Embed Size (px)
Citation preview

Virginia Commonwealth University Virginia Commonwealth University
VCU Scholars Compass VCU Scholars Compass
Theses and Dissertations Graduate School
1969
Inhibition of Cell Division, Protein Synthesis and Nucleic Acid Inhibition of Cell Division, Protein Synthesis and Nucleic Acid
Synthesis in Escherichia coli W by Tetracycline Antibiotics Synthesis in Escherichia coli W by Tetracycline Antibiotics
George Henry Miller
Follow this and additional works at: https://scholarscompass.vcu.edu/etd
Part of the Pharmacy and Pharmaceutical Sciences Commons
© The Author
Downloaded from Downloaded from https://scholarscompass.vcu.edu/etd/5216
This Dissertation is brought to you for free and open access by the Graduate School at VCU Scholars Compass. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected].

Inhibition of Cell D iv i s ion , Prote in Synthes i s
and Nuc leic Ac id Synthes i s in
Escherichia coli W by
Tetracyc l ine Antibiotics
by
George Henry Mi l ler
B . Sc . , Philadelphia College of P harmacy and Science , 196 0
M . Sc . , Phi ladelphia Col lege of Pharmacy and Science , 196 2
Degree of
The s i s
Submitted i n partial ful f i l lment of
the requirements for the
Doc tor of Phi losophy in the Depar tment of
Pharmacy at the Med ical Col lege of Virginia
Richmond , Virg inia
June , 1969
100561

ii
This thesis by Mr . George H. Mil ler is accepted in its
present f orm as satisfying the thesis requirement for the
degree of Doctor of Philosophy
Date : Approved :
. {fle:t. -�' �- . I?. t. J: ... .
. . o.J. .. .It' . . (! ?. � ...... . Dr . Arthur A . Cammarata
Dr . Edward S . Kline
... -� . . �� J.<i 6. r. .... .
.. _(!. 4. . t � . J.? � r: .... . � D . Punch
.Dr . ;� Richard
APPROVED :

iii
Curriculum Vitae

iv

v
Acknowledgments
The author wishes to expre s s h i s apprec i ation and h i s s incere
gratitude to :
Dr . A . N . Martin for providing the opportun ity to study
and learn in a stimulating and plea sant environment and for
h i s counseling , superv i s ion , patience and espec ially h i s
expre ss ions o f confidence during the course of thi s work ;
Dr . A . A . Cammarata and Dr . F . Perade j ordi for the many
stimulating and helpful d i s cus s ion s ;
Dr . S. A . Khal il for h i s help and encouragement durina
the development o f the exper imental procedures;
Luisa C . Mmller for her excellent technical a s s i stance
in performing these experimental procedure s ;
Sung Ho Hahn for h i s a s s i stance in preparing the materials
for the experiments ;
John Andrako Jr . , Willi am Crouthamel , John Gill , Douglas
Martin and Teresa Taylor for the i r help i n the v iable cell
determinations as well as other experiments ;
the members o f my Adv i sory Committee , Dr . E. Kline ,
Dr . Punch and Dr . Richard in addition to Dr . Cammarata and
Dr . Martin for the i r advice and especiali1 for the i r
reading of thi s the s i s ;
the staf f o f the Computer Center , Department o f B i ometry ,
Medical College o f Virginia for the prompt stat i stical
analy s i s o f the experimental data and for the i r a s s i stance
in studying the proposed model of tetracycline act ion;
my parents , Arthur and P r i sc illa , for the i r encouragement

Acknowledgments
and sacr i fices in my behal f ;
my daughter , Claudia , for the inspiration and joy she
provided during this work ;
vi
my wi fe , L i lo , for providing the encouragement to
undertake thi s s tudy and for her patience , understanding and
sacri fices during its complet ion in addi tion to her excel lent
technical a s s i s tance acknowledged above .

TABLE OF CONTENTS
I, I ntroduction
I I . Literature Survey
A. Tetracycline s - Nomenclature , General P roperties
and B iological Activity
A,l, A-Rmng Modi fications
A.2. B-Ring Modi fications
A . 3 . C-Ring Modif ications
A,4, D-Ring Modi fications
A . S . Structure-Activity General i z ations
A,6, Acidity Constants o f the Tetracyclines
B . Tetracycl ine s - Biochemical B a s i s of The ir
Antibiotic Activity
v i i
Page
3
3
s
6
6
7
7
8
1 0
B . l . E f fects o n Rate o f Cell Reproduction 10
B . 2. E f fects on Oxidation and Respirat ion 1 2
B . 3, Ef fects on Protein Synthe s i s in Culture s 13
B . 4 . E f fects on Cell-Free Nitro-Reductase Systems 16
B . S . The Nature o f Cell-Free Protein Synthe s i z ing
Systems- Translation of the Genetic Code
B . S . a . Ribosome s
B . S.b . Mes senger Ribonucle ic Acid
B . S . c. Trans fer Ribonucle ic Ac id
B . S . d . A Model of Cell-Free Prote in Synthe s i s
2 3
24
2 9
31
33
S�ep 1 . - Binding of a 30 S Subunit to mRNA 40
Step 2 . - Formation of the I nitiation Complex 40
Step 3 . - Attachment of the SO S Subunit to

the Initiation Complex
S tep 4.- Bind ing to the Aminoacyl S ite on
the 70 s Ribosome
S tep 5,- Format ion o f the Peptide Bond
S tep 6.- Pul sation ( Movement ) of the
Ribosome
S tep 8 . - Chain Termination and Release
B.6. The E f fects of Tetracycl ines on C e l l - Free
Protein Synthe s i z ing Systems
B.6.a. E f fects of Tetracycl ines in Terms of the
Mode l o f Cel l-Free Protein Synthe sis
Inte rference with S tep I.-
I nterference with S tep 2 . -
I nter ference with Step 3 .-
Interference with Step 4 . -
Interference with S tep 5 . -
I nter ference with S tep 6 . -
I nterference with S tep 8.-
B . 6.b . Binding o f Tetracycl ines to Ribosomes
I I I. Experimental
A . Materials and Equipment
A . l. Tetracy c line Derivatives
A. 2 . Escherichia c o l i
A . 3 . Culture Broths
A . 4. Culture Turbid ity and Colorime try
A . 5. Total C e l l Counts
B . Hethods
viii
Page
4 6
5 0
5 3
5 4
5 4
5 6
5 8
5 9
6 0
6 1
6 2
6 3
6 3
6 3
6 6
6 6
6 6
6 9
7 0
7 0

B . l. E s che richia col i Growth
B . 2. Viable Count Method
B.3 . Total Count Method
B . 4. Nuc leic Acid and Protein Determinations
Nuc leic Acid Determinations
Prote in Determinations
I V . Results and Discuss ion
A . E f fect o f Tetracyc l ines on Rate Constants for
C e l l Div i s ion
B . E f fect o f Tetracyc l ines on Rate Cons tants for
Protein and Nuc l e i c Acid Synthe s i s
c . T ime o f Onset o f Tetracyc l ine I nhibition
D . E f fect o f .Broth pH Change s on Te tracyc l ine
I nhibition
E. Kinetic Mode l s o f the I nhibi tion of Bacterial
Culture s by Tetracy c l ines
V . Summary
VI . B ib l iography
ix
Page
7 1
7 1
7 2
7 2
7 3
7 6
7 9
9 2
1 0 5
111
1 1 9
1 3 6
1 4 0

AA
DPN
DPNH
EDTA
f l , f 2
FMN
FAD
G
ked
kv
L i s t of Abbreviations
ami no acids
ala a lanine
a s p asparagine
cys cysteine
leu leuc i ne
lys lysine
met methionine
phe phenyla lanine
ser ser ine
thr threonine
tyr tyros ine
va l val ine
d iphosphopyr idine nuc leotide
reduced d iphosphopyrid ine nuc leotide
ethylenediaminetetraacetic a c id
X
r ibosoma l proteins which are neces sary for prote in
synthe s i s and which are removed from r ibosome s by
wa shing with NH4C l
f lavin mononuc leotide
flavin adenine d i nucleotide
a supernatant enzyme which cata lyzes the hydrolysis
of guanos i ne - 5 ' - tr iphosphate in the pre sence of
r ibosome s
generat ion rate constant for c e l l d iv i s ion
generation rate constant for cell d iv i s ion a s
determined b y the v iable cell count method

kt
kp
kna
LDH
MDH
na
List of Abbreviations
generation rate constant for c e l l div i s ion as
determined by the tota l c e l l count method
generation rate constant for protein synthes i s
generation rate constant for the synthe s i s o f
nuc leic a c id
xi
the proportiona l i ty constant r elating generation
rate constants to tetracyc l i ne concentration; the
subscripts v , t , p , na i nd icate the type of
generation constant used , the subscript avg
ind icates that a l l g eneration rate constants were
u sed col lectively
lactic ac id dehydrogenas e
mal ic a c i d dehydrog ena se
nuc leic acids
DNA
RNA
mRNA
rRNA
sRNA
tRNA
AAtRNA
met- tRNAf
N-F-met-tRNAf
deoxyr ibonuc leic acid
r i bonuc leic a c id
me s senger r ibonuc leic acid
r ibosoma l r i bonuc l e ic a c id
soluble r i bonuc leic a c id
�ransfer r i bonuuleic acid
aminoacyl transfer r ibonuc leic acid
methionyl transfer r ibonuc l e ic acid
which a llows the addi tion of a
formyl group to the methionyl amine
N-formylmethionyl transfer
r ibonuc leic acid

L
L i st of Abbreviations
met- tRNAm
ATP
CTP
GOP
GMP
GTP
UTP
poly A
poly c
poly G
poly u
methionyl transfer r ibonuc leic
a c id which does not a l low the
addi tion of a formyl group to
the methionyl amine
adenos i ne- 5 ' -tr iphosphate
cyto s ine- 5 ' -triphosphate
guano s i ne- 5 ' -d iphosphate
guanos ine - 5 ' -monophosphate
guanos i ne - 5 ' - tr iphosphate
ur idine - 5'-tr iphosphate
polyadenyl i c acid
polycytidyl i c acid
polyguanyl i c a c id
polyur idylic ac id
x i i
" Nuc leic Acids" the absorbance of an orc inol reaction ( 4 ml . )
Pm
" Proteins "
T
which i s due to the nuc leic acids of E . coli
in a 1 2 . 5 ml . culture sample
puromyc in
the absorbance of a Fol in-Lowry reac tion
( 6 . 5 ml . ) which i s due to the protein of
E . c o l i in a 2 5 ml . culture sample
supernatant prote ins which are nec e s sary for
protein synthe s i s

TCA
e
L i s t of Abbreviations
trichloroacetic a c id
the frac tion of total c e l l s in a culture which
are found to be viable
x i i i

F igure I .
F igure I I .
F igure I I I .
F igure IV .
F igure V .
F igure VI .
F igure V I I .
F igure V I I I .
F igure I X .
F igure X .
F igure XI .
F igure XI I .
F igure X I I I .
L i s t of F igures
The Tetracyc l i ne s
the Ioni zable Groups of Tetracyc l i ne
Ribosome s . Components , Subunits ,
x iv
Page
4
9
Associati on and D i s sociation Reac tions 2 5
The Formation o f Ami noacyl Transfer
Ribonuc leic Ac id
Schema tic D iagram o f the Proposed C lover
leaf Structure of Transfer Ribonuc leic
Acids
Codon - Anticodon Interaction
A Mode l of C e l l-Free Protein Synthes i s
Ca l ibrat ion Curves for "Nuc l e ic Ac id s "
and " Protei n " Determina t ions
An Exam�le of Viable Cell Genera tion Rate
Curves
An Example o f Tota l Cell Generation Rate
Curves
Examples of the D i f f erence Between Tota l
3 2
3 4
3 5
3 6
7 5
8 0
8 1
and Viable C e l l Generation Rate Curves 9 0
The L inear Dependence o f Generation Rate
Constants , Kv , kp and kna , upon Tetracy-
c l ine Concentration
The Linear Dependence of Generation Rate
Constants , kv , kt , kp and kna , upon
9 3
9-N i tro-dm-do- tetracyc l i ne Concentration 9 4

