Embed Size (px)
Citation preview

Discoveries made over the past two decades have mark‑edly changed the way we study immunological processes. Not the least of these was the identification and clon‑ing of several new cytokines, including interleukin‑17A (IL‑17A)1 (BOX 1). These discoveries provided compelling reasons to explore outside the T helper 1 (TH1)– TH2 cell paradigm in search of answers to explain the effector T cell responses that occur independently of known TH1 and TH2 cell signalling pathways. It was clear that a strong T cell‑ dependent response can occur even in the absence of the TH1 cell promoting factors interferon‑γ (IFNγ), IL‑12p35, signal transducer and activator of transcription 1 (STAT1) and STAT4 or the TH2 cell promoting factors IL‑4, STAT6 and GATA binding protein 3 (GATA3)2.
Through the study of the role of IL‑23 in autoimmu‑nity, it was discovered that an alternative T cell subset can promote chronic inflammation and tissue damage3–6. A rapid succession of studies defined the TH17 cell para‑digm, in which IL‑6–STAT3 activation of the transcrip‑tional regulator retinoic acid receptor‑related orphan receptor‑γt (RORγt) controls the lineage fate of IL‑17A‑, IL‑17F‑, IL‑21‑ and IL‑22‑producing T cells (known as TH17 cells) that are highly responsive to IL‑1 receptor 1 (IL‑1R1) and IL‑23R signalling4–10. However, the TH17 cell pathway was inadequate to explain the early IL‑17‑mediated immune responses that have crucial roles during stress responses and host defence. The IL‑17‑mediated immune pathway is induced within hours following epithelial cell injury or activation of pattern recognition receptors (PRRs)8,11–13, which is not enough time for the development of TH17 cells. It is important to remember that the earliest reported functions of IL‑17A were its effects on innate immune responses 14,15.
Specifically, IL‑17A produced within 4–8 hours after microbial infection was shown to enhance neutrophil chemotaxis by promoting IL‑6, granulocyte colony‑stim‑ulating factor (G‑CSF) and CXC‑chemokine ligand 8 (CXCL8; also known as IL‑8) production and to trigger rapid, nonspecific immunity to infectious agents12.
So, what cell types are driving these early IL‑17‑dependent innate immune responses? It is notable that the organ systems that protect the host from the environ‑ment, specifically the lungs, intestinal mucosa and skin, seem to contain most of the IL‑17‑producing ‘sentinel’ cells16. Because, these sentinel cells would not have time to sense processed antigens, undergo developmental and expansion programmes and traffic to the site of injury, they probably reside in barrier tissues at the interface of host and environment, possess a pre‑activated pheno‑type and express an array of sensory receptors that allow them to sample their surrounding milieu. Innate IL‑17‑producing cells can also interact with memory cells during chronic inflammation17,18. The importance of TH17 cells in immune responses and diseases has been reviewed in detail elsewhere2,19,20, so we focus here on the innate sources of IL‑17 and how these cells are involved in sensing stress and injury.
Evidence for innate sources of IL‑17Following the discovery that IL‑23 promotes the secre‑tion of IL‑17 by memory CD4+ T cells21,22, we observed that IL‑23 also induced IL‑17 expression in recombination- activating gene (RAG)‑deficient mice, which lack both B and T cells, suggesting that innate IL‑17‑producing cells are an integral part of IL‑17‑mediated immune responses23. Using a T cell‑independent inflammatory
Merck Research Laboratories, DNAX Discovery Research, 901 California Avenue, Palo Alto, California 94304, USA.Correspondence to D.J.C. e-mail: [email protected]:10.1038/nri2800Published online 18 June 2010; corrected online 25 June 2010
Recombination-activating gene (RAG). Expressed by developing lymphocytes, mice that are deficient for either Rag1 or Rag2 fail to produce B or T cells owing to a developmental block in the gene rearrangement that is necessary for receptor expression.
Innate IL-17-producing cells: the sentinels of the immune systemDaniel J. Cua and Cristina M. Tato
Abstract | The cytokine interleukin‑17 (IL‑17) has received considerable attention since the discovery of a distinct CD4+ T helper (T
H) cell subset that produces it, known as the T
H17 cell subset.
Despite the fact that most of the recent literature describes IL‑17 as a T cell‑secreted cytokine, much of the IL‑17 released during an inflammatory response is produced by innate immune cells. In this Review, we explore the many innate immune cell populations that are an early source of IL‑17 in response to stress, injury or pathogens. These early sources have been shown to have a central role in the initiation of IL‑17‑dependent immune responses, even before the first CD4+T cell sees its cognate antigen and initiates the T
H17 cell developmental programme.
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 479
© 20 Macmillan Publishers Limited. All rights reserved10

γδ T cell A T cell that expresses a TCR consisting of a γ-chain and a δ-chain. These T cells are present in the intestinal epithelium as intraepithelial lymphocytes. γδ T cells are also present in the lungs, skin, liver and other tissue sites during inflammation. γδ T cells are thought to have an important role in innate immune responses, particularly in mucosal tissues.
Invariant natural killer T (iNKT) cell A T cell that expresses a particular variable gene segment, Vα14 (in mice) and Vα 24 (in humans), precisely rearranged to a particular Jα (joining) gene segment to yield T cell receptor α-chains with an invariant sequence. Typically, these cells co-express cell-surface markers that are encoded by the natural killer (NK) locus, and they are activated by recognition of CD1d, particularly when α-galactosylceramide is bound in the groove of CD1d.
disease model, in which RAG‑deficient mice are injected with a CD40‑specific monoclonal antibody (which mimics T cell–myeloid cell interaction), we found roles for IL‑12 and IL‑23 in the regulation of systemic and tissue‑specific inflammation, respectively23. Treatment of RAG‑deficient mice with CD40‑specific antibody induced a systemic cytokine storm response and multi‑organ inflammation and, unexpectedly, we found that IL‑23, but not IL‑12, was essential for local innate inflammation in the colon; however, IL‑23 had no role in systemic inflammatory changes, such as splenomegaly and elevated levels of tumour necrosis factor (TNF) and IL‑1 in the serum. Importantly, induction of IL‑17 expression was evident in the colon but not the spleen following CD40‑specific antibody treatment, correlating with the finding that IL‑23 promotes the activation of resident immune cells in the gut23.
The ability of IL‑23‑specific antibody to inhibit local innate IL‑17‑mediated responses is consistent with the observation that nearly all innate IL‑17‑producing cells express IL‑23R, and this signalling pathway can amplify
the inflammatory response by affecting activated mac‑rophages and dendritic cells23,24. In the ensuing years since their discovery, numerous reports have uncov‑ered the nature of these IL‑23‑dependent innate IL‑17‑producing cells. As predicated by the original studies, it has been shown that innate IL‑17‑producing cells pre‑dominantly reside in the skin and mucosal tissues that separate the host from the environment — where they serve as the sentinels of the immune system8,17,25–28.
Main sources of innate IL‑17what are the early producers of IL‑17 following tissue injury or exposure to pathogens? Recent studies have shown that γδ T cells are important innate IL‑17‑producing cells during autoimmune inflammation and infectious diseases17,29–34. The fact that γδ T cells express PRRs such as CLeC7A (also known as dectin 1) and Toll‑like recep‑tor 2 (TLR2) suggests that they are well suited for this role35. Additional innate IL‑17‑producing cells described in the literature include CD3+ invariant natural killer T (iNKT) cells, lymphoid‑tissue inducer (LTi)‑like cells, natural
Box 1 | The origins of the IL‑17 pathway
Since the discovery of interleukin‑17A (IL‑17A) in 1993 (REF. 1) (initially termed cytotoxic T lymphocyte antigen 8), several family members have been identified and are found to be highly conserved among vertebrate organisms129. Orthologous genes have not been identified for IL17A in primitive life, and none has been found in Drosophila melanogaster or Caenorhabditis elegans — species in which we typically find primordial genes that have crucial immune functions. Nevertheless, given that homology is only 16–50% between family members, perhaps it is not surprising that IL17A‑like genes across phyla may not be entirely recognizable. Although Agnathan orthologues have not been identified for interferon (IFN), IL‑2, IL‑4 or IL‑7, an Agnathan orthologue of IL‑17‑like factor has been described130. Interestingly, Agnathans seem to use IL‑17D, rather than IL‑17A, the function of which is yet to be identified in mammals. In addition, IL‑17D showed some degree of homology to IL‑17‑like factors in primitive phyla such as insects and worms.
Another key protein in the IL‑17 pathway is retinoic acid receptor‑related orphan receptor‑γt (RORγt), a transcriptional factor encoded by Rorc. Orthologous Rorc is only recognizable as recently as marsupials, although a Rorc‑like sequence is also present in bony fish. Given the importance of RORγt in regulation of IL‑17A and IL‑17F responses, it is possible that both the innate and adaptive IL‑17‑mediated immune response might not be as ‘ancient’ as previously thought; indeed, following careful analysis of phylogenic and sequence data, it is apparent that the signature genes of the IL‑17‑mediated immune response are not likely to be found in insects and worms.
It remains unknown if IL‑17A evolved before the development of the adaptive immune system. Indeed, it is possible that IL‑17A may have evolved as an innate cytokine following its development as an adaptive cytokine by virtue of the fact that it is most abundantly expressed by thymus‑dependent lymphocytes, including adaptive αβ T cells, as well as the innate γδ T cells, invariant natural killer T (iNKT) cells and lymphoid‑tissue inducer (LTi)‑like cells. Regardless of the evolutionary past of this cytokine, the importance of IL‑17A production by innate cell populations and the integral role these cells have in shaping the immune response are becoming clear.
