different spatio-temporal scales and various
ecological levels. The main components into which it can be decomposed are respectively: 1)
population level variability which expresses the differences in trait values between populations
of a single species, 2) between-individual variability, defined as the trait variability within a
certain population and finally 3) within-individual variability, known as the capacity of trait
values to vary within individuals (Albert et al., 2011).
Normally, it is expected that trait values will vary between and within species as well as between
and within plots. That is why signals for community assembly have been often tested by
accounting for both between species variation using species-fixed means and the combination of
between species and within species variation. Within species variation is achieved through plot-
specific means or trait values per individual (Jung et al., 2010; Siefert, 2012). It is known that the
variation observed in community-level trait patterns (mean and spread) is exclusively due to
species turnover when using species-fixed means. However, if the study is based on plot-specific
means, variation in these trait patterns can originate from both species turnover and within
species variation. In this situation, the “intra-specific variability effect” can be derived simply by
separating the role of within species variation on the variation of community-level trait patterns
(Albert et al., 2011).

21
2.4.3.3 Magnitude of intra-specific plant trait variability
Traits differ greatly in their plasticity in plants. For instance, SLA seems to be more plastic than
leaf dry matter content (LDMC). Leaf pH is known to be less plastic whereas traits linked to
resource uptake are often largely variable (Albert et al., 2011). Substantial differences in
plasticity have been also reported for reproductive traits when they were considered to have very
little plasticity. Seed nitrogen content for example presents larger variability than seed mass
which is less plastic (Albert et al., 2011).
Plant traits appear to be unevenly variable. The knowledge of trait plasticity does not necessarily
give a final indication on whether intra-specific trait variation will be relatively important such
that it can significantly influence community level patterns. In addition to intra-specific
variability’s magnitude, other parameters such as intra-specific’s structure might as well affect
these patterns. It is therefore important to know under what conditions it is worthy quantifying
the intra-specific trait variability. Albert et al., (2011) proposed a set of rules that could be
helpful in that respect. They first came up with a hypothesis termed as “spatial variance
partitioning” (SVP) (Figure 2.3) and explained its core tenets. The hypothesis states that the
relative importance of intra-specific trait variation and inter-specific trait variation changes in a
predictable way with the studied organizational and spatial scale. They then concluded that at the
largest organizational or spatial scales, inter-specific trait variability is relatively larger than
intra-specific trait variability. As the scale of the study decreases, intra-specific trait variability
expands. At intermediate spatial scales, intra-specific trait variability can be both greater (Figure
2.3, scenario 2 or 3) or smaller (scenario 1) than inter-specific variability due to the fact that both
intra-specific trait variability and inter-specific variability have quite large magnitudes.

22
Figure 2.3 Hypothetical changes in the magnitude of inter-specific (INTER) and intra-specific
(INTRA) trait variability over geographical scales (and a gradient of increasing environmental
heterogeneity): the spatial variance partitioning (SVP) assumption. INTRA is saturating when scale
is widening, following an asymptotic function: for each study species, a broad scale means that its whole
range is included and thus its whole potential INTRA. INTER keeps on increasing, until the whole
biosphere is included: studies at broader scales include contrasting biomes and the studied species are
typically functionally more different. At broader scales, INTER thus becomes relatively larger than
INTRA (Reproduced from Albert et al., 2011).

23
CHAPTER THREE:
MATERIALS AND METHODS
3.1 Study location
These investigations were conducted in the Arboretum of the agronomic research center
(INERA) of Yangambi, 100 Km West of Kisangani, Orientale province, Democratic Republic of
Congo (0° 38' et 1° 10' N, 24° 16' et 25° 08' E; 470 m of altitude) (Figure 3.1). The center was
established in 1937 and covers an area estimated at 6297 km2 of which 737 ha were devoted to
tree plantations. The study location is characterized as humid tropical forest with an equatorial
climate. The temperature is relatively constant, ranging from 22.4 to 29.3 °C, with an annual
average of about 25°C (Figure 3.2). Annual precipitation varies between 1500 mm and 2000
mm, with an average of 1750 mm. According to Köppen’s classification, the area belongs to the
Af climatic type (Peel et al., 2007). The humid and tropical climatic conditions of the region
favored a strong weathering of the primary minerals leading to the formation of ferallitic soils
(Ngongo et al., 2009).
Figure 3.1 Study site localization (source: Boyemba, 2011)

24
Figure 3.2 Monthly average (from 2000-2008) for precipitation and temperature in the Yangambi
region. The shadowed part corresponds to the months during which precipitation is higher than
the annual average (Source: Boyemba, 2011).
3.2 Experimental set-up
The Yangambi arboretum covers an area currently estimated at about 50 ha. This tree plantation
consists of different experimental plots, planted around 1940. The lay-out consists of
monospecific plots and plots with a mixture of up to six species. However, experimental plots
planted with tree species alone or in combination of two dominated the arboretum. The size of
the plots were either 60 by 60 m (0.36 ha) in the western part of the arboretum or 40 by 40 m
(0.16 ha) within the eastern section of the arboretum.
Since investigating the functional trait variability among co-existing tree species was the
principal stake of this study, only tree species which were present in this arboretum in at least
one monospecific and one two-species mixture were selected. This resulted in a total of 12 target
tree species. Plots were subsequently pooled based on these selected target species in 12 groups,
comprising each time one target species planted alone (monocultures) and a mixture of the target
species with another species (mixed-cultures) as shown in Table 3.1.

25
Table 3.1 Experimental design of the Yangambi arboretum
It should be specified that each of these different configurations is unique as no repetition could
be recognized in the arboretum in its current state. In some cases, an admixed species is also
found as a monoculture in the arboretum and is thus also considered as a target species. The
overall set of planted trees, including both target and admixed species, was made of 23 tree
Abbreviation Scientific name
Plot
ID
Plot size
(ha) Target
species
Admixed
species
A.c. - Autranella congolensis (De Wild.) A. Chev 6 0.36
D.l. Drypetes likwa J. Leonard 14 0.16
E.c. - Entandrophragma cylindricum (Sprague) Sprague 16 0.16
A.n. Antrocaryon nannanii De Wild. 9 0.36
E.a. Entandrophragma angolense (Welw. ex C. DC.) C. DC. 25 0.36
G.c. - Guarea cedrata (A. Chev.) Pellegr. 12 0.16
L.t. Lovoa trichilioides Harms 23 0.36
. P.e. Pericopsis elata (Harms) Meeuwen 3 0.36
L.t. - Lovoa trichilioides Harms 21 0.36
K.a. Khaya anthotheca (Welw.) C.DC. 22 0.36
M.a. - Mammea africana Sabine 28 0.16
S.g. Strombosia grandifolia Hook. f. 29 0.36
M.e. - Milicia excelsa (Welw.) C.C. Berg 11 0.36
P.sp. Phyllanthus species 18 0.36
P.t. - Pachyelasma tessmannii (Harms) Harms 24 0.36
C.a. Chrysophyllum africanum A. DC. 27 0.16
P.o. - Panda oleosa Pierre 15 0.16
P.e. Pericopsis elata (Harms) Meeuwen 5 0.16
P.m. - Pentaclethra macrophylla Benth. 17 0.16
C.p. Carapa procera DC. 10 0.36
Z.g. Zanthoxylum gilletii (De Wild.) P.G. Waterman 4 0.36
P.e. - Pericopsis elata (Harms) Meeuwen 2 0.36
- Pericopsis elata (Harms) Meeuwen 13 0.16
G.c. Guarea cedrata (A. Chev.) Pellegr. 3 0.36
B.w. Blighia welwitschii (Hiern) Radlk. 1 0.36
P.o. Panda oleosa Pierre 5 0.16
S.t. Strombosiopsis tetrandra Engl. 20 0.16
P.s. - Pterocarpus soyauxii Taub. 8 0.36
T.a. Treculia africana Decne. 7 0.36
S.t. - Strombosiopsis tetrandra Engl. 19 0.16
P.e. Pericopsis elata (Harms) Meeuwen 20 0.16

26
species as listed in Table 3.1. In total, 28 plots were inventoried encompassing 13 monocultures
and 15 two-species mixtures.
To prevent the occurrence of spontaneous in-growth, these plots were regularly examined and
tended. This management went on for about 20 years following plantation for plots planted with
the tree species P. elata and 10 years for plots planted with all the other species. After these
periods, the setup was no longer managed. As a result, spontaneously in-growing species are also
now found alongside planted species in the arboretum.
3.3 Leaf sampling
For the leaf sampling, plots were divided in either 4 or 9 subplots of 20 m x 20 m depending on
the initial plot-size, resulting in a total of 201 subplots for the whole arboretum. Five individuals
for each target species present in monoculture plots and in two-species mixture plots were
selected within the arboretum. The individuals were randomly distributed over the plots. From
each individual of the species, and depending on the leaf size, 10 to 20 fully-expanded, non-
senescent and non-juvenile leaves from two different places in the sub-canopy were collected
and placed in A4 size brown envelopes. Upon return from the field, samples were oven dried for
48 hours at 80 ºC. The samples were then shipped to the Isotope Bioscience laboratory
(ISOFYS) of Ghent University, where all the chemical analyses were performed.
3.4 Functional traits and measurement
The first trait studied was the DBH which is commonly used as a measure of tree growth. The
diameter of live stems of the selected individuals of the target species was measured at 1.3 m
above the ground using a measuring tape and following the RAINFOR protocol (Marthews et al.,
2012).
The second functional trait examined in this study was the H. It represents the distance between
the uppermost part of the photosynthetic tissue and the ground. The height of the selected
individuals of the target species was measured using a hypsometer (Vertex III, Haglöf, Sweden).
To reflect the nutrient status of trees, three more traits were measured: LPC, LNC and the stable
nitrogen isotope δ15
N. LPC and LNC both relate to photosynthetic capacity whereas δ15
N
provides information about plant N acquisition and N ecosystem cycling. In addition to the three
mentioned traits, LCC and leaf carbon isotope discrimination (δ13
C) were also analysed. LCC
characterizes the carbon economy of tree leaves whereas δ13
C gives an indication on the WUE.
After the LPC, LNC and LCC of the sampled individuals of the target tree species were
determined, the LCC:LNC and LCC:LPC ratios were computed.
The foliar P determination was performed following the Chapman and Pratt (1961) procedure as
modified by Ryan (2001). 0.5 to 1.0 g of the powdered leaf materials was weighed into 50 mL
Pyrex glass beakers. The beakers were placed in a cool muffle furnace, and the temperature of
the furnace was gradually raised to 550 ºC. The leaf materials were hashed at this temperature
(550 ºC) for 5 hours. After ashing the samples, the beakers were cooled and the ashed samples
were dissolved in 5 mL 2 N hydrochloric acid (HCl) and thoroughly mixed. The mixture was

27
allowed to equilibrate for 20 minutes after which deionized (DI) water was added to the mixture
to make up the volume to 25 mL. The solution was thoroughly mixed again and allowed to stand
for an additional 30 minutes. After that, the solution was filtered through a phosphorus-free
Whatman filter paper (No.42) and the filtrate collected into a new beaker.
10 ml of the digest filtrate were subsequently pipetted into a 100 mL volumetric flask to which
10 mL of ammonium vanadomolybdate reagent were added and the solution was diluted to 100
mL with DI water. Standard solutions were prepared in the same way: 1, 2, 3, 4, and 5 mL of the
standard stock solution in 100 mL volumetric flasks to which 10 ml of the reagent were added
and the solutions diluted to 100 mL with DI water. A blank was also prepared which consisted of
10 ml of the reagent diluted to 100 mL with DI water.
The absorbance of the blank, standards and samples were read by means of a photospectrometer
at 410 nm wavelength. A calibration curve for standard was prepared by plotting absorbance
against the respective P concentrations. The P concentrations of the unknown samples were read
from the calibration curve and expressed in percentage (%) total phosphorus in the leaf samples.
This was later converted to mg P g-1
dry mass of leaf.
The total foliar N and C contents as well as their stable isotope compositions were analyzed
using an elemental analyzer coupled to a continuous flow isotope ratio mass spectrometer (EA-
IRMS). Duplicate aliquots of dried and pulverized leaf material in the range of 1.3-1.7 mg were
weighed from each sample to ensure highest result accuracy and placed in round tin cups. The
packed tin cups containing the leaf samples were then introduced into the EA’s auto sampler unit
for the analysis. Expressed in percentage after chemical analysis, the contents of leaf nitrogen
and carbon were converted to units of dry leaf mass Nmass and Cmass (mg g-1
). The delta notation
relative to Vienna Pee Dee Belemnite standard for δ13
C and atmospheric air for δ15
N served to
express isotope ratios. The reference material for the stable isotope analysis consisted of a flour
laboratory standard with an isotopic composition of -27.01 ± 0.04 ‰ δ13
C and 2.69 ± 0.15 ‰
δ15
N.
3.5 Statistical analysis
3.5.1 Variance partitioning
The first objective of this study was to determine how variability in the nine traits studied was
structured between and within species. In order to achieve this, the trait variability was
decomposed in different levels of variation using both single trait analyses and multi-trait
analyses.
For the single trait analyses, a nested analysis of variance was used to estimate how the variance
in the studied traits was partitioned as described by Hulshof and Swenson (2010). The choice of
this analytical method was motivated by the hierarchical structure of the collected data resulting
from the experimental set up. Three nested levels, namely among species (between species),
among plots (between population) and among individual trees (within population) were included

28
in the analysis for all the studied traits. The factors species had 12 levels; the factor population
(plot) had 31 levels.
For the multi trait analyses, between-group principal component analysis (BPCA) on the set of
nine traits measured at the individual level was used. The method is described in Albert et al.
(2010). When using BPCA, the correlation matrix is built based on the group’s means weighted
by their sample size (Albert et al., 2010). By proceeding in this manner, it becomes possible to
minimize the linear combinations of variables that capture the overall variance and maximize
those that account for the between group variance. The between group variance in this case is
none other than the computed between inertia. This corresponds to the fraction of the overall
variance attributed to the difference between groups (Albert et al., 2010). It should be noted that
the whole procedure is built-in the statistical package that was used (ADE4). Below, the main
steps that were followed to decompose the variance using the multi-trait analyses are outlined
and the whole procedure is sketched in the Figure 3.3.
1) Given the small number of individuals sampled per species and as the dataset contained a few
missing data, a multiple imputation method (Fichman and Cummings, 2003) was performed to
estimate the missing data and balance the data set (equal number of individuals);
2) BPCA was run on the whole dataset. In this case, the groups were made of the different
species. This resulted in the estimation of the between species variance and the within species
variance for the overall data set (the twelve target species and the nine functional traits or
variables included);
3) To estimate the variance between and within population, BPCAs were run for each species
taken individually and with the nine variables (functional traits) considered. Here, the groups
consisted of the different populations for each target species. Since each target species had at
least two different populations (one monoculture population and one two-species mixture
population), it was possible by running a BPCA to decompose the variance in between and
within population for each species.
4) The contribution of each of the nine variables (functional traits) to the total variance was
computed for the BPCA performed with the whole data set averaged by species and all the
BPCAs performed with each species individually (data sets averaged by populations). Following
the same procedure described above, it was possible to decompose the variance for each variable
into between-species variance, between-population variance and within-population variance. It
should however be mentioned that to obtain the overall decomposition into between- and within-
population variances for each variable, the between- and within-population variances obtained
for each species individually were summed.

