Embed Size (px)
Citation preview

Aquaculture 147 ( 1996) 2 I-35
Integrated cultivation of the red alga Kappu@zycus alvarezii and the pearl oyster Pinctada martensi
Pei-Yuan Qian a* * , C. Y. Wu b, Madeline Wu a, Y. K. Xie ’
a Department of Biology, The Hong Kong University of Science and Technology, Clear Water Bay, Hong Kong b Institute of Oceanology. Chinese Academy of Science, Qingdao , China
’ South China Sea Institute of Oceanology, Chinese Academy of Science, Guangzhaw , China
Accepted 20 June 1996
Abstract
The effect of simultaneously cultivating the pearl oyster Pincfada martensi and the red alga Ku~pup~ycus alvarezii on growth rates of both species was investigated in laboratory and field studies conducted from December 1993 to June 1995. The two study sites were in subtidal areas 100 km apart off the east coast of Hainan Wand, China. Pearl oysters were cultivated in the center of an algal farm and red alga was cultivated in the center of the pearl oyster farm. These field experiments showed higher growth rates of both P. martensi and K. alvarezii in a co-culture system than in a monospecies culture system. Laboratory studies showed that the algae removed nitrogenous wastes released by pearl oysters. Algae treated with pearl oyster wastes grew much faster than those without oyster wastes. Algae treated with the seawater to which NH&l, NaNO, and NaNO, were added grew at the same rate as those treated with natural seawater containing oyster nitrogenous wastes, suggesting that enhanced growth of algae in the co-culture system was largely due to nitrogenous metabolites of the pearl oysters. In the co-culture, growth of pearl oysters was positively influenced by the presence of rapidly growing algae but when seawater temperature decreased below 2O”C, the algae grew slowly and there was no measurable benefit of mixed culture to either algae or pearl oyster.
Keywords: Pearl oyster Pinctada martensi; Red alga Kappaphycus alvarezii; Microenvironment; Ecological farming; Nitrogen release and uptake
’ Corresponding author. Tel: 852-2358-7331; Fax: 852-2358-1559, E-mail: [email protected]
00448486/%/$15.00 Copyright 0 1996 Elsevier Science B.V. All rights reserved. PII SOO44-8486(96)01393-2

22 P.-Y. Qian et al./Aquaculture 147 (1996) 21-35
1. Introduction
Marine pollution and environmental degradation are by-products of industrialization and intensification of marine aquaculture in Asia. The noticeable decline in water quality affects public health in coastal regions and directly threatens development of the mariculture industry. In addition, massive monospecies mariculture systems often cause the degradation of species quality and result in a progressive decline in species resistance to pathogenic infections. For example, the recent collapse of the prawn industry in China and Taiwan has been attributed to outbreaks of pathogenic bacteria (Liao, 1992).
The combined culture of marine algae and animals has recently been tested in China and Taiwan. This system is based on the concept that algae actively uptake CO,, release 0, to the surrounding environment, and utilize the metabolic wastes from the maricul- ture animals as nutrients. As a result, the algae can create a favorable environment for animal growth (Shari and Wang, 1985). Several studies have reported (Chiang, 1980; Chang and Wang, 1985; Lou and Wei, 1986; Tian et al., 1987; Chunhan, 1989; Trono, 1989; Wei, 1990) enhanced growth rate of seaweed and animals (shrimp, mussel and crabs) in integrated culture.
In this study, we chose Kappuphycus alvarezii (Doty) and Pincfudu murtensi (Dumker) as our experimental materials because of their availability and economic potential. K. ulvurezii is an important source of carrageenan (Li, 1990) and is harvested from sea or grown in many mariculture farms in South-East Asia (Doty, 1973; Doty and Alvarez, 1973). This species was introduced to Hainan Island, China in 1980’s (Wu et al., 1989) and currently supports a substantial local carrageenan industry. P. martensi is the key pearl farming species of Hainan. Large scale pearl oyster farms are located in semi-enclosed bay areas along both the east and west coasts of Hainan province. Our hypothesis was that metabolites of seaweed and shellfish are mutually beneficial. We examined the effects of mixed culture of K. ulvurezii and P. murtensi on the growth rates of both species and the effects of pearl oyster nitrogenous wastes on the growth of seaweed in field and laboratory experiments.
2. Materials and methods
2.1. Species
K. ulvurezii (Doty) was obtained from an alga farm in Lian Bay of Hainan province (where we cultivated the experimental pearl oyster in the middle of the farm) prior to each experiment below. Juveniles of P. murtensi (Dumker) with average size of about 2.5 cm in shell length were obtained from a commercial pearl oyster farm in Luhuitou, Sanya City, Hainan province.
2.2. Co-culture conditions in the open sea
The experiments began in the 10 m deep (at high tide) water in front of Luhuitou Tropical Marine Station (Fig. 1) in October 1993 and ended in June 1994 due to the

P.-Y. Qian et al./Aquaculture 147 (1996) 21-35 23
I
Fig. 1. Map of the experimental sites at Luhiutou and Liau Bay on Hainan Island, China. The Luhiutou Tropical Marine Station is marked as Luhiutou on the map. Field studies were conducted in open water in front of the station. In Lian Bay, field studies were conducted at the opening of the bay as indicated by the dot. The two sites were about 100 km apart.