Il‑17 family members
Cytokine Receptor Sources Function
IL‑17A IL‑17RA–IL‑17RC αβ T cells, γδ T cells, iNKT cells and LTi‑like cells
Pro‑inflammatory; exacerbates organ‑specific autoimmune inflammation; promotes mobilization of neutrophils and cytokine production by epithelial cell for protective immunity to extracellular pathogens; tight junction integrity; angiogenesis129
IL‑17B Undefined Intestine and pancreas129 Unknown
IL‑17C IL‑17RE Thymus and spleen80, 131 Unknown
IL‑17D Undefined Secreted by T cells, smooth muscle cells and epithelial cells129
Unknown
IL‑17E (also known as IL‑25)
IL‑17RA–IL‑17RB Mast cells and epithelial cells
Promotes T helper 2 cell differentiation protective immunity to parasitic worms and other extracellular pathogens132
IL‑17F IL‑17RA–IL‑17RC T helper 17 cells Enhances neutrophil recruitment at high concentrations129
R E V I E W S
480 | jULy 2010 | vOLUme 10 www.nature.com/reviews/immunol
© 20 Macmillan Publishers Limited. All rights reserved10

Common cytokine-receptor γ-chain A chain common to type I cytokine receptors. It was first discovered as the γ-chain of the IL-2 receptor and was subsequently shown also to be present in the receptors for IL-4, IL-7, IL-9, IL-15 and IL-21. It is mutated in humans with X-linked severe combined immunodeficiency.
killer (NK) cells and myeloid cells26,28,36,37. Not surpris‑ingly, each of these populations has an important role in tissue surveillance, most notably in the gut, lung and skin. These cell populations share some common activating signals, as well as unique pathways, and their function is vital to maintain the integrity of the target tissue (TABLE 1). How these distinct subsets of innate cell populations help shape T cell effector and memory responses and promote rapid antimicrobial immunity remains an area of active research.
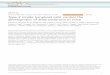
γδ T cells. The γδ T cell subset is an innate immune cell population that has an important role at the mucosal bar‑rier. However, the functional characteristics of γδ T cells have been difficult to elucidate. It is now becoming clear that the function of γδ T cells, in addition to their anti‑genic specificity, is influenced by a unique thymic selec‑tion process38. Because peripheral γδ T cells can recognize both self and non‑self ligands, it is thought that they can be divided into two broad categories of ‘antigen experienced’ and ‘antigen naive’ γδ T cells38. In turn, these subsets can also be sorted based on several cell surface markers and according to whether they readily produce IFNγ or IL‑17 (FIG. 1). Consistent with this hypothesis, fetal thymocytes can be segregated into IL‑17‑ or IFNγ‑producing γδ T cells based on their expression of CD27 (a TNF family mem‑ber). CD27– γδ T cells from the thymi of e18 mouse embryos were shown to constitutively express RORγt and runt‑related transcription factor 1 (RUNX1) and could be stimulated with phorbol 12‑myristate 13‑acetate (PmA) and ionomycin to produce IL‑17 (REF. 39). By contrast, CD27 co‑stimulation of γδ T cells seems to be required for the generation of IFNγ‑producing γδ T cells, as well as NKT cells. CD27+ γδ T cells were found to express Tbx21 (which encodes T‑bet) and thus, in turn, make IFNγ39. The CD27 ligand CD70 is expressed constitutively by thymic
epithelial cells, therefore CD27+ γδ T cells are thought to emerge from the thymus already ‘preactivated’ to make IFNγ38. CD25 (also known as IL‑2Rα) and CD122 (also known as IL‑2Rβ) are also expressed by IL‑17‑ and IFNγ‑producing γδ T cells, respectively40,41. CD25 expression may be downregulated on γδ T cells that experience their antigen in the thymus41, making them less sensitive to endogenous IL‑2 production. As IL‑2 is known to inhibit early TH17 cell development42, maintaining expression of CD25, the high‑affinity receptor for IL‑2, on this subset of thymic γδ T cells may also help to regulate the homeo‑stasis of peripheral IL‑17‑secreting γδ T cells41 (FIG. 1). IFNγ‑secreting γδ T cells in the periphery also express NK1.1, whereas IL‑17‑secreting γδ T cells express CC‑chemokine receptor 6 (CCR6), which is also known to be expressed by TH17 cells40. Finally, a recent study showed that the scavenger receptor SCART2 was exclusively expressed by peripheral γδ T cells that could be stimulated to pro‑duce IL‑17A43. As the phenotype of IL‑17A‑producing γδ T cells becomes more clearly defined, we should gain a better understanding of the role of these first‑responder cells to stress and injury.
LTi cells. Seminal work establishing the importance of Rorc (which encodes RORγt) in promoting IL‑17 pro‑duction by CD4+ T cells was paramount to our under‑standing of the biology of the TH17 cell developmental programme. As a result, it led to the investigation of other cells that were already known to express the tran‑scription factor RORγt, such as LTi cells44,45, which reside in peripheral lymph nodes, spleen and gut lam‑ina propria. In general, LTi cells depend on expression of the transcription factor inhibitor of DNA binding 2 (ID2)46, common cytokine-receptor γ-chain signalling and IL‑7R expression47–49 for their development and homeo‑stasis (TABLE 1). LTi‑like cells in the spleen28 share many
Table 1 | Innate IL‑17 producing cells
Cell type Inducing signal (ligand–receptor)
main effector cytokines
location or effector site
Functions Transcript‑ion factors
Refs
CD3+CD27– γδ T cell
IL‑23–IL‑23R; IL‑1– IL‑1R; RAE1 or MICA–NKG2D; β‑glucan–dectin 1; bacterial product–TLR
IL‑17 Gut and skin Surveillance; rapid host defence; maintenance of barrier function
RORγt, RUNX1, AHR and IRF4?
16,17, 35,89
CD1d+CD3+NK1.1–
iNKT cellIL‑23–IL‑23R; glycolipid–CD1d IL‑17 Liver, lung and
skinSurveillance; rapid host defence
RORγt 35,58,84
CD3–NKp46+ cell IL‑23–IL‑23R; RAE1 or MICA–NKG2D; IL‑15–IL‑15R
IL‑22 (IL‑17 in humans)
Gut and skin Production of pro‑ inflammatory mediators
RORγt, AHR, IRF4 and ID2
26,27,61, 62,64,65
CD3–
CD4+KIT+THY1+ LTi‑like cell
IL‑23–IL‑23; IL‑7–IL‑7R; bacterial product–TLR
IL‑22 and IL‑17 Lamina propria and spleen
Surveillance; lymphoid aggregate formation?
RORγt, ID2, AHR and STAT3
28,54, 79,133
THY1+SCA1+CD3–
CD4–KIT– cellIL‑23–IL‑23R; IL‑7–IL‑7R IL‑17, IL‑22
and IFNγLamina propria Immune surveillance RORγt and
T‑bet (AHR–)79
Paneth cell TNF–TNFR; bacterial product–NOD2
IL‑17 Intestinal crypts Surveillance; amplification of immune response
Not known 74
GR1+CD11b+ cell Bacterial product–TLR IL‑17 Lung and kidney Host defence Not known 13,75
AHR, aryl hydrocarbon receptor; ID2, inhibitor of DNA binding 2; IFN, interferon; IL, interleukin; iNKT, invariant natural killer T; IRF4, interferon‑regulatory factor 4; LTi, lymphoid‑tissue inducer; MICA, MHC class I polypeptide‑related sequence A; NKG2D, natural killer group 2, member D; NOD2, nucleotide‑binding oligomerization domain protein 2; R, receptor; RAE1, retinoic acid early transcript 1; RORγt, retinoic acid receptor‑related orphan receptor‑γt; RUNX1, runt‑related transcription factor 1; SCA1, stem cell antigen 1; STAT3, signal transducer and activator of transcription 3; TLR, Toll‑like receptor; TNF, tumour necrosis factor.
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 481
© 20 Macmillan Publishers Limited. All rights reserved10

Nature Reviews | Immunology
γδ T cell γδ T cell
TCR
IL-1R1
CD44
CD44 CD122
NK1.1
IL-12RIL-18R
CCR6
SCART2IL-23R
IL-17
Stressreceptor
No or weak TCR engagement TCR engagement
Thymus
TCR
IFNγ
RORγt T-bet
phenotypic similarities with LTi cells. Although LTi cells have been shown to be required for lymph node develop‑ment50–53, it is unknown whether LTi‑like cells can also induce lymph node organogenesis; therefore, they cannot yet be termed true LTi cells. when stimulated with either exogenous IL‑23 or the TLR2‑agonist zymosan, LTi‑like cells can produce IL‑17 (REF. 28); however, expression of specific TLRs by these cells has not been shown. In addi‑tion, both mouse and human LTi cell populations can produce IL‑22 following stimulation ex vivo with PmA and ionomycin or with TLR agonists54,55.
In mice, LTi and LTi‑like cells have been defined as leukocyte lineage marker (LIN)–CD4+CD127+ cells that express lymphotoxin‑α (LTα) and LTβ and the chemo‑kine receptors CCR7 and CXC‑chemokine receptor 5 (CXCR5)56. By contrast, human LTi cells are LIN–CD4–
CD127+CD45mid cells54. Following more rigorous exam‑ination, other disparities between human and mouse LTi cell surface markers have been noted: both CD7 and CD161 (also known as KLRB1) are expressed by human LTi cells, markers that are more characteristically expressed by human NK cells54. In fact, the development of CD56+ NK cells expressing Rorc transcripts could be induced from human LTi precursor cells27,54, suggesting a common lineage for these IL‑17‑ and IL‑22‑producing populations.
iNKT cells. iNKT cells are part of the innate immune system and are activated in response to self and non‑self glycolipid antigens presented by the non‑polymorphic mHC class I‑like molecule CD1d. iNKT cells rapidly produce immunoregulatory cytokines following acti‑vation by foreign or self glycolipids57. iNKT cells have a key role in immune surveillance, as well as immune suppression, and have been divided into subsets that produce either IL‑4 or IFNγ. Recently, a new IL‑17‑producing iNKT cell subset that develops in the thymus has been described58. Treatment of CD44hiNK1.1–CD4– iNKT thymocytes with the glycolipid α‑galactoceramide (α‑GalCer) stimulated IL‑17 production, indicating that a subset of iNKT thymocytes are already committed to making IL‑17 (REF. 58). Furthermore, NKT cells in the peripheral blood of simian immunodeficiency virus‑infected rhesus macaques were found to secrete IL‑17 and express CD161 and CCR6 (REF. 59), consistent with other IL‑17‑producing populations. IL‑17+ iNKT cells express RORγt and IL‑23R but not T‑bet (TABLE 1). By contrast, T‑bet+ iNKT cells express NK1.1 and produce IFNγ36. Of note, IL‑6, which is important for lineage commitment of TH17 cells, is not required for IL‑17‑producing iNKT cell development and func‑tion60. However, the precise signals that induce RORγt expression and lineage commitment of IL‑17‑producing iNKT cells are not known.