F igure XIV .
F igure XV .
F igure XVI .
F igure XVI I .
F igure XVI I I .
F igure XIX .
F igure XX .
F igure XXI .
F igure XXI I .
L i s t of F igure s
The Linear Dependence of Generation
Rate Constants , kv , kt , kp and kna '
upon Concentrat ion of 9-D imethylamino
dm-do-tetracyc l i ne
The L inear Dependence o f Generation
Rate Constants , kv , kt , kp and kna '
upon Concentration of Several
Tetracyc l ines
An Example of Generati on Rate Curves
for Protei n Synthes i s
An Example o f Generation Rate Curves
for Protei n Synthe s i s
An Example of Genera tion Rate Curves
for Nuc leic Ac id Synthe s i s
An Example of Genera t ion Rate Curves
for Nuc leic Ac id Synthe s i s
T ime o f Onset of and Recovery from
Oxytetracyc l ine Inhibition in Peptone
Broth Culture s
T ime of Onset of Oxytetracyc l i ne
Inhibition of Cultures in Dextrose
Salts Broth
The L inear Dependence of Generation
Ra te Constants upon Concentration of
Zwitter ionic Chlor tetracyc l i ne
XV
Page
9 5
9 6
9 9
1 0 0
1 0 2
1 0 3
1 0 8
1 0 9
1 1 3

L i s t of F igure s
F igure XXI I I . The Effec t of Broth pH on T ime of
Onset of Oxytetracyc l ine Inhibition
F igure XXIV .
F igure XXV .
F igure XXVI .
Propo s ed Mode l of Tetracycline
Inhib i tion of Protein Synthe s i s in
Bac teri a l Cul ture s
A n Unsealed Analog C omputer Program for
the So lution of the D i f ferenti a l
Equations of the Proposed Model o f
Tetracyc l ine Inhibition of Prote i n
Synthes i s i n Bac teri a l Cultur e s
The Continuous Sy stems Mode l i ng Package
Program Used in the Search for Solutions
to the D i f ferential Equations from the
Proposed Mode l of Tetracyc l i ne Inhibition
xv i
Page
1 1 6
1 2 3
1 2 7
o f Pro�e in Synthe s i s in Bac ter i a l Culture s 1 2 9

Table I .
Table I I .
Table I I I .
Table IV .
Table V .
Table VI .
L i s t of Tables
The Tetracyc l ine s
First Order Generation Rate Constants ,
-1 4 sec . x 1 0 , for E . coli W Culture s
Inhibi ted by Var ious Tetracyc l i ne
Antibiotic s , M x 1 0 6
F i r s t Order Generation Rate Cons tants ,
sec . - 1 x 1 0 4 , for E . c o l i W Cultures
Inhibited by Var ious Tetracyc l i ne
Antibiot ic s , � x 1 0 6 , and the Frac tion
of Viable to Tota l Cel l s Present at
the Start , e1 , and at the End , e5 , of
the Exper iments
I nhibitory Rate Constant s , L/M- sec . ,
for Inhibition of E . c o l i W by
Var ious Tetracyc l ine s
F ir s t Order Generat ion Rate C onstants ,
sec . - 1 x 1 0 4 , and Inhibitory Ra te
Con s tants , L/M-sec . , for E . c o l i W in
Var ious Broth Culture s Inhibited by
Setiera l Tetracyc l ine s
The Differential Equat ions from a Model
of Tetracyc l i ne Inhibition o f Prote in
Synthes i s in Bacter ial Cultures
xvi i
Page
6 7
8 2
8 5
9 7
1 1 4
1 2 6

L i s t of Tables
Table V I I . C omp i la tion of Condi t ions Which Were
Employed to Test the Model of
Tetracyc l ine Inhibition of Prote in
Synthes i s i n Bac ter i a l Cultures
xvi i i
Page
1 3 0

I . INTRODUCTION
Tetracyc lines , or igina l ly isolated i n the late 1 9 4 0's
and early 1 9 5 0 ' s from the myce l ium of S treptomyces , have
achieved maj or importance as therapeutic and prophylactic
agents agai ns t a wide range of i nfections i n human and
veter i nary med i c ine . They have a l s o achieved great
importance in agriculture where they are widely used to
promote weight gain in l ivestock . Commerc i a l production ,
duri ng 1 9 5 8 i n the United States a lone was 1 2 0 tons with a
value of approximately $ 1 m i l lion per ton .
The c l inical importance of these compounds has
s t imulated e fforts to def i ne thei r mode of action a s
inhibitor s of bacteri a l reproduction . Antibiotics having
a s many func t i onal groups a s the tetracyclines may have many
modes of action . The problem faci ng the re search worker i s
t o determine the relative contribution of each mode o f action
in a g iven b i ological system a s a function of antibiotic
concentration . I f a l l parts of a biological system are
exposed to an equiva lent antibiotic concentration , one might
expect that the degree of i nhibition of the vari ous
sub-component systems would be propor tional to the relative
stabi lities of the reaction products of the antibiotic and
the sub-component system . The mos t critical reaction from
the s tandpoint of c e l lular reproduct i on would then represent
the point of the pr imary b i ochemic a l les ion or the s i te of

2
action . The type of reaction would be the mode of ac tion for
the biolog ical sys tem . For a g iven biological system i t i s
probable that there i s one c r i tical reaction which i s most
sensitive to the antibiotic , but thi s reac tion may not be the
same for every biolog ical sys tem ( see Sne l l and Cheng ( l ) for
an exce l lent d i scuss ion of the d i f f i culties of def ining a mode
of tetracyc l ine action) . Many investigator s presently feel
that the pr imary b i ochemical l e s ion inf l icted upon susceptible
bac ter ia by tetracyc l ines is a genera l inhibition of protein
synthe s i s ( 2 , 3 , 4 ) . Therefore it was felt that the study of
severa l measures o f inhibi tion , particularly cell reproduc tion
and pro�e in synthe s i s , in the presenc e of several tetracyc l ine s
might be a u seful way o f studying modes of tetracyc l i ne action .
Biological activ i t i e s of a large number of compounds
obtained quantina t ively under identical dond i tions and in a
prec i s e manner are required to estab l i sh structure-activity
relationships . Presently ava i lable activities for tetracyc l ine
antibiotics have been summa r i zed by Barrett ( 5 ) , Boothe ( 6 ) and
Plakunov ( 7 ) . Many of the se activities have been obtained
under cond i tions such that the results par a l l e l c linical
activ i t i e s . Thu s , some compounds may not have achieved
equ i l ibr ium with the test system . Some activities have been
obta i ned for the purpose of s tudyi ng antibiotic r e s i stance
whi l e s t i l l o ther s have been obta ined in widely varying and not
ea s i ly i nterrelatable bio log i ca l te st systems . Consequently ,
quantitative ac tivit�es suitable for s truc ture-activity
relationships have not been reported for most tetracyc l i ne s .

3
The present work was undertaken to obtain activities suitable
for s tructure-activi ty relationships and which are
pertinent to both bacterial reproduction and the proposed
mode of action . The activi ties obtained should inc lude
both clinically active and "inactive " tetracyc l ine antibiotics .
I I . Literature Survey
A . Tetracyc l i ne s - Nomenclature , General Properties and
Biological Activity
The i s olation in 1 9 4 7 o f the broad spectrum antibiotic ,
chlorte tracyc l ine , from Streptomyces aureofac iens was followed
by the dis covery of oxytetracycl ine in 1 950 and tetracycl ine
in 1953 . The c l inical use fulne s s o f the se compounds prompted
the i s olation , synthes i s and characteri z ation of a large number
of derivatives , degradation products and s imilar compounds with
a wide range o f antibiotic properties . The e lucidation of the
s tructure of the tetracycl ine s has been a ma j or achievement in
natural product chemi s try ( 8). The structure o f chlortetracy
cl ine , e s tab l ished on chemi cal grounds and confirmed by X-ray
analy s i s ( 9 ) , i s shown as ( I ) in Fig . I . The ring numbering
and lettering system shown serve as the bas i s for the
nomenc lature of mos t of the tetracycline s . Several analogs
with altered nuc l e i are also shown in Fig . I .

I.
II ][
F igure I . The Tetracyc l ine s
( I ) 7 -Chlorotetracyl ine
( I I ) diD-do-tetracyc l i ne
( I I I ) 5a ( 6 ) -Anhydrotetracyc l ine
( IV ) 7 -Chloro-Sa ( l la ) -dehydrotetracyc line
(V ) I sotetracyc l i ne
4

5
The s tereochemi s try of the compounds i s depicted in ( l )
of F ig . I . The 4 - d imethylamino-group , the 4a- and Sa-hydrogens ,
the 6-methyl - a nd the l 2 a - hydroxy- are a l l in the a lpha
or i entation . Epimer i zation by general ac id-ba se cata lys i s of
the 4 - d imethylamino -group in neu tr a l solutions occur s rapidly
and g ives r i s e to the so-ca l led 4 -epitetracyc l ines ( 1 0 ) .
Synthetic compound s epimer ic a t both the Sa- and 6a- position
have been reported . The Sa-epitetracyc l i ne i s e s sentially
inac tive whi l e the 6 -deoxy- 6-epite tracycl ines are reported to
be only s l ightly d i f f erent from the norma l 6-deoxy- der ivative
( 6} .
A . l . A-Ring Mod if ications
The 2 -carboxamide-group has been mod if ied in severa l ways .
The 2-acety l - 2-decarboxamido-tetracyc l ines have been prepared
biosynthetically and are e s sent i a l ly i nactive as antibac ter ia l s
( S ) . Dehydration of the carboxamide yields the nitr i le , a
modif ication which i s a l so s a id to result in loss of activity
( S ) . Sever a l compound s which are c l inica l ly useful because of
their i ncreased wa ter solubi l ity have been prepared by employing
the Mannich reaction to form a methylene br idge between the
amide nitrogen and various primary amine s . Hydro lys i s back to
the parent compound is probably respons ible for the ac tivity of
these der ivative s ( 6 ) .
The d imethylamino group at carbon 4 can be epimer i z ed ; it
can be quatern i z ed by treatment with methyl iodide or other
agents and it can be replaced by hydrogen . All of these
compounds are r epor ted to have reduced ant ibacter i a l activity .

