Embed Size (px)
Citation preview

1
INJURY AND THE BRAIN BARRIERS
Co-Chairs: Thomas P. Davis1, Gary A. Rosenberg2
Contributors: Ingolf E. Blasig3, Adam Chodobski4, Joanna Szmydynger-Chodobska5, Paula
Dore-Duffy6, Joe Fenstermacher7, Damir Janigro8, Eng Lo9, David Miller10, Sukriti Nag11, Martha
O’Donnell12, Darryl Peterson13, Jane Preston14, Norman Saunders15, Danica Stanimirovic16
1University of Arizona, Pharmacology Department, 1501 North Campbell Avenue, LSN 542,
Tucson, Arizona, USA 85724, [email protected], 520-626-7643
2University of New Mexico, Department of Neurology, HSC Neurology MSC10 5620 1 UMN,
Albuquerque, New Mexico, USA 97131, [email protected], 505-272-3315
3 Leibnizinstitute of Molecular Pharmacology, Berlin-Buch, Germany
4 Rhode Island Hospital and Brown University School of Medicine, Providence, Rhode Island,
USA
5 Rhode Island Hospital and Brown University School of Medicine, Providence, Rhode Island,
USA
6 Wayne State University, Detroit, Michigan, USA
7 Henry Ford Health System, Detroit, Michigan, USA
8Cleveland Clinic Foundation, Cleveland, Ohio, USA
9Massachusetts General Hospital, Charlestown, Massachusetts, USA
10National Institutes of Health/National Institute of Environmental Health Sciences, Research
Triangle Park, North Carolina, USA
11 University of Toronto, Toronto, Ontario, Canada
12 University of California Davis, California, USA
13 The Chicago Medical School, North Chicago, Illinois, USA
14 King’s College London, London, United Kingdom
15 University of Melbourne, Parkville, Australia

2
16National Research Council Canada, Ottawa, Ontario, Canada

3
Introduction
Neurotrauma and related neuropathological states (stroke, head injury,
ischemia/reperfusion, hemorrhage, infarction and hypertension) share several common features,
one of which is an alteration of the blood-brain barrier in the area of injury, 1 which may extend to
surrounding tissues. These barriers play an important role in overall CNS homeostasis including
regulation of the flow of nutrients, ions, water, metabolites and xenobiotics from blood to CNS and
vice versa 2,3 as well as in preventing CNS entry of potentially neurotoxic plasma constituents.4,5
Thus, such changes can profoundly affect the progression of injury (including loss of neurons and
specific CNS functions), responses to therapy and the time course and extent of recovery.
Clearly, it is important to understand both the basic, underlying biology (at the functional and
molecular levels) of the barrier tissues and how these change following injury.2,3,6 As discussed
below, these issues are complex in that the barriers reflect the combined function of multiple
proteins, cellular structures and cell types and these can both sense and respond to their
immediate microenvironment, with a range of responses from subtle to dramatic.
The past two decades have seen the development of several fundamental concepts that
have collectively changed our view of barrier function. In some cases these concepts have been
integrated into our understanding of brain injury; in others they have not, leaving open the
possibility that this new knowledge can be of benefit in the clinic by providing additional markers
of barrier alteration and new therapeutic targets.
It is now recognized that blood-brain barrier permeability to solutes reflects the
contributions of both passive and carrier-mediated processes and that these contributions (and
thus integrated barrier function) are dynamic rather than static. The passive barrier is
dominated by paracellular tight junctions 6,7 and very low cellular passive permeability.8
Transporters add the capability to increase or decrease apparent permeability for certain solutes
(selectivity). Given this complex mix of determinants, it should not be surprising that the blood
brain barrier does not behave like a binary system, either open or closed. Rather it is able to
express a number of permeability states, depending on the nature of the solute under study, the
state of the tight junction proteins and the activity of appropriate transporters.