Typhoon season. We investigated the potential benefit of multi-species mixed culture system on growth of both K. alvarezii and P. martensi. For this experiment, eighteen cylindrical experimental cages of 50 cm in diameter and 40 cm in length were constructed with stainless steel wire and nylon mesh. All cages were divided into three groups. One group of six cages contained only pearl oysters (20 individuals/cage). Pearl oysters were randomly selected, tagged, and assigned to the cages. Prior to the experiment, height, width, and length of each oyster were measured with a calliper and wet weight was determined to 0.1 mg. A second group of six cages each only contained 200 grams of algae (divided into four 50 gram clumps which were then suspended perpendicularly in the cages). The third group of six cages received both oysters and algae (20 pearl oysters and 200 grams of algae). All the cages were suspended at 2 m depth from bamboo rafts, the different groups being set at 25 m intervals. Cages were checked and brushed by hand twice a week during the experimental period. Growth of pearl oysters and algae were monitored by direct measurements of wet weight (algae and oyster) and height, width and length (pearl oyster only) every 2-3 weeks. The daily growth rates of algae were determined by the following equation:
%R = [ ( lnWj - hWi)/T] X 100%
Where Wi is the initial wet weight, Wj is the wet weight on day j, T is the interval between day i and day j, and R is the daily growth rate of algae.

24 P.-Y. Qian et al./Aquaculture 147 (19%) 21-35
2.3. Tagging and maintenance
For tagging pearl oysters, the outer shell surface of each selected individual was cleaned with a test-tube brush to remove debris or fouling organisms on the shell. The flat comer of the shell was dried and a thin layer of Krazy Glue was added to the dried area. A small plastic sheet with a given number was placed on top of the glue and pressed gently against the shell. Thus, each pearl oyster was individually tagged. Over 95% of tagged individuals survived the nine month experimental period.
2.4. Cultivation of algae in the oyster farm
In a separate experiment, growth of algae was compared within and outside of an oyster farm. Ten cages containing algae K. alvarezii (200 g per cage) were suspended at 2 m depth from the rafts in the center of the Luhuitou pearl P. martensi farm (treatment groups) while another ten cages of algae were suspended at 2 m depth in the water 20 m outside of the farm (control groups). Growth of algae in these cages was determined by monitoring the net increase in algae wet weight every 2-3 weeks. The daily growth rate was determined according to the equation mentioned above.
2.5. Cultivation of pearl oyster in the algae farm
In a third experiment, growth of pearl oysters within and outside of a K. alvarezii farm were compared. Two hundred and forty pearl oysters with an initial length of approximately 2.5 cm were randomly selected, tagged, measured, and divided into twelve cages. Six cages of pearl oysters (20 oysters/cage) were suspended in the middle of the farm at 2 m depth in Lian Bay of Hainan province, while another six cages of pearl oysters were suspended 2 km away in the same bay where no algal farm exists. Growth of pearl oysters in and outside the algal farm was monitored every month.
2.6. Efsects of nitrogen release of the pearl oyster on nitrogen uptake and growth of algae
Fertilization of algae by oyster metabolites (including nitrogenous wastes) is a potential mechanism for increasing the growth of algae. Experiments were therefore performed to study the effects of nitrogenous waste from pearl oysters on growth of algae.
In a laboratory experiment at Luhuitou Tropical Marine Station, 80 randomly selected pearl oysters P. martensi of a size range of 3-4 cm in shell length were each placed in a glass container with 20 1 natural seawater. Concentrations of NH:, NO; and NO; in the seawater were determined according to Gao (1980). After 6 h, the oysters were removed and concentrations of NH:, NO; and NO; in the seawater were determined again. The amount of NH:, NO; and NO; re eased by the oysters was estimated by 1 subtracting the initial concentrations from the final concentrations. A 50 g wet weight of K. alvarezii was placed in the pearl oyster treated seawater for one hour. After removal of the algae from the container, the concentration of NH:, NO; and NO; in the

P.-Y. Qian et al./Aquaculture I47 (1996121-35 25
seawater were determined again. Amounts of NH:, NO; and NO; uptaken by the algae were estimated by subtracting the amount of post-algal uptake from that of pre-algal uptake. The algae treated with oyster nitrogenous waste were then placed in cages and suspended in the field to monitor their growth. The treatment was repeated on day 4,7, 10 and 13. Growth of algae was estimated by wet weight measurement on days 7 and 13 (total growth for each period of 6 days). The daily growth rate was determined according to the equation mentioned above. This experiment was started on April 8, 1995 and completed on April 20, 1995. A similar experimental series was conducted from April 10-22, 1995. In this second series one more group was added. In the additional group, the concentrations of NH:, NO; and NO; in the seawater released by oysters were simulated by addition to seawater of NH&l, NaNO,, and NaNO,. Uptakes of NH:, NO; and NO; by algae and algal growth rates were determined as in the first series.