The 4 -N-ethyl-methyl-amino a s wel l a s the 4 -N-methyl-amino
der ivative have been prepared and are r eported to pos s e s s
biological activ ity .
6
The �2a-hydroxy-group may be e l iminated by reduction with
z i nc and ammonium hydroxide or by catalytic reduc tion of the
l 2 a - formyloxy der ivative . In these antibiotics nhe phenolic
diketone moiety is conjuga ted with the ketone at carbon l .
Such conjugation i s said to be re spons ible for the loss of
activ i ty by this c ompound . The 4a ( l 2 a ) -anhydrotetracyc l ines
have been prepared by pyrolysi s of the l 2a-acetoxy der ivatives
and are r eported to have minimal antibac ter ial activity . The
l 2 a- f ormyloxy- a nd l 2a-acetoxy- der iva t ives are r eported to
pos se s s activ i ty s o l e ly becau s e of their hydr o ly s i s to the
parent compound s .
A . 2 . B-Ring Modi f ications
5 - Hydroxytetracyc l ine wa s the second tetracyc l ine antibiotic
i solated from Streptomyce s and wa s the f ir s t t o have its
structure d e l i neated ( 8 ) . I t s c l inical u t i l i ty ha s been proven
throughout the last 2 0 year s . The Sa ( l l a ) -dehydrotetracyc l i ne s ,
( IV ) i n F ig . I , are a l so produced biosynthe tica l ly and are
g ener a l ly r egarded a s being inactive ( 6 ) .
A . 3 . C-Ring Modif ications
In s trong a c id solutions , tetracyc l ines are dehydrated to
form anhydro te tracyc line s , ( I I I ) in F ig . I , whi l e in a lka l ine
solutions i sotetracyc line s , ( V ) in F ig . I , are formed . Both
modi f ications r e sult in compounds which are tradi ti ona lly
regarded a s inact ive or a s pos s e s si ng an a l tered spec trum of

7
antibacter ial activ i ty ( 5 , 6 ) . The i sotetracyc l i ne nuc leu s has
such d i f f erent conf orma t ions avai lable to it that its lack of
activity should probably not be u t i l i zed i n s truc ture-ac tivity
relationships . The c linically useful 6 -demethyltetracyc l ine s
are produced biosynthetica l ly , whi le the 6-deoxy-6-epitetra
cyc l ines are the result o f catalytic hydrog enation reac tions .
Both type s of compound s a s we l l a s the 6-deoxy- 6 -demethyl
tetracyc l i ne s , ( I I ) i n F ig . I , are potent ant ibac ter ial agents .
In addi tion , the 6 -methylene tetracyc l ines have been prepared
and are a l s o ac tive antibacter ial substanc e s .
A . 4 . D-Ring Modif ications
7 -Chlortetracyc l ine , ( I ) i n F ig . I , and 7 -bromotetracycl i ne
are produced biosynthe t ic a l ly and are among the mo st potent
tetracyc l i ne antibiotic s . The acid stabi l i ty of the
6-d eoxy - 6 -demethyl-te tracyc l ines ha s a l lowed the chemical
substitution of the aromatic D-r ing at positions 7 and 9 and
a number of antibac ter i a l substances with a wide range of
activ i ties have been reported . The ava i labi l i ty of the se
der ivatives made possible our present attempt at quantitative
correlations of struc ture and activ i ty .
A . S . Struc ture-Ac tiv i ty Generali zations
Modif ications of the A-r ing which are not readily rever s ible
by s imple aqueous hydrolys i s lead to drastic reduc tions in
biolog ical activ i ty . The mos t notable exceptions are the
4 -dedimethylamino-tetracyc l ine s which retain about 5 0 % of the
biolog ic a l activ i ty of the ir parent compound . In contras t , the
4 - epitetracyc l ines are e s sentially inac tive . P lakunov ( 7 )

8
s tudied s everal tetracycl ines with altered A-r ing systems a s
well a s several compound s s truc turally analogous t o the A-r ing
of tetracycl ine and concluded that thi s r ing i s the most
important part of the tetracycl ine nucleu s . He f ound that
alterations of it may produce compounds with a d i f f erent
antibac ter ial spec trum which are probably exerting an effec t
by a dif ferent mode of ac tion (v itamin K antagon i sm i s proposed ) .
Modif ications of the substituents at carbons 5 , 6 , 7 and
9 produce der ivatives with quantitatively d i f f erent activities
but do not produce the extreme d i f f erences in activ i ty seen
with A-r ing modif ications . These compound s would seem to offer
the best pos s ibil i ty for structure-act ivity correlations and
Colai z z i , Knevel and Martin ( ll ) have repor ted l imi ted
correlations of biolog ical activity with Hammett- s igma value s
for these der ivatives .
The oxygen func t i ons at pos i t ions 1 0 , ll and 1 2 appear to
be important for biolog ical activ i ty s i nc e alterations of thi s
chromophoric group lead t o very inac tive compounds . The se
grouping s are also one of the mos t l i kely s i te s for metal ion
chelation , a phenomena which ha s always been expec ted to play
a large role mn the activ i ty of the se antibiotic s ( 1 2 ) .
A . 6 . Ac idity Constants of the Tetracycl ines
The normal tetracycl ine nucleus , ( I ) and ( I I ) in F ig . I,
ha s three mea surable pKa value s i n the vic inity of 3 , 7 . 5
and 9 , Table I , see page 61j Stephens et . al . , ( 1 3 ) have
def ined three ioni zable groups , (A ) , ( B ) and ( C ) correspond i ng
to these values a s ind icated in F ig . I I . Thi s a s s ignment o f

8 H@
CH3 OH N(CH3)2
OH
CONH2
OH OH A c
F igure I I. The Ioni zable Groups of Tetracyc l ine
(A ) Tricarbonylmethane
( B ) Ammonium Cation
(C ) Phenolic diketone
9

1 0
the observed pKa values t o spec i f ic functions ha s been the
subject of some controversy ( 1 4 , 1 5 ) , but it is now genera lly
agreed that the f ir s t pKa i s attr ibutable to the
tr icarbonylmethane sys tem (A) , the second to the phenolic
d iketone moiety ( C ) and the third to the ammonium cation ( B )
I t i s recog n i zed that in many compounds con s iderable over lap
may exi s t between the ioni zation of the second and third groups
( 1 6 ) •
B . Tetracyc lines - Biochemica l Ba s i s of their Antibiotic
Act iv i ty
I t i s g ener a l ly a s sumed that a l l the tetracyc l ines have
s im i lar mode s of ac tion . Thi s view i s ba sed upon the
s imilarity of their s truc ture s , the fact that the bac ter iostatic
effects of the mos t active compound s occur at s imi lar
concentrations and the common occurrence of cross-resi stance .
Never thele s s , i nve s tiga tor s have occas iona l ly noted
d i s s imilar i t ie s , part icular ly in the ni tro-reduc ta se system and
in c e l l wal l synthe s i s . Cross-re s i stance i s seen with a great
many other struc tur a l ly d i s s imi lar antibiotics and it probably
does not have any bearing on the mode of bac ter iostatic action .
Also , there i s a large d i f f erence in the concentrations at which
bac ter iosta tic e f fects are seen in some le ss common tetracyc l i ne s .
The resolution o f these problems , undoubtedly , depend s o n the
test system .
B . l . Effect on Rate of C e l l Reproduc tion
The ant ibac ter i a l activ i ty of the tetracyc l ines ha s been

1 1
evaluated i n various media against many pathogenic and
non-pathogenic organ i sms . The data ha s usually been presented
as the minimum concentration of drug necessary to prevent
v i s ible growth dur i ng a spec i f i ed time per iod . These value s ,
the so-c a l led MIC or p value s , have been tabulated by La skin
( 1 7 ) . The wide range o f MIC va lue s obtained for a g iven
tetracyc l ine by d i f f erent i nvestiga tors is the reason why
these activ i t i e s are not useful in making structure-activity
correlations .
The effec t of the antibiotics on the rate of cell d iv i s ion
ha s been studied by turbidimetr ic , viafule plate and tota l cell
count methods . Rates of growth mea sured by turbid imetr ic
method s f orm the bas i s for the biological activities compi led
by Boothe ( 6 ) , Barrett ( 5 ) , P lakunov ( 7 ) and Benbough and
Morr i s son ( 1 8 ) . These methods were c r i t i c i zed by Ha sh et . a l . ,
( 1 9 ) s i nc e they are sensi tive to c e l l volume in addition to
cell number . They c la im that cell vo lume may change during
tetracyc l i ne inhibition . The Coulter Counter can be used to
observe chang e s i n c e l l volume . C e l l vo lume changes , other
than those norma l ly a s sociated with the exponential growth
cyc l e , however , were not seen in cultures inhibi ted with low
tetracyc l ine concentrations ( 2 0 ) . Jones and Morr i sson ( 2 1 )
have pointed out that the ir turbidimetr ic data were correlated
wi th c e l l weight , which to them is the mos t meani ngful and
only exponential growth parameter . They be l ieve that changes
in cell number s are only exponential becau se of exponential
chang e s in cell mas s and that therefore turbidimetr ic measures

1 2
were more meaning fu l . Data to be pre sented later make it c lear
that measurements of culture turbidity and c e l l numbers may be
d i fferent dur ing normal and inhibited c e l l growth .
C iak a nd Hahn ( 2 2 ) , Jone s and Mor r i s son ( 2 1 ) and Garrett
and Brown ( 2 3 ) d emonstrated that exponenti a l growth i s
ma inta ined in cultures conta ining low tetracyc l ine concentrations .
They propo sed k inetic equa tions which expre s sed the dependence
of the growth rate constants upon tetracyc l ine concentration .
Changes in this dependence at higher antibiotic concentrations
have been interpre ted as ind icating a change in the predominant
mode of action .
B . 2 . Effec ts on Oxidation and Re spiration
Eagle and Saz ( 2 4 ) , Snel l and Cheng ( 1 ) as wel l a s Laskin
(1 7 ) r eviewed the effects of the antibiotics on respiration and
on the oxidation of var ious substrates . The pronounced
inhibition in mo st cultures of the se parameter s was one of the
f ir s t areas s tudi ed . Cheng and Sne l l ( 2 5 ) reported 5 0 %
inhibition o f o0 2 in resting E scher ichia coli and
Staphylococcu s aureus by a s l ittle a s 2 x 1 0 - 6 M
oxytetracyc l ine . At the same concentration severa l
tetracyc l ine analogs : 5a ( lla ) -anhydro-5-hydroxy-tetracycline ,
4 -dedimethylam i no-oxytetracycl ine and 2-acety l - 2 -decarboxamido-
oxytetracyc l ine , compounds not normal ly cons idered
bacter i o static , cau s ed inhibitions of 4 6 , 59 and 2 3 % ,
r e spect ively . Crema and Barol i ( 2 6 ) repor t that oxytetracyc l i ne
was a lmo st without e f f ec t on the r espiration of S . curcio when
the organism wa s in the resting state , whi l e a s l ittle a s

1 3
0 . 1 to 1 . 0 mcg . /ml . o f drug inhibited re spiration by 5 0 to 6 0 %
at the beginning of the log pha s e . This observat ion wa s
conf irmed by Wong , Barban and Aj l ( 2 7 ) who r eport 9 0 %
inhibition o f re spiration i n pro l i f erating cultur e s , by 0 . 1 3
mcg . /ml o f oxytetracycl ine .
A large number o f r eport s documenting inhibition of
oxidat ion of compounds involved in carbohydrate metabo l i sm
have been reviewed ( 1 7 , 2 4 ) . The se effects are now thought
to be due to an inhibition of enzyme f ormation or to the
appearance of an add i t iona l mode of ac tion due to the extremely
high concentrations of antibiotic employed . S ince inhibition
o f severa l o f these system s , pyruv ic " hydrogenlya se " , amino
ac id dec arboxy l a s e ,oC-amylase , a lanine dehydrogena se and
polynuc l eotide phosphoryla s e , occur in in v i tro systems wi th
preformed enzyme s , the inhibi tions can not be solely a scribed
to faulty protein synthe s i s . La skin ( 17 ) , however , ha s
conc luded that " in no ins tance i s there compell ing evidence
to sugg e s t that the primary s ite of ac tion is involved among
thes e effects on oxidation and r e spiration . "
B . 3 . Effects on Protein Synthe s i s in Cultur e s
I n l 9 5 0 , Ga le and Paine ( 2 8 ) reported that chlortetracyc l ine
inhibited protein synthe s i s in S . aureus. Thi s s tudy showed that
the antibiotic inhibi ted " c ombined g lutamate " formation , but not
free g lutamate accumu la tion in wa shed c e l l suspens ions . Hahn and
Wi s seman ( 2 9 ) noted that adaptive enzyme f ormation wa s inhibited
in E . c o l i by both chlortetracyc l ine and oxytetracyc l i ne . They
a ttr ibuted this to a general inhibition of protein synthe s i s .

