Embed Size (px)
Citation preview

Institut für Nutzpflanzenwissenschaften und Ressourcenshutz der
Rheinischen Friedrich-Wilhelms-Universität Bonn
___________________________________________________________________________
Isolation, characterization and interactions of soil microorganisms involved in the enhanced biodegradation of non-fumigant organophosphate nematicides
Inaugural – Dissertation
Zur
Erlangung des Grades
Doktor der Agrarwissenschaften (Dr. agr.)
der Hohen Landwirtschaftlichen Fakultät
der
Rheinischen Friedrich-Wilhelms-Universität
zu Bonn
vorgelegt am 12. 06. 2009
von
José Alfonso Cabrera Motta
aus Guatemala

Diese Dissertation ist auf dem Hochschulschriftenserver der ULB Bonn http://hss.ulb.uni-bonn.de/diss_online elektronisch publiziert (2009). Referent: Prof. Dr. R.A. Sikora Korreferent: Prof. Dr. M. Becker Tag der mündliche Prüfung: 24.08.2009

I dedicate this work to my parents Elizabeth and Alfonso, to my sisters Vicky and
Mercedes, to their families, and to my life’s partner Véronique.

Isolation, characterization and interactions of soil microorganisms involved in the enhanced biodegradation of non-fumigant organophosphate nematicides
The most widely used pesticides utilized for the management of plant-parasitic nematodes belong to the organophosphorus group. Their efficacy may be reduced in areas where adapted microorganisms accumulate that are capable of rapidly degrading the active ingredients. The enhanced biodegradation process of non-fumigant nematicides is of particular concern in intensive agriculture. However, it remains unclear which microorganisms play the most important role in the rapid metabolization and how and why this process develops. Furthermore little is known as to whether the biodegradation process may be slowed down, stopped or reversed. Studies using soils with different nematicide history collected in four banana fields in the Atlantic region of Costa Rica demonstrated that the non-fumigant organophosphate nematicide terbufos had lower levels of efficacy and shorter effective activity against the burrowing nematode Radopholus similis when the soil had a prolonged terbufos application history. Lower levels of efficacy were related to the microorganisms capable of rapidly degrading the active ingredient. The analysis of soils collected in Germany with different nematicide application history demonstrated that fenamiphos, another organophosphate non-fumigant nematicide, was not rapidly biodegraded in soil with no previous pesticide exposure. This study also demonstrated that Pseudomonas spp. does not accumulate upon fenamiphos applications and may not be involved at all in fenamiphos degradation. The lack of surfactant production of the isolated Pseudomonas spp. could be a reason for their absence in the biodegradation process. Bacteria capable of degrading fenamiphos were isolated from another German soil with a large fenamiphos-history. These bacteria utilized fenamiphos as a sole carbon source. By comparison of the partial sequences of their 16S rRNA coding genes with those genes present in the GenBank sequence database, a fully resolved phylogenetic tree could be generated, showing that these fenamiphos degrading (Fd) isolates belonged to closely related Microbacterium, Sinorhizobium, Brevundimonas, Ralstonia, or Cupriavidus species. The Fd bacteria did not cross-degrade the novel organophosphate nematicide fosthiazate, thus suggesting that they are fenamiphos-specific. However, a combination of all microorganisms of the same soil from which the fenamiphos-degrading bacteria was isolated, was capable of degrading fosthiazate, thus demonstrating that there are other microorganisms capable of degrading nematicides even in the absence of an application history. This also revealed that the nematicide-history of one organophosphate nematicide does not intrinsically influence the degradation of another pesticide of this same chemical group. The application of plant revitalizers enhanced soil microbial biomass over time which resulted in an enhanced biocontrol activity against the root-knot nematode Meloidogyne incognita and a delayed biodegradation process of fenamiphos. In conclusion, this research demonstrated that many different soil bacteria can adapt when frequently exposed to a particular nematicide, thus offering them an alternative carbon source to grow. This effect can be slowed down by altering the microbial soil diversity through the application of natural plant enhancers that benefit nematicide non-degrading strains and simultaneously reduce nematode damage.

Isolierung, Charakterizierung und Wechselwirkungen von Bödenmikroorganismen verantworlich für den beschleunigten biologischen Abbau von nicht gas förmigen
organophosphatishce Nematiziden Die weitverbreitesten Pestizide gehören zur Wirkstoffgruppe der Organophosphate. Jedoch kann deren Wirkung durch das verstärkt Auftreten von Mikroorganismen, welche in der Lage sind diesen Wirkstoff zu degradieren, gemindert werden. Die verstärkte Degradierung von nicht gasförmigen Nematiziden betrifft vor allem Anbaugebiete mit intensiver Landwirtschaft. Bis heute ist ungeklärt welche Mikroorganismen bei dem Prozess der beschleunigten Metabolisierung von nicht gasförmigen organophosphatischen Nematiziden eine wichtige Rolle spielen oder wie und warum diese Prozess entsteht. Auch gibt es wenige Erkenntinsse darüber ob der Prozess der Bio-Degradierung verzögert, gestoppt oder umgekehrt werden kann. In diesen Untersuchungen wurden Böden von vier Bananenfeldern Costa Ricas, die zuvor mit verschiedenen Nematiziden behandelt wurden, genauer betrachtet. Es zeigte sich das die Behandlung mit dem Nicht-Begasungs Organophosphate Nematizid Terbufos einen Bekämpfungserfolg gegen den Nematoden Radopholus Similis zur Folge hatte sofern die Böden zuvor nicht so häufig mit dem Nematizid Terbufos behandelt wurden. Dieser Effekt konnte auf den hohen Anteil von Mikroorganismen in den Böden zurückgeführt werden, die den Wirkstoff im Boden schnell abbauten. Weiter Versuche mit verschiedenen Böden aus Deutschland zeigten, dass Böden die erstmals mit dem Nicht-Begasungs Organophosphate Nematizid Fenamiphos behandelt wurden, den Wirkstoff im Boden nicht ausreichend schnell biologisch abgebauen konnten. Verschiedene Bakterien der Gattung Pseudomonas konnten den Wirkstoff hier nicht metabolisieren. Ein Anstieg der Pseudomonas Population wurde nach einer Fenamiphos Behandlung nicht ermittelt. Der Mangel der Surfactant Produktion der bodenbürtigen Bakterien könnte ein Grund für den fehlenden biologischen Abbau sein. Folglich, könnten nur vereinzelte Pseudomonas spp. Stämme Nematizide abbauen. In weiteren Versuchen wurden aus deutschen Böden, die zuvor häufig mit Fenamiphos behandelt wurden, 17 Fenamiphos abbauende Bakterienstämme isoliert. Diese Bakterien bauten den Fenamiphos schnell ab. Weitere Versuche zeigten, dass ein Bakterienstamm den Wirkstoff als Kohlenstoffquelle für sein Wachstum nutzte. DNA Profile der Fenamiphos abbauenden Bakterienstämme wiesen 5 verschiedene RFLP Muster auf. Diese Bakterien wurden als Microbacterium, Sinorhizobium, Brevundimonas, Ralstonia oder Cupriavidus Spezies anhand ihrer partiellen 16S rRNA Gensequenzen identifiziert. Phylogenetische Analysen mit die Bakterien zeigten enge Verwandtschaft mit einander und haben gezeigt dass die Bakterien stammten von dem gleichen Vorfahren ab. Multiple Sequenz Analyse von den Fenamiphos abbauenden Bakterien ergaben identische Nucleotide Regionen mit Bakterien von ein Genebank. Die Fenamiphos abbauenden Bakterien bauten das neuartige Organophosphate Nematizid Fosthiazate nicht ab wodurch eine Fenamiphos Spezifizierung der Bakterien nachgewiesen werden konnte. Jedoch, in den Böden, in denen zuvor die Fenamiphos abbauenden Bakterien isoliert wurden, wurde der Wirkstoff Fosthiazate, aufgrund des hohen Mikroorganismen Anteil im Boden, abgebaut. Applikationen von Pflanzen revitalisierenden Mitteln erhöhte die mikrobielle Biomasse im Boden. Das frühe Eindringen des Wurzelgallen Nematoden Meloidogyne incognita wurde gehemmt. Der Abbau von Fenamiphos wurde verzögert. Zusammenfassend zeigte diese Arbeit, dass spezifische bodenbürtige Bakterien sich an bestimmte Nematizide anpassen und deren Wirkstoff als Kohlenstoffquelle für sich nutzen können. Dieser Effekt verlangsamte sich mit veränderter Populationsdichte der Mikroorganismen. Die Diversität durch Applikation von biologischen Pflanzenfördern hemmte den Nematodenbefall selbst wenn nicht Nematizid abbauende Stämme im Boden vorkommen.

i
1. GENERAL INTRODUCTION...........................................................................................1 1.1. IMPORTANCE OF PLANT-PARASITIC NEMATODES.........................................................1 1.2. USE OF NEMATICIDES IN AGRICULTURE.......................................................................1 1.3. ENHANCED BIODEGRADATION OF NON-FUMIGANT NEMATICIDES ...............................3 1.4. CROSS BIODEGRADATION............................................................................................4 1.5. NATURAL PLANT ENHANCERS .....................................................................................4 1.6. SCOPE OF THE STUDY ..................................................................................................5 1.7. REFERENCES ...............................................................................................................5
2. GENERAL MATERIALS AND METHODS .................................................................10 2.1. LOCATION OF FIELD RESEARCH IN COSTA RICA ........................................................10 2.2. GERMAN SOILS USED FOR ENHANCED BIODEGRADATION SCREENING........................11 2.3. CULTURE MEDIA, ANTIBIOTICS, FUNGICIDES AND REAGENTS....................................11 2.4. ISOLATION OF BACTERIA FROM SOIL .........................................................................13 2.5. ORIGIN AND CULTURE OF PLANT-PARASITIC NEMATODES .........................................14
2.5.1. Radopholus similis ..............................................................................................14 2.5.2. Meloidogyne incognita........................................................................................14
2.6. NEMATICIDES ...........................................................................................................15 2.7. IDENTIFICATION OF SOIL BACTERIA...........................................................................15
2.7.1. Gas chromatography technique (GC-FAME)......................................................15 2.7.2. Molecular characterization and identification technique (16S rRNA)................16
2.7.2.1. Bacterial culture and preparation of compounds for the PCR Master Mix 17 2.7.2.2. PCR Master Mix preparation ......................................................................17 2.7.2.3. PCR procedure.............................................................................................17 2.7.2.4. Gel electrophoresis analysis ........................................................................18 2.7.2.5. Restriction enzyme analysis .........................................................................18 2.7.2.6. DNA purification..........................................................................................19 2.7.2.7. DNA Sequencing and bacterial identification .............................................20
2.8. HIGH PRESSURE LIQUID CHROMATOGRAPHY (HPLC ANALYSIS)...............................20 2.8.1. Quantification of nematicides in soil extract liquid medium...............................21 2.8.2. Extraction and quantification of nematicides in soil extract agar medium.........21
2.9. REFERENCES .............................................................................................................22
3. FIELD EVIDENCE OF TERBUFOS ENHANCED BIODEGRADATION IN BANANA CULTIVATION...............................................................................................23
3.1. INTRODUCTION .........................................................................................................23 3.2. MATERIALS AND METHODS ......................................................................................24
3.2.1. Soil and root collection........................................................................................24 3.2.2. Determination of terbufos biodegradability and side-effect................................26 3.2.3. Determination of nematode diversity in banana roots ........................................27 3.2.4 Statistical analysis ...............................................................................................27
3.3. RESULTS ...................................................................................................................27 3.3.1 Biodegradability of terbufos ................................................................................27 3.3.2. Terbufos side-effect..............................................................................................29 3.3.3. Nematode diversity in banana roots ....................................................................29
3.4. DISCUSSION ..............................................................................................................30 3.5. REFERENCES .............................................................................................................33

ii
4. INVOLVEMENT OF MICROORGANISMS OTHER THAN PSEUDOMONADS ON FENAMIPHOS ENHANCED DEGRADATION ....................................................37 4.1. INTRODUCTION .........................................................................................................37 4.2. MATERIALS AND METHODS ......................................................................................38
4.2.1. Effect of compost on total soil bacteria and Pseudomonas spp...........................38 4.2.2. Efficacy of fenamiphos after three consecutive treatments..................................38 4.2.3. Analysis of biosurfactant production ...................................................................39
4.2.3.1. Drop collapse test.........................................................................................39 4.2.3.2. Blue media test.............................................................................................39
4.2.4. Identification of Pseudomonas spp. .....................................................................39 4.2.5. Fenamiphos metabolization and presence of Pseudomonads in the degrading microorganisms................................................................................................................40
4.3. RESULTS ...................................................................................................................41 4.3.1. Effect of compost on total soil bacteria and Pseudomonas spp...........................41 4.3.2. Efficacy of fenamiphos on nematode control.......................................................42 4.3.3. Biosurfactant production tests .............................................................................43 4.3.4. Identification of single bacterial colonies............................................................44 4.3.5. Pseudomonads and the conversion of fenamiphos ..............................................45
4.4. DISCUSSION ..............................................................................................................47 4.5. REFERENCES .............................................................................................................50
5. ISOLATION AND CHARACTERIZATION OF FENAMIPHOS-DEGRADING MICROORGANISMS.......................................................................................................54 5.1. INTRODUCTION .........................................................................................................54 5.2. MATERIALS AND METHODS ......................................................................................55
5.2.1. Determination of microorganisms responsible for enhanced fenamiphos degradation......................................................................................................................55 5.2.2. Metabolization of fenamiphos by bacterial single colonies.................................56 5.2.3. Use of fenamiphos as sole carbon source............................................................57 5.2.4. Molecular characterization and identification of fenamiphos degrading bacteria 58 5.2.5. Phylogenetic analysis and sequence comparison ................................................58
5.3. RESULTS ...................................................................................................................59 5.3.1. Determination of microorganism responsible for enhanced fenamiphos degradation......................................................................................................................59 5.3.2. Metabolization of fenamiphos by bacterial single colonies.................................60 5.3.3. Use of fenamiphos as sole carbon source............................................................62 5.3.4. Molecular characterization and identification of fenamiphos degrading bacteria 63 5.3.5. Phylogenetic analysis and sequence comparison ................................................65
5.4. DISCUSSION ..............................................................................................................67 5.5. REFERENCES .............................................................................................................71
6. FOSTHIAZATE CROSS-DEGRADATION AND SPECIFICITY OF NEMATICIDE DEGRADING BACTERIA...............................................................................................76 6.1. INTRODUCTION .........................................................................................................76 6.2. MATERIALS AND METHODS ......................................................................................77
6.2.1. Degradation of fosthiazate...................................................................................77 6.2.2. Cross-degradation essays ....................................................................................77
6.3. RESULTS ...................................................................................................................78

iii
6.3.1. Degradation of fosthiazate...................................................................................78 6.3.2. The effect of fosthiazate on fenamiphos degradation ..........................................79 6.3.2. Degradation of fosthiazate by fenamiphos-degrading bacteria ..........................80
6.4. DISCUSSION ..............................................................................................................81 6.5. REFERENCES .............................................................................................................82
7. EFFECT OF NATURAL PLANT ENHANCERS ON SOIL BACTERIA, MELOIDOGYNE INCOGNITA AND NEMATICIDE DEGRADATION....................85 7.1. INTRODUCTION .........................................................................................................85 7.2. MATERIALS AND METHODS ......................................................................................86
7.2.1. Effect of different M. incognita inoculum densities on lettuce cv. Milan ..............86 7.2.2. Effect of plant enhancers on soil bacteria, Meloidogyne incognita and biodegradation.................................................................................................................87
7.2.2.1. Natural plant enhancers and nematicides .................................................................................. 87 7.2.2.2. Soil treatment with plant enhancers and nematicides ................................................................ 87 7.2.2.3. Effect on soil bacteria population densities ............................................................................... 88 7.2.2.4. Effect on M. incognita early root penetration............................................................................ 88 7.2.2.5. Effect on enhanced biodegradation of fenamiphos.................................................................... 88
7.2.3. Statistical analysis ...............................................................................................89 7.3. RESULTS ...................................................................................................................89
7.3.1. Effect of different M. incognita inoculum densities on lettuce cv. Milan ..............89 7.3.2. Effect of plant enhancers on soil bacteria, Meloidogyne incognita and biodegradation.................................................................................................................90
7.3.2.1. Effect on soil bacteria population densities ....................................................90 7.3.2.2. Effect on M. incognita early root penetration.................................................91 7.3.2.3. Effect on enhanced biodegradation of fenamiphos.........................................92
7.4. DISCUSSION ..............................................................................................................93 7.5. REFERENCES .............................................................................................................96

Chapter 1 General Introduction
1
1. GENERAL INTRODUCTION
1.1. Importance of plant-parasitic nematodes
Nematodes are microscopic, aquatic, elongated, tubular, spindle shaped worms that live in
moist surfaces, films of water within soil and in moist tissues of different organisms and
plants (Dropkin, 1989). Most plant-parasitic nematodes attack underground plant parts,
especially roots (Whitehead, 1998). Other species are predominantly shoot parasites,
attacking stems, leaves, flowers, seeds or combinations thereof. Nematodes may feed ecto-,
semi-endo-, or endo-parasitically on host tissue using a narrow mouth spear or stylet. Some
nematodes can transmit pathogenic viruses with their stylet (Evans et al., 1993). Most plant-
parasitic nematodes are obligate parasites. The opening in the root tissue made with the stylet
to penetrate or feed may be used later by pathogenic bacteria and fungi for secondary
infections (Luc et al., 2005).
Plant parasitic nematodes are an important limiting factor to crop production in temperate,
tropical and sub-tropical agriculture (Evans et al., 1993; Luc et al., 2005). The damage done
to a plant depends on the nematode species and the number of nematodes feeding on it
(Whitehead, 1998). Most crops including cereals, vegetables, fruit trees and fibre plants are
susceptible to several nematode species. Yield losses and crop quality reduction caused by
nematodes have negative economic consequences on farmers, consumers and society
(Webster, 1972).
The aim of nematode control is to restrict significant yield losses and quality in vulnerable
crop plants and, in the longer term, to keep plant-parasitic nematode populations under the
threshold level (Whitehead, 1998). Despite the use of crop rotation, soil amendments,
resistant/tolerant varieties, catch crops and biocontrol agents, the control of plant parasitic
nematodes still relies heavily on the use of chemical nematicides worldwide.
1.2. Use of nematicides in agriculture
The role of chemicals in nematode control has been well reviewed by Whitehead (1998),
Hague and Gowen (1987) and Johnson (1985). Sikora and Marczok (2005) provided a list of
the most common chemicals available on the market used for nematode control. Chemicals

Chapter 1 General Introduction
2
which paralyse or kill nematodes are referred to as nematicides (Whitehead, 1998). They are
classified as fumigant or non-fumigant types. Fumigant nematicides have large vapour
pressures and diffuse rapidly through the network of soil pores in the gas phase. Most of
these chemicals are either halogenated aliphatic hydrocarbons or methyl isothiocyanate
precursor compounds with toxic effect on almost all living organisms including bacteria,
fungi and plants. The fumigant nematicides are highly effective in nematode control. To
prevent plant damage they must be applied long before planting or transplanting. However,
the use of fumigant nematicides, such as methyl bromide, has decreased in modern
agriculture due to an environmental concerns which resulted in a restriction of use (Santos et
al., 2006; Webster et al., 2001).
Non-fumigant nematicides are granular or liquid compounds which are water soluble and
have either contact or nematistatic and systemic activity against nematodes (Sikora and
Fernandez, 2005). Some of these nematicides are also used as insecticides. The non-fumigant
nematicides are applied to soil at concentrations that paralyze nematodes and do not kill
them. This type of nematicides can be divided into two groups, the organophosphates and the
carbamates, according to their molecule structure. In most cases, the mechanism of action of
both groups is associated with suppression of nematode mobility during the period when
adequate concentrations are present in the soil. These non-volatile types of nematicides are
preferred nowadays in modern agriculture since they are more specific than the fumigants,
have less environmental risk and are generally not phytotoxic (Cabrera et al., 2009a). They
can be applied to the soil even when the crop has been established or as seed treatment
(Cabrera et al., 2009b). The lack of new non-fumigant nematicide molecules has lead to the
repetitive application of the same compounds for nematode management. It takes about 12
years to develop a new non-fumigant nematicide for the market. The repeated application of
the same nematicide is practiced specially in monoculture systems, for example in banana
production, where nematicide treatments are performed according to an application calendar
that can vary from 2 to 3 times per year. These repeated applications may influence the
emergence of specific soil microorganisms that can degrade the active substances at
accelerated rates (Smelt et al., 1987; Ou et al., 1994).

Chapter 1 General Introduction
3
1.3. Enhanced biodegradation of non-fumigant nematicides
The accelerated microbial degradation of pesticides was first described in 1951 with the
herbicide 2,4-D (Audus, 1951). Since then over 375 publications have appeared which
describe the biodegradation of more than 50 different soil applied crop protection products
(Anderson et al., 1998) such as insecticides (Felsot et al., 1981; Read, 1983; Racke and
Coats, 1988; Morel-Chevillet et al., 1996) fungicides (Walker, 1987; Thom et al., 1997) and
herbicides (Kirkland and Fryer, 1972; Torstensson et al., 1975; Wilson, 1984; Gray and Joo,
1985; Skipper et al., 1986).
Enhanced biodegradation is the rapid microbial degradation of a pesticide, in the present case
nematicides, by a specialized fraction of the soil microflora (Karpouzas and Giannakou,
2002). Soil microorganisms have adapted in such a way that they can rapidly metabolize
specific nematicides. The degradation products (metabolites) of nematicides have been
implicated in an enrichment emergence of soil microflora capable of accelerated degradation
of parent compounds (Ou and Rao, 1986; Jones and Estes, 1995). The nematicide
biodegradation in monoculture systems was reported in the early 1990’s from soil cultivated
with tomato after repetitive nematicide applications using the same active substance (Stirling
et al., 1992). A similar effect was observed later in soil cultivated with turfgrass (Ou et al.,
1994) and potato (Karpouzas et al., 1999). Mclean and Lawrence (2003) found a lack of
nematicide efficacy in a soil cultivated with cotton which was treated often with the same
chemical.
At the present time, the most widely used pesticides belong to the organophosphorus group
(Singh and Walker, 2006). The enhanced biodegradation of several insecticides of this
chemical group, such as diazinon, chlorpyrifos, parathion and dimethoate has been reported
(Sethunathan and Pathak, 1972; Singh and Walker, 2006; Drufovka et al., 2008; Li et al.,
2008). The organophosphate herbicide glyphosphate and the acaricide coumaphos have also
been found to rapidly biodegrade (Singh and Walker, 2006). However, studies on
organophosphate nematicides are rarer. Even though there are reports of biodegradation on
this nematicide group it remains unclear which microorganisms play the most important role
in the rapid metabolization and how and why this process develops. To date, there is little
information whether the enhanced biodegradation process can be slowed down or reversed by
manipulating the natural soil microbial populations.

Chapter 1 General Introduction
4
1.4. Cross biodegradation
Cross biodegradation is referred to when microorganisms have been frequently exposed to
one nematicide and the microbes are able to simultaneously degrade other nematicides. The
cross-enhancement of selected pesticides in soil can be dependent on the structural similarity
of the compounds (Singh et al., 2005). For example, the bacterial population of a cadusafos-
adapted soil was able to rapidly degrade the chemically related nematicide ethoprophos
(Karpouzas et al., 2004b). The cross adaptation of microorganisms decreases exponentially
the efficacy of nematicides but it also can be considered an important process in the rapid
degradation of pesticides from the soil environment (Ankumah et al., 2008). However, not all
microorganisms that can degrade one nematicide are able to metabolize another one
(Karpouzas and Walker, 2000; Karpouzas et al., 2004a). To avoid the onset of
biodegradation, and therefore cross-degradation, sufficient chemical rotation, i.e. the use of
active ingredients from different chemical groups, in combination with crop rotation and the
use of resistant cultivars is recommended (Karpouzas and Giannakou, 2002). However, the
cross-degradation process is still poorly understood (Suett and Jukes, 1988) and whether
adapted microorganisms to old molecules can also degrade the new nematicides at
accelerated rates is currently unknown.
1.5. Natural plant enhancers
Application of residue amendments derived from crops or animals is known to improve crop
yield. The use of amendments has become an important agronomic tool and an efficient way
of improving plant nutrition and therefore they are considered to be plant growth enhancers
(Mulawarman et al., 2001; Mulawarman, 2002). A beneficial effect of adding extra nutrients
to soil is to increase crop yield and performance (Muller and Gooch, 1982; Mullins and
Mitchell, 1995). Most of these residue derived compounds are organic amendments. The use
of organic amendments which can provide extra nutrients to soil, increase crop yield and
control plant-parasitic nematodes has been previously reviewed (Mankau, 1962/1968; Muller
and Gooch, 1982). More recently, it has been reported that in some cases the benefits of
adding organic material or residues to the soil is mainly due to a decrease in the levels of soil
pathogens (Mullawarman et al., 2001). This is caused by an alteration in soil structure and
ecology, and/or by the action of residue derived chemicals on the soil fauna and flora.
Organic amendments deliver nutrients which specifically support the growth of antagonistic

Chapter 1 General Introduction
5
fungi on other organisms (Muller and Gooch, 1982; Rodriguez-Kabana, 1986). However, it
has still not been reported whether the addition of organic amendments or plant enhancers
can also support or suppress the growth of microorganisms responsible for the enhanced
degradation of nematicides. It is known that once the enhanced biodegradation of nematicides
is present in a soil it can stay active for prolonged periods after the last application (Smelt et
al., 1996). If the microorganisms involved in the degradation process could be reduced
through the application of organic amendments or plant enhancers and nematicide-efficacy
re-establish is still unknown.
1.6. Scope of the study
The overall goal of the present study was to investigate the enhanced biodegradation process
in soils with different histories of non-fumigant nematicide applications. The objectives of
the research in this thesis were:
1. Demonstrate the enhanced biodegradation process occurring in soil of a banana
monoculture system.
2. Study the microorganisms involved in the rapid nematicide metabolization.
3. Isolate, indentify and characterize nematicide-degrading microorganisms.
4. Determine the occurrence of cross-degradation from old to new nematicide
molecules.
5. Determine whether natural plant enhancers can slow down or reverse the rapid
nematicide breakdown process caused by microorganisms.
1.7. References
Anderson, J.P.E., Nevermann, K. and Haidt, H. 1998. Accelerated microbial degradation of
nematicides in soils: problem and its management. In: Proceedings of the XIII Acorbat
Meeting. Guayaquil, Ecuador, pp. 568–579.
Ankumah, R.O., Joshua, C., Egiebor, E.S., Hamilton, J., Watt, I., Bambele, L., Gaskin, N.,
Sutherland, Y. and Brown, D.D. 2008. Implications of cross-enhancement and influence

Chapter 1 General Introduction
6
of carbon sources in the enhanced degradation of EPTC, butylate and fenamiphos by
isolated soil microorganisms. Recent Advances in Agriculture, 289-306.
Audus, L.J. 1951. The biological detoxication of hormone herbicides in soil. Plant and Soil
3:170-192.
Cabrera, J.A., Kiewnick, S., Grimm, C., Dababat, A.A. and Sikora, R.A. 2009a. Efficacy of
abamectin seed treatment on Pratylenchus zeae, Meloidogyne incognita, and
Heterodera schachtii. Journal of Plant Diseases and Protection 116:124-128.
Cabrera, J.A., Kiewnick, S., Grimm, C., Dababat, A.A. and Sikora, R.A. 2009b. Effective
concentration and range of activity of abamectin as seed treatment against root-knot
nematodes on tomato under glasshouse conditions. Nematology in press.
Dropkin, V.H. 1989. Introduction to plant nematology. 2nd edition. John Wiley & Sonst, Inc.
USA. p. 304.
Drufovka, K., Danevčič, T., Trebše, P. and Stopar, D. 2008. Microorganisms trigger chemical
degradation of diazinon. International Biodeterioration and Biodegradation 62:293-296.
Evans, K., Trudgill, D.L. and Webster, J.M., 1993. Plant parasitic nematodes in temperate
agriculture. CAB International, Wallingford United Kingdom. pp 648.
Felsot, A.S., Maddox, J.V. and Bruce, W. 1981. Enhanced microbial degradation of carbofuran
in soils with histories of Furadan use. Bulletin of Environmental Contamination and
Toxicology 26:781-788 .
Gray, R.A. and Joo, G.K. 1985. Reduction in weed control after repeated applications of
thiocarbamate and other herbicides. Weed Science 33:698-702.
Hague, N.G.M. and Gowen, S.R. 1987. Chemical control of nematodes. In: Brwon, R.H. and
Kerry, B.R. (eds) Principles and Practices of Nematode Control. Academic Press, New
York, pp. 131-173.
Johnson, A.W. 1985. The role of nematicides in nematode management. In: Sasser, J.N. and
Carter, C.C. (eds) An Advanced Treatise on Meloidogyne. Volume I. Biology and
Control. North Carolina University Graphics, Raleight, North Carolina, USA. pp. 249-
267.
Jones, R.L. and Estes, T.L. 1995. Summary of aldicarb monitoring and research programs in
the United States. Journal of Contaminant Hydrology 18:107-140.
Karpouzas, D.G. and Giannakou, I.O. 2002. Biodegradation and enhanced biodegradation: a
reason for reduced biological efficacy of nematicides. Russian Journal of Nematology
10:59-78.

Chapter 1 General Introduction
7
Karpouzas, D.G., Giannakou, I.O., Walker, A. and Gowen, S. 1999. Reduction in biological
efficacy of ethoprophos in a soil from Greece due to enhanced biodegradation:
comparing bioassay with laboratory data. Pesticide Science 55:1089-1094.
Karpouzas, D.G., Hatziapostolou, P., Papadopoulou-Mourkidou, E., Giannakou, I.O. and
Georgiadou, A. 2004a. The enhanced biodegradation of fenamiphos in soils from
previously treated sites and the effect of soil fumigants. Environmental Toxicology
and Chemistry 23:2099-2107.
Karpouzas, D.G., Karanasios, E., Menkissoglu-Spiroudi, U. 2004b. Enhanced microbial
degradation of cadusafos in soils from potato monoculture: demonstration and
characterization. Chemosphere 56:549-559.
Karpouzas, D.G. and Walker, A. 2000. Aspects of the enhanced biodegradation and
metabolism of ethoprophos in soil. Pest Management Science 56:540-548.
Kirkland, K. and Fryer, J.D. 1972. Degradation of several herbicides in a soil previously
treated with MCPA. Weed Research 12:90-95.
Li, X.H., Jiang, J.D., Gu, L.F., Ali, S.W., He, J. and Li, S.P. 2008. Diversity of chlorpyrifos-
degrading bacteria isolated from chlorpyrifos-contaminated samples. International
Biodeterioration and Biodegradation 62:331-335.
Luc, M., Sikora R.A. and Bridge J. 2005. Plant parasitic nematodes in tropical and subtropical
agriculture, second edition. CAB International, Wallingford, UK. pp 871.
Mankau, R. 1962. The effect of some organic additives upon a soil nematode population and
associated natural enemies. Nematologica 7:65-73.
Mankau, R. 1968. Reduction of root-knot disease with organic amendments under semifield
conditions. Plant Disease Reporter 52:315-319.
Mclean, K.S. and Lawrence, G.W. 2003. Efficacy of aldicarb to Rotylenchus reniformis and
biodegradation in cotton field soils. Journal of Nematology 35:65-72.
Morel-Chevillet, C., Parekh, N., Pautrel, and D., Fournier, J.C. 1996. Cross-enhancement of
carbofuran biodegradation in soil samples previously treated with carbamate
pesticides. Soil Biology and Biochemistry 28:1767-1776.
Mulawarman, 2002. Use of natural products based on renewable raw materials to stimulate
soil health and control Meloidogyne incognita. PhD Thesis, University of Bonn,
Germany. p.115.
Mulawarman, Hallmann, J., Bell, D., Kopp-Holtwiesche, B. and Sikora, R.A. 2001. Effects of
natural products on soil organisms and plant health enhancement. Mededelingen -

Chapter 1 General Introduction
8
Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, Universiteit
Gent. 66:609-617.
Muller, R. and Gooch, P.S. 1982. Organic amendments in nematode control. An examination
of the literature. Nematropica 12:319-326.
Mullins, G.L. and Mitchell, C.C. 1995. Crops. In: J.E. Reccigl. Soil Amendments: Impact on
Biotic System. Agriculture Ecology, pp. 2-30.
Ou, L.T. and Rao P.S.C. 1986. Degradation and metabolism of oxamyl and fenamiphos in
soils. Journal of Environmental Sciences and Health B 21:25-40.
Ou, L.T., Thomas, J.E. and Dickson D.W. 1994. Degradation of fenamiphos in soil with a
history of continuous fenamiphos applications. Soil Science Society of American
Journal 58:1139-1147.
Racke, K.D. and Coats J.R. 1988. Enhanced degradation and the comparative fate of
carbamate insecticides in soil. Journal of Agricultural and Food Chemistry 36:1067-
1072.
Read, D.C. 1983. Enhanced microbial degradation of carbofuran and fensulfothion after
repeated applications to acid mineral soil. Agriculture Ecosystems and Environment
10:37-46.
Rodriguez-Kabana, R. 1986. Organic and inorganic amendments to soil as nematode
suppressants. Journal of Nematology 18:129-135.
Santos, B.M., Gilreath, J.P., Motis, T.N., Noling, J.W., Jones, J.P. and Norton, J. A. 2006.
Comparing methyl bromide alternatives for soilborne disease, nematode and weed
management in fresh market tomato. Crop Protection 25:690-695.
Sethunathan, N. and Pathak, M.D. 1972. Increased biological hydrolysis of diazinon after
repeated application in rice paddies. Journal of Agricultural Food Chemistry 20:586-
589.
Sikora, R.A. and Fernandez, E. 2005. Nematodes parasites of vegetables. In: Luc, M., Sikora,
R.A. and Bridge, J. (Eds). Plant parasitic nematodes in tropical and subtropical
agriculture, second edition. CAB International, Wallingford, UK, pp. 319-392.
Sikora, R.A. and Marczok, P. 2005. Apendix A, nematicides. In: Luc, M., Sikora, R.A. and
Bridge, J. (Eds). Plant parasitic nematodes in tropical and subtropical agriculture,
second edition. CAB International, Wallingford, UK, pp. 827-829.
Singh, B.K. and Walker, A. 2006. Microbial degradation of organophosphorus compounds.
FEMS Microbiological Reviews 30:428-471.