NK cells. NK cells have also drawn considerable interest; however, to date, only one report has shown evidence for the ability of NK cells to produce IL‑17 (REF. 37). Peritoneal NK1.1+ NK cells isolated from Toxoplasma gondii‑infected mice were shown to produce IL‑17A in an IL‑6‑ and IL‑23‑dependent manner37. However, it has been suggested that mucosal NK cell populations expressing NKp46 or NKp44 markers are a different lineage from the classical NK cell population found in the periphery27, and they may instead be related to LTi cells61,62. whether or not these cells can produce IL‑17 has yet to be determined; however, several studies have reported that CD3–NKp46+ cells can constitutively secrete substantial amounts of the TH17 cell‑associated cytokine IL‑22 (REFs 26,63–65). This NK cell subset is dependent on both RORγt and ID2 and are poor IFNγ producers61,66 (TABLE 1). Further investigation of NK cells that can rapidly produce IL‑17 and/or IL‑22 is warranted, given that they will probably be an impor‑tant population for mediating efficient early immunity under various conditions.
Paneth cells. Paneth cells are highly specialized epi‑thelial cells thought to be involved in mucosal homeo‑stasis and immunity. Known for their production of antimicrobial peptides, they have been intimately linked with innate immune responses in the gut67,68. In addition, Paneth cells have been shown to produce various pro‑inflammatory mediators, such as TNF and nitric oxide, and can respond to inflammatory stim‑uli through cytokine receptors or intracellular PRRs, such as nucleotide‑binding oligomerization domain 2 (NOD2)69–73. Thus, Paneth cells may actively promote
Figure 1 | Developmental programming of γδ T cell subsets. Certain subsets of γδ T cells may leave the thymus programmed to secrete either interleukin‑17 (IL‑17) or interferon‑γ (IFNγ) once they enter the periphery. It is hypothesized that the predilection towards a particular functional programme by a γδ T cell is determined in part by the strength of signal as it develops within the thymus. T cell receptor (TCR) engagement and/or coincident co‑stimulation in the thymus will bias the cell towards an IFNγ‑secreting programme. By contrast, lack of TCR engagement or weak TCR signalling will promote an IL‑17‑secreting programme. Whether or not these subsets are truly mutually exclusive has yet to be shown. However, several surface receptors have been identified by various laboratories as being associated with either IL‑17‑ or IFNγ‑producing γδ T cells. CCR6, CC‑chemokine receptor 6; R, receptor; RORγt, retinoic acid receptor‑related orphan receptor‑γt.
R E V I E W S
482 | jULy 2010 | vOLUme 10 www.nature.com/reviews/immunol
© 20 Macmillan Publishers Limited. All rights reserved10

Ischaemia–reperfusion injury An injury in which the tissue first suffers from hypoxia as a result of severely decreased, or completely arrested, blood flow. Restoration of normal blood flow then triggers inflammation, which exacerbates the tissue damage.
chronic inflammation after exposure to inflammatory cytokines such as TNF through a positive feedback loop that induces increased pro‑inflammatory cytokine production in the mucosa.
Paneth cells activated in a model of TNF‑induced shock have been shown to rapidly secrete IL‑17 (REF. 74) and are therefore another innate source of this cytokine. Furthermore, IL‑17 expression detected at the bottom of crypts in the jejunum of naive mice was found to co‑localize with Paneth cells74. These specialized epithelial cells constitutively express intracellular IL‑17 and can efficiently release cytokine stores following stimula‑tion. IL‑17 could then feed forward to promote more TNF production followed by increased IL‑6 and nitric oxide. In addition, IL‑17‑encoding mRNA is rapidly upregulated to replace the spent store of IL‑17 (REF. 74). Paneth cell‑derived IL‑17 was shown to be responsible for increased chemokine production and recruitment of neutrophils74. Taken together, these data provide an additional mechanism for acute production of IL‑17. Given the role of these specialized epithelial cells in immune surveillance, it is conceivable that these stores of IL‑17 could provide a rapid and efficient mechanism to respond to mucosal pathogens.
Neutrophils. A recent report75 suggests that myeloid cells can produce IL‑17 in response to ‘sterile’ injury such as ischaemia, in which there is recruitment of neutrophils into the damaged tissue. In a model of acute kidney ischaemia–reperfusion injury, early production of IL‑23 acti‑vates downstream IL‑17‑mediated pathways, including chemo kine production and recruitment of neutrophils. most of the initial IL‑17 in the kidney was produced by bone marrow‑derived CD11b+GR1+ cells and not by lympho cytes75. This IL‑17 production directly promoted kidney damage as measured by serum creatinine levels after injury induction. Furthermore, transfer of CD11b+GR1+ cells to Il17a–/– mice promoted an injury response, which could be subsequently blocked by IL‑17‑specific anti‑body75. Interestingly, IFNγ production was also decreased in the absence of IL‑17, suggesting that IL‑17 promoted pathogenic levels of IFNγ that exacerbated tissue injury.
An earlier study also shows that Il17 mRNA can be detected in purified neutrophils after administration of anti‑neutrophil cytoplasmic autoantibodies (ANCA) to induce arthritis76. GR1hiCD11bmidF4/80– neutrophils iso‑lated after ANCA injection could be stimulated ex vivo with PmA and ionomycin in the presence or absence of myeloperoxidase‑specific ANCA to produce IL‑17 (REF. 76), suggesting these cells may be contributors to the pathogenesis of the disease. However, there remain many open questions; for example, it is unknown whether neutrophils express IL‑23R, through which signalling is required for most IL‑17‑producing cells, or whether these cells exhibit IL‑17‑inducing transcription factors, such as RORγt or RORα. whether these cells respond to stimuli distinct from those already known to induce IL‑17 production also remains to be determined.
Although we have described many subsets of innate IL‑17‑producing cells, it is likely that more will be uncov‑ered. The observation that these innate cells reside in
diverse anatomical locations — mainly in the lungs, liver, skin, gut and secondary lymphoid organs — suggests that they have a broad range of immune regulatory func‑tions that has yet to be fully elucidated. The gut mucosal tissue seems to harbour the highest numbers and types of innate IL‑17‑producers77,78. Indeed, a new subset of innate lymphoid cells has been recently described that is characterized as THy1hiSCA1+LIN–KIT–CD3–CD4– (REF. 79) (TABLE 1). These cells are CCR6+ and reside in the intestinal lamina propria and can readily expand more than 100‑fold during microbial infection. Following stimulation with IL‑23, this innate population pro‑duces high levels of IL‑17, IL‑22 and IFNγ and seems to be uniquely regulated by RORγt and T‑bet but not by the TH17 cell‑associated transcription factor aryl hydrocarbon receptor (AHR) (see below)79.
Although many innate lymphoid subsets are found in the tissues with barrier functions, during a tissue‑specific inflammatory response these cells can also migrate from secondary lymphoid organs to immune‑privileged sites, such as the central nervous system (CNS), where they participate in the disease process17. we now have a fundamental understanding of how the IL‑17R signalling pathway, through the adaptor molecule ACT1 (also known as CIKS) and the tran‑scription factor nuclear factor‑ κB (NF‑κB), induces downstream mediators, including IL‑6, CXCL8 and CC‑chemokine ligand 20 (CCL20)80. we next discuss the common activating signals shared among the innate IL‑17 producing cells, as well as the unique pathways that regulate them.
Regulation of innate IL‑17‑producing cellsTranscriptional regulation by RORγt and AHR. The orphan nuclear receptor RORγt has been shown to induce the transcription of genes encoding IL‑17. Although IL‑6‑dependent STAT3 activation is thought to be crucial for RORγt expression and development of TH17 cells, careful examination of IL‑6‑deficient mice revealed several sub‑sets of IL‑17‑producing cells that arise independently of this cytokine60,81,82, including iNKT cells, γδ T cells, LTi‑like cells and NK‑like cells (FIG. 2). The fact that these cells do not require IL‑6 stimulation raises an interesting ques‑tion: what promotes preferential RORγt expression in these innate populations? In αβ TCR thymocytes, RORγt is first induced as a crucial survival factor83 for CD4+CD8+ (double positive) thymocytes; as these cells mature and go through the TCR selection process, RORγt is repressed. Subsequent activation with TGFβ and IL‑6 is required to induce the re‑expression of RORγt for TH17 cell dif‑ferentiation10. By contrast, the subsets of γδ T cells and iNKT cells that exit the thymus without going through TCR selection do not repress RORγt expression and seem to preferentially develop into IL‑17‑producing cells58,84. Interestingly, the subsets of γδ T cells and iNKT cells that do go through TCR selection preferentially express T‑bet and produce IFNγ38. However, more work is needed to confirm this TCR selection hypothesis for the ‘default’ development of innate IL‑17‑producing cells. Another hypothesis that may be more applicable for CD3– LTi and NKp46+ cells is that other STAT3 activators, such as
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 483
© 20 Macmillan Publishers Limited. All rights reserved10

Nature Reviews | Immunology
APC
a TH17 cell development
Naive CD4+ T cell
3–5 days
TGFβ and IL-6
Differentiationin lymph node
Polarized effector TH17 cell
IL-21IL-1IL-23
IL-17IL-22
↑ STAT3 RORγt
STAT3RORγtAHR
Antigen-presentation and co-stimulation
APC
b Innate IL-17-producing cell development
γδ T or iNKT cell
4–8 hours
IL-1βIL-23
PAMPsStress proteinsPathogen metabolitesGlycolipids
Activation intarget tissue
↑ IL-17IL-17IL-22(?)
RORγtSTAT3AHR
RORγtSTAT3AHR
IL‑21 and IL‑23, could initiate the RORγt‑mediated pro‑gramme (FIG. 2). Further studies are needed to formally test whether IL‑21‑ and/or IL‑23‑induced STAT3 signal‑ling is necessary and sufficient for LTi and NKp46+ cells to turn on RORγt expression.