1 4
Ga le and Folkes ( 3 0 ) conf irmed that wa shed suspens ions of
S. aureus do not incorporate g lutamate into protein in the
pre s ence of growth inhibitory concentrations of tetracyc line ;
a 9 0 % inhibition o f rate o f incorporation a t 1 mcg . /ml . of
e i ther chlortetracyc l ine or oxytetracyc line wa s observed .
They a l so noted that inhibition of nuc leic ac id synthe s i s ,
rate of free g lutamate accumu la tion and rate of glucose
fermentation took place at higher antibiotic concentrations
and that the two antibiotic s d i f f ered quantitatively in the ir
effect on thes e parameter s . Park ( 3 1 ) and Sa zykin and Bori sova
( 3 2 ) in shor t communications , conf irmed the preferential
inhibition of protein synthe s i s . Park also noted that
funhibition of c e l l wa l l synthe s i s required higher antibiotic
concentrat i ons , 50 mcg . /ml . Neverthe l es s , Cheng and Snel l
( 1 ) , unaware of Park's r eport , have c la imed that the use of
D , L-Glutamate by Gale and co-worker s had not e l iminated the
pos s ib i li ty of a preferenti a l inhibition of D-Glutamate into
c e l l wal l mater i a l . Ha sh , Wishnick and Miller ( 1 9 ) have
r eported that c e l l s treated with tetracyc l ine show increased
c e l l wal l , hexosamine s and r i bonuc�e ic acid ( RNA) but decreased
protein and deoxyr ibonuc leic ac id ( DNA) as compared with
contr o l c e l l s . S imilarly , the incorporation of c�4 -labeled
lys ine , a lanine and g lutamic a c id into the cytoplasmic protein ,
i s o lated in a manner so that c e l l wa l l protein wa s e l iminated ,
wa s greatly inhibited by tetracyc l ine at 0 . 1 , 0 . 5 and 1 . 0
mcg . /ml . and chlortetracyc l ine at 1 . 0 mcg/ml . Incorporation
of c l 4_a lanine into c e l l wa l l material wa s not inhibited at

1 5
1 . 0 mcg . /mL o f c hlortetracyc l ine , but wa s inhibited at
50 mcg . /ml . ; tetracyc l i ne had no effec t on this incorporation
at 5 0 mcg . /m l . The kinetic s of incorporation of the amino
acids into cytoplasmic prote in revea led that inhibitrnon was
immediate . Mor r i s son et . a l . , ( 1 8 , 2 1 ) provided further
evidence in support of the theory that the ma j or effec t of
the tetracyc l ine s is an impa irment of protein synthe s i s .
v / Cer ny and Haberman (3 3 ) have made kinetic mea surements of the
inhibition of protein and nuc l e ic a c id synthes i s in cultures
o f E . coli C at tetracyc l i ne concentrations of 20 to 5 0
mcg . /ml . They found an immediate c e s sation of protein
s ynthe s i s f o l lowed by a latter , complete inhibition of nuc leic
acid synthe s i s . The s e concentrations are suff ic i ent to cause
c e l l death and it is unfortunate that the se workers d id not
employ addi t iona l lower antibiotic concentrations . Holmes and
W i ld ( 3 4 ) , dur i ng the course of investigatroons of the synthes i s
o f abnorma l r ibosomal RNA , have reported a l s o a t high
chlortetracycl ine concentra tions , results s imi lar to those of
v / Cerny and Haberman . They observed an immediate cessation of
protein synthes i s and continued RNA synthe s i s . Several
addi t iona l tetracyc l ine ana log s have a l so been stud ied and
s imilar f inding s reported ( 3 5 , 3 6 ) . Their mea surements were
completed approximately within the time of one culture
g eneration cycle and they did not observe the inhibition of
muc leic ac id synthes i s which occur s at longer growth time s .
These authors seem to feel that r ibosoma l precur sor RNA
accumu lated under antibiotic treatment i s not only the result

o f an inhibition of protein synthe s i s , but a l so due to a
spec i f ic s t imulatory effect o f the tetracyc line s .
Numerous investigator s have reported that the
16
tetracyc l ine s inhibit the formation of adaptive enzyme s . Sne l l
and Cheng ( l ) a s we l l a s La skin ( 1 7 ) have l i s ted these
inves t igat ions and have empha s i zed the low concentrations of
antibiotic needed to inhibit these phenomena . S�rm et . a l,, ( 3 7 )
i n addi t ion to Hahn and Wis seman ( 2 9 ) , have tr ied to interpret
their data as meaning that tetracyc l i ne caused an impairment of
the over a l l protein synthe s i z ing abi l i ty . These authors showed
that the inhibition o f g lutamic ac id decarboxylation was re lated
to the synthes i s of the enzyme and not to a lack of nec e s sary
co-factor s .
Sne l l and Cheng ( l ) have cautioned against drawing such
general conc lus ions from thi s type of data . They be l ieve that
an inhibition of adaptive enzyme f ormation might not be
suf f ic iently critical to account for bac ter iosta s i s . They were
a l so not sure at that t ime , 1 9 6 1 , that an inhibition of protein
synthe s i s would be able to cause the effects observed in bac ter i a l
culture s . They , however , f e l t that blocking a reaction in the
e lectron transport sys tem or an impairment of c e l l wa l l synthe s i s
wou ld b e suf f ic iently cr i tica l . A n over a l l inhibition of
protein synthes i s is presently thought to be c r itica l enough to
cause bac ter iosta s i s .
B . 4 . Effects on Cel l-Free Nitro-reduc ta se Systems
Ear ly reports ind icated that high concentrations of
chlortetracyc l ine were capable of uncoupling oxidative pho s
phorylation in norma l mitochondr ia ( 3 8 ) a s we l l a s i n r a t liver

17
and brain s l ic e s ( 3 9 ) . An interference with orthophosphate
uptake wa s conf irmed in the s tud i e s of Van Meter et . a l . , ( 4 0 ) .
The inhibition by c hlortetracyc line in rat l iver mitochondr ia
at low adenos ine tr iphosphate (ATP ) a nd phosphate concentrations
wa s shown to be reversed by added Mg ions . S imilar inhibi tions
and meta l ion r ever s a l were reported for oxytetracyc l ine and
tetracyc l ine by Brody , Hurwitz and Bain ( 4 1 ) . The se reports
suggested that the inhibition of protein synth e s i s repor ted by
Gal e and Pa ine ( 2 8 ) might be due to di sruption of the e lectron
transport system , a lthough a s imilar inhibition of nuc l e ic a c id
synthe s i s might have been expected . Inhibition wa s postulated
to occur because of interactions with meta l lo-enzyme s known to
be important in these systems . Miura e t . a l . , ( 4 2 ) showed that
oxytetracyc l ine wa s able to r educe incorporation of P3 2 into
a c id soluble phosphorus in intac t r e sting cultures of S. aureus
at growth inhibitory concentrations . However , these
experimental r e sults in add ition to being highly non- spec i f ic ,
could be interpreted a s due to the improper use of controls .
Contr o l c e l l s were a l lowed to enter the log pha se of growth
whi le inhibited c e l l s d id not leave the stationary pha se of
growth . In a r ev i ew of oxidative phosphorylat ion , Brody ( 4 3 )
reached the conc lus ion that the uncoupling caused by the se
antibiotic s wa s non-spec i f ic and was most probably the result
of r emoval of e s sential metal ions. Hunter and Lowry ( 4 4 ) in
a later rev iew d i sagreed with thi s v iewpoint because of the large
Mg ion/tetracyc l ine ratio r equired to rever se the inhibition .
However , they be l i eved that inhibition of these mamma l ian

18
sys tems by tetracyc lines occur s at such high concentrations
as to make unlike ly the postulate that this type of activity
is r esponsible for the compound ' s antibiotic ac tivity
Goma z kov ( 4 5 ) , Santi ( 4 6 ) and Morikawa e t . a l . , ( 4 7 ) have
recently studied thes e inhibitions and a l s o noted that the
high antibiotic concentra tions required for inhibition make
this an unlikely mode of action in bac teria l c e l l s .
The pos sibility sugge s ted by this ear ly work , that
tetracyc line s f unc tion as inhibitor s by chelation of e s s entia l
meta llo-enzyme system s , prompted Sa z and co-worker s to search
for other similar bacteria l systems . Saz and Maramur ( 4 8 )
r eported the inhibition of a c e l l-free nitro-reducta se system
by chlortetracyc line in 1 9 5 3 . The r ever sal of this inhibition
by manganese ions was reported by S a z and S lie ( 4 9 , 5 0 ) .
The se extrac ts were capabl e of mediating the r eduction of
a large number of aryl compounds ( 5 1 ) a s we l l a s the reduc tion
of nitrate to nitrite . The aryl nitro reduction wa s inhibited
by chlortetracyc line , but not oxytetracyc line and neither
compound interfered with nitrate r educ tion .
In addition to chlortetracyc line , sever a l of its
degradation products were active inhibitor s of the reduc ta s e ,
i . e . aureone amide , aureomycinic acid , anhydrotetracyc line ,
isochlortetracyc line and 4 -dedimethylamino-aureomycinic acid .
Several o f these compounds were a l s o ac tive in reducta se
experiments performed with whole c e l l s , a l though they are not
a l l bacteriostatic agents .

1 9
Further s tud i e s ( 5 2 ) with more pur if ied nitro-reductase
systems showed that they r equired ATP , d iphosphopyridine
nuc leotide (DPN ) a nd a d i carboxyl ic ac id a s wel l a s cyste ine
and manganese ions . If r educed d iphosphopyr idine nuc leotide
(DPNH ) wa s u sed in place of DPN and L-ma la te , the r educ tion
of aryl n i tro compounds wa s no long er inhibited by
chlortetracyc l ine .
As a r e su l t of these experiments , S a z and co-worker s
proposed that chlortetracyc l ine wa s an inhibitor of the
formation of DPNH from L-ma late and DPN . They thought that
it accomp l ished this inhibi tion by preventing fur ther
metabol i sm of oxa loacetic a c id , the ma j or product of the
DPN-malate reac tion . I t wa s known tha t oxa loacetic
decarboxyl a s e ha s a manganese ion requirement and this enzyme
wa s the pre sumed s ite of action . The s ite of action wa s
proposed to be spec i f ic for the chlor ine conta ining
tetracyc l ines a s oxytetracyc l ine or tetracyc l ine did not
inhibit the s y stem . The a b i l i ty of the inactive as we l l a s
active chlor ine containing tetracyc l ines t o inhibit the system
made i t a poor choice for the s i te of antibacterial action of
these compound s .
Saz and a s soc iates next d irected their attention to
purif ied extracts obta ined from both sens it ive and r e s i s tant
E . co l i . These more highly pur if ied extrac t s reta i ned some
but not a l l of the or ig ina l nitro-reducta se activ i ty . While
the extracts were no longer able to reduce DPN to DPNH , i . e . ,
they were devoid of mal ic dehydrogenase activity , they were