Chapter 1 General Introduction
9
Singh, B.K., Walker, A. and Wright, D.J. 2005. Cross-enhancement of accelerated
biodegradation of organophosphorus compounds in soils: dependence on structural
similarity of compounds. Soil Biology and Biochemistry 37:1675-1682.
Skipper, H.D., Murdock, E.C., Gooden, D.T., Zublena, J.P. and Amakiri, M.A. 1986.
Enhanced herbicide biodegradation in South Carolina soils previously treated with
butylate. Weed Science 34:558-563.
Smelt, J.H., Crum, S.J.H., Teunissen, W. and Leistra, M. 1987. Accelerated transformation of
aldicarb, oxamyl and ethoprophos after repeated soil treatments. Crop Protection
6:295-303.
Smelt, J.H., van de Peppel-Groen, A.E., van der Pas, L.J.T., and Dijksterhuis, A. 1996.
Development and duration of accelerated degradation of nematicides in different soils.
Soil Biology and Biochemistry 28:1757-1765.
Stirling, A.M., Stirling, G.R. and Macrae, I.C. 1992. Microbial degradation of fenamiphos
after repeated application to tomato growing soil. Nematologica 38:245-254.
Suett, D.L. and Jukes, A.A. 1988. Accelerated degradation of aldicarb and its oxidation
products in previously treated soils. Crop Protection 7:147-152.
Thom, E., Ottow, J.C.G. and Benckiser, G. 1997. Degradation of the fungicide
difenoconazole in a silt loam soil as affected by pretreatment and organic matter.
Environmental Pollution 96:409-414.
Torstensson, N.T.L., Stark, J. and Göransson, B. 1975. The effect of repeated applications of
2,4-D and MCPA on their breakdown in soil. Weed Research 15:159-164.
Walker, A. 1987. Enhanced degradation of iprodione and vinclozolin in soil: a simple
colorimetric test for identification of rapid-degrading soils. Pesticide Science 21:233-
240.
Webster, J.M. 1972. Economic nematology. Academic Press Inc. London, UK. p.563.
Webster, T.M., Csinos, A.S., Johnson, A.W., Dowler, C.C., Summer, D.R. and Fery, R.L. 2001.
Methyl bromide alternatives in a bell pepper-squash rotation. Crop Protection 20:605-
614.
Whitehead, A.G. 1998. Plant nematode control. CAB International Wallingford, Oxford, UK. p
384.
Wilson, R.G. 1984. Accelerated degradation of thiocarbamate herbicides in soil with prior
thiocarbamate exposure. Weed Science 32:264-268.

Chapter 2 General Materials and Methods
10
2. GENERAL MATERIALS AND METHODS
2.1. Location of field research in Costa Rica
To investigate the enhanced biodegradation in a mono culture system, 4 commercial banana
plantations were selected in the Atlantic region of Costa Rica (Figure 1). This region contains
95% of all commercial banana plantations of the country. The Reventazon River (RR) was a
natural soil dividing factor. Soils of banana fields located west of the river (F1, F3 and F4)
were of volcanic origin and the east one (F2) of river sediments (Serrano et al., 2006). The
average mean temperature and the average relative humidity in this area between 2000 and
2005 was 25.5 °C and 83.6% as reported by the meteorological station at Siquirres, Costa
Rica, respectively.
F1
F2F3
F4
Other banana farms
Other banana farms
Reventazon River
Atlantic Ocean
F1
F2F3
F4
Other banana farms
Other banana farms
Reventazon River
Atlantic Ocean
Figure 1. Enlargement of the Atlantic region of Costa Rica showing fields with banana cultivation in the area (dark gray blocks). Collection of soil and roots was performed in the four commercial banana plantations indicated as F1, F2, F3 and F4. Map of Costa Rica (above right). This map was adapted from Serrano et al. (2006).

Chapter 2 General Materials and Methods
11
2.2. German soils used for enhanced biodegradation screening
To investigate the enhanced biodegradation phenomena at laboratory and greenhouse level
samples from four soils were used in this investigation. All soil samples were collected from
the top 20 cm of soil in fields of experimental stations in Nordrhein Westfalen, Germany.
Soil 1 (S1): was obtained from the field 1 of the experimental farm Laacher Hof of Bayer
CropScience AG. This soil was composed of 24.5% clay, 62.6% silt and 12.9% sand and had
a pH of 4.06. This soil was treated 42 times with fenamiphos in the last 16 years.
Soil 2 (S2): was obtained from the same field as S1 but has received 25 fenamiphos
treatments in 12 years. This soil was composed of 23.1% clay, 60.8% silt and 16.1% sand,
and had a pH of 4.51.
Soil 3 (S3): was obtained from the field Am Hohenseh 4a, Burscheid, of the experimental
farm Höfchen of Bayer CropScience AG. This soil was composed of 16.2% clay, 78.4% silt
and 5.4% sand, and had a pH of 6.17. This soil has received 25 fenamiphos treatments in 12
years.
Turf grass was grown in these three soils before arrival to our laboratory. The soil samples
collected in these soils were stored at 20 ± 2 °C in the dark. The last fenamiphos treatment
was performed about 1 month before this investigation started.
Soil 4 (S4): was soil obtained from the research station of the University of Bonn at Klein
Altendorf, Rheinbach. This soil was composed of 14.6% clay, 77.6% silt and 7.8% sand, and
had a pH of 6.75. The field was previously cultivated with wheat during summer season and
has never received fenamiphos or other nematicide treatment before arrival to the laboratory.
During the research period all soils were kept in slightly capped plastic containers at 8 °C in
the dark and have received autoclaved water to their holding capacity to maintain moisture.
2.3. Culture media, antibiotics, fungicides and reagents
Acetonitrile: HPLC grade (Sigma-Aldrich).
Agar: AppliChem GmbH.
Ampicillin: A stock solution was prepared dissolving Ampicillin (AppliChem) in autoclaved
(121 ºC for 20 min) distilled water to a desired concentration following filter-sterilization (0.4
µm). The stock solution was kept at -20 °C.
Blue media: 20 g Mannitol (C6H14O6), 0.7 g Potassium dihydrogen (KH2PO4), 0.9 g
Sodiumphosphatedibasic heptahydrate (NA2HPO4), 2.0 g Sodiumnitrate (NaNO3), 0.4 g

Chapter 2 General Materials and Methods
12
Magnesiumsulfate heptahydrate (MgSO4 7H2O), 0.1 g Calcium chloride anhydrous (CaCl2
2H2O), 2 ml Trace Elements solution, 0.2 g CTAB (Cetyl trimetrilamonium bromide,
Cl9H42NBr), 0.005 g Methylene blue and 15 g bacteriological agar dissolved in 1 L
autoclaved distilled water.
Buffer phosphate solution (BPS): 1.39 g HK2 O4P (136.09 g mol-1, Merk) and 1.08 g
H2KO4P (136.09 g mol-1, Merk) dissolved in 800 ml autoclaved distilled water.
Chloramphenicol: A stock solution was prepared dissolving Chloramphenicol (AppliChem)
in 80% ethanol (v:v) to a desired concentration following filter-sterilization. The stock
solution was kept at -20 ºC.
Cycloheximide: A stock solution was prepared dissolving Cycloheximide (Sigma) in 80%
ethanol (v:v) to a desired concentration following filter-sterilization. The stock solution was
kept at -20 ºC.
Ethanol: HPLC grade (Merck).
Glycerol: (Merck).
GC-TSB: 24 g purified and fine Trypticase Soy Broth (Becton Dickson) made for gas
chromatography and 16 g Bacto Agar in 800 ml autoclaved distilled water.
Hexan: HPLC grade >95% purity (Merck).
KBM: 26.3 g King B Agar (Fluka, Biochemika) and 8 ml glycerol in 790 ml autoclaved
distilled water.
KMB+: 26.3 g King B Agar (Fluka, Biochemika) and 8 ml de glycerol in 790 ml autoclaved
distilled water amended with filter-sterilized stock solutions of 13 µg Chloramphenicol ml-1,
40 µg Ampicillin ml-1 and 100 µg Cycloheximide ml-1 final concentrations.
Methanol: HPLC grade (Sigma-Aldrich).
PDA+: 19.2 g potato dextrose broth (Oxid LTD) and 14.4 g agar in 800 ml autoclaved
distilled water amended with 150 ppm of Streptomycin sulphate and 150 ppm of
Chloranphenicol.
Reagent 1: 45 g sodium hydroxide (certified ACS) pellets dissolved in 150 ml of methanol
(reagent grade) and 150 ml of deionised distilled water.
Reagent 2: 325 ml 6.0 N hydrochloric acid supplemented with 275 ml methanol (reagent
grade).
Reagent 3: 200 ml Methyl-tert Butyl Ether in 200 ml hexane (HPLC grade).
Reagent 4: 10.8 g sodium hydroxide pellets dissolved in 900 ml deionised distilled water.
Soil extract agar medium (SEAM): This medium was used since it contains low levels of
carbon (close to 0). Every 500 g of autoclaved S4 were mixed with 900 ml of autoclaved

Chapter 2 General Materials and Methods
13
distilled water in a Duncan bottle and shaken vigorously by hand for 1 minute. After shaking,
the solution was kept at room temperature for 72 hours. After this period, the solution was
transferred into cylindrical tubes and centrifuged for 10 min at 20 ºC, 10 brake at 2000 G.
The liquid was then filtered through folded filter paper (ø 385 mm, Whatman, Schleicher and
Schuell) and collected in glass flasks. 20 g of Difco Bacto Agar were added per liter of soil
extract, stirred and the pH was adjusted to 6.0. This mixture was autoclaved and poured in
sterile conditions into sterile 5 cm diameter Petri dishes. Each Petri dish was filled with 5 ml
of medium.
Soil extract liquid medium (SELM): The soil extract liquid medium was prepared as
described in the SEAM except for the addition of agar. pH = 6.
Streptomycinsulphate: A stock solution was prepared by dissolving Streptomycinsulphate
(AppliChem) in autoclaved distilled water to a desired concentration following filter-
sterilization. The stock solution was kept at -20 °C.
Trifluoroacetic acid: 99% spectrophotometric grade (Sigma-Aldrich).
TSA 100%: 9.6 g agar (Agar Bacteriology grade, AppliChem) and 24 g Tryptone Soya broth
(Oxoid) in 800 ml autoclaved distilled water.
TSA+: 9.6 g agar (Agar Bacteriology grade, AppliChem) and 24 g Tryptone Soya broth
(Oxoid) in 800 ml autoclaved distilled water amended with a filter-sterilized stock solution of
Cycloheximide to a final concentration of 100 µg ml-1.
Trace elements: 2 g iron sulphate heptaydrate (FeSO4 7H2O), 1.5 g Magnese sulphate
monohydrate (MNS4) and 0.6 g Amonium molybdate tetrahydrate [(NH4)6 Mo7O24 H2O] in 1
L of distilled water.
2.4. Isolation of bacteria from soil
Bacteria were isolated by transferring 10 g of soil to an autoclaved 300 ml Erlenmeyer flask
containing 100 ml of autoclaved distilled water. Flasks were shaken at 120 rpm for 16 h at 28
± 2 °C in an incubator in the dark. After incubation and under laminar flow, 1 ml of the
solution was transferred into 9 ml autoclaved BPS in a glass test tube. Serial dilutions to 1 X
10-3 in BPS were prepared. From the final dilution, 50 µL were transferred to TSA 100% in
10 cm diameter Petri dishes. About 10 autoclaved glass beads were placed in every Petri dish
which was closed and shaken by hand for 10 s. After shaking, the beads were removed and
the Petri dishes were sealed with Parafilm and incubated in the dark at 28 ± 2 ºC for 48 h.
Single colonies were picked with a sterile 4 mm loop and transferred onto fresh TSA+.

Chapter 2 General Materials and Methods
14
2.5. Origin and culture of plant-parasitic nematodes
2.5.1. Radopholus similis
Radopholus similis was isolated from banana roots in a commercial farm in Costa Rica. A
pure R. similis culture was maintained and multiplied in sterile carrot discs, as described by
Speijer and De Waele (1997). Carrots were cleaned with tap water, dried with paper tissue
and sprayed with 70% ethanol. The ethanol was burned off in the laminar flow. Both carrot
ends were removed and the carrot was peeled with a sterile scalpel. The carrots were cut into
discs and placed in sterile 3 cm Ø Petri dishes. R. similis was treated with Streptomycin
sulphate (2000 ppm) on a 20 µm aperture sieve. 2 hours after treatment, the nematodes were
collected from the sieve and inoculated at the outer edge of the carrot discs. Petri dishes were
sealed with Parafilm and kept in an incubator at 25 ºC in dark conditions for 6 weeks. After
this period of time, Petri dishes were taken out of incubation and opened under laminar flow.
Nematodes on the surface of the carrot disc were washed and transferred with tap water into a
beaker. The remaining carrot disc was macerated twice for 10 s at low speed in a commercial
kitchen blender with tap water. The macerated suspension was poured onto a 100 µm
aperture sieve to separate nematodes from carrot tissues. Nematodes that passed through the
sieve were collected in a beaker and mobile nematodes were counted and used in the
experiments as required.
2.5.2. Meloidogyne incognita
Meloidogyne incognita race 3 was obtained from a natural infested soil in Florida, USA, and
maintained on the tomato cv. Furor permanently cultivated in a green house at 27 ± 5 °C.
Tomato seedlings, 3-4 weeks old, were planted in autoclaved field soil mixed with sand (2:1,
v:v) inoculated with high numbers of M. incognita juveniles and eggs. Plants were fertilized
with 0.2% Polycrescol (14:10:14, N:P:K) once a week and watered as needed. Nematode
eggs were extracted from 8 weeks old tomato roots using 1.5% NaOCl following the method
described by Hussey and Barker (1973). Plants were removed from soil and uprooted. Roots
were gently washed with tap water, cut in 1-2 cm pieces and macerated 2 times for 10 s each
time in a Warring blender (Bender and Hohbein) with tap water. Every 500 ml of the
macerated solution was mixed with 258 ml of 4% NaOCl (AppliChem) and manually shaken
for 3 min. This suspension was poured over four nested sieves; 250 µm on the top, followed

Chapter 2 General Materials and Methods
15
by 100 µm, 45 µm and 25 µm aperture sieve. Eggs remaining in the 25 µm sieve were rinsed
with tap water to separate nematodes from NaOCl and were collected on a beaker for
experimental use.
2.6. Nematicides
DiTera® DF: Biological nematicide with 90% of non-viable Myrothecium verrucaria strain
AARC-0255 its metabolites and production substrates (Valent BioSciences U.S.A.).
Fenamiphos: Ethyl 4-methylthio-m-tolyl isopropylphosphoramidate analytical grade (Sigma-
Aldrich) was mixed with methanol to a desired concentration and filter sterilized (0.4 µm).
Fosthiazate 150 EC: (RS)-S-sec-butyl-O-ethyl 2-oxo-1,3-thiazolidin-3-ylphosphonothioate
liquid formulation 15% active ingredient (Syngenta Crop Protection, Switzerland).
Nemacur 5 GR: Ethyl 4-methylthio-m-tolyl isopropylphosphoramidate. Commercial
fenamiphos granular formulation 5% active ingredient (Bayer Crop Science, Germany) was
mixed with methanol to a desired concentration and filter sterilized (0.4 µm).
Nemathorin 10 WG: granular formulation 10% fosthiazate ((RS)-S-sec-butyl-O-ethyl 2-oxo-
1,3-thiazolidin-3-ylphosphonothioate) active ingredient (Syngenta Crop Protection,
Switzerland) was mixed with methanol to a desired concentration and filter sterilized (0.4
µm).
Terbufos 10 GR: S-tert-butylthiomethyl O, O-diethylphosphorodithioate granular
formulation 10% active ingredient (Industrias Bioquim Centroamericana S.A.).
2.7. Identification of soil bacteria
2.7.1. Gas chromatography technique (GC-FAME)
Well separated bacterial single colonies were identified by bacterial fatty acids extraction as
described by Sasser (1990). Single bacterial colonies were transferred from TSA 100% to
GC-TSB with a sterile plastic loop in 4 quadrants. The bacteria were scratched onto the GC-
TSB in 4 quadrants. The first and second quadrants were performed with the same loop and a
new sterile plastic loop was used to perform a third and fourth quadrant. Plates were
incubated for 24 h in the dark at 28 °C. After incubation, to harvest bacterial cells, a 4 mm
sterile loop was used to transfer the cells from the third quadrant to the bottom of culture
polypropylene tubes (13 X 100 mm) which were previously cleaned with hexane for 10 min

Chapter 2 General Materials and Methods
16
in a tube rotator. To release the fatty acids the cells were lysed by adding 1.0 ml of Reagent 1
to each tube containing cells. The tubes were closely with Teflon lined caps, vortexed briefly
and heated in a water bath for 5 min at 100 °C. After heating, the tubes were vigorously
vortexed for 10 s and returned to the water bath at 100 °C for 25 min and then cooled slowly
to room temperature.
To generate fatty acid methyl esters 2 ml of Reagent 2 were added to every sample. The tubes
were capped, briefly vortexed and heated for 10 min at 80 °C in a water bath. Subsequently
the tubes were cooled rapidly in cold tap water. To transfer the fatty acids from the aqueous
phase to an organic stage, each cooled sample received 1.25 ml of Reagent 3. After capping
the tubes were gently mixed on a tube rotator for 10 min. The tubes were uncapped and the
aqueous (lower) phase was discarded. The organic phase was washed by adding 3 ml of
Reagent 4 to every sample. The tubes were recapped and mixed for 5 minutes. About 2/3 of
the organic phase was transferred into a GC glass vial and stored at -20 °C until the analysis
by gas chromatography was performed.
The purified samples were identified by gas chromatographic analysis of bacterial fatty acids
methyl esters using the Gaschromatograph Hewlett Packard Co. (HP) 5890E Serie II Plus,
with electronic pressure programming. The chromatographer used a FED Split (Splitless-
Einlassblock) detector and a 25 m + 0,2 mm + 0,33 mm Kap Soile Ultra 2 column. The
identification program used was the Sherlock MIDI Identification system, Version 3.9, TSBA
(Microbial Identification System, Newark, Delaware, USA). The program conditions for
sample identification were; Injector A at 250 °C, Det Temp A at 300 °C, Aux E Press: 38,0
Psi and Inj A Press: 9,0 Psi. A calibration mix sample (commercial FAME calibration
mix:Reagent 3, 1:1, v:v) was used prior running samples and after every ten samples. GC-
FAME peaks were annotated by the microbial identification system software.
2.7.2. Molecular characterization and identification technique (16S rRNA)
To characterize and identify soil bacteria Restriction Fragment Length Polymorphism (RFLP)
patterns of the 16S rRNA gene and partial 16S rRNA sequence analysis was performed. For
the characterization and identification the following procedure was developed based on Chen
and James (2002) adapted to the conditions of the Security 1 (S1) laboratory of the INRES-
Phytomedizin University of Bonn, Germany.

Chapter 2 General Materials and Methods
17
2.7.2.1. Bacterial culture and preparation of compounds for the PCR Master Mix
Single colonies were grown for 48 h on TSA+ under dark conditions at 28 °C. Two different
primers were used to amplify the 16S rRNA genes by Polymerase Chain Reaction (PCR).
The forward primer was 27F (5’-AGAGTTTGATCCTGGCTCAG-3’) and the reverse primer
was the 9rev (5’-AAGGAGGTGATCCAGCC-3’) both obtained from Sigma.
The primers were originally dissolved in milliQ water to a concentration of 100 μM and were
diluted to a final concentration of 10 μM with milliQ water. The PCR buffer was prepared
mixing 5 X Green GoTaq Flexi Buffer (Promega) with Magnesium Chloride (MgCl2 at 25
miliMolar, Promega) in a 10:3, v:v ratio. Deoxynucleotide triphosphate (dNTP) at 2.5 mM
was prepared by adding 10 μL 100 mM dCTP (Promega), 10 μL 100 mM dATP (Promega),
10 μL 100 mM dGTP (Promega) and 10 μL 100 mM dTTP (Promega) to 1160 μL autoclaved
milliQ water on an autoclaved Eppendorf tube. Taq Polymerase (Promega) was the enzyme
used for PCR and was kept on ice prior adding it to the PCR Master Mix.
2.7.2.2. PCR Master Mix preparation
The PCR Master Mix was prepared in an autoclaved Eppendorf tube kept on ice, by mixing
10 μL 5 X PCR buffer, 4 μL dNTP (2.5 mM), 1 μL 27F (10 μM), 1 μL 9rev (10 μM), 0.25
μL Taq polymerase and 33.75 μL autoclaved MilliQ water for every bacterial sample.
From the TSA+ plates containing the growing bacteria a small colony was picked with a
sterile pipette tip and transferred to the bottom of an autoclaved plastic microfuge PCR vial.
To each PCR vial 50 μL of the PCR Master Mix was added. All microfuge vials were closed
properly with autoclaved plastic caps. Centrifugation for 2 s was performed in a
microcentrifuge (Labnet International) to bring all PCR Master Mix to the bottom of the
plastic vials where bacterial cells were located.
2.7.2.3. PCR procedure
The microfuge vials were placed on a PCR thermal cycler (T Gradient, Biometric).
Amplification was performed by an initial denaturation at 95 °C for 4 minutes, followed by
35 cycles of 95 °C for 1 minute, 50 °C for 1 minute and 72 °C for 1 minute, and with a final
extension cycle of 72 °C for 5 minutes. After the PCR procedure, samples were kept at 4 °C.

Chapter 2 General Materials and Methods
18
2.7.2.4. Gel electrophoresis analysis
After PCR the amplification was verified by agarose gel electrophoreses. The gel was
prepared with 1 X Tris-Acetate EDTA-Buffer (TAE, AppliChem). To every 100 ml 1 X
TAE, 1 g Agarose (Sigma) was added in an Erlenmeyer flask and heated for 5 min in a
microwave (MW800, Continent) at 650 Watts. After cooling at aprox. 50 °C every 100 ml of
this mixture received 1 μL of 10 mg ml-1 Ethidium Bromide (AppliChem). This solution was
poured into an electrophoresis tray and left in rest for 30 min until the gel had solidified. The
gel was subsequently transferred to the gel electrophoresis chamber filled with 1 X TAE
solution. From every PCR sample obtained, 5 μL were taken and transferred into the wells of
the agarose gel. In the first slot of the agarose gel the 1 kb DNA Ladder (Promega) was
loaded. After transferring all samples to the gel, the electrophoresis analysis was performed
for 60 min at 120 V and 400 mA. To visualize and analyze the DNA bands, the gel was
placed on ultraviolet transilluminator and a picture with a digital camera (S9500, Finepix,
Fujifilm) was taken. The remaining 45 μL of PCR product of each sample was kept on
refrigeration at 4 °C until further use.
2.7.2.5. Restriction enzyme analysis
In order to digest the bacterial DNA bands to detect different bacterial species it was
necessary to conduct a restriction enzyme analysis. The enzyme Cfo1 (Promega) was used to
prepare the Restriction Enzyme Master Mix (REMM) since it has been shown to be
informative for 16S RFLP analysis (Karpouzas et al., 2000). In a sterile Eppendorf tube the
REMM was prepared by adding 0.2 μL acetylated bovine serum albumin (BSA, Promega), 2
μL Buffer B (Promega) and 1 μL Cfo1 enzyme to 1.8 μL autoclaved distilled milliQ water for
each bacterial sample.
From the 45 μL PCR product left per sample 15 μL were taken and transferred to a new
autoclaved microfuge PCR plastic vial and mixed with 5 μL of REMM. The plastic vials
were closed, centrifuged for 2 s and incubated for 3 h at 37 °C. After incubation, the samples
were stored at 4 °C. To analyze the restriction enzyme analysis the procedure described in
2.7.2.4. for gel electrophoreses analysis was performed with two modifications. Firstly, the
time for electrophoresis was extended to a total of 120 min, and secondly, the DNA marker

Chapter 2 General Materials and Methods
19
used was the 50 bp ladder (Promega). The remaining PCR product was kept in refrigeration
and used later for purification.
2.7.2.6. DNA purification
The purification of the amplified DNA fragment was performed using the GFX PCR DNA
and Gel Band Purification Kit (Illustra, General Electric, Health Biosciences) following the
procedure described in the user manual. Briefly, in a sterile Eppendorf tube 150 μL Capture
Buffer type 2 were added to every 30 μL PCR product sample. The mixture was directly
pipette onto the filter units of the plastic vials of the purification kit and centrifuged at 14000
rpm for 30 s at 28 °C on a 5402-Eppendorf Centrifuge. The supernatant in the filter was
recovered and used further. 25 ml Buffer type 1 were mixed with 100 ml ethanol (analytical
grade) and 500 μL of this mixture was added to each sample on the filter and centrifuged as
previously described. The supernatant in the filter was used further. The filter containing the
DNA was transferred to a new Eppendorf tube. 50 μL Elutio-Buffer 4 was added to each
sample and incubated for 60 s at room temperature. Subsequently samples were centrifuged
for 60 s (14000 X G at 28 °C) and the DNA was eluted.
From every purified sample 2 µl were taken and mixed with 1 µl loading dye (Sigma) to
quantify the amount of purified DNA by gel electrophoresis. Agarose gel electrophoreses
analysis was performed as previously described in section 2.7.2.4. To calculate the amount of
DNA present in the samples the molecular marker Lambda DNA (Promega) digested with
Eco RI, Hind III, and BamH I was used. To prepare this marker every 100 μL of Lambda
DNA were mixed with 20 μL Buffer E (Promega), 3.3 μL Eco RI (Promega), 3.3 μL Hind III
(Promega), 3.3 μL BamH I (Promega) and 70 μL distilled milliQ water and incubated for 3 h
at 37 °C. To the digested Lambda DNA loading dye was added. To prepare the loading dye 4
mg Bromophenol blue were mixed with 1 ml milliQ water in an Eppendorf tube. 650 μL of
this mixture were transferred to 350 μL milliQ water in another Eppendorf tube and from this
dilution 250 μL were transferred to another clean Eppendorf tube. The same procedure was
performed with Xylene cyanol and the final 250 μL were mixed with the 250 μL obtained
previously with Bromophenol blue. The 500 μL dye solution was mixed with 300 μL of
glycerol and 200 μL milliQ water to obtain a final volume of 1000 μL loading dye. Every 4
μL of Lambda DNA (250 ng/μL) was mixed with 1 μL of the loading dye to obtain a final
concentration of 200 ng of DNA per μL.
The band intensity obtained with the bacterial samples was related to individual bands of the
marker.
Chapter 2 General Materials and Methods
20
2.7.2.7. DNA Sequencing and bacterial identification
Samples containing the bacterial DNA were sent to GATC Biotech AG (Konstanz, Germany)
and sequenced. Sequences were edited with the BioEdit Sequence Alignment Editor (Hall,
1999) and compared to the sequences of the National Center for Biotechnology Information
(http://www.ncbi.nlm.nih.gov/) to identify the bacterial isolate using blast analysis.
2.8. High pressure liquid chromatography (HPLC analysis)
To separate, identify and quantify nematicides High Pressure Liquid Chromatography
analysis (HPLC analysis) was performed. The HPLC analysis was performed on a
HEWLETT PACKARD (HP) system using a LiChrospher®100 C18 reversed phase column
(250 by 4.0 mm, 5 µm), preceded by a LiChrospher® C18 reversed phase guard column (4.0
by 4.0 mm, 5 µm). The HPLC was controlled by ChemStation for LC 3D systems and
consisted of a HP 1050 pump unit, HP 1050 diode array detector, HP 1046A fluorescence
array detector, and 1050 (717S) autosampler. Before samples were injected, the column had
been equilibrated with 90% (v/v) water and 0.1% (v/v) trifluoroacetic acid (TFA) (solvent A)
and 10% acetonitril (solvent B). After injection the samples were eluted at a flow rate of 1.0
ml/min using an isocratic flow of solvent A for 2 min, a linear gradient to 90% solvent B for
28 minutes, followed by a isocratic flow for 5 minutes with 90% solvent B. Before the next
sample was injected the column was re-equilibrated by a 1 minute linear gradient to 10 %
solvent B, followed by a 4 minute isocratic flow of 4 minutes. The retention time for
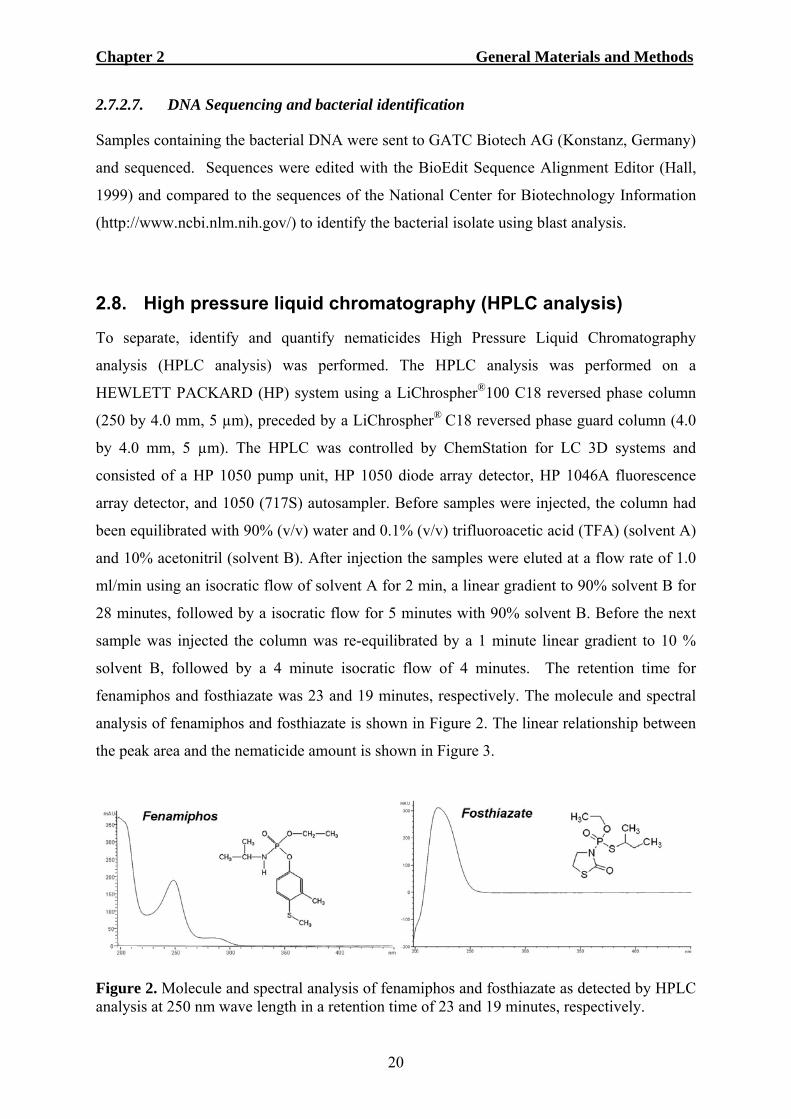
fenamiphos and fosthiazate was 23 and 19 minutes, respectively. The molecule and spectral
analysis of fenamiphos and fosthiazate is shown in Figure 2. The linear relationship between
the peak area and the nematicide amount is shown in Figure 3.
Figure 2. Molecule and spectral analysis of fenamiphos and fosthiazate as detected by HPLC analysis at 250 nm wave length in a retention time of 23 and 19 minutes, respectively.