29
Figure 3.3 Variance partitioning using multi-trait approach. The BPCA on the whole dataset
gave the relative importance of between- and within-species variances (A) while BPCAs on each
species data resulted in the decomposition into between and within populations (B). Populations
of each species are represented by squares (C). Ac: A. congolensis; Ec: E. cylindricum; Gc:G.
cedrata;Lt: L. trichilioides.Ma: M. Africana;Me: M. excelsa; Pt: P. tessmannii; Po: P. oleosa;
Pm: P. macrophylla; Pe: Pericopsis elata; Pt: Pterocarpus soyauxii ; St: S. tetandra.
3.5.2 Main functional trade-offs and strategies
The second objective of this study was to determine whether the main functional trade-offs and
strategies adopted by the target species at the intra-specific level were similar to that at the inter-
specific level. To this end, a within-group principal component analysis (WPCA) on the set of
nine traits measured at individual level was used in addition the BPCA mentioned above.
The correlations between traits at the intra-specific levels obtained using the WPCA analysis was
compared with the correlations between traits at the inter-specific level obtained by running the
BPCA. The correlation between traits at the inter-specific level seeks for axes that discriminate
species in the trait space whereas the correlation between traits at the intra-specific level searches
for axes that discriminate individuals within species in the trait space (Albert et al., 2010).
The WPCA is associated to the BPCA. While the BPCA focuses on the differences between
groups (here between species), WPCA separates the structure within groups (here species) by

30
taking into account the data centered on species means (Dodelec and Chessel, 1991; Albert et al.,
2010). Again, the procedure to obtain the WPCA is built-in the software that was used (ADE4).
The correlation between traits was also examined for each of the species taken individually using
the BPCAs that were performed for each of them. Then, these were compared to check for
similarities.
3.5.3 Trait differences between target species in monocultures and two-species mixtures
The third objective of this study was to examine whether there were significant differences in
plant traits values and/or in multivariate trait distributions between target species in
monocultures and in two-species mixtures.
The differences in trait values were assessed by comparing the mean trait values of target species
from monoculture populations (plots) and those from two-species mixture populations (plots).
Mann Whitney test (U) was used whenever a specific target species was present in one
monoculture plot and one two-mixture plot. In instances where the target species happened to be
present in more than two plots, the Kruskall Wallis test (K-W) was used instead. This was
followed by Dunn’s multiple comparison tests where a statistical difference was detected.
The differences in multivariate trait distributions were assessed by testing the significance of
grouping (here populations) for the BPCAs performed on each species. The between analysis test
built-in the software AD4 was used for this purpose. This analysis tests for the statistical
significance of the dispersion of the centers of gravity of each group. The decision was based on
the number of random values out of 1000 permutations higher than the observed value. The level
of significance was set at 5 %.
It should be mentioned that for this third objective the overall dataset that was balanced by
performing the multiple imputation method (Fichman and Cummings, 2003) was used. However,
in a few cases the original data set contained variables (traits) with only one data point. To avoid
distorting the overall data set, variables with a single data point were maintained during the
multiple imputation method. This resulted in data points that were all equals for some variables
so that there was no variation among them during the analysis and the standard deviation was
equal to zero.
3.5.4 Statistical packages
The nested anova was performed using the GLM procedure with the software Minitab 17
(Minitab Inc, Coventry, UK). The multiple imputation method for the estimation of missing data,
the Mann Whitney test (U) and the Kruskall Wallis test (K-W) were run using the software XL
STAT 2014 (Addinsoft, New York, USA). BPCAs and WPCA were computed using the
software ADE4 version 2001 (CNRS, Toulouse, France).

31
CHAPTER FOUR:
RESULTS
4.1. Variance partitioning
The following trends emerged from the results of the variance decomposition using either the
single- or the multi-trait approach. The intra-specific trait variation was higher than the inter-
specific trait variation for three of the nine traits studies. These are H, DBH and LPC. When the
single trait analysis was used, the variance partitioning resulted in 69.08 % for intra-specific
variation against 30.92 % for inter-specific variation for the trait H. The variance portioning
resulted in 70.40 % for intra-specific variation against 29.60 % for inter-specific variation for the
trait DBH. As for the trait LPC, the intra-specific trait variation accounted for 73.98 % against
26.02 % for the inter-specific variation (Figure 4.1a). The multi-traits analyses led to almost
similar patterns with slight lower intra-specific vs. inter-specific partitions. The intra-specific
variation accounted for 56.87 % of the overall variation for the trait H, 64.97 % of the overall
variation for the trait DBH and 59.44 % of the overall variation for the trait LPC (Figure 4.1b).
For all the other traits examined, the intra-specific trait variation was lower than the inter-
specific trait variation (Fig 4.1a and 4.1b). The traits LCC:LNC and LNC had the greatest inter-
specific variation accounting for 93.40 % and 87.65 % of the total trait variation respectively
when the single approach was used (Fig 4.1a). When the multi-trait approach was used, the
fraction of inter-specific trait variation to the overall trait variation was estimated at 93.81 % for
the trait LCC:LNC and 87.42 % for the trait LNC (Fig 4.1b). For the remaining four traits
examined, the fraction of the intra-specific trait variation to the total trait variation ranged from
from 23.29 % to 49.71 % when the single trait approach was used (Fig 4.1a) and from 26.36 %
to 38.33 % when the multi-trait analysis was used (Fig 4.1b).
Within species, and for all the traits studied, the variation within populations was greater than the
one between populations (Figure 4.1a and 4.1b). When the single trait approach was used, the
traits δ15
N had the greatest between population variation accounting for 18.81 % of total
variation. It was followed by the traits LPC and H with 17.25 % and 14.42 % respectively of the
total variation (Figure 4.1a). The multi-trait analyses also resulted in more less the same pattern
(Figure 4.1b).
The multi-trait variance decomposition taking into account the nine traits and twelve species
together resulted in partition of 63.13 % vs. 36.87 % for inter-specific vs. intra-specific variation
(Figure 4.1c). The contribution of the within population variation to the total within species
variability was about the same as the one of the between population variability for the species L.
trichilioides and M. excelsa. It was lower than the between population variability for the species
E. cylindricum (12.52 % vs. 24.35 %). For the rest of the species studied, the within population
variability was greater than the between population variability (Figure 4.1c).

32
HD
BHLPC
LNC N15
LCC C13
C:N C
:P
0
20
40
60
80
100P
erce
nta
ge
of v
aria
nce
HD
BH
LPC
LN
C N15
LC
C C13
C:N C
:P
0
20
40
60
80
100
Per
cen
tag
e of
var
ian
ce
A. c
ongo
lens
is
E. c
ylin
dric
um
G. c
edra
ta
L. t
rich
ilioi
des
M. a
fric
ana
M. e
xcel
sa
P. t
essm
anni
i P
. ole
osa
P. m
acro
phyl
la
P. e
lata
P
. soy
auxi
i S.
tetr
andr
a
0
20
40
60
80
100
Per
cen
tag
e o
f va
rian
ce
Figure 4.1 Variance decomposition in inter-specific and intra-specific contributions for single-
trait and multi-trait patterns. Variance decomposition in different levels: species, populations and
within populations. (a) Single trait analyses: decompositions, resulting from nested analysis of
variance, are given for each of the nine measured traits. (b) Multi-trait analyses: decomposition
takes into account the contribution of each of the nine variables (traits) to the total variance. (c)
Multi-trait analyses: decomposition takes into account the nine variables (traits) together. The
BPCA on the whole dataset gave the relative importance of between- and within-species
variances while BPCAs on each species data resulted in the decomposition into between and
within populations.
a) b)
c)

33
4.2 Main functional trade-offs and strategies
The loading plot of the BPCA performed on the whole data set revealed that the dataset was
structured by a fairly strong first axis explaining 40.53 % of the total variance, as against 18.03
% for the second axis. The first axis of variation of the loading plot was mainly explained by
LCC:LPC, LCC:LNC, LNC and LPC whereas the second axis was mostly explained by δ13
C,
DBH and H. There was a moderate positive correlation between DBH and H (r = 0.36), between
LNC and LPC (r = 0.47), between LCC:LPC and LCC:LNC (r = 0.65) and a strong negative
correlation between LCC:LNC and LNC (r = -0.92) and LCC:LPC and LPC (r = -0.85). Thus the
first dimension opposed LCC:LPC and LCC:LNC to LPC and LNC (Figure 4.2a).
The WPCA showed that the dataset was characterized by a relatively moderate contribution of
the first and second axes to the total variance (29.01 % and 22.31 %, respectively). In the first
dimensions, most of the variation were explained by LCC, LCC:LNC, LNC and δ15
N. In the
second dimension, the variation was mainly explained by δ13
C, H and DBH (Figure 4.2b).
The BPCAs conducted on each species one by one exhibited a certain inconsistency in the
functional traits across the twelve species studied as shown in Figure 4.3. Depending on the
species, the first axis of ordination accounted for 29.41 % to 68.03 % of the variance. For most
species, the first axis was driven by LCC:LPC, LCC:LNC and LPC. The second axis of
ordination (from 13.61 % to 29.06 % of variance), for its part, was driven for the majority of
species by LCC, δ15
N and DBH.
The nine considered functional traits significantly (p < 0.000) segregated different species within
the trait space when the BPCA on the whole dataset was used (Figure 4.4). Species adopting
different trade-offs in terms of leaf economics, tree size and water use efficiency could be
recognized among species. This can be exemplified by the species M. africana (Ma) that was
characterized by high LCC and low LPC and LNC in the first dimension. It was opposed to E.
cylindricum (Ec). In the second dimension, the species P. elata (Pe) had a high value for δ13
C
and was opposed to S. tetandra (St). P. tessmannii (Pt), for its part, was characterized by a big
size and low δ15
N content, and was opposed to E. cylindricum (Ec). The majority of the
remaining species were located around the center of origin of the plane (Figure 4.4).

34
Figure 4.2 Multidimensional structure within the trait space: Inter-specific and intra-specific
trade-offs. Between and within PCA analysis using the nine measured traits; the correlation
circles and the two-first PCA axes at (a) the inter-specific level (BPCA) and (b) at the intra-
specific level (WPCA) with the whole dataset.

35
Figure 4.3 Multidimensional structure within the trait space: Intra-specific trade-offs. BPCA
analyses using the nine measured traits on the data of each species; the correlation circles and the
two-first PCA axes at intra-specific level within each species.

36
Figure 4.4 Dispersion of species and individuals of each species in the trait space (BPCA on the
whole dataset) as function of the nine trait variables: Variability of individuals among species.
Individuals are identified by squares. Lines link individuals to the corresponding species. Ac: A.
congolensis; Ec: E. cylindricum; Gc:G. cedrata;Lt: L. trichilioides.Ma: M. Africana;Me: M.
excelsa; Pt: P. tessmannii; Po: P. oleosa; Pm: P. macrophylla; Pe: Pericopsis elata; Pt:
Pterocarpus soyauxii ; St: S. tetandra.
Ac
EcGc
Lt
Ma
Me
Pe
Pm
Po
Ps
Pt
St
-3
3.4-4.9 4.3
Axis 2: 18.03%
Axis 1: 40.53% p<0.000
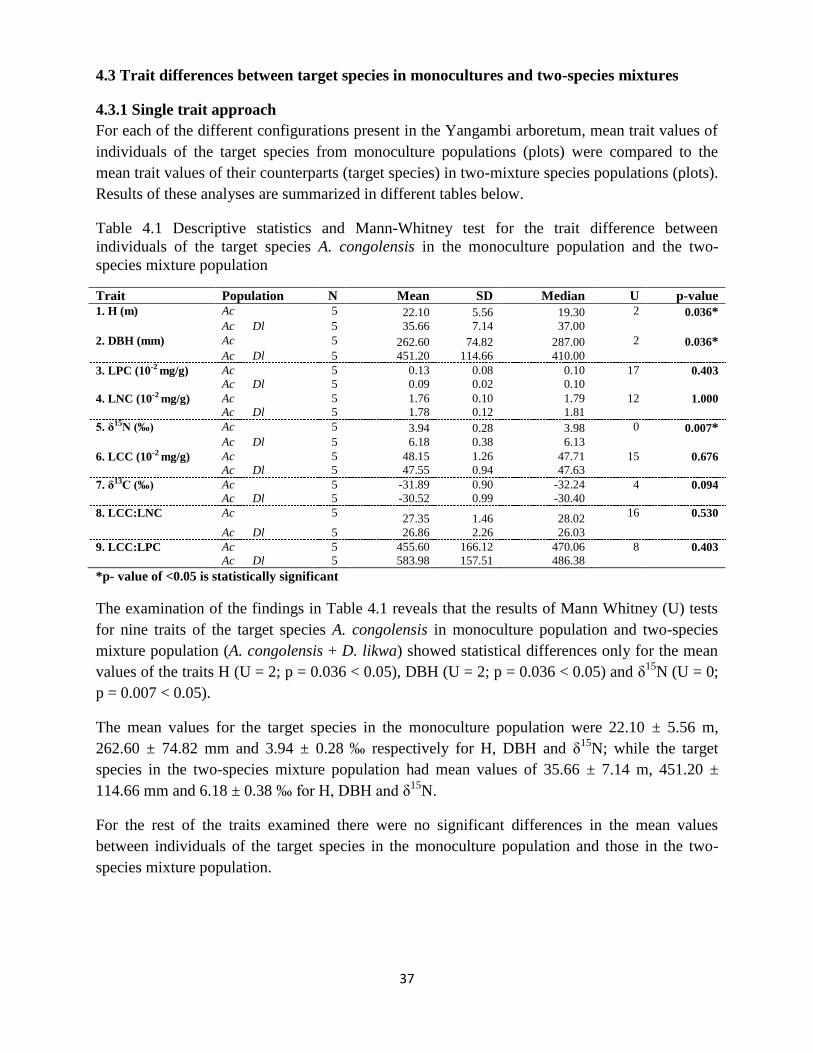
37
4.3 Trait differences between target species in monocultures and two-species mixtures
4.3.1 Single trait approach
For each of the different configurations present in the Yangambi arboretum, mean trait values of
individuals of the target species from monoculture populations (plots) were compared to the
mean trait values of their counterparts (target species) in two-mixture species populations (plots).
Results of these analyses are summarized in different tables below.
Table 4.1 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species A. congolensis in the monoculture population and the two-
species mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Ac 5 22.10 5.56 19.30 2 0.036*
Ac Dl 5 35.66 7.14 37.00
2. DBH (mm) Ac 5 262.60 74.82 287.00 2 0.036*
Ac Dl 5 451.20 114.66 410.00
3. LPC (10-2 mg/g) Ac 5 0.13 0.08 0.10 17 0.403
Ac Dl 5 0.09 0.02 0.10
4. LNC (10-2 mg/g) Ac 5 1.76 0.10 1.79 12 1.000
Ac Dl 5 1.78 0.12 1.81
5. δ15N (‰) Ac 5 3.94 0.28 3.98 0 0.007*
Ac Dl 5 6.18 0.38 6.13
6. LCC (10-2 mg/g) Ac 5 48.15 1.26 47.71 15 0.676
Ac Dl 5 47.55 0.94 47.63
7. δ13C (‰) Ac 5 -31.89 0.90 -32.24 4 0.094
Ac Dl 5 -30.52 0.99 -30.40
8. LCC:LNC Ac 5 27.35 1.46 28.02
16 0.530
Ac Dl 5 26.86 2.26 26.03
9. LCC:LPC Ac 5 455.60 166.12 470.06 8 0.403
Ac Dl 5 583.98 157.51 486.38
*p- value of <0.05 is statistically significant
The examination of the findings in Table 4.1 reveals that the results of Mann Whitney (U) tests
for nine traits of the target species A. congolensis in monoculture population and two-species
mixture population (A. congolensis + D. likwa) showed statistical differences only for the mean
values of the traits H (U = 2; p = 0.036 < 0.05), DBH (U = 2; p = 0.036 < 0.05) and δ15
N (U = 0;
p = 0.007 < 0.05).
The mean values for the target species in the monoculture population were 22.10 ± 5.56 m,
262.60 ± 74.82 mm and 3.94 ± 0.28 ‰ respectively for H, DBH and δ15
N; while the target
species in the two-species mixture population had mean values of 35.66 ± 7.14 m, 451.20 ±
114.66 mm and 6.18 ± 0.38 ‰ for H, DBH and δ15
N.
For the rest of the traits examined there were no significant differences in the mean values
between individuals of the target species in the monoculture population and those in the two-
species mixture population.