2 0
s t i l l able t o r educe chloramphenicol o r �-nitrobenzoic acid .
S i nce this r educ tion of p-nitrobenzoic a c id wa s inhibited by
chlortetracyc l ine they were f orced to conc lude that the ir
prev i ously proposed mechan i sm was not correct for the new
r educ tase ( 5 3 ) .
Extracts obta ined from sensit ive c e l l s were shown to be
greatly stimulated by f lavin mononuc leotide and manganese ions
whi l e tho s e from r e s i stant c e l l s were not stimulated . Kinetic
exper iment s demonstrated that the extract s from sens itive
c e l l s ox id i z ed stoichiometr ic amounts of DPNH in the reduction
o f FMN and the produc tion of p-aminobenzoic acid . Saz and
Martinez ( 5 4 ) interpreted the se results to mean that f lavin
and mangane se were loos e ly a s soc iated as co-factor s with
enzymes in sens itive c e l l s while they were more tightly bound
in r e s i s tant c e l l s . Plakunov ( 5 5 ) i n a s tudy o f resi stant
s trains has , however , r eported that the d i f ferences are mor e
l ikely a scr ibable t o d i f f erenc e s in the relative amounts of
FMN and f lav in adenine dinuc leotide ( FAD ) f ound in r e s i s tant
and s en s itive strains of S . aureus .
The hypothes i s that tetracyc l ines inhibit by interfering
with f lavins ha s been s tudied by Yag i e t . a l . ( 5 6 ) . They
showed that the mode of ac tion of the se compound s in the
inhibit ion of D-amino ac id oxida se was by complexation with
FAD . They f ound that chlortetracyc l ine formed highly s table
complexes with r ibof lav i n , FMN and FAD . They a l so showed that
the kinetic s of inhibition of the oxidation of D , L-alanine by
the enzyme-FAD preparation could be explained by a s suming the

rever s ible formation of an inactive chlor tetracyc l ine-FAD
complex .
Continued puri f ication of the reduc tase system by S a z
2 1
and Martinez ( 57 , 5 8 ) has not demonstrated that i t i s the s i te
of bac teri a l res i s tance to chlortetracyc l ine . They i solated
a protein- stab i l i z ed enzyme preparation which catalyzed the
reduc tion of 2 , 6 -dichloropheno lindophenol . This enzyme
preparation wa s inhibited by chlor tetracyc l i ne when prepared
from sensitive but not when prepared from r e s i s tant strains .
However , chlortetracyc l ine inhibited the reduct ion of
ferricyanide and s timulated the reduction of cytochrome c by
extracts prepared from both sens itive and r e s i s tant strains .
V i l lanueva ( 5 9 ) in a study of the nitro-reducta se of extracts
from both sensi t ive and r e s i s tant Nocardia ha s recently
reached the conc lus ion that there is no difference in
reductase activ i ty .
Krfmery and a s sociates ( 6 0 ) have continued work on the
hypothe s i s that development of tetracyc l ine r e s i s tance as we l l
a s a poss ible mode of action l i e s in the e lectron transport
cha i n . They have shown that triphenyltetrazol ium chlor ide
reduction by whole c e l l s of sens i t ive stra ins but not by who le
cells of r e s i s tant strains is inhibi ted by oxytetracyc l ine .
The degree of inhibi t ion var ied markedly with the type of
carbon source in the growth media . Further s tudi e s ( 6 1 ) showed
that malate a nd lacta te dehydrogena s e (MDH , LDH ) activities
dif fered in s en s i t ive and r e s i s tant s trains . Re s i s tant c e l l s
( 6 2 ) were a l so not able t o reduce tel lurite ( K2
Te03
) whi le

2 2
the sensitive stra ins had this abi l i ty , but were inhibited by
oxytetracyc l i ne . Crude c e l l-free preparations which catalyzed
MDH , LDH and N03 reduc tion showed s imi lar dif ferenc e s among
sensitive and resistant s trains . Recently ( 6 3 ) these worker s
have reported that cytochrome oxidas e activity in sensi tive
strains of S . aureus , E . coli and Brucella abortis are
inhibited by short exposure to oxytetracyc l ine . Re si s tant
s tra ins are not affected under the u sual aerobic conditions
but are inhibited only under anaerobic cond itions . The se
worker s be lieve that oxytetracyc l i ne interferes with microbial
enzyme systems involved in elec tron transport and that
resistant s trains have developed a l ternate pathways . Severa l
other worker s bel ieve , however , that resi stance to �etracy
c lines is a s soc iated with an a l tered permeab i l i ty of the c e l l s
( 6 4 , 6 5 , 6 6 ) .
Colai z z i , Knevel and Mar tin ( 1 1 ) used mammal ian cytochrome
C oxido-reductase system to obta in biolog ical activities for
use in struc ture-ac tivity corre lations . Whi le many of the
active tetracyc l ine s were inhibitor s of this system , severa l
tetracyc l ine s which a r e not considered bac teriostatic were
equ a l ly good inhibitor s . The author s felt that this might
be explained by dif ferences in abi l i ty to permeate c e l l wal l s .
However , S a z and co-worker s ( 5 1 ) have previou s ly noted that
such reasoning wa s not va l id in exp laining difference s in
activity in the ir nitro-reductase system . The high
c oncentrations ( lo - 3 �) of antibiotic nece s sary for inhibition
of this sys tem are typical of results obta ined with mammal ian
system s .

B . 5 . The Nature of C e l l -Free Protein S ynthe s i z i ng
Systems - Translation of the Genetic Code
2 3
The purpose o f thi s section i s to summar i z e the exi sting
knowledge of c e ll-free protein synthe s i z ing systems . In this
way , the reader may have a better under stand ing of the effects
of tetracyc l ines upon the se systems and can judge whether
the se effects are suf f ic ient to account for the inhibition of
growth of whole c e l l s . In add i tion , several recent experiments
that have led to new and detai led mode l s of thi s system wi l l be
d i scus sed .
Trans lation refers to the proc e s s in the synthesis of
proteins by which a particular sequenc e of nuc leotide ba ses
in mes s enger RNA (mRNA) determine a specific sequence of
amino acids in a polypeptide cha i n . Translation i s
accomp l i shed b y the interac tion o f three c la s ses of
macromo lecules : mRNA , the spec i f ied amino ac id cova lently
l i nked to transfer RNA ( tRNA ) and a comp lex r ibonuc leoprotein
cal led a r ibosome . Severa l exc e l lent review ar ticles have
appeared recently which descr ibe exi sting knowledge of the
system in deta i l (67, 68 , 69, 70).
The bas ic components of a s imple c e l l-free protein
synthes i z ing system are :
l . Ribosome s
2 . mRNA
3 . Arninoacyl tRNA (AAtRNA)
4 . Ribosome a s sociated ini tiation factor s , f 1 and f 2
». Non-r ibosome a s soc iated ( supernatant ) enzyme s , T and G

2 4
6 . Guanos ine - 5� Triphosphate ( GTP )
I n addition to these bas ic components , a ce l l - free
system usual ly includes a buf fer , pH 7-8 ; monovalent inorganic
s a l ts ; divalent salts or a polyamine polypeptide ; and a
sul fhydryl reducing agent .
More complex sys tems , or alternate ly more crude systems ,
may generate within the sys tems AAtRNA and these sys tems
require addi tional ly ATP , pyruvate kinase (PK ) , phosphoenol
pyruvate ( PEP ) , amino acids (AA ) , soluble RNA ( sRNA) and
aminoacyl synthetase s . A few experiments have been performed
with systems that generate mRNA s imul taneously and these
require DNA , DNA-dependent RNA polymerase and appropriate
nucleotide s .
B . 5 . a . Ribo somes
Bacterial ribosome s are complex aggregates which cons i s t
of 6 0 - 6 5 % RNA and 3 0 - 3 5 % prote in . They serve a s a template
for the assemb ly of peptide bonds between AAtRNA molecules
held on the ribosomal surface . I n addition to thi s inactive
role , it is thought that peptide synthetase enzyme s as we l l
as initiation enzymes may b e an integral part of the normal
ribosomal surface . Because the s i te of action of tetracycl ines
is presumed to be on the ribosomal surface , a detailed
des c ription of ribosomal structure wi l l be presented .
Ribosomal particles partic ipate in a series of revers ible
as sociation reactions which are i llustrated in Fig . I I I .
The species active ly associated with protein synthe s i s has
been shown to be the 70 S ribosomes ( 71) , which have a

506
36S �
'I� 70S
R I�OM E
t.,. :I
� 2 6 S
C OR E !lOS ssJSsmm S U B UN I T
S PL �0
P�OTE INS( · · �
+
2JS r RNA 6S r R NA +
JJ ) ) � ) � � � ) ) � ) �0 S PRO TE I N S
�_, JOS � C�� S U B Ui T SSJ!fffffff \� J OS f S P L I T P R O T EINS
� ISS r R NA
J O S P RO TE I N S
2 5
Figure I I I . Ribosomes , Components , Subunits , As sociation and
D i s sociation Reactions , Shaded c ircles depic t the ribosoma l
partic les , scrambled l ine s depict the ribosoma l ribonuc leic
acids and s - shaped lines depict the ribosomal proteins for
which the ind icated S values refer to the sedimentation
values of the origina l ribosome subuni ts .

molecular weight of approximately 2 . 7 x 1 06 and a d iameter 0
of about 2 0 0 A . They dimeri z e i n solutions of high Mg ion
concentration in an apparently non-phys iolog ica l reac tion ,
2 6
they a l so undergo an apparent shape change i n EDTA solutions .
The d i s sociation of 7 0 S r ibosome s to 5 0 S and 3 0 S subunits
occur s at Mg ion concentrations s imi lar to those found in
bacteri a ( 7 0 , 7 2 ) . The d i s soc iation of 70 S r ibosomes to
subunits i s inhibited by sulfhydry l blocking agents or
polycationic polyamines a s we l l as high Mg ion concentrations
( 7 3 , 7 4 ) . Moore ( 7 5 ) , in a s tudy of the effects of chemica l
reagents on subunit a s soc iation , ha s conc luded that hydrogen
bond formation e i ther between nuc leotide ba s e s or between a
nuc leotide base and a phosphate group are respons ible for
this a s soc iation .
The 3 0 S submm i t cons i s ts of a s ing le RNA molecule ,
1 6 S rRNA , with a molecular weight of approximately 0 . 6 x 1 06
,
and a t lea st 1 5 spec i f ic prote ins ( 7 6 ) . The 5 0 S subunit
cons i s ts of 2 molecu l e s of RNA , one a 2 3 S rRNA and the other
a 5 S rRNA , as wel l as at leas t 19 spec i f ic prote ins ( 7 7 ) .
The 5 S spec i e s of rRNA wa s f ir st i so lated in 1 9 6 4 by
Ros sett et . a l . ( 7 8 ) and wa s shown to be d i s t inc t from sRNA
which ha s a s imilar sedimentation coe f f ic ient . The entire
nuc leotide sequence of 5 S rRNA from KB carc inoma c e l l s ha s
been determined by Suther land , Carr ier a nd Setlow ( 7 9 ) , and
a l s o that of E . col i , by Brownlee and Sanger ( 8 0 ) and Brownlee ,
Sanger and Barre l l ( 8 1 ) . This r i bosoma l RNA l ike 1 6 S and
2 3 S rRNA conta ins no unusua l nuc leotide s .

2 7
The secondary s tructure of thi s relative ly new rRNA i s
also thought t o b e s i milar t o that of both 1 6 S and 2 3 S rRNA .
I t cons i s ts partly of double stranded hel ical regions and
partly of non-hydrogen bonded s i ngle s trands ( 8 2 ) . Extens ive
s tudi e s on the phy s i cal and chemical properties of rRNA have
been reviewed ( 8 5 ) .
The 5 ' terminal nuc leotide sequences of E . col i 1 6 S and
2 3 S rRNA have been determined by Takanami ( 8 3 , 8 4 ) . These
determinations show that the two rRNA ' s are d i s tinct species .
The prote in components of r ibosome s have been s tudied in
considerable detai l . The large number of prote ins indicated
i n Fig . I I I , have been i solated and s tudied by : end-group
ana lys i s ( 8 6 ) , ultracentri fugation ( 8 7 , 8 8 ) , s tarch gel
electrophore s i s ( 8 6 ) , polyacrylami de gel electrophores i s
( 7 6 , 7 7 , 8 7 , 9 0 , 9 1 ) and chromatography on
carboxyme thylcel lulose ( 9 1 ) and Sephadex ( 9 1 ) They are unique
proteins wi th d i s t inct amino acid analys i s ( 8 9 , 9 1 , 9 2 ) and are
not the result of aggregate s formed during preparation . The
large number o f r ibosomal proteins argues against the theory
that rRNA could serve as a mRNA for their synthe s is . Ribosomal
RNA doe s not contain enough trinucleotide s to synthe s i ze thi s
many large , d i s tinct prote ins ( 7 0 ) .
I t s eems l ikely that ribosomal proteins play d i s tinct
and non- i nterchangeable roles ( 9 3 , 9 4 , 9 5 ) . Thei r functional
spe c i f i c i ty has been inve s tigated by a degradation -
recomb ination technique . In these experiments the proteins are
partially spl i t from the ribosomal subuni ts with C sC l ( Fig . I I I )