AHR is another transcriptional regulator that con‑trols IL‑17‑associated immune responses85. It is a nuclear receptor that senses environmental toxins, such as dioxin and FICZ (6‑formylindolo[3,2‑b]carbazole)86,87. Recently, AHR was shown to be expressed by TH17 and regulatory T cells, suggesting a link between toxic environmental pollutants and tissue inflammation18,88. Importantly, AHR is also expressed by γδ T cells, LTi cells and NK cells and is important for their effector functions, including IL‑22 production26,89 (TABLE 1). whether other IL‑17‑producing myeloid cells also express AHR is currently unknown. It has been suggested that AHR can cooperate with RORγt to induce maximal amounts of IL‑17 and IL‑22 produc‑tion and to inhibit TGFβ‑induced FOXP3 expression90. In addition, AHR was shown to co‑immuno precipitate with STAT1 and STAT5. This interaction was thought to be relevant for suppressing STAT1‑ and STAT5‑mediated signalling and thus enhancing the effects of IL‑23‑ and RORγt‑dependent functions91. The expres‑sion of AHR by innate sentinel cells seems fortuitous
owing to their proximity to the environment. However it is conceivable that this sensor capable of detecting toxic pollutants may predispose individuals to chronic inflammatory disorders.
Cytokines that regulate innate IL‑17‑producing cells. A common characteristic of many of these innate IL‑17‑producing cells is the constitutive expression of IL‑1R1 and IL‑23R36,82,89,92–94, both of which are also important to potentiate the TH17 cell differentiation programme (FIG. 2). Studies have now shown that in vitro stimula‑tion of γδ T cells with IL‑1β or IL‑23 can promote rapid IL‑17 secretion17,89; optimal IL‑17 production is induced following culture with both IL‑1β and IL‑23 (REF. 17). Furthermore, 4 hours after injection of IL‑1β and IL‑23 into the footpad of mice, γδ T cells in the draining lymph node were already stimulated to secrete IL‑17 (REF. 17). Ex vivo stimulation of LTi cells with IL‑23 also induced the secretion of IL‑17 (REFs 28,54). Similar results were obtained in studies culturing iNKT cells, from the drain‑ing lymph nodes of type II collagen‑immunized mice, with IL‑23 (REF. 93). Furthermore, human NKT cells treated with IL‑23 in vitro produced IL‑17 (REF. 82). Although not strictly dependent on CD1d ligation, co‑activation of iNKT cells with IL‑23 and α‑GalCer was
Figure 2 | Activation of innate Il‑17‑producing cells. a | Development of adaptive immunity and a polarized effector T helper 17 (T
H17) cell population takes up to 5 days in vivo. This process requires antigen presentation by antigen‑presenting
cells (APCs), using classical MHC molecules, in the presence of transforming growth factor‑β (TGFβ), interleukin‑6 (IL‑6), IL‑21, IL‑23 and IL‑1β. This initial activation results in the upregulation of signal transducer and activator of transcription 3 (STAT3) and retinoic acid receptor‑related orphan receptor‑γt (RORγt) expression to enhance IL‑23 responsiveness and induce IL‑17 production. After rounds of proliferation and differentiation in the lymph node, polarized T
H17 cells, with
stable expression of RORγt and aryl hydrocarbon receptor (AHR), are ready to migrate into target tissues. b | By contrast, innate cells that reside in peripheral tissues can be activated in the presence of cytokines, such as IL‑23 and IL‑1β, alone or in combination with direct pathogen‑associated molecular pattern (PAMP) recognition by pattern recognition receptors (PRRs) or through their T cell receptor (TCR), when expressed. Because these cells constitutively express transcriptional regulators for IL‑17 production, they can produce IL‑17 protein within hours of stimulation. iNKT, invariant natural killer T.
R E V I E W S
484 | jULy 2010 | vOLUme 10 www.nature.com/reviews/immunol
© 20 Macmillan Publishers Limited. All rights reserved10

Nature Reviews | Immunology
Panethcell
Pathogensand commensals
Epithelial cell
DC
IL-17AIL-17FIL-22
Stress proteins such as MICA and RAE1
Innate IL-17-producing cell• γδ T cell• iNKT cell• LTi-like cell• NKp46+ cell• THY1hi SCA1+ cell
IL-1βIL-23
TNF
IL-17
Defensins, REG proteins,S100A, lipocalin, lactoferrin
CCL20, G-CSFand IL-6
Tight junctionproteins
shown to have synergistic effects on IL‑17 production82,93. Conventional NK cells activated in vivo after challenge with the intracellular parasite T. gondii, can produce a substantial amount of IL‑17 in response to exogenous IL‑6 and IL‑23 (REF. 37). These studies show that the con‑stitutive expression of IL‑1R1 and IL‑23R provides an efficient mechanism to induce effector cytokine produc‑tion in innate IL‑17‑producing cells. Additional inflam‑matory cytokines, not typically associated with a TH17 cell response, could also promote IL‑17 production in subsets of innate cell populations. For example, the IL‑1 family member IL‑18 is known to induce iNKT cell acti‑vation, and thus may in turn promote IL‑17 secretion by these cells59. Similarly, Paneth cells have been shown to be responsive to TNF leading to IL‑17 protein synthesis74.
Physiological roles of innate IL‑17Early IL‑17 and infection. A hallmark of innate immu‑nity is its rapid response to pathogens. Innate IL‑17‑producing cells can induce epithelial cell secretion of granulopoietic factors such as G‑CSF and CCL20, which recruit large numbers of neutrophils crucial for effective and rapid control of bacterial and fun‑gal pathogens (FIG. 3). IL‑17 also synergizes with other cytokines, such as IL‑1, IL‑6 and TNF, that promote
activation of tissue infiltrating neutrophils to effec‑tively eliminate extracellular pathogens. A key feature of neutrophil recruitment is the requirement for an early response — within 4–8 hours after exposure — for effec‑tive immunity against bacterial and fungal pathogens such as Citrobacter rodentium, Klebsiella pneumoniae, Staphylococcus aureus and Candida albicans11,12,95–97. which innate populations provide this early IL‑17 dur‑ing infectious challenge? The answer probably depends on the target tissue of the pathogen, such as the lungs and skin and whether it induces a systemic infection involving the liver, spleen and/or peritoneum. It is also likely that several innate cell subsets could cooperate to promote this IL‑17‑mediated response.
In the lung, the γδ T cell subset has been implicated as a primary source of early IL‑17 production in sev‑eral in vivo models of infection. Initial studies looked at extracellular pathogens that target the lungs, such as K. pneumoniae and Mycobacterium tuberculosis, and showed that IL‑23 was a main stimulator of innate IL‑17 production33,34,98; γδ T cells were later identified as being an important source of this cytokine in the lungs of Mycobacterium bovis‑infected mice99, as well as in patients with tuberculosis100. Consequences of γδ T cell deficiency during M. tuberculosis infection include dys‑regulated granuloma formation in the lung characterized by impaired recruitment of other lymphocytes101, suggest‑ing an important early role for γδ T cells in promoting efficient antibacterial immunity in the lung. Furthermore, there is increasing evidence that early IL‑17 production may also influence the subsequent effector CD4+ T cell response, particularly TH1 cells102, by inducing chemo‑kines that attract TH1 cells and induce optimal production of IFNγ in the lung99. In this way, innate IL‑17‑producing cells promote a more potent adaptive immune response for a more efficient resolution of infection.
In the skin, γδ T and iNKT cells are two distinct populations of innate IL‑17 producers. Deficiency of γδ T cells can result in decreased control of S. aureus infec‑tion103. A more recent study showed that in the absence of γδ T cells, there is a specific defect in IL‑17 produc‑tion 8 hours after intra‑dermal challenge with S. aureus, whereas αβ T cell deficiency did not affect IL‑17 pro‑duction at this early timepoint104. Furthermore, vγ5+ γδ T cells isolated from the epidermis of naive mice could be stimulated with IL‑1β and IL‑23 to readily produce large amounts of IL‑17 in vitro104, suggesting that this skin‑homing population of innate cells are poised to respond rapidly. Although there are as yet no examples showing iNKT cells can participate in antimicrobial responses in the skin, resident iNKT cells can expand rapidly and pro‑duce IL‑17 following mitogen‑induced injury caused by 12‑O‑tetradecanoylphorbol‑13‑acetate (TPA) skin paint‑ing84, suggesting that these cells could contribute to early IL‑17 production in response to tissue injury.
Infection of peripheral, non‑mucosal tissues can also be controlled by innate IL‑17‑producing cells. A substantial population of peritoneal IL‑17‑producing γδ T cells was identified after intraperitoneal challenge with Escherichia coli or Salmonella enterica subsp. enterica serovar enteritidis105,106. After E. coli challenge, IL‑17
Figure 3 | Innate Il‑17‑producing cells have key roles in orchestrating mucosal barrier functions during health and disease. Innate interleukin‑17 (IL‑17)‑producing cells are important sentinels of the immune system. Cells such as γδ T cells, invariant natural killer T (iNKT) cells, LTi‑like cells and NK cells reside in the intestinal lamina propria and intraepithelial compartment close to the host–environment interface. Paneth cells reside in intestinal crypts in close contact with bacterial microbiota. When activated, these different groups of cells produce factors, including IL‑17, that are known to maintain intestinal integrity. For example, IL‑17 can enhance claudin and zona occludens 1 synthesis. These tight junction proteins form interconnecting ultra‑structures between epithelial cells to keep out gut luminal contents and infectious organisms. IL‑17 also acts directly on epithelial cells to promote release of defensins, regenerating (REG) proteins and S100 proteins, which have important antimicrobial activities. Finally, IL‑17 also promotes epithelial cell secretion of chemokines such as CC‑chemokine ligand 20 (CCL20) for recruitment of neutrophils when the mucosal barrier is breached. DC, dendritic cell; G‑CSF, granulocyte colony‑stimulating factor; MICA, MHC class I polypeptide‑related sequence A; RAE1, retinoic acid early transcript 1; SCA1, stem cell antigen 1; TNF, tumour necrosis factor.