4
Passive Permeability Markers
Progress:
The earliest markers used for determining disruption of the blood-brain barrier in a variety
of pathological states, which date back to the early 20th Century, were dyes such as trypan blue
or Evans blue.9,10 They have the advantage of being easily visualized, which is perhaps why they
continue to be used in spite of serious limitations. These dyes have been shown to bind firmly to
proteins in vitro,11 but may be exchangeable with tissues with higher affinity constituents in vivo.12
Thus it is not always clear whether the visualized marker is the small dye molecule (mw
<1000Da) or the albumin,11 or other plasma proteins to which it is bound.13 This is important
because there is recent evidence suggesting that the barriers in brain and spinal cord may show
increased permeability to small molecules for several days after injury,14,15 whereas the barrier to
large molecules in most investigations has been found to be closed by 3-6 hours post injury.
Since the 1960s the plant enzyme horseradish peroxidase16 has been used extensively to test
barrier integrity following neurotrauma.17,18 It has the practical advantages of being relatively easy
to visualize at the light microscopical and electronmicoscopical level. However, it has a number of
potential artifactual problems that were well recognized when it was first introduced into studies of
blood-brain barriers in the 1960s (outlined in 14)14, but which seem often to be overlooked in
more recent studies. This may explain some of the discrepancies in the literature. For example
the duration of barrier opening to HRP following neurotrauma has been variously reported as 6
hours to 24 hours plus.14 The mechanism of barrier disruption is also disputed, some authors
reporting leaks of HRP via tight junctions and others by transcellular mechanisms. A few studies
have used immunocytochemical staining for endogenous plasma proteins,9,14 such as albumin,19
fibronectin14 or IgG,20 to measure barrier integrity.9,14 These can show whether “breakdown” has
occurred, but are not suitable for defining its time course or extent.
Unresolved and New Questions:

5
The time course of “barrier breakdown” to different sized molecules is important for two
reasons: (a) While barrier permeability is increased, its protective function is reduced and the
brain/spinal cord is vulnerable to entry of potentially neurotoxic agents occurring naturally in the
blood (i.e. albumin). (b) The period when the barrier is more permeable can represent a
therapeutic opportunity to use neuroprotectant agents that would not normally enter the CNS.
Indeed, there is reasonable evidence suggesting a period of 4-6 hours following several types of
trauma to the brain when the barrier is permeable to large protein-sized molecules. In the spinal
cord this period may be as long as 24 hours. Recent evidence also suggests that for molecules
<10 kDa there is a period of 4 days or more after injury when the both blood-brain and blood-
spinal cord barriers are more permeable at the site of injury.14,15 The spatial resolution of most
methods is such that it is unclear whether increased permeability is confined to the blood vessels
injured in the original trauma, although there are claims that blood vessels in surrounding neural
tissue are also affected. These claims remain controversial and largely unresolved.
Tight Junction Proteins
Progress:
At the molecular level, recent work has identified many new, critical proteins in endothelial tight
junctions3,6 and caveolae.21 The tight junction proteins: occludin, claudins 3, 5 and 12, junctional
adhesion molecule A and zonula occludens proteins have been localized in cerebral endothelium
by immunohistochemistry.22-24 In vitro studies of brain injury or stress, such as hypoxia25 and in
vivo studies of systemic inflammation26 show redistribution and reduced expression of tight
junctional proteins, which increases the paracellular barrier permeability.1,3,26 These data show
that the blood-brain barrier tight junctions are compromised with specific changes occurring at the
level of specific tight junction proteins. However, ultrastructural studies of the pathways for
visualisable tracers such as HRP in blood-brain barrier experiments in vivo have given conflicting
results concerning the route by which macromolecules such as HRP cross the disrupted blood-
brain barrier.1,17