38
Table 4.2 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species L. trichilioides in the monoculture population and the two-
species mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Lt 5 17.74 9.99 14.80 7 0.296
Lt Ka 5 22.80 3.23 24.40
2. DBH (mm) Lt 5 168.00 38.35 159.00 7 0.296
Lt Ka 5 209.20 72.88 238.00
3. LPC (10-2 mg/g) Lt 5 0.09 0.01 0.10 7 0.296
Lt Ka 5 0.10 0.01 0.10
4. LNC (10-2 mg/g) Lt 5 2.91 0.13 2.89 25 0.007*
Lt Ka 5 2.47 0.02 2.47
5. δ15N (‰) Lt 5 7.36 0.35 7.43 25 0.007*
Lt Ka 5 4.18 0.63 4.03
6. LCC (10-2 mg/g) Lt 5 45.66 0.40 45.62 2 0.036*
Lt Ka 5 46.31 0.32 46.27
7. δ13C (‰) Lt 5 -31.19 0.92 -31.46 1 0.026*
Lt Ka 5 -29.17 1.01 -29.27
8. LCC:LNC Lt 5 15.69 0.71 15.71 0 0.007*
Lt Ka 5 18.70 0.29 18.68
9. LCC:LPC Lt 5 507.87 93.12 482.25 15 0.676
Lt Ka 5 467.24 24.99 463.16
*p- value of <0.05 is statistically significant
An examination of the findings in Table 4.2 shows that the results of the Mann Whitney U tests
applied to the nine traits examined for individuals of the target species L. trichilioides in the
monoculture population and in the two-species mixture population (L. trichilioides + K.
anthotheca) revealed statistically differences at level of p < 0.05 for the mean values of five
traits. These are LNC (U = 25; p = 0.007 < 0.05), δ15
N (U = 25; p = 0.007 < 0.05), LCC (U = 2;
p = 0.036 < 0.05), δ13
C ((U = 1; p = 0.026 < 0.05), LCC:LNC (U = 0; p = 0.007 < 0.05).
The compared mean values were respectively (2.91 ± 0.13) x 10-2
mg/g, 7.36 ± 0.35 ‰, (45.66 ±
0.40) x 10-2 mg/g, -31.19 ± 0.92 ‰ and 15.69 ± 0.71 for LNC, δ
15N, LCC, δ
13C, LCC:LNC in the
monoculture population, whereas in the two-species mixture population they were (2.47 ± 0.02)
x 10-2 mg/g, 4.18 ± 0.63 ‰, (46.31 ± 0.32) x 10
-2 mg/g, -29.17 ± 1.01 ‰ and 18.70 ± 0.29. These
mean values were higher for individuals of the target species in the monoculture population than
in the two species mixture population for the traits LNC and δ15
N. They were lower for for
individuals of the target species in the monoculture population than in the two species mixture
population for the traits LCC, δ13
C and LCC:LNC.
No significant differences in the mean values between individuals of the target species in the
monoculture population and those in the two-species mixture population were detected for all the
other traits.

39
Table 4.3 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species M. africana in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Ma 5 32.04 8.85 36.60 10 0.676
Ma Sg 5 35.04 6.01 33.20
2. DBH (mm) Ma 5 387.00 167.18 343.00 7 0.296
Ma Sg 5 480.80 112.85 532.00
3. LPC (10-2 mg/g) Ma 5 0.07 0.01 0.07 6 0.210
Ma Sg 5 0.08 0.01 0.08
4. LNC (10-2 mg/g) Ma 5 1.75 0.11 1.72 19 0.210
Ma Sg 5 1.65 0.14 1.64
5. δ15N (‰) Ma 5 4.55 0.79 4.78 17 0.403
Ma Sg 5 4.42 1.49 4.13
6. LCC (10-2 mg/g) Ma 5 50.00 1.10 50.11 15 0.676
Ma Sg 5 49.50 0.22 49.57
7. δ13C (‰) Ma 5 -31.52 0.54 -31.47 5 0.143
Ma Sg 5 -31.05 0.56 -30.94
8. LCC:LNC Ma 5 28.73 1.58 28.97 7 0.296
Ma Sg 5 30.18 2.57 30.24
9. LCC:LPC Ma 5 786.18 143.71 752.58 20 0.153
Ma Sg 5 655.32 96.45 663.02
*p- value of <0.05 is statistically significant
The results of the Mann Whitney U tests in Table 4.3 indicate that there were no significant
differences between individuals of the target species M. africana in the monoculture population
and individuals of the same target species in the two species-mixture population (M. Africana +
S. grandifolia) for the mean values of all the traits examined at level of p < 0.05.
Table 4.4 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species M.excelsa in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Me 5 32.60 8.79 28.40 16 0.531
Me Psp 5 28.46 5.83 30.90
2. DBH (mm) Me 5 278.00 49.12 290.00 5 0.144
Me Psp 5 420.80 150.58 409.00
3. LPC (10-2 mg/g) Me 5 0.13 0.01 0.13 25 0.007*
Me Psp 5 0.11 0.00 0.11
4. LNC (10-2 mg/g) Me 5 2.56 0.13 2.56 25 0.007*
Me Psp 5 2.37 0.00 2.37
5. δ15N (‰) Me 5 6.35 0.48 6.44 25 0.009*
Me Psp 5 4.92 0.00 4.92
6. LCC (10-2 mg/g) Me 5 43.18 0.72 43.21 1 0.022*
Me Psp 5 43.93 0.00 43.93
7. δ13C (‰) Me 5 -29.08 1.42 -29.47 10 0.666
Me Psp 5 -29.43 0.00 -29.43
8. LCC:LNC Me 5 16.86 0.58 16.86 0 0.007*
Me Psp 5 18.53 0.00 18.53
9. LCC:LPC Me 5 353.96 29.99 363.67 0 0.009*
Me Psp 5 409.79 0.00 409.79
*p- value of <0.05 is statistically significant

40
The results of the Mann Whitney U tests in Table 4.4 indicate that there were significant
differences at level of p < 0.05 between individuals of the target species M.excelsa in the
monoculture population and individuals of the same target species in the two species-mixture
population (M.excelsa + Phyllanthus sp.) for the mean values of the following traits: LPC (U =
25; p = 0.007 < 0.05), LCN (U = 25; p = 0.007 < 0.05), δ15
N (U = 25; p = 0.009 < 0.05), LCC (U
= 1; p = 0.022 < 0.05), C:N (U = 0; p = 0.007 < 0.05), LCC:LPC (U = 0; p = 0.007 < 0.05).
The mean trait values for individuals of the target species from the monoculture population were
higher than the mean trait values for individuals of the target species from the two species
mixture population for LPC: (0.13 ± 0.01) x 10-2 mg/g > (0.11 ± 0.00) x10
-2 mg/g; LNC: (2.56 ±
0.13) x 10-2 mg/g > (2.37 ± 0.00) x 10
-2 mg/g and δ15
N: (6.35 ± 0.48 ‰ > 4.92 ± 0.00 ‰). The
opposite was true for LCC: (43.18 ± 0.72) x 10-2
mg/g < (43.93 ± 0.00) x10-2
mg/g, LCC:LNC:
16.86 ± 0.58 < 18.53 ± 0.00) and LCC:LPC (353.96 ± 29.99 < 409.79 ± 0.00).
No significant differences in the mean values between individuals of the target species in the
monoculture population and those in the two-species mixture population were detected for the
remaining traits (H, DBH and δ13
C).
Table 4.5 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. oleosa in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Po 5 20.50 10.51 19.00 7 0.296
Po Pe 5 28.82 3.04 29.88
2. DBH (mm) Po 5 279.00 146.05 286.00 8 0.403
Po Pe 5 344.40 64.69 355.00
3. LPC (10-2 mg/g) Po 5 0.11 0.01 0.11 12 1.000
Po Pe 5 0.13 0.05 0.11
4. LNC (10-2 mg/g) Po 5 3.08 0.21 3.09 5 0.144
Po Pe 5 3.29 0.03 3.29
5. δ15N (‰) Po 5 7.51 0.74 7.61 19 0.210
Po Pe 5 7.08 0.30 7.11
6. LCC (10-2 mg/g) Po 5 47.63 0.85 47.46 10 0.676
Po Pe 5 47.70 0.43 47.66
7. δ13C (‰) Po 5 -32.93 0.92 -32.75 13 1.000
Po Pe 5 -32.84 1.12 -33.08
8. LCC:LNC Po 5 15.50 0.97 15.23 21 0.095
Po Pe 5 14.45 0.24 14.41
9. LCC:LPC Po 5 421.33 47.33 451.95 14 0.834
Po Pe 5 428.19 131.11 437.71
*p- value of <0.05 is statistically significant
The results of the Mann Whitney U tests in Table 4.5 indicate that there were no significant
differences between individuals of the target species P. oleosa in the monoculture population and
individuals of the same target species in the two species-mixture population (P. oleosa + P.
elata) for the mean values of all the traits examined at level of p < 0.05.

41
Table 4.6 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. soyauxii in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H (m) Ps 5 26.63 12.79 23.85 9 0.531
Ps Ta 5 29.80 2.73 29.30
2. DBH (mm) Ps 5 187.80 43.47 163.00 0 0.007*
Ps Ta 5 419.40 51.68 429.00
3. LPC (10-2 mg/g) Ps 5 0.10 0.01 0.10 5 0.143
Ps Ta 5 0.18 0.06 0.18
4. LNC (10-2 mg/g) Ps 5 3.20 0.33 3.22 9 0.531
Ps Ta 5 3.38 0.15 3.39
5. δ15N (‰) Ps 5 2.70 0.78 2.77 15 0.671
Ps Ta 5 2.62 0.83 2.60
6. LCC (10-2 mg/g) Ps 5 47.79 0.69 47.86 24 0.021*
Ps Ta 5 44.89 1.53 44.66
7. δ13C (‰) Ps 5 -31.53 0.49 -31.59 6 0.210
Ps Ta 5 -30.90 0.79 -31.15
8. LCC:LNC Ps 5 15.02 1.73 14.53 20 0.141
Ps Ta 5 13.43 1.05 13.46
9. LCC:LPC Ps 5 467.69 27.54 478.59 19 0.210
Ps Ta 5 374.13 125.51 385.75
*p- value of <0.05 is statistically significant
As revealed by the results of the Mann Whitney U tests in Table 4.6, there were significant
differences at level of p < 0.05 between individuals of the target species P. soyauxii in the
monoculture population and individuals of the same target species in the two species-mixture
population (P. soyauxii + T. africana) only for the mean values of two out of the nine trait
examined, namely DBH and LCC.
Individuals of the target species in the monoculture population had a lower DBH (Mean = 187.80
± 43.47 mm) compared to individuals of the same target species in the two-species mixture
population (Mean = 419.40 ± 51.68 mm), U = 0, p = 0.007 < 0.05. Individuals of the target
species in the monoculture population exhibited a higher LCC (Mean = (47.79 ± 0.69) x 10-2
mg/g) than those present in the two-species mixture population (Mean = (44.89 ± 1.53) x10-2
mg/g), U = 24, p = 0.021 < 0.05.
For the remaining seven traits examined, there were no significant differences in the mean values
between individuals of target species in the monoculture population and those in the two-species
mixture population.

42
Table 4.7 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species P. tessmannii in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H(m) Pt 5 33.18 3.10 32.60 5 0.143
Pt Ca 5 38.36 8.09 38.93
2. DBH(mm) Pt 5 546.00 235.71 577.00 11 0.834
Pt Ca 5 624.20 360.56 546.00
3. LPC (10-2 mg/g) Pt 5 0.06 0.01 0.06 7 0.295
Pt Ca 5 0.09 0.05 0.06
4. LNC (10-2 mg/g) Pt 5 2.34 0.19 2.34 5 0.144
Pt Ca 5 2.64 0.27 2.68
5. δ15N (‰) Pt 5 2.08 0.92 1.98 4 0.0947
Pt Ca 5 3.12 0.56 3.07
6. LCC(10-2 mg/g) Pt 5 50.24 0.46 50.15 25 0.007*
Pt Ca 5 47.34 0.17 47.36
7. δ13C (‰) Pt 5 -30.32 0.89 -30.45 3 0.060
Pt Ca 5 -29.06 0.30 -29.11
8. LCC:LNC Pt 5 21.46 1.65 21.24 24 0.021*
Pt Ca 5 18.22 1.87 18.41
9. LCC:LPC Pt 5 840.01 96.06 844.06 19 0.021*
Pt Ca 5 567.56 160.55 576.82
*p- value of <0.05 is statistically significant
As shown by the results of Mann Whitney U tests set out in Table 4.7, the mean values of
individuals of the target species P. tessmannii in the monoculture population and those in the two
species-mixture population (P. tessmannii + Chrysophyllum africanum) were statistically
different at level of p < 0.05 for the traits LCC, LCC:LNC and LCC:LPC.
Individuals of the target species in the monoculture population had a higher LCC (Mean = (50.24
± 0.46) x 10-2 mg/g) compared to individuals of the same target species in the two-species
mixture population (Mean = (47.34 ± 0.17) x10-2 mg/g), U = 25, p = 0.007< 0.05. Similarly,
individuals of the target species in the monoculture population exhibited higher LCC:LNC and
LCC:LPC ratios (Mean = 21.46 ± 1.65 and Mean = 840.01 ± 96.06 respectively) than those
encountered in the two-species mixture population (Mean =18.22 ± 1.87 and Mean = 567.56 ±
160.55 respectively), U = 24, p = 0.02 < 0.05 for LCC:LNC and U = 19, p = 0.021 < 0.05 for
LCC:LPC.
The mean values of the individuals of the target species in the monoculture population and those
in the two-species mixture population did not show any statistical differences for the six
remaining traits investigated.
The results of the Mann Whitney U tests in Table 4.8 indicate that there were no significant
differences between individuals of the target species S. tetrandra in the monoculture population
and individuals of the same target species in the two species-mixture population (S. tetrandra +
P. elata) for the mean values of all the traits examined at level of p < 0.05.