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 485
© 20 Macmillan Publishers Limited. All rights reserved10

Tight junction A belt-like region of adhesion between adjacent epithelial or endothelial cells that regulates paracellular flux. Tight junction proteins include the integral membrane proteins occludin and claudin, in association with cytoplasmic zonula occludens proteins.
production can be detected a few hours after exposure and can be maintained for days, suggesting that these cells may be required for enhancing the antimicrobial response105. IL‑17‑producing γδ T cells are also found in the liver during the first three days of infection with intracellular pathogens such as Listeria mono-cytogenes and T. gondii 107,108. This early IL‑17 produc‑tion is required for optimal neutrophil recruitment to the liver and resistance to the infection. It is important to note that innate IL‑17 populations not only interact with pathogens during infection, but also interact with the commensal flora to maintain mucosal homeo stasis. Recent studies have suggested that this interaction with the intestinal microbiota is an important defining fea‑ture of IL‑17‑producing cells — the sentinels of the mucosal barrier.
Early IL‑17 and surveillance. Although there is compel‑ling evidence in the literature of the destructive nature of dysregulated IL‑17‑mediated inflammation, it has long been known that IL‑17 also has an important role in main‑taining mucosal barrier integrity both during homeostasis and pathogenic insults109,110. One way IL‑17 can preserve tissue integrity is by enhancing synthesis of tight junction proteins, such as claudin111. These tight junction proteins form interconnecting ultra‑structures between epithelial cells to keep out gut luminal contents and commensal organisms (FIG. 3). The importance of IL‑17 in sustaining the integrity of the intestinal cell wall was shown in stud‑ies where neutralization of IL‑17 exacerbated epithelial destruction in a colonic injury model112.
Another mechanism for the protective effects of IL‑17 is the induction of antimicrobial agents such as β‑defensins, regenerating (ReG) proteins, S100 proteins, lipocalins and lactoferrins (FIG. 3). These microbicidal agents are predominantly produced by epithelial cells, as well as neutrophils and macrophages that constitu‑tively express IL‑17R. It is notable that IL‑17 often works cooperatively with IL‑22. These two cytokines can be co‑produced by γδ T cells, LTi cells and THy1hiSCA1+ cells, although they may preferentially produce only IL‑17 or IL‑22 at a given time point17,26–28,79. exactly what regulates differential IL‑17 and IL‑22 production is not known. In the airways, IL‑17 and IL‑22 have been shown to work together to promote the secretion of β‑defensin 2, β‑defensin 3 and calgranulin by bronchial epithelial cells25,113. Induction of lipocalin‑2 expression by IL‑17 and IL‑22 in tracheal epithelial cells is essential for killing of the Gram‑negative pathogen K. pneumoniae25,114.
In the gut mucosa, IL‑17 and IL‑22 have synergistic effects on the induction of antimicrobial proteins such as ReG3γ by gastrointestinal epithelial cells, which has an important role in the control of pathogens and limits dis‑semination of commensal bacteria that could penetrate a disrupted epithelial barrier8,115. ReG3γ is a soluble C‑type lectin produced by Paneth cells and interacts directly with the bacterial cell wall to promote its anti‑microbial activity116. mice lacking IL‑22 and IL‑23 have reduced amounts of S100A8, S100A9, ReG3β and ReG3γ and were highly susceptible to C. rodentium infection but can be rescued by treatment with recombinant ReG3γ8. The
production of ReG3γ is maintained at low levels by com‑mensal flora and is strongly induced by pathogens such as C. rodentium and L. monocytogenes8,116,117.
One intriguing implication of these studies is that the composition of gut commensal species can skew the mucosal immune effector cell balance. For exam‑ple, a recent study has shown that a single group of commensal bacteria, segmented filamentous bacteria (SFB), preferentially induced intestinal IL‑17‑producing cells77. mice that are free of SFB have few gut‑resident IL‑17‑producing cells and were shown to be more sus‑ceptible to Citrobacter spp. infection. Transfer and coloni‑zation of SFB in these mice increased the number of gut‑ resident IL‑17‑producing cells and enhanced resistance to the inflection77. These findings suggest that increases in mucosal IL‑17 and IL‑22 production — which induce a subset of antimicrobial peptides that target specific groups of bacteria — may ‘re‑shape’ the gut microbiota. In turn, gut commensal organisms can specifically promote IL‑17‑secreting cells by inducing IL‑1R1 expression on their sur‑face92. During SFB colonization, it is likely that the sentinel innate IL‑17 producing cells, such as LTi cells, NKp46+ cells and γδ T cells, are the first responder populations and influence the development of TH17 cells. These innate and memory cells would act in concert to produce a basal level of IL‑17 and IL‑22, which then maintain a consti‑tutive level of antimicrobial proteins. In addition, there are many checks and balances in this system to maintain mucosal homeostasis, and FOXP3+ regulatory T cells have an important role in opposing the effects of the IL‑17‑producing cells118. Given the pro‑inflammatory activities of IL‑17 and IL‑22 in this model, it is conceivable that a dysregulated interaction between sentinel IL‑17‑producers and pathogenic organisms could lead to skewing of the effector versus regulatory cell balance and predispose an individual to chronic inflammatory diseases.
Early IL‑17 and autoimmunity. Recent studies have begun to elucidate the role of innate IL‑17‑producing cells in priming and perpetuating autoimmunity. Although innate cells do not recognize specific peptide antigens presented by accessory cells and cannot, therefore, initi‑ate an immune response against self antigen, they may still have an important role in shaping the cytokine milieu in a target organ to favour the induction of chronic inflam‑mation. The immune ‘feedforward’ loop between mucosal pathogens and an inflammatory ‘TH17‑type’ response is a good example. Among the many subsets of innate IL‑17 producing cells, γδ T cells have received the most atten‑tion regarding their participation in both the induction, as well as the effector, phase of autoimmune inflamma‑tion17,29,30,31. For example, in a model of collagen‑induced arthritis, γδ T cells were found to be an important source of early IL‑17 produced in response to IL‑1β and IL‑23 signalling30. It is thought that γδ T cells are activated directly by complete Freund’s adjuvant (CFA), which is used in the protocol to help induce collagen‑induced arthritis29. Cooperation between IL‑17‑producing γδ T cells and antigen‑specific CD4+ αβ T cells was shown to enhance the pathogenesis of experimental auto‑immune uveitis after immunization with the autoantigen
R E V I E W S
486 | jULy 2010 | vOLUme 10 www.nature.com/reviews/immunol
© 20 Macmillan Publishers Limited. All rights reserved10

Blood–brain barrierA physiological barrier between blood vessels and brain parenchyma. It is formed by specialized tight junctions between endothelial cells of the blood vessel wall, which is surrounded by a basement membrane and an additional membrane formed from astrocyte feet and microglial cells, known as the glial limitin.
IL-23ReGFP and RORγteGFP reporter miceMice expressing enhanced green fluorescent proteins (eGFP) under the control of the IL-23R (encoded by Il23a) or RORγt (encoded by Rorc) promoter. These mice can be used to track the differentiation, proliferation, migration and effector function of IL-17-producing cells. Gene targeting vectors are generated by introducing eGFP sequences into bacterial artificial chromosome (BAC) clones containing the Il23a or Rorc gene. The targeting constructs are introduced into embryonic stem cells and injected into mouse blastocysts to generate eGFP transgenic mice. The eGFP is co-expressed with IL-23R or RORγt, which allows cell fate analysis of IL-17-producing cells during an immune response.
interphotoreceptor retinoid‑binding protein31. In a CNS autoimmune model, when γδ T cells were depleted dur‑ing immune priming, fewer myelin‑oligodendrocyte glycoprotein (mOG)‑derived antigen specific TH17 cells develop in vivo, suggesting that γδ T cells may promote the expansion of antigen‑specific TH17 cells17. In addition, treatment with IL‑1β and IL‑23 could induce γδ T cell secretion of IL‑17 without the need for TCR engagement. Remarkably, in vivo injection of IL‑1 and IL‑23 could induce γδ T cell expression of Il17 mRNA in just 4 hours17. Together, these results indicate that early production of cytokines by innate IL‑17‑producers may enhance devel‑opment of pathogenic TH17 cells, although the precise mechanism has not been elucidated. One suggestion was that innate cell‑produced IL‑17 could act as a paracrine factor to promote activation of TH17 cells17. whether IL‑17 can signal and activate CD4+ T cells as an autocrine and/or paracrine factor remains controversial. A recent study, using surface plasmon resonance analysis, showed that once an IL‑17 molecule is bound by an IL‑17RA mol‑ecule, the binding affinity for a second IL‑17RA is reduced by three logs, whereas the affinity of IL‑17RC for IL‑17 remains high, thereby promoting heterodimeric rather than homodimeric receptor formation119. Fibroblasts, epithelial cells and endothelial cells express both IL‑17RA and IL‑17RC, whereas T cells express IL‑17RA but not IL‑17RC120. These findings raise the possibility that T cells, which bear the homodimeric IL‑17RA receptor complex, may have a lower affinity for IL‑17 than macrophages and epithelial cells that express high‑affinity heterodimeric IL‑17RA–IL‑17RC receptor complexes119,120.
An alternative suggestion for the mechanism by which innate cell‑produced IL‑17 can enhance autoimmune inflammation, is that early IL‑17 can act on antigen‑pre‑senting cells (APCs), such as macrophages and subsets of dendritic cells17, which could express both IL‑17RA and IL‑17RC120. IL‑17 has been shown to directly induce APC production of IL‑23, IL‑1, IL‑6 and TGFβ17, which are the crucial factors for development of pathogenic TH17 cells. In this way, innate IL‑17 produced early during immune priming could influence the generation of antigen‑specific TH17 cells and exacerbate autoimmunity. In humans, γδ T cells have been found to accumulate in acute multiple sclerosis lesions121, as well as in cerebrospinal fluid of patients with recent‑onset multiple sclerosis122, suggesting that innate cells may also have a role in human disease.