Chapter 2 General Materials and Methods
21
y = 2306.287143x - 1.675238R2 = 0.999981
0
500
1000
1500
2000
2500
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Fenamiphos [μg]
Area
und
er th
e cu
rve
[mAu
*s]
Fenamiphos
Figure 3. Standard curve of fenamiphos and fosthiazate detected by HPLC analysis at 250 nm wave length.
2.8.1. Quantification of nematicides in soil extract liquid medium
To quantify the nematicides in soil extract liquid media, a 400 µL sample of each treatment
was mixed with 1600 µL methanol in an Eppendorf tube under laminar flow. This mixture
was filter-sterilized (0.4 µm) and transfer to a HPLC glass vial. The glass vials were capped
tightly. From each sample, 20 µL was injected onto the column for HPLC analysis. The
nematicide area under the peak was determined. Using the standard curve the amount of
nematicide in the sample was determined.
2.8.2. Extraction and quantification of nematicides in soil extract agar medium
Under laminar flow, 5 cm diameter Petri dishes containing soil extract agar amended with
nematicides were opened. A triangular shaped sample was taken with a sterile scalpel from
an area near the bacterial colony, transferred to an Eppendorf tube and weighted. For every
0.1 g of agar 100 µL of methanol was added and incubated over night. Subsequently, the
tubes containing the agar-methanol mixture were centrifuged for 5 min at 14000 X G at 24
°C. After centrifugation, 200 µL of the solution were transferred to HPLC plastic vials and
caped tightly. From each sample, 20 µL was injected onto the column and the remaining
nematicide was detected by retention time and spectral pattern. The amount of nematicide
was calculated using the standard curve obtained with the pure compound.
y = 25.553x - 0.8484R2 = 0.9979
020406080
100120140160180200
0 1 2 3 4 5 6 7
Fosthiazate [μg]
Area
und
er th
e cu
rve
[mAu
*s]
Fosthiazate

Chapter 2 General Materials and Methods
22
2.9. References
Chen, B.Y. and James, H.W. 2002. Methods in Molecular Biology - PCR Cloning protocols. 2nd
edition. Humana Press Inc.Totowa, New Jersey, USA. p 439.
Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT. Nucleic Acids Symposium Series 41:95-98.
Hussey, R. and Barker, K. 1973. A comparison of methods of collecting inocula of
Meloidogyne spp., including a new technique. Plant Disease Reporter 57:1025-1028.
Karpouzas, D.G. Morgan, J.A. and Walker, A. 2000. Isolation and characterization of
ethoprophos-degrading bacteria. FEMS Microbiology Ecology 33:209-218.
Sasser, M. 1990. Identification of bacteria by gas chromatography of cellular fatty acids.
Technical Note 101. Newark, DE:MIDI.
Serrano, E., Sandoval, J., Pocasangre, L.E., Rosales, F. and Delgado, E. 2006. The importance
of physical-chemical indicators in the soil quality for the sustainable production of
banana in Costa Rica. In: Soprano, E., Adami, F., Lichtemberg, L.A. and Silva, M.C.
editors. Proceedings of the 17th ACORBAT International meeting; Banana: A
sustainable business, October 15-20, 2006, Joinville, Santa Catarina, Brasil. 1:207-221.
Speijer, P.R. and De Waele, D. 1997. Screening of Musa Germplasm for resistance and
tolerance to nematodes. INIBAP Technical Guidelines. Montpellier, France. p.48.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
23
3. FIELD EVIDENCE OF TERBUFOS ENHANCED BIODEGRADATION IN BANANA CULTIVATION
3.1. Introduction
The burrowing nematode, Radopholus similis (Cobb 1893) Thorne 1949, is the most common
and damaging nematode in banana cultivation worldwide (Araya, 1995; Araya et al., 1995;
Araya and Moens, 2005; Gowen et al., 2005). This nematode is responsible for restricted root
growth and diminished bunch weight, leading to overall decreases in yield (Frison et al.,
1998; Moens and Araya, 2002; Araya, 2003; Araya and Moens, 2005). Consequently, R.
similis root infestations are correlated to a decline in banana yield (Sarah et al., 1996; Gowen
et al., 2005; Cabrera et al., 2006). Because of the lack of effective alternative management
tools in this perennial crop, the control of R. similis in commercial plantations is usually
based on up to three applications of non-fumigant nematicides on a yearly basis (Araya,
2003). One of the most commonly used nematicides currently being applied is the
organophosphate terbufos. However, after terbufos application, root decline associated with
high nematode densities were observed, suggesting that the efficacy of the nematicide is
declining, which is most likely caused by an elevated nematicide degradation process in the
soil.
Accelerated degradation of the nematicides fenamiphos, carbofuran and ethoprophos has
been shown to occur in soils with a history of repeated exposure to these specific types of
nematicides (Ou, 1991; Ou et al., 1994; Smelt et al., 1996; Karpouzas et al., 1999; Karpouzas
and Walker, 2000). In soils treated with aldicarb, oxamyl and fenamiphos a specific
microflora established which was capable of rapidly degrading the active nematicidal
components (Ou and Rao, 1986; Jones and Estes, 1995). For example, the fungi Bjerkandera
adusta, Pleurotus ostreatus and Phanerochaete chrysosporium have been shown to degrade
50 to 96% of terbufos after four days of exposure (Jauregui et al., 2003). The performance of
terbufos in differently treated soils was reported in maize cultivation in relation to the corm
root worm (Little et al., 1992). However, little is known about the biodegradation of terbufos
in soils cultivated with banana where other agroecological conditions and use of pesticides
are encountered.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
24
In the commercial cultivation of Musa, the use of tissue-culture bananas instead of
transferring the follower production suckers is an attractive alternative when fields have to be
replanted (Quénéhervé, 1993). The in-vitro propagated bananas are propagated under sterile
laboratory conditions and, consequently, these healthy plants, generally promise higher
yields, when compared to suckers, which acquired almost always nematodes and other pests
and diseases during their development in the field. The growth of the delicate in-vitro
propagated plants could be affected by the repeated application of non-fumigant nematicides.
Previous reports in other crops have shown a phytotoxic side effect caused by terbufos
treatments (Sinclair et al., 1992; Kennedy, 2002). Therefore, it is of importance to the banana
production industry to elucidate whether or not terbufos applications negatively affect the
quality and functioning of the banana root system.
Knowledge about the level of both nematicide efficacy and biodegradation of a pesticide or
pesticide group is important for the grower in order to develop management strategies that
can effectively reduce crop losses caused by plant-parasitic nematodes. The current lack of
knowledge with respect to these ongoing processes are leading to an increase in numbers of
nematicide applications, thus increasing the production costs and the chemical pressure on
human health and the environment. Therefore, the aims of this study were to:
1. Determine the presence of terbufos biodegradation in soils infested with R. similis
from banana plantations in Costa Rica.
2. Establish the effect of the nematicide application history on the current level of
nematicide biodegradation.
3. Investigate whether terbufos has phytotoxicity side-effects on banana root
development.
4. Elucidate the plant parasitic nematode diversity in banana roots.
3.2. Materials and Methods
3.2.1. Soil and root collection
Soil samples were collected from four commercial banana plantations (Musa AAA) of the
province of Limon along the Atlantic coast of Costa Rica. The soil samples of these
plantations will be named F1, F2, F3 and F4 hereafter. The samples were taken from
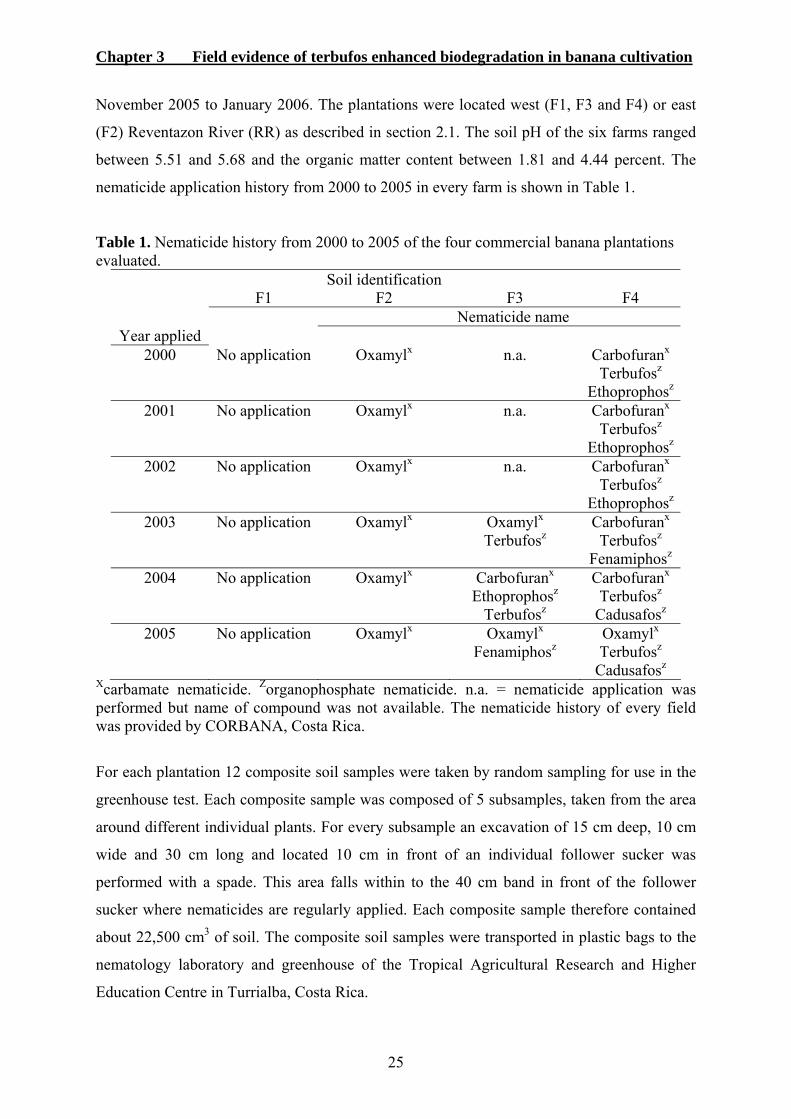
Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
25
November 2005 to January 2006. The plantations were located west (F1, F3 and F4) or east
(F2) Reventazon River (RR) as described in section 2.1. The soil pH of the six farms ranged
between 5.51 and 5.68 and the organic matter content between 1.81 and 4.44 percent. The
nematicide application history from 2000 to 2005 in every farm is shown in Table 1.
Table 1. Nematicide history from 2000 to 2005 of the four commercial banana plantations evaluated.
Soil identification F1 F2 F3 F4 Nematicide name Year applied
2000 No application Oxamylx n.a. Carbofuranx Terbufosz Ethoprophosz
2001 No application Oxamylx n.a. Carbofuranx Terbufosz Ethoprophosz
2002 No application Oxamylx n.a. Carbofuranx Terbufosz Ethoprophosz
2003 No application Oxamylx Oxamylx Carbofuranx Terbufosz Terbufosz Fenamiphosz
2004 No application Oxamylx Carbofuranx Carbofuranx Ethoprophosz Terbufosz Terbufosz Cadusafosz
2005 No application Oxamylx Oxamylx Oxamylx Fenamiphosz Terbufosz Cadusafosz
Xcarbamate nematicide. Zorganophosphate nematicide. n.a. = nematicide application was performed but name of compound was not available. The nematicide history of every field was provided by CORBANA, Costa Rica.
For each plantation 12 composite soil samples were taken by random sampling for use in the
greenhouse test. Each composite sample was composed of 5 subsamples, taken from the area
around different individual plants. For every subsample an excavation of 15 cm deep, 10 cm
wide and 30 cm long and located 10 cm in front of an individual follower sucker was
performed with a spade. This area falls within to the 40 cm band in front of the follower
sucker where nematicides are regularly applied. Each composite sample therefore contained
about 22,500 cm3 of soil. The composite soil samples were transported in plastic bags to the
nematology laboratory and greenhouse of the Tropical Agricultural Research and Higher
Education Centre in Turrialba, Costa Rica.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
26
In every field 12 composite root samples were taken to extract the plant-parasitic nematodes.
Roots were taken from follower production suckers performing a similar excavation as the
one used for soil sampling. The follower production suckers for extracting root samples were
different from those used for soil sampling. The follower suckers to be sampled were selected
according to their mother plant stage by using the method described by Speijer and De Waele
(1997).
3.2.2. Determination of terbufos biodegradability and side-effect
To determine the presence of enhanced biodegradability of terbufos and its effect on root
growth a modification of the green house bioassay described by Pattison et al. (2000) was
used. All 12 composite soil samples from the same plantation received one of the following
three treatments; i) non-autoclaved and no nematicide treated control, ii) non-autoclaved and
nematicide treated, or iii) autoclaved and nematicide treated. Soil autoclaving was performed
for 60 minutes twice at 121 °C over two consecutive days. One-hundred µg of Terbufos 10
GR per gram of soil was applied once to the respective treatments at the beginning of the
experiment (day 0). The nematicide granules were applied to a plastic bag containing the soil
and mixed thoroughly. From each treatment six sub-samples were taken at 0, 15, 30, 45 and
60 days after nematicide application. Each sub-sample was composed of 60 g soil and was
transferred to a 10 cm diameter plastic pot. One in-vitro propagated banana (Musa AAA) cv.
Grande Naine, 15 to 20 cm long from the first leaf to the crown, was planted in each pot at
the time the soil-subsamples were taken. After planting, treatments were arranged in a
randomized block design at the greenhouse. After nematicide application, all soil treatments
were stored in the dark at room temperature.
Nematodes were inoculated at the same moment when the six soil-subsamples per treatment
were taken. Per pot, a suspension containing 500 mixed stages of R. similis was inoculated
into three holes 3 cm deep previously made with a 1 ml pipette tip around the banana stem.
The plants were carefully watered daily as required with tap water. Seven days after
nematode inoculation, the banana plants were harvested. The roots were gently rinsed free of
soil with tap water and root fresh weigh was measured. Nematode extraction from the roots
was performed as described by Araya (2002) and quantified visually, using a microscope.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
27
3.2.3. Determination of nematode diversity in banana roots
Every composite sample was gently washed and dead roots were discarded. Plant parasitic
nematodes were extracted from 25 g of functional roots, which were macerated for 10 s at
high speed followed by 5 s at low speed in a commercial blender containing 300 ml tap
water. The macerated solution was filtered using nested sieves with apertures of 250 µm, 106
µm and 0.25 µm, respectively. Nematodes recovered on the 0.25 µm sieve were transferred to
a glass beaker and the volume adjusted to 200 ml using tap water. Of each sample, 4 ml was
taken and all nematodes encountered in the first 2 ml were counted and identified. For
identification the key described by Hunt et al. (2005) was used.
3.2.4 Statistical analysis
To conform to assumptions of normally distributed data, nematode numbers were
transformed using a log(x+1) transformation before statistical analysis was performed. One-
way analysis of variance (ANOVA) was performed for each set of data separated. One set of
data corresponded to one evaluation date of the five dates possible. Nematode numbers and
root fresh weight were analyzed in the same way. Fisher’s Least Significant Difference
(LSD) was performed where significant differences (P < 0.05) were detected. The statistical
software SPSS 12.0 was used to perform the analysis.
3.3. Results
3.3.1 Biodegradability of terbufos
The number of recovered nematodes from roots in the non-treated control soil differed
significantly (p < 0.05) from roots which were treated with nematicides from the plantation
F1 (Figure 1A). In this soil a high level of nematode control was obtained from 15 days after
nematicide treatment up to 60 days. In the bioassays where soil from F2 and F3 were used,
nematode numbers in banana roots were significantly lower in both nematicide treatments
when compared to the non-treated control during all the evaluations period (Figures 1B and
1C, respectively). The number of nematodes recovered from the banana roots in the non-
treated control soil from F4 were not significantly different from the non-autoclaved terbufos
treated soil 0, 15, 30 and 60 days after nematicide treatment (Figure 1D). Forty five days after
nematicide application there was no significant difference between the non-autoclaved
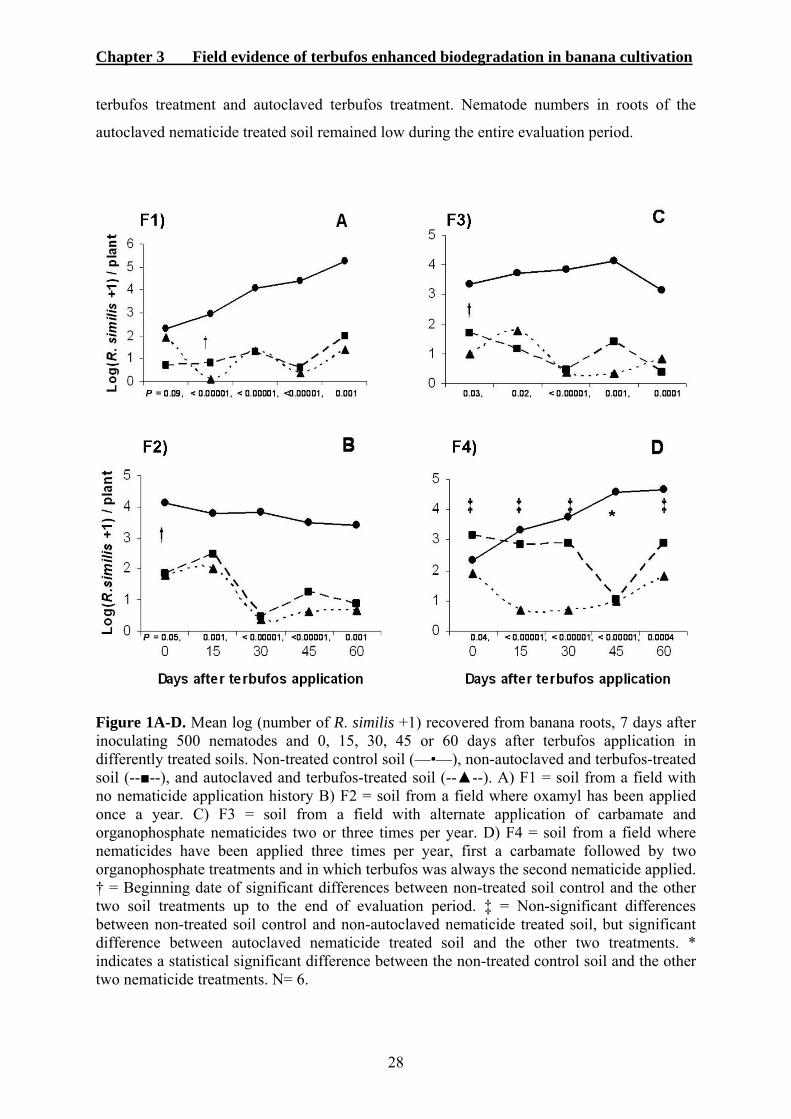
Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
28
terbufos treatment and autoclaved terbufos treatment. Nematode numbers in roots of the
autoclaved nematicide treated soil remained low during the entire evaluation period.
Figure 1A-D. Mean log (number of R. similis +1) recovered from banana roots, 7 days after inoculating 500 nematodes and 0, 15, 30, 45 or 60 days after terbufos application in differently treated soils. Non-treated control soil (—•—), non-autoclaved and terbufos-treated soil (--■--), and autoclaved and terbufos-treated soil (--▲--). A) F1 = soil from a field with no nematicide application history B) F2 = soil from a field where oxamyl has been applied once a year. C) F3 = soil from a field with alternate application of carbamate and organophosphate nematicides two or three times per year. D) F4 = soil from a field where nematicides have been applied three times per year, first a carbamate followed by two organophosphate treatments and in which terbufos was always the second nematicide applied. † = Beginning date of significant differences between non-treated soil control and the other two soil treatments up to the end of evaluation period. ‡ = Non-significant differences between non-treated soil control and non-autoclaved nematicide treated soil, but significant difference between autoclaved nematicide treated soil and the other two treatments. * indicates a statistical significant difference between the non-treated control soil and the other two nematicide treatments. N= 6.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
29
3.3.2. Terbufos side-effect
The terbufos application did not significantly affect root fresh weight of the in-vitro
propagated bananas, cv. Garnde Naine, with three exceptions (Table 2). The banana root
weight in the tests using F1 and F4 were significantly higher in autoclaved and nematicide
treated soil than in the non-treated control, but not significantly different from the nematicide
treated soil 45 days after terbufos application. Similar results occurred in tests using F2,
evaluated thirty days after terbufos application. In all these cases a positive effect was found,
resulting in an increase in root weight when compared to roots extracted from the non-treated
soils. Banana roots in the tests with F3 were not significantly different at any time of
evaluation.
Table 2. Comparison of root fresh weight (g) of in-vitro propagated Musa (AAA) cv. Grande Naine after 7 days planted in different terbufos treated and untreated soils with different nematicide history.
Soil identification
F1 F2 F3 F4
Days after terbufos application
0 15 30 45 60 0 15 30 45 60 0 15 30 45 60 0 15 30 45 60
Root fresh weight [g]
Soil treatment
Not-treated soil 1.60 2.08 3.43 1.85b 2.52 3.50 4.05 4.20b 4.63 2.67 4.10 4.20 4.40 4.47 1.63 1.52 1.58 3.25 1.70b 2.52 Nematicide treated soil 1.85 1.88 2.95 2.08ab 2.88 2.82 4.05 5.05ab 4.80 2.02 4.02 3.48 3.80 4.70 1.68 1.43 1.68 2.95 2.48a 2.30 Autoclaved and nematicide treated soil 1.67 1.85 2.95 2.38a 2.77 3.10 4.58 5.93a 4.68 2.43 3.93 3.77 5.22 4.60 2.07 1.95 1.75 4.25 2.32a 2.68 Fisher’s LSD n.s. n.s. n.s. 0.43 n.s. n.s. n.s. 0.90 n.s. n.s. n.s. n.s. n.s. n.s. n.s. n.s. n.s. n.s. 0.44 n.s.
P 0.67 0.40 0.65 0.05 0.32 0.15 0.50 0.01 0.87 0.24 0.97 0.45 0.32 0.98 0.33 0.97 0.80 0.25 0.004 0.84
Numbers in same column with different letter are significantly different. n.s. = not significantly different. A) F1 = soil from a field with no nematicide application history. F2 = soil from a field where oxamyl has been applied once a year. F3 = soil from a field with alternate application of carbamate and organophosphate nematicides two or three times per year. F4 = soil from a field where nematicides have been applied three times per year, first a carbamate followed by two organophosphate treatments and in which terbufos was always the second nematicide applied. N = 6.
3.3.3. Nematode diversity in banana roots
Radopholus similis was the most abundant plant-parasitic nematode in banana roots,
representing over 70% of the total population in all four fields evaluated (Figure 2).
Helicotylenchus spp. was present in all fields been the 18, 30, 2 and 5% in the farms F1, F2,
F3 and F4, respectively. Pratylenchus spp. was present only in F2 conforming 1% of the

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
30
total nematode population. Meloidogyne incognita was found only in F4 with a total
population of 8%.
0%10%20%30%40%50%60%70%80%90%
100%
F1 F2 F3 F4
Soil name
Perc
enta
ge o
f the
tota
l pla
nt
para
sitic
nem
atod
es
Meloidogyne spp.
Pratylenchus spp.
Helycotylenchus spp.
R. similis
Figure 2. Plant-parasitic nematode diversity in roots of follower production suckers Musa AAA taken in four different commercial plantations of Costa Rica. F1 = soil from a field with no nematicide application history. F2 = soil from a field where oxamyl has been applied once a year. F3 = soil from a field with alternate application of carbamate and organophosphate nematicides two or three times per year. F4 = soil from a field where nematicides have been applied three times per year, first a carbamate followed by two organophosphate treatments and in which terbufos was always the second nematicide applied. N = 12.
3.4. Discussion
The bioassay indicated that terbufos was rapidly degraded in F4, the soil sample from the
plantations where this nematicide was applied on a yearly basis. Since terbufos applied to the
non-autoclaved treatment lost effectiveness at all evaluation times, except when the soil had
been autoclaved in advance, microbial activity must have been responsible for this
degradation process. Similar results in a comparable bioassay were obtained from soil
frequently treated with Furadan®, Mocap®, Nemacur® and Vydate® (Moens et al., 2004).
Our results are in agreement with the statements by Smelt and Leistra (1992), who concluded
that nematicides are vulnerable to accelerated degradation by adapted microorganisms,
whenever regular and frequent applications of the same compound occur and Stirling et al.
(1992), who postulated that repeated exposure to a particular pesticide can lead to enhanced
microbial degradation. Furthermore, indigenous bacteria and a microbial mat were shown to
be responsible for mineralization of carbofuran in the soils of banana plantations in St.
Vincent, in the West Indies (Murray et al., 1997). In banana plantations of the Ivory Coast
where fenamiphos was applied every 4 months, a step-by-step build up of microbial

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
31
populations was detected which was responsible for the highly accelerated degradation of the
nematicide (Anderson and Lafuerza, 1992). In laboratory tests and in commercial banana
plantations, treatment of soils every 4 months with 3 grams of active ingredient per plant
caused the gradual build-up of bacteria and fungi which could degrade fenamiphos
(Anderson, 1989). The results in the present investigation proved that terbufos is no
exception to the biodegradation rule.
In the present study microbial degradation of terbufos was absent in the nematicide treated
and non-autoclaved soils from F1, F2 and F3. The effectiveness of the nematicide in these
soils was stable for the entire 60 day evaluation period in the autoclaved and the non-
autoclaved treatments. Previous to the present research, the F1 soil had never been treated
with a nematicide. The F2 soil was treated once a year with oxamyl, a carbamate nematicide.
Soil F3 had only been treated previously twice with terbufos, but one year had passed since
the last application. These results suggested that the low level of nematicide used in these
fields prevented or delayed the selection of microorganisms responsible for the
biodegradation of terbufos. Similar results were obtained by Pattison et al. (2000) who found
high efficacy of fenamiphos against R. similis for up to 8 weeks after nematicide treatment on
a sandy loam soil with no previous nematicide use history as compared to increased
biodegradation after 2 weeks on a silty loam soil with 15 previous fenamiphos treatments.
Additionally, in one field cultivated with banana where alternative nematicide applications
were used Moens et al. (2004) did not find enhanced biodegradation of Counter® after 5
consecutively applications.
The reduction in the number of nematodes in F4 in the non-autoclaved terbufos treatment 45
days after nematicide application was inconsistent with the preceding and successive
sampling dates. This suggests that an experimental error at treatment establishment or during
nematode inoculation had occurred. However, the data obtained in the four other sampling
dates were consistent, thus supporting the conclusion that biodegradation is a very important
factor in reducing terbufos efficacy.
Terbufos had no negative side effects on banana root growth. Under green house conditions
terbufos reduced seedling emergence to pearl millet (Kennedy, 2002), diminished carrot
seedlings emergence, decreased onion and carrot seedling size and delayed emergence of
both crops (Sinclair et al., 1992). In contrast to the above results, terbufos treatment to