2.7. Data analysis
Unpaired t-tests were used to compare the growth rates of: 1) algae within and outside of the pearl oyster farm; 2) algae alone in cages with algae in cages containing oysters; 3) pearl oysters within and outside of the algae farm and 4) pearl oysters alone in cages with pearl oysters in cages containing algae. To investigate the effects of pearl oyster nitrogenous waste release on the nitrogenous waste uptake and growth of algae, we used two-way ANOVAs to determine the effect of treatment of pearl oyster metabolites and different experimental days on the release rate of NH:, NO; and NO; by oysters and their uptake rate by algae. Since the analyses indicated no significant interaction between experiments and times or experimental treatment effects, the data from Experiments 1 and 2 were pooled for the further analysis. To determine the relationship between the release rate of NH:, NO; and NO; from oysters and the uptake rate by the algae, non-parametric correlation coefficient analysis (due to small sample size) was conducted for each of NH:, NO; and NO; and total nitrogen. One-way ANOVAs and Sheffe multiple comparison tests were used to determine the effects of time on the release or uptake of NH:, NO; and NO; and to determine the effects of treatments on growth rate of algae.
3. Results
3.1. Effect of co-culture on growth of P. martensi and K. alvarezii
Growth of K. alvarezii in the experimental cages suspended in the subtidal region in front of Luhuitou Tropical Research Station, Sanya city is summarized in Fig. 2. It should be pointed out that in November and December 1993 when the seawater temperature decreased below 2O”C, the algae in most cages were rotten and therefore no data were included in this figure. In January 1994 when monthly averaged seawater temperature rose to 22S”C, the algae in the cages with pearl oysters (treatment) showed positive but slower growth than those in the cages without pearl oysters (control) (Fig.

26 P.-Y. Qian et al./Aquaculture 147 (1996) 21-35
6
S
4
3
2
1
0 Jan. Feb. Mar. Apr.
Month
Fig. 2. Growth rate of red algae Kappaphycus aluarezii (% increase in wet weight per day) in caged experiments conducted in the subtidal area in front of Luhuitou Tropic Research Station, Sanya. All values are mean f s.e. for 24 algal samples. Significant differences (at 0.05 level) behveen the treatment group (pearl oyster Pinctada murtensi co-cultivated with K. alvarezid and the control group (K. aluarezii cultivated alone) are indicated by * on top of each paired bars. For all groups, the initial amount of K. aluarezii was 50 g. Averaged seawater temperatures at the study site for January, February, March, and April 1994 were 22.5, 23.5, 24.6, and 27.1”C, respectively (data provided by Sanya City observatory).
2). However, in February, March and April 1994 when seawater temperature rose to 23S”C, 24.6”C and 27.1”C respectively, algae in treatment groups grew much faster than those in control groups. On average, algae grew at least 20% more in the treatment
1 --C- Pearl oyster alone
I -O- Pearl oyster & alga
SO-
4.0-
3.0-1 m Dec. Jan. Feb. Mar. Apr.
Month
Fig. 3. Growth of pearl oysters Pinctada martensi (change in shell length) in caged experiments conducted in the subtidal area in front of Luhuitou Tropical Research Station, Sanya. All values are mean f s.d. of 118 tagged pearl oysters.

P.-Y, Qian et al./Aquacubure 147 (1996) 21-35 27
group than those in the control. In April, the increase in wet weight of algae in cages containing oysters was up to 45% greater than algae in cages without oysters, but on average was 23 + 5% greater. The low values recorded here for March was largely due to heavy grazing by fish on the algae in cages. The actual growth rate shall be much higher.
Fig. 3 shows growth of P. martensi in terms of changes in shell length when enclosed with or without K. nluarezii. Pearl oysters kept with K. aluarezii grew faster than their siblings kept in the cages without algae. The presence of red algae in culture cages enhanced the growth of pearl oysters by 12.5 +_ 3.0%.
3.2. Growth of K. alvarezii in the pearl oyster farming area
Growth of red algae K. aluarezii is summarized in Fig. 4. For each month from March to June 1994 (seawater temperatures ranged from 24.6 to 29XC, see figure legend), the growth of K. aluarezii suspended in the center of the pearl oyster farm was significantly higher than growth of algae outside the pearl oyster farm (unpaired t-test: t = 3.2, p < 0.05). Pearl oysters enhanced the growth of this red algae by 19.2 -t 2.9%. Again, the low values recorded here for March were largely due to heavy grazing by fish on the algae. The actual growth rate shall be much higher than the values in Fig. 4.
Mar. Apr.
Month
Cl Inside farm
q Outside farm
May Jun.
Fig. 4. Growth rate of red algae Kappaphycus alvarezii (% increase in wet weight per day) in caged
experiments conducted within and outside a pearl oyster farm in front of Luhuitou Tropical Research Station,
Sanya. All values are mean-C s.e. for twenty algal samples. Significant differences (at 0.05 level) between the
treatment group (cages suspended in the center of pearl oyster farm) and the control group (cages suspended outside pearl oyster farm) are indicated by * on top of the paired bars and were detected by unpaired f-tests.
