Embed Size (px)
Citation preview

ELSEVIER Neuroscience Letters 190 (1995) 29-32
N[UROSCliNC[ IEIIEgS
Inhibition of nitric oxide synthesis potentiates the responsiveness of carotid chemoreceptors to systemic hypoxia in the rat
Andrzej Trzebski a, Yuko Sato b, Atsuko Suzuki c, Akio Sato c,*
aDepartment of Physiology, Medical Academy, Warsaw, Poland bLaboratory of Physiology, Tsukuba College of Technology, Tsukuba 305, Japan
CDepartment of the Autonomic Nervous System, Tokyo Metropolitan Institute of Gerontology, Tokyo 173, Japan
Received 15 March 1995; accepted 22 March 1995
Abstract
Carotid sinus nerve afferent activity was recorded in the peripheral end of the cut carotid sinus nerves in rats anesthetized with urethane, paralyzed and artificially ventilated with pure oxygen in order to abolish any resting chemoreceptor activity. Hypoxic stimuli were applied by switching pure oxygen to a nitrogen/oxygen gas mixture in the inspiratory line, reducing end-tidal oxygen concentrations to 10% FEXo2, 8% Fwro2 and 6% FE'rO2 respectively. Each stimulus was applied for 60 s and ventilation was switched again to pure oxygen. Increases in the carotid sinus nerve activities were due to chemo- and not to baroreceptor stimulation as arterial blood pressure decreased during hypoxia. After administration of nitric oxide synthase blocker L-NG-nitroarginine methyl ester, 30 mg/kg weight i.v., chemoreceptor excitatory response to all hypoxic stimuli increased significantly. Subsequent administration of L- arginine, 300 mg/kg weight i.v., restored chemoreceptor response to hypoxia to initial magnitude. It is concluded that NO is generated in the carotid body and attenuates chemoreceptor responsiveness in rats in vivo, as reported on isolated carotid bodies in cats in vitro.
Keywords: Nitric oxide; Nitric oxide synthase inhibition; Carotid chemoreceptors; Hypoxic responsiveness; Systemic hypoxia; Rat
Nitric oxide (NO), a powerful activator of guanylate cyclase [1], synthesized by constitutive NO-synthase (NOS) in the carotid body [21], is identified by immuno- cytochemical methods in the nerves supplying carotid glomic cells and carotid body vessels [5,6,12,13,20,21]. As guanylate cyclase is synthesizing a secondary messen- ger, cyclic GMP (cGMP), which appears to be a key me- diator of chemoreceptor inhibition [19], a NO-cGMP sig- naling pathway in the carotid body has been postulated recently as a tonic inhibitory mechanism for chemorecep- tion [21]. Experiments performed in vitro on isolated ca- rotid body preparations of cats superfused and/or perfused with inhibitors of NOS enzymatic activity or NO donors; nitroglycerine and nitroprusside or NOS substrate, L- arginine support this view [5,13,21].
Rat carotid body chemoreceptors studied in the whole animal are very sensitive to systemic hypoxia [3]. The purpose of the present study was to check the influence of
* Corresponding author, Tel.: +81 3 3964 3241, ext. 3087; Fax: +81 3 3964 1415.
inhibition of nitric oxide synthesis on the chemoreceptor excitatory response to brief hypoxia in vivo in the rat.
Carotid chemoreceptor responsiveness to systemic hy- poxia appeared to be significantly augmented by inhibi- tion of NOS, provided that the dose of L-N~-nitroarginine methyl ester (L-NAME) a NOS blocker, was much higher by i.v. administration than used in our preliminary ex- periments [18].