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
32
autoclaved soil in the present study increased root weight of banana plants on three
occasions. This effect was found 45 days after terbufos application in soils F1 and F4, and 30
days after nematicide treatment in F2. On these dates R. similis was highly controlled in the
autoclaved soils with a terbufos application history. This suggests that high invasion of the
nematodes negatively affected root weight in case the nematicide was not applied in the
bioassay. The root weight of in-vitro propagated bananas planted in virgin soils with 5
consecutive applications of Counter® was higher than from non-treated soil (Araya and
Lakhi, 2004). Moens and Araya (2002) also found that R. similis reduced root weight by 66%
when compared to non-nematode inoculated controls, supporting the results obtained in the
present work. However, the lack of root phytotoxicity in banana could be related to the
limited exposure time of 7 days, the amount of active ingredient, and the size of the plant at
treatment time. Terbufos probably did not negatively affect root development in our assays
because of the resilient root system of the banana plant. Therefore phytotoxic effects in the
field are not to be expected.
All banana plants grew for 7 days in the treated soils. However there were differences in
growth between evaluation periods, such as 60 days after terbufos treatment in F2 and F3,
where a decrease in the overall root weight could be observed. The original bioassay
described by Pattison et al. (2000) uses corn seeds, which grow more uniformly than the
young in-vitro propagated bananas used in this study. The variation in growth may thus be
due to a lack of uniformity of the in-vitro plants used for each set of experiments. These
plants were received from different banana production batches and differed in size. However
for each period of evaluation all banana plants came from the same batch, thus making a
comparison among them is possible.
Radopholus similis was found in all banana fields, accounting over 70% of the total plant
parasitic nematode population in roots. Comparable high R. similis densities were found by
Araya et al. (1995) in the same Atlantic region of Costa Rica. Both studies confirm the
presence of economical important populations of the burrowing nematode in this area. This
work reveals that even with several chemical treatments on a year basis, R. similis continues
to be the main nematode problem in banana cultivation, as it has been for decades.
Furthermore, Menjivar (2005) showed that R. similis numbers increased in the banana roots,
even when non-fumigant nematicides were applied in the field. Consequently, research that is
focussed on controlling this nematode will remain of prime importance.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
33
Terbufos should only be used in an integrated nematode management program to retain high
efficacy, which will be lost with frequent application as demonstrated in this study.
Application of nematicides having different active ingredients can reduce and in some cases
prevent the onset of increased biodegradation (Anderson and Lafuerza, 1992 ). Such chemical
rotations are, however, becoming difficult to maintain, due to the use-restriction of many
nematicides that have been on the market for a long time and the lack of availability
regarding new compounds that can be used for nematode control in banana production.
Increasing the time interval between chemical applications, e.g. by utilizing alternative
biocontrol agents (Pocasangre et al., 2000; Mendoza et al. 2004/2008; zum Felde et al., 2006)
could reduce the emergence of enhanced microbial nematicide degradation through
sustaining a non-biased and diverse microbial population over time and thus support the
maintenance of nematicide efficacy.
3.5. References
Anderson, J.P.E. 1989. Accelerated microbial degradation of nematicides and other plant
protection chemicals in soils. Proceedings of the 20th annual meeting of OTAN, 7-11
November, 1988, San Jose, Costa Rica. Nematropica 19:1.
Anderson, J.P.E. and Lafuerza, A. 1992. Microbiological aspects of accelerated pesticide
degradation. Proceedings of the international symposium on environmental aspects of
pesticide microbiology, 17-21 August 1992, Sigtua, Sweden. 184-192.
Araya, M. 1995. Efecto depresivo de ataques de Radopholus similis en banano (Musa AAA).
CORBANA 20:3-6.
Araya, M. 2002. Metodología utilizada en el laboratorio de nematología de CORBANA S.A.
para la extracción de nematodos de las raices de banano (Musa AAA) y plátano (Musa
AAB). CORBANA 28:97-110.
Araya, M. 2003. Situación actual del manejo de nematodos en banano (Musa AAA) y plátano
(Musa AAB) en el trópico americano. In: Rivas G. and Rosales F.E. editors. Manejo
convencional y alternativo de la Sigatoka negra, nematodos y otras plagas asociadas al
cultivo de Musaceas en los tropicos. INIBAP, Montpellier, France . 79-102.
Araya, M., Centeno, M. and Carillo, W. 1995. Densidades poblacionales y frecuencia de los
nematodos parásitos del banano (Musa AAA) en nueve cantones de Costa Rica.
CORBANA 20:6-11.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
34
Araya, M. and Lakhi, A. 2004. Response to consecutive nematicide applications using the
same product in Musa AAA cv. Grande Naine originated from in vitro propagative
material and cultivated in virgin soil. Nematologia Brasileira 28:55-61.
Araya, M. and Moens, T. 2005. Parasitic nematodes on Musa AAA (Cavendish subgroups cvs
Grande naine, Valery and Williams. In: Turner DW, Rosales FE, editors. Banana root
system; towards a better understanding for its productive management. Proceedings of
an International Symposium, 3-5 November 2003, San José, Costa Rica. 201-223.
Cabrera, J.A., Pocasangre, L.E., Rosales, F., Najera, J., Vargas, R., Serrano, E. and Sikora,
R.A. 2006. Relation between Radopholus similis (Cobb) Thorne in follower suckers
and yield decline of mother plants in commercial banana plantations (Musa AAA)
with different yield structures. Nematropica 36:111-112.
Frison, E.C., Gold, C.S., Karamura, E.B. and Sikora, R.A. 1998. Mobilizing IPM for
sustainable banana production in Africa. Proceedings of a workshop on banana IPM,
23-28 November 1998, Nelspruit, South Africa. INIBAP, Montpellier, France. 356p.
Gowen, S.R., Quénéhérvé, P. and Fogain, R. 2005. Nematodes parasites of bananas and
plantains. In: Luc M., Sikora R.A. and Bridge J. editors. Plant parasitic nematodes in
tropical and subtropical agriculture, second edition, Wallingford: CAB International.
611-643.
Hunt, D.J., Luc, M. and Manzanilla-López, R.H. 2005. Identification, morphology and
biology of plant parasitic nematodes. In: Luc M., Sikora R.A. and Bridge J. editors.
Plant parasitic nematodes in tropical and subtropical agriculture, second edition,
Wallingford: CAB International. pp. 11-52.
Jauregui, J., Valderrama, B., Albores, A. and Vazquez-Duhalt R. 2003. Microsomal
transformation of organophosphorus pesticides by white rot fungi. Biodegradation
16:397-406.
Jones, R.L. and Estes, T.L. 1995. Summary of aldicarb monitoring and research programs in
the United States. Journal of Contaminant Hydrology 18:107-140.
Karpouzas, D.G., Walker, A., Froud-Williams, J.R. and Drennan, D. 1999. Evidence for the
enhanced biodegradation of ethoprophos and carbofuran in soils from Greece and the
UK. Pesticide Science 55:301-311.
Karpouzas, D.G. and Walker, A. 2000. Aspects of the enhanced biodegradation and
metabolism of ethoprophos in soil. Pest Management Science 56:540-548.
Kennedy, C.W. 2002. Phytotoxicity in pearl millet varies among in-furrow insecticides. Crop
Protection 21:799-802.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
35
Little, R.J., Devine, J.M., Tenne, F.D., Walgenbach, P.J. and Belcher DW. 1992. Performance
of terbufos on corn-rootworm (Coleoptera, Chrysomelidae) in the corn belt. Journal of
Economic Entomology 85:1413-1424.
Mendoza, A.R., Sikora, R.A. and Kiewnick, S. 2004. Efficacy of Paecilomyces lilacinus
(strain 251) for the control of Radopholus similis in banana. Communications in
Agricultural and Applied Biological Sciences 69:365-372.
Mendoza, A.R., Sikora, R.A. and Kiewnick, S. 2007. Influence of Paecilomyces lilacinus
strain 251 on the biological control of the burrowing nematode Radopholus similis in
banana. Nematropica 37:203-213.
Menjivar, R.D. 2005. Estudio del potencial antagonista de hongos endofiticos para el
biocontrol del nematodo barrenador Radopholus similis en plantaciones de banano en
Costa Rica. MSc Thesis, CATIE, Turrialba, Costa Rica. 65 p.
Moens, T. and Araya, M. 2002. Efecto de Radopholus similis, Meloidogyne incognita,
Pratylenchus coffeae y Helicotylenchulus multicinctus en la producción de Musa AAA
cv. Grande Naine. CORBANA 28:43-56.
Moens, T., Araya, M., Swennen, R. and De Waele, D. 2004. Enhanced biodegradation of
nematicides after repetitive applications and its effect on root and yield parameters in
commercial banana plantations. Biology and Fertility Soils 39:407-414.
Murray, R., Phillips, P. and Bender, J. 1997. Degradation of pesticides applied to banana farm
soil: comparison of indigenous bacteria and a microbial mat. Environmental
Toxicology and Chemistry 16:84-90.
Ou, L.T. 1991. Interactions of microorganisms and soil during fenamiphos degradation. Soil
Science Society of America Journal 55:716-722.
Ou, L.T. and Rao, P.S.C. 1986. Degradation and metabolism of oxamyl and phenamiphos in
soils. Journal of Environmental Sciences and Health B 21:25-40.
Ou, L.T., Thomas, J.E. and Dickson, D.W. 1994. Degradation of fenamiphos in soil with a
history of continuous fenamiphos applications. Soil Science Society of America
Journal 58:1139-1147.
Pattison, A.B., Stanton, J.M. and Cobon, J.A. 2000. Bioassay for enhanced biodegradation of
nematicides in soil. Australasian Plant Pathology 29:52-58.
Pocasangre, L.E., Sikora, R.A., Vilich, V. and Shuster, R.P. 2000. Survey of banana
endophytic fungi from Central America and screening for biological control of
Radopholus similis. Acta Horticulturae 531:283-289.

Chapter 3 Field evidence of terbufos enhanced biodegradation in banana cultivation
36
Quénéhervé, P. 1993. Nematode management in intensive banana agrosystems: comments
and outlook from the Cote d'Ivoire experience. Crop Protection 12:164-172.
Sarah, J.L., Pinochet, J. and Stanton, J. 1996. The burrowing nematode of bananas,
Radopholus similis Cobb, 1913. Musa Pest Fact Sheet No. 1. INIBAP, Montpellier,
France.
Sinclair, P.J., Neeson, R.J. and Williams, P.A. 1992. Phytotoxicity of some organophosphate
insecticides to onions and carrots during germination and emergence. Plant Protection
Quarterly 7:23-25.
Smelt, J.H. and Leistra, M. 1992. Availability, movement and accelerated transformation of
soil-applied nematicides. In: Gommers FJ, Maas PW, editors. Nematology from
molecule to ecosystem. European Society of Nematologist, Inc. Inwergrowie, Dundee,
Scotland. 266-280.
Smelt, J.H., Van de Peppel-Groen, A., Vand der Pas, L. and Dijksterhuis, A. 1996.
Development and duration of accelerated degradation of nematicides in different soils.
Soil Biology and Biochemestry 28:1757-1765.
Speijer, P.R. and De Waele, D. 1997. Screening of Musa Germplasm for resistance and
tolerance to nematodes. INIBAP Technical Guidelines, Montpellier, France.
Stirling, A.M., Stirling, G.R. and Macrae, I.C. 1992. Microbial degradation of fenamiphos
after repeated application to tomato growing soil. Nematologica 38:245-254.
zum Felde, A., Pocasangre, L.E., Cañizares-Monteros, C.A., Sikora, R.A., Rosales, F.E. and
Riveros, A.S. 2006. Effect of combined inoculations of endophytic fungi on the
biological control of Radopholus similis. Infomusa 15:12-18.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
37
4. Involvement of microorganisms other than Pseudomonads
on fenamiphos enhanced degradation
4.1. Introduction
Fenamiphos is a broad spectrum, non-volatile, systemic, organophosphorus nematicide
extensively used throughout the world to control plant-parasitic nematodes in fruit and
vegetable crops (Stirling et al., 1992). The activity tends to be nematostatic rather than
nematotoxic, inhibiting movement of nematodes and delaying egg hatching (Van Gundy and
Mckenry, 1977).
A result of continuous or repeated exposure to a particular pesticide, microbial populations in
the soil are capable of developing degradative traits, which leads to an accelerated
degradation of the active components as shown in the previous chapter of this thesis. Such
events have also been demonstrated in tomato cultivation, in which the soil was frequently
treated with fenamiphos (Stirling et al., 1992).
Several microorganisms such as fungi, yeasts and bacteria are capable to produce surface-
active molecules, known as biosurfactants, which can facilitate the uptake of an insoluble
substrate (Fiechter, 1992; De Souza, 2002). Biosurfactant producing bacteria have, for
example, been shown to rapidly degrade xenobiotics (Johnsen et al., 1996). Among the
bacteria genera, Pseudomonas sp. was reported to produce biosurfactants (Fiechter, 1992;
Ron and Rosenber, 2001; De Souza, 2002). The genus Pseudomonas is a common inhabitant
of the rhizosphere and has received considerable attention in bioremediation of xenobiotics
(De Souza, 2002). Arino et al. (1996) and Deziel et al. (1996) have shown the implications of
surfactant-producing Pseudomonas in facilitating degradation of pollutants such as polycyclic
aromatic hydrocarbons and n-alkanes.
To date, however, little is known about the influence of Pseudomonas species on the
enhanced biodegradation of non-fumigant nematicides and its importance on supplying a key
emulsifier to the soil for starting the rapid microbial degradation process. For these reasons,
the objectives of the research in this chapter were to:

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
38
1. Determine the Pseudomonas population and growth in soil amended with different
concentrations of organic matter.
2. Investigate the enhanced biodegradation of fenamiphos in soil with high densities
of Pseudomonas spp. after several consecutive fenamiphos treatments.
3. Establish the production of biosurfactants by Pseudomonas spp.
4. Study the presence of Pseudomonas spp. in the degrading microorganisms of
fenamiphos.
4.2. Materials and Methods
4.2.1. Effect of compost on total soil bacteria and Pseudomonas spp.
Soil 4, which had no nematicide treatment history, was used to determine the Pseudomonas
population density and evaluate the effect of the addition of different compost concentrations.
One day prior to the addition of compost, all culturable soil bacteria and Pseudomonas spp.
were extracted and cultivated on TSA+ and KBM+, respectively, as described in section 2.4.
After adding 1, 4 or 7% (w/w) green compost, the soil was thoroughly mixed by hand to
obtain a homogenized mixture. Tap water was added to increase the moisture content and the
soil was mixed again. Soil bacteria were isolated 1 and 3 weeks after compost amendment.
S4 with no addition of compost served as control. Each treatment had 4 replicates.
4.2.2. Efficacy of fenamiphos after three consecutive treatments
Soil 4-composte mixtures were split into three subsamples and each subsample was (i) not
treated any further (control), (ii) treated with fenamiphos or (iii) autoclaved and subsequently
treated with fenamiphos. Autoclaving was performed twice for 60 min at 121 °C in two
consecutive days. In case of nematicide treatment, 10 µg a.i. of Nemacur 5GR per gram of
soil was applied three times with 6 weeks interval. To all samples, water was added to field
capacity after each nematicide treatment. After the first nematicide treatment, four 250 ml
soil samples were transferred to plastic pots from each soil every two weeks until 6 weeks
after nematicide application. Three holes were made per pot in the soil using a plastic 1 ml
pipette tip and a suspension containing 2500 M. incongita eggs was inoculated. Then, 50
seeds of lettuce cv. Milan were sown per pot and covered with a thin layer of autoclaved sand
to enhance germination. Lettuce plants were regularly irrigated with tap water as needed and

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
39
30 days after sowing the seeds the lettuce plants were harvested. The roots were gently
washed with tap water and percentage of galled roots was determined. Efficacy of Nemacur
5GR was calculated according to the formula described by Abbott (1925).
4.2.3. Analysis of biosurfactant production
4.2.3.1. Drop collapse test
A modification of the drop collapse test described by Jain et al. (1991) was performed to
evaluate biosurfactant production capacity of bacteria. Therefore, 100 bacterial single
colonies were isolated from S4 amended with 7% compost and grown on KBM+ plates. A full
loop of each colony was transferred into autoclaved distilled water in a 2 ml Eppendorf tube.
The tube with the bacterial suspension was vortexed vigorously for 15 s and 20 µL of the
bacterial suspension were carefully pipetted on Parafilm paper (Parafilm M, Laboratory Film,
Pechiney, Plastic Packaging, Chicago, IL). Collapse of the drop was evaluated up to one
minute after application on the Parafilm and compared to the drops of autoclaved distilled
water, autoclaved distilled water mixed with the biosurfanctant producer, Pseudomonas
SS101 (positive control), and autoclaved distilled water mixed with Pseudomonas 10.24
(negative control) which does not produce the biosurfactant.
4.2.3.2. Blue media test
Since biosurfactant production may be media dependent, a second analysis was performed to
verify biosurfactant production. Therefore, every single bacterial colony used in the drop
collapse test was further grown on KBM+ for 48 h at 28 °C in the dark. Subsequently a full
loop containing the bacterial colony was transferred onto ‘blue media’ (Siegmund and
Wagner, 1991). Petri dishes were incubated at 28 °C in the dark and the halo formation was
evaluated after 5 days.
4.2.4. Identification of Pseudomonas spp.
A total of 16 well separated randomly selected bacterial single colonies, from the 100 isolates
tested for biosurfactant production, were identified to verify whether the bacteria growing on
KBM+ were Pseudomonas species. Every isolate was transferred from KBM+ onto CG-TSB
with a sterile 4 mm loop under laminar flow. After 48 h of incubation in the dark at 28 °C,
the bacteria were identified by GC-FAME analysis as described in section 2.7.1.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
40
4.2.5. Fenamiphos metabolization and presence of Pseudomonads in the degrading microorganisms
To investigate the role of Pseudomonas spp. on the biodegradation of fenamiphos, two
different tests were performed. In the first test, two of the GC-FAME identified soil bacteria
were analyzed for their ability to metabolize fenamiphos. One full loop containing
Pseudomonas syringae or P. putida was inoculated on a 10 cm diameter plate containing
SEAM supplemented with 100 or 200 μg of fenamiphos ml-1. As control a fenamiphos-
SEAM plate was incubated without bacteria. The plates were sealed with Parafilm and all
treatments were incubated for 10 days in the dark at 28 °C. Then, one triangular shaped agar
block was transferred to a reaction tube using a sterile scalpel. Fenamiphos extraction and
quantification was performed by HPLC analysis as described in section 2.8. Each treatment
had 4 replicates. Analysis of Variance (ANOVA, P < 0.05) was performed to compare the
amounts of remaining fenamiphos in the plates. For statistical analysis the program GenStat
Discovery Edition 3 was used.
In the second test, a mixture of all the different microorganisms present in S1, S2, S3 or S4
were used. To obtain these microorganisms, 10 g of each soil was transferred to 100 ml of
autoclaved distilled water and incubated in the dark for 16 h at 28 °C on an orbital shaker set
at 120 rpm. Then, 1 ml of the suspension was transferred to 19 ml soil extract liquid medium
supplemented with fenamiphos (analytical grade) to a final concentration of 100 µg ml-1 in a
115 X 28 mm sterile plastic tube (Sarstedt). SELM treated with fenamiphos without bacterial
inoculation served as control. All plastic tubes were tightly capped, placed vertically in an
orbital shaker set at 60 rpm and incubated in the dark at 28 °C. After 0, 1, 6, 10 and 14 days
of incubation, a 400 µl sample of each treatment was transferred and mixed with 1600 µL of
methanol. This mixture was filter-sterilized (0.4 µm) and analyzed by HPLC as described
section 2.8. Each treatment was done in triplicate. The Standard Error of the Mean was
calculated for every treatment.
In addition, to investigate the total soil bacterial populations and verify the presence of
Pseudomonads in the fenamiphos-degrading microorganisms, 50 µl suspension of each of the
four German soils was plated on TSA+ or KBM+ 14 days after nematicide treatment. The
plates were incubated in the dark for 48 h at 28 °C and subsequently the number of CFU was
determined. Every treatment was repeated six times.
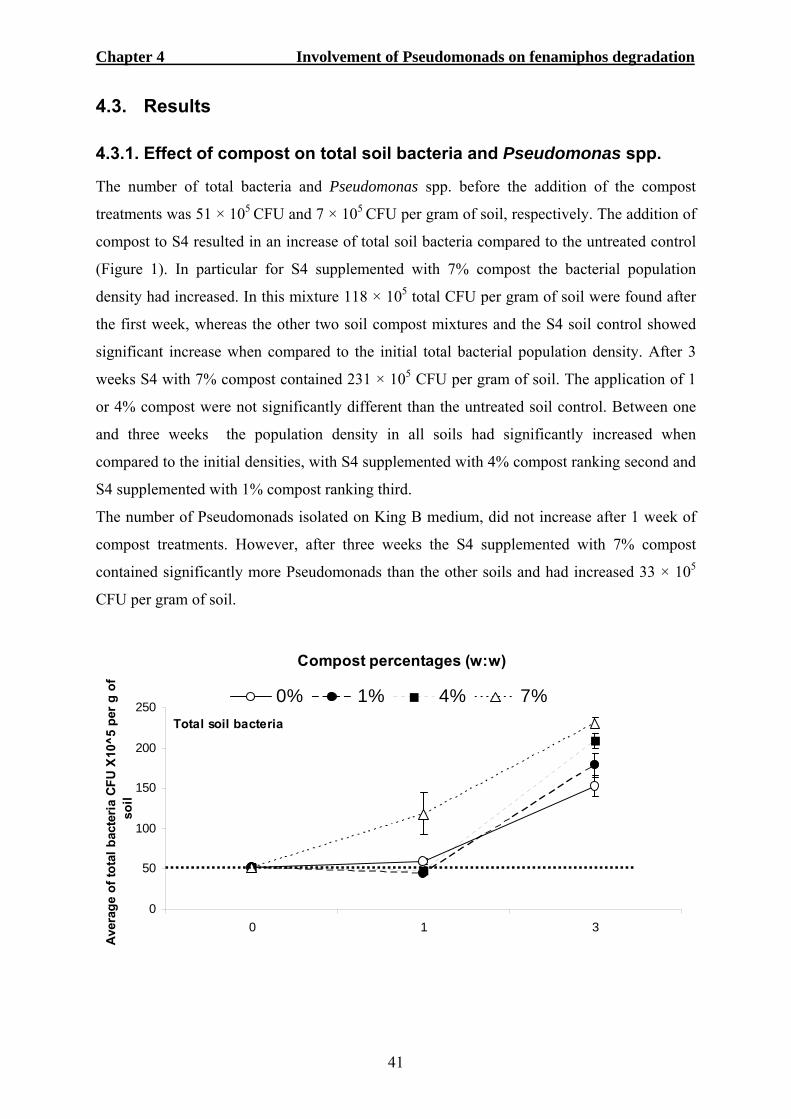
Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
41
4.3. Results
4.3.1. Effect of compost on total soil bacteria and Pseudomonas spp.
The number of total bacteria and Pseudomonas spp. before the addition of the compost
treatments was 51 × 105 CFU and 7 × 105 CFU per gram of soil, respectively. The addition of
compost to S4 resulted in an increase of total soil bacteria compared to the untreated control
(Figure 1). In particular for S4 supplemented with 7% compost the bacterial population
density had increased. In this mixture 118 × 105 total CFU per gram of soil were found after
the first week, whereas the other two soil compost mixtures and the S4 soil control showed
significant increase when compared to the initial total bacterial population density. After 3
weeks S4 with 7% compost contained 231 × 105 CFU per gram of soil. The application of 1
or 4% compost were not significantly different than the untreated soil control. Between one
and three weeks the population density in all soils had significantly increased when
compared to the initial densities, with S4 supplemented with 4% compost ranking second and
S4 supplemented with 1% compost ranking third.
The number of Pseudomonads isolated on King B medium, did not increase after 1 week of
compost treatments. However, after three weeks the S4 supplemented with 7% compost
contained significantly more Pseudomonads than the other soils and had increased 33 × 105
CFU per gram of soil.
0
50
100
150
200
250
0 1 3
Ave
rage
of t
otal
bac
teria
CFU
X10
^5 p
er g
of
soil
0% 1% 4% 7%
Compost percentages (w:w)
Total soil bacteria
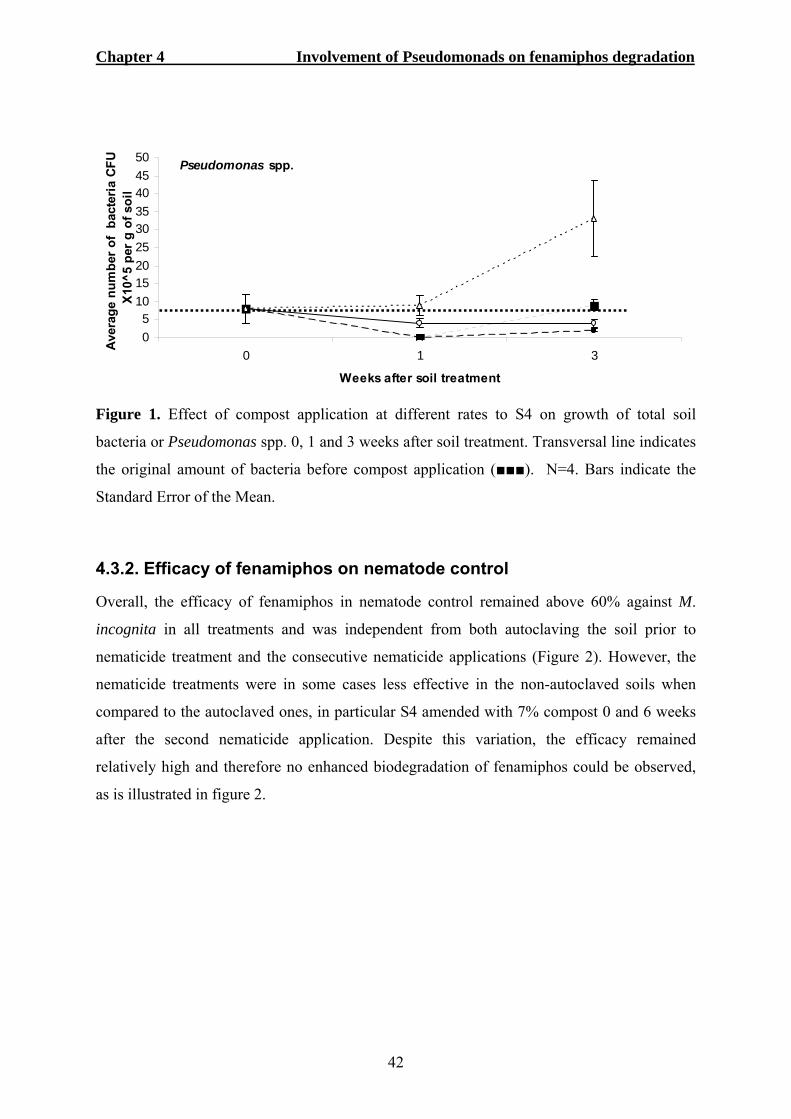
Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
42
05
101520253035404550
0 1 3
Weeks after soil treatment
Aver
age
num
ber o
f ba
cter
ia C
FU
X10^
5 pe
r g o
f soi
l
Pseudomonas spp.
Figure 1. Effect of compost application at different rates to S4 on growth of total soil
bacteria or Pseudomonas spp. 0, 1 and 3 weeks after soil treatment. Transversal line indicates
the original amount of bacteria before compost application (■■■). N=4. Bars indicate the
Standard Error of the Mean.
4.3.2. Efficacy of fenamiphos on nematode control
Overall, the efficacy of fenamiphos in nematode control remained above 60% against M.
incognita in all treatments and was independent from both autoclaving the soil prior to
nematicide treatment and the consecutive nematicide applications (Figure 2). However, the
nematicide treatments were in some cases less effective in the non-autoclaved soils when
compared to the autoclaved ones, in particular S4 amended with 7% compost 0 and 6 weeks
after the second nematicide application. Despite this variation, the efficacy remained
relatively high and therefore no enhanced biodegradation of fenamiphos could be observed,
as is illustrated in figure 2.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
43
0102030405060708090
100
0 2 4 6
Effic
acy
of N
emac
ur [%
]After 1 nematicide application
0102030405060708090
100
0 2 4 6
Effi
cacy
of N
emac
ur [%
]
After 2 nematicide applications
0102030405060708090
100
0 2 4 6
Weeks after nematicide treatment
Effic
acy
of N
emac
ur [%
]
After 3 nematicide applications
Figure 2. Efficacy of Nemacur GR on root galling of lettuce cv. Milan 0, 2, 4 and 6 weeks after one, two or three consecutive nematicide treatments. Autoclaved (----) or non-autoclaved (—) soil amended with 1% (●), 4% (▲) or 7% (■) compost. N = 4. Bars indicate the Standard Error of the Mean.
4.3.3. Biosurfactant production tests
One hundred single colonies isolated from S4 and grown on KBM+ were tested for
biosurfactant production. Besides the positive biosurfactant control Pseudomonas SS101,
none of the one hundred single colonies was able to collapse the water drop on the Parafilm
paper (Figure 3).

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
44
C
BA
C
BA
Figure 3. Drop collapse test on Parafilm paper after 1 minute of placing the drops. Drop caused by autoclaved distilled water alone (A). Drop caused by the mixture of autoclaved distilled water and the negative biosurfactant producing control Pseudomonas 10.24 (B). Drop collapse caused by the mixture of autoclaved distilled water and the biosurfactant producing control Pseudomonas SS101 (C). The rest of the drops were caused by the mixture of individual bacterial single colonies isolated on KBM+ from S4, a soil with no fenamiphos application history, and autoclaved distilled water.
The same one hundred bacterial isolates used in the drop collapse test were further tested on
‘blue media’ for biosurfactant production. During the 5 days of incubation none of the
isolates was able to produce a halo, confirming that all bacterial single colonies were not
capable of producing biosurfactants (Figure 4).
Figure 4. Plate with ‘blue medium’ containing 13 bacterial single colonies isolated on KBM+
from S4, a soil with no fenamiphos application history, with no halo formation after 5 days of incubation.
4.3.4. Identification of single bacterial colonies
From the 100 isolates that were tested for biosurfactant production grown on King B
medium, 16% were identified by their fatty acid composition. GC-FAME analysis

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
45
demonstrated that all isolates belonged to the genus Pseudomonas. Fifty percent of the
isolates were identified as Pseudomonas putida, 31% corresponded to P. fluorescens, 13%
were identified as P. syringae and 6% were P. chlororaphis (Figure 5).
31%
13%
50%
6%
Pseudomonas fluorescens Pseudomonas syringaePseudomonas putida Pseudomonas chlororaphis
Figure 5. Diversity of different non-biosurfactant Pseudomonas isolated on KBM+ from S4, a soil with no fenamiphos application history, amended with 7% compost and identified by GC-FAME.
4.3.5. Pseudomonads and the conversion of fenamiphos
In the first in-vitro nematicide assay, individual bacteria were tested for their fenamiphos
metabolizing abilities on culture plates supplemented with 100 or 200 µg µl-1. No significant
differences (P < 0.05) were observed with respect to the remaining amounts of fenamiphos
between SEAM inoculated with bacteria and the not inoculated control for both
concentrations 10 days after incubation (Figure 6). The final fenamiphos concentration in
agar medium with initially applied at 100 μg ml-1 fenamiphos was of 99 and 98 μg ml-1 when
inoculated with Pseudomonas syringae and P. putida, respectively, whereas the non-
inoculated control had reached 99 μg ml-1. When the initial fenamiphos concentration was of
200 μg ml-1 the final concentrations were 198, 193 and 200 μg ml-1, respectively. Thus, both
Pseudomonads were not capable of converting fenamiphos over a period of 10 days.
In the second assay, all microorganisms were tested together for the ability to convert
fenamiphos in liquid cultures. HPLC analysis showed that in the presence of microorganisms,
extracted from the soils S2 and S3, fenamiphos was completely degraded after incubating for
14 or 6 days, respectively (Figure 7). In the same assay the microorganisms from the soils S1
and S4 were not able to convert fenamiphos, even after incubating for 14 days. In S1 and S2
no Pseudomonads were found (Figure 8). In contrast, in S3 and S4 17 × 104 and 32 × 104

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
46
CFU of Pseudomonas were found, respectively. Thus, the results obtained from soil S2
suggest that bacterial genera, other than Pseudomonas are capable of degrading fenamiphos.
0
50
100
150
200
250
Pseudomonas syringae Pseudomonas putida No bacteria present
Fena
mip
hos
conc
entra
tion
in S
EAM
(μ
g/m
l)
100 µg/ml 200 µg/ml
A A A
a a a
Figure 6. Fenamiphos concentrations in soil extract agar medium (SEAM) after 10 days of incubation which was initially applied at 100 or 200 μg ml-1 and inoculated with Pseudomonas syringae, P. putida or not inoculated. These bacteria were isolated on KBM+
from S4, a soil with no fenamiphos application history, and were not able to produce surfactants. N = 4. Bars indicate the Standard Error of the Mean.
0102030405060708090
100
0 1 6 10 14
Number of days of incubation after nematicide treatment
Fena
mip
hos
conc
entra
tion
(%) c
ompa
red
to tr
eatm
ent
with
no
soil
mic
roor
gani
sms
S1 S2 S3 S4
Figure 7. Metabolization of fenamiphos by microorganisms obtained from the German soils S1, S2, S3 and S4, in samples taken 0, 1, 6, 10 or 14 days of incubation after nematicide treatment. Different than soils S1, S2, and S3, soil S4 had no fenamiphos application history. N = 3. Bars indicate the Standard Error of the Mean.
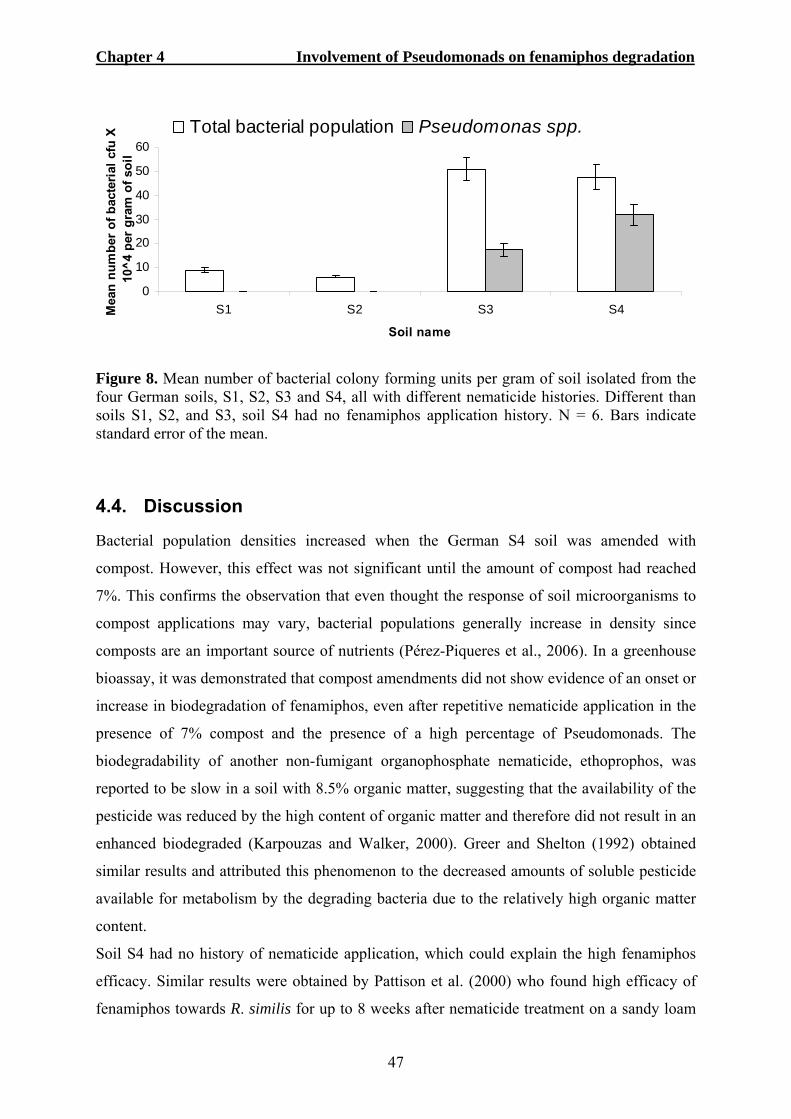
Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
47
0
10
20
30
40
50
60
S1 S2 S3 S4
Soil name
Mea
n nu
mbe
r of
bac
teria
l cfu
X
10^4
per
gra
m o
f soi
lTotal bacterial population Pseudomonas spp.
Figure 8. Mean number of bacterial colony forming units per gram of soil isolated from the four German soils, S1, S2, S3 and S4, all with different nematicide histories. Different than soils S1, S2, and S3, soil S4 had no fenamiphos application history. N = 6. Bars indicate standard error of the mean.
4.4. Discussion
Bacterial population densities increased when the German S4 soil was amended with
compost. However, this effect was not significant until the amount of compost had reached
7%. This confirms the observation that even thought the response of soil microorganisms to
compost applications may vary, bacterial populations generally increase in density since
composts are an important source of nutrients (Pérez-Piqueres et al., 2006). In a greenhouse
bioassay, it was demonstrated that compost amendments did not show evidence of an onset or
increase in biodegradation of fenamiphos, even after repetitive nematicide application in the
presence of 7% compost and the presence of a high percentage of Pseudomonads. The
biodegradability of another non-fumigant organophosphate nematicide, ethoprophos, was
reported to be slow in a soil with 8.5% organic matter, suggesting that the availability of the
pesticide was reduced by the high content of organic matter and therefore did not result in an
enhanced biodegraded (Karpouzas and Walker, 2000). Greer and Shelton (1992) obtained
similar results and attributed this phenomenon to the decreased amounts of soluble pesticide
available for metabolism by the degrading bacteria due to the relatively high organic matter
content.
Soil S4 had no history of nematicide application, which could explain the high fenamiphos
efficacy. Similar results were obtained by Pattison et al. (2000) who found high efficacy of
fenamiphos towards R. similis for up to 8 weeks after nematicide treatment on a sandy loam