6
Unresolved and New Questions:
It is important to discover the structure, function and regulation of these tight junction
proteins in the maintenance of blood-brain barrier homeostasis and the extent to which they are
affected in brain injury or stress associated with diseases of the CNS. Such studies could be very
useful for the implementation and development of approaches that could protect the barrier. It is
also important to understand the fate and role that tight junction proteins play in maintaining the
integrity of the barrier during health and disease. Areas such as tight junction protein trafficking,
associated with barrier formation and maintenance, are critical to understand and study. Defining
the mechanism of tight junction formation of the barrier is also highly significant to understanding
the consequence of blood-brain barrier “leak” and the potential for repair.
Influx and Efflux Transporters
Progress:
A literature search reveals that this aspect of barrier trauma is largely unstudied, although
recent work with animal models suggests important implications. One study on morphine
pharmacokinetics in patients with severe brain injury found that morphine permeability was
increased in “worse” affected parts of the injured brains.27 14 C-� – Aminoisobutyric acid has been
used in a few studies but at saturating concentrations,15 so that what was being measured was
non-specific transfer of a small molecule. These transporter-based influx and efflux mechanisms
are essential for control of the normal internal environment of the brain and for the supply of
essential nutrients. Disruption of these transport mechanisms is likely to contribute to secondary
consequences of neurotrauma and their repair will contribute to the damage control and to
recovery.
Striking examples of the importance of transporters as potential targets of therapy comes
from animal models of cerebral ischemia and epilepsy. First, edema following middle cerebral
artery occlusion is substantially attenuated when rats are treated with an inhibitor of the blood-
brain barrier Na-K-Cl cotransporter, a major contributor to the movement of Na+ and water into
the brain that is upregulated following ischemia.28 Second, temporary cerebral ischemia was

7
found to upregulate a blood-brain barrier drug efflux pump, p-glycoprotein. Pump inhibition
enhanced both the brain uptake and the effects of two neuroprotective drugs.29 Third, epileptic
seizures appear to increase brain capillary expression of multiple drug efflux pumps and to
reduce brain uptake of antiepileptic drugs (AEDs).30 Preliminary data in animal models and
patients show that inhibition of efflux transporter activity increases both the CNS accumulation
and the effectiveness of AEDs.31
Unresolved and New Questions:
Following brain injury, there are many unresolved and new questions such as: What is
the time course of barrier disruption and repair for different size molecules? Is this similar in
different regions of the brain and spinal cord?
Which elements of the barrier (tight junction proteins, transcytosis receptors, specific
transporters) are altered and when are they altered in the process? What are the molecular
targets that underlie barrier disruption? How does this change in a time course manner?
Can information on time course, brain regions and mechanisms of barrier injury be used
to facilitate delivery of neuroprotective drugs? Would it be important to develop barrier-protective
drugs with barrier tissues as therapeutic targets? Last of all, how do patterns of barrier disruption
depend upon the nature of the injury, (i.e., ischemic stroke versus hemorrhagic stroke versus
traumatic brain injury?
Each of these unresolved questions is critical to address and when done so will cause
many more new questions which need to be answered.
Signals Disrupting Barrier Function
Progress:
Complexity dominates here. Brain injury releases a cocktail of chemical signals, including
hormones, cytokines, chemokines, enzymes, free radicals, neurotransmitters, and ions. In
addition, injury activates the brain‘s innate immune response and a variety of peripheral
neuroinflammatory responses, both chemical and cellular (macrophage infiltration; see Report on

8
Inflammation and Brain Barriers). These can target all elements of the CNS including barrier
tissues. The latter are unique in that they are directly exposed to signals from both the CNS and
the periphery and can act as a conduit for exchange of signaling molecules, e.g. cytokines,
between the two compartments.
Moreover, although the major focus of previous studies has been the cerebral
endothelium, it has been clear for some time that other elements of the neurovascular unit, such
as neurons, pericytes and astrocytes, communicate directly with the endothelium and can
profoundly affect barrier function. Thus, with respect to brain injury, barrier research requires a
broader focus to define the changes occurring in other components of the neurovascular unit and
the relationship of these changes to those in and around the endothelium. Consider, for example,
the role of the extracellular matrix in barrier injury. It has been recognized for many years that
there is a biphasic opening of the blood-brain barrier in reperfusion injury.32 Correlating structure
with function has shown that the early reversible barrier opening is caused by the activation of the
constitutive matrix metalloproteinase-2 (MMP-2) by the hypoxia/ischemia. MMP-2 degrades
claudin-5 and occludin, and the breakdown of the tight junctions can be blocked by synthetic
MMP inhibitors.33 A delayed disruption of the blood-brain barrier that results from multiple factors
involved in the inflammatory response occurs after one to two days and is only slowly reversed.
This phase is associated with cytokines, free radicals and MMP-3 and MMP-9. Control of this
phase of injury has been a challenge and should be a major target of novel therapies.
Given the plethora of signals generated and the capability of barrier tissues to respond to
so many factors, one looks for a rational way to decide which signals/receptors, etc, should be
considered as priority targets in therapy and when. This point brings up an important and
troubling issue. So many stroke treatments that were promising in extensive animal studies
proved to be disappointing when brought to clinical trials. Indeed, we still have only one effective
treatment to limit stroke damage, tissue plasminogen activator, administered within 3 hours of an
ischemic stroke. From this one could easily conclude that available animal models are inadequate
to mimic all the critical aspects of stroke in patients. Could this be a result of species differences,
the age of the animals, or a lack of appreciation of the complexity of events, signals and targets?