Experiments were carried out on 12 male Wistar rats weighing 350--420 g anesthetized with urethane (I. 1 g/kg weight i.p.). During the experiment, additional doses 1/5 of the initial dose, were administered i.v. at 1-2 h inter- vals to maintain anesthesia at a relatively constant level. The animals were artificially ventilated with pure oxygen by respirator pump (model 683, Harvard, USA) after im- mobilization with gallamine triethiodide (Sigma, USA). A polyethylene catheter for blood pressure measurement was inserted in the femoral artery. Another catheter for injection of drugs was inserted in the femoral vein. Rectal temperature was maintained at 37.0-38.0°C using a ther- mostatically-regulated heating pad and an infrared lamp
0304-3940/95/$09.50 © 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0304-3940(95)11492-C
30 A. Trzebski et al. / Neuroscience Letters 190 (1995) 29-32
control after L-NAME after L-Arg (30mg/kg) (300mg/kg) imp/5s
0
.~100
6%02
•1150 ~1100
stim., 60s
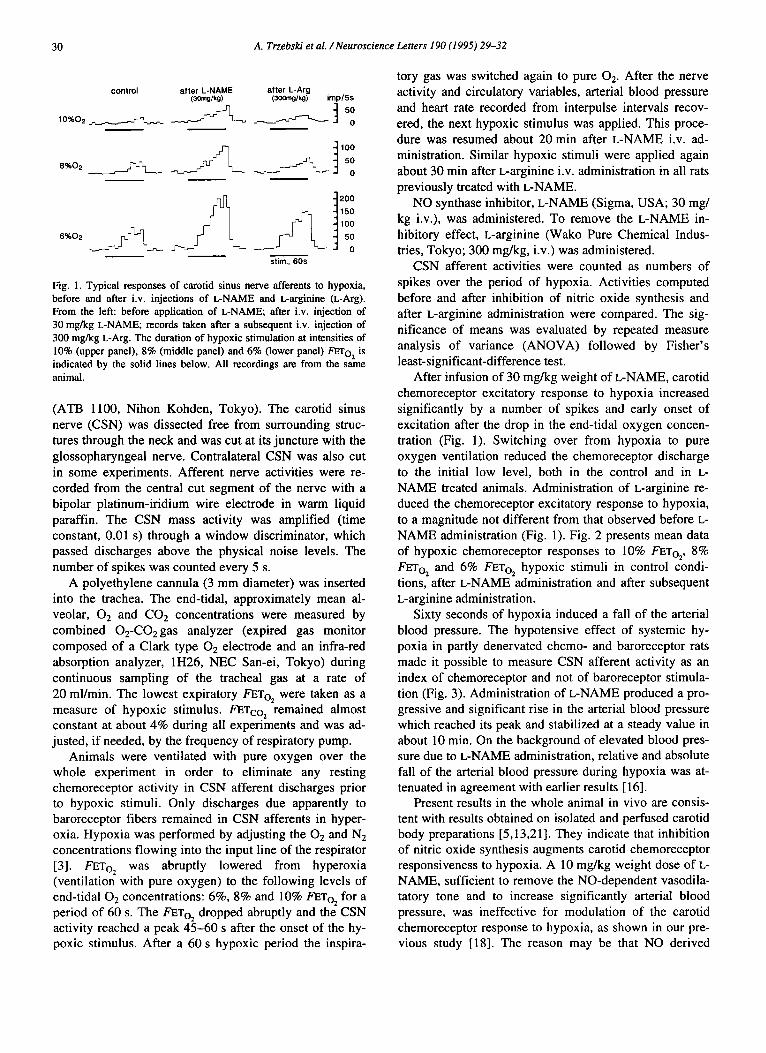
Fig. 1. Typical responses of carotid sinus nerve afferents to hypoxia, before and after i.v. injections of L-NAME and L-arginine (L-Arg). From the left: before application of L-NAME; after i.v. injection of 30 mg/kg L-NAME; records taken after a subsequent i.v. injection of 300 mg/kg L-Arg. The duration of hypoxic stimulation at intensities of 10% (upper panel), 8% (middle panel) and 6% (lower panel) FETo2 is indicated by the solid lines below. All recordings are from the same animal.
(ATB 1100, Nihon Kohden, Tokyo). The carotid sinus nerve (CSN) was dissected free from surrounding struc- tures through the neck and was cut at its juncture with the glossopharyngeal nerve. Contralateral CSN was also cut in some experiments. Afferent nerve activities were re- corded from the central cut segment of the nerve with a bipolar platinum-iridium wire electrode in warm liquid paraffin. The CSN mass activity was amplified (time constant, 0.01 s) through a window discriminator, which passed discharges above the physical noise levels. The number of spikes was counted every 5 s.