For all groups, the initial amount of K. alvarezii was 50 g. Averaged seawater temperature at the study site for
March, April, May, and June 1994 were 24.6, 27.1, 29.6, and 29.8”C. respectively (data provided by Sanya
City observatory).

28 P.-Y. Qian et al./Aquaculture 147 (1996) 21-35
3.3. Growth of P. martensi in the algae farming area
In the first three months of the experiment (November 1993 to January 1994) pearl oysters suspended in the center of the algal farm grew more slowly than those suspended outside the algae farm (Fig. 5). However, in the following three months, pearl oyster suspended in the center of the algal farm grew faster than those outside the algal farm. This growth pattern corresponded to the growth pattern of the red algae.
3.4. Relationship between the nitrogen release rate of pearl oysters and the nitrogen uptake rate of algae
Table 1 summarizes the concentrations of NH:, NO; and NO; and total nitrogen released in 6 h by 80 P. martensi and taken up by K. alvarezii (200 g) in one hour on five different occasions. Concentrations differed considerably, but the ratios of NH:, NO; and NO; released by pearl oysters remained consistent over the different sampling dates as follows: NH::NO;:NO;= 25:20:1. Amounts of NH:, NO; and NO; taken up by algae followed a similar trend. The ratio of NH:, NO; and NO; uptake was NH::NO;:NO; = 2615: 1, slightly different from the ratio released by oysters. Nonparametric correlation coefficient analysis revealed significant correlations between the amount of release by oysters and the amount taken up by the algae for NH:, NO; and NO;, and for total nitrogen (p < 0.05 for all tests).
Table 2 summarizes the NH:, NO; and NO;, and total nitrogen uptake by algae from natural seawater and from the seawater to which NH:, NO; and NO; were
60- -o- In the center of algal farm
2 -O- Outside algal farm
* *
g 50- 8 z % z t 40. a B a ‘b 5
30‘
z s
20> ’ Nov. Dec. Jan. Feb. Mar. Apr.
Month
Fig. 5. Growth of pearl oyster Pinctada martensi (changes in shell length) in caged experiments conducted in
the algae farm at Lian site. All values are mean f s.d. of 120 tagged pearl oysters. Significant differences (at
0.05 level) between the treatment group (pearl oysters cultivated in the center of Kappuphycw aluarezii farm)
and the control group (pearl oysters cultivated outside K. aluarezii farm) are indicated by * on top of the
paired data points.
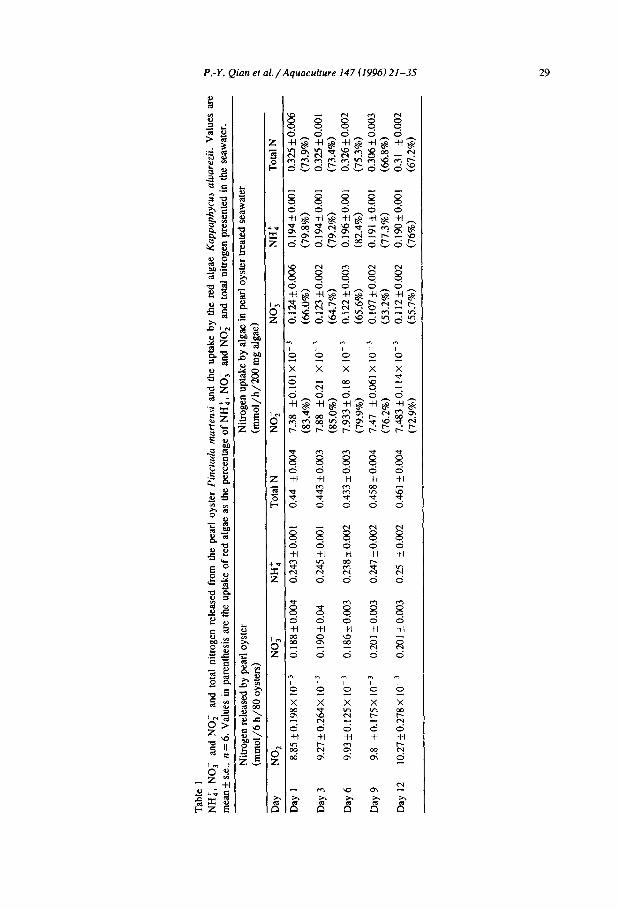
Tab
le I
N
H;,
NO
; an
d N
O;
and
tota
l ni
trog
en r
elea
sed
from
the
pea
rl o
yste
r P
incr
uda
mar
tens
i an
d th
e up
take
by
the
red
alga
e K
appu
phyc
us
ulua
rezi
i. V
alue
s ar
e T
m
ean
f se
., n
= 6
. V
alue
s in
par
enth
esis
are
the
upt
ake
of r
ed a
lgae
as
the
perc
enta
ge o
f N
H:,
NO
; an
d N
O;
and
tota
l ni
trog
en p
rese
nted
in
the
seaw
ater
. .Y
Nitr
ogen
rel
ease
d by
pea
rl o
yste
r a
Nitr
ogen
upt
ake
by a
lgae
in
pear
l oy
ster
tre
ated
sea
wat
er
f:
(mm
ol/d
h/
80
oyst
ers)
(m
mol
/h/2
00
mg
alga
e)
5
Day
N
O;
NO
; N
H:
Tot
al N
N
O;
NO
; N
H;
Tot
al N
K
\
Day
I
8.85
+0.