9
Unresolved and New Questions:
Following injury, what is the time course of release of signaling molecules (CNS and
systemic) and how does this correlate with barrier disruption (tight junctions and transporters) and
repair? What is the role of the barrier tissues in hemorrhagic transformation?
Another important unresolved question is: can synergy be obtained by targeting more
than one signaling event, such as free radical damage, MMPs and proinflammatory cytokines,
and at the optimal times.
In the area of model development, can we develop better models for defined CNS
injuries, models that more closely mimic the human condition?
Stroke and Head Injury
Progress:
Stroke and head injury are two major areas of clinical blood-brain barrier (BBB) research.
The insights from in vitro studies of the molecular aspects of the BBB have been translated into in
vivo studies where disruption of the tight junction proteins is seen in experimental models of
injury, including reperfusion after stroke. Greater understanding of the extracellular matrix has
come from recent studies on the proteolytic disruption of basal lamina proteins, such as laminin,
fibronectin, and type IV collagen, and the tight junctions.34
In the past five years, the role of magnetic resonance imaging (MRI) has emerged as an
important method to visualize and quantify BBB opening in vivo. Fenstermacher and colleagues
adapted an autoradiographic graphical method to the MRI.35,36 Using Gadolinium-DTPD and fast
T1-weighted scanning sequences, they were able to measure the permeability coefficient, ki, in
rats with stroke. This is an important advance over the invasive methods that need to have tissue
samples because it provides visual information on the sites of BBB disruption, which is otherwise
not available with quantitative methods, and it is noninvasive. In addition, MRI can be used to
measure infarct size and cerebral perfusion in the intact animal so that long-term behavioral
measurements can be made. This combination of measurements on the same animal provides a
powerful means to assess therapies, which may have beneficial early effects, but later

10
detrimental ones. More importantly, the MRI graphical method that has been developed for
animals (Fig. 1) can be applied to studies in humans.
Unresolved and New Questions:
There are unresolved and new questions that need to be answered to achieve the goals
associated with advancing the area of stroke and head injury, one major obstacle is the expense
of the imaging equipment and the need for teams of scientists and engineers to carry out these
studies. This will necessitate designation of specialized centers for human studies and other
centers for molecular studies. A major problem is the lack of effective treatments for either head
trauma or stroke. Collaboration will be needed between academia and by big pharmaceutical
companies to speed discovery of therapies in these areas. This may require studies of multiple
agents to block several aspects of BBB damage; such studies provide unique challenges. These
obstacles can be overcome by the joining together of multiple centers for consolidation of studies,
sharing of information, and training of the next generation of experts on the BBB. The need to
train this next generation of BBB experts is critical to advance the field but remains unresolved.
Overall Concepts that need to be Developed
The prevailing medical textbook vision of brain barrier tissues as static structures whose
properties reflect solely tight junctional permeabilities is misleading and detrimental to research.
To fully understand the role that barrier tissues play in brain injury, this vision must be changed to
incorporate the following major concepts:
1) Barrier properties are a function of both paracellular and cellular elements, including tight
junctions, transcytotic mechanisms and specific influx and efflux transporters and
channels.
2) Barrier properties are dynamic, responding to signals received from both the CNS and the
periphery.