Conclusions and future directionsIt is evident that both adaptive T cells and innate cell populations can produce IL‑17, and this has crucial roles in mucosal tissues. Signalling through IL‑17RA–IL‑17RC
receptor complexes (reviewed in REF. 80) — which are highly expressed by epithelial cells — recruits neu‑trophils, promotes secretion of defensin‑like factors and maintains tight junction structure of the intestinal wall; together, these factors maintain mucosal tissue barrier functions. Despite these important insights, we still do not fully understand the effects of IL‑17 in immune privileged sites such as the CNS, where IL‑17 is produced by both adaptive TH17 cells and innate γδ T cells17. A recent report suggested that the IL‑17R complex is expressed by astrocytes that form part of the blood–brain barrier and that these cells are an important initial IL‑17‑responder population that control inflam‑matory responses in the CNS123. whether additional subsets of glial cells also express the IL‑17R complex and how early IL‑17 signalling affects brain inflammation remain important areas of research.
we still do not fully appreciate the roles of IL‑23R+RORγt+ innate cells in the mucosal tissues. One issue is that many of these cells lack lineage mark‑ers, making it difficult to study their developmental pathway and function. It is likely that they are impor‑tant tissue‑resident first responders during microbial or toxin insults. with the availability of IL-23ReGFP and RORγteGFP reporter mice, it may be possible to dissect the in vivo functions of these lineage marker‑negative cells. The fact that LTi‑like and NK‑like cells are also found in the spleen and other lymphoid tissue is interesting. Perhaps these cells produce early IL‑17 and induce APCs to secrete IL‑1, IL‑6 and IL‑23, thereby enhancing TH17 cell priming17,120. Are these LTi‑like and NK‑like cells part of the same population of mucosal residents that then migrate throughout the host organism? If so, what factor (or factors) controls their decision to reside in the peripheral tissue or to migrate to lymphoid organs? These are important areas of research that will shed light on the complex regulation and function of these cells.
Finally, the clinical relevance of innate IL‑17‑producing cells is now better understood. IL‑23R poly‑morphisms are associated with psoriasis, ankylosing spondylitis and inflammatory bowel disease124,125,126 — a cluster of diseases with a well recognized clinical asso‑ciation and overlap. It has become evident that animal models of these inflammatory diseases can be induced in the complete absence of adaptive immune cells. This is likely to have important consequences for therapeutic treatments as current clinical studies have shown efficacy for targeting IL‑17 and IL‑23 in human autoimmune dis‑orders127,128. Understanding these early innate immune responses will be the key to designing rational therapies targeting these important immune pathways.
1. Rouvier, E., Luciani, M. F., Mattei, M. G., Denizot, F. & Golstein, P. CTLA-8, cloned from an activated T cell, bearing AU-rich messenger RNA instability sequences, and homologous to a herpesvirus saimiri gene. J. Immunol. 150, 5445–5456 (1993).
2. Weaver, C. T., Hatton, R. D., Mangan, P. R. & Harrington, L. E. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu. Rev. Immunol. 25, 821–852 (2007).
3. Langrish, C. L. et al. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 201, 233–240 (2005).
4. Harrington, L. E. et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nature Immunol. 6, 1123–1132 (2005).
5. Park, H. et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nature Immunol. 6, 1133–1141 (2005).
6. Veldhoen, M., Hocking, R. J., Atkins, C. J., Locksley, R. M. & Stockinger, B. TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
7. Wei, L., Laurence, A., Elias, K. M. & O’Shea, J. J. IL-21 is produced by Th17 cells and drives IL-17 production in a STAT3-dependent manner. J. Biol. Chem. 282, 34605–34610 (2007).
8. Zheng, Y. et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nature Med. 14, 282–289 (2008).This is the first study to suggest that RORγt+ lymphocytes can produce IL‑22, which promotes epithelial cell production of REG3γ required for host defence against extracellular gut pathogens.
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 487
© 20 Macmillan Publishers Limited. All rights reserved10

9. Korn, T. et al. IL-21 initiates an alternative pathway to induce proinflammatory TH17 cells. Nature 448, 484–487 (2007).
10. Ivanov, I. I. et al. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 126, 1121–1133 (2006).The data in this report show RORγt is the key transcription factor promoting IL‑17A, IL‑17F, and IL‑22 expression by TH17 cells. Subsequent studies showed that RORγt also regulates innate lymphocytes.
11. Happel, K. I. et al. Cutting edge: roles of Toll-like receptor 4 and IL-23 in IL-17 expression in response to Klebsiella pneumoniae infection. J. Immunol. 170, 4432–4436 (2003).
12. Happel, K. I. et al. Divergent roles of IL-23 and IL-12 in host defense against Klebsiella pneumoniae. J. Exp. Med. 202, 761–769 (2005).
13. Ferretti, S., Bonneau, O., Dubois, G. R., Jones, C. E. & Trifilieff, A. IL-17, produced by lymphocytes and neutrophils, is necessary for lipopolysaccharide-induced airway neutrophilia: IL-15 as a possible trigger. J. Immunol. 170, 2106–2112 (2003).
14. Fossiez, F. et al. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J. Exp. Med. 183, 2593–2603 (1996).
15. Kennedy, J. et al. Mouse IL-17: a cytokine preferentially expressed by αβ TCR + CD4–CD8– T cells. J. Interferon Cytokine Res. 16, 611–617 (1996).
16. Lochner, M. et al. In vivo equilibrium of proinflammatory IL-17+ and regulatory IL-10+ Foxp3+ RORγt+ T cells. J. Exp. Med. 205, 1381–1393 (2008).This study used RORγteGFP reporter mice to determine the proportion of ‘type 17’ [Au: TH17 type?] αβ and γδ T cell subsets in skin, lungs and gut. It shows the expansion of IL‑17+RORγt+ cells in situ during inflammatory responses.
17. Sutton, C. E. et al. Interleukin-1 and IL-23 induce innate IL-17 production from γδ T cells, amplifying Th17 responses and autoimmunity. Immunity 31, 331–341 (2009).This report shows that γδ T cells have an early role in promoting CNS inflammation. The authors suggest that innate cell‑produced IL‑17 directly enhances development of MOG‑specific TH17 cells.
18. Veldhoen, M. et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 453, 106–109 (2008).The data presented here suggest that AHR is a transcriptional regulator associated with regulatory T cells and IL‑17‑producing cells. Subsequent studies showed that AHR promotes IL‑22 production in both adaptive and innate cell populations.
19. Korn, T., Bettelli, E., Oukka, M. & Kuchroo, V. K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 27, 485–517 (2009).
20. McGeachy, M. J. & Cua, D. J. Th17 cell differentiation: the long and winding road. Immunity 28, 445–453 (2008).
21. Aggarwal, S., Ghilardi, N., Xie, M. H., de Sauvage, F. J. & Gurney, A. L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 278, 1910–1914 (2003).
22. Murphy, C. A. et al. Divergent pro- and antiinflammatory roles for IL-23 and IL-12 in joint autoimmune inflammation. J. Exp. Med. 198, 1951–1957 (2003).This early study shows that IL‑23‑deficient mice lack IL‑17‑producing cells following antigenic challenge, which suggests a role for IL‑17+ TH cells in promoting autoimmune inflammation.
23. Uhlig, H. H. et al. Differential activity of IL-12 and IL-23 in mucosal and systemic innate immune pathology. Immunity 25, 309–318 (2006).This study shows that IL‑23 can activate innate IL‑17‑producing cells in RAG‑deficient mice. IL‑23 acts locally in gut mucosal tissues whereas IL‑12 has a dominant role in promoting systemic inflammation.
24. Cua, D. J. et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 421, 744–748 (2003).IL‑23 directly promotes innate cell production of inflammatory cytokines. This is the first suggestion that IL‑23 acts on both innate and adaptive populations.
25. Aujla, S. J. et al. IL-22 mediates mucosal host defense against Gram-negative bacterial pneumonia. Nature Med. 14, 275–281 (2008).
26. Cella, M. et al. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 457, 722–725 (2009).This study shows NK‑like cells that produce IL‑22 are essential for host defence against gut bacterial infection. In this mouse system, the NKp46+ IL‑22‑producing cells do not seem to produce IL‑17.
27. Crellin, N. K., Trifari, S., Kaplan, C. D., Cupedo, T. & Spits, H. Human NKp44+IL-22+ cells and LTi-like cells constitute a stable RORC+ lineage distinct from conventional natural killer cells. J. Exp. Med. 207, 281–290 (2010).This study shows that human RORγt+ NK‑like cells can be isolated from peripheral blood mononuclear cells. These cells produce both IL‑17 and IL‑22 and are probably the counterparts of mouse NKp46+ IL‑22‑producing cells. A subsequent study from the same group showed that NKp46+ cells may develop from LTi‑like cells in the human system.
28. Takatori, H. et al. Lymphoid tissue inducer-like cells are an innate source of IL-17 and IL-22. J. Exp. Med. 206, 35–41 (2009).The is the first study to show that KIT+LIN– LTi‑like cells are an innate source of IL‑17 and IL‑22 during an immune response to fungal antigens.
29. Roark, C. L. et al. Exacerbation of collagen-induced arthritis by oligoclonal, IL-17-producing γδ T cells. J. Immunol. 179, 5576–5583 (2007).
30. Ito, Y. et al. γδ T cells are the predominant source of interleukin-17 in affected joints in collagen-induced arthritis, but not in rheumatoid arthritis. Arthritis Rheum. 60, 2294–2303 (2009).
31. Cui, Y. et al. Major role of γδ T cells in the generation of IL-17+ uveitogenic T cells. J. Immunol. 183, 560–567 (2009).
32. Fenoglio, D. et al. Vδ1 T lymphocytes producing IFN-γ and IL-17 are expanded in HIV-1-infected patients and respond to Candida albicans. Blood 113, 6611–6618 (2009).
33. Aujla, S. J., Dubin, P. J. & Kolls, J. K. Th17 cells and mucosal host defense. Semin. Immunol. 19, 377–382 (2007).
34. Lockhart, E., Green, A. M. & Flynn, J. L. IL-17 production is dominated by γδ T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J. Immunol. 177, 4662–4669 (2006).
35. Hayday, A. C. γδ T cells and the lymphoid stress-surveillance response. Immunity 31, 184–196 (2009).
36. Michel, M. L. et al. Identification of an IL-17-producing NK1.1neg iNKT cell population involved in airway neutrophilia. J. Exp. Med. 204, 995–1001 (2007).
37. Passos, S. T. et al. IL-6 promotes NK cell production of IL-17 during toxoplasmosis. J. Immunol. 184, 1776–1783 (2010).
38. Jensen, K. D. et al. Thymic selection determines γδ T cell effector fate: antigen-naive cells make interleukin-17 and antigen-experienced cells make interferon γ. Immunity 29, 90–100 (2008).
39. Ribot, J. C. et al. CD27 is a thymic determinant of the balance between interferon-γ- and interleukin 17-producing γδ T cell subsets. Nature Immunol. 10, 427–436 (2009).