43
Table 4.8 Descriptive statistics and Mann-Whitney test for the trait difference between
individuals of the target species S. tetrandra in the monoculture population and the two-species
mixture population
Trait Population N Mean SD Median U p-value
1. H(m) St 5 23.50 6.90 27.30 20 0.116
St Pe 5 17.50 3.67 15.40
2. DBH(mm) St 5 282.00 101.00 242.00 16 0.543
St Pe 5 256.00 82.13 214.00
3. LPC (10-2 mg/g) St 5 0.16 0.03 0.16 21 0.095
St Pe 5 0.12 0.03 0.11
4. LNC (10-2 mg/g) St 5 3.48 0.21 3.44 13 1.000
St Pe 5 3.47 0.44 3.58
5. δ 15 N (‰) St 5 6.77 0.97 6.93 14 0.834
St Pe 5 6.48 1.52 6.21
6. LCC(10-2 mg/g) St 5 48.60 0.64 48.30 12 1.000
St Pe 5 48.27 1.45 48.96
7. δ 13 C (‰) St 5 -35.29 0.43 -35.32 13 1.000
St Pe 5 -34.85 1.33 -35.69
8. LCC:LNC St 5 14.02 0.90 14.41 13 1.000
St Pe 5 14.08 1.56 13.71
9. LCC:LPC St 5 304.28 58.66 299.78 4 0.094
St Pe 5 424.07 109.71 448.59
*p- value of <0.05 is statistically significant
Table 4.9 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species E. cylindricum in the monoculture population and the two-
species mixture populations
Trait Population N Mean SD Median Mean
rank
K-W p-value
1. H (m) Ec 5 10.92 2.51 10.70 5.7 2.27 0.322
Ec An 5 18.70 12.78 10.80 8.4
Ec Ea 5 22.08 9.48 19.70 9.9
2. DBH (mm) Ec 5 160.60 71.37 146.00 5.4 2.54 0.281
Ec An 5 210.60 47.25 225.00 9.2
Ec Ea 5 201.40 44.34 220.00 9.4
3. LPC (10-2 mg/g) Ec 5 0.20 0.02 0.20 10.4a 9.38 0.009*
Ec An 5 0.21 0.03 0.20 10.6a
Ec Ea 5 0.12 0.01 0.12 3.0b
4. LNC (10-2 mg/g) Ec 5 3.51 0.00 3.51 3.0a 10.59 0.005*
Ec An 5 4.68 0.45 4.77 9.2ab
Ec Ea 5 5.13 0.31 5.06 11.8b
5. δ15N (‰) Ec 5 6.95 0.00 6.95 4.0a 10.28 0.006*
Ec Ea 5 7.53 0.68 7.59 7.2ab
Ec An 5 9.40 1.14 8.84 12.8b
6. LCC (10-2 mg/g) Ec An 5 43.90 1.01 43.80 4.0a 10.88 0.004*
Ec 5 45.02 0.00 45.02 7.0ab
Ec Ea 5 46.53 0.44 46.42 13b
7. δ13C (‰) Ec Ea 5 -30.19 0.42 -30.13 4.6 5.88 0.052
Ec 5 -29.81 0.00 -29.81 8.0
Ec An 5 -29.16 0.61 -29.10 11.4
8. LCC:LNC Ec 5 12.82 0.00 12.82 13.0a 9.97 0.007*
Ec An 5 9.58 1.00 9.76 6.2b
Ec Ea 5 9.12 0.63 9.18 4.8b
9. LCC:LPC Ec 5 183.28 0.00 183.28 4.0a 10.88 0.004*
Ec An 5 214.05 24.32 210.64 7.0ab
Ec Ea 5 413.44 28.62 407.54 13.0b
*p- value of <0.05 is statistically significant; Values within treatments for each analysis in the column Mean rank followed
by the same letter are not significantly different at p < 0.05 according to Dunn’s multiple range test.

44
The results of Kruskal Wallis tests in Table 4.9 indicate that there were significant differences
among individuals of the target species E. cylindricum present in the monoculture population,
those present in the first two-species mixture population (E. cylindricum + A. nannanii) and
those present in the second two-species mixture population (E. cylindricum + E. angolense) for
the mean values of six of the nine traits considered. These are LPC (K-W = 9.38; DF = 2; p =
0.009 < 0.05), LNC (K-W = 10.59; DF = 2; p = 0.005 < 0.05), δ15
N (K-W = 10.28; DF = 2; p =
0.006 < 0.05), LCC (K-W = 10.88; DF = 2; p = 0.004 < 0.05), LCC:LNC (K-W = 9.97; DF = 2;
p = 0.007 < 0.05) and LCC:LPC (K-W = 10.88; DF = 2; p = 0.004 < 0.05).
The multiple pairwise comparisons using Dunn’s procedure revealed that the LPC of individuals
of the target species in the monoculture population was statistically higher (Mean = (0.20 ± 0.02)
x 10-2 mg/g) than that of the individuals of the target species in the second two-species mixture
population (Mean = (0.12 ± 0.01) x 10-2 mg/g) but was not statistically different from the one of
individuals of the target species in the first two-species mixture population.
The same comparison procedure showed that the LNC of individuals of the target species in the
monoculture population was statistically lower (Mean = (3.51 ± 0.00) x 10-2 mg/g) than that of
individuals of the target species in the second two-species mixture population (Mean = (5.13 ±
0.31) x10-2 mg/g), but was not statistically different from the one of individuals of the target
species in the first two-species mixture population. In the case of the leaf trait δ15
N, a
significantly lower value was detected in leaves of individuals of the target species present in the
monoculture population (Mean= 6.95 ± 0.00 ‰) compared to leaves individuals of the target
species present in the first two-species mixture population (Mean = 9.40 ± 1.14 ‰).
For the LCC, individuals of the target species in the first two-species mixture population had a
statistically lower content (Mean = (43.90 ± 1.01 x 10-2 mg/g) than those in the second two
species-mixture population (Mean = (46.53 ± 0.44) x 10-2 mg/g). However, there were no
significant differences between the mean values of the individuals of the target species in the
monoculture population and those in both the first and second two-species mixture populations
for this trait.
The LCC:LNC ratio was statistically higher for individuals of the target species in the
monoculture population (Mean = 12.82 ± 0.00 ) than for individuals present in both the first two-
species mixture population (Mean = 9.58 ± 1.00) and the second two-species mixture population
(Mean = 9.12 ± 0.63). As for the LCC:LPC ratio, it was significantly lower for individuals of the
target species in the monoculture population (Mean = 183.28 ± 0.00) than for individuals present
in the second two-species mixture population (Mean = 413.44 ± 28.62).
For the remaining traits examined, there were no significant differences in the mean values
between individuals of the target species in the monoculture population and those in the first or
second two-species mixture populations.

45
Table 4.10 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species G.cedrata in the monoculture population and the two-species
mixture populations
Trait Population N Mean SD Median Mean
rank
K-W p-value
1. H (m) Gc 5 27.74 4.61 28.00 5.4 5.18 0.075
Gc Lt 5 34.64 1.41 35.12 11.6
Gc Pe 5 27.76 8.30 28.90 7.0
2. DBH (mm) Gc 5 223.00 71.53 213.00 4.5 4.62 0.099
Gc Lt 5 338.20 107.86 323.00 9.9
Gc Pe 5 307.80 54.43 337.00 9.6
3. LPC (10-2 mg/g) Gc 5 0.15 0.02 0.15 10.4 4.58 0.101
Gc Lt 5 0.12 0.01 0.12 4.6
Gc Pe 5 0.14 0.05 0.13 9.0
4. LNC (10-2 mg/g) Gc 5 3.33 0.30 3.41 7.2 3.23 0.198
Gc Lt 5 3.16 0.00 3.16 6.0
Gc Pe 5 3.64 0.39 3.76 10.8
5. δ15N (‰) Gc Lt 5 3.70 0.00 3.70 3.0a 10.14 0.006*
Gc Pe 5 8.16 0.38 8.18 9.6ab
Gc 5 8.46 1.41 8.99 11.4b
6. LCC (10-2 mg/g) Gc 5 48.14 1.13 48.29 7.4 0.39 0.821
Gc Lt 5 48.66 0.00 48.66 7.6
Gc Pe 5 48.22 0.99 48.04 9.0
7. δ13C (‰) Gc Lt 5 -31.60 0.00 -31.60 3.0a 10.16 0.006*
Gc 5 -31.11 0.59 -31.42 9.4ab
Gc Pe 5 -30.83 0.45 -31.02 11.6b
8. LCC:LNC Gc Pe 5 13.36 1.37 13.15 4.0a 6.62 0.036*
Gc 5 14.53 1.14 14.39 9.0ab
Gc Lt 5 15.39 0.00 15.39 11.0b
9. LCC:LPC Gc 5 333.52 44.97 337.75 5.0a 6.62 0.036*
Gc Pe 5 370.46 125.39 368.48 7.0ab
Gc Lt 5 460.79 0.00 460.79 12.0b
*p- value of <0.05 is statistically significant; Values within treatments for each analysis in the column Mean rank followed
by the same letter are not significantly different at p < 0.05 according to Dunn’s multiple range test.
As shown by the results of Kruskal Wallis tests in Table 4.10, there were significant differences
among individuals of the target species G.cedrata present in the monoculture population, those
present in the first two-species mixture population (G.cedrata + L. trichilioides) and those
present in the second two-species mixture population (G.cedrata + P. elata) for the mean values
of four out the nine traits measured. These are δ15
N (K-W = 10.14, DF = 2, p = 0.006 < 0.05),
δ13
C (K-W = 10.16, DF = 2, p = 0.006 < 0.05), LCC:LNC (K-W = 6.62, DF = 2, p = 0.036 <
0.05) and LCC:LPC (K-W = 6.62, DF = 2, p = 0.036 < 0.05).
The multiple pairwise comparisons using Dunn’s procedure showed that individuals of the target
species in the monoculture population had a significantly higher δ15
N value (Mean = 8.46 ± 1.41
‰) than the one of those in the first two-species mixture population (Mean = 3.70 ± 0.00 ‰).
The LCC:LPC ratio for the individuals of the target species in the monoculture population was
statistically lower (Mean = 333.52 ± 44.97) than the one for the individuals of the target species
in the first two-species mixture population (Mean = 460.79 ± 0.00).
There were no significant differences between the mean values of the individuals of the target
species in the monoculture population and those in both the first and second two-species mixture

46
populations for the traits δ13
C and LCC:LNC. Significant differences in the mean values were
rather detected between individuals of the target species present in the first and those in the
second two-species mixture populations.
For the remaining traits examined, there were no significant differences in the mean values
between individuals of the target species in the monoculture population and those in the first or
second two-species mixture populations.
Table 4.11 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species P. macrophylla in the monoculture population and the two-
species mixture populations
Trait Population N Mean SD Median Mean rank K-W p-value
1. H (m) Pm Cp 5 17.76 4.47 17.47 5.4a 8.66 0.013*
Pm 5 17.80 4.89 20.80 5.8a
Pm Zg 5 27.90 3.43 28.80 12.8b
2. DBH (mm) Pm 5 310.60 173.79 380.00 6.8 2.54 0.281
Pm Cp 5 461.20 103.15 443.00 10.6
Pm Zg 5 359.80 168.19 336.00 6.6
3. LPC (10-2 mg/g) Pm 5 0.10 0.00 0.10 7.5 0.40 0.818
Pm Cp 5 0.10 0.00 0.10 7.5
Pm Zg 5 0.12 0.03 0.13 9.0
4. LNC (10-2 mg/g) Pm 5 3.46 0.36 3.37 5.6 2.26 0.323
Pm Cp 5 3.63 0.00 3.63 9.0
Pm Zg 5 3.64 0.48 3.66 9.4
5. δ15N (‰) Pm 5 4.87 0.75 4.99 5.0a 9.85 0.007*
Pm Cp 5 5.29 0.00 5.29 6.0ab
Pm Zg 5 6.72 0.48 6.74 13.0b
6. LCC (10-2 mg/g) Pm 5 46.94 0.90 47.08 4.8a 6.97 0.031*
Pm Zg 5 47.34 0.29 47.28 7.2ab
Pm Cp 5 47.75 0.00 47.75 12b
7. δ13C (‰) Pm Cp 5 -32.65 0.00 -32.65 4.0a 7.55 0.023*
Pm 5 -31.29 1.64 -31.40 8.4ab
Pm Zg 5 -30.94 0.34 -30.88 11.6b
8. LCC:LNC Pm 5 13.78 1.21 14.11 10.4 2.26 0.323
Pm Cp 5 13.15 0.00 13.15 7.0
Pm Zg 5 13.01 1.90 12.87 6.6
9. LCC:LPC Pm 5 486.85 10.73 487.87 7.0 1.53 0.460
Pm Cp 5 494.82 0.00 494.82 10.0
Pm Zg 5 434.46 89.82 440.22 7.0
*p- value of <0.05 is statistically significant; Values within treatments for each analysis in the column Mean rank followed
by the same letter are not significantly different at p < 0.05 according to Dunn’s multiple range test.
An examination of the findings in the Table 4.11 indicates that that the results of the Kruskal
Wallis tests applied to the nine traits examined for the individuals of the target species P.
macrophylla in the monoculture population and in the two two-species mixture populations (P.
macrophylla + C. procera and P. macrophylla + Z. gilletii) revealed statistically differences at
level of p < 0.05 for the mean values of four traits. These are H (K-W = 8.66, DF = 2, p = 0.013
< 0.05), δ15
N (K-W = 9.85, DF = 2, p = 0.007 < 0.05), LCC (K-W = 6.97, DF = 2, p = 0.031 <
0.05) δ13
C (K-W = 7.55, D F= 2, p = 0.023 < 0.05).
As shown by the Dunn’s multiple pairwise comparisons, individuals of the target species in the
monoculture population were significantly shorter (Mean = 17.80 ± 4.89 m) and had a
statistically lower δ15
N value (Mean = 4.87 ± 0.75 ‰) than those in the two-species mixture

47
population composed of a combination of P. macrophylla and Z. gilletii (Mean = 27.90 ± 3.43 m
for H and Mean = 6.72 ± 0.48 ‰ for δ15
N).
In addition to that, individuals of the target species in the monoculture population exhibited a
significantly low LCC (Mean = (46.94 ± 0.90) x 10-2 mg/g) compared to the individuals of the
target species encountered in the two-species mixture population composed of a combination of
P. macrophylla and C. procera (Mean = (47.75 ± 0.00) x 10-2 mg/g).
Individuals of the target species in the monoculture population did not significantly differ from
the individuals of the target present in both two-species mixture population as far as the mean
values of the trait δ13
C was concerned. Instead, it is the individuals of target species present in
the first and second two-species mixture populations that significantly differed for the mean
values of this trait (δ13
C).
For the remaining traits examined, there were no significant differences in the mean values
between individuals of the target species in the monoculture population and those in the first or
second two-species mixture populations.
The results of Kruskal Wallis tests in Table 4.12 indicate that there was a significant difference
in the mean values among individuals of the target species P.elata present in the two
monoculture populations (plot No. 13 and No. 2) and those present in the four two-species
mixture populations (P.elata + G. cedrata; P.elata + B. welwitschii; P.elata + P. oleosa; P.elata
+ S. tetrandra) for the trait H (K-W =11.48, DF = 5, p = 0.043 < 0.05).
The multiple pairwise comparisons using Dunn’s procedure revealed a significant difference in
the height between individuals of the target species in the first monoculture populations (Plot
No.13) (Mean = 28.60 ± 6.91 m) and those in the two-species mixture population made of a
combination of P.elata + G. cedrata (Mean = 37.73 ± 0.39 m).
For the remaining traits examined, there were no significant differences in the mean values
between individuals of the target species in neither of the two monoculture populations and those
in the four two-species mixture populations.