11
3) The brain capillary endothelium is itself one element of a neurovascular unit (along with
pericytes, astrocytes and neurons) that integrates environmental signals and together
determines blood-brain barrier function.
4) Brain barrier properties (functional and molecular), susceptibility to injury (response to signals)
and inherent neuroprotective mechanisms likely vary with region.
5) Brain barrier disruption is an important contributor to secondary injury following neurotrauma
and thus barrier protective therapeutics could provide an opportunity for neuroprotection.
Summary Points
What needs to be done?
1. Resources specifically focused on brain barrier research
• Funding for international (cross continent), translational research projects at the level of
2-3 laboratories within the same discipline but having complementary knowledge.
• Increased highly targeted funding opportunities with specified set-asides
• Generation of databases listing BBB specific genes, reagents, antibodies, primers, siRNA
etc. Development of a BBB- Gene-Map similar to the brain-map published in Nature
January 2007
2. Training/Personnel
• Too few groups investigating BBB trauma at the mechanistic level (driven traditionally by
physiologists and pharmacologists). There is a critical need for cellular and molecular
biologists, neurobiologists and immunologists investigating trauma and the BBB.

12
• A critical mass of BBB researchers is lacking. This limits the intellectual advance of the
field and does not give scope for replication of key findings or even dissemination to
peripheral fields. Establish a graduate student program specifically designed to attract
high quality students to work on the BBB with the opportunity to spend time “rotating” in
other BBB laboratories in the USA or abroad.
• BBB research in neurotrauma should be developed as a distinct area – until today it is
hidden in many different disciplines (i.e. research on barrier trauma damage is mostly a
minor component of neurotrauma research using out of date methods).
• Cell biologists and protein biochemists working on tight junctions and isolation of
transporter biology need to be attracted into the BBB field. Most research by this group of
researchers is performed in epithelial cells, which are distinct from endothelial cells.
• Better behavioral methods are required for evaluating effects of barrier dysfunction during
development. This will probably require attracting behavioral scientists into the field.
Conclusion and Single Most Important Issue
Future directions of research in the BBB of trauma and stroke will involve studies of both
the natural history of brain barrier disruption in human subjects and parallel studies using
proteomics 37,38 and genomics,39 of the molecular events in animal models and in vitro.3,6,7 It is
crucial to define the natural history in humans to improve the experimental models used today,
but it is also important to use modern genomic and proteomic methods to discover new
mechanisms that could lead to successful human therapy. Pathway analysis should identify
upstream genes that will be potential therapeutic targets for amelioration of the deleterious effects
of blood-brain barrier disruption in trauma and other pathologies. In parallel, the structure,
function and regulation of the proteins tightening the blood-brain barrier have to be elucidated to
define novel target molecules and modulators for barrier protection (protein structure
factories).3,6,40 To limit brain injury and speed recovery, there is an urgent need for high
resolution, non-invasive, in vivo tools (molecular probes and detection technology) that will
facilitate monitoring of brain barrier function (tight junctions, transporters and channels) within the

13
components of the neurovascular unit. These tools should include CNS-deliverable, molecular
level reporters of expression (small molecules that bind specifically) and of activity (substrates) as
well as inhibitors that can be used in therapy. To aid translational studies, these molecules should
be usable in both cell-based and animal-based models as well as in patients. In addition,
appropriate imaging technology (PET, MRI multiphoton/confocal) should permit both time-based
and spatially based monitoring over a range of resolutions. Advances in imaging are the single
most critical issue for the advancement of understanding in the field of Brain Injury and for the
development of new treatments.

14
References
1. Nag S. Pathophysiology of blood-brain barrier breakdown. Methods Mol Med 2003;89:97-
119.
2. Davson H, Segal MB. Physiology of the CSF and Blood-Brain Barriers. New York CRC
Press, 1995.
3. Hawkins BT, Davis TP. The blood-brain barrier/neurovascular unit in health and disease.
2005;57(2):173-185.
4. Nordborg C, Sokrab TE, Johansson BB. The relationship between plasma protein
extravasation and remote tissue changes after experimental brain infarction. Acta
Neuropathol (Berl) 1991;82(2):118-26.
5. Wagner KR, Packard BA, Hall CL, Smulian AG, Linke MJ, De Courten-Myers GM,
Packard LM, Hall NC. Protein oxidation and heme oxygenase-1 induction in porcine white
matter following intracerebral infusions of whole blood or plasma. Dev Neurosci
2002;24(2-3):154-60.
6. Huber JD, Egleton RD, Davis TP. Molecular physiology and pathophysiology of tight
junctions in the blood-brain barrier. Trends Neurosci 2001;24(12):719-25.
7. Nag S. Morphology and molecular properties of cellular components of normal cerebral
vessels. Methods Mol Med 2003;89:3-36.
8. Ek CJ, Dziegielewska KM, Stolp H, Saunders NR. Functional effectiveness of the blood-
brain barrier to small water-soluble molecules in developing and adult opossum
(Monodelphis domestica). J Comp Neurol 2006;496(1):13-26.
9. Nag S. Blood-brain barrier permeability using tracers and immunohistochemistry.
Methods Mol Med 2003;89:133-44.