198X
IO
-’
0. I8
8 f
0.00
4 0.
243
f 0.
00 I
0.44
+
0.00
4 7.
38
*O.lO
l X
lO-3
0.
124~
0.00
6 0.
194*
0.00
1 0.
325
f 0.
006
B E
(83.
4%)
(66.
0%)
(79.
8%)
(73.
9%)
a D
ay 3
9.
27*0
.264
X
IO-’
0.
190*
0.04
0.
245
f 0.
001
0.44
3 *
0.00
3 7.
88
kO.2
1 X
IO
-’
0.12
3 f
0.00
2 0.
194*
0.00
1 0.
325
f 0.
001
5
(85.
0%)
(64.
7%)
(79.
2%)
(73.
4%)
S Z
Day
6
9.93
&0.
125X
lO
-3
0.18
6 f
0.00
3 0.
238
f 0.
002
0.43
3 f
0.00
3 7.
933f
0.18
X
lo-’
0.
122
f 0.
003
0.19
6rtO
.001
0.
326
f 0.
002
3 (7
9.9%
) (6
5.6%
) (8
2.4%
) (7
5.3%
) 9.
8 &
0.17
5X 1
O-3
2
Day
9
0.20
1 f
0.00
3 0.
247
k 0.
002
0.45
8 *
0.00
4 7.
47
*0.0
61 x
10-
s 0.
107f
0.00
2 0.
191*
0.00
1 0.
306
f0.0
03
g (7
6.2%
) (5
3.2%
) (7
7.3%
) (6
6.8%
) Z
!
Day
12
10.2
7&0.
278X
1O
-3
0.20
1 f
0.00
3 0.
25
~0.0
02
0.46
1 f
0.00
4 7.
483*
0.11
4X
lO-3
0.
112*
0.00
2 0.
190f
0.00
1 0.
31
f0.0
02
7 (7
2.9%
) (5
5.7%
) (7
6%)
(67.
2%)
2

Upt
ake
of N
H:,
NO
; an
d N
O;
and
tota
l nitr
ogen
by
the
red
alga
e K
uppu
phyc
us a
fuar
etii
. In
the
las
t fo
ur c
olum
us,
the
valu
es i
n pa
rent
hesi
s ar
e th
e co
ncen
trat
ions
z
in t
he s
eaw
ater
pri
or t
o al
gal
upta
ke, w
hich
wer
e ob
tain
ed b
y ad
ding
sal
ts o
f N
H&
I,
NaN
O,
and
NaN
O,
to t
he n
atur
al s
eaw
ater
. T
hese
sim
ulat
ed c
once
ntra
tions
of
NH
:, N
O;
and
NO
; an
d to
tal
nitr
ogen
wer
e si
mila
r to
tho
se r
elea
sed
by t
he p
earl
oys
ters
in
Tab
le
I. T
he c
once
ntra
tions
of
NH
:, N
O;
and
NO
; in
nat
ural
B
2
seaw
ater
wer
e 0.
4-0.
7,
2.3-
2.7
and
0.04
-O
.+
mm
ol/m
3.
resp
ectiv
ely.
g
Nitr
ogen
upt
ake
by a
lgae
fro
m n
atur
al s
ea w
ater
\
Nitr
ogen
upt
ake
by a
lgae
in
sim
ulat
ed n
itrog
en c
once
ntra
tion
of s
ea w
ater
(m
mol
/hr/
200
g al
gae)
(m
mol
/hr/
200
g al
gae)
2 6
Day
N
O;
NO
; N
H:
Tot
al N
N
O;
NO
; N
H:
Tot
al N
P S
Day
1
0.6x
IO
- 3
0.02
4 0.
010
0.03
5 7.
l(
8.8)
X 1
0-s
0.12
3(0.
188)
0.
I %(0
.244
) 0.
326(
0.44
1)
2
Day
3
0.7x
10
-l
0.02
6 0.
007
0.03
4 7.
7( 8
.9)x
IO
- 3
0.13
1(0.
185)
0.
193(
0.24
4)
0.33
2tO
.438
) S
Day
6
0.7x
10
-s
0.02
4 0.
009
0.03
4 8.
1(10
.0)X
lO
-3
0.11
4(0.
179)
0.
193(
0.24
4)
0.3
15cO
.427
) 2
Day
9
0.9
x 10
-S
0.02
7 0.
008
0.04
4 7.
l(
9.6)
x 10
-3.
0.11
1(0.
199)
0.
191(
0.23
8)
0.30
9(0.
457)
%
Day
I2
0.6X
lO
-3
0.03
0 0.
009
0.04
0 7.
6(10
.4)x
10
-s
O~l
l3(0
.198
) 0.