48
Table 4.12 Descriptive statistics and Kruskal Wallis test for the trait difference between
individuals of the target species P.elata in the monoculture populations and the two-species
mixture populations
Trait Population N Mean SD Median Mean
rank
K-W p-value
1. H (m) Pe13 5 28.60 6.91 25.70 9.0a 11.48 0.043*
Pe2 5 32.52 1.95 32.00 11.2ab
Pe Bw 5 34.24 3.55 33.40 15.9ab
Pe Po 5 33.64 3.02 34.70 13.4ab
Pe St 5 34.24 5.30 35.90 17.6ab
Pe Gc 5 37.73 0.39 37.77 25.9b
2. DBH (mm) Pe13 5 279.40 52.92 279.00 10.0 2.7361 0.7406
Pe St 5 367.00 172.88 303.00 14.6
Pe2 5 352.00 85.48 335.00 16.5
Pe Bw 5 336.40 50.35 345.00 16.7
Pe Gc 5 425.40 222.85 409.00 17.4
Pe Po 5 365.80 86.78 344.00 17.8
3. LPC (10-2 mg/g) Pe Po 5 0.12 0.01 0.12 9.2 6.35 0.274
Pe2 5 0.13 0.01 0.13 13.8
Pe Bw 5 0.13 0.01 0.12 14.2
Pe Gc 5 0.13 0.02 0.13 15.8
Pe13 5 0.15 0.03 0.13 17.4
Pe St 5 0.15 0.02 0.15 22.6
4. LNC (10-2 mg/g) Pe13 5 4.20 0.15 4.21 9.2 6.44 0.266
Pe Gc 5 4.18 0.52 4.06 12.6
Pe Bw 5 4.35 0.21 4.30 14.6
Pe2 5 4.37 0.10 4.37 15.7
Pe Po 5 4.45 0.21 4.51 19.5
Pe St 5 4.60 0.33 4.48 21.4
5. δ15N (‰) Pe Gc 5 3.86 0.31 3.91 7.6 7.40 0.192
Pe13 5 4.27 0.84 4.20 14.2
Pe St 5 4.39 0.35 4.41 14.4
Pe Po 5 4.41 1.52 5.46 16.8
Pe2 5 4.66 0.24 4.69 18.1
Pe Bw 5 5.06 0.96 4.96 21.9
6. LCC (10-2 mg/g) Pe Gc 5 47.59 1.09 47.73 11.0 2.71 0.744
Pe St 5 47.88 0.51 48.04 13.0
Pe2 5 48.10 0.45 48.14 16.3
Pe13 5 48.20 1.60 48.18 16.4
Pe Bw 5 48.56 1.08 48.13 18.0
Pe Po 5 48.32 0.70 48.23 18.3
7. δ13C (‰) Pe13 5 -29.98 0.50 -30.01 7.8 11.22 0.05
Pe Gc 5 -30.12 1.61 -29.54 10.8
Pe Po 5 -29.43 0.29 -29.46 12.6
Pe Bw 5 -28.81 0.82 -28.64 18.6
Pe2 Pe2 5 -28.47 0.83 -28.54 21.4
Pe St 5 -28.54 1.22 -28.21 21.8
8. LCC:LNC Pe St 5 10.46 0.79 10.77 9.4 6.55 0.256
Pe Po 5 10.86 0.41 10.70 11.2
Pe2 5 11.01 0.26 10.89 15.2
Pe Bw 5 11.19 0.54 11.26 17.4
Pe Gc 5 11.51 1.26 12.07 18.60
Pe13 5 11.47 0.42 11.58 21.2
9. LCC:LPC Pe St 5 323.58 45.31 332.15 8.4 6.86 0.231
Pe13 5 342.33 69.17 364.08 14.4
Pe2 5 362.16 20.85 361.55 14.6
Pe Gc 5 366.47 63.81 375.96 15.2
Pe Bw 5 373.69 25.41 380.64 18.0
Pe Po 5 406.79 47.31 417.26 22.4
*p- value of <0.05 is statistically significant; Values within treatments for each analysis in the column Mean rank followed
by the same letter are not significantly different at p < 0.05 according to Dunn’s multiple range test.

49
The following highlights capture the key points of all the results presented and described above:
The interaction of the individuals of the admixed species D. likwa with those of the target
species A. congolensis in the mixture population led the latter to increase their size (H and DBH)
and as well as their δ15
N values in comparison with the monoculture population.
The interaction of individuals of the admixed species K. anthotheca with those of the target
species L. trichilioides in the mixture population made the latter to lower their LNC and their
δ15
N value whereas they increased their LCC, their δ13
C value and their LCC:LNC ratio value in
the mixture population compared to the monoculture population.
The interaction of the individuals of the admixed species Phyllanthus sp. with the individuals of
target species M.excelsa in the mixture population resulted in the decrease of LPC, LNC and that
of the δ15
N value of the target species in the mixture population compared to the monoculture
population. On the contrary, the LCC, LCC:LNC and LCC:LPC ratio values were increased.
The interaction of the individuals of the admixed species T. africana with the individuals of
target species P. soyauxii in the mixture population resulted in the increase of the target species
DBH and in the decrease of LCC of the target species in the mixture population as compared to
the monoculture population.
The interaction of the individuals of the admixed species C. africanum with those of the target
species P. tessmannii led to the decrease of the LCC of the target in the mixture population in
comparison to the monoculture population. The LCC:LNC and LCC:LPC ratios values of target
species were also decreased in the mixture population in comparison to the monoculture
population.
The interaction of the individuals of the admixed species A. nannanii with the individuals of
target species E. cylindricum in the mixture population resulted in an increase of the δ15
N value
and a decrease of the LCC:LNC ratio value of the target species in the mixture population in
comparison to the monoculture population.
The interaction of the individuals of the target species E. cylindricum with the individuals of the
admixed species E. angolense resulted in an increased LNC and an decreased LCC:LNC ratio
value for the target species in the mixture population when compared to monoculture population.
On the contrary, the LPC of the target species was lowered while its LCC:LPC ratio value was
augmented in the mixture population as compared to the monoculture population.
The interaction of the individuals of the admixed species L. trichilioides with those of the target
species G. cedrata induced a decrease in the δ15
N value of the target species while increasing its
LCC:LPC ratio value in the mixture population compared to the monoculture population.
The interaction of the individuals of the admixed species C. procera and the individuals of the
target species P. macrophylla made the latter to increase their LCC in the mixture population as
compared to the monoculture population.

50
The interaction of the individuals of the admixed species Z. gilletii with those of the target
species P. macrophylla resulted in an increased δ15
N value of the target species in the mixture
population as compared to the monoculture population. This interaction also induced an increase
in the H of the target species in the mixture population compared to the monoculture.
The interaction of the individuals of the admixed species G. cedrata with the individuals of
target species P. elata resulted in an increase in the H of the target species in the mixture
population compared to the second monoculture population (i.e plot No.13).
4.3.2 Multi-trait approach
The results of the between analysis tests for detecting the significance of grouping for the
BPCAs performed on each species are presented in Table 4.13.
Table 4.13 Between analysis tests for detecting the significance of grouping for the BPCAs
performed on each species
Population No. Perm. Obs. X < Obs. X ≥ Obs. P-value Ac 1000 2.86 995 5 < 0.005
Ac Dl
Ec 1000 5.94 1000 0 < 0.000
Ec An
Ec Ea
Gc 1000 3.35 991 9 < 0.009
Gc Lt
Gc Pe
Lt 1000 4.28 987 13 < 0.013
Lt Ka
Ma 1000 1.23 696 304 0.304ns
Ma Sg
Me 1000 4.26 990 10 <0.010
Me Psp
Pe2 1000 2.15 957 43 <0.043
Pe13
Pe Gc
Pe Bw
Pe Po
Pe St
Pm 1000 2.95 1000 0 <0.000
Pm Cp
Pm Zg
Po P 1000 1.34 764 236 0 .236ns
Po Pe
Ps 1000 2.99 983 17 <0.017
Ps Ta
Pt 1000 3.68 982 18 <0.018
Pt Ca
St 1000 1.12 618 382 0.382ns
St Pe
No. Perm= Number of permutations; Obs.= Observed value; X < Obs.= number of random values strictly
lower than the observed value; X ≥ Obs= number of random values strictly higher than the observed value,
p- value of <0.05 is statistically significant.

51
An examination of the findings in Table 4.13 shows that at significance level of 5 %, the nine
functional traits considered in the present study were able to significantly segregate the different
populations of target species (monocultures and two-species mixture populations) for nine out
twelve species examined (p-values ranging from <0.000 to <0.018).
The grouping of target species individuals in either the monoculture population or two-species
mixture population was not significant for the remaining three species, namely M. Africana (p =
0.304), P. oleosa (p = 0.236) and S. tetandra (p = 0.382).
For the grouping of target species with more than two populations (i.e. E. cylindricum, G.
cedrata, P. elata, P. macrophylla), additional between test analyses were performed to detect
whether the individuals in the monoculture populations could be significantly discriminated from
those in the two-species mixture populations. The results of these tests are summarized in table
4.14.
Table 4.14 Between analysis tests for detecting the segregation of monoculture populations from
two-species mixture populations for the grouping of target species with more than two
populations
Populations No. Perm. Obs. X < Obs. X ≥ Obs. P-value Ec vs. Ec/An 1000 4.12 998 2 < 0.002*
Ec vs. Ec/Ea 1000 5.85 999 1 <0.001*
Gc vs. Gc/Lt 1000 3.92 993 7 < 0.007*
Gc vs. Gc/Pe 1000 0.93 495 505 0.505ns
Pe2 vs. Pe/Bw 1000 0.48 140 860 0.860ns
Pe2 vs. Pe/Gc 1000 2.21 942 58 0.058ns
Pe2 vs. Pe/Po 1000 1.11 648 352 0.352ns
Pe2 vs. Pe/St 1000 1.48 855 145 0.145ns
Pe13 vs. Pe/Bw 1000 1.68 900 100 0.100ns
Pe13 vs. Pe/Gc 1000 0.99 597 403 0.403ns
Pe13 vs. Pe/Po 1000 2.17 960 40 0.040*
Pe13 vs. Pe/St 1000 1.73 916 84 0.084ns
Pe2 vs. Pe13 1000 1.98 949 51 0.051ns
Pm vs. Pm/Cp 1000 1.68 919 81 0.081ns
Pm vs. Pm/Zg 1000 2.081 983 17 <0.017*
No. Perm= Number of permutations; Obs.= Observed value; X < Obs.= number of random values strictly
lower than the observed value; X ≥ Obs= number of random values higher or equal than the observed value,
*p- value of <0.05 is statistically significant, ns= not significant.
These results in table 4.14 indicate that the nine functional traits taken together were able to
significantly segregate the individuals of the target species E. cylindricum present in the
monoculture population and those present in the first two-species mixture population composed
of E. cylindricum and A. nannanii (p < 0.002) on one hand, and those present in the second two-
species mixture population composed of E. cylindricum and E. angolensis (p < 0.001) on the
other hand.
Regarding the target species G. cedrata, the individuals in the monoculture population were
significantly discriminated from those in the first two-species mixture population constituted of
G.cedrata and L. trichilioides by the nine trait considered (p < 0.007), but not from those in the
second two-species mixture population constituted of G.cedrata and P. elata (p = 0.505).

52
The same pattern could also be recognized for the target species P. macrophylla whereby the
individuals constituting the monoculture population were significantly discriminated from those
making the two-species mixture population, P. macrophylla and Z. gilletii by the nine traits
considered (p < 0.017). However, the individuals belonging to the monoculture population could
not be significantly discriminated from the individuals constituting the two-species mixture
population, P. macrophylla and P. carapa by the traits considered (p = 0.081).
As for the target species P. elata, the individuals belonging to the first monoculture population
(plot No.2) did not significantly segregated from those in all the two-species mixture populations
in relation to the nine traits considered. The individuals in the second monoculture population
(plot No.13) could also not be discriminated from the individuals in the two-species mixture
populations by the nine traits considered, except for the individuals belonging to the two-species
mixture population made of P.elata and P.oleosa (p < 0.040). Additionally, there was no
significant discrimination between the individuals of the target species belonging to the first
monoculture population and those belonging to the second monoculture population (p = 0.051).