15
10. Bradbury M. The Concept of a Blood-Brain Barrier. New York: John Wiley and Sons,
1979.
11. Tschirgi RD. Protein complexes and the impermeability of the blood-brain barrier to dyes.
Am J Physiol 1950;163:756.
12. Dallal MM, Chang SW. Evans blue dye in the assessment of permeability-surface are
product in perfused rat lungs. J Appl Physiol 1994;77(2):1030-5.
13. Hansen P, Nielsen NC. The Binding of Evans Blue to Plasma Proteins. an Evaluation
Using Adsorption onto Sephadex. Scand J Clin Lab Invest 1964;16:491-7.
14. Habgood MD, Bye N, Dziegielewska KM, Ek CJ, Lane MA, Potter A, Morganti-Kossmann
C, Saunders NR. Changes in blood-brain barrier permeability to large and small
molecules following traumatic brain injury in mice. Eur J Neurosci 2007;25(1):231-8.
15. Popovich PG, Horner PJ, Mullin BB, Stokes BT. A quantitative spatial analysis of the
blood-spinal cord barrier. I. Permeability changes after experimental spinal contusion
injury. Exp Neurol 1996;142(2):258-75.
16. Reese TS, Karnovsky MJ. Fine structural localization of a blood-brain barrier to
exogenous peroxidase. J Cell Biol 1967;34(1):207-17.
17. Lossinsky AS, Shivers RR. Structural pathways for macromolecular and cellular transport
across the blood-brain barrier during inflammatory conditions. Review. Histol Histopathol
2004;19(2):535-64.
18. Stewart PA, Farrell CR, Farrell CL, Hayakawa E. Horseradish peroxidase retention and
washout in blood-brain barrier lesions. J Neurosci Methods 1992;41(1):75-84.

16
19. Abraham CS, Harada N, Deli MA, Niwa M. Transient forebrain ischemia increases the
blood-brain barrier permeability for albumin in stroke-prone spontaneously hypertensive
rats. Cell Mol Neurobiol 2002;22(4):455-62.
20. Hoane MR, Kaplan SA, Ellis AL. The effects of nicotinamide on apoptosis and blood-
brain barrier breakdown following traumatic brain injury. Brain Res 2006;1125(1):185-93.
21. Frank PG, Woodman SE, Park DS, Lisanti MP. Caveolin, caveolae, and endothelial cell
function. Arterioscler Thromb Vasc Biol 2003;23(7):1161-8.
22. Ballabh P, Hu F, Kumarasiri M, Braun A, Nedergaard M. Development of tight junction
molecules in blood vessels of germinal matrix, cerebral cortex, and white matter. Pediatr
Res 2005;58(4):791-8.
23. Virgintino D, Errede M, Robertson D, Capobianco C, Girolamo F, Vimercati A, Bertossi
M, Roncali L. Immunolocalization of tight junction proteins in the adult and developing
human brain. Histochem Cell Biol 2004;122(1):51-9.
24. Wolburg H, Wolburg-Buchholz K, Kraus J, Rascher-Eggstein G, Liebner S, Hamm S,
Duffner F, Grote EH, Risau W, Engelhardt B. Localization of claudin-3 in tight junctions of
the blood-brain barrier is selectively lost during experimental autoimmune
encephalomyelitis and human glioblastoma multiforme. Acta Neuropathol (Berl)
2003;105(6):586-92.
25. Abbruscato TJ, Davis TP. Combination of hypoxia/aglycemia compromises in vitro blood-
brain barrier integrity. J Pharmacol Exp Ther 1999;289(2):668-75.
26. Brooks TA, Ocheltree SM, Seelbach MJ, Charles RA, Nametz N, Egleton RD, Davis TP.
Biphasic cytoarchitecture and functional changes in the BBB induced by chronic
inflammatory pain. Brain Res 2006;1120(1):172-82.