A polyethylene cannula (3 mm diameter) was inserted into the trachea. The end-tidal, approximately mean al- veolar, 02 and CO2 concentrations were measured by combined O2-CO2gas analyzer (expired gas monitor composed of a Clark type 02 electrode and an infra-red absorption analyzer, 1H26, NEC San-ei, Tokyo) during continuous sampling of the tracheal gas at a rate of 20 ml/min. The lowest expiratory FETo2 were taken as a measure of hypoxic stimulus. FETco 2 remained almost constant at about 4% during all experiments and was ad- justed, if needed, by the frequency of respiratory pump.
Animals were ventilated with pure oxygen over the whole experiment in order to eliminate any resting chemoreceptor activity in CSN afferent discharges prior to hypoxic stimuli. Only discharges due apparently to baroreceptor fibers remained in CSN afferents in hyper- oxia. Hypoxia was performed by adjusting the 02 and N2 concentrations flowing into the input line of the respirator [3]. FETo2 was abruptly lowered from hyperoxia (ventilation with pure oxygen) to the following levels of end-tidal 02 concentrations: 6%, 8% and 10% FETo2 for a period of 60 s. The FETo2 dropped abruptly and the CSN activity reached a peak 45-60 s after the onset of the hy- poxic stimulus. After a 60 s hypoxic period the inspira-
tory gas was switched again to pure 02. After the nerve activity and circulatory variables, arterial blood pressure and heart rate recorded from interpulse intervals recov- ered, the next hypoxic stimulus was applied. This proce- dure was resumed about 20 rain after L-NAME i.v. ad- ministration. Similar hypoxic stimuli were applied again about 30 min after L-arginine i.v. administration in all rats previously treated with L-NAME.
NO synthase inhibitor, L-NAME (Sigma, USA; 30 mg/ kg i.v.), was administered. To remove the L-NAME in- hibitory effect, L-arginine (Wako Pure Chemical Indus- tries, Tokyo; 300 mg/kg, i.v.) was administered.
CSN afferent activities were counted as numbers of spikes over the period of hypoxia. Activities computed before and after inhibition of nitric oxide synthesis and after L-arginine administration were compared. The sig- nificance of means was evaluated by repeated measure analysis of variance (ANOVA) followed by Fisher's least-significant-difference test.
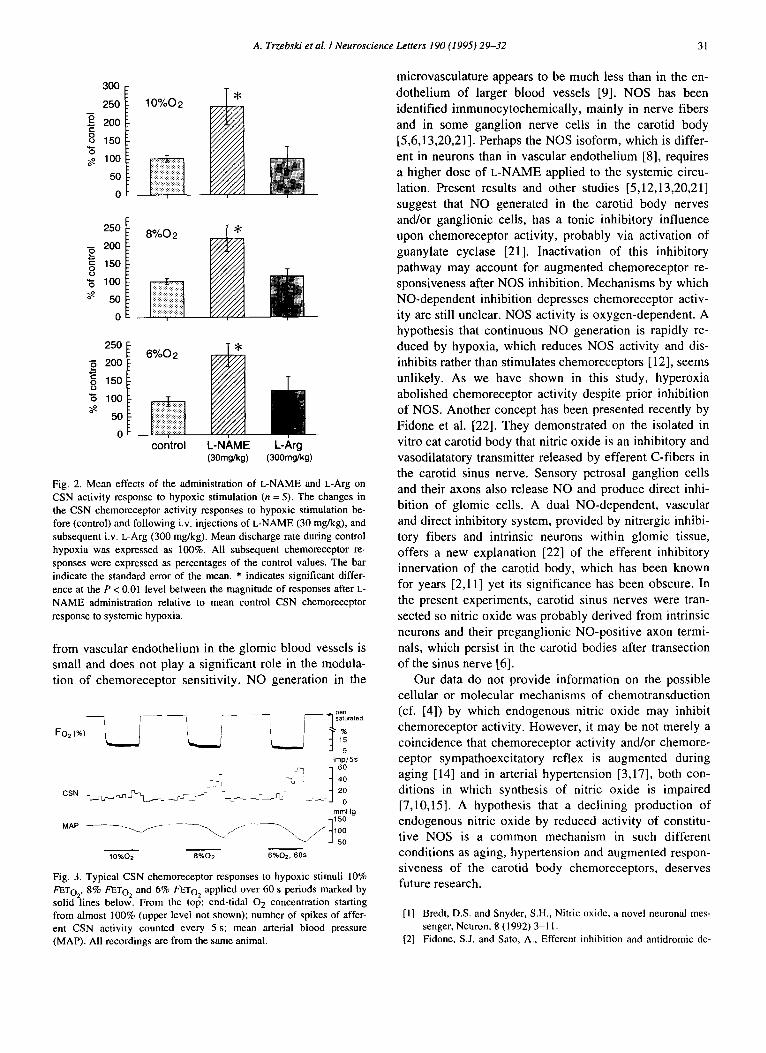
After infusion of 30 mg/kg weight of L-NAME, carotid chemoreceptor excitatory response to hypoxia increased significantly by a number of spikes and early onset of excitation after the drop in the end-tidal oxygen concen- tration (Fig. 1). Switching over from hypoxia to pure oxygen ventilation reduced the chemoreceptor discharge to the initial low level, both in the control and in L- NAME treated animals. Administration of L-arginine re- duced the chemoreceptor excitatory response to hypoxia, to a magnitude not different from that observed before L- NAME administration (Fig. 1). Fig. 2 presents mean data of hypoxic chemoreceptor responses to I0% FETo2, 8% FETo2 and 6% FETo2 hypoxic stimuli in control condi- tions, after L-NAME administration and after subsequent L-arginine administration.