2 8
and the l i berated core particles and split prote ins are
separated , isolated and identif ied . Normal r ibosome func tion
can be achieved by mixing the isolated components . Ribosomes
reconstituted in thi s manner but lacking one or more split
proteins have been shown i n some cases to lack one or more ,
but not a l l , r ibosoma l functions ( 9 4 ) . This technique ha s
been u sed to show that streptomycin sensi tivity , resistance
and dependence l i e with the 2 3 S core particle and not with
the split prote ins ( 9 6 , 9 7 ) . Sensitivi ty and resistance have ,
however , been shown to l i e i n the prote in frac tion of the
core partic l e s and not in the rRNA ( 9 8 ) .
EDTA treated r ibosomes conta in a l l known r ibosome
components but probably because they have dif ferent hydro
dynamic proper ties they lack certa in spec ific ribosoma l functions
( 9 9 ) •
Treatment of r ibo some s with LiC l causes a loss of
protein components s imi lar to that which i s produced by cSC l .
L iC l -r ibosomes are now known to be identical to the particles
found in bacterial culture s inhibited by chloramphenicol ( 1 0 0 ) .
The se r ibosoma l particles are probably the same a s those found
in tetracyc l ine i nhibited cultures ( 3 4 , 3 6 ) . Such r ibosomes
are thought to be norma l precur sor par tic les .
The effect of Mg ion concentration on the a s soc iat ion
d i s sociation of r i bosomes is outl ined in F ig . I II . I t ha s
recently been shown that Mg ion i s required to neutra l i ze the
phosphate g roups in the r ibosoma l RNA . "Norma l " 7 0 S r ibosome s
ex i s t i n an ionic environment in which the ratio of

2 9
r ibosomal-bound Mg ion to ioni zed r ibosomal pho sphate is 0 . 5
( 7 2 , 1 0 1 , 1 0 2 , 1 0 3 ) . Cationic groups , such a s ca++ , NH4+ ,
spermidine or putr e sc ine , in the proper concentrations are
a l so capable of cata l y z i ng the formation of functiona l
r ibosomes ( 7 2 , 1 0 2 , 1 0 4 , 1 0 5 , 1 0 6 , 1 0 7 ) .
B . 5 . b . Mes senger Ribonuc leic Acid
The existence of a spec ies of rapidly synthe s i zed , unstable
RNA , c a l led mRNA , wa s pred icted in a review by Jacob and
Monod ( 1 0 8 ) in 1 9 6 1 . The demonstration of the exi stence of
such a spec i e s ha s been accomp l i shed by exper iments ba sed on
two phenomena : i t s rapid rate of synthe s i s and turnover a s
compared with rRNA o r sRNA and a l s o the synthe s i s of thi s type
of RNA in phag e - i nf ec ted c e l l s . The pred icted function of mRNA
wa s that i t carr ied the information of the genetic code from
DNA to the r ibo somal template , where it directed the sequence
of add ition of amino acids into protein . Evidence in support
of this predic tion ha s been rev i ewed by S impson ( 1 0 9 ) and a l s o
b y S i nger and Leder ( 1 1 0 ) . Conv inc ing ev idence for t h i s role
has been found in the demonstration of mRNA in peptide
synthe s i z ing polysome s ( r ibosome-mRNA aggregate s ) ( 1 1 1 ) , in
the synthe s i s of phage proteins of known sequence by bacter ia l
systems d irec ted by phage mRNA ( 1 1 2 ) and in the coinc idence of
amino ac id sequence and nuc leotide sequence of synthetic
polyr ibonuc leotides ( 1 1 3 ) .
Trans lation of the code i s not accomp l i shed by a d irect
interaction of an amino ac id with a spec i f ic tr inuc leotide
sequence ( codon ) of a mRNA , but rather through an intermed iate

3 0
adaptor mo lecule , tRNA , cova lently linked to a n amino acid .
The tRNA conta ins a spec i f ic trinuc leotide sequence ( anticodon)
that recogn i z e s the codon . The codons which direct the
incorporation into protein of particular amino acids have been
determined by studies of the bind ing of AAtRNA ' s to r ibosoma l
codon , where the codon wa s e i ther a trinue leotide ( 1 1 4 ) o r a
polyr ibonuc leotide of known sequence ( 1 1 5 ) . S imilar i ly
incorpora tion of AAtRNA ' s into protein in systems directed by
polyr ibonuc leotide s of known s equence ( 1 1 6 ) has a l so been used
to determine the codons which d irect the incorporation of
part icular amino acids . The four common nuc leotide bases
when uti l i z ed in a trinuc leotide code can be arranged in 6 4
poss ible combina tions . S ince there a r e only 2 0 amino acids ,
degenerac ie s might be expected and indeed are found in the
code . The i solation of s everal d i f f erent tRNA ' s capable of
accepting a g iven amino a c id ha s led to the speculation that
there may be a particular tRNA corresponding to each dodon .
D i ff erent leuc ine-tRNA spec i e s have been shown to s elec tively
bind to d if f erent codons ( 1 1 7 ) . However , i t i s unlikely that
this is the c a s e for a l l tRNA molecules ( 1 1 8 ) .
To account for the known degenerac ies in the coae , Crick
has introduced a theory of codon - anticodon pa ir ing , cal led
the " Wobble Hypothe s i s " ( 11 9 ) . This theory states that the
bind ing of the f ir s t two bas e s of the codon ( 5 ' to 3 ' ) with the
appropr iate bases of the anticodon ( 3 ' to 5' ) fol lows s trict
Watson-Crick hydrogen bond ba se pair ing . The interaction of
the third ba se of the codon with the third ba se of the anticodon

3 1
i s not spec i f ic , but ha s certain a l lowable exception s . These
exceptions are not random but may be pred icted by a cons idera
tion of the bond d i s tances as soc iated with the non-classical
hydrogen bondi ng of codon to anticodon . The sing le exception
to the known c ode degenerac i e s occurr i ng i n the third ba se of
the codon is in those sequence s capable of cod ing for the
initiator , N-formyl -methiony l - tRNA ( N-F-met-tRNAf ) where
degenerac i e s in the f ir s t ba se of the codon exi st ( 12 0 )
I nterpretation o f this phenomena has not been made .
S i nce i t i s known that hydrogen bondi ng between
trinuc leotides is not suff ic i ently s trong to s tabi l i z e mRNA
AAtRNA complexes , the s tabl i l i z i ng role of the ribosome must be
empha s i zed .
B . B . c . Transfer Ribonuc leic Ac id
The adaptor mol ecule , tRNA , i s a re latively smal l molecule ,
4 - 5 S , which forms an e ster with activaned ami no acids i n the
pre s ence of amino ac id syntheta s e . Thi s reaction i s a
relatively wel l under stood area of biochemistry ; it i s depicted
in schematic form in F ig . IV and ha s been de scr ibed in severa l
reviews ( 1 2 1 , 1 2 2 ) . That tRNA func tions a s an adaptor molecule
ha s been shown by demonstrating that cysteine-tRNA , chemic a l ly
converted to a lanine - tRNA , leads to the incorporation of a lanine
i n place of cyste ine into protein ( 1 2 3 ) .
The nuc leotide sequence s of at leas t four tRNA mo lecules
have been determined complete ly ( 1 2 4 , 1 2 5 , 1 2 6 , 1 2 7 ) . A common
feature of thes e and a l l other tRNA ' s which have been s tud ied
is a termina l . . . pCpCpA nuc leotide s equence . I t i s thi s part

A 8
D E
+AMP
F igure IV . The Formation of Aminoacyl Transfer Ribonuc leic
Ac id . (A ) Binding of ATP and an amino acid to the enzyme .
( B ) Forma tion o f a n " activated amino ac id . " ( C ) Binding
of tRNA to the enzyme . ( D ) Displacement of the origina l
adenylate bound t o the enzyme b y the termina l adenylate of
the tRNA . ( E ) Forma tion of the e ster , AAtRNA .
3 2

3 3
of the molecule which accepts the amino ac id ( see Fig . IV)
and which is probably involved in the binding of the AAtRNA to
the 5 0 S r ibosomal subunit ( 1 2 8 ) .
I n add i tion to the above mentioned amino acid accepting
s i te , each spec ies of tRNA mus t conta in a region which enables
it to accept , exc lu s ively , the proper amino a c id ( recogni tion
s i te ) and a l so a reg ion which conta ins the appropr iate anticodon
for the amino a c id accepted . Reg ions in tRNA which might contain
these s ites have been predicted on the ba s i s of a c lover - leaf
secondary s truc ture postulated for tRNA . This is shown in a
general manner , fol lowing Ful ler and Hodg son ( 1 2 9 ) in Fig . V .
This structure was f ir s t proposed by Holley et . a l . ( 1 2 4 ) for
yeast ala-tRNA . I t i s ba sed on s tudies of hydrogen bonding and
the results of X-ray d i ffrac tion studies which have shown that
tRNA ' s exi s t in par tly helical and partly non-helical conforma
tions . A pos s ible codon - anticodon interaction , shown in
F ig . VI ha s been g iven by Ful ler and Hodg son ( 1 2 9 ) .
B . S . d . A Mode l of C e l l-Free Protein Synthe s i s
The sequence of events i n a cell-free protein synthe s i z ing
system is shown in F ig . V I I . They may be outl i ned a s fol lows :
Step 1 . - Binding of a 3 0 S r ibosomal subunit to a mRNA .
Step 2 . - Formation of the ini tiation complex .
a . - Al ignment of the codon for N-F-met- tRNAf on
the r i bosomal surface so that i t may bind to
the anticodon , i . e . , creation of the N-F-met
tRNAf or peptidyl binding s i te .

AMINO ACID ARM
I' p
�
f c I c I 3' X
�H'\\, ... ,� XTRA ARM
J=t ' / t 7 � ANTICODON ARM
"W0811U" �NTlCOOON BAS[
F igure V . Schematic Diagram o f the Proposed C lover-Leaf
Structure of Transfer Ribonuc leic Ac id s . Hydrogen bond s
are i nd icated a s l ines l i nking the strand s . The number of
nuc leotides i n each s i ng le stranded loop i s indicated
within the loop . X ind icates a nuc leotide which var i e s
with the tRNA spec ies , r i s pseudour idylic a c id , T i s
thymidylic ac id , DMG i s d imethylguanylic acid .
3 4

3'
P E P T I D Y L
T R NA M R N A
A M I N O ACYL
TRNA
5'
F igure V I . Codon - Ant icodon Interaction . Hydrogen
bonds are i ndicated by double l ine s l inking the r ibo
nuc l e ic ac ids ; depicted by shaded l i ne s . A hel ical
relat ionship i s impl ied between the anticodon loops
of the tRNA ' s and the mRNA . Only the anticodon arms
of the tRNA ' s are shown , see F igure V .
3 5

3 6
Figure V I I . A Model o f Cell-Free Protein Synthe s i s .
Shaded half-circles depict 3 0 S r ibosoma l subunits ,
shaded circles depict 5 0 S ribosomal subunits , a wavy
line depicts mRNA and tRNA ' s are depicted by the
symbo l , � , the s-shaped curve depicts a protein .

3 7
ST E P I . - ST EP 2 A .-
M- F - t.I E T
STE P 2 B.- ST E P 3 . -
ST E P 8. -
F igure VI I .