53
CHAPTER FIVE:
DISCUSSION
5.1. Variance partitioning
Part of the current study was concerned with knowing whether the majority of trait variation, for
the nine traits considered and across the twelve co-existing target species in the setup, was
situated within species or between species.
The variability within species (intra-specific) in the present study was higher than the variability
between species (inter-specific) for three of the nine traits considered, namely H and DBH and
LPC. For the above three mentioned traits, the intra-specific variability accounted for 69.08 %,
70.40 % and 73.98 % respectively of the overall variability when the single trait approach was
used whereas it accounted for 56.87 %, 64.97 % and 59.44 % respectively when the multi-trait
analysis taking into account the contribution of each of the nine variables (traits) was used.
At first sight, these results seem surprising considering the widely spread assumption that intra-
specific trait variation is negligible compared to variation among species (Garnier et al., 2001;
Baraloto et al., 2010; Auger and Shipley, 2013). However, studies where higher intra-specific
variations were measured in functional traits compared to inter-specific variations have already
been reported before.
Studying the variation in the functional traits LMA and LDMC across six nested ecological
scales, along a precipitation gradient for 119 tree species and 1910 leaves in lowland tropical
rainforests of East-West Panama, Messier et al. (2010) estimated the contribution of the intra-
specific variation to the overall variation for both traits at 48 % whereas the strictly inter-specific
variation fraction accounted for 21 % and 35 % respectively for LMA and LMDC. In their study
on variation in leaf functional trait values within and across individuals and species in a Costa
Rican dry forest, Hulshof and Swenson (2010) estimated the fraction of total variance within
species of 62 % and 68 % for the trait LWC when the leaves were collected at high (sun) and
low (shade) canopy positions, respectively.
These two studies that have reported a higher proportion of intra-specific variation relative to the
inter-specific variation for certain traits involved tree species in tropical forests as it is also the
case of the present study. However, a strong and valid generalization cannot be drawn only on
the basis of these few studies.
The intra-specific variability for the six remaining traits considered in the present study remained
lower than the inter-specific variability. However, except for the traits LNC and LCC:LNC, the
fraction of total variance within species for these traits was not marginal at all as it was
hypothesized at beginning of this study. It ranged from 23.29 % to 49.71 % when the single trait
approach was used and from 26.36 % to 38.33 % when the multi-trait analysis taking into
account the contribution of each of the nine variables (traits) was used. When the nine traits were

54
considered as a whole, the multi-trait variance decomposition resulted in fraction of 36.87 % for
the intra-specific variation against 63.13 % for inter-specific variation for the twelve species
studied. Using the same approach in a study involving thirteen herbaceous and shrub species and
the functional trait H, SLA, LDMC, LNC and LCC in the French Alps, Albert et al. (2010) also
noted a relatively strong component of intra-specific variability of about 30 % against 70 % for
the inter-specific component.
These results seem also to corroborate observations made previously that the fraction of total
variance between species relative to that within species is strongly dependent on the species and
traits considered. For instance, Wilson et al. (1999) noted that the intra-specific variability for
the trait SLA was about 8 % when all the plants of the considered dataset were incorporated into
the analysis. However, when only angiosperm species were taken into account in the analysis,
the fraction of total variance within species raised to 32 %. The same was true for the trait
LMDC for which the intra-specific variability increased from 6 % when all the plants were
considered to 14 % when only angiosperms were taken into account. As already stated by Albert
et al., (2010), it can be reaffirmed here that any decision seeking to know whether or not the
intra-specific variability can be neglected should be based on the system considered but also on
the selected trait and species.
A probable explanation for the fairly strong proportion of the intra-specific trait variation
obtained in this study could come from the fact that there might have been occurrence of biotic
interactions and niche complementarily between individuals of the target species at the
neighborhood scale. As explained by Violle et al. (2012), the appearance of biotic interactions
and niche complementarily at the neighborhood scale might force individuals of the same species
to adjust their trait values in response to the activity of their closest neighbor. As a result, the
relative importance of intra-specific trait variation to the overall trait variation is expected to
increase.
For the traits like δ15
N, LPC and H for which a fairly good fraction of between population
variation was observed, it can be assumed that this was as a result of the biotic interaction of the
target species with different admixed tree species to which were associated. The difference in the
species composition of different populations (plots) in this arboretum must have mediated a
relative higher between population variation in the particular case of the above mentioned traits.
5.2 Main functional trade-offs and strategies
The BPCA confirmed the well established leaf economics spectrum (Wright et al., 2004). In the
first dimension, species were separated according to their LCC and nutrient content (LPC and
LNC). Species with low nutrient content are expected to have thicker leaves. They are
characterized by a long life span and a slow return on investment. Species exhibiting high
nutrient content on the contrary are supposed to bear thinner leaves and are characterized by a
short life span and a fast return on investment. This illustrates well the acquisitive and
exploitative ecological strategies adopted by the species under investigation (Diaz et al., 2004).

55
In the second dimension, variations were mainly led by δ13
C, H, DBH. This reflects the strategies
utilize by these species in terms of water use efficiency, accessibility to light and investment in
perennial structures (Faster and Westoby, 2005). Plant H in particular has been mentioned as the
second main axis of variation in many previous studies (Diaz et al., 2004; Gross et al., 2007;
Albert et al., 2010).
The functional diversity exhibited by the considered species as portrayed by the BPCA axes
seems to suggest that within this planted tropical forest of Yangambi, target species co-exist with
diverse strategies that allow them to occupy different niches. However, the fact that a relatively
good number of the species investigated were situated around the centre of origin of the plane
(trait space) is an indication that they are characterized by an average values for the traits driving
the main axes variation in both the first and second dimension. Therefore, the co-existence of
these species in that arboretum may be explained by both the niche theory (Silvertown, 2004)
and the neutral theory (Hubblel, 2005).
The WPCA which focuses on axes that discriminate individuals within species in the trait space
seems to indicate that the main functional strategies adopted by the sampled species at the inter-
specific level were also maintained at the intra-specific level supporting the second hypothesis
made in this study. Though less marked, trait tradeoffs in terms of leaf economics and
accessibility to light and investment in perennial structures within species were broadly
analogous to that of between species.
Albert et al. (2010) in their study on herbaceous and shrub species in the French Alps made the
same observation that inter-specific functional strategies were broadly conserved at intra-specific
level. Maire et al. (2013) in a study among grass species involving two traits negatively
correlated along the LES (SLA and LLS) and two other negatively correlated traits (H and tiller
density: TD) but independent from the LES also indicated that that the trade-offs observed at the
inter-specific level were preserved at the intra-specific level as well. Contrary that these findings,
Boucher et al. (2013) in their study on the functional variability in the wild populations of the
herbaceous species Polygonum viviparum L. seemed to suggest that the main functional trade-
offs exhibited by plants at inter-specific level over large geographical scale could be different
from the ones at the population level.
5.3 Trait differences between target species in monocultures and two-species mixtures
5.3.1 Single trait approach
Using the single trait approach, significant differences in mean values for at least one trait (and
up to six traits) out of the nine traits examined were detected between individuals present in
monoculture populations and those in two-species mixture populations for nine species out of the
twelve sampled.
In the light of the results obtained from this part of the study, there seems to be phenotypic
plasticity for some of the studied traits in this planted tropical forest in line with the third
hypothesis of this work. Since it was the individuals of the same species occupying different

56
plots (i.e. belonging to different populations) that were each time compared, the trait differences
observed are presumably induced by changes in the growth environment rather than by genotype
differences.
The biotic interaction of some target species present in this arboretum with particular admixed
species might have led them to adjust their functional some of their functional trait values in
order to cope with the changing environmental conditions to which they were subjected. Because
the target species and their respective admixed species are native to the same region and co-exist
together, there is reason to believe that the new environmental conditions created by their
interaction were most probably driven by inter-specific competition for resources. The
availability of light, water and nutrients has been identified as the main cause responsible for
plant strategies in space and time (Ordonez, 2010).
Depending on the target species, the admixed species to which it was associated and the trait
involved, the mean trait value either increased or decreased in the mixture population compared
to the monoculture population. The traits that regularly showed a significant difference in the
mean values between individuals in different monoculture populations and those in two-species
mixture population were δ15
N, LCC and LCC:LNC. In their study on functional leaf trait
diversity of tree species in a Congolese secondary tropical forest, Verbeeck et al. (2014) had also
noted that the leaf trait δ15
N was strongly influenced by plot location.
In the case of this study, a pattern seemed to emerge throughout different configurations of the
arboretum pointing out an interplay between the LCC and LNC and its isotope δ15
N. Whenever
the interaction of an admixed species with a target species led to an increase in the LNC and/or
the δ15
N value in the mixture population in comparison to the monoculture population, the LCC
value was almost always lowered and vice-versa. This could be as a result of the competition
from the admixed species on the target species for N, water or light resulting, perhaps, in a
divergent allocation of C and N within the leaves of the target species in the mixture populations
in comparison to those in monoculture populations.
N is considered as one of the most important nutrients steering plant composition and richness,
and plants need C as source of energy for their growth (biomass accumulation). The C economy
and the N economy are known to be closely linked. It has been demonstrated for instance that
plants allocate more biomass to roots versus leaves and increase their N concentration when
water is in short supply (Ye et al., 2015). It is also known that in low light environments, plants
exhibit higher leaf-to-root ratios and lower their N content than in high light environments
(Suguira and Tateno, 2011). In a similar way, fast growing plant species are believed to allocate
more biomass to the roots in suboptimal N supply conditions (Aerts and chapin, 2000).
The observed interplay of LCC and LNC in target species as a result of their interaction with
admixed species seems also to be a good illustration of how phenotypic plasticity may lead to
niche differentiation, thus avoiding competitive exclusion. Through phenotypic plasticity,
organisms are able to adjust to an array of conditions without evolutionary changes. Siefert
(2012) explained that apart from genetic variation, phenotypic plasticity could also make

57
species’ trait values to differ among locations as response to environmental filters and interaction
with neighboring species. Niche differentiation among species can be interpreted as the capacity
of species to exploit diverse environments, and it is sometimes expressed as changes in species
performance (Roscher et al., 2015). It has been identified as one of the main factors driving
species co-existence (Levine et al., 2009).
The interaction of the individuals of the target species M. Africana, P. oleosa and S. tetandra
with their respective admixed species resulted in no significant difference in the mean values of
all the traits examined between the individuals of these target species in monoculture populations
and those in mixture populations. A plausible explanation to that could come from the neutral
theory. It states that co-existing species are functionally the same; therefore, they share similar
niches (Hubbell, 2005). The interaction of the individuals of the above mentioned target species
with their respective admixed species did probably not lead to any change in their respective
environmental conditions. Therefore, there was no reason for them to differ from their
counterparts in monoculture populations with respect to the considered functional traits.
5.3.2 Multi-trait approach
Another interesting lesson learnt from the results of this study came from multi-trait analyses that
were used to test the significance of grouping for the BPCAs performed on each species. The
results of these tests suggest that the nine functional traits used in this study significantly
segregated the different monoculture populations from the mixture populations for nine target
species out of the twelve tested. This also implies that the individuals of the different populations
that were compared were significantly different in terms of the considered functional traits.
The three target species for which the monoculture populations could not be significantly
segregated from the mixture population were M. africana, P. oleosa and S. tetandra. These
results were consistent with those obtained when the single trait analyses were used. They seem
to suggest that a difference in a single trait between a monoculture population and a mixture
population for a particular target species was enough to make the two groups different when the
nine traits were considered together. Since many plant functional traits tend to co-vary together,
it is quite possible that a plastic response of single trait to changing environmental conditions
could also impact functionally correlated traits when a suite of traits are considered together
(Roscher et al., 2015). Therefore, co-variations among species traits could be playing an
important role in the overall performance of plants.

58
CHAPTER SIX:
GENERAL CONCLUSION AND RECOMMENDATIONS
6.1 General conclusion
The variance decomposition performed in this study showed that the inter-specific variation
dominated for all the traits examined except for the trait DBH and H and LPC. Despite this fact,
the intra-specific variation was not negligible. The overall partition taking into account the nine
functional traits and twelve species studied together was estimated at 36.87 % for the intra-
specific variation against 63.13 % for inter-specific variation. This fairly good proportion of
intra-specific variation vs. inter-specific variation can be attributed the appearance of biotic
interactions and niche complementarily at the neighborhood scale between individuals of the
same species. The consequence of this is an increase in the relative importance of intra-specific
variation. These results point to the importance of incorporating intra-specific trait variation into
community assembly analyses.
The functional dimensions identified in this study were in accordance with the well known leaf
economy spectrum. Species were segregated depending on the LCC, LNC and LPC in the first
dimension. In the second axis, variation was led by δ13
C, H, DBH. It was also shown that the
main functional strategies adopted by the investigated species at the inter-specific level were also
maintained at the intra-specific level.
Using single trait-analyses, significant differences in trait values were detected between target
species in monocultures and in two-species mixtures for nine species out of the twelve
investigated. An apparent interplay between the traits δ15
N, LNC and LCC was recognized
throughout different configurations of the arboretum. An increase in LNC and/or δ15
N value in
the mixture population of a specific target species due the interaction of its individuals with those
of the admixed species was almost always followed by a reduction in the LCC and vice versa.
This tends to indicate the occurrence of phenotypic plasticity among the individuals of the target
species in the mixture populations as a result of their competition with their respective admixed
species allowing a divergent allocation of C and N within their respective leaves. The occurrence
of this phenotypic plasticity was probably responsible for the niche differentiation explaining the
co-existence of these target species with the admixed species which they were associated to.
For three target species, namely M. africana, P. oleosa and S. tetandra, no significant differences
in trait values were detected between individuals in monocultures and those in two-species
mixtures for all the nine traits examined. Most probably the interaction of these target species
with their respective admixed species resulted in no major inter-specific competition for
resources. Hence, the neutral theory was proposed as the possible cause explaining the co-
existence of these target species with the respective admixed species with which they were
combined.

59
The results of the multi-trait analyses that were used to test if the nine functional traits taken
together could significantly differentiate between the monoculture populations and mixture
populations of the target species were in line with the results obtained from the single trait
analyses. It was found that a difference in a single trait between a monoculture population and a
mixture population for a particular target species as detected through the single trait analysis was
enough to make the two groups different when the nine traits were considered together.
Altogether this study has shown how intra-specific variation was mediating the co-existence of
target species in this arboretum by accounting for good proportion of the total variation for most
traits, by maintaining the same functional trade-offs and strategies at the intra-specific level and
at the inter-specific level, and through the occurrence of phenotypic plasticity.
These results have highlighted the important role that the intra-specific trait variation may play in
determining tree species co-existence. Therefore, the intra-specific trait variation should not be
systematically neglected in quantitative functional trait-based analyses. The decision on whether
or not to neglect the intra-specific trait variation should be made on a case-by-case basis taking
into account the trait, the species and the system under investigation.
6.2. Recommendations
To be complete and to confirm some of the outcomes of this study, these investigations should
be extended to the admixed species. It will be of particular importance to compare the trait
values of the target species in monoculture populations and in mixture populations with those of
the admixed species in mixture populations. In addition, studies on nutrient cycling and the
pattern of C and N allocation to roots and leaves of the species in the arboretum will also be
needed. This will allow elucidating further the role of inter-specific competition between the
target species and admixed species as a probable cause of the observed variation in the trait
values of the target species in monoculture populations and mixture populations.

60
REFERENCES
Ackerly, D. D., Dudley, S. A., Sultan, S. E., Schmitt, J., Coleman, J. S., Linder, R., Sandquist,
D. R., Geber, M. A., Evans, A. S., Dawson, T. E., Lechowicz M. J. 2000. The evolution of
plant ecophysiological traits: recent advances and future directions. Bioscience 50:979-995.
Aerts, R. and Chapin, F. S. 2000. The mineral nutrition of wild plants revisited: a re-evaluation
of processes and patterns. Advance in Ecological Research 30: 1-67.
Albert C. H., Grassein F., Schurr F. M., Vieilledent G. and Violle C. 2011. When and how
should intraspecific variability be considered in trait-based plant ecology? Perspectives in
Plant Ecology, Evolution and Systematics 13:217-225.
Albert, C. H., Thuiller, W., Yoccoz, N. G., Douzet, R., Aubert, S. and Lavorel, S. 2010. A multi-
trait approach reveals the structure and the relative importance of intra- vs. interspecific
variability in plant traits. Functional Ecology 24:1192-1201.
Alcoverro, T., Manzanera, M. and Romero, J. 2000. Nutrient mass balance of the sea grass
Posidonia oceanica: the importance of nutrient retranslocation. Marine Ecology Progress
Series 194: 13-21
Arndt, S. K and Wanek, W. 2002. Use of decreasing foliar carbon isotope discrimination during
water limitation as a carbon tracer to study whole plant carbon allocation. Plant, Cell and
Environment 25: 609-616.
Auger, S. and Shipley, B. 2013. Inter-specific and intra-specific trait variation along short
environmental gradients in an old-growth. Journal of Vegetation Science 24: 419-428.
Badyaev, A. V. 2009. Evolutionary significance of phenotypic accommodation in novel
environments: an empirical test of the Baldwin effect. Philosophical Transactions of the
Royal Society B 364:1125-1141.
Baker, T. R., Phillips, O. L., Malhi, Y., Almeida, S., Arroyo, L. Arroyo, L., Di Fiore, A.,
Killeen, T. J., Laurance, S. G., Laurance, W. F., Lewis, S. L., Lloyd, J., Monteagudo, A.,
Neill, D. A., Patino, S., Pitman, N. C. A., Macedo Silva, J. N. and Vasquez Martinez, R.
2004. Variation in wood density determines spatial patterns in Amazonian forest biomass.
Global Change Biology 10:545-562.
Baraloto, C., Hardy, O.J., Paine, C. E. T., Dexter, K. G., Cruaud, C., Dunning, L. T., Gonzalez,
M., Molino, J., Sabatier, D., Savolainen, V. and Chave, J. 2012. Using functional traits and
phylogenetic trees to examine the assembly of tropical forest tree communities. Journal of
Ecology 100:690-701.
Baraloto, C., Paine, C. E. T., Poorter, L., Beauchene, J., Bonal, D., Domenach, A. M., Hérault,
B., Patiño, S., Roggy, J. C. and Chave, J. 2010. Decoupled leaf and stem economics in rain
forest trees. Ecology Letters 13: 1338-1347.
Bazzaz, F. A. and Pickett, S. T. A. 1980. Physiological ecology of tropical succession: A
comparative review. Annual Review of Ecology and Systematics 11:287-310.