Sixty seconds of hypoxia induced a fall of the arterial blood pressure. The hypotensive effect of systemic hy- poxia in partly denervated chemo- and baroreceptor rats made it possible to measure CSN afferent activity as an index of chemoreceptor and not of baroreceptor stimula- tion (Fig. 3). Administration of L-NAME produced a pro- gressive and significant rise in the arterial blood pressure which reached its peak and stabilized at a steady value in about 10 min. On the background of elevated blood pres- sure due to L-NAME administration, relative and absolute fall of the arterial blood pressure during hypoxia was at- tenuated in agreement with earlier results [16].
Present results in the whole animal in vivo are consis- tent with results obtained on isolated and perfused carotid body preparations [5,13,21]. They indicate that inhibition of nitric oxide synthesis augments carotid chemoreceptor responsiveness to hypoxia. A 10 mg/kg weight dose of L- NAME, sufficient to remove the NO-dependent vasodila- tatory tone and to increase significantly arterial blood pressure, was ineffective for modulation of the carotid chemoreceptor response to hypoxia, as shown in our pre- vious study [18]. The reason may be that NO derived
A. Trzebski et al. / Neuroscience Letters 190 (1995) 29-32 31
300 250
"~ 200 t- O 150 "5
lOO 50
10%O2
i i 250
-- 200 = 150 8 "5 100 o~ 50
8 % 0 2 >~
L i
250 200
~, 15o o "5 100 o~ 50
6%O~, 2
control L-NAME (30mg/kg)
L-/~,rg (300mg/kg)
Fig. 2. Mean effects of the administration of L-NAME and L-Arg on CSN activity response to hypoxic stimulation (n = 5). The changes in the CSN chemoreceptor activity responses to hypoxic stimulation be- fore (control) and following i.v. injections of L-NAME (30 mg/kg), and subsequent i.v. L-Arg (300 mg/kg). Mean discharge rate during control hypoxia was expressed as 100%. All subsequent chemoreceptor re- sponses were expressed as percentages of the control values. The bar indicate the standard error of the mean. * indicates significant differ- ence at the P < 0.01 level between the magnitude of responses after L- NAME administration relative to mean control CSN chemoreceptor response to systemic hypoxia.
from vascular endothel ium in the glomic blood vessels is small and does not play a significant role in the modula- tion of chemoreceptor sensitivity. NO generation in the
p e n / j - - - - - F saturated Fo2,., /I } 1}
imp/5s
~m I 60 y r-~ L 40
CSN F L ~ ~ . ~ , ~ _ - ~ ' ~ L ~ ~ ___ - ~ 20 0
mmHg _ _ _ _ -115o
MAP ~ ~ ~- ~ ~1100
~ ' J 50
10%O2 8%02 6%02, 60S
Fig. 3. Typical CSN chemoreceptor responses to hypoxic stimuli 10% FETo2, 8% FETo2 and 6% FETo2 applied over 60 s periods marked by solid lines below. From the top: end-tidal 02 concentration starting from almost 100% (upper level not shown); number of spikes of affer- ent CSN activity counted every 5 s; mean arterial blood pressure (MAP). All recordings are from the same animal.