3 8
b . - Bind i ng o f N-F-met- tRNAf to the r ibosome-codon
in a reac tion which i s s t imula ted by and may
require the initiation factor s and GTP to form
the so-cal led initiation complex .
Step 3 . - Attachment of the 5 0 S subunit to the initiation
complex to form a monosome . Thi s binding s imulna-
neou s ly creates a binding s i te for AAtRNA . The
aminoacyl bidding s ite corre spor.ds to the 3 ' end of
the me s senger . l
Step 4 . - Binding to the aminoacyl s ite on the 7 0 S r ibosome .
E i ther AAtRNA a lone or the complex AAtRNA-GTP-T may
f i l l thi s s ite , where T is a supernatant protein .
Step 5 . - Formation of the peptide bond . N-F-met-AAtRNA would
be the f ir s t peptide formed on the ribosome . Thi s
react ion may b e cata lyzed b y a tightly bound r ibosomal
peptidyl syntheta s e .
S tep 6 . - Pulsation (Movement) of the r ibosome .
a . Remova l of tRNAf from the peptidyl s i te , either
to an exi t s i te or directly runto the surrounding
solution .
1 r f attachment of a 7 0 S r ibosome to mRNA occur s in Step l rather than a 3 0 S subuni t , then both peptidyl and aminoacyl binding s ites may be created at that t ime . Al ignment of the codon may be such that N-F-met- tRNA may bind to either site . I f bind ing to the aminoacyl s i te occur s , then trans location of N-F-met-tRNA is required before the next step . Step 3 is then , of cour se , not nece s sary . Steps 1 to 3 may be repeated to form a polysome with d i fferent r ibosomes attaching to the same mRNA a f ter incorporation of about ten amino acids into a polypeptide .

3 9
b . - A s imul taneous shift o f the mRNA and
N-F-met-AAtRNA on the r ibosome surface so
that a new codon is a l igned under the
aminoacyl s ite and the peptidyl tRNA occupies
the peptidyl s i te .
Step 6 i s accompanied by hydroly s i s of GTP
catalyzed by G , a r ibosome-dependent GTPase .
Step 7 . - Steps 4 , 5 and 6 are repeated to form an increas ing ly
longer peptide cha i n .
Step 8 . - Chain termination and release .
a . - E i ther repetition of Step 4 i s prevented by
a l ignment of a nonsense codon wi th the aminoacyl
s i te or Step 5 is prevented by occupation of the
aminoacyl s ite by a non-func tional sRNA bound by
the nonsense codon . ( Such a nonfunc tional sRNA
which bind s to r ibosome-nonsense codon ha s not
as yet been found ) .
b . - Hydroly s i s of the peptidyl-tRNA on the r ibosome
followed by e j ec tion of the peptide and tRNA or
removal of an intact peptidyl-tRNA from the
r i bosome and subsequent hydroly s i s to peptide and
tRNA .
Several s teps in the proposed mode l have been s tudied in
great deta i l and their nature i s at least partially unders tood .
A lack of spec if ic techniques has prevented thorough inves tigation
of some of the other steps .

4 0
Step l . - B ind ing o f a 3 0 S subunit to mRNA .
Attachment of mRNA to the 3 0 S subunit rather than the
5 0 S subuni t wa s demons trated by Okamoto and Takanami ( 1 3 0 ) .
Bind ing of mRNA to 7 0 S r ibosomes has been presumed to occur
through the attachment to the 3 0 S subunit and not because of
the forma tion of some new s i te which does not exi s t on either
subunit a lone . Moore ( 1 3 1 , 1 3 2 ) has presented evidence that
there i s only one binding s ite for mRNA on 7 0 S ribosomes and
that poly C and poly U compete for this site . The attachement
of mRNA ' s to r ibosome s is thought to invo lve only sing le
s trand ed regions of the mRNA , s ince double s tranded mRNA ' s
do not bind ( 1 3 3 ) . Thi s observation accounts for the strong
bind ing of poly U to r ibosomes since i t ex ists a s a random coil
above 4 ° C . The length of mRNA bound to ribosome s has been
e s t imated to be about 2 7 nuc leotide s . This estimate is ba sed
upon the number of nuc leotide s which are protec ted from
r i bonuc lease hydroly s i s by the ir attachment to r ibosomes . Moore
( 7 5 ) ha s u sed chemical modif ications of r ibosome s and/or mRNA
to study their interac tion . He found that the mo st likely
groups involved in the bind ing were amine groups in the rRNA
and the phes phate backbone of the mRNA .
Step 2 . - Format ion of the initiation complex .
S ince r ibosomes d irec ted by natural mRNA ' s synthe s i z e
comp lete prote ins there mu st b e control mechani sms whereby initial
read ing of mRNA by r ibosomes i s restric ted to a specif ic s i te .
Such control might resu l t from attachment of r ibosomes at a
spec i f ic si te on mRNA . This mechanism would not be suf f ic iently

4 1
strict to a l low proper pha s i ng of the mRNA readout , s ince the
only requirement for binding i s that the mRNA be s i ng l e
s tranded . ( Pha s ing i s the a l ignment of the f irst ba se of a
tr inuc leotide codon whos e trans lation i s des ired on the
r ibosome , so that it spec i f ie s the readout . When a l ignment
is such that the second or third bas e specify the code
readout then the read ing of the mRNA is said to be out of pha se . )
In 1 9 6 1 , Wa l ler ( 1 3 4 ) d i scovered that me thionine wa s the
NH2 -termina l group in at least 4 5 % of a l l E . co l i proteins .
Fol lowing this repor t , Marcker and Sanger ( 1 3 5 ) observed that
one spec ies of met- tRNA could be formylated (met- tRNAf ) while
other spec ies could not be formy lated (met-tRN� ) . A l ikely
f ormyl donor wa s s hown to be N 1 0-Formy l - tetrahydrofolic a c id
( 1 3 6 ) . They showed that formylation occurred only a f ter
formation of the AAtRNA and that an enzyme d i s tinc t from
methionyl synthe ta s e wa s involved . C lark and Marcker ( 1 3 7 )
observed that N-F-met from N-F-met-tRNAf was incorporated into
the NH2 -terminal position of prote ins synthes i zed by a c e ll-free
extract . Later ( 1 3 8 ) they showed tha t c e l l-free extracts
directed by poly UG and capable of formylating met-tRNAf
produced proteins in which more than 5 0 % of the NH2 - terminal
groups were N -F -met . This f inding seemed to explain Wa l ler ' s
early repor t ( 1 3 4 ) .
Adams and Capecchi ( 1 3 9 ) showed tha t N-F-met-ala- ser-a sp-
-phe- thr- was the initial amino acid sequence in R 17 phage
coat protein produced by an extract directed by R 17 me s senger .
S ince the identical s equence without the initial N-F-met wa s

4 2
found i n endogenous coat protein , these worker s proposed
that N-F-met-tRNAf was the univer sal bac ter i a l chain initiator ,
but that in some cases e i ther the termina l N-Formyl or the
N-F-met groups were removed by hydrolysi s . Webster , Eng lehardt
a nd Z inder ( 1 4 0 ) �eported s imilar f inding s . Capecchi ( 1 4 1 ) ,
We i sbach and Red f i e ld ( 1 4 2 ) and Lugay and Aronson ( 1 4 3 ) have
s tudi ed the hydroly s i s of NH2 - termina l formate in cell - free
extracts and have attempted to i solate the respons ible enzyme . �
Vinuela , Sala s and Ochoa ( 14 4 ) found that a l l v iral prote ins
as wel l as the coat proteins synthe s i zed in systems directed
by MS- 2 phage RNA had N-F-met as the NH2 -terminal group .
In add i t ion to demonstrating that N-F-met from
N-F-met-tRNAf wa s the f ir s t amino a c id incorporated into
protein by c e l l -free extract s . C lark and Marcker ( 1 3 7 , 1 3 8 )
proposed that the d i f f iculty in a s s igning a codon to met-tRNA
might be due to the ex i s tence of two dif ferent codons , one
codon for met- tRNAf a nd another for met-tRNAm . Sundararaj an
and Thach ( 1 4 5 ) have estab l i shed that binding of met-tRNAf to
r i bosome s i s best when AUG i s the codon , but other triplets
a l s o code for it at high Mg ion concentrations . Met-tRNAm
i s only coded for by AUG . They showed that N-F-met- tRNAf
wa s c apable of pha s i ng the reading of synthetic mRNA ' s ,
such a s AUGG ( U l n · I n the presence of N-F-met-tRNAf bind ing
of val-tRNA in response to the second triplet , GUU , wa s
s t imu lated . Out o f pha s e bind ing of other AAtRNA ' s in
response to UGG or GGU wa s suppressed . Binding experiments
with other synthetic mRNA' s , inc lud ing several with the AUG
codon not a t the 5 ' termini , gave s imilar results . B inding

4 3
above background d id not occur at low Mg ion concentrations
if the AUG codon wa s located at the 3 ' end of the mRNA .
Thach , Dewey , Brown and Doty ( 14 6 ) reported s imi lar results
when incorporation of amino ac id i nto protein wa s used rather
than r ibosome-codon bindi ng . They noted that if an AUG
codon appeared in a mRNA at a position internal to another
AUG , thi s second codon a lways d irected the incorporation of
met-tRNAm . They felt tha t a l ignment of the AUG codon at the
peptidyl s ite spec i f ied chain initiation by N-F-met-tRNAf ,
whi l e a lignment of the codon at the aminoacy l s i te spec i f i ed
chain continuation with met-tRNAm · Leder and Bur s z tyn ( 1 4 7 )
have confirmed the se f ind ing s on phas ing o f readout by
u s i ng AUG ( U ) n as a mRNA and measur ing the incorporation of
met , val and phe . They a l so showed that N-F-met-tRNA bound
dur i ng binding exper iments wa s not displaced from r ibosomes
by the addition of other AAtRNA ' s . Thi s f indi ng in conj unction
with later results of Nomura , Lowry and Guthr ie ( 1 4 8 ) show that
binding of N -F -met-tRNAf ha s relevance in peptide synthesis .
A solution to the pu z z l ing enigma that c e l l - free extrac ts
d irected by synthetic mRNA ' s requ ire higher Mg ion concentra
tions than those direc ted by endogenous mRNA was sugge sted by
Clark and Marcker ( 13 7 ) . They noted that met-tRNAf bind s to
r ibosome-codon at much lower Mg ion concentrations than those
which are nec es sary for bind i ng met-tRN� . Thi s observation
was a l s o reported by Sundararaj an and Thach ( 1 4 5 ) . Nakamoto
and Kolakof sky ( 1 4 9 ) have shown that the Mg ion optimum of a
system incorporating phe-tRNA but d irec ted by a mRNA which

4 4
a l lows incorporation of N-F-met- tRNAf wa s greatly lowered by
inc luding the l atter AAtRNA in the reaction mixture .
Kolakof sky and Nakamoto ( 1 5 0 ) studied the Mg ion dependency
of extracts d irec ted by MS- 2 viral RNA . They found a lowered
Mg ion optimum and a large s t imulation of over a l l
incorporation when a formyl donor system wa s added t o the
extract . The synthes i s of coat prote in and a l s o other viral
proteins wa s shown to fol low this pattern .
Clark and Marcker ' s original suppos i tion ( 1 3 7 ) that
N-F-met-tRNAf wa s capable of initiating prote in synthe s i s
because the formamide bond resembl ed a peptide bond wa s shown
to be f a l s e by these same worker s ( 1 3 8 , 1 5 1 ) . They
demonstrated that met- tRNAf as wel l as N-F-met-tRNAf could act
a s an initiator , Leder and Bur s z tyn ( 1 4 7 ) and a l so Sa l a s
et . a l . , ( 1 5 2 ) conf irmed t h i s r e su l t . Leder and Bur s z tyn ( 1 4 7 )
sugges ted that formylat ion might take place af ter attachment
to the r ibosome-codon . They showed that this type of r ibosoma l
formylation wa s pos s ible . Economou and Nakamoto ( 1 5 3 ) have
stud ied the kinetic s of the formylat ion reac tion and conc luded
that the reaction is so fast that met- tRNAf would be formylated
before binding to r ibosomes in any extrac t capable of
performing the reaction . I t mu st , however , be conc luded that
the unique abi l i ty of N-F-met-tRNAf to ini tiate protein
synthes i s l ie s in the tRNAf port ion of the mo lecule and not
in the formamide bond itself .
Incorporation of met- from met-tRNAm into interna l portions
of peptides synthe s i z ed by c e l l-free extracts wa s reported by
Thach et . a l . , ( 1 4 6 ) . Clark and Marcker ( 1 5 1 ) conf irmed thi s