193(
0.24
7)
0.3
I4(0
.455
) 2 !z
I 2

P.-Y. Qian et al./Aquaculture 147 (1996) 21-35 31
added. The concentrations of NH:, NO; and NO; in natural seawater were very low (0.4-0.7, 2.3-2.7 and 0.04-0.06 mmol/m3 for NH:, NO; and NO, respectively), as were uptake rates by the algae. Correlation analyses showed a concentration dependence of uptake rate by algae. Levels of NH:, NO, and NO; and total nitrogen in seawater with artificially enhanced concentration were similar to those resulting from release by the oyster natural metabolite treatment (Table 2). NH:, NO; and NO; and total nitrogen uptake by algae in the seawater with a chemical source of nitrogen was also similar to that in the seawater containing oyster metabolites. The uptake ratio was NH::NO;:NO; = 26: 16: 1. There was also a significant correlation between uptake and concentration ( p < 0.05).
One-way ANOVA: F=11.17, PcO.001 Day 1 to Day 6
61
D One-way ANOVA: Fz41.3, P-eO.0001 E ._ 61 Day 7 to Day 12
waste + nitrogen
Treatment groups
Fig. 6. Effects of NH:, NO; and NO; a dd’t’ I 10” on the growth rate of Kappaphycus ulvarezii. Plotted values
are mean + s.e. with sample size given at the top of each error bar. The “oyster nitrogenous wastes” treatment
consisted of natural seawater plus the nitrogenous wastes released by 80 pearl oysters Pinctada murtensi during 6 h; the “simulated seawater” treatment consisted of natural seawater with added NH,CI. NaNO, and
NaNO, to obtain concentrations of NH:, NO; and NO; similar to those released by pearl oysters;
“control” consisted of untreated natural seawater.
32 P.-Y. Qian et al./Aquaculture 147 (19%) 21-35
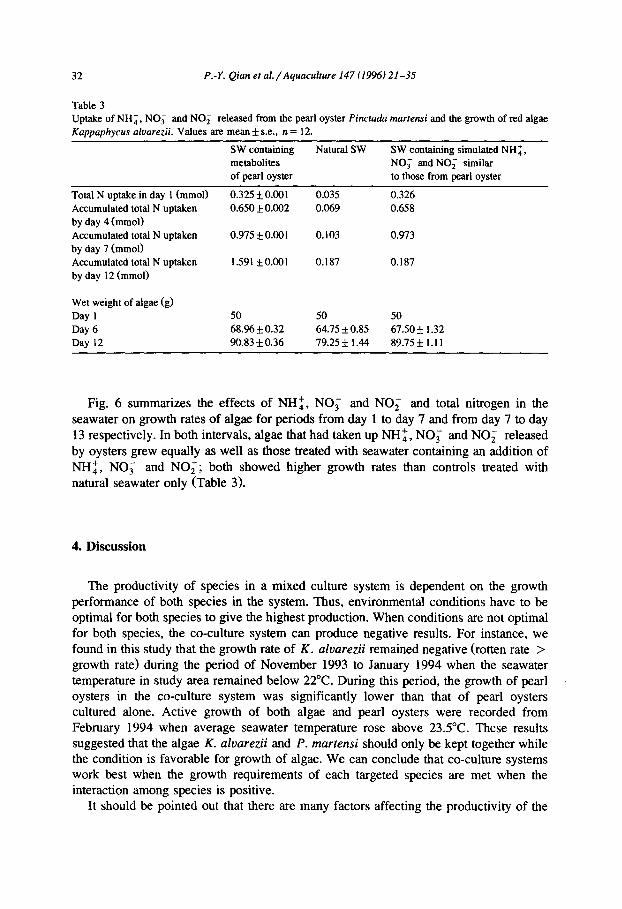
Table 3 Uptake of NH:, NO; and NO; released from the pearl oyster Pinctadu martensi and the growth of red algae Kappaphycus aluarezii. Values are mean f s.e.. n = 12.
Total N uptake in day 1 (mmol) Accumulated total N uptaken by day 4 (mmol) Accumulated total N uptaken by day 7 (mmol) Accumulated total N uptaken by day 12 (mmol)
SW containing metabolites of pearl oyster
0.325~0.001 0.650 & 0.002
0.975 f 0.001
1.591*0.001
Natural SW
0.035 0.069
0.103
0.187
SW containing simulated NH:, NO; and NO; similar to those from pearl oyster
0.326 0.658
0.973
0.187
Wet weight of algae (g) Day 1 Day 6 Day 12
50 50 50 68.% f 0.32 64.75 kO.85 67.50 f 1.32 90.83 kO.36 79.25 f 1.44 89.75f 1.11
Fig. 6 summarizes the effects of NH:, NO; and NO, and total nitrogen in the seawater on growth rates of algae for periods from day 1 to day 7 and from day 7 to day 13 respectively. In both intervals, algae that had taken up NH:, NO; and NO; released by oysters grew equally as well as those treated with seawater containing an addition of NH:, NO; and NO;; both showed higher growth rates than controls treated with natural seawater only (Table 3).