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
48
soil with no previous nematicide application history. Pesticide pre-treatment history is a main
factor need to detect the expression of the biodegradation processes of non-fumigant
nematicides (Karpouzas et al., 1999). Apparently a prolonged application over a substantial
period is essential for the soil microorganisms to adapt and start degrading a nematicide.
Smelt and Leistra (1992) stated that the most commonly used nematicides are all vulnerable
to conversion by adapted microorganisms, which arise by frequent applications. However, the
rate at which the microorganisms adapt to the nematicide cannot be predicted. This
unpredictability is illustrated by the present findings on the fenamiphos biodegradation by
microorganisms from soils with different fenamiphos application history. In the soil with the
longest fenamiphos application history, S1, no fenamiphos biodegradation could be observed,
whereas, the other two soils with a less fenamiphos application history, S2 and S3, did
contain biodegradation activity.
It is difficult to explain why microorganisms of S1 and S4 were not able to degrade
fenamiphos. It may be important to consider that S1 had the lowest pH of all soils (4.0),
which is regarded unfavourable for bacterial growth (Smelt et al., 1996). This could be the
reason why, even with a significant fenamiphos application history, the microorganisms
living in this soil were not able to transform into biodegrading isolates. It has been previously
demonstrated that soils with high pH are more likely to adapt and build-up large numbers of
bacteria that quickly degrade nematicides when compared to soils with a low pH (Smelt et
al., 1996). Suett et al. (1996) found that carbofuran degrading microorganisms were present
in all soils previously treated with the nematicide, but that the degrading microbes were
predominant in soils with the highest pH. More specific, Singh et al. (2003) found that
biodegradation of fenamiphos developed mainly in soils with high pH and a slow degradation
process developed in soil with a pH value of less than 5. Based on pH, soil S4 was optimal to
build-up to high populations of nematicide biodegrading isolates. It maybe that further
nematicide treatments are required to build up such microbial populations. However, next to
the pH, other biotic and abiotic factors may also play an important role, like the competition
or collaboration with other microorganisms, the availability of minerals and the composition
of root exudates. Soil S4 contained the highest percentage of Pseudomonads, of which
Pseudomonas putida was the most dominant species, representing half of all isolates
characterized. The other three species identified were P. fluorescens, P. syringe and P.
chlororaphis. All these four Pseudomonas species have been previously reported to degrade
organophosphate and/or carbamate non-fumigant nematicides. For example, Pseudomonas

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
49
putida, P. fluorescens and P. syringae were capable of degrading carbofuran (Karpouzas et
al., 2000b). P. putida was responsible for ethoprophos degradation (Karpouzas et al., 2000a).
In addition, P. chlororaphis has shown to degrade 1,2,3,4-tetrachlorobenzene (Portawfke et
al., 1998). As demonstrated by the biotest, none of the Pseudomonas species in S4 were
capable of degrading and this capability did not developed with fenamiphos application.
Also, none of the individually tested Pseudomonas isolates did produced surfactants, which
are surface-active molecules that may facilitate the uptake of an insoluble substrate (Fiechter,
1992) and/or may facilitate degradation of certain chemicals by emulsifying them (Guerra-
Santos et al., 1984; Zhang and Miller, 1992/1994). Biosurfactant producing Pseudomonads,
are known to be present in the rhizosphere at relatively low densities, ranging between 5 to
14% of the total Pseudomonads population (De Souza et al., 2003). Furthermore not all soils
contain biosurfactant Pseudomonads at detectable levels (Raaijmakers et al., 1997). The lack
of surfactant producing Pseudomonads present in S4 may make this soil intrinsically
incapable of developing a fenamiphos degrading microbial population.
According to HPLC analysis in the first metabolization test, the two bacterial isolates P.
syringae and P. putida were not able to degrade fenamiphos. The absence of degrading
capability could thus be related to the lack of biosurfactant production. Furthermore, the
results obtained in this investigation can also be related to the lack of degradation ability of
Pseudomonas when it is grown in the absence of a suitable helper bacterium. In a previous
study with a bacterial consortium where Pseudomonas spp. was present, a rapid degradation
of fenamiphos occurred (Singh et al., 2003). However, the Pseudomonads themselves were
not able to degrade fenamiphos without the presence of two other bacteria of the genus
Flavobacterium and Caulobacter. Perhaps Pseudomonas strains that can not produce
surfactants can still profit from the surfactant production of other bacteria. Flavobacterium,
for example, is known to produce surfactants supporting this theory (Bodour et al.,
2003/2004).
Overall, the exact role of neither the Pseudomonads nor the surfactants in the facilitation of
fenamiphos degradation could be demonstrated in the present studies. Further
characterization of the Pseudomonas population, the nematicide biodegradation ability and
the presence of surfactant producing isolates in S3, a soil with demonstrated fenamiphos
degrading abilities and containing Pseudomonads, may reveal more information on this
matter.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
50
Nevertheless, the results obtained with S2, a soil with no detectable numbers of
Pseudomonads, do demonstrate that microorganisms other than Pseudomonads can be
involved in the efficient degradation of fenamiphos. Until now, few microorganisms have
been identified that can degrade fenamiphos, e.g. Brevibacterium sp. MM1 (Megharaj et al.,
2003). Perhaps soils S2 and S3 contain significant numbers of this type of bacteria resulting
in efficient nematicide degradation. Furthermore, since in this study all microorganisms of
the soil were tested, other microorganisms than bacteria may be involved in the degradation
of fenamiphos. Fungi, for example, have also been reported to be involved in the fenamiphos
degradation process (Anderson, 1989).
In conclusion, this study demonstrated that a prolonged nematicide application does not
necessarily lead to an increase in biodegradation of the active compound and that
microorganisms other that Pseudomonads can be important factors in the in the degradation
of the nematicides. These microorganisms are the focus of the next chapter.
4.5. References
Abbott, W.S. 1925. A method of computing the effectiveness of an insecticide. Journal of
Economic Entomology 18:265-267.
Anderson, J.P.E. 1989. Accelerated microbial degradation of nematicides and other plant
protection chemicals in soils. Proceedings of the 20th annual meeting of OTAN, 7-11
November, 1988, San Jose, Costa Rica. Nematropica 19:1.
Arino, S., Marchal, R. and Vandecasteele, J.P. 1996. Identification and production of a
rhamnolipid biosurfactant by Pseudomonas species. Applied Microbiology
Biotechnology 45:162-168.
Bodour, A.A., Drees, K.P. and Maier, R.M. 2003. Distribution of biosurfactant-producing
bacteria in undisturbed and contaminated arid Southwestern soils. Applied and
Environmental Microbiology 69:3280-3287.
Bodour, A.A., Guerrero-Barajas, C., Jiorle, B.V., Malcomson, M.E., Paull, A.K., Somogyi,
A., Trinh, L.N., Bates, R.B. and Maier, R.M. 2004. Structure and characterization of
flavolipids, a novel class of biosurfactants produced by Flavobacterium sp. strain
MTN11. Applied and Environmental Microbiology 70:114-120.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
51
De Souza, J.T. 2002. Distribution, diversity and activity of antibiotic-producing Pseudomonas
spp. PhD Thesis. Wageningen University, The Netherland. p.161.
De Souza J.T., Weller, D.M. and Raaijmakers, J.M. 2003. Frequency, diversity and activity of
2,4-diacetylphloroglucinol-producing Fluorescent Pseudomonas spp. in Dutch Take-
all decline soils. Phytopathology 93:54-62.
Deziel, E., Paquette, G., Villemur, R., Lepine, F. and Bisaillon, J.G. 1996. Biosurfactant
production by a soil Pseudomonas strain growing on polycyclic aromatic
hydrocarbons. Applied Environmental Microbiology 62:1908-1912.
Fiechter, A. 1992. Biosurfactants: moving towards industrial application. Trends in
Biotechnology 10:208-217.
Guerra-Santos, L., Käppeli, O. and Fiechter, A. 1984. Pseudomonas aeruginosa biosurfactant
production in continuous culture with glucose as carbon source. Applied and
Environmental Microbiology 48:301-305.
Greer, L.E. and Shelton, D.R. 1992. Effect of inoculant strain and organic matter content on
kinetics of 2,4-dichlorophenoxyacetic acid degradation in soil. Applied and
Environmental Microbiology 58:1459-1465.
Jain, D.K., Collins-Thompson, D.L., Lee, H., and Trevors, J.T. 1991. A drop-collapsing test
for screening surfactant-producing microorganisms. Journal of Microbiological
Methods 13:271-279.
Johnsen, K., Andersen, S., and Jacobsen, C.S. 1996. Phenotypic and genotypic
characterization of phenanthrene-degrading fluorescent Pseudomonas biovars.
Applied and Environmental Microbiology 62:3818-3825.
Karpouzas, D.G. and Walker, A. 2000. Factors influencing the ability of Pseudomonas putida
epI to degrade ethoprophos in soil. Soil Biology and Biochemistry 32:1753-1762.
Karpouzas, D.G., Walker, A., Foud-Williams, R. J. and Drennan, D.S.H. 1999. Evidence for
the enhanced biodegradation of ethoprophos and carbofuran in soils from Greece and
the UK. Pesticide Science 55:1089:1094.
Karpouzas, D.G., Alun, J., Morgan, J.A.W. and Walker, A. 2000a. Isolation and
characterisation of ethoprophos-degrading bacteria. FEMS Microbiology Ecology
33:209-218.
Karpouzas, D.G., Morgan, J.A.W. and Walker, A. 2000b. Isolation and characterization of 23
carbofuran-degrading bacteria from soils from distant geographical areas. Letters in
Applied Microbiology 31:353-358.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
52
Megharaj, M., Singh, N., Kookana, R.S., Naidu, R. and Sethunathan, N. 2003. Hydrolysis of
fenamiphos and its oxidation products by a soil bacterium in pure culture, soil and
water. Applied Microbiology and Biotechnology 61:252–256.
Pattison A.B., Stanton J.M. and Cobon J.A. 2000. Bioassay for enhanced biodegradation of
nematicides in soil. Australasian Plant Pathology 29:52-58.
Pérez-Piqueres, V.E.H., Alabouvette, C. and Steinberg, C. 2006. Response of soil microbial
communities to compost amendments. Soil Biology and Biochemistry 38:460:470.
Portawfke, T., Timmis, K.N. and Wittich R.M. 1998. Degradation of 1,2,3,4-
Tetrachlorobenzene by Pseudomonas chlororaphis RW71. Applied and
Environmental Microbiology 64:3798-3806.
Raaijmakers, M.M., Weller, D.M. and Thomashow, L.S. 1997. Frequency of antibiotic-
producing Pseudomonas spp. in natural environments. Applied and Environmental
Microbiology 63:881-887.
Ron, E.Z. and Rosenberg, E. 2001. Natural roles of biosurfactants. Environmental
Microbiology 3:229-236.
Siegmund, I., and Wagner, F. 1991. New method for detecting rhamnolipids excreted by
Pseudomonas species during growth on mineral agar. Biotechnology Technology
5:265-268.
Singh, B.K., Walker, A., Morgan, J.A.W. and Wright, D. 2003. Role of soil pH in the
development of enhanced biodegradation of fenamiphos. Applied and Environmental
Microbiology 69:7035-7043.
Smelt, J.H. and Leistra, M. 1992. Availability, movement and accelerated transformation of
soil-applied nematicides. In: Nematology from molecule to ecosystem. Gommers, F.J.
and Maas, P.W. (eds). European Society of Nematologist, Inc. Inwergrowie, Dundee,
Scotland. 266-280.
Smelt, J.H., Van de Peppel-Groen, A.E., Van der Pas, L.J.T. and Dijksterhuis, A. 1996.
Development and duration of accelerated degradation of nematicides in different soils.
Soil Biology and Biochemistry 28:1757-1765.
Stirling, A.M., Stirling, G.R. and Macrae, I.C. 1992. Microbial degradation of fenamiphos
after repeated application to tomato growing soil. Nematologica 38:245-254.
Suett, D.L., Jukes, A.A. and Parekh, N.R. 1996. Non-specific influence of pH on microbial
adaptation and insecticide efficacy in previously-treated field soils. Soil Biology and
Biochemistry 28:1783-1790.

Chapter 4 Involvement of Pseudomonads on fenamiphos degradation
53
Van Gundy, S.D. and Mckenry, M.V. 1977. Action of nematicides. In: Plant disease: An
advanced treatise. Horsfall, J.D. and Cowling, E.B. editors. Academic Press, New
York, USA. 263-283.
Zhang, Y. and Miller, R.M. 1992. Enhanced octadecane dispersion and biodegradation by a
Pseudomonas rhamnolipid surfactant (biosurfactant). Applied and Environmental
Microbiology 58:3276-3282.
Zhang, Y. and Miller, R.M. 1994. Effect of a Pseudomonas rhamnolipid biosurfactant on cell
hydrophobicity and biodegradation of octadecane. Applied and Environmental
Microbiology 60:2101-2106.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
54
5. Isolation and characterization of fenamiphos-degrading
microorganisms
5.1. Introduction
Microbial breakdown has been reported for several soil-applied herbicides, fungicides,
insecticides (Somasundram et al., 1989) and, in particular, for a large number of different
non-fumigant nematicides (Somasundram et al., 1989; Pattison et al., 2000; Mclean and
Lawrence, 2003; Moens et al., 2004). Fenamiphos, an organophosphate non-fumigant
nematicide, can be degraded by soil microorganisms into non-nematoxic compounds
(Johnson, 1998). Several microorganisms, either in pure or mixed culture, have been isolated
from soils exhibiting the biodegradation of organophosphorus nematicides, like
Flavobacterium spp., Penicillium waksmani Zaleski, Trichoderma viride Persoon,
Pseudomonas alcaligenes and Pseudomonas putida (Sethunthan and Yoshida, 1973; Rao and
Sethunathan, 1974; Sheela and Pai, 1983; Matsumura and Boush, 1986; Karpouzas et al.,
2000a).
In most cases, degrading microorganisms have been characterized using standard
microbiological taxonomic techniques, which are based on morphological and biochemical
characteristics (Ramanand et al., 1988). More recently, molecular based methods have been
added to the process of characterization (Feng et al., 1997). Information about the DNA
sequence of certain genomic regions is particularly valuable since there are extensive
sequence databases available, which can be used for taxonomic purposes. In this way,
Karpouzas et al. (2000b) molecularly characterized 23 carbofuran-degrading bacteria from
soils collected in UK and Greece.
Repetitive fenamiphos treatments accumulated bacteria and fungi capable of degrading the
active compound (Anderson, 1989; Anderson and Lafuerza, 1992). However, the
microorganisms involved in the degradation process were not identified. The degradation of
fenamiphos seems to be a complex process, since it was not possible to isolate one
microorganism which was solely responsible of the degradation activity. Fenamiphos
degradation could only be achieved by a mixed bacterial culture (Ou, 1991). Singh et al.
(2003) also found a bacterial consortium capable to rapidly degrade fenamiphos and, again,
the individual isolates were not capable of degrading the nematicide. Brevibacterium sp.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
55
MM1 was the first reported individual bacterium capable of rapidly hydrolyzing fenamiphos,
but the bacterium was not molecularly characterized (Megharaj et al., 2003). To date, there is
little information about fenamiphos degrading microorganisms especially of those that can
perform the metabolization process individually. Knowledge about the different species that
can be responsible for the degradation of fenamiphos and nematicides in general, will
facilitate prediction of how successful nematicides can behave in certain soils and how a
rapid break down of the nematicidal activity can be avoided. In the previous chapter it was
demonstrated that soils were found containing microbes, other than Pseudomonads, which
could degrade fenamiphos. Therefore, the objectives of this study were to:
1. Determine which type of microorganism is primarily involved in the fenamiphos
metabolization, bacteria or fungi.
2. Analyze the fenamiphos biodegrading capacity of individual purified isolates.
3. Elucidate if adherent compounds in the formulation, Nemacur GR, enhance or interfere
with fenamiphos metabolization.
4. Investigate if individual bacterial colonies utilize fenamiphos as sole source of carbon to
grow.
5. Characterize and identify fenamiphos degrading bacteria.
5.2. Materials and Methods
5.2.1. Determination of microorganisms responsible for enhanced fenamiphos degradation
To determine whether or not bacteria and fungi were involved in fenamiphos metabolization
an in-vitro degradation assay was set up. Since microorganisms living in soil S3 were able to
rapidly metabolize fenamiphos, as demonstrated in the previous chapter, this soil was
selected for this assay. 10 g of S3 were mixed with 100 ml of autoclaved distilled water. This
mixture was incubated in dark conditions for 16 h at 28 °C in an orbital shaker at 120 rpm.
Subsequently, 1 ml of the water soil mixture containing all soil microorganisms of S3 was
transferred to 19 ml SELM in sterile plastic tubes, supplemented with a stock solution of
fenamiphos analytical grade to a final concentration of 100 µg ml-1. This mixture was further
treated with Cycloheximide (150 µg ml-1), Streptomycin sulfate (150 µg ml-1) and

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
56
Chloramphenicol (150 µg ml-1) or untreated. SELM containing fenamiphos and no
microorganisms from S3 was used as control. All treatments were maintained vertically in
dark conditions at 28 °C in an orbital shaker at 60 rpm. At day 0, 3, 6, 9, 12, 15 and 20 after
nematicide treatment, a 400 µl sample was taken from each treatment, mixed with 1600 µl of
methanol, filter-sterilized and analyzed by HPLC as described in section 2.8. Each treatment
was done in triplicate.
To determine whether fungi and bacteria had been affected by the fungicide and antibiotic
applications, respectively, 50 µl of each treatment was transferred onto TSA+ or PDA+ in 10
cm diameter Petri dishes 20 days after nematicide application. The plates were incubated for
24 h in dark conditions at 28 °C and subsequently the number of CFU were determined. Each
treatment had three replicates.
5.2.2. Metabolization of fenamiphos by bacterial single colonies
From the previous treatments, 50 µl SELM supplemented with fenamiphos, with and without
Cycloheximide and inoculated with the microorganisms obtained from soil S3, were plated
onto 10 cm TSA+. The plates were sealed with Parafilm and incubated for 48 h in the dark at
28 °C. A total of 20 well separated single colonies obtained from each of the two treatments
were picked with a sterile 4 mm loop and transferred to fresh TSA+ and further incubated at
28 °C for 48 h in the dark. One full 4 mm loop of each purified colony was taken under
laminar flow and transferred onto SEAM supplemented with fenamiphos at 100 µg ml-1 in 5
cm diameter Petri dishes. Samples from the agar medium were taken 0, 7 and 14 days after
bacterial inoculation as shown in Figure 1. To determine the remaining fenamiphos
concentration the agar samples were extracted by adding methanol to the agar samples in a
1:1 ratio (v:w) and analyzed by HPLC as described in section 2.8. After every sampling date,
the plates were sealed again and further incubated at 28 °C in the dark until the next sampling
moment. Each sampling was done in triplicate.
The single colonies that were able to metabolize fenamiphos were further cultured on TSA+,
grown in the dark at 28 °C and used in a second metabolization assay, similar as the one
described above, but now using the formulation Nemacur 5GR instead of fenamiphos
analytical grade. Plates contained Nemacur 5 GR with an equivalent of 100 µg ml-1 of active
ingredient. The incubation, sampling and fenamiphos extraction was identical to the plate
assay with the purified fenamiphos. Each sampling was done in triplicate.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
57
Control Bacterial single colony
A
B
C
Control Bacterial single colony
A
B
C
Figure 1. Bioassay for determining nematicide degradation in agar medium. Petri dishes containing soil extract agar medium amended with 100 μg ml-1 of fenamiphos without bacteria (Control) or with a bacteria single colony growing in the center. Arrows show the 3 samples taken for nematicide quantification at 0 (A), 7 (B) and 14 (C) days after nematicide application. Nematicides were extracted from the agar by adding methanol to the agar sub-samples in a 1:1 ratio (v:w) and analyzed by HPLC.
5.2.3. Use of fenamiphos as sole carbon source
To investigate whether soil bacteria of S3 can use fenamiphos as sole carbon source, one
bacterium of those previously isolated was grown in the minimal SELM in the presence and
absence of fenamiphos. A bacterium isolated from the non-degrading S4 was used as a
negative degrading control. A single colony of each bacterium was slected and grown in 10
cm diameter Petri dishes for 72 h in dark conditions at 28 °C. From these plates, 0.1 g of each
bacterial colony was transferred with a sterile 4 mm loop to 10 ml of sterile SELM in a
plastic tube and mixed. From this suspension, 1 ml of the mixture was transferred to 19 ml of
SELM with and without 100 µg ml-1 fenamiphos analytical grade. SELM with and without
fenamiphos served as controls. The medium was incubated in the dark at 28 °C, in an orbital
shaker set at 60 rpm. From each treatment two 1.6 ml samples were taken at 0, 4, and 72 h
after inoculation. For the first sample the optical density (OD) of the medium was determined
using a Ultrospect III Spectrophotometer (Pharmacia Biosystems) set at 600 nm, in which the
OD600 of the non inoculated medium without fenamiphos was set at zero. Each treatment had
3 replicates.
Parallel to the in-vitro assay described above, a second SELM sample was taken from every
treatment to quantify the amount of fenamiphos present in the medium according to the
procedure as described in the second test in section 4.2.5. HPLC analysis was performed to
every sample as described in section 2.8. Each sampling was done in triplicate.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
58
5.2.4. Molecular characterization and identification of fenamiphos degrading bacteria
To molecularly characterize and identify the fenamiphos-degrading bacterial single colonies,
each isolate was grown individually on TSA+ in 10 cm Petri dishes for 48 h at 28 °C in the
dark. Restriction Fragment Length Polymorphism (RFLP) patterns of the 16S rRNA gene and
partial 16S rRNA coding sequence analysis was performed as described in section 2.7.2. The
RFLP analysis was performed as described in section 2.7.2.5. using Cfo1 as restriction
endonuclease. Chromatographic sequencing data was visually/manually inspected for
sequence quality using BioEdit (Hall, 1999). The DNA sequences were analyzed using a
Blast search on the National Center for Biotechnology Information (NCBI) server
(http://www.ncbi.nlm.nih.gov/).
5.2.5. Phylogenetic analysis and sequence comparison
The phylogenetic analysis was performed as described by Kurtz (2009). The analysed dataset
consisted of a total of 25 bacterial strains. The 17 sequences of fenamiphos-degrading
bacteria were aligned with NCBI sequences of Microbacterium, Sinorhizobium,
Brevundimona, Ralstonia and Cupriavidus species resembling the taxonomic groups
identified by the score of highest identity at NCBI Blast. The sequence of Pseudomonas
fluorescence was used as reference. Staphylococcus aureus was used as outgroup.
Chromatographic sequencing data was visually/manually inspected for sequence quality
using BioEdit. Assembling of sequences and transformation into fasta format was performed
with Vector NTI Advance 10.0 (Invitrogen). Aligning, editing and converting of fasta files
into nexus format was performed with ClustalX 1.81 (Thomson et al., 1997).
To identify the best fitting model for nucleotide substitution of 16S rRNA datasets
MrModeltest was used. According to AIC (-lnL = 4693.9512; AIC = 9405.9023)
implemented in MrModeltest general time reversible model of nucleotide substitution with
gamma rates (GTR+G) and base frequencies freq A = 0.2353; freq C = 0.2597; freq G =
0.2791; freq T = 0.2260 was selected for the Baysian inference and analysis settings for Paup
and MrBayes. Baysian inference started from a random tree defining Staphylococcus as for
outgroup. The burn period started after 25% of the 5.000.000 cycles performed, discarding all
trees generated prior burnin. Paup started from a random tree using Staphylococcus as
outgroup. Convergence amongst chains was followed by log-likelihood values using Tracer

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
59
v.1.3. Bayesian semi-strict 50% majority-rule consensus tree was generated in Paup*. A
Parsimony Jackknife analysis was performed with default settings, except for gaps, which
were handled as a new fifth state. Jackknife was performed with 37% deletion of data, 10.000
replicates searching under fast stepwise addition.
5.3. Results
5.3.1. Determination of microorganism responsible for enhanced fenamiphos degradation
To determine what type of microorganisms were involved in fenamiphos degradation an in-
vitro bioassay was set up in which the degradation of the nematicide was followed over time.
In parallel assays the development of either fungi or bacteria was repressed by
Cycloheximide or antibiotics, respectively. In the absence of Cycloheximide and antibiotics
the degradation of fenamiphos started 3 days post inoculation (d.p.i.) and was completed 12
d.p.i. (Figure 2). In the presence of the fungicide Cycloheximide, the degradation of
fenamiphos started 12 d.p.i. and was almost completed 20 d.p.i. In the presence of the
antibiotics Streptomycin sulfate and Chloramphenicol, fenamiphos was not degraded at all
during the 20 days incubation period. These results indicated that the metabolization of
fenamiphos was primarily mediated by soil bacteria and that fungi are less involved.
0
20
40
60
80
100
0 3 6 9 12 15 20
Days of incubation after fenamiphos treatment
Fena
mip
hos
conc
entra
tion
[%]
com
pare
d to
the
trea
tmen
t with
no
mic
roor
gani
sms
S3+Antibiotics S3+Fungicide S3
Figure 2. The remaining amount of fenamiphos during incubation in the presence of microorganisms isolated from the German soil S3. Samples from medium plates, initially containing 100 µg ml-1 fenamiphos analytical grade from plates supplemented with antibiotics (Streptomycin sulfate at 150 µg ml-1 and Chloramphenicol at 150 µg ml-1), fungicide (Cycloheximide at 150 µg ml-1) or untreated (S3), were taken 0, 3, 6, 9, 12, 15 and 20 days after inoculating the microorganisms from S3. N = 3. Bars indicate the Standard Error of the Mean.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
60
Samples from the fungicide, antibiotic and control treatments were taken 20 days after
inoculating the microorganisms of S3 and plated on TSA+ and PDA. Although the treatments
were supplemented with either antibiotics or a fungicide, bacteria and fungi were isolated
from both treatments (Figure 3). However, the amount of bacteria in the treatment with
antibiotics was significantly lower than in the fungicide treated and untreated control, being
120 × 104, 963 × 104 and 1071 × 104 CFU per gram of soil, respectively. Thus, the number of
bacteria in the fungicide treatment was not significantly different from the untreated control.
Similarly to what happened with soil bacteria, the number of fungi in the treatment with
fungicide was significantly lower than in the treatment with antibiotics and the untreated
control with 42 × 104, 823 × 104 and 830 × 104 CFU per gram of soil, respectively. Here, the
number of fungi in the antibiotic treatment was not significant different that the untreated
control.
0200
400600
8001000
12001400
S3+Antibiotics S3+Fungicide S3
Treatment
Mea
n of
CFU
X10
^4 p
er g
of s
oil Bacteria Fungi
B b
a a
A A
Figure 3. Mean number of bacteria and fungi colony forming units (CFU) per gram of soil growing after 24 h of incubation on TSA+ or PDA+ and isolated 20 days after nematicide application from SELM amended with antibiotics (Streptomycin sulfate at 150 µg ml-1 and Chloramphenicol at 150 µg ml-1), fungicide (Cycloheximide at 150 µg ml-1) or untreated (S3), inoculated with microorganisms from Soil 3. N = 3. Bars indicate the Standard Error of the Mean.
5.3.2. Metabolization of fenamiphos by bacterial single colonies
From the 40 bacterial single colonies tested 17 isolates with the numbers 1, 2, 3, 4, 6, 10, 12,
15, 17, 20, 22, 23, 24, 32, 34, 35 and 38 were able to metabolize 100 µg ml-1 fenamiphos
analytical grade almost to completion in a plate assay with the nematicide as sole carbon
source after 14 days of incubation (Figure 4). After 7 days all these isolates had degraded
fenamiphos by more that 50%. The bacterial isolates 1 to 20 were isolated from the treatment