40. Haas, J. D. et al. CCR6 and NK1.1 distinguish between IL-17A and IFN-γ-producing γδ effector T cells. Eur. J. Immunol. 39, 3488–3497 (2009).
41. Shibata, K. et al. Identification of CD25+ γδ T cells as fetal thymus-derived naturally occurring IL-17 producers. J. Immunol. 181, 5940–5947 (2008).
42. Laurence, A. et al. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity 26, 371–381 (2007).
43. Kisielow, J., Kopf, M. & Karjalainen, K. SCART scavenger receptors identify a novel subset of adult γδ T cells. J. Immunol. 181, 1710–1716 (2008).
44. Sun, Z. et al. Requirement for RORγ in thymocyte survival and lymphoid organ development. Science 288, 2369–2373 (2000).
45. Eberl, G. et al. An essential function for the nuclear receptor RORγt in the generation of fetal lymphoid tissue inducer cells. Nature Immunol. 5, 64–73 (2004).
46. Fukuyama, S. et al. Initiation of NALT organogenesis is independent of the IL-7R, LTβR, and NIK signaling pathways but requires the Id2 gene and CD3–
CD4+CD45+ cells. Immunity 17, 31–40 (2002).47. Adachi, S. et al. Essential role of IL-7 receptor α in the
formation of Peyer’s patch anlage. Int. Immunol. 10, 1–6 (1998).
48. Yoshida, H. et al. IL-7 receptor alpha+ CD3– cells in the embryonic intestine induces the organizing center of Peyer’s patches. Int. Immunol. 11, 643–655 (1999).
49. Meier, D. et al. Ectopic lymphoid-organ development occurs through interleukin 7-mediated enhanced survival of lymphoid-tissue-inducer cells. Immunity 26, 643–654 (2007).
50. Cupedo, T., Kraal, G. & Mebius, R. E. The role of CD45+CD4+CD3– cells in lymphoid organ development. Immunol. Rev. 189, 41–50 (2002).
51. Cupedo, T. & Mebius, R. E. Cellular interactions in lymph node development. J. Immunol. 174, 21–25 (2005).
52. Finke, D., Acha-Orbea, H., Mattis, A., Lipp, M. & Kraehenbuhl, J. CD4+CD3– cells induce Peyer’s patch development: role of α4β1 integrin activation by CXCR5. Immunity 17, 363–373 (2002).
53. Mebius, R. E. Organogenesis of lymphoid tissues. Nature Rev. Immunol. 3, 292–303 (2003).
54. Cupedo, T. et al. Human fetal lymphoid tissue-inducer cells are interleukin 17-producing precursors to RORC+ CD127+ natural killer-like cells. Nature Immunol. 10, 66–74 (2009).Human NK‑like cells produce both IL‑17 and IL‑22 and may share a common lineage with LTi‑like cells. Their ability to produce both IL‑17 and IL‑22 is in contrast to mouse NKp46+ cells, which only produce IL‑22.
55. Lane, P. et al. Lymphoid tissue inducer cells in adaptive CD4 T cell dependent responses. Semin. Immunol. 20, 159–163 (2008).
56. Mebius, R. E., Rennert, P. & Weissman, I. L. Developing lymph nodes collect CD4+CD3– LTβ+ cells that can differentiate to APC NK cells, and follicular cells but not T or B cells. Immunity 7, 493–504 (1997).
57. Kronenberg, M. & Engel, I. On the road: progress in finding the unique pathway of invariant NKT cell differentiation. Curr. Opin. Immunol. 19, 186–193 (2007).
58. Michel, M. L. et al. Critical role of ROR-γt in a new thymic pathway leading to IL-17-producing invariant NKT cell differentiation. Proc. Natl Acad. Sci. USA 105, 19845–19850 (2008).
59. Campillo-Gimenez, L. et al. AIDS progression is associated with the emergence of IL-17-producing cells early after simian immunodeficiency virus infection. J. Immunol. 184, 984–992 (2010).
60. Grajewski, R. S. et al. Activation of invariant NKT cells ameliorates experimental ocular autoimmunity by a mechanism involving innate IFN-γ production and dampening of the adaptive Th1 and Th17 responses. J. Immunol. 181, 4791–4797 (2008).
61. Luci, C. et al. Influence of the transcription factor RORγt on the development of NKp46+ cell populations in gut and skin. Nature Immunol. 10, 75–82 (2009).One of the first reports suggesting NKp46+ cells are the early innate IL‑22‑producing cells that have a role in skin and mucosal tissue repair and homeostasis.
62. Satoh-Takayama, N. et al. IL-7 and IL-15 independently program the differentiation of intestinal CD3–NKp46+ cell subsets from Id2-dependent precursors. J. Exp. Med. 207, 273–280 (2010).
63. Dhiman, R. et al. IL-22 produced by human NK cells inhibits growth of Mycobacterium tuberculosis by enhancing phagolysosomal fusion. J. Immunol. 183, 6639–6645 (2009).
64. Sanos, S. L. & Diefenbach, A. Isolation of NK cells and NK-like cells from the intestinal lamina propria. Methods Mol. Biol. 612, 505–517 (2010).
65. Satoh-Takayama, N. et al. Microbial flora drives interleukin 22 production in intestinal NKp46+ cells that provide innate mucosal immune defense. Immunity 29, 958–970 (2008).
66. Satoh-Takayama, N. et al. The natural cytotoxicity receptor NKp46 is dispensable for IL-22-mediated innate intestinal immune defense against Citrobacter rodentium. J. Immunol. 183, 6579–6587 (2009).
67. Ayabe, T. et al. Secretion of microbicidal α-defensins by intestinal Paneth cells in response to bacteria. Nature Immunol. 1, 113–118 (2000).
68. Wilson, C. L. et al. Regulation of intestinal α-defensin activation by the metalloproteinase matrilysin in innate host defense. Science 286, 113–117 (1999).
69. Bultinck, J., Brouckaert, P. & Cauwels, A. The in vivo contribution of hematopoietic cells to systemic TNF and IL-6 production during endotoxemia. Cytokine 36, 160–166 (2006).
70. Keshav, S. et al. Tumor necrosis factor mRNA localized to Paneth cells of normal murine intestinal epithelium by in situ hybridization. J. Exp. Med. 171, 327–332 (1990).
71. Lala, S. et al. Crohn’s disease and the NOD2 gene: a role for paneth cells. Gastroenterology 125, 47–57 (2003).
R E V I E W S
488 | jULy 2010 | vOLUme 10 www.nature.com/reviews/immunol
© 20 Macmillan Publishers Limited. All rights reserved10

72. Rumio, C. et al. Degranulation of paneth cells via toll-like receptor 9. Am. J. Pathol. 165, 373–381 (2004).
73. Salzman, N. H. et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nature Immunol. 11, 76–83 (2010).
74. Takahashi, N. et al. IL-17 produced by Paneth cells drives TNF-induced shock. J. Exp. Med. 205, 1755–1761 (2008).
75. Li, L. et al. IL-17 produced by neutrophils regulates IFN-γ-mediated neutrophil migration in mouse kidney ischemia-reperfusion injury. J. Clin. Invest. 120, 331–342 (2010).
76. Hoshino, A. et al. MPO-ANCA induces IL-17 production by activated neutrophils in vitro via classical complement pathway-dependent manner. J. Autoimmun. 31, 79–89 (2008).
77. Ivanov, I. I. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139, 485–498 (2009).Comparative analysis of microbiota from TH17 cell‑deficient and TH17 cell‑sufficient mice identified SFB as inducers of TH17 cells in the gut. This study suggests that commensal bacteria can shape the balance of immune cell subsets.
78. Ivanov, I. I. & Littman, D. R. Segmented filamentous bacteria take the stage. Mucosal Immunol. 3, 209–212 (2010).
79. Buonocore, S. et al. Innate lymphoid cells drive interleukin-23-dependent innate intestinal pathology. Nature 464, 1371–1375 (2010).This study identified THY1hiSCA1+ innate lymphoid cells that produce IL‑17 and IL‑22. In contrast to LTi‑like and NKp46+ cells, this newly described subset also produces IFNγ and is regulated by both RORγt and T‑bet.
80. Gaffen, S. L. Structure and signalling in the IL-17 receptor family. Nature Rev. Immunol. 9, 556–567 (2009).
81. Tanaka, S. et al. Natural occurring IL-17 producing T cells regulate the initial phase of neutrophil mediated airway responses. J. Immunol. 183, 7523–7530 (2009).
82. Rachitskaya, A. V. et al. Cutting edge: NKT cells constitutively express IL-23 receptor and RORγt and rapidly produce IL-17 upon receptor ligation in an IL-6-independent fashion. J. Immunol. 180, 5167–5171 (2008).
83. He, Y. W., Deftos, M. L., Ojala, E. W. & Bevan, M. J. RORγt, a novel isoform of an orphan receptor, negatively regulates Fas ligand expression and IL-2 production in T cells. Immunity 9, 797–806 (1998).
84. Doisne, J. M. et al. Skin and peripheral lymph node invariant NKT cells are mainly retinoic acid receptor-related orphan receptor γt+ and respond preferentially under inflammatory conditions. J. Immunol. 183, 2142–2149 (2009).
85. Esser, C., Rannug, A. & Stockinger, B. The aryl hydrocarbon receptor in immunity. Trends Immunol. 30, 447–454 (2009).
86. Schecter, A., Birnbaum, L., Ryan, J. J. & Constable, J. D. Dioxins: an overview. Environ. Res. 101, 419–428 (2006).
87. Rannug, A. et al. Certain photooxidized derivatives of tryptophan bind with very high affinity to the Ah receptor and are likely to be endogenous signal substances. J. Biol. Chem. 262, 15422–15427 (1987).
88. Quintana, F. J. et al. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
89. Martin, B., Hirota, K., Cua, D. J., Stockinger, B. & Veldhoen, M. Interleukin-17-producing γδ T cells selectively expand in response to pathogen products and environmental signals. Immunity 31, 321–330 (2009).
90. Zhou, L. & Littman, D. R. Transcriptional regulatory networks in Th17 cell differentiation. Curr. Opin. Immunol. 21, 146–152 (2009).