17
27. Ederoth P, Tunblad K, Bouw R, Lundberg CJF, Ungerstedt U, Nordstrom C-H,
Hammarlund-Udenaes M. Blood-brain barrier transport of morphine in patients with
severe brain trauma. British Journal of Clinical Pharmacology 2004;57(4):427-435.
28. Pedersen SF, O'Donnell ME, Anderson SE, Cala PM. Physiology and pathophysiology of
Na+/H+ exchange and Na+ -K+ -2Cl- cotransport in the heart, brain, and blood. Am J
Physiol Regul Integr Comp Physiol 2006;291(1):R1-25.
29. Spudich A, Kilic E, Xing H, Kilic U, Rentsch KM, Wunderli-Allenspach H, Bassetti CL,
Hermann DM. Inhibition of multidrug resistance transporter-1 facilitates neuroprotective
therapies after focal cerebral ischemia. Nat Neurosci 2006;9(4):487-8.
30. Oby E, Janigro D. The blood-brain barrier and epilepsy. Epilepsia 2006;47(11):1761-74.
31. Loscher W, Potschka H. Drug resistance in brain diseases and the role of drug efflux
transporters. Nat Rev Neurosci 2005;6(8):591-602.
32. Rosenberg GA, Estrada EY, Dencoff JE. Matrix metalloproteinases and TIMPs are
associated with blood-brain barrier opening after reperfusion in rat brain. Stroke
1998;29(10):2189-2195.
33. Yang Y, Estrada EY, Thompson JF, Liu W, Rosenberg GA. Matrix metalloproteinase-
mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic
matrix metalloproteinase inhibitor in focal ischemia in rat. J Cereb Blood Flow Metab
2007;27(4):697-709.
34. Cunningham LA, Wetzel M, Rosenberg GA. Multiple roles for MMPs and TIMPs in
cerebral ischemia. Glia 2005;50(4):329-339.
35. Ewing JR, Knight RA, Nagaraja TN, Yee JS, Nagesh V, Whitton PA, Li L, Fenstermacher
JD. Patlak plots of Gd-DTPA MRI data yield blood-brain transfer constants concordant

18
with those of 14C-sucrose in areas of blood-brain opening. Magn Res Med
2003;50(2):283-292.
36. Patlak CS, Blasberg RG, Fenstermacher JD. Graphical evaluation of blood-to-brain
transfer constants from multiple-time uptake data. J Cereb Blood Flow Metab
1983;3(1):1-7.
37. Enerson BE, Drewes LR. The rat blood-brain barrier transcriptome. J Cereb Blood Flow
Metab 2006;26(7):959-73.
38. Haqqani AS, Kelly J, Baumann E, Haseloff RF, Blasig IE, Stanimirovic DB. Protein
markers of ischemic insult in brain endothelial cells identified using 2D gel
electrophoresis and ICAT-based quantitative proteomics. J Proteome Res 2007;6(1):226-
39.
39. Pardridge WM. Blood-brain barrier genomics. Stroke 2007;38(2 Suppl):686-90.
40. Blasig IE, Winkler L, Lassowski B, Mueller SL, Zuleger N, Krause E, Krause G, Gast K,
Kolbe M, Piontek J. On the self-association potential of transmembrane tight junction
proteins. Cell Mol Life Sci 2006;63(4):505-14.