microvasculature appears to be much less than in the en- dothelium of larger blood vessels [9]. NOS has been identified immunocytochemical ly , mainly in nerve fibers and in some ganglion nerve cells in the carotid body [5,6,13,20,21]. Perhaps the NOS isoform, which is differ- ent in neurons than in vascular endothelium [8], requires a higher dose of L-NAME applied to the systemic circu- lation. Present results and other studies [5,12,13,20,21] suggest that NO generated in the carotid body nerves and/or ganglionic cells, has a tonic inhibitory influence upon chemoreceptor activity, probably via activation of guanylate cyclase [21]. Inactivation of this inhibitory pathway may account for augmented chemoreceptor re- sponsiveness after NOS inhibition. Mechanisms by which NO-dependent inhibition depresses chemoreceptor activ- ity are still unclear. NOS activity is oxygen-dependent . A hypothesis that continuous NO generation is rapidly re- duced by hypoxia, which reduces NOS activity and dis- inhibits rather than stimulates chemoreceptors [ 12], seems unlikely. As we have shown in this study, hyperoxia abolished chemoreceptor activity despite prior inhibition of NOS. Another concept has been presented recently by Fidone et al. [22]. They demonstrated on the isolated in vitro cat carotid body that nitric oxide is an inhibitory and vasodilatatory transmitter released by efferent C-fibers in the carotid sinus nerve. Sensory petrosal ganglion ceils and their axons also release NO and produce direct inhi- bition of glomic cells. A dual NO-dependent , vascular and direct inhibitory system, provided by nitrergic inhibi- tory fibers and intrinsic neurons within glomic tissue, offers a new explanation [22] of the efferent inhibitory innervation of the carotid body, which has been known for years [2,11] yet its significance has been obscure. In the present experiments, carotid sinus nerves were tran- sected so nitric oxide was probably derived from intrinsic neurons and their preganglionic NO-posi t ive axon termi- nals, which persist in the carotid bodies after transection of the sinus nerve [6].
Our data do not provide information on the possible cellular or molecular mechanisms of chemotransduction (cf. [4]) by which endogenous nitric oxide may inhibit chemoreceptor activity. However, it may be not merely a coincidence that chemoreceptor activity and/or chemore- ceptor sympathoexcitatory reflex is augmented during aging [14] and in arterial hypertension [3,17], both con- ditions in which synthesis of nitric oxide is impaired [7,10,15]. A hypothesis that a declining production of endogenous nitric oxide by reduced activity of constitu- tive NOS is a common mechanism in such different conditions as aging, hypertension and augmented respon- siveness of the carotid body chemoreceptors, deserves future research.
[1] Bredt, D.S. and Snyder, S.H., Nitric oxide, a novel neuronal mes- senger, Neuron, 8 (1992) 3-11.
[2] Fidone, S.J. and Sato, A., Efferent inhibition and antidromic de-

32 A. Trzebski et al. / Neuroscience Letters 190 (1995) 29-32
pression of chemoreceptor A-fibers from the cat carotid body, Brain Res., 22 (1970) 181-193.
[3] Fukuda, Y., Sato, A. and Trzebski, A., Carotid chemoreceptor discharge responses to hypoxia and hypercapnia in normotensive and spontaneously hypertensive rats, J. Auton. Nerv. Syst., 19 (1987) 1-I 1.
[4] Gonzalez, C., Almaraz, L., Obeso, A. and Rigual, R., Carotid body chemoreceptors: from natural stimuli to sensory discharges, Physiol. Rev., 74 (1994) 829-898.
[5] Grimes, P.A., Lahiri, S., Stone, R., Mokashi, A. and Chug, D., Nitric oxide synthase occurs in neurons and nerve fibers of the ca- rotid body. In R.G. O'Regan, P. Nolan, D.S. McQueen and D.J. Paterson (Eds.), Arterial Chemoreceptors - Cell to System, Ad- vances in Experimental Medicine and Biology, Vol. 360, Plenum Press, New York, 1994, pp. 221-224.
[6] Hrhler, B., Mayer, B. and Kummer, W., Nitric oxide synthase in the rat carotid body and carotid sinus, Cell Tissue Res., 276 (1994) 559-564.