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
61
with Cycloheximide and isolates number 21 to 40 came from the untreated control. Isolates
number 5, 7, 8, 9, 11, 13, 14, 15, 18, 19, 21, 25, 26, 27, 28, 29, 30, 31, 33, 36, 37, 39 and 40
were not able to significantly metabolize fenamiphos after 14 days of incubation.
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40
Bacterial Code
Fena
mip
hos
conc
entr
atio
n [%
] co
mpa
red
to th
e tr
eatm
ent w
ith n
o ba
cter
ia
7 days 14 days
Figure 4. Metabolization of fenamiphos in soil extract agar medium originally amended with 100 µg ml-1 fenamiphos analytical grade by 40 bacterial single colonies isolated from S3 after 7 and 14 days of incubation. N =3. Bars indicate the Standard Error of the Mean. All 17 fenamiphos analytical grade degrading isolates were further tested for their ability to
metabolize fenamiphos in the formulation Nemacur 5GR. With the exception of isolate
number 3 all isolates degraded the active ingredient, although degradation was not as efficient
compared to the analytic grade (Figure 5). After 7 days some isolates had not yet degraded at
all (isolate 38) or only degraded fenamiphos by 30 to 40%. However, after 14 days most
isolates had degraded 80% of fenamiphos. The isolate number 38 was able to metabolize only
half of the nematicide concentration 14 days after incubation. Apparently, components
present in the formulation affect the development of the bacteria or their degradation efficacy.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
62
0
20
40
60
80
100
1 2 3 4 6 10 12 16 17 20 22 23 24 32 34 35 38
Bacterial codeFena
mip
hos
conc
entra
tion
[%]
com
pare
d to
the
treat
men
t with
no
bac
teria
Figure 5. Metabolization of fenamiphos in soil extract agar medium originally amended with Nemacur 5 GR applied at 100 µg ml-1 active ingredient by 17 fenamiphos-analytical grade degrading bacterial single colonies isolated from S3 after 7 and 14 days of incubation. N =3. Bars indicate the Standard Error of the Mean.
5.3.3. Use of fenamiphos as sole carbon source
The fenamiphos degrading bacterium isolate 32 was not able to grow in SELM that was not
amended with fenamiphos, resulting in no increase in bacterial density (Figure 6). When 100
µg ml-1 fenamiphos was present the OD600 increased from 0.12 to 0.16 nm at 4 hours after
application, indicating bacterial growth. However, after this optical density increment, the
OD600 of the culture decreased to 0.08 nm after 72 hours. The bacterium that was isolated
from the fenamiphos non-degrading soil S4 did not show an increase in density, both in the
absence and presence of fenamiphos. Apparently this isolate was not able to use fenamiphos
as a carbon source and no alternative carbon sources were available for this bacterium.
In the same assay the fate of the fenamiphos was monitored by HPLC analysis. This showed
that the fenamiphos degrading isolate 32 had already converted 95% of the total fenamiphos
after 4 hours of incubation (Figure 7). As expected, in the medium containing the non-
degrading isolate no fenamiphos degradation was observed, thus remaining at the same level
as the non inoculated control. The almost complete conversion of fenamiphos 4 hours after
the start of the incubation explains the observation that the bacterial biomass only increases in
these first 4 hours.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
63
0.00
0.05
0.10
0.15
0.20
0 4 72
Hours of incubation after fenamiphos application
UV li
ght a
bsor
banc
e [n
m]
Isolate 32 Isolate 32+F BS4 BS4+F SELM+F
Figure 6. Optical density analysis at 600 nm wave length to determine growth behaviour of the fenamiphos-degrading isolate number 32 isolated from S3 and a bacterium isolated from non-degrading S4 (BS4) in soil extract liquid media (SELM) in the presence (+F) or absence of fenamiphos. N = 3. Bars indicate the Standard Error of the Mean.
0102030405060708090
100
0 4 72
Hours of incubation after nematicide application
Fena
mip
hos
conc
entra
tion
[%] f
rom
inic
ital a
pplic
atio
n
Isolate 32 SELM BS4
Figure 7. Metabolization of fenamiphos initially applied at 100 µg ml-1 in soil extract liquid media (SELM) by fenamiphos-degrading isolate number 32 isolated from S3 or a bacterium isolated from non-degrading S4 (BS4). N = 3. Bars indicate the Standard Error of the Mean.
5.3.4. Molecular characterization and identification of fenamiphos degrading bacteria
Of all 17 fenamiphos-degrading bacterial isolates the genomic 16S rRNA coding regions
were amplified by PCR and analyzed by restriction length polymorphism and DNA sequence
analysis. The isolated bacteria produced a variety of 16S rRNA RFLP profiles (Figure 8). The

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
64
restriction enzyme analysis provided an initial clustering of strains which divided into two
groups with three or more members within the same profile. The first group of isolates, or
group A, comprised eleven members; 1, 3, 4, 10, 12, 22, 23, 24, 34, 35 and 38. All these
isolates showed high homology to the internal fragment of the 16S rRNA gene. A second
group, group D, was conformed by three isolates; 16, 17 and 20. There were three individual
isolates with unique fragmentation patterns, been number 2, 6 and 32.
L 1 2 3 4 6 10 12 16 17 20 22 23 24 32 34 35 38
A B A A C A A D D D A A A E A A A
Fd1 Fd2 Fd3 Fd4 Fd5 Fd6 Fd7 Fd8 Fd9 Fd10 Fd11 Fd12 Fd13 Fd14 Fd15 Fd16 Fd17
Figure 8. RFLP patterns of the 1.5 kb PCR amplified fragments of the 16S rRNA gene of 17 fenamiphos-degrading bacterial isolates, digested with the restriction endonuclease Cfo1. Lane L is the 50bp molecular size marker (Promega). Numbers in the upper line indicate the number of the isolate in the previous fenamiphos metabolization tests. Letters in the middle line show the group to which each isolate correspond according to its RFLP type. Fd in the lower line shows the new code assigned for its capacity of fenamiphos degradation.
Almost complete 16S rRNA sequences of 17 fenamiphos-degrading bacterial strains were
obtained, ranging in length from 1080 to 1397 nucleotides. Blast analysis revealed that
isolates 1, 3, 4, 10, 12, 22, 23, 24, 34, 35 and 38 had high similarities, ranging from 95 to
99% of match identity, with the sequences of two Microbacterium sp. from the NCBI library
(Table 1). The isolates 16 and 20 had a match identity of 95 and 96% with the sequence of
Ralstonia sp., respectively. The partial sequence of isolate 2 was similar to Sinorhizobium
spp. with 93% identity match. The sequence similarity between isolate 6 and Brevundimonas
mediterranea was of 95%. Isolates 17 and 32 had similar sequences as Cupriavidus sp. and
C. necator with 95 and 92% identity match, respectively.
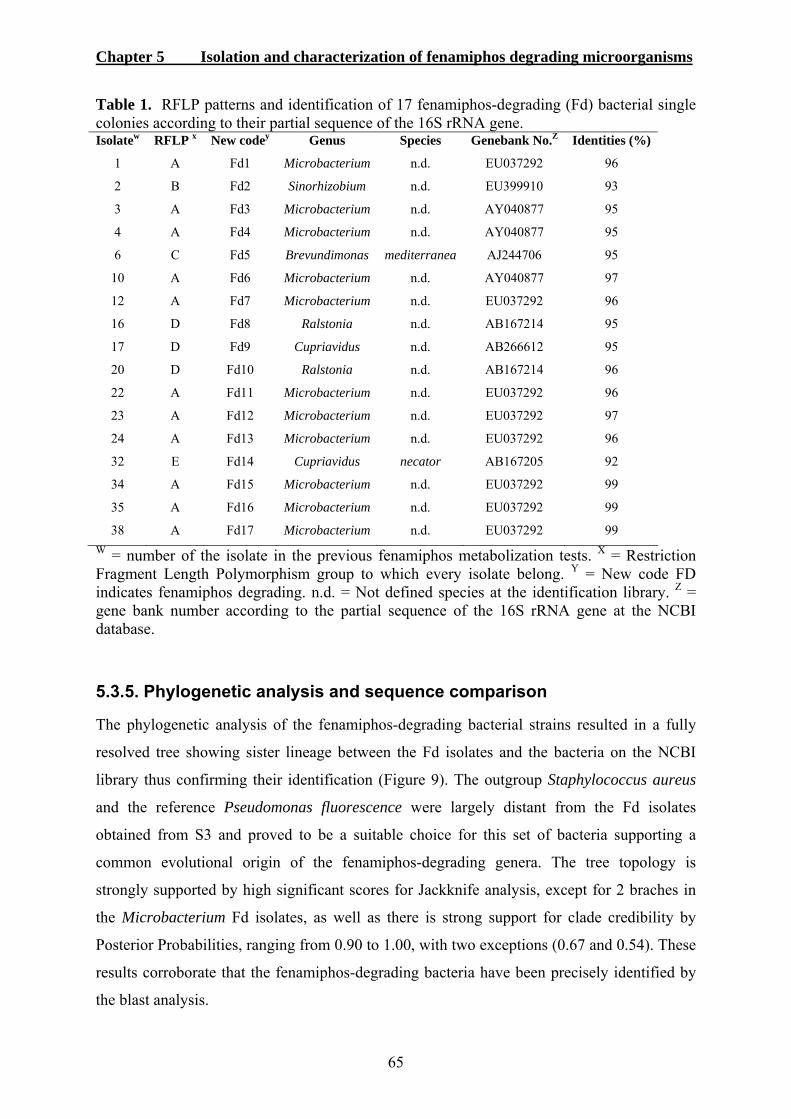
Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
65
Table 1. RFLP patterns and identification of 17 fenamiphos-degrading (Fd) bacterial single colonies according to their partial sequence of the 16S rRNA gene. Isolatew RFLP x New codey Genus Species Genebank No.Z Identities (%)
1 A Fd1 Microbacterium n.d. EU037292 96
2 B Fd2 Sinorhizobium n.d. EU399910 93
3 A Fd3 Microbacterium n.d. AY040877 95
4 A Fd4 Microbacterium n.d. AY040877 95
6 C Fd5 Brevundimonas mediterranea AJ244706 95
10 A Fd6 Microbacterium n.d. AY040877 97
12 A Fd7 Microbacterium n.d. EU037292 96
16 D Fd8 Ralstonia n.d. AB167214 95
17 D Fd9 Cupriavidus n.d. AB266612 95
20 D Fd10 Ralstonia n.d. AB167214 96
22 A Fd11 Microbacterium n.d. EU037292 96
23 A Fd12 Microbacterium n.d. EU037292 97
24 A Fd13 Microbacterium n.d. EU037292 96
32 E Fd14 Cupriavidus necator AB167205 92
34 A Fd15 Microbacterium n.d. EU037292 99
35 A Fd16 Microbacterium n.d. EU037292 99
38 A Fd17 Microbacterium n.d. EU037292 99 W = number of the isolate in the previous fenamiphos metabolization tests. X = Restriction Fragment Length Polymorphism group to which every isolate belong. Y = New code FD indicates fenamiphos degrading. n.d. = Not defined species at the identification library. Z = gene bank number according to the partial sequence of the 16S rRNA gene at the NCBI database.
5.3.5. Phylogenetic analysis and sequence comparison
The phylogenetic analysis of the fenamiphos-degrading bacterial strains resulted in a fully
resolved tree showing sister lineage between the Fd isolates and the bacteria on the NCBI
library thus confirming their identification (Figure 9). The outgroup Staphylococcus aureus
and the reference Pseudomonas fluorescence were largely distant from the Fd isolates
obtained from S3 and proved to be a suitable choice for this set of bacteria supporting a
common evolutional origin of the fenamiphos-degrading genera. The tree topology is
strongly supported by high significant scores for Jackknife analysis, except for 2 braches in
the Microbacterium Fd isolates, as well as there is strong support for clade credibility by
Posterior Probabilities, ranging from 0.90 to 1.00, with two exceptions (0.67 and 0.54). These
results corroborate that the fenamiphos-degrading bacteria have been precisely identified by
the blast analysis.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
66
Figure 9. Phylogenetic analysis of the partial 16S rRNA of fenamiphos-degrading and closely related bacteria using Staphylococcus aureus (FJ609418) as outgroup and Pseudomonas fluorescence (EU73092) as reference. 50% majority rule semi-strict consensus tree generated in MrBayes is displayed as dendrogram. Model of nucleotide substitution was GTR+G. Values annotated to each node indicate: Jackknife (37% deletion of data) / Posterior probabilities. Nodes with Jackknife values below 67 are generally considered as weak supported (-). Bars indicate the relative number of substitutions per site. The accession number of the 16S rRNA sequences obtained from NCBI database showing a high identity with Fd bacteria are shown in parenthesis.
The sequence alignment of 17 fenamiphos-degrading and other 8 closely related bacteria,
showed different regions of similarity among isolates (Table 2). Based on 1092 bp of the 16S
rRNA coding sequence information this study revealed that there were 29 similar nucleotide
regions among all the isolates compared. The Fd sequences were divided in two groups
according to their sequence homology. The first group was composed by the Microbacterium
sp. and Brevundimonas mediterranea isolates and the second group by the rest of bacteria.
However, every sequence of the Fd isolates had more similar nucleotide regions with their
respective identified genera than with the rest of bacteria. These results support the blast
identification and the phylogenetic dendrogram and re-confirm the high similarity between
the Fd isolates from S3 and the bacteria reported on the NCBI library.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
67
Table 2. Sequence alignment of 17 fenamiphos-degrading and closely related bacteria based on 1092 bp of 16S rRNA information.
Microbacterium sp. EU037292 (Mb.), Microbacterium sp. AY040877 (Microbacter), Brevundimonas mediterranea AJ244706 (Brevundimon), Cupriavidus necator AB167205 (Cupriavidus), Ralstonia sp. AB167214 (Ralstonia), Pseudomonas fluorescence EU73092 (Pseudomonas), Staphylococcus aureus FJ609418 (Staphylococ), and Sinorhizobium sp. EU399910 (Sinorhizobi). *Identical nucleotide position in all bacteria. Bacteria were aligned according to similarity. Different colours highlight the homology and changes in the nucleotide.
5.4. Discussion
In this study, it is further demonstrated that metabolization of fenamiphos was performed by
microorganisms of S3, a soil with a 12 year long fenamiphos application history. In contrast,
there was no nematicide metabolization where antibiotics were applied. These results
strongly suggest the involvement of bacteria rather than fungi in fenamiphos degradation.
However, microorganisms in the treatment where neither antibiotics nor fungicide were
applied degraded fenamiphos at the highest rate suggesting that the combination of bacteria
and fungi accelerate even more fenamiphos degradation than bacteria alone. Similar results
were reported for another organophosphate non-fumigant nematicide ethoprophos. Karpouzas
et al. (1999) found that an untreated degrading-soil metabolized more rapidly the nematicide
than soil treated with Cycloheximide and this degraded faster than soil treated with
Chloramphenicol. Both studies are in agreement that bacteria are the main microbes involved
in the rapid degradation process of organophosphate nematicides in soil. Additionally,
bacteria have been reported to be capable of degrading other organophosphate pesticides such
as parathion, fensulfothion and diazinon (Sheela and Pai, 1983; Serdar et al.,
1982; Sethunthan and Yoshida, 1973). Furthermore, Rosenberg and Alexander (1979)
demonstrated that the breakdown of organophosphate pesticides result from the synthesis of

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
68
an enzyme or enzyme complex which support the organism in using the organophosphates as
phosphorus sources for growth and development.
Even though the soil extract liquid medium was treated with either fungicide or antibiotics, it
was still possible to isolate fungi and bacteria from the corresponding amended treatments.
However, the amount of bacteria growing on TSA+ obtained from the antibiotic amended
treatment and the number of fungi growing on PDA+ obtained from the fungicide amended
treatment were 10 times lower than those obtained from the opposite treatment or the
untreated control. It is well known that Cycloheximide is an inhibitor of protein biosynthesis
in eukaryotic organisms. Streptomycin sulfate inhibits protein synthesis leading to death of
microbial cells and Chloramphenicol is a bacteriostatic antimicrobial compound, explaining
the results herein. These results illustrated that fungi and bacteria suffered and their growth
and development was strongly inhibited by the application of Cycloheximide or Streptomycin
sulfate together with Chloramphenicol, respectively. Thus this confirms the conclusion that
bacteria are the dominating factor in the fenamiphos degradation process.
From the 40 bacterial single colonies tested, obtained from the biodegrading soil S3, 17 were
able to metabolize fenamiphos analytical grade at an accelerated rate during the 14 days of
incubation period. In addition, these 17 isolates were capable to metabolize fenamiphos as
active ingredient of Nemacur 5GR, with the exception of isolate number 3 (Fd3), confirming
their biodegrading capability. This conclusion agrees with other studies that have reported
bacteria as principal factors for fenamiphos enhanced degradation in soil (Ou, 1991;
Megharaj et al., 2003; Singh et al., 2003).
Isolate number 3 (Fd3) was the only bacteria not capable of degrading fenamiphos in
Nemacur 5GR. Additionally, most of the isolates were not as efficient in degrading
fenamiphos in the commercial product as in the analytical grade, although the amount of
fenamiphos in both tests was identical (100 µg ml-1). Nemacur 5GR formulation contains 5%
of the active ingredient and 95% of other unspecified aggregate compounds and it may have
been that these additives directly interfered with the degradation process or supplied an
alternative nutrient source for the bacteria, thus reducing the metabolization of fenamiphos.
All fenamiphos-degrading bacteria were identified. According to the amplification of the 16S
rRNA gene and blast analysis, eleven isolates from the seventeen fenamiphos-degrading

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
69
corresponded to Microbacterium spp. showing high similarity to Microbacterium species
reported at the NCBI library. Restriction enzyme analysis with Cfo1 revealed that these
bacteria had high homology of their internal DNA fragment. This first bacterial group
represented 64.7% of all the fenamiphos-degrading isolates which composed the RFLP type
A. The phylogenetic analysis confirmed this identification showing sister lineage between
Microbacterium Fd isolates and the Microbacterium sp. from NCBI. Sequence comparison
reconfirmed this result. Previous phylogenetic analyses of most Microbacterium species have
shown that this genus has a common ancestor and that the different species are close relatives
(Schippers et al., 2005: Takeuchi and Hatano, 1998). The present study is in accordance with
a recent short communication which reported that Microbacterium esteraromaticum was able
to hydrolyze fenamiphos and its toxic oxidation products (Cáceres et al., 2009). Furthermore
Microbacterium species have been previously reported to degrade the carbamate non-
fumigant nematicide carbofuran and the herbicide isoxaben at an accelerated rate (Karpouzas
et al., 2000b; Arrault et al., 2002). For the congruence between the present work and the
above cited, the genus Microbacterium appears to be an important component of the pesticide
enhanced biodegradation process.
A second group of fenamiphos-degrading bacteria was composed by isolates 16 (Fd8), 17
(Fd9) and 20 (Fd10) which had high homology of the internal fragment RFLP type D. The
partial sequence of this group was identified to belong to the genus Ralstonia or Cupriavidus.
Isolate 32 (Fd14) was identified as Cupriavidus necator but its RFLP pattern was unique
(type E). Ralstonia eutropha is currently considered Cupriavidus necator and therefore the
sequence similarities exist (Manzano et al., 2007). However, the species of these two genera
can significantly differ from one another thus encoding different DNA internal fraction.
Nevertheless, the phylogenetic analysis and sequence comparison confirmed the
identification of these bacteria as it occurred with the Microbacterium Fd species.
Cupriavidus and Ralstonia species can degrade herbicides such as 2,4-dichlorophenoxyacetic
acid (2,4-D) and 4-chloro-2-methylphenoxyacetic acid (Huong, et al., 2007; Manzano et al.,
2007; Martineza et al., 2008; Nicolaisen et al., 2008). Until now, there have been described
15 genes that play a role in the 2,4-D degradation (Streber et al., 1987; Perkins et al., 1990;
Matrubutham and Harker, 1994; You and Ghosal, 1995; van der Meer, 1997; Leveau et al.,
1999; Laemmli et al., 2000; Perez-Pantoja et al., 2000). Perhaps these genes are also
necessary for the degradation of fenamiphos, thus explaining the results obtained in this
work. Moreover this is the first time that Ralstonia and Cupriavidus species are reported to
metabolize a nematicide.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
70
There were other two isolates with a unique RFLP pattern. The first one was isolate number 6
(Fd5) identified as Brevundimonas mediterranea. Its identification was also confirmed by
phylogenetic analysis and sequence comparison. Bacteria from this genus have been
previously found to degrade other organophosphate pesticides such as the insecticide
chlorpyrifos and the herbicide trifluralin (Bellinaso et al., 2003; Li et al., 2008).
Brevundimonas diminuta contains the bacterial organophosphorus-hydrolyzing enzyme,
which is a zinc-containing homodimeric protein (Horne, 2002). Since B. mediterranea and B.
diminuta are close related bacteria, it can be expected that the isolate 6 (Fd5) found in this
research also posses an enzyme involved in organophosphates degradation and utilize it to
degrade fenamiphos. This is the first report of Brevundimonas mediterranea rapidly
degrading a nematicide.
The last bacteria with unique RFLP pattern was isolate 2 (Fd2) identified as Sinorhizobium
sp. according to its partial sequence of the 16S rRNA gene. As the other 16 Fd isolates, this
bacterium was able to metabolize fenamiphos in soil extract agar medium as sole source of
carbon. The phylogenetic analysis and sequence alignment confirmed that this isolate belongs
to the Sinorhizobium bacteria. Sinorhizobium has been reported to degrade the polycyclic
aromatic hydrocarbon Phenanthrene (Seo et al., 2007) and posses the herbicide degrading
genes which are able to mineralize atrazine (Devers et al., 2007). Perhaps this bacterium also
uses these genes to rapidly degrade fenamiphos. The nematicide degradation ability of
Sinorhizobium sp. was not previously reported.
One fenamiphos degrading isolate, Cupriavidus necator Fd14, revealed that the bacterium
utilized fenamiphos as sole carbon source for growth. The bacterium seemed to respond very
quickly to the availability of fenamiphos resulting in a rapid and complete degradation of the
organophosphate. The fenamiphos molecule can be degraded through different pathways into
several products. Fenamiphos can be oxidized into fenamiphos sulfoxide (FSO) which is
further oxidized into fenamiphos sulfone (Singh and Walker, 2006). Fenamiphos can be
degraded into FSO and then turned into FSO-phenol and subsequently mineralized into CO2
(Chung and Ou, 1996). Fenamiphos can also be directly converted into fenamiphos phenol,
where probably the first oxidation step is replaced by hydrolysis (Singh et al., 2003).
In conclusion, this study demonstrated for the firs time that gram-positive and gram-negative
bacteria are the principal microorganisms responsible for fenamiphos degradation in soil.
These bacteria belong to six different genera, namely Microbacterium, Ralstonia,

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
71
Brevundimonas, Cupriavidus and Sinorhizobium. Additionally, this study revealed that the
combination of all soil microorganisms degraded the organophosphate nematicide faster than
bacteria alone. Synthetic organophosphates contain three phosphoester linkages and the
phosphorus is also linked by a double bond to either an oxygen in oxon-organophosphates or
a sulfur in thion-organophosphates (Horne et al., 2002). Hydrolysis of one of the
phosphoester bonds reduces the toxicity of an organophosphate therefore reducing its
efficacy for agricultural purposes. However, despite the fact that enhanced biodegradation
has a negative effect on the control of plant-parasitic nematodes, it is a beneficial process
from an ecological point of view since it eliminates potentially dangerous products from soil,
which otherwise could cause serious problems to animal life and human health through
accumulation in the environment (Moens et al., 2004). Fenamiphos activity has been found to
persist for 3–4 months or more in soil (Homeyer, 1971) and for several years in groundwater
(Franzman et al., 2000) thus the fenamiphos-degrading bacteria isolated in this study are
good candidates for bioremediation of fenamiphos in areas where this organophosphate
nematicide persist.
5.5. References
Anderson J.P.E. 1989. Accelerated microbial degradation of nematicides and other plant
protection chemicals in soils. Proceedings of the 20th annual meeting of OTAN, 7-11
November, 1988, San Jose, Costa Rica. Nematropica 19:1.
Anderson, J.P.E. and Lafuerza, A. 1992. Microbiological aspects of accelerated pesticide
degradation. In J.P.E. Anderson, D.J. Arnold, F. Lewis and L. Torstensson (ed.),
Proceedings of the International Symposium of Environmental Aspects of Pesticide
Microbiology, 17-21 August 1992, Sigtua, Sweden. p. 184-192.
Arrault S., Desaint S., Catroux C., Se´mon E., Mougin C. and Fournier J.C. 2002. Isolation
and characterization of efficient isoxaben-transforming Microbacterium sp. strains
from four European soils. Pest Management Science 58:1229-1235.
Bellinaso, M.D.L., Greer, C.W., do Carmo-Peralba, Pegas-Henriques M.J.A. and Gaylarde, C.
C. 2003. Biodegradation of the herbicide trifluralin by bacteria isolated from soil.
FEMS Microbiology Ecology 43:191-194.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
72
Cáceres, T.P., Megharaj, M., Malik, S., Beer, M., and Naidu, R. 2009. Hydrolysis of
fenamiphos and its toxic oxidation products by Microbacterium sp. in pure culture and
groundwater. Bioresource Technology 100:2732-2736.
Chung, K.Y. and Ou, L.T. 1996. Degradation of fenamiphos sulfoxide and fenamiphos
sulfone in soil with history of continous application of fenamiphos. Archives of
Environmental Contamination and Toxicology 30:452-458.
Devers, M., El-Azhari, N., Kolic, N.U., Martin-Laurent, F. 2007. Detection and organization
of atrazine-degrading genetic potential of seventeen bacterial isolates belonging to
divergent taxa indicate a recent common origin of their catabolic functions. FEMS
Microbiology Letters 273:78-86.
Feng, X., Ou, L.T. and Ogram, A. 1997. Plasmid mediated mineralization of carbofuran by
Sphingomonas sp. Train CF06. Applied and Environmental Microbiology 63:1332-
1337.
Franzmann, P.D., Zappia, L.R., Tilbury, A.L., Patterson, B.M., Davis, G.B. and Mandelbaum,
R.T. 2000. Bioaugmentation of atrazine and fenamiphos impacted groundwater:
laboratory evaluation. Bioremediation Journal 4:237-248.
Homeyer, B. 1971. Nemacur, a highly effective nematocide for protective and curative
application. Pflanzenschutz Nachrichten Bayer 24:48-68.
Horne, I., Sutherland T.D., Harcourt, R.L., Russell, R.J., and Oakeshott J.G. 2002.
Identification of an opd (Organophosphate Degradation) gene in an Agrobacterium
isolate. Applied and Environmental Microbiology 68:3371-3376.
Huong, N.L., Itoh, K. and Suyama, K. 2007. Diversity of 2,4-dichlorophenoxyacetic acid
(2,4-D) and 2,4,5-trichlorophenoxyacetic acid (2,4,5-T)-degrading bacteria in
Vietnamese soils. Microbes and Environments 22:243-256.
Johnson, A.W. 1998. Degradation of fenamiphos in agricultural production soil. Supplement
to the Journal of Nematology 30:40-44.
Karpouzas, D.G., Alun, J., Morgan, J.A.W. and Walker, A. 2000a. Isolation and
characterisation of ethoprophos-degrading bacteria. FEMS Microbiology Ecology
33:209-218.
Karpouzas, D.G., Morgan, J.A.W. and Walker, A. 2000b. Isolation and characterization of 23
carbofuran-degrading bacteria from soils from distant geographical areas. Letters in
Applied Microbiology 31:353-358.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
73
Karpouzas, D.G., Walker, A., Foud-Williams, R. J. and Drennan, D.S.H. 1999. Evidence for
the enhanced biodegradation of ethoprophos and carbofuran in soils from Greece and
the UK. Pesticide Science 55:301-311.
Kurtz, A. 2009. Endophytic Fusarium oxysporum: Phylogeny and induced defense responses
in banana plants (Musa AAA) against Radopholus similis (Cobb) Thorne. PhD Thesis,
University of Bonn. In press.
Laemmli, C.M., Leveau, J., Zehnder, A.J.B., and Van der Meer, J.R. 2000. Characterization
of a second tfd gene cluster for chlorophenol and chlorocatechol metabolism on
plasmid pJP4 in Ralstonia eutropha JMP134 (pJP4). Journal of Bacteriology
182:4165-4172.
Leveau, J.H.J., König, F., Füchslin, H.P., Werlen, C., and van der Meer, J.R. 1999. Dynamics
of multigene expression during catabolic adaptation of Ralstonia eutropha JMP134
(pJP4) to the herbicide 2,4-dichlorophenoxyacetate. Molecular Microbiology 2:389-
395.
Li X.H., Jiang J.D., Gu L.F., Ali, S. W. He J. and Li S.P. 2008. Diversity of chlorpyrifos-
degrading bacteria isolated from chlorpyrifos-contaminated samples. International
Biodeterioration and Biodegradation 62:331-335.
Manzano, M. Morán, A. C. Tesser, B. and González, B. 2007. Role of eukaryotic microbiota
in soil survival and catabolic performance of the 2,4-D herbicide degrading bacteria
Cupriavidus necator JMP134. Antonie van Leeuwenhoek 91: 115-126.
Martineza, C.O., Silvaa, C.M.M.S., Francisconi-Faya, E., Nunes-Maiaa, A.H., Blotta-
Abakerlia, R. and Durrantb, L.R. 2008. Degradation of the herbicide sulfentrazone in a
Brazilian Typic Hapludox soil. Soil Biology and Biochemistry 40:879-886.
Matrubutham, U. and Harker, A.R. 1994. Analysis of duplicated gene sequences associated
with tfdR and tfdS. Alcaligenes eutrophus JMP134. Journal of Bacteriology 176:2348-
2353.
Matsumura, F. and Boush, G.M. 1986. Degradation of insecticides by a soil fungus,
Trichoderma viride. Journal of Economic Entomology 61:610-612.
Megharaj, M., Singh, N., Kookana, R.S., Naidu, R. and Sethunathan, N. 2003. Hydrolysis of
fenamiphos and its oxidation products by a soil bacterium in pure culture, soil and
water. Applied Microbiology and Biotechnology 61:252-256.
Moens, T., Araya, M., Swennen, R. and De Waele, D. 2004. Enhanced biodegradation of
nematicides after repetitive applications and its effect on root and yield parameters in
commercial banana plantations. Biology and Fertility Soils 39:407-414.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
74
Mclean, K.S. and Lawrence, G.W. 2003. Efficacy of Aldicarb to Rotylenchus reniformis and
biodegradation in cotton field soils. Journal of Nematology 35:65-72.
Nicolaisen, M.H., Bælum, J., Jacobsen, C.S. and Sørensen, J. 2008. Transcription dynamics
of the functional tfdA gene during MCPA herbicide degradation by Cupriavidus
necator AEO106 (pRO101) in agricultural soil. Environmental Microbiology 10:571-
579.
Ou, L.T. 1991. Interactions of microorganisms and soil during fenamiphos degradation. Soil
Science Society of American Journal 55:716-722.
Pattison A.B., Stanton, J.M. and Cobon, J.A. 2000. Bioassay for enhanced biodegradation of
nematicides in soil. Australasian Plant Pathology 29:52-58.
Perez-Pantoja, D., Guzman, L., Manzano, M., Pieper, D.H. and Gonzalez, B. 2000. Role of
tfd CIDIEIFI and tfdDIICIIEIIFII gene modules in catabolism of 3-Chlorobenzoate by
Ralstonia eutropha JMP134(pJP4). Applied and Environmental Microbiology
66:1602-1608.
Perkins, E.J., Gordon, M.P., Caceres, O. and Lurquin, P.F. 1990. Organization and sequence
analysis of the 2,4-dichlorophenol hydroxylase and dichlorocatechol oxidative
operons of plasmid pJP4. Journal of Bacteriology 172:2351–2359.
Ramanand, K., Sharmila, M., and Sethunathan, N. 1988. Mineralization of carbofuran by a
soil bacterium. Applied and Environmental Microbiology 54:2129-2133.
Rao, A.V. and Sethunathan, N. 1974. Degradation of parathion by Penicillium waksmani
Zaleski isolated from flooded acid sulphate soil. Archives of Microbiology 97:202-
208.
Rosenberg, A. and Alexander, M. 1979. Microbial cleavage of various organophosphorus
insecticides. Applied and Environmental Microbiology 37:886-891.
Schippers, A., Bosecker, K., Spröer, C. and Schumann, P. 2005. Microbacterium oleivorans
sp. nov. and Microbacterium hydrocarbonoxydans sp. nov., novel crude-oil-degrading
Gram-positive bacteria. International Journal of Systematic and Evolutionary
Microbiology 55:655-660.
Seo, J.S., Keum, Y.S., Harada, R.M. and Li, Q.X. 2007. Isolation and characterization of
bacteria capable of degrading polycyclic aromatic hydrocarbons (PAHs) and
organophosphorus pesticides from PAH-contaminated soil in Hilo, Hawaii. Journal of
Agricultural and Food Chemistry 55:5383-5389.