19
Figure Legend
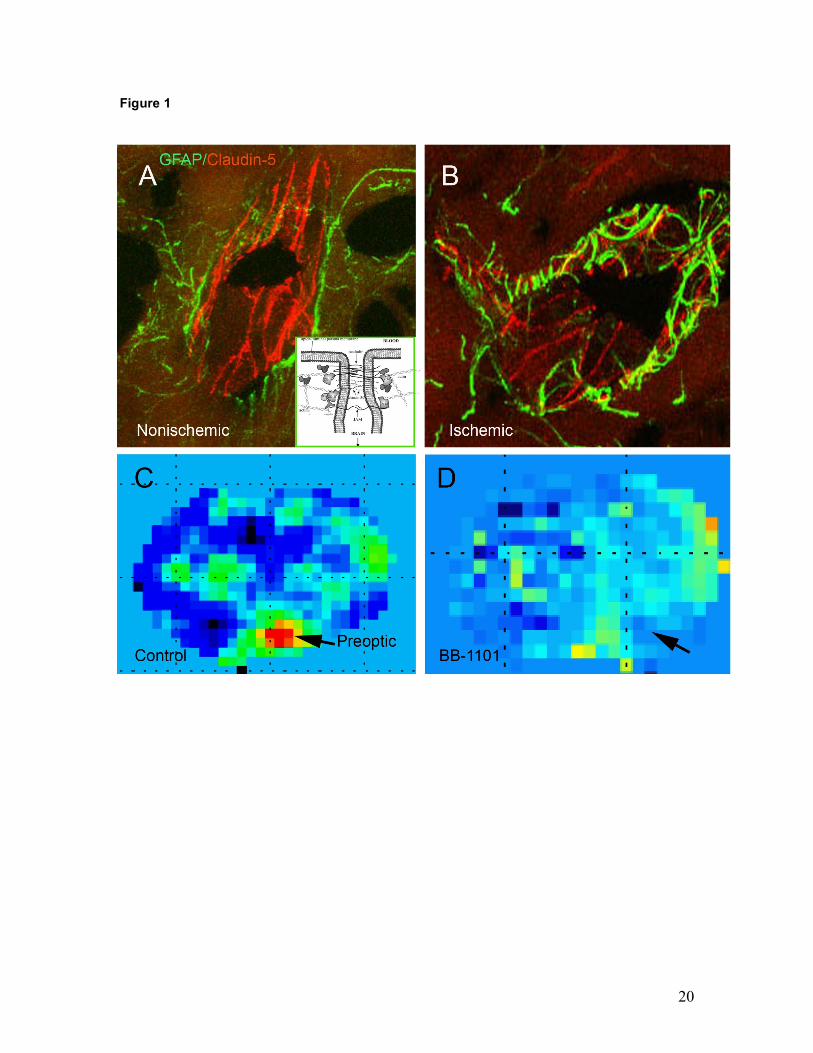
Figure 1: In vivo studies of the blood-brain barrier in rat with transient ischemia and reperfusion,
using the suture method. A) Brain section of a spontaneously hypertensive rat after 90 min of
ischemic middle cerebral artery occlusion (MCAO) with a suture and 3 h of reperfusion. A
confocal image of a blood vessel in the non-ischemic piriform cortex is shown. The astrocytes are
immunolabeled with GFAP (green) and the tight junction protein, claudin-5 is immunostained red.
Note the linear bands of claudin-5 surrounded by the GFAP-positive astrocytic foot processes. B)
A similar image from the ischemic piriform cortex showing disruption of the claudin-5 positive
bands. C) An MRI of an untreated Wistar rat with a 2 h MCAO followed by 3 h of reperfusion. The
red represents regions of high permeability calculated from the Gd-DTPA contrast enhanced
image using the Patlak Plot method. D) An MRI permeability map of a rat treated with the matrix
metalloproteinase inhibitor, BB-1101. The region of increased permeability is absent. The insert
shows the molecular components of the tight junction. (Modified from Yang et al.33 and Hawkins
and Davis3; MRI courtesy of Rohit Sood, PhD, MD. University of New Mexico, U.S.A.)
20
Figure 1











![Programa curt[1] · Ulla Knaus (University College, Dublin, Ireland): ROS at mucosal barriers. WORKSHOP: Redox-Omic Technologies (Sponsored by Promega) Chairs: Elena Hidalgo (Universitat](https://img.pdfslide.us/doc/110x75/5f3b3aeb5990ba394b68f559/programa-curt1-ulla-knaus-university-college-dublin-ireland-ros-at-mucosal.jpg)

