4. Discussion
The productivity of species in a mixed culture system is dependent on the growth performance of both species in the system. Thus, environmental conditions have to be optimal for both species to give the highest production. When conditions are not optimal for both species, the co-culture system can produce negative results. For instance, we found in this study that the growth rate of K. aluarezii remained negative (rotten rate > growth rate) during the period of November 1993 to January 1994 when the seawater temperature in study area remained below 22°C. During this period, the growth of pearl oysters in the co-culture system was significantly lower than that of pearl oysters cultured alone. Active growth of both algae and pearl oysters were recorded from February 1994 when average seawater temperature rose above 23.5”C. These results suggested that the algae K. aluarezii and P. martensi should only be kept together while the condition is favorable for growth of algae. We can conclude that co-culture systems work best when the growth requirements of each targeted species are met when the interaction among species is positive.
It should be pointed out that there are many factors affecting the productivity of the

P.-Y, Qian et al./Aquaculture 147 (19%) 21-35 33
red algae in our study sites although the temperature appears to be major limiting factor in algae growth. For instance, although very active growth of K. aluarezii was observed in our experimental cages in March and April, due to heavy grazing by small fish on the algae, the total biomasses obtained from the cages at the end of experiments were small, resulting in lower daily growth rate for March and April. Therefore, closer monitoring on algae growth is necessary for our future study.
In this study, we found that within one hour, 200 g of K. aluarezii took up the total amount of nitrogen released by 80 pearl oysters in 6 h. This information is important for determining the density of both pearl oysters and algae in a mixed culture system.
Results similar to ours have been reported in other co-culture farming systems. Chiang (1980) found higher growth rates of both Gruciluria spp. and prawn in co-culture. Higher growth rates were also reported for a mixed culture of Gruciluriu spp. and crab as well as Gruciluriu spp. and grass shrimps (Chiang, 1980). Chang (1987) reported higher growth rates of the bay scallop Argopecten irrudiuns and the kelp tuminuria juponicu in a rotated culture system than in a monospecies culture system. Trono (1989) found that the productivity of both Grucifuriu spp. and crabs were increased when they were cultivated in the same pond. Shan and Wang (1985) found that in a mixed culture system of prawn and algae Gruciluriu spp., the survival rate of prawn increased by 51.6% compared with monospecies culture, while the growth rate of both prawn and algae was almost doubled. All results support the idea that mixed culture of animals with seaweeds enhances productivity of both groups.
It is a generally accepted hypothesis that the seaweeds in co-culture systems incorporate the metabolite wastes of animals into algal biomass resulting in a high growth rate (Harlin et al., 1978). The animals in the system presumably benefit from improved water quality. For instance, Asare (1980) found that animal waste can be used as the nitrogen source for cultivated Gruciluriu tikvuhiue and Neougurdhiellu buileyi. Wei (1990) found that decomposed feces and/or feed residuals of shrimp, Penueus penicillutus, and of the crab, Scyllu serrutu, increased the nitrogen and phosphate concentrations in pond culture and in turn enhanced photosynthesis of Gruciluria tenuistzpitufu (Yin, 1987). It has also been demonstrated that growth and carrageenan content of K. uluurezii can be affected by ammonia concentration in seawater (Wu et al., 1984; Li, 1990).
In this study, correlation coefficient analyses showed a strong relationship between algal growth and uptake nitrogenous wastes within the tested ranges, proving that the algae remove nitrogenous wastes very efficiently when conditions (e.g. light, tempera- ture, turbidity) are suitable for a high growth of algae. Simulation of the concentrations of NH:, NO; and NO, in this experiment further confirmed that the pattern and amount of NH:, NO; and NO; removed by the seaweeds were similar to those released by oysters. Furthermore, seaweeds that took up NH:, NO; and NO; from either oyster-treated seawater or nitrogen-added seawater grew faster than those treated with natural seawater. More importantly, we found that K. ulvurezii removed NI-: more effectively than NO; and NO;. These results indicate that at our study site, nitrogenous compounds limit K. ulvurezii growth and the presence of the algae in the system improved seawater quality by reducing nitrogenous waste levels, resulting in a higher growth rate of pearl oysters.

34 P.-Y. Qian et al./Aquacultwe 147 11996) 21-35
5. Conclusion
Both pearl oyster P. mrrensi and red alga K. aluarezii in a co-culture system grew faster than in a monospecies culture system. Our laboratory experiments revealed that K. aluarezii removed nitrogenous wastes released by pearl oysters very efficiently and that algae treated with pearl oyster wastes grew much faster than those without oyster wastes. Algae treated with seawater with elevated concentrations of NH:, NO; and NO; through adding NH&l, NaNO, and NaNO, into natural seawater grew as well as those treated with seawater containing oyster nitrogenous wastes. These results suggest that enhanced growth of algae in the co-culture system is largely due to the nitrogenous metabolites of the pearl oysters. The red algae K. uluurezii can therefore be used as a nitrogenous waste remover in pearl oyster farming systems to improve the seawater quality and pearl oyster production. Since the number of sites truly favorable to aquaculture are limited, it is advantageous to find ways of increasing productivity within existing sites. The solution to conflicting demands between development of high-density mariculture and protection of a healthy marine environment may rely on co-culture and our understanding of the interactions between primary producers and consumers.