61
Bolnick, D. I., Amarasekare, P., Araújo, M. S., Bürger, R., Levine, J. M., Novak, M., Rudolf, V.
H. W., Schreiber, S. J., Urban, M. C., Vasseur, D. A. 2011. Why intra-specific trait
variation matters in community ecology. Trends in Ecology and Evolution 26(4):183-192.
Bonan, G. B. 2008. Forests and climate change: forcings, feedbacks, and the climate benefits of
forests. Science 320:1444-1449.
Boucher, F. C., Thuiller, W., Arnoldi, C., Albert, C. H and Lavergne, S. 2013. Unravelling the
architecture of functional variability in wild populations of Polygonum viviparum L.
Functional Ecology 27: 382-391.
Boyemba, B.F. 2011. Ecologie de Pericopsis elata (Harms) Van Meeuwen (Fabaceae), arbre de
forêt tropicale africaine à répartition agrégée. PhD thesis, Université Libre de Bruxelles,
Bruxelles, 181p.
Butler, R. 2014. Tropical rainforest of the world. www. rainforest.mongabay.com, retrieved on
24.07.2015
Carvalho, J. A., Higuchi, N., Araújo, T. M and Santos, J. C. 1998. Combustion completeness in a
rainforest clearing experiment in Manaus, Brazil. Journal of geophysical research 103:
13195-13199.
Cattanio, J. H., Kuehne, R. and Vlek, P. L. 2008. Organic material decomposition and nutrient
dynamics in a mulch system enriched with leguminous trees in the amazon. The Revista
Brasileira de Ciência do Solo32:1073-1086.
Chapman, H. D. and Pratt, F. P. 1961. Ammonium Vandate-molybdate method for determination
of phosphorus: Methods of analysis for soils, plants and water. 1st edition, Agriculture
Division, California University, USA, pp. 184-203.
Chave, J., Coomes, D., Jansen, S., Lewis, S. L., Swenson, N. G. and Zanne, A. E. 2009. Towards
a worldwide wood economics spectrum. Ecology Letters 12:351-366.
Clark, J. S., Bell, D., Chu, C., Courbaud, B., Dietze, M., Hersh, M., HilleRisLambers, J., Ibáñez,
I., LaDeau, S., McMahon, S., Metcalf, J., Mohan, J., Moran, E., Pangle, L., Pearson, S.,
Salk, C., Shen, Z., Valle, D. and Wyckoff, P. 2010. High-dimensional coexistence based on
individual variation: a synthesis of evidence. Ecological Monographs 80:569-608
Cornelissen, J. H. C., Lavorel, S., Garnier, E., Díaz, S., Buchmann, N., Gurvich, D. E., Reich, P.
B., ter Steege, H., Morgan, H. D, van der Heijden, M.G.A, Pausas, J. G.and Poorter, H.
2003. A handbook of protocols for standarised and easy measurement of plant functional
traits worldwide. Australian Journal of Botany 51:335-380.
Cornwell, W. K. and Ackerly, D. D. 2009. Community assembly and shifts in plant trait
distributions across an environmental gradient in coastal California. Ecological
Monographs 79:109-126.
Dale, V. H., Joyce, L. A., McNulty, S., Neilson, R. P., Ayres, M. P., Flanningan, M. D., Hanson,
P. J., Irland, L. C., Lugo, A. E., Peterson, C. J., Simberloff, D., Swanson, F. J., Stocks, B. J.
and Wotto, B. M. 2001. Climate change and forest disturbances. BioScience 51:723-34

62
de Bello, F., Lavorel, S., Albert, C.H., Thuiller, W., Grigulis, K., Dolezal, J., Janecek, S. and
Leps, J. 2011. Quantifying the relevance of intraspecific trait variability for functional
diversity. Methods in Ecology and Evolution 2: 163-174.
Díaz, S. and Cabido, W. 2001. Vive la différence: plant functional diversity matters to ecosystem
processes. Trends in Ecology and Evolution 16(11):646-655.
Diaz, S., Hodgson, J. G., Thompson, K., Cabido, M., Cornelissen, J. H. C., Jalili, A., Montserrat
Marti, G., Grime, J. P., Zarrinkamar, F., Asri, Y., Band, S. R., Basconcelo, S., Castro-Diez,
P., Funes, G., Hamzehee, B., Khoshnevi, M., Perez-Harguindeguy, N., Perez-Rontome,
M.C., Shirvany, F. A., Vendramini, F., Yazdani, S., Abbas-Azimi, R., Bogaard,A.,
Boustani, S., Charles, M., Dehghan, M., De Torres-Espuny, L., Falczuk, V.,Guerrero-
Campo, J., Hynd, A., Jones, G., Kowsary, E., Kazemi-Saeed, F., Maestro- Martinez, M.,
Romo-Diez, A., Shaw, S., Siavash, B., Villar-Salvador, P. and Zak,M. R. 2004. The plant
traits that drive ecosystems: evidence from three continents. Journal of Vegetation Science
15: 295-304.
Dodélec, S. and Chessel, D. 1991. Recent developments in linear ordination methods for
environmental sciences. Advances in Ecology, India, 1: 133-155.
Ernst, C., Mayaux, P., Verhegghen, A., Bodart, C., Musampa, C. and Defourny, P. 2013.
National forest cover change in Congo Basin: deforestation, reforestation, degradation and
regeneration for the years 1990, 2000 and 2005. Global Change Biology 19:1173-1187.
Falster, D. S. and Westoby, M. 2005. Alternative height strategies among 45 dicot rain forest
species from tropical Queensland, Australia. Journal of Ecology 93:521-535.
FAO. 2010. Global forest resources assessment 2010. Forestry paper 163, FAO, Rome. 340p.
Fichman, M. and Cummings, J. M. 2003. Multiple imputation for missing data: Making the most
of what you know. Organizational Research Methods 6(3):282-308.
Field, C., and Mooney, H. A. 1986.The photosynthesis-nitrogen relationship in wild plants. In:
On the economy of plant form and function: adaptative patterns of energy capture in
plants. (T. J. Givnish, ed.), pp. 25-56, Cambridge University Press, Cambridge.
Fonseca, C. R., Overton, J. M., Collins, B. and Westoby, M. 2000. Shifts in trait combinations
along rainfall and phosphorous gradients. Journal of Ecology 88:964-977.
Fortunel, C., Garnier, E., Joffre, R., Kazakou, E., Quested, H., Grigulis, K., Lavorel, S., Ansquer,
P., Castro, H., Cruz, P., Dolezal, J., Eriksson, O, Freitas, H., Golodets, C., Jouany, C,
Kigel, J, Kleyer, M., Lehsten, V, Leps, J, Meier, T., Pakeman, R., Papadimitriou, M.,
Papanastasis, V. P., Quétier F., Robson, M., Sternberg, M., Theau, J.P., Thébault, A. and
Zarovali, M. 2009. Leaf traits capture the effects of land use changes and climate on litter
decomposability of grasslands across Europe. Ecology 90 (3): 598-611.
Franklin, J. F., Spies, T. A., Van Pelt, R., Carey, A. B., Thornburgh, D.A., et al., 2002.
Disturbances and structural development of natural forest ecosystems with silvicultural
implications, using Douglas fir forests as an example. Forest Ecology and Management
155:399-423

63
Fridley, J. D., Grime, J. P. and Bilton, M. 2007. Genetic identity of interspecific neighbours
mediates plant responses to competition and environmental variation in a species-rich
grassland. Journal of Ecology 95:908-915.
Gadow, K., Zhang, C. Y., Wehenkel, C., Pommerening, A., Corral-Rivas, J., Korol, M.,
Myklush, S., Hui, G. Y., Kiviste, A. and Zhao, X. H. 2012. Forest structure and diversity.
Managing Forest Ecosystems 23:29-83.
Garnier, E., Cordonnier, P., Guillerm, J. L. and Sonié, L. 1997. Specific leaf area and leaf
nitrogen concentration in annual and perennial grass species growing in Mediterranean old-
fields. Oecologia, 111: 490-498.
Garnier, E., Laurent, G., Bellmann, A., Debain, S., Berthelier, P., Ducout, B., Roumet, C., Navas
and M. L. 2001. Consistency of species ranking based on functional leaf traits. New
Phytologist 152: 69-83.
Gibbs, H. K., Brown, S., Niles, J. O. and Foley, J. A. 2007. Monitoring and estimating tropical
forest carbon stocks: making REDD a reality. Environmental Research Letters 2(4):054023
Giller, K.E. 2001. Nitrogen fixation in tropical cropping systems, 2nd
edition, CABI Publising,
Wallingford, UK. 423p.
Grime, J. P. and Pierce, S. 2012. The evolutionary strategies that shape ecosystems. Wiley-
Blackwell, Chichester, UK. 244p.
Grime, J. P., Thompson, K., Hunt, R., Hodgson, J. G., Cornelissen, J. H. C., Riorison, I. H.,
Hendry, G. A .F., Ashenden, T. W., Askew, A. P., Band, S. R., Booth, R. E., Bossard, C.
C., Campbell, B. D, Cooper, J.E., Davison, A. W., Gupta, P. L., Hall, W., Hand, D. W,
Hannah, M. A., Hillier, S. H., Hodkinson, D. J., Jalili, A., Liu, Z., Mackey, J. M. L.,
Matthews, N., Mowforth, M. A., Neal, R. J., Reader, R. J., Reiling, K., Ross-Fraser, W.,
Spencer, R. E., Sutton, F., Tasker, D. E., Thorpe, P. C. and Whitehouse, J. 1997. Integrated
screening validates primary axes of specialization in plants. 1997. Integrated screening
validates primary axes of specialization in plants. Oikos 79: 259-281.
Gross, N., Suding, K. N., Lavorel, S. and Roumet, C. 2007. Complementarity as a mechanism of
coexistence between functional groups of grasses. Journal of Ecology 95: 1296-1305.
Güsewell, S. 2004. N : P ratios in terrestrial plants: variation and functional significance. New
Phytologist 164(2): 243-266.
Hubbell, S. P. 2005. Neutral theory in community ecology and the hypothesis of functional
equivalence. Functional Ecology 19(1):166-172.
Hughes, A. R. and Stachowicz. J. J. 2004. Genetic diversity enhances the resistance of a seagrass
ecosystem to disturbance. Proceedings of the National Academy of Sciences USA
101:8998-9002.
Hulshof, C. M. and Swenson, N. G. 2010. Variation in leaf functional trait values within and
across individuals and species: an example from a Costa Rican dry forest. Functional
Ecology 24: 217-223.

64
IPCC, 2007. Climate Change 2007: Impacts, Adaptation and Vulnerability: Contribution of
Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on
Climate Change. Cambridge University Press, Cambridge, United Kingdom and New
York, NY, USA. 976p.
Jung, V., Violle, C., Mondy, C., Hoffmann, L. and Muller, S. 2010. Intraspecific variability and
trait-based community assembly. Journal of Ecology 98:1134-1140.
Kang, M., Chang, S. X., Yan, E-R. and Wang, X-H. 2014. Trait variability differs between leaf
and wood tissues across ecological scales in subtropical forests. Journal of Vegetation
Science 25: 703-714.
Kooyman, R. M. and Westoby, M. 2009. Costs of height gain in rainforest saplings: main-stem
scaling, functional traits and strategy variation across 75 species. Annals of Botany
104:987-993
Kotowska, A. M., Cahill, J. F. and Keddie., B. A. 2010. Plant genetic diversity yields increased
plant productivity and herbivore performance. Journal of Ecology 98:237-245.
Kraft, N. J. B., Valencia, R. and Ackerly, D. D. 2008. Functional traits and niche-based tree
community assembly in an Amazonian forest. Science 322:580-582.
Krieger, D. 2001. Economic value of forest ecosystem services: A review. The Wilderness
Society, Washington, D.C. 31p.
Laughlin, D. C. 2014. The intrinsic dimensionality of plant traits and its relevance to community
assembly. Journal of Ecology 102:186-193.
Lee, J.A. and Verleyson, M. 2007. Nonlinear Dimensionality Reduction. Springer, New York,
NY. 309p.
Levine, J. M., Hille Ris Lambers, J. 2009. The importance of niches for the maintenance of
species diversity. Nature 461: 254-257.
Linhart, Y. B. and Grant, M. C. 1996. Evolutionary significance of local genetic differentiation
in plants. Annual Review of Ecology and Systematics 27:237-277.
Liu, J., Zeng, D., Lee, D. K., Fan, Z. and Zhong, L. 2008. Leaf traits and their interrelationship
of 23 plant species in southeast of Keerqin Sandy Lands, China. Frontiers of Biology in
China 3: 332-337.
Lloyd, J., Bloomfield, K., Domingues, T. F. and Farquhar, G. D. 2013. Photosynthetically
relevant foliar traits correlating better on a mass vs an area basis: of ecophysiological
relevance or just a case of mathematical imperatives and statistical quicksand? New
Phytologist 199:311-321.
Maire, V., Gross, N., Hill, D., Martin, R., Wirth, C., Wright, I. J and Soussana, I. J. 2013.
Disentangling coordination among functional traits using anindividual-centred model:
Impact on plant performance at intra- and inter-specific levels. PLoS ONE 8(10): e77372.
doi:10.1371/journal.pone.0077372.