Chapter 5 Isolation and characterization of fenamiphos degrading microorganisms
75
Serdar, C.M., Gibson, D.T. Munnecke, D.M. and Lancaster, J.H.1982. Plasmid involvement
in parathion hydrolysis by Pseudomonas diminuta. Applied and Environmental
Microbiology 44:246-249.
Sethunthan, N. and Yoshida, T. 1973. A Flavobacterium spp. that degrades diazinon and
parathion. Canadian Journal of Microbiology 19:873-875.
Sheela, S. and Pai, S.B. 1983. Metabolism of fensulfothion by a soil bacterium, Pseudomonas
alcaligenes C1. Applied and Environmental Microbiology 46:475-479.
Singh, B.K., Walker, A., Morgan, J.A.W. and Wright, D.J. 2003. Role of soil pH in the
development of enhanced biodegradation of fenamiphos. Applied and Environmental
Microbiology 69:7035-7043.
Singh, B.K. and Walker, A. 2006. Microbial degradation of organophosphorus compounds.
FEMS Microbiological Review 30:428-471.
Somasundaram, L., Coats, J.R. and Racke, K.D. 1989. Degradation of pesticides in soil as
influenced by the presence of hydrolysis metabolites. Journal of Environmental
Science and Health, Part B 24:457-478.
Streber, W.R., Timmis, K.N., and Zenk, M.H. 1987. Analysis, cloning, and high-level
expression of 2,4- ichlorophenoxyacetate monooxygenase gene tfdA of Alcaligenes
eutrophus JMP134. Journal of Bacteriology 169:2950–2955.
Takeuchi, M. and Hatano, K. 1998. Proposal of six new species in the genus Microbacterium
and transfer of Flavobacterium marinotypicum ZoBell and Upham to the genus
Microbacterium as Microbacterium maritypicum com. nov. International Journal of
Systematic Bacteriology 48:973-982.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., and Higgins, D.G. 1997. The
CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment
aided by quality analysis tools. Nucleic Acids Research 25:4876-4882.
van der Meer, J.R. 1997. Evolution of novel metabolic pathways for the degradation of
chloroaromatic compounds. Antonie Van Leeuwenhoek 71:159–178.
You, I.S., and Ghosal, D. 1995. Genetic and molecular analysis of a regulatory region of the
herbicide 2,4-dichlorophenoxyacetate catabolic plasmid pJP4. Molecular
Microbiology 16:321-331.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
76
6. Fosthiazate cross-degradation and specificity of nematicide degrading bacteria
6.1. Introduction
Repetitive applications of the same nematicide in soil have been shown to enhance its
biodegradation process, especially in monocultures systems, as shown in the previous
chapters and elsewhere (Karpouzas et al., 1999a; Karpouzas et al., 1999b). Adapted
microorganisms, such as soil bacteria, are able to use the nematicides’ active ingredients as a
carbon source (Karpouzas and Walker, 2000; Karpouzas et al., 2005). In agricultural
production, the enhancement of biodegradation is a great concern since it directly reduces
nematicide efficacy (Rich et al., 2004).
The biodegradation of pesticides is not always restricted to the applied pesticide, but can
extend to other pesticides that were never used in the field before. There is already evidence
for this “cross-conditioning” of microbial populations in the soil (Morel-Chevillet et al.,
1996). The efficiency of pesticides such as benfuracarb, carbosulfan and furathiocarb was
reduced by prior treatment of carbofuran (Suett, 1987). Pest control by aldicarb in the field
was reduced by prior treatment with carbofuran rather than prior treatments with aldicarb
itself (Suett, 1989). Laboratory experiments have shown that several successive treatments
with carbofuran led to increased rates of degradation of other compounds (Harris et al.,
1984). Sphingomonas paucimobilis isolated from a soil often treated with cadusafos was able
to cross-degrade ethoprophos (Karpouzas et al., 2005).
Fosthiazate is a relatively new organophosphate non-fumigant nematicide and effective in the
control of potato cyst and root-knot nematodes (Rich et al., 1994; Woods et al., 1999; Woods
and Haydock, 2000; Hafez and Sundararaj, 2006; Tobin et al., 2008). It is considered an
alternative to nematicides that have been on the market for quite some time. However, it was
reported that soil biotic factors can degrade fosthiazate (Pantelelis et al., 2006). Fosthiazate
and fenamiphos belong to the same organophosphate family and it may therefore be possible
that fenamiphos-degrading microorganisms can cross-degrade fosthiazate. The cross-
degradation process is of importance since it could make new pesticides immediately
ineffective, thus offering no alternative for the farmer.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
77
The objectives of this investigation were to:
1. Monitor the degradation of fosthiazate by fenamiphos-degrading microorganisms.
2. Elucidate whether the presence of another organophosphate affects the degradation
process of fenamiphos.
3. Determine whether fenamiphos-degrading bacteria can quickly degrade fosthiazate.
6.2. Materials and Methods
6.2.1. Degradation of fosthiazate
A stock solution of Fosthiazate 150 EC was applied to SELM to a final concentration of 100
µg ml-1 and either not inoculated or inoculated with a mixture of fenamiphos-degrading
microorganisms extracted from soil S3. The inoculation conditions, sampling process and
HPLC analysis were performed as described in the second test of section 4.2.5. Samples were
taken 0, 3, 6, 9, 13, 16, 20 and 30 days after inoculation. Each treatment was done in
triplicate.
6.2.2. Cross-degradation essays
Two cross-degradation assays were set up to investigate whether fenamiphos-degrading
microorganisms can also degrade fosthiazate. In the first assay, to elucidate if all the
fenamiphos-degrading microorganisms isolated from S3 would still enhance degrade
fenamiphos in the presence of fosthiazate, the following treatments were performed. SELM
was supplemented with fenamiphos analytical grade to a final concentration of 100 µg ml-1,
and Nemathorin 10WG to a final concentration of 100 µg ml-1 a.i. SELM supplemented with
fenamiphos was used as control treatment. The SELM was then not inoculated, inoculated
with a mixture microorganisms extracted from soil S3 or inoculated with an autoclaved
mixture of microorganisms extracted from soil S3. To obtain the microorganisms from soil
S3, the same process as described in the second test section 4.2.5. was performed.
Autoclaving was performed for 60 min at 121 °C in the respective treatments. The incubation
conditions, sampling and HPLC analysis were performed as described in section 4.2.5.
Samples were taken 0, 3, 6, 9, 13 and 20 days after inoculation. Each treatment was done in
triplicate.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
78
In the second essay, the 17 fenamiphos degrading bacteria that were isolated from soil S3 in
chapter 5 were grown on TSA+ for 48 h in dark conditions and used for inoculation onto
SEAM plates supplemented with Nemathorin 10WG to a final concentration of 100 µg ml-1
a.i. The incubation conditions, sampling and HPLC analysis were performed as described in
section 5.2.2. Samples were taken 0, 7 and 14 days after incubation. All samples were
compared to the treatment with Nemathorin 10WG and no single colony inoculation. Each
treatment was done in triplicate.
6.3. Results
6.3.1. Degradation of fosthiazate
The microorganisms present in the fenamiphos-degrading soil S3 were tested for their
fosthiazate degrading abilities in an assay with soil extract liquid medium, supplemented with
100 µg ml-1 fosthiazate. This showed that in non-inoculated medium the fosthiazate was
degraded to approximately 50% of the initial concentration after a 30 days incubation period
(Figure 1). Fosthiazate can also be degraded by abiotic factors. However, in the presence of
the microorganisms obtained from S3, fosthiazate was reduced by 70% of the initial
concentration after a 30 days incubation period. This accelerated degradation in the presence
of microorganisms was observed throughout the whole incubation period. Apparently, biotic
factors present in soil S3 are involved in the accelerated degradation of fosthiazate even
though this is the first time that the microorganisms are in contact with this specific
nematicide.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
79
0102030405060708090
100
0 3 6 9 13 16 20 30
Days of incubation after nematicide application
Fost
hiaz
ate
conc
entra
tion
[%]
from
inic
ial a
pplic
atio
nInoculated with all microorganisms of S3 Not inoculated
Figure 1. Degradation of fosthiazate applied to an initial concentration of 100 µg ml-1 in soil extract liquid medium alone or when inoculated with microorganisms of soil S3, which contained fenamiphos-degrading organisms. N = 3. Bars indicate the Standard Error of the Mean.
6.3.2. The effect of fosthiazate on fenamiphos degradation
To determine the effect on biodegradation when two organophosphates were present, the
microorganisms extracted from S3 were incubated in the presence of both fenamiphos and
fosthiazate and the degradation of fenamiphos was followed over time and compared to the
S3 microbial extracts, incubated with fenamiphos alone. In the control treatments, in which
no microorganisms were inoculated or the microbial extracts were autoclaved prior to
inoculation, in the presence of only fenamiphos or both organophosphates, no fenamiphos
metabolization could be observed during the 20 days incubation period (Figure 2). When
fenamiphos was present alone, the degradation of the organophosphate was visible 6 days
after the start of the incubation. At this time, the fenamiphos degradation was not yet visible
when both organophosphates were present. However, for both treatments the fenamiphos was
completely degraded after 13 days. Although, there was some delay in the initiation of the
fenamiphos biodegradation in the presence of fosthiazate, this delay was apparently
compensated by a higher biodegradation activity when finally initiated.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
80
0
20
40
60
80
100
0 3 6 9 13 20
Days of incubation after nematicide treatment
Fena
mip
hos
cone
cntra
tion
[%]
com
pear
ed to
the
treat
men
t with
no
mic
roor
gani
sms
Foz+Fen alone Fenamiphos with autoclaved S3 Foz+Fen with autoclaved S3 Fenamiphos with S3 Foz+Fen with S3
Figure 2. Metabolization of fenamiphos (Fen) in soil extract liquid medium, in the absence or presence of fosthiazate (Foz) and which was not inoculated with microorganisms, inoculated with microorganisms extracted from S3, a soil containing fenamiphos degrading microorganisms or inoculated with an autoclaved suspension containing the microorganisms extracted from S3. N = 3. Bars indicate the Standard Error of the Mean.
6.3.2. Degradation of fosthiazate by fenamiphos-degrading bacteria
The 17 fenamiphos degrading bacteria that were isolated from soil S3 in chapter 5 were
tested for their ability to degrade fosthiazate in an assay with soil extract liquid medium at
100 µg ml-1. HPLC analysis showed that throughout the 14 days incubation period none of
these isolates was capable of degrading fosthiazate (Figure 3).
0102030405060708090
100
Microb
acter
ium
Sinorhi
zobiu
m
Microb
acter
ium
Microb
acter
ium
Brevun
dimon
as
Microb
acter
ium
Microb
acter
ium
Ralston
ia
Cupria
vidus
Ralston
ia
Microb
acter
ium
Microb
acter
ium
Microb
acter
ium
Cupria
vidus
Microb
acter
ium
Microb
acter
ium
Microb
acter
ium
Genus of the fenamiphos-degrading bacteria
Fost
hiaz
ate
conc
entr
atio
n [%
] co
mpa
red
to tr
eatm
ent w
ith n
o ba
cter
ia
0 7 14
Figure 3. Percentage of remaining fosthiazate applied to an initial concentration of 100 µg ml-1 as detected by HPLC analysis in soil extract agar medium 0, 7 and 14 days after inoculation with one of the 17 fenamiphos-degrading bacteria isolated and identified in chapter 5. N = 3.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
81
6.4. Discussion
Fosthiazate is a novel organophosphate which is also marketed in nematicidal formulation.
The results in this investigation suggest that abiotic factors can cause degradation of
fosthiazate. However, the degradation rate is not considered enhanced since the half-life of
this organophosphate nematicide in soil has been reported to range between 0.5 to 1.5 months
(Pantelelis et al., 2006). Furthermore it is also shown that fosthiazate may be degraded by
biotic factors. These results are in accordance with other fosthiazate studies, since the
presence of microorganisms has been shown to accelerate the degradation process.
Elimination of microorganisms by sterilization resulted not only in an increase of fosthiazate
persistence in soil but also in a decrease of its degradation rate (Qin et al., 2004).
Additionally, soil autoclaving was reported to double the half-life of fosthiazate (Pantelelis et
al., 2006).
The soil (S3) which was frequently exposed to fenamiphos and contains microorganisms that
can degrade this organophosphate is also capable of biodegrading fosthiazate, although not as
efficient. Even though currently these microorganisms may not be that important in the
fosthiazate degradation, it is still remarkable that such organisms are already present in a soil
that had never been exposed to this organophosphate. This observation may indicate that the
application of fosthiazate will quickly lead to the accumulation of fosthiazate-degrading
organisms and, consequently, to the accelerated biodegradation of the compound.
The bacteria that were found to be very effective in the degradation of fenamiphos could not
degrade fosthiazate, thus no indications were found of cross-biodegradation. Apparently,
fenamiphos degrading bacteria were rather specialized. Completely different microorganisms
or types of microorganisms are involved and the degradation of each organophosphate may
therefore require different enzymes (pathways). Bacteria from the genus Enterobacter,
Flavobacterium, Agrobacterium, Bacillus and Rhizobium, among others, have been reported
to degrade organophosphorus compounds (Singh and Walker, 2006). In addition, not only
bacteria have been shown to rapidly degrade xenobiotics. Fungi from the genus
Phanerochaete, Hypholama, Coriolus, Spergillus, Trichoderma and Pencillium, for example,
have also bee reported to degrade organophosphorus pesticides (Singh and Walker, 2006).
The results obtained in the present study suggest that the phenomena of cross-conditioning is
not that important in the soil and therefore, the application history of one organophosphate
nematicide for a soil is not informative regarding the development of biodegradation for

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
82
another organophosphate. This theory is supported by similar results of pesticide specificity
degrading microorganisms reported with other organophosphate pesticides. Racke and Coats
(1988) isolated Artrhobacter spp. from a soil with large history of isofenphos use. This
bacterium was capable to degrade isofenphos at accelerated rates but could not degrade
chlorpyrifos, fonofos, ethoprop, terbufos and phorate. In soils with large ethoprophos history
this nematicide was rapidly biodegraded but degradation of fenamiphos, fonofos, cadusafos,
isazofos, aldicarb and oxamyl did not occurred (Karpouzas and Walker, 2000). Furthermore,
the evidence in all the above studies supports the hypothesis of bacterial adaptation to a
specific compound. This theory would also explain why the presence of both
organophosphates at the same time did not affect the degradation of fenamiphos, either
positively or negatively, since different organisms are involved in the degradation process.
Although some delay in the initiation of fenamiphos biodegradation was observed in the
presence of fosthiazate, the complete biodegradation occurred in the same time frame.
Fosthiazate may have some initial restrictive impact on the proliferation of the fenamiphos
degrading microorganisms, but this effect disappears completely in the course of time.
It can be concluded from this study that fosthiazate biodegradation process can be developed
by microorganisms even if they are exposed to this nematicide for the first time, that the
fosthiazate-degrading microorganisms are different from those degrading fenamiphos and
that the nematicide application history in soil of one nematicide does not necessarily
influence the degradation rate of another one. From an agricultural point of view, these
observations are of importance since it would suggest that the efficacy of nematicides with
fosthiazate could rapidly decline even if it is applied for the first time. The emergence of new
alternatives to slow down the enhanced biodegradation process is important and therefore is
the focus of the next chapter of this thesis.
6.5. References
Harris, C.R., Chapman, R.A., Harris, C. and Tu, C. 1984. Biodegradation of pesticides in soil:
rapid induction of carbamate degrading factors after carbofuran treatment. Journal of
Environmental Sciences and Health, Part B 19:1-11.
Hafez, S.L. and Sundararaj, P. 2006. Efficacy of fosthiazate for the control of
Paratrichodorus spp. and Meloidogyne chitwoodi on potato. International Journal of
Nematology 16:157-160.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
83
Karpouzas, D.G., Giannakou, I.O., Walker, A. and Gowen, Simon. 1999a. Reduction in
biological efficacy of ethoprophos in a soil from Greece due to enhanced
biodegradation: comparing bioassay with laboratory incubation data. Pesticide Science
55:1089:1094.
Karpouzas, D.G., Walker, A., Foud-Williams, R. J. and Drennan, D.S.H. 1999b. Evidence for
the enhanced biodegradation of ethoprophos and carbofuran in soils from Greece and
the UK. Pesticide Science 55:1089:1094.
Karpouzas, D.G. and Walker, A. 2000. Factors influencing the ability of Pseudomonas putida
strains epI and II to degrade the organophosphate ethoprophos. Journal of Applied
Microbiology 89:40-48.
Karpouzas, D.G., Fotopoulou, A., Menkissoglu-Spiroudi, U. and Singh, B.K. 2005. Non-
specific biodegradation of the organophosphorus pesticides, cadusafos and
ethoprophos, by two bacterial isolates. FEMS Microbiology Ecology, 53:369-378.
Morel-Chevillet, C., Parekh, N.R., Pautrel, D. and Fournier, J.C.1996. Cross-enhancement of
carbofuran biodegradation in soil samples previously treated with carbamate
pesticides. Soil Biology and Biochemistry 28:1767-1776.
Pantelelis I., Karpouzas D.G., Menkissoglu-Spiroud U. and Tsiropoulos, N. 2006. Influence
of soil physicochemical and biological properties on the degradation and adsorption of
the nematicide fosthiazate. Journal of Agricultural Food Chemistry 54:6783-6789.
Racke, K.D. and Coats, R.J. 1988. Comparative degradation of organophosphorus insecticides
in soil: specificity of enhanced microbial degradation. Journal of Agricultural Food
and Chemistry 36:193-199.
Rich, J.R. Dunn, R.A. and Noling, J.W. 2004. Nematicides: past and present uses. In:
Nematology: advances and perspectives. Volume 2: Nematode management and
utilization. Chen, Z.X., Chen, S.Y. and Dickson D.W. (eds.), 1179-1200.
Rich, J. R. and Dunn, R.A., Thomas, W.D., Breman, J.W. and Tervola, R.S. 1994. Evaluation
of fosthiazate for management of Meloidogyne javanica in Florida flue-cured tobacco.
Journal of Nematology 26:701-704.
Suett, D.L. 1987. Influence of treatment of soil with carbofuran on the subsequent
performance of insecticides against cabbage root fly (Delia radicum) and carrot fly
(Psila rosae). Crop Protection 6:371-378.
Suett, D.L. 1989. Accelerated degradation of soil-applied insecticides: implications for insect
pest control. Mededelingen van de Faculteit Landbouwwetenschappen
Rijksuniversiteit Gent 54:983-991.

Chapter 6 Fosthiazate cross-degradation and nematicide specificity of bacteria
84
Tobin, J.D., Haydock, P.P.J., Hare, M.C., Woods, S.R. and Crump, D.H. 2008. Effect of the
fungus Pochonia chlamydosporia and fosthiazate on the multiplication rate of potato
cyst nematodes (Globodera pallida and G. rostochiensis) in potato crops grown under
UK field conditions. Biological Control 46:194-201.
Woods, S.R., Haydock, P.P.J. and Edmunds, C. 1999. Mode of action of fosthiazate used for
the control of potato cyst nematode Globodera pallida. Annals of Applied Biology
135:409-415.
Woods, S.R. and Haydock, P.P.J. 2000. The effect of granular nematicide incorporation depth
and potato planting depth on potatoes grown in land infested with the potato cyst
nematodes Globodera rostochiensis and G. pallida. Annals of Applied Biology
136:27-33.
Qin, S. Gan, J., Liu, W. and Becker, J.O. 2004. Degradation and adsorption of fosthiazate in
soil. Journal of Agricultural and Food Chemistry 52:6239-6242.

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
85
7. Effect of natural plant enhancers on soil bacteria, Meloidogyne incognita and nematicide degradation
7.1. Introduction
The root-knot nematode Meloidogyne incognita is distributed worldwide (Sasser and Carter,
1985) and is known to be a major pest in a wide variety of crop plants. Among the host
plants, lettuce is one of the vegetables that can be seriously affected by M. incognita
(Sikora and Fernandez, 2005). To control plant-parasitic nematodes non-fumigant
nematicides are presently preferred over the fumigant nematicides due to their lower
environmental hazard (Hague and Gowen, 1987). However, the efficacy of non-fumigant
nematicides has been reduced in recent times due to adapted soil microorganisms capable of
degrading the nematicides, which is caused by the prolonged and intensive application
(Karpouzas et al., 2000; Karpouzas and Giannakou, 2002) as described in previous chapters.
At present, there are some strategies available to maintain the efficacy of nematicides or to
delay the onset of biodegradation. In soils of the Ivory Coast, the degradation rate of
fenamiphos returned to the level found in previously untreated soils when the prolonged
application of this pesticide were interrupted for 12 to 16 months (Anderson et al., 1992).
Furthermore, to avoid redevelopment of accelerated degradation, fenamiphos application was
alternated every second treatment with a different organophosphate nematicide. In Australia,
a rotation in nematicide applications showed to delay the development of enhanced
biodegradation in soils (Pattison and Cobon, 2003). Ample time intervals between application
of the same nematicide, and repressing the disease incidence by crop rotation, could reduce
the need for pesticide applications and thus further delay the development of an increased
pesticide biodegradation (Smelt and Leistra, 1992). However, in intensive agriculture it may
in some cases be unpractical to postpone nematicide applications for a long time, especially
when the nematode numbers increase exponentially within time, thus drastically reducing
crop yield. Additionally, in regions where farmers get attractive prices when buying products
from the same chemical company, there is often little rotation in the application of pesticides.
Organic amendments have been used worldwide to positively affect soil properties and,
ultimately, crop quality (Muller and Gooch, 1982). Among the organic amendments, plant

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
86
revitalizers, also known as plant enhancers, have been identified to stimulate microbial
activity in soil and promote plant growth (Mulawarman et al., 2001). Additionally, it has
been reported that the plant revitalizers promote growth of plant-beneficial microorganisms,
such as mycorrhizal fungi, and reduce to some extent infection of M. incognita (Mulawarman
et al., 2000). Soil amendments can thus reduce the presence of soil born pathogens such as
plant-parasitic nematodes.
There is an increasing demand for plant enhancers with multiple beneficial effects, like to
provide nutrients to the plant, increase the antagonistic potential of the soil and delay, stop or
reverse the nematicide enhanced biodegradation process. Application of the plant revitalizers
may be capable of shifting a nematicide-degrading microbial community into a non-
degrading community and/or provide additional carbon sources, that could be used by the
microorgasnisms in order to reduce the use of nematicides as carbon source. Therefore the
objectives of this research were to:
1. Determine the effect of different inoculum densities of Meloidogyne incognita on
lettuce cv. Milan.
2. Quantify the shifts in bacterial populations caused by the application of natural plant
enhancers.
3. Establish the biocontrol activity of plant revitalizers on early root penetration of M.
incognita.
4. Evaluate the effect of natural plant enhancers on the biodegradation of fenamiphos.
7.2. Materials and Methods
7.2.1. Effect of different M. incognita inoculum densities on lettuce cv. Milan
To determine the susceptibility of lettuce cv. Milan to Meloidogyne incognita a greenhouse
bioassay was initiated, in which plastic pots with a diameter of 10 cm were filled with 250 ml
of autoclaved mixture of soil S4 and sand, in a 1:1 (v:v) ratio and supplemented with 7%
compost. Per pot 50 lettuce seeds cv. Milan were sown. After sowing, approximately 40 ml
of autoclaved sand was put on top of the seeds for germination and the pots were gently
watered. 1000, 3000, 5000 or 7000 M. incongita J2 were inoculated per pot through 3 cm

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
87
deep holes in the soil around the lettuce plants 1 week after sowing. Non infested pots were
used as control. The lettuce plants were harvested 4 weeks past nematode inoculation and the
shoot fresh weight was recorded. Roots were gently washed, weighted and the gall index
(Zeck, 1971) with a scale from 0 (no galls) to 10 (completely galled root) was determined per
root system. Each treatment had 6 replicates. All treatments were distributed in a randomized
block design under greenhouse conditions at 26 ± 2 ºC. Plants were watered with tap water
when necessary.
7.2.2. Effect of plant enhancers on soil bacteria, Meloidogyne incognita and biodegradation
7.2.2.1. Natural plant enhancers and nematicides
The tested plant commercial enhancers were MagicWet®, TerraPy® (Cognis Deutschland
GmbH, Germany), Azet® (W. Neudorff GmbH KG, Germany), and Oscorna® (Oscorna-
Dunger GmbH & Co. KG., Germany). MagicWet® is a liquid formulation of sugar based
surfactants and fatty acid derivatives and TerraPy® is a liquid formulation composed of plant
lipids, sugar based surfactants and organic bounds of nitrogen and phosphorous
(Mulawarman et al., 2001). Azet® and Oscorna® are granular formulated plant enhancers.
Azet® is a natural compost produced using animal and plant material. Oscorna® is composed
of coral algae lime, clay minerals and extracts of primary rocks. Chicken manure and green
compost were produced in the University of Bonn, Germany. As nematicides the biological
DiTera® DF and the non-fumigant nematicide Nemacur® 5G were used in this research.
7.2.2.2. Soil treatment with plant enhancers and nematicides
Soil S4 was supplemented with 7% (w:w) organic matter and plastic bags were filled with 6
kg of soil each. Every bag was treated with Oscorna® (450 kg ha-1), Azet® (300 kg ha-1),
Compost (5,000 kg ha-1, Joshi et al., 2000), Chicken manure (5,000 kg ha-1, Tüzel et al.,
2003), Diterra® (112 kg ha-1), Magic Wet® (200 kg ha-1, Mulawarman et al., 2001),
Terrapy® (200 kg ha-1 Mulawarman et al., 2001) or Nemacur® 5G (120 kg ha-1) once at the
beginning of the experiment. As a control, one plastic bag with soil was not treated. Tap
water was added to all soils to reach field capacity immediately after treatment. After
watering, all plastic bags containing the soil were vigorously mixed, thus distributing the
amendments evenly and incubated in the green house at 27 ± 5 ºC until further use.

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
88
7.2.2.3. Effect on soil bacteria population densities
One day before the soil was treated, the bacteria present in the soil were isolated by spreading
it onto TSA+ as described on section 2.4. in triplicate. One and three weeks after soil
treatment, bacteria were isolated as described above. After 24 h of incubation the number of
colony forming units was determined in every treatment.
7.2.2.4. Effect on M. incognita early root penetration
From every soil treatment 250 ml of soil was taken and transferred to 10 cm diameter plastic
pots at 0, 2 and 4 weeks after soil treatment. Every pot was inoculated through three 3 cm
deep holes with a suspension containing 2500 well developed M. incongita eggs. The holes
were closed, approximately 50 seeds of lettuce cv. Milan were sown per pot and the seeds
were covered with autoclaved sand to provide adequate germination conditions. Plants were
watered with tap water when necessary. Plants were harvested 20 days after planting and the
roots were gently washed with tap water, stained with lactic acid-fuchsin (768 ml lactic acid,
56 ml glycerol, 1 g acid fuchsin and 154 distilled water) and heated in a microwave for 2 min.
Then roots were cut into 1 cm pieces and further fragmented in tap water using an Ultra-
Turrax (IKA-Werk) at 11,000 rpm for 1.5 minutes. The suspension was adjusted to 100 ml
and the number of penetrated nematodes per root system was determined. The experiment
was arranged in a randomized complete block design with each treatment replicated four
times and conducted under greenhouse conditions at 26 ± 5 °C.
7.2.2.5. Effect on enhanced biodegradation of fenamiphos
Soil S3, containing fenamiphos-degrading microorganisms, was used to determine whether
the plant enhancers could delay or stop the biodegradation process. For this, 20 g soil samples
of S3 were taken and transferred to sterile plastic beakers. Every sample was treated with one
of the plant enhancers at the equivalent dosage described in section 7.2.2.2. with the
exception of chicken manure which was not utilized in this test. A sample with no plant
enhancer application was used as control. Sterile water was added to the soil samples up to
holding capacity. The beakers were closed with a sterile cap, vigorously shaken and kept in
the green house at 26 ± 5 ºC for one week. Then, 10 g of each sample were transferred to 100
ml of sterilized distilled water in Erlenmeyer flasks. The suspensions were incubated for 16