Acknowledgements
The authors wish to thank the director and staff of Shanya Tropical Marine Station, the South China Sea Institute of Oceanology, Academy of Science, China, for providing laboratory space as well as access to the sea in the front of the station. The authors also wish to thank Mrs. Y.C. Zhow, Z.G. Shen, G.C. Zhang for their hard work in helping with field works, Drs. Patrick Bryan, Louis Gosselin and Dan Rittschof for their comments on the previous version of the manuscript, and anonymous reviewers for providing helpful comments on the manuscript. This research has been supported by a research grant (No. 3927014) of the Natural Science Research Foundation of China and a research grant (R5069) from the Hainan Institute of the Hong Kong University of Science and Technology.
References
Asare, SO., 1980. Animal waste as a nitrogen source for Gracilaria tikuahiae and Neoagardhiella baileyi in culture. Aquaculture, 21: 87-91.
Chang, C.X. and Wang, N.C., 1985. Mixed culture system of shrimp and seaweeds. Mar. Sci., 9(3): 32-35 (In Chinese).
Chang, F.H., 1987. Preliminary study on rotated culture system of Argopecten irradians Lamarck and
Lmninaria japonica, Mar. Sci., 6: l-6 (In Chinese).
Chiang, Y.M., 1980. Cultivation of Gracilaria (Rhodophycophyta, Gigartinales) in Taiwan. In: Proc. Intl.
Seaweed Symp. 10, Walter de Grugter, Berlin, New York, pp. 569-574.
Chunhan, V.D., 1989. Seminar on Gracilaria production and utilization in the Bay of Bengal. Anti-bacterial properties. In: Fishing chimes, R. 20-00, pp. 43.
Doty, M.S., 1973. Farming tbe red seaweed, Akimbo, for carrageenans. Micronesica, 9(l): 59-73.
Doty, M.S. and Alvarez, V.B., 1973. Seaweed farms: a new approach for U.S. industry. Mar. Tech. Sot., 9th Aunu. Conf., pp. 701-708.

P.-Y. Qian et nl./Aquaculture 147 (1996) 21-35 35
Gao, F.M., 1980. Determination of nitrates in seawater by oxidation method. Transact. Gceanol. Limnol., 4: 41-45.
Harhn, M.M., Thome-Miller, B. and Thursby, G.B., 1978. Ammonium uptake by Gracilaria sp. (Floridophyceae) and Ulua Zactuca (Chlorophyceae) in closed system fish cuhure. Proc. Int. Seaweed Symp., 9: 285-292.
Li, R., 19%. Effect of ammonia on growth and carrageenan content in Kappaphycus alvarezi; (Gigarrinales. Rhodophyta). Hydrobiologia, 204: 499-503.
Lou, Y.Y. and Wei, S.Q., 1986. Experimental ecology study of crab ScyUa serruta (Forkal). East Sea Oceano., 4: 91-95 (In Chinese).
Liao, I.C., 1992. Aquaculture in Asia: status, constraints, strategies, and prospects. In: I.C. Liao, C.Z. Shyu and N.H. Chao (Editors), Aquaculture in Asia: Proceedings of the 1990 APO symposium in aquaculture, Taiwan Fisheries Research Institute, Taiwan, pp. 13-27.
Shan, Q.X. and Wang, L.C., 1985. Study of the mixed cuhure of algae and prawn. Mar. Sci., 9(3): 32-35. Tian, Z.P., Gao, F.M., Sun, S., Liu, S.Q., Zhang, Y.L. and Li, L.X., 1987. Effects of the interculture of
Mytilus edulis and Laminaria on the environment condition. Transact. Gceanol. Linmol., 2: 60-66. Trono, G., 1989. Management of natural resources of tropical agarophytes. In: Report of the seminar on
Gracilaria production and utilization in the Bay of Bengal region, BOBP/REP/45, pp. 92-96. Wei, S.Q., 1990. Study of mixed culture of Gracilaria tenuistipitata, Penaeus peniciilatus, and Scylla serrata.
Acta Ckeanol. Sinica, 12(3): 388-394. Wu, C.Y., Li, J.J., Xian, E.Z., Peng, Z.S., Tan, S.Z., Wen, L.J., Yang, Z.C., Huang, X.H., Cai. Z.L. and Chen,
G.J.. 1989. On the transplantation and cultivation of Kappaphycus aluarezii in China. Chin. J. Oceanol. Limnol., 7: 1-8.
Wu, C.Y., Zhang. Y.X., Li, R.Z., Peng, Z.S., Zhang, Y.F., Liu, WC, Zhang, J.P. and Fan, X., 1984. Utilization of ammonium-nitrogen by Porphyra yezoensis and Gracilaria uerrucosa. Hydrobiologia, 116/l 17: 475-477.
Yin, Z.Q., 1987. Mixed culture. of the prawn Penaeus motwdon and Gracilaria tenuistipitatu. Chin. Aquaculture, 6: 17.





