91. Kimura, A., Naka, T., Nohara, K., Fujii-Kuriyama, Y. & Kishimoto, T. Aryl hydrocarbon receptor regulates Stat1 activation and participates in the development of Th17 cells. Proc. Natl Acad. Sci. USA 105, 9721–9726 (2008).
92. Duan, J., Chung, H., Troy, E. & Kasper, D. L. Microbial colonization drives expansion of IL-1 receptor 1-expressing and IL-17-producing γ/δ T cells. Cell Host Microbe 7, 140–150 (2010).
93. Yoshiga, Y. et al. Invariant NKT cells produce IL-17 through IL-23-dependent and -independent pathways with potential modulation of Th17 response in collagen-induced arthritis. Int. J. Mol. Med. 22, 369–374 (2008).
94. Riol-Blanco, L. et al. IL-23 receptor regulates unconventional IL-17-producing T cells that control bacterial infections. J. Immunol. 184, 1710–1720 (2010).
95. Curtis, M. M. & Way, S. S. Interleukin-17 in host defence against bacterial, mycobacterial and fungal pathogens. Immunology 126, 177–185 (2009).
96. Iwakura, Y., Nakae, S., Saijo, S. & Ishigame, H. The roles of IL-17A in inflammatory immune responses and host defense against pathogens. Immunol. Rev. 226, 57–79 (2008).
97. Kolls, J. K. & Linden, A. Interleukin-17 family members and inflammation. Immunity 21, 467–476 (2004).
98. Ye, P. et al. Interleukin-17 and lung host defense against Klebsiella pneumoniae infection. Am. J. Respir. Cell. Mol. Biol. 25, 335–340 (2001).
99. Umemura, M. et al. IL-17-mediated regulation of innate and acquired immune response against pulmonary Mycobacterium bovis bacille Calmette-Guerin infection. J. Immunol. 178, 3786–3796 (2007).
100. Peng, M. Y. et al. Interleukin 17-producing γδ T cells increased in patients with active pulmonary tuberculosis. Cell. Mol. Immunol. 5, 203–208 (2008).
101. D’Souza, C. D. et al. An anti-inflammatory role for γδ T lymphocytes in acquired immunity to Mycobacterium tuberculosis. J. Immunol. 158, 1217–1221 (1997).
102. Khader, S. A. et al. IL-23 and IL-17 in the establishment of protective pulmonary CD4+ T cell responses after vaccination and during Mycobacterium tuberculosis challenge. Nature Immunol. 8, 369–377 (2007).
103. Molne, L., Corthay, A., Holmdahl, R. & Tarkowski, A. Role of γ/δ T cell receptor-expressing lymphocytes in cutaneous infection caused by Staphylococcus aureus. Clin. Exp. Immunol. 132, 209–215 (2003).
104. Cho, J. S. et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Invest. 120, 1762–1773 (2010).
105. Shibata, K., Yamada, H., Hara, H., Kishihara, K. & Yoshikai, Y. Resident Vδ1+ γδ T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J. Immunol. 178, 4466–4472 (2007).
106. Siegemund, S. et al. Differential IL-23 requirement for IL-22 and IL-17A production during innate immunity against Salmonella enterica serovar Enteritidis. Int. Immunol. 21, 555–565 (2009).
107. Meeks, K. D., Sieve, A. N., Kolls, J. K., Ghilardi, N. & Berg, R. E. IL-23 is required for protection against systemic infection with Listeria monocytogenes. J. Immunol. 183, 8026–8034 (2009).
108. Kelly, M. N. et al. Interleukin-17/interleukin-17 receptor-mediated signaling is important for generation of an optimal polymorphonuclear response against Toxoplasma gondii infection. Infect. Immun. 73, 617–621 (2005).
109. Chen, Y., Chou, K., Fuchs, E., Havran, W. L. & Boismenu, R. Protection of the intestinal mucosa by intraepithelial γδ T cells. Proc. Natl Acad. Sci. USA 99, 14338–14343 (2002).
110. Kuhl, A. A. et al. Aggravation of intestinal inflammation by depletion/deficiency of γδ T cells in different types of IBD animal models. J. Leukoc. Biol. 81, 168–175 (2007).
111. Kinugasa, T., Sakaguchi, T., Gu, X. & Reinecker, H. C. Claudins regulate the intestinal barrier in response to immune mediators. Gastroenterology 118, 1001–1011 (2000).
112. Ogawa, A., Andoh, A., Araki, Y., Bamba, T. & Fujiyama, Y. Neutralization of interleukin-17 aggravates dextran sulfate sodium-induced colitis in mice. Clin. Immunol. 110, 55–62 (2004).
113. Liang, S. C. et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 203, 2271–2279 (2006).
114. Chan, Y. R. et al. Lipocalin 2 is required for pulmonary host defense against Klebsiella infection. J. Immunol. 182, 4947–4956 (2009).
115. Ismail, A. S., Behrendt, C. L. & Hooper, L. V. Reciprocal interactions between commensal bacteria and γδ intraepithelial lymphocytes during mucosal injury. J. Immunol. 182, 3047–3054 (2009).
116. Cash, H. L., Whitham, C. V., Behrendt, C. L. & Hooper, L. V. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science 313, 1126–1130 (2006).REG3γ is a potent antimicrobial agent that kills Gram‑positive bacteria. Subsequent studies showed that REG3γ is strongly induced by IL‑22.
117. Brandl, K., Plitas, G., Schnabl, B., DeMatteo, R. P. & Pamer, E. G. MyD88-mediated signals induce the bactericidal lectin RegIIIγ and protect mice against intestinal Listeria monocytogenes infection. J. Exp. Med. 204, 1891–1900 (2007).
118. Littman, D. R. & Rudensky, A. Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 140, 845–858 (2010).
119. Ely, L. K., Fischer, S. & Garcia, K. C. Structural basis of receptor sharing by interleukin 17 cytokines. Nature Immunol. 10, 1245–1251 (2009).
120. Ishigame, H. et al. Differential roles of interleukin-17A and -17F in host defense against mucoepithelial bacterial infection and allergic responses. Immunity 30, 108–119 (2009).
121. Wucherpfennig, K. W. et al. γδ T-cell receptor repertoire in acute multiple sclerosis lesions. Proc. Natl Acad. Sci. USA 89, 4588–4592 (1992).
122. Shimonkevitz, R., Colburn, C., Burnham, J. A., Murray, R. S. & Kotzin, B. L. Clonal expansions of activated γ/δ T cells in recent-onset multiple sclerosis. Proc. Natl Acad. Sci. USA 90, 923–927 (1993).
123. Kang, Z. et al. Astrocyte-restricted ablation of interleukin-17-induced Act1-mediated signaling ameliorates autoimmune encephalomyelitis. Immunity 32, 414–425 (2010).
124. Duerr, R. H. et al. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science 314, 1461–1463 (2006).First study to show that IL23R gene polymorphism is linked to human inflammatory bowel disease susceptibility. Numerous studies have since confirmed and extended this initial observation.
125. Capon, F. et al. Sequence variants in the genes for the interleukin-23 receptor (IL23R) and its ligand (IL12B) confer protection against psoriasis. Hum. Genet. 122, 201–206 (2007).
126. Rueda, B. et al. The IL23R Arg381Gln non-synonymous polymorphism confers susceptibility to ankylosing spondylitis. Ann. Rheum. Dis. 67, 1451–1454 (2008).
127. Griffiths, C. E. et al. Comparison of ustekinumab and etanercept for moderate-to-severe psoriasis. N. Engl. J. Med. 362, 118–128 (2010).
128. Steinman, L. Mixed results with modulation of TH-17 cells in human autoimmune diseases. Nature Immunol. 11, 41–44 (2010).
129. Hurst, S. D. et al. New IL-17 family members promote Th1 or Th2 responses in the lung: in vivo function of the novel cytokine IL-25. J. Immunol. 169, 443–453 (2002).
130. Guo, P. et al. Dual nature of the adaptive immune system in lampreys. Nature 459, 796–801 (2009).
131. Franke, A. et al. Genome-wide association study for ulcerative colitis identifies risk loci at 7q22 and 22q13 (IL17REL). Nature Genet. 42, 292–294 (2010).
132. Fort, M. M. et al. IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 15, 985–995 (2001).
133. Colonna, M. Interleukin-22-producing natural killer cells and lymphoid tissue inducer-like cells in mucosal immunity. Immunity 31, 15–23 (2009).
Acknowledgements.We thank M. McGeachy and K. Boniface for insightful discus-sion and R. Kastelein for critical reading of the manuscript.
Competing interests statementThe authors declare no competing financial interests.
DATABASESUniProtKB: http://www.uniprot.orgACT1 | AHR | CD7 | CD25 | CD27 | CD70 | CD122 | CD161 | CLEC7A | CXCL8 | GATA3 | G‑CSF | IFNγ | IL‑1R1 | IL‑4 | IL‑6 | IL‑12p35 | IL‑17A | IL‑17F | IL‑21 | IL‑22 | IL‑23R | RORγt | RUNX1 | STAT1 | STAT3 | STAT4 | STAT6 | T‑bet | TLR2
All lInkS ARe ACTIve In The onlIne pDF
R E V I E W S
NATURe RevIewS | Immunology vOLUme 10 | jULy 2010 | 489
© 20 Macmillan Publishers Limited. All rights reserved10

CORRIGENDUM
Innate IL‑17‑producing cells: the sentinels of the immune system Daniel J. Cua and Cristina M. TatoNature Reviews Immunology 10, 479–489 (2010), doi:10.1038/nri2800; published online 18 June 2010; corrected online 25 June 2010
In the version of the article initially published, the table in BOX 1 incorrectly listed IL-17RB as the receptor for IL-17E (also known as IL-25). The correct receptor should have been IL-17RA– IL-17RB.
© 20 Macmillan Publishers Limited. All rights reserved10









![Pulmonary type2 innate lymphoid cells in paediatric severe ... · the induction of type 2 innate lymphoid cells (ILCs) [12-14]. ILCs are a rare population of cells of lymphoid lineage,](https://img.pdfslide.us/doc/110x75/5e17e22a8563b16def417337/pulmonary-type2-innate-lymphoid-cells-in-paediatric-severe-the-induction-of.jpg)