[7] Hongo, K., Nakagomi, T., Kassell, N.F., Sasaki, T., Lehman, M., Vollmer, D.G., Tsukahara, T., Ogawa, H. and Torner, J., Effects of aging and hypertension on endothelium-dependent vascular relaxation in rat carotid artery, Stroke, 19 (1988) 892-897.
[8] Janssens, S.P., Shimouchi, A., Quertermous, T., Bloch, D.B. and Bloch, K.D., Cloning and expression of a cDNA encoding human endothelium-derived relaxing factor/nitric oxide synthase, J. Biol. Chem., 267 (1992) 14519-14522.
[9] Katusic, Z.S. and Cosentino, F., Nitric oxide synthase: from mo- lecular biology to cerebrovascular physiology, News Physiol. Sci., 9 (1994) 64-67.
[10] Moritoki, H., Yoshikawa, T., Hisayama, T. and Takeuchi, S., Possible mechanisms of age-associated reduction of vascular re- laxation caused by atrial natriuretic peptide, Eur. J. Pharmacol., 210 (1992) 61-68.
[11] Neil, E. and O'Regan, R.G., The effects of electrical stimulation of the distal end of the cut sinus and aortic nerves on peripheral arterial chemoreceptor activity in the cat, J. Physiol., 215 (1971) 15-32.
[12] Prabhakar, N.R., Neurotransmitters in the carotid body. In R.G. O'Regan, P. Nolan, D.S. McQueen and D.J. Pate~on (Eds.), Ar-
terial Chemoreceptors - Cell to System, Advances in Experimen- tal Medicine and Biology, Vol. 360, Plenum Press, New York, 1994, pp. 57-69.
[13] Prabhakar, N.R., Kumar, G.K., Chang, C.H., Agani, F.H. and Haxhiu, M.A., Nitric oxide in the sensory function of the carotid body, Brain Res., 625 (1993) 16-22.
[14] Sato, A., Sato, Y., Suzuki, H. and Trzebski, A., Reflex responses in adrenal sympathetic nerves to stimulation of glucoreceptors and chemoreceptors in aging rats, J. Auton. Nerv. Syst., 32 (1991) 63-68.
[15] Sato, I., Morita, I., Kaji, K., Ikeda, M., Nagao, M. and Murota, S., Reduction of nitric oxide producing activity associated with in vitro aging in cultured human umbilical vein endothelial cells, Biochem. Biophys. Res. Commun., 195 (1993) I070-1076.
[16] Sun, M.-K. and Reis, D.J., Evidence nitric oxide mediates the vasodepressor response to hypoxia in sino-denervated rats, Life Sci., 50 (1992) 555-565.
[17] Trzebski, A., Arterial chemoreceptor reflex and hypertension, Hypertension, 19 (1992) 562-566.
[18] Trzebski, A., Sato, A., Sato, Y. and Suzuki, A., Carotid chemore- ceptor activity and heart rate responsiveness to hypoxia after in- hibition of nitric oxide synthase. In R.G. O'Regan, P. Nolan, D.S. McQueen and D.J. Paterson (Eds.), Arterial Chemoreceptors - Cell to System, Advances in Experimental Medicine and Biology, Vol. 360, Plenum Press, New York, 1994, pp. 285-288.
[19] Wang, W.-J., He, L., Chen, J., Dinger, B. and Fidone, S., Mecha- nisms underlying chemoreceptor inhibition induced by atrial natriuretic peptide in rabbit carotid body, J. Physiol., 460 (1993) 427-441.
[20] Wang, Z.-Z., Bredt, D.S., Fidone, S.J. and Stensaas, L.J., Neurons synthesizing nitric oxide innervate the mammalian carotid body, J. Comp. Neurol., 336 (1993) 419-432.
[21] Wang, Z.-Z., Stensaas, L.J., Bredt, D.S., Dinger, B. and Fidone, S.J., Localization and actions of nitric oxide in the cat carotid body, Neuroscience, 60 (1994) 275-286.
[22] Wang, Z.-Z., Stensaas, L.J., Dinger, B.G. and Fidone, S.J., Nitric oxide mediates chemoreceptor inhibition in the cat carotid body, Neuroscience, 65 (1995) 217-229.





