65
Marthews, T. R., Metcalfe, D., Malhi, Y., Phillips, O., Huaraca, H. W., Riutta, T., Ruiz, J.
M.,Girardin, C., Urrutia, R., Butt, N., Cain, R., Oliveras, M. I and colleagues from the
RAINFOR and GEM networks. 2012. Measuring tropical forest carbon allocation and
cycling: A RAINFOR-GEM field manual for intensive census plots (v2.2). Manual, Global
Ecosystems Monitoring network, http://gem.tropicalforests.ox.ac.uk/
McGroddy, M. E., Daufresne, T. and Hedin, L. O. 2004. Scaling of C : N : P stoichiometry in
forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 85(9): 2390-
2401.
MECNT, DRC/UN-REDD. 2012. Synthèse des études sur les causes de la déforestation et de la
dégradation des forêts en République Démocratique du Congo, Version Finale. MECNT,
Kinshasa. 40p.
Mediavilla, S. and Escudero, A. 2003. Leaf life span differs from retention time of biomass and
nutrients in the crowns of evergreen species. Functional Ecology 17: 541-548.
Meir, P., Kruijt, B., Broadmeadow, M., Barbosa, E., Kull, O., Carswell, F., Nobre, A. and
Jarvis, P.G. 2002. Acclimation of photosynthetic capacity to irradiance in tree canopies in
relation to leaf nitrogen concentration and leaf mass per unit area. Plant Cell Environment
25:343-357.
MENCT, RDC. 2009. The Democratic Republic of Congo’s REDD+ Potential. MENCT,
Kinshasa. 62p.
Messier, J., McGill, B. J. and Lechowicz, M. J. 2010. How do traits vary across ecological
scales? A case for trait-based ecology. Ecology Letters 13:838-848.
Miner, B. G., Sultan S. E, Morgan, S. G., Padilla, D. K., Relyea, R. A. 2005. Ecological
consequences of phenotypic plasticity. Trends in Ecology and Evolution 20(12):685-692.
Moreira, B., Tavsanoglu , Ç. and Pausas, J. G. 2012. Local versus regional intraspecific
variability in regeneration traits. Oecologia 168:671-677.
Ngongo, M. L., van Ranst, E., Baert, G., Kasongo, E. L., Verdoodt, A., Mujinya, B. B. and
Mukalay, J. M. 2009. Guide des sols en R.D. Congo. Tome I: Etude et Gestion. UGent -
HoGent-UNILU, 262 p.
Onoda, Y., Richards, A. E. and Westoby, M. 2010. The relationship between stem biomechanics
and wood density is modified by rainfall in 32 Australian woody plant species. New
Phytologist 185:493-501.
Ordonez, J. C., van Bodegom, P. M., Witte, J. P. M., Bartholomeus, R. P., van Hal, J. R., Aerts,
R. 2010. Plant strategies in relation to resource supply in mesic to wet environments: Does
theory mirror nature? The American Naturalist 175 (2): 225-239.
Osnas, J. L. D., Lichstein, J. W., Reich, P. B. and Pacala, S. W. 2013. Global leaf trait
relationships: mass, area, and the leaf economics spectrum. Science 340:741-744.

66
Pan, Y., Birdsey, A. R., Philipps, O. L. and Jackson, B. R. 2013. The structure, distribution and
biomass of the world’s forests. Annual Review of Ecology, Evolution, and Systematics.
44:593-662.
Pan, Y., Birdsey, R. A., Fang, J., Houghton, R., Kauppi, P. E., Kurz, W. A., Phillips, O. L.,
Shvidenko, A., Lewis, S. L., Canadell, J. G., Ciais, P., Jackson, R. B., Pacala, S. W.,
McGuire, A. D., Piao, S., Rautiainen, A., Sitch, S. and Ha, D. 2011. A large and persistent
carbon sink in the world’s forests. Science 333:988-993.
Pardo, L. H., Semaoune, P., Schaberg, P. G., Eagar, C. and Sebilo, M. 2013. Patterns in δ15
N in
roots, stems, and leaves of sugar maple and American beech seedlings, saplings, and
mature trees. Biogeochemistry, 112:275-291.
Pardo, L. H., Templer, P. H., Goodale, C. L., Duke, S., Groffman, P. M., Adams, O. M., Boeckx,
P., Boggs, J., Campbell, J. L., Colman, B., Compton, J., Emmett, B., Gundersen, P.,
Kjonaas, J., Lovett, G., Mack, M., Magill, A., Mbila, M., Mitchell, M. J., McGee, G.,
McNulty, S., Nadelhoffer, K., Ollinger, S., Ross, D., Rueth, H., Rustad, L., Schaberg, P.,
Schiff, S., Schleppi, P., Spoelstra, J., Wessel, W. 2006. Regional assessment of N saturation
using foliar and root δ15
N. Biogeochemistry 80:143-171.
Patiño, S., Fyllas, N. M., Baker, T. R., Paiva, R., Quesada, C. A., Santos, A. J., Schwarz, M., ter
Steege, H., Phillips, O. L. and Lloyd, J. 2012. Coordination of physiological and structural
traits in Amazon forest trees. Biogeosciences 9:775-801.
Pearce, D. W. 2001. The economic value of forest ecosystems. Ecosystem Health 7 (4): 284-296.
Peel, M. C., Finlayson, B. L., and McMahon, T. A. 2007. Updated world map of the Köppen-
Geiger climate classification. Hydrology and Earth System Sciences 11, 1633-1644.
Pérez-Harguindeguy, N., Díaz, S., Garnier, E., Lavorel, S., Poorter, H., Jaureguiberry, P., Bret-
Harte, M. S., Cornwel, W. K., Craine, J. M., Gurvich, D. E., Urcelay, C., Veneklaas, E. J.,
Reich, P. B., Poorter, L., Wrigh,t I. J., Ray, P., Enrico, L., Pausa,s J. G., de Vos, A. C.,
Buchmann, N., Funes, G., Quétier, F., Hodgson, J. G., Thompson, K., Morgan, H. D., ter
Steege, H., van der Heijden, M. G. A., Sack, L., Blonder, B., Poschlod, P., Vaieretti, M. V.,
Conti, G., Staver, A. C., Aquino, S. and Cornelissen J. H. C. 2013 New handbook for
standardized measurement of plant functional traits worldwide. Australian Journal of
Botany 61:167-234.
Peri, P. L., Ladd, B., Pepper, D. A., Bonser, S. P., Laffan, S. W. and Amelungi, W. 2012. Carbon
(δ13
C) and nitrogen (δ15
N) stable isotope composition in plant and soil in Southern
Patagonia’s native forests. Global Change Biology 18:311-321.
Poorter, L. and Bongers, F. 2006. Leaf traits are good predictors of plant performance across 53
rain forest species. Ecology 87(7):1733-1743.
Poorter, L., Bongers, F., Sterck, F. J. and Hannsjorg, W. 2003. Architecture of 53 rainforest trees
differing in adult stature and shade tolerance. Ecology 84:602-608.

67
Poorter, L., Wright, S. J., Paz, H., Ackerly, D. D., Condit, R., Ibarra-Manríquez, G., Harms, K.
E., Licona, J. C., Martínez-Ramos, M., Mazer, S. J., Muller-Landau, H. C., Peña-Claros,
M., Webb, C. O. and Wright, I. J. 2008. Are functional traits good predictors of
demographic rates? Evidence from five Neotropical forests. Ecology 89 (7):1908-1920.
Reich, P. B. and Oleksyn, J. 2004. Global patterns of plant leaf N and P in relation to
temperature and latitude. Proceedings of the National Academy of Sciences 101:11001-
11006.
Reich, P. B., Ellsworth, D. S., Walters, M. B., Vose, J. M., Gresham, C., Volin, J. C. and
Bowman, W. D. 1999. Generality of leaf trait relationships: a test across six biomes.
Ecology 80: 1955-1969.
Reich, P. B., Walters, M. B. and Ellsworth, D. S. 1992. Leaf life-span in relation to leaf, plant,
and stand characteristics among diverse ecosystems. Ecological Monographs 62: 365-392.
Reich, P. B., Wright, I. J. and Lusk, C. H. 2007. Predicting leaf physiology from simple plant
and climate attributes: a global GLOPNET analysis. Ecological Applications 17:1982-
1988.
Reich, P. B., Wright, I. J., Cavender Bares, J., Craine, J. M., Oleksyn, J., Westoby, M. and
Walters., M. B. 2003. The evolution of plant functionnal variation: traits, spectra, and
strategies. International Journal of Plant Science 164 (3): S143-S164.
Richards, R.A., 2005. Physiological traits used in the breeding of new cultivars for water scarce
environment. Agricultural Water Management 80:197-211.
Roche, P., Diaz-Burlinson, N. and Gachet, S. 2004. Congruency analysis of species ranking
based on leaf traits: which traits are more reliable? Plant Ecology 174: 37-48.
Roscher, C., Schumacher, J., Schmid, B. and Schulze, E. D. 2015. Contrasting effects of
intraspecific trait variation on trait-based niches and performance of legumes in plant
mixtures. PLoS ONE 10(3): e0119786. doi:10.1371/journal. pone.0119786.
Ryan, J., Estefan, G. and Rashid, A. 2001. Soil and plant analysis laboratory manual, 2nd
edition, International Center for Agricultural Research in the Dry Areas (ICARDA),
Aleppo, Syria, pp. 46-48.
Ryser, P. and Eek, L. 2000. Consequences of phenotypic plasticity vs interspecific differences in
leaf and root traits for acquisition of aboveground and belowground resources. American
Journal of Botany 87: 402-411.
Semenova, G. V. and van der Maarel, E. 2000. Plant functional types: A strategic perspective.
Journal of Vegetation Science 11(6): 917-922.
Siefert, A. 2012. Incorporating intraspecific variation in tests of trait-based community assembly.
Oecologia 170: 767-775.
Silvertown, J. 2004. Plant coexistence and the niche. Trends in Ecology and Evolution 19: 605-
611.

68
Sugiura, D. and Tateno, M. 2011. Optimal leaf-to-root ratio and leaf nitrogen content determined
by light and nitrogen availabilities. PLoS ONE 6(7): e22236.
doi:10.1371/journal.pone.0022236.
Sumida, A., Miyaura, T. and Torri, H. 2013. Relationships of tree height and diameter at breast
height revisited: analyses of stem growth using 20-year data of an even-aged
Chamaecyparis obtusa stand. Tree Physiology 33(1):106-118.
Swaine, M. D. and Whitmore, T. C. 1988. On the definition of ecological species groups in
tropical rain forests. Vegetatio 75:81-86.
Swanson, M. E., Franklin, J. F., Beschta, R. L., Crisafulli. C. M, DellaSala, D. A., Hutto, R. L.,
Lindenmayer, D. B. and Swanson, F. J. 2011. The forgotten stage of forest succession:
early-successional ecosystems on forest sites. Frontiers in Ecology and the Environment
9:117-125.
Swenson, N. G. and Enquist, B. J. 2007. Ecological and evolutionary determinants of a key plant
functional trait: Wood density and its community wide variation across latitude and
elevation. American Journal of Botany 94(3):451-459.
Thomas, S. C. 1996. Asymptotic height as a predictor of growth and allometric characteristics
Malaysian rain forest trees. American Journal of Botany 83:556- 566.
Thomas, S. C. and Malczewski, G. 2007. Wood carbon content of tree species in eastern China:
interspecific variability and the importance of the volatile fraction. Journal of
Environmental Management 85:659-662.
van der Werf, G. R., Morton, D. C., DeFries, R. S., Olivier, J. G. J., Kasibhatla, P. S., Jackson,
R. B., Collatz, G. J. and Randerson., J. T. 2009. CO2 emissions from forest loss. Nature
Geoscience 2:737-738.
van Geffen, K. G., Poorter, L., Sass-Klaasssen, U., van Logtestijn, R. S. P. and Cornelissen, H.
C. 2010. The trait contribution to wood decomposition rates of 15 neotropical tree species.
Ecology 91(12):3686-3697.
van Gelder, H. A., Poorter, L. and Sterck, F. J. 2006. Wood mechanics, allometry, and life-
history variation in a tropical rain forest community. New Phytologist 171:367-78.
Vendramini, F., Diaz, S., Gurvich, D. E., Wilson, P. J., Thompson, K. and Hodgson, J. G. 2002.
Leaf traits as indicators of resource-use strategy in floras with succulent species; New
Phytologist 154:147-157.
Verbeeck, H., Betehndoh, E., Maes, W. H., Hubau, W., Kearsley, E., Buggenhout, L., Hufkens,
K., Huygens, D., Van Acker, J., Beeckman, H., Mweru J. P. M., Boeckx, P. and Steppe, K.
2014. Functional leaf trait diversity of 10 tree species in Congolese secondary tropical
forest. Journal of Tropical Forest Science 26(3): 409-419.
Violle, C., Enquist, B. J.,McGill, B. J., Jiang, L., Albert, C. H., Hulshof, C., Jung, V. and
Messier, J. 2012. The return of the variance: intraspecific variability in community
ecology. Trends in Ecology and Evolution 27:244-252.

69
Violle, C., Navas, M. L., Vile, D., Kazafou, E., Fortunel, C., Hummel, I. and Garnier, E. 2007.
Let the concept of trait be functional! Oikos 116:882-892.
Weiher, E. and Keddy, P. A. 1995. Assembly rules, null models, and trait dispersion - New
questions from old patterns. Oikos 74:159-164.
Weiher, E., van der Werf, A., Thompson, K., Roderick, M., Garnier, E. and Eriksson, O.1999.
Challenging Theophrastus: A common core list of plant traits for functional ecology.
Journal of Vegetation Science 10: 609-620.
Welden, C. W., Hewett, S. W., Hubbell, S. P. and Foster, R. B. 1991. Sapling survival, growth,
and recruitment - relationship to canopy height in a neotropical forest. Ecology 72:35-50.
Westoby, M. 1998. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil 199:
213-227.
Westoby, M., Falster, D. S., Moles, A. T., Vesk, P. A. and Wright, I. J. 2002. Plant ecological
strategies: some leading dimensions of variation between species. Annual Review of
Ecology and Systematics 33:125-159.
Westoby, M., Reich, P. B. and Wright, I. J. 2013. Understanding ecological variation across
species: area-based vs mass-based expression of leaf traits. New Phytologist 199:322-323.
Wilson, P. J., Thompson, K. and Hodgson, J. G. 1999. Specific leaf area and leaf dry matter
content as alternative predictors of plant strategies. New Phytologist 143: 155-162.
Woodward, F. I., Lomas, M. R., Kelly, C. K. 2004. Global climate and the distribution of plant
biomes. Philosophical Transactions of the Royal Society B 359:1465-1476.
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z. Bongers, F., Cavender-
Bares, J., Chapin, T., Cornelissen, J. H., Diemer, M., Flexas, J., Garnier, E., Groom, P. K.,
Gulias, J., Hikosaka, K., Lamont, B. B., Lee, T., Lee, W., Lusk, C., Midgley, J. J., Navas,
M. L., Niinemets, U., Oleksyn, J., Osada, N., Poorter, H., Poot, P., Prior, L., Pyankov, V. I.,
Roumet, C., Thomas, S. C., Tjoelker, M. G., Veneklaas, E. J., Villar, R. 2004. The
worldwide leaf economics spectrum. Nature 428:821-827.
Wright, S. J., Kitajima, K., Kraft, N. J. B., Reich, P. B., Wright, I. J., Bunker, D. E., Condit, R.,
Dalling, J. W., Davies, S. J., Díaz, S., Engelbrecht, B.M., Harms, K. E, Hubbell, S. P.,
Marks, C. O., Ruiz-Jaen, M. C., Salvador, C. M. and Zanne, A. E. 2010. Functional traits
and the growth-mortality trade-off in tropical trees. Ecology 91(12):3664-3674.
Ye, X. H., Pan, X., Cornwell, W. K., Gao, S. Q., Dong, M. and Cornelissen, J. H. C 2015.
Divergence of above- and belowground C and N pool within predominant plant species
along two precipitation gradients in North China. Biogeosciences 12: 457-465.





