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
89
hours in the dark on a rotary shaker at 60 rpm at 28 ˚C. Subsequently, 1 ml suspension was
transferred into 19 ml of SELM supplemented with 100 µg ml-1 fenamiphos analytical grade.
From this medium samples were taken 0, 6, 10 and 14 days after inoculation. Incubating
conditions, sampling and HPLC analysis were performed as described in the second test of
section 4.2.5.
7.2.3. Statistical analysis
To determine the effect of each treatment in time the data obtained in the greenhouse and
laboratory tests was analyzed by linear regression using Sigma Plot (version 8.02, SPSS, Inc.,
Chicago). The number of bacterial colony forming units was evaluated by one-way ANOVA
(p<0.05). When significant differences were detected Fisher’s Least Significant Difference
(LSD) was performed.
7.3. Results
7.3.1. Effect of different M. incognita inoculum densities on lettuce cv. Milan
In a greenhouse bioassay, pots containing lettuce plants were inoculated with different
number of M. incognita J2 and the gall index was determined 4 weeks after nematode
inoculation. This assay revealed that the gall index made by Meloidogyne incognita increased
from 4.0 when 1000 nematodes were inoculated to 6.8 when 7000 nematodes were inoculated
(Figure 1). The non-linear regression analysis showed a significant relationship, with a
correlation coefficient (r2 ) of 0.97, between increasing nematode inoculum densities and a
higher gall index.
Shoot fresh weight was significantly reduced from 36 to 29 g in the treatments with 0 and
7000 nematodes inoculated (Figure 2). The linear regression analysis showed a significant
relationship with r2 value of 0.91, between the increase nematode inoculum densities and the
reduction in shoot fresh weight. In addition, root fresh weight was significantly reduced by
the different inoculation treatments from 10 g in the non-inoculated control to 8.8 g when
inoculated with 7000 nematodes. However, the root weight was less affected than the shoot
weight and only showed an r2 value of 0.51.
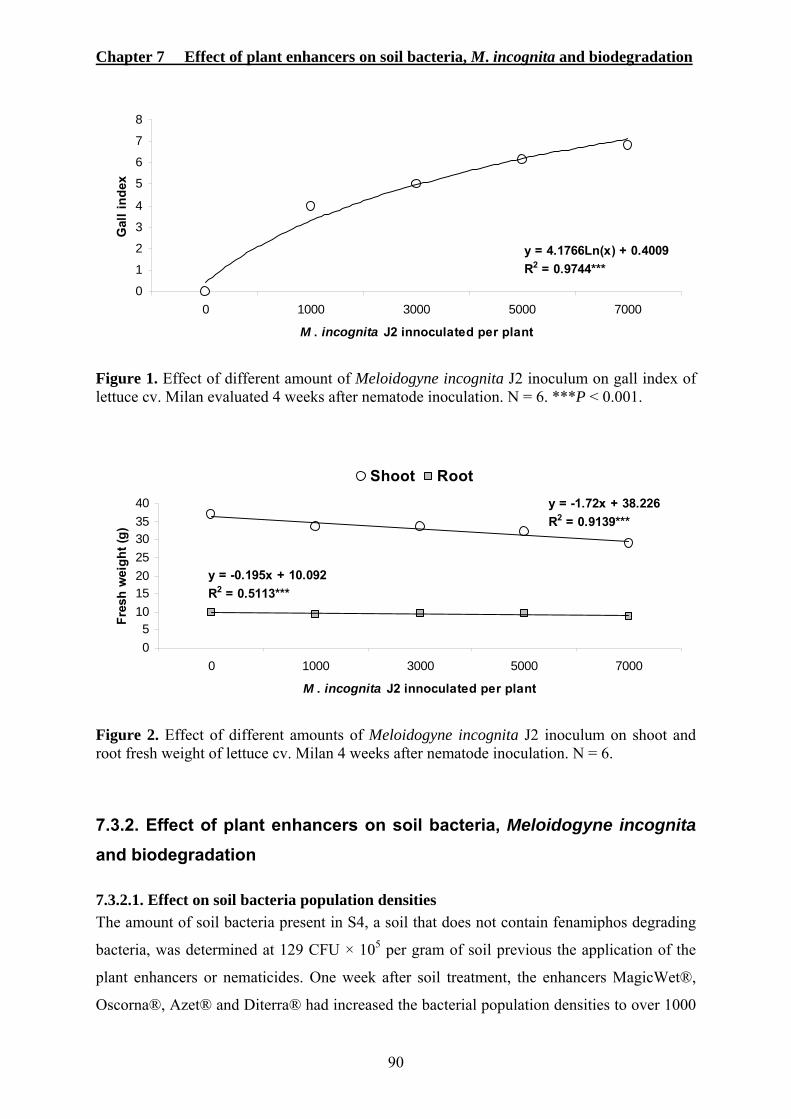
Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
90
y = 4.1766Ln(x) + 0.4009R2 = 0.9744***
0
1
2
3
4
5
6
7
8
0 1000 3000 5000 7000
M . incognita J2 innoculated per plant
Gal
l ind
ex
Figure 1. Effect of different amount of Meloidogyne incognita J2 inoculum on gall index of lettuce cv. Milan evaluated 4 weeks after nematode inoculation. N = 6. ***P < 0.001.
y = -1.72x + 38.226R2 = 0.9139***
y = -0.195x + 10.092R2 = 0.5113***
05
10152025303540
0 1000 3000 5000 7000
M . incognita J2 innoculated per plant
Fres
h w
eigh
t (g)
Shoot Root
Figure 2. Effect of different amounts of Meloidogyne incognita J2 inoculum on shoot and root fresh weight of lettuce cv. Milan 4 weeks after nematode inoculation. N = 6.
7.3.2. Effect of plant enhancers on soil bacteria, Meloidogyne incognita and biodegradation
7.3.2.1. Effect on soil bacteria population densities The amount of soil bacteria present in S4, a soil that does not contain fenamiphos degrading
bacteria, was determined at 129 CFU × 105 per gram of soil previous the application of the
plant enhancers or nematicides. One week after soil treatment, the enhancers MagicWet®,
Oscorna®, Azet® and Diterra® had increased the bacterial population densities to over 1000
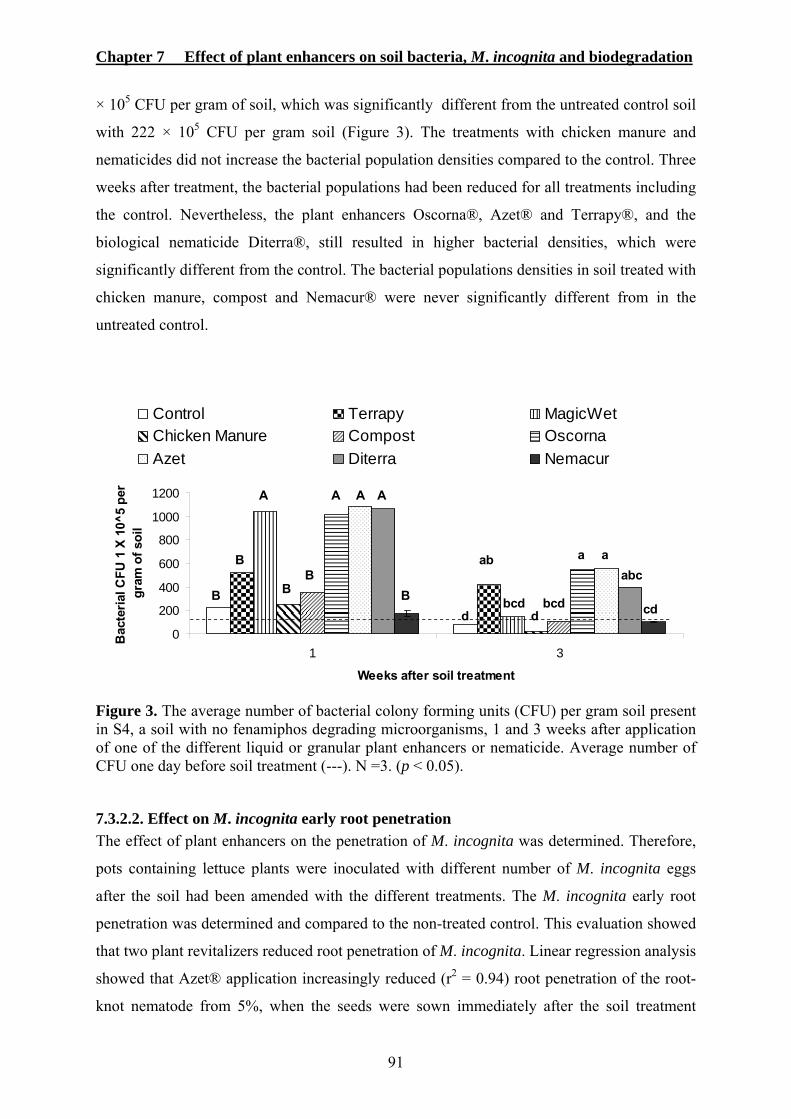
Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
91
× 105 CFU per gram of soil, which was significantly different from the untreated control soil
with 222 × 105 CFU per gram soil (Figure 3). The treatments with chicken manure and
nematicides did not increase the bacterial population densities compared to the control. Three
weeks after treatment, the bacterial populations had been reduced for all treatments including
the control. Nevertheless, the plant enhancers Oscorna®, Azet® and Terrapy®, and the
biological nematicide Diterra®, still resulted in higher bacterial densities, which were
significantly different from the control. The bacterial populations densities in soil treated with
chicken manure, compost and Nemacur® were never significantly different from in the
untreated control.
0
200
400
600
800
1000
1200
1 3
Weeks after soil treatment
Bac
teria
l CFU
1 X
10^
5 pe
r gr
am o
f soi
l
Control Terrapy MagicWetChicken Manure Compost OscornaAzet Diterra Nemacur
B
A A A A
BBB
d d cdbcdbcd
abcab a aB
Figure 3. The average number of bacterial colony forming units (CFU) per gram soil present in S4, a soil with no fenamiphos degrading microorganisms, 1 and 3 weeks after application of one of the different liquid or granular plant enhancers or nematicide. Average number of CFU one day before soil treatment (---). N =3. (p < 0.05).
7.3.2.2. Effect on M. incognita early root penetration The effect of plant enhancers on the penetration of M. incognita was determined. Therefore,
pots containing lettuce plants were inoculated with different number of M. incognita eggs
after the soil had been amended with the different treatments. The M. incognita early root
penetration was determined and compared to the non-treated control. This evaluation showed
that two plant revitalizers reduced root penetration of M. incognita. Linear regression analysis
showed that Azet® application increasingly reduced (r2 = 0.94) root penetration of the root-
knot nematode from 5%, when the seeds were sown immediately after the soil treatment
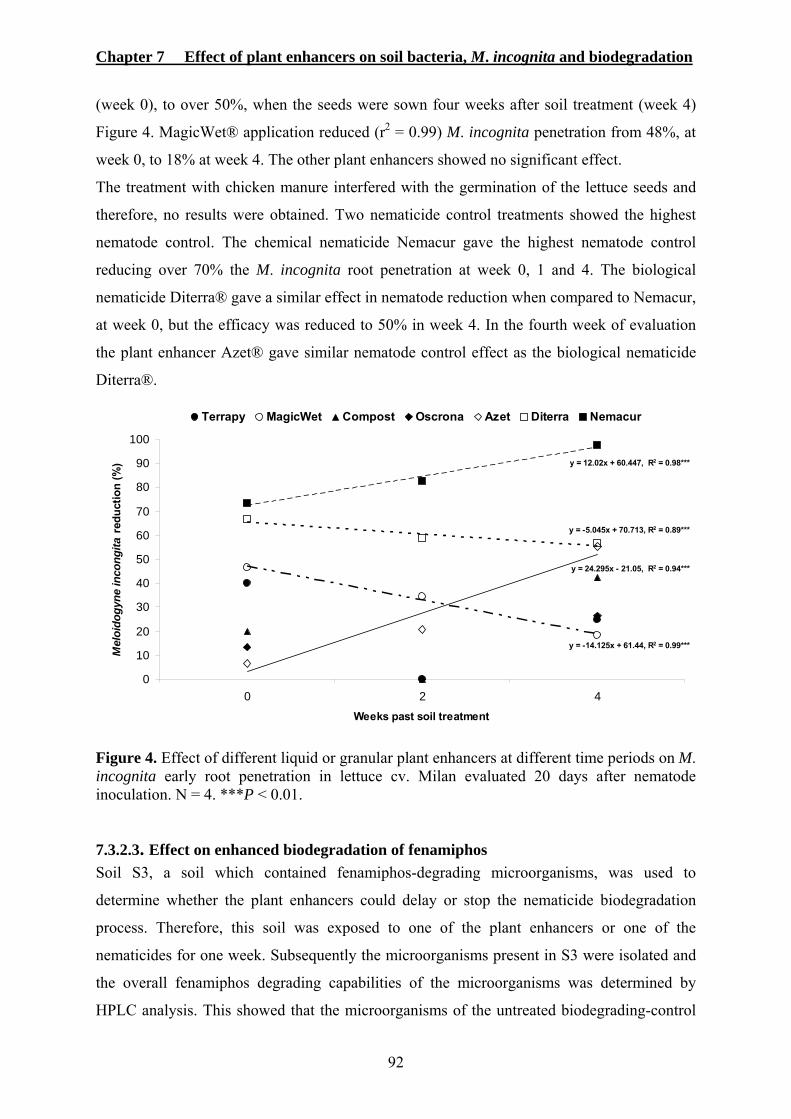
Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
92
(week 0), to over 50%, when the seeds were sown four weeks after soil treatment (week 4)
Figure 4. MagicWet® application reduced (r2 = 0.99) M. incognita penetration from 48%, at
week 0, to 18% at week 4. The other plant enhancers showed no significant effect.
The treatment with chicken manure interfered with the germination of the lettuce seeds and
therefore, no results were obtained. Two nematicide control treatments showed the highest
nematode control. The chemical nematicide Nemacur gave the highest nematode control
reducing over 70% the M. incognita root penetration at week 0, 1 and 4. The biological
nematicide Diterra® gave a similar effect in nematode reduction when compared to Nemacur,
at week 0, but the efficacy was reduced to 50% in week 4. In the fourth week of evaluation
the plant enhancer Azet® gave similar nematode control effect as the biological nematicide
Diterra®.
y = 12.02x + 60.447, R2 = 0.98***
y = -14.125x + 61.44, R2 = 0.99***
y = 24.295x - 21.05, R2 = 0.94***
y = -5.045x + 70.713, R2 = 0.89***
0
10
20
30
40
50
60
70
80
90
100
0 2 4Weeks past soil treatment
Mel
oido
gyne
inco
ngita
redu
ctio
n (%
)
Terrapy MagicWet Compost Oscrona Azet Diterra Nemacur
Figure 4. Effect of different liquid or granular plant enhancers at different time periods on M. incognita early root penetration in lettuce cv. Milan evaluated 20 days after nematode inoculation. N = 4. ***P < 0.01.
7.3.2.3. Effect on enhanced biodegradation of fenamiphos Soil S3, a soil which contained fenamiphos-degrading microorganisms, was used to
determine whether the plant enhancers could delay or stop the nematicide biodegradation
process. Therefore, this soil was exposed to one of the plant enhancers or one of the
nematicides for one week. Subsequently the microorganisms present in S3 were isolated and
the overall fenamiphos degrading capabilities of the microorganisms was determined by
HPLC analysis. This showed that the microorganisms of the untreated biodegrading-control
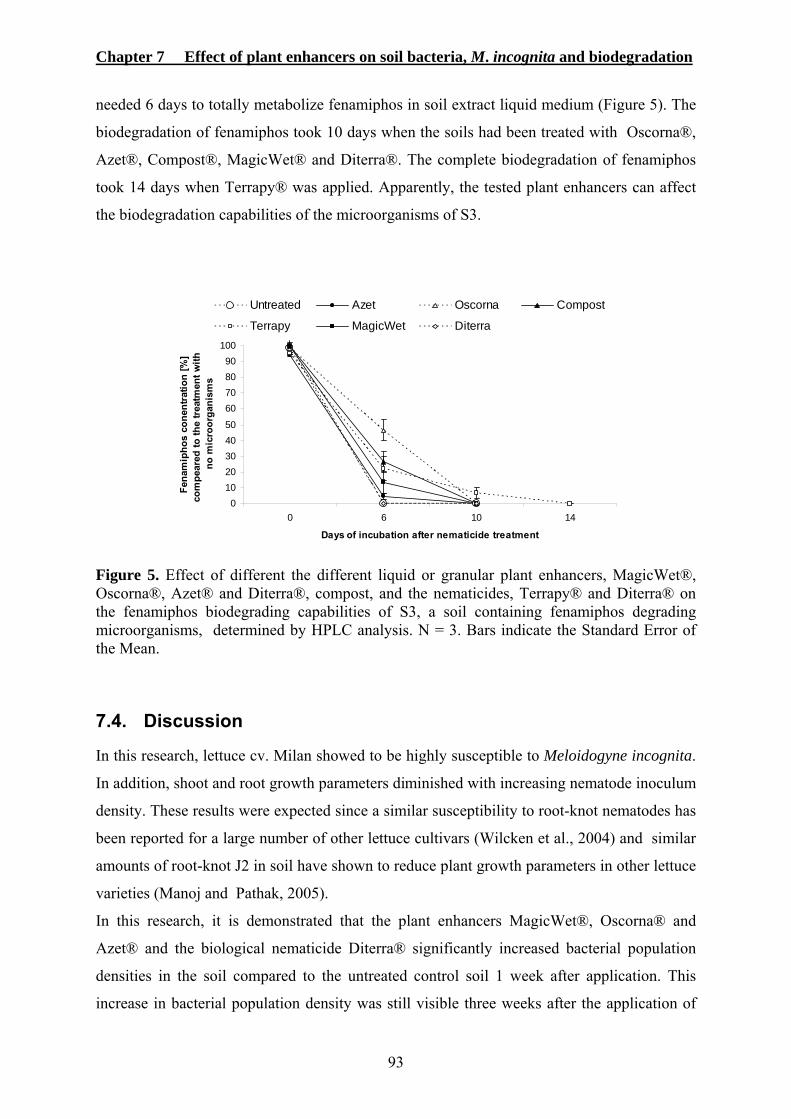
Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
93
needed 6 days to totally metabolize fenamiphos in soil extract liquid medium (Figure 5). The
biodegradation of fenamiphos took 10 days when the soils had been treated with Oscorna®,
Azet®, Compost®, MagicWet® and Diterra®. The complete biodegradation of fenamiphos
took 14 days when Terrapy® was applied. Apparently, the tested plant enhancers can affect
the biodegradation capabilities of the microorganisms of S3.
01020304050
60708090
100
0 6 10 14
Days of incubation after nematicide treatment
Fena
mip
hos
cone
ntra
tion
[%]
com
pear
ed to
the
treat
men
t with
no
mic
roor
gani
sms
Untreated Azet Oscorna Compost
Terrapy MagicWet Diterra
Figure 5. Effect of different the different liquid or granular plant enhancers, MagicWet®, Oscorna®, Azet® and Diterra®, compost, and the nematicides, Terrapy® and Diterra® on the fenamiphos biodegrading capabilities of S3, a soil containing fenamiphos degrading microorganisms, determined by HPLC analysis. N = 3. Bars indicate the Standard Error of the Mean.
7.4. Discussion
In this research, lettuce cv. Milan showed to be highly susceptible to Meloidogyne incognita.
In addition, shoot and root growth parameters diminished with increasing nematode inoculum
density. These results were expected since a similar susceptibility to root-knot nematodes has
been reported for a large number of other lettuce cultivars (Wilcken et al., 2004) and similar
amounts of root-knot J2 in soil have shown to reduce plant growth parameters in other lettuce
varieties (Manoj and Pathak, 2005).
In this research, it is demonstrated that the plant enhancers MagicWet®, Oscorna® and
Azet® and the biological nematicide Diterra® significantly increased bacterial population
densities in the soil compared to the untreated control soil 1 week after application. This
increase in bacterial population density was still visible three weeks after the application of

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
94
Oscorna® and Azet®. Also Terrapy® and Diterra® still showed a significant accumulation
of bacteria. These results confirm that plant enhancers can increase the bacterial populations
rapidly to about 5 times of its original amount, perhaps by providing large amounts of extra
nutrients that soil microorganisms can use to proliferate. These results are in accordance with
those of Mulawarman et al. (2001) who reported that some of these commercially available
plant enhancers, such as Terrapy® and MagicWet®, stimulate soil microbial activity. In a
previous study, Diterra® showed to stimulate, fungi and bacteria in soil and rhizosphere over
time (Fernandez et al., 2001.). Moreover, even though the response of microbial communities
depends on different factors, such as the amendment composition (Pereq-Piqueres et al.,
2006), it could be expected that the effect of the plant revitalizers on microorganisms would
be similar in different soils since the plant enhancers are uniformly produced.
The two plant enhancers that reduced the root infection of M. incognita to lettuce roots were
Azet® and MagicWet®. Azet® gave an increasing nematode reduction, over time, that ended
with similar levels of control as that given by the biological nematicide Diterra®. This
suggests that Azet® can enhance the antagonistic potential of the soil for biological control of
M. incognita by enhancing beneficial microorganisms over time. MagicWet® reduced M.
incognita penetration to 50% immediately after treatment but its effectiveness was reduced
significantly 2 and 4 weeks after application. These results confirm that MagicWet®
application stimulates soil microbial activity and thereby the antagonistic potential in soils
leading to a reduction in nematode infestation as previously reported by Mulawarman et al.
(2001). However, its effect on root penetration is not maintained for a long period of time.
This is the first time that Azet® is found to reduce nematode penetration and it is believed
that this biocontrol activity is similar to that of MagicWet®. However, the biocontrol activity
of Azet® appears to increase continuously over time and therefore it may be important to
evaluate its effect over a longer time period. Moreover, it was believed that the other plant
revitalizers would not affect nematode root penetration because the microorganisms involved
in this process do not utilize or profit from their formulations.
The most effective treatment for controlling M. incognita penetration was still the chemical
nematicide Nemacur, with fenamiphos as active ingredient. The high efficacy of fenamiphos
can be explained, since it is well known that it inhibits nematode movement and suppresses
the hatching of nematode eggs (Van Gundy and Mckenry, 1977). However, the efficacy of
fenamiphos could rapidly deteriorate when the soil is frequently treated with fenamiphos for
an extensive period since then adapted fenamiphos degrading microorganisms may

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
95
accumulate (Ou and Rao, 1986; Ou, 1991; Ou et al., 1994; Pattison et al., 2000; Stirling et al.,
1992).
It was shown that the application of plant revitalizers delayed the biodegradation of
fenamiphos. In particular, the application of the liquid plant enhancer Terrapy® delayed the
complete metabolization of fenamiphos by a factor of 2.5. The previous two tests showed that
plant revitalizers can stimulate specific bacterial population densities. Perhaps the non-
biodegrading microorganisms living in S3 increased substantially with Terrapy® treatment
while the degrading ones were suppressed. The actual mode of action that supports such a
specific repression is however difficult to explain at the current moment, but it may be that
fungi, with their more restricted biodegrading activity, become the more dominating group in
the soil as a consequence of the Terrapy® treatment. Fungi from this soil lacks in fenamiphos
degrading ability as shown in the Chapter 5 of this thesis supporting this theory. However,
even though Terrapy® delayed the complete metabolization to 14 days, it is still not enough
for managing the enhanced biodegradation process under field conditions. A nematicide
should remain effective in soil for the first 6 weeks after application in order to provide
adequate nematode control supporting plant development. From the current alternatives
known today to slow down the biodegradation process one is to not apply nematicides in a
period of over 12 months (Anderson et al., 1992). However, this is unpractical in intensive
agriculture. Therefore, the alternative of nematicide rotation remains as an essential strategy
for the management of enhanced nematicide biodegradation (Karpouzas and Walker, 2000b).
Furthermore, the results in this thesis suggest that rotating the use of nematicides together
with the application of natural plant enhancers, such as Azet, could also further suppress
nematode root penetration and delay the onset of enhanced biodegradation.
In conclusion, lettuce cv. Milan was highly susceptible to M. incognita but the root-knot
nematode damage can be reduced by the application of natural plant enhancers such as Azet®
and MagicWet®. The plant enhancers increased soil bacteria populations providing a
biocontrol activity and delayed the total metabolization process of fenamiphos.

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
96
7.5. References
Anderson, J.P.E., Arnold, D.J, Lewis, F. and Torstensson, L., 1992. Microbiological aspects
of accelerated pesticide degradation. Proceedings of the international symposium on
environmental aspects of pesticide microbiology, Sigtuna. Sweden Uppsala:
Department of Microbiology, Swedish University of Agricultural Sciences. Pp. 184-
192.
Fernandez, C., Rodriguez-Kabana, R., Warrior, P. and Kloepper, J.W. 2001. Induced soil
suppressiveness to a root-knot nematode species by a nematicide. Biological Control
22:103-114.
Hague N.G.G. and Gowen R.S. 1987. Chemical Control of nematodes. In: Principles and
Practice of Nematode Control in Crops. Academic Press Australia. Brown, R.H. and
Kerry, B.R editors. 131-178.
Hussey, R. and Barker, K., 1973. A comparasion of methods of collectin inocula of
Meloidogyne spp., including a new technique. Plant Disease Reporter 57:1025-1028.
Joshi, S.D., Jadhav, A.S., Patil, M.B and Kurundkar, B.P. 2000. Effect of organic
amendment and fly ash on root-knot disease of tomato. Journal of Maharashtra
Agricultural Universities 25:84-85.
Karpouzas, D.G., Morgan, J.A.W. and Walker, A. 2000. Isolation and characterization of 23
carbofuran-degrading bacteria from soils from distant geographical areas. Letters in
Applied Microbiology 31:353-358.
Karpouzas, D.,G. and Giannakou, I.O. 2002. Biodegradation and enhanced biodegradation: a
reason for reduced biological efficacy of nematicides. Russian Journal of Nematology
10:59-78.
Karpouzas, D.G. and Walker, A. 2000a. Factors influencing the ability of Pseudomonas
putida strains epI and II to degrade the organophosphate ethoprophos. Journal of
Applied Microbiology 89:40-48.
Karpouzas, D.G. and Walker, A. 2000b. Aspects of the enhanced biodegradation and
metabolisms of ethoprophos in soil. Pest Management Science 56:540-548.
Manoj K. and Pathak, K. N. 2005. Influence of Meloidogyne incognita on germination,
seedling emergence and plant growth of lettuce, Lactuca sativa Linn. Annals of Plant
Protection Sciences 13:450-453.

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
97
McSorley, R. Stansly, P.A., Noling, J.W., Obreza, T.A. and Conner, J. N. 1997. Impacts of
organic soil amendments and fumigation on plant-parasitic nematodes in a southwest
Florida vegetable field. Nematropica 27:181-189.
Mulawarman, Hallmann, J., Shuster, R.P., Bell, D., Kopp-Holtwiesche, B. and Sikora, R.A.
2000. TerraPy®, a natural product with plant growth enhancing effect and nematode
control. Mededelingen Faculteit Landbouwkundige en Toegepaste Biologische
Wetenschappen, University of Gent 65:537-544.
Mulawarman, Hallmann, J., Bell, D., Kopp-Holtwiesche, B. and Sikora, R.A. 2001. Effects of
natural products on soil organisms and plant heath enhancement. Mededelingen
Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, University of
Gent 66:609-617.
Muller, R. and Gooch, P.S. 1982. Organic amendments in nematode control. An examination
of the literature. Nematropica 12:319-326.
Pattison, A.B. and Cobon, J.A. 2003. Integrated systems for managing nematodes on bananas.
Final Report. Horticulture Australia Limited. Queensland Department of Primary
Industries.
Ou, L.T. 1991. Interactions of microorganisms and soil during fenamiphos degradation. Soil
Science Society of America Journal 55:716-722.
Ou, L.T. and Rao P.S.C. 1986. Degradation and metabolism of oxamyl and phenamiphos in
soils. Journal of Environmental Sciences and Health B 21:25-40.
Ou, L.T., Thomas J.E., Dickson, D.W. 1994. Degradation of fenamiphos in soil with a history
of continuous fenamiphos applications. Soil Science Society of America Journal
58:1139-1147.
Pattison, A.B., Stanton, J.M. and Cobon, J.A. 2000. Bioassay for enhanced biodegradation of
nematicides in soil. Australasian Plant Pathology 29:52-58.
Perez-Piqueres, A., Edel-Hermann, V., Alabouvette, C. and Steiberg, C. 2006. Response of
soil microbial communities to compost amendments. Soil Biology and Biochemistry
38:460-470.
Sasser, J.N. and Carter, C.C. 1985. An advanced treatise on Meloidogyne. Vol 1.: Biology
and Control. North Carolina, United States of America.
Sikora, R.A. and Fernandez, E. 2005. Nematodes parasites of vegetables. In Plant parasitic
nematodes in tropical and subtropical agriculture, second edition. Luc, M., Sikora, R.,
and Bridge, editors. CAB International, Wallingford, U.K. 319-392.

Chapter 7 Effect of plant enhancers on soil bacteria, M. incognita and biodegradation
98
Smelt, J.H. and Leistra, M. 1992. Availability, movement and accelerated transformation of
soil-applied nematicides. In: Nematology from molecule to ecosystem. Gommers, F.,J.
and Maas, P.W. (eds). European Society of Nematologist, Inc. Inwergrowie, Dundee,
Scotland. 266-280.
Striling, G.R. 1991. Biocontrol of plant parasitic nematodes. CAB International, Wallingford,
U.K. 283pp.
Stirling, A.M., Stirling, G.R. and Macrae I.C. 1992. Microbial degradation of fenamiphos
after repeated application to tomato growing soil. Nematologica 38:245-254.
Tüzel, Y., Yagmur, B. and Gümüs, M. 2003. Organic tomato production under greenhouse
conditions. Acta Horticulturae 614:775-780.
Van Gundy, S.D. and Mckenry, M.V. 1977. Action of nematicides. In: Plant disease: An
advanced treatise. Horsfall, J.D. and Cowling, E.B. editors. Academic Press, New
York, USA. 263-283.
Wilcken, S.R.S., Garcia, M.J. de M. and Silva, N. da. 2004. Reproduction of Meloidogyne
incognita race 2 in lettuce (Lactuca sativa L.). Arquivos do Instituto Biológico (São
Paulo) 71:379-381.
Wilcken, S.R.S., Garcia, M.J. de M. and Silva, N. da. 2005. Resistance of lettuce to
Meloidogyne incognita race 2. Nematologia Brasileira 29:267-271.
Zeck, W.M. 1971. A rating scheme for field evaluation of root knot nematode infestation.
Pflanzenschutz-Nachrichten Bayer 24:141-144.

99
Acknowledgements
Firstly, I extend my sincere and eternal gratitude and thankfulness to Prof. Dr. Richard Sikora for supporting and guiding me at all times through the entire process of the doctoral research in Germany and Costa Rica. I appreciate and acknowledge Prof. Dr. M. Becker for accepting to review this thesis work and support me during my studies in Bonn. My special thanks to Dr. Alexander Schouten for his guidance and orientation in the molecular and biochemical tests on this research as well as for his comments on this manuscript. I am grateful with Dr. Luis Pocasangre for receiving me in his research group at Bioversity International/CATIE, Costa Rica, and for his guidance during the experimental field period of this work. I acknowledge the entire INRES-Phytomedizin Böden Ökosystem Team 2006-09 for their outstanding support, cooperation and useful discussions. Many thanks to the INIBAP/CATIE Laboratory Team 2005-06 and CORBANA for their excellent collaboration and assistance during the investigations in Costa Rica. I am very grateful with the German Academic Exchange Service (DAAD) for funding this research throughout a scholarship. Special thanks to Peter Marczok and Olaf Wuestner of Bayer CropScience, Monheim, Germany, for providing soil samples and Nemacur® 5GR used in this investigation. My appreciation to Dr. Brigitte E. Slaats of Syngenta Crop Protection, Stein, Switzerland, for providing the nematicide Fosthiazate used in this research. Valent BioSciences, USA, and Cognis, Germany, are acknowledge for providing DiTera®, and MagicWet® and TerraPy®, respectively. I thank all my colleagues and friends from all around the world who supported me because they made my life in Germany a pleasant and memorable experience. Finally, I thank all my family members for their encouragement and support to reach this goal in my life, because without it, it would have been impossible.


101
Publications
Cabrera, J.A., Sikora, R.A. and Schouten, A. 2009. Involvement of microorganisms other than Pseudomonads on the enhanced degradation of the organophosphate non-fumigant nematicide fenamiphos. Communications in Agricultural and Applied Biological Sciences, in press. Cabrera, J.A., Pocasangre, L.E., Pattison, A.B. and Sikora, R.A. 2009. Terbufos biodegradability and efficacy against Radopholus similis in soils from banana cultivation having different histories of nematicide use, and the effect of terbufos on plant growth of in-vitro propagated Musa AAA cv. Grande Naine. International Journal of Pest Management, in press. Cabrera, J.A., Kiewnick, S., Grimm, C., Dababat, A.A. and Sikora, R.A. 2009. Effective concentration and range of activity of Abamectin as seed treatment against root-knot nematodes in tomato under glasshouse conditions. Nematology, in press. Cabrera, J.A., Kiewnick, S., Grimm, C., Dababat, A.A. and Sikora, R.A. 2009. Efficacy of abamectin seed treatment on Pratylenchus zeae, Meloidogyne incognita and Heterodera schachtii. Journal of Plant Diseases and Protection 116(3):124-128. Cabrera, J.A. and Sikora, R.A. 2009. Effect of natural plant enhancers on soil bacteria and control of plant-parasitic nematodes in lettuce. Journal of Plant Diseases and Protection 116(1):42. Cabrera, J.A., Pocasangre, L.E., Rosales, F., Najera, J., Vargas, R., Serrano, E. and Sikora, R.A. 2006. Relation between Radopholus similis (Cobb) Thorne in production suckers and yield decline of mother plants in commercial banana plantations (Musa AAA) of Costa Rica with different yield structures. Nematropica 36(2):111-112. Kiewnick, S., Grimm, C., Cabrera, J.A. and Sikora, R.A. 2005. Range of activity and efficacy of abamectin-seed coating for control of root-knot nematodes on tomato. Phytopathology 95(6):S53.

102
Presentations in international conferences
Cabrera, J.A., Sikora, R.A. and Schouten, A. 2009. Involvement of microorganisms other than Pseudomonads on the enhanced degradation of the organophosphate non-fumigant nematicide fenamiphos. Abstracts of the 61st International Symposium on Crop Protection, May 19, 2009, Ghent Univesrity, Gent, Belgium. p. 21. Cabrera, J.A. and Sikora, R.A. 2008. Effect of natural plant enhancers on soil bacteria and control of plant-parasitic nematodes in lettuce. Proceedings of the International Conference on International Research on Food Security, Natural Resource Management and Rural Development, Tropentag: Competition for Resources in a Changing World, New Drive for Rural Development, Tropentag, October 7 - 9, 2008, University of Hohenheim, Stuttgart, Germany. p. 262. Cabrera, J.A., Sikora, R.A. and Schouten, A. 2008. Cross-degradation of novel non-fumigant nematicides by soil biotic factors. Proceedings of the 5th International Nematology Congress, July 13 - 18, 2008, Brisbane, Australia. p. 307. Cabrera, J.A., Pocasangre L.E. and Sikora, R.A. 2006. Impact of plant parasitic nematodes in following suckers on a root necrosis index in six commercial banana plantations of Costa Rica. Proceedings of the International Conference on International Research on Food Security, Natural Resource Management and Rural Development, Tropentag: Prosperity and Poverty in a Globalized World, Challenges for Agricultural Research, October 11 - 13, 2006, Rheinische Friedrich Wilhelms Universität, Bonn, Germany. p. 202. Cabrera, J.A., Pocasangre, L.E. and Sikora R.A. 2005. Importance and strategies of screening for enhanced biodegradation of pesticides in banana plantations. Proceedings of the International Conference on International Research on Food Security, Natural Resource Management and Rural Development, Tropentag: The Global Food and Product Chain, Dynamics, Innovations, Conflicts and Strategies, October 11 - 13, 2005, University of Hohenheim, Stuttgart, Germany. p. 303.





