4 5
f indi ng i n systems directed by bac ter iophage RNA , Stanley
et . a l . , ( 1 5 4 ) and Salas et . a l . , ( 1 5 5 ) reached s imi lar
conc lusion s u s i ng synthetic mRNA ' s conta ining the AUG codon .
In summary , these results have been interpreted a s
r e f l ec ting the exi stence of two binding s ites o n the ribo some .
This theory states that N-F-met-tRNAf can unique ly bind to
the peptidyl s i te in response to the AUG codon , whi le
met-tRNAm is bound when the AUG codon is a l igned with the
ami noacyl s i te . Ma tthaei and Voig� ( 1 5 6 ) have demonstrated
the exi stence of bind i ng s ites on the r ibosome-codon with
two d i f ferent Mg ion optima , one of which binds N-F-met-tRNAf
and the other which bind s met-tRNAm .
The reac tiv i ty , in the absence of supernatant factor s ,
of r ibosoma l bound -N-F-met-tRNAf with puromyc in ( Pm) is
cons i stent with the conc lus ion that N-F-met- tRNAf occupies
the peptidyl s ite ( 1 5 1 , 1 5 8 ) . Other r ibosoma l -codon bound
AAtRNA ' s do not have thi s reactivity in the absence of super
natant . Her shey and Thach ( 1 5 9 ) have repor ted that the Pm
reaction i n their extrac ts , a s we l l a s the forma tion of the
d ipeptide N-F-met-phe wa s strongly stimulated by GTP and
supernatant enzyme s . They propose that N-F-met-tRNAf i s
initia l ly bound t o the aminoacyl si te o f the r ibo some and
is then transferred i n a GTP-dependent step to the peptidyl
site - this ha s been termed the sing l e entry s i te model .
Alternate ly , b i nd i ng may occur d irectly with the peptidyl s i te
but a GTP-dependent chemical activation of the bound met-tRNAf
must then occur before d ipeptide formation - thi s ha s been

4 6
termed the peptidyl s ite activation mod e l .
Stanley et . a l . , ( 1 5 4 ) and Salas et . a l . , ( 1 5 5 ) have
i solated two protei n components , f 1 and f 2 , from salt washings
of E . coli r ibosomes . These proteins are required for the
trans lation of natura l mes seng er s , but not for synthetic mRNA ,
unle s s they conta in the AUG codon . Trans lation of synthetic
mRNA ' s which conta i n AUG i s greatly s t imulated by the se factor s .
This stimulation results from an i ncreased i ncorpora tion of
met-tRNAf but not met-tRNAro . The se f ac tors s timulate
incorporation only at low Mg ion concentrations and do not
promote the bindi ng of other AAtRNA ' s . Ander sen , Bretscher ,
C lark and Marcker ( 1 6 0 ) and Thach , Dewey and Myko la j ewyc z
( 1 6 1 ) and Leder and Nau ( 1 6 2 ) have substantiated these results
and have shown that the binding promoted by thes e f ac tors is
stimulated by addi tion of GTP . I t seems l ikely that in the
presence of the se factors and GTP , N-F-met-tRNAf is bound
e ither directly to , or rapidly transferred to , the peptidyl
s ite . Bind i ng , in their absence , may be to the aminoacyl s ite .
S im i lar f ac tor s have been i so lated by Eisenstadt and
Brawerman ( 1 6 3 , 1 6 4 ) and Revel and Gros ( 1 6 5 , 1 6 6 ) . The se
authors have also noted the pos s ible exi stence of a third
factor , retarded on DEAE-c e l lulose , which may s t imu late or
may be an e ssentia l requirement for the bind i ng of r ibosome s
to mRNA in the presence of DNA .
Step 3 . - Attachment of the 5 0 S subunit to the initiation
complex .
Bind i ng of AAtRNA ' s to 3 0 S subuni t s ha s been s tud ied
by Kaj i , Su zuka and Ka j i ( 1 6 7 ) and Pestka and Nirenberg ( 1 6 8 ) .

4 7
They have f ound that a t high Mg ion concentrations , speci f ic
binding of AAtRNA ' s occur s with one s ite of the subunit .
The add i tion of 5 0 S subunits ( 1 6 9 ) causes a 2 - 3 fold increase
in the observed bind i ng , pre sumably by creating an additiona l
bind i ng area . This may be an extra point of attachment of
the tRNA at the same bind ing s i te or an entirely new binding
s ite . S ince bind i ng in the presence of 5 0 S subunits has
d i f f erent metal ion cofactor requirements ( 1 6 8 ) , it most
like ly ref lec ts a new binding s i te . Matthae i and Milberg
( 1 7 0 ) have interpreted the ir results on the kinetics of AAtRNA
binding in terms of the creation of a new s ite upon addition
of 5 0 S subunits . Igarashi and Ka j i ( 17 1 ) have shown that
phe- tRNA bound to 3 0 S subunit-codon is d i s soc iated by
add i tion of 5 0 S subunits and tha t such prebound AAtRNA ' s are
not incorporated into peptides . This result indicated that
bind i ng of AAtRNA to 3 0 S subunits is non- func tiona l and
extraneous .
Recently , however , Nomura and Lowry ( 1 7 2 ) have
reinvestigated the area . They showed that f - 2 phage RNA
directs the binding of N-F-met-tRNAf to 3 0 S subunits at low
Mg ion concentrations and that such bind ing i s not stimulated
by the add ition of 5 0 S subunits . Thi s bind ing was stimulated
by the initiation factor s , f� and f 2 , and by GTP when
salt-wa shed r ibo soma l subunits were s tud ied . The viral RNA
d id not d ir ec t the binding of phe- , leu- , lys- , tyr- , or
va l -tRNA or met-tRNAro to 3 0 S subunits . These AAtRNA ' s were
bound however to 3 0 S subunits at high Mg ion concentrations ,

4 8
when appropr iate synthetic polyribonuc leotides were used .
This binding wa s s t imulated by the addition of 5 0 s subunits
a s had been prev iously reported ( 1 6 8 , 1 6 9 ) . The se results
ind icated that b i nd ing of N-F-met-tRNAf to 30 S subunits is
d i f f erent from the binding of other AAtRNA ' s . Nomura , Lowry
and Guthrie ( 1 4 8 ) in a s imi lar contrast , have repor ted that
the addi t ion of 5 0 S subunits to N-F-met-tRNAf bound to
3 0 S subunit-codon does not cause d i s soc iation of the bound
AAtRNA ; rather the N-F-met-tRNAf is now bound to 7 0 S
r ibosome s .
Nakada and Kaj i ( 1 7 3 ) had shown earl ier that native ( a s
opposed t o d i s soc iated ) 3 0 S and 5 0 S subunits do not combine
in 1 0 - 2 � Mg ion , but are able to synthe s i z e polyphenyla lanine
in the presence of poly U . The r ibosome found assoc iated
with poly U in these exper iments is a 7 0 S r ibosome . The se
resu lts indicate that mRNA catalyzed the assoc iation of native
subunits . Eisenstadt and Brawerman ( 1 7 4 ) have found s imilar
a s soc iation of native subunits i n the presence of f - 2 phage
RNA and bel ieve such monosomes are more active in prote in
synthe s i s than 7 0 S r ibosome s . They bel ieve that 7 0 S r ibosomes
must d i s soc iate pr ior to partic ipating in peptide synthe s i s .
Mangiarotti and Schles s inger ( 1 7 5 ) developed very gentle
extract ive techniques to s tudy polysome d i s tr ibution in
E . col i . With the se technique s , they showed ( 17 6 ) that E . coli .
do not conta i n free 7 0 S ribosomes but rather have a pool of
d i s sociated 30 S and 50 S subunits . Kaempfer , Me selson and
Ra skas ( 17 7 ) conf irmed the exi s tence of such a subunit pool by

4 9
growing E . c o l i i n heavy and l ight i sotope media . They found
that r ibosomes produced in heavy media d i s soc iated and
reas soc iated in very short t ime periods with r ibosoma l
subunits synthe s i z ed subsequently i n l ight media . Subunits ,
however , once synthes i z ed did not equi librate with rRNA or
r ibosoma l proteins .
Mang iarotti and Schle s s inger ( 1 7 6 ) and Nomura and Lowry
( 17 2 ) were led by their quite d i f f erent exper iments to
propose that protein synthes i s is initiated by attachment of
a 3 0 S subunit to mRNA af ter which a 5 0 S subunit is added .
H i l le , Mi l ler , Iwaki and Wahba ( 1 7 8 ) have conf irmed the
b i nd ing stud i e s of Nomura and Lowry . They a l so showed that
N-F-met-tRNA bound to 30 S subuni t s - codon ( in the presence
o f f 1 , f 2 and GTP ) wa s able to react with Pm when 5 0 S
subunits were added . Ghosh and Khorana ( 17 9 ) have studi ed
the binding of N-F-met-tRNAf to 3 0 S subuni t s in the presence
of poly (AUG) n or poly ( UG ) n . They conf irmed the bind ing ,
the requ irement for initiation f ac tor s and GTP in the bind ing
and were able to i so late the ternary complex by sucrose
dens i ty gradient c entr ifugation . The i solated complex wa s
ab�e to form a d ipeptide , N-F-met-me t , when 5 0 S subunits and
met-tRNAm were added to i t . A s imi l ar complex conta i ning 7 0 S
r ibosomes instead of the 3 0 S subunit wa s a l s o capable of
forming the d ipeptide . They a lso showed that binding of cys- ,
val-tRNA or met-tRNAm to 3 0 S subunits occurred only at high
Mg ion concentration . A complex formed at high Mg ion
concentration with 3 0 S subunits , poly (AUG ) n , N-F-met - tRNAf

5 0
and met-tRNAm wa s a l so i solated . This complex wa s not able
to form a d ipeptide upon the addition of 50 s subunits .
These results support the conclusion that only N-F-met-tRNAf
b i nd s in a func tiona l manner to 3 0 S subunits - codon .
Ohta , Sarkar and Thach ( 1 8 0 ) have observed a lower Mg ion
optimum for GTP - f l - f 2 s t imula ted binding of N-F-met- tRNAf
to 7 0 S r ibosomes ( 5m�) than to 3 0 S subunits ( 8 - 9mM) . They
postulate that this might ind icate that the 7 0 S r ibosome is
attached to codon easier than the 3 0 S subunit . They are
aware , however , that extrac tive procedure s and storage
cond i t ions play a role in r ibosomal Mg ion dependency and that
the d i f f erence in the two optima i s not large .
Step 4 . - Bind ing to the aminoacyl s i te on the 7 0 S
r ibosome .
The bind i ng of AAtRNA ' s to r ibosomes i s a wel l known
phenomena which ha s been reviewed by several author s ( 1 2 8 ,
1 8 1 , 1 8 2 ) . Exper iments designed to test for thi s binding have
been u sed to e stabl ish the genetic code ( 1 1 4 , 1 8 3 , 1 8 4 ) .
Gi lbert ( 7 1 , 1 8 5 ) , Cannon , Krug and Gi lber t ( 1 8 6 ) and Bretscher
( 18 2 ) wer e among the early worker s to establ i sh that AAtRNA ' s
were a s s oc i a ted with ribosome s dur ing protein synthe s i s in
bacter ial extra c t s . Nirenberg and Leder ( 1 1 4 ) showed that
such spec if ic binding occurred in the absence of prote in
synthes i s and developed a convenient a s say method to te st
the b i nd ing . This method , which involves adsorption of the
complex on c e l lulose ester f i lter s , has been widely used in
binding investigations . Igarashi and Ka j i ( 1 7 1 ) showed that

5 1
AAtRNA ' s bound to 7 0 S r ibo somes i n the absence o f prote in
synthes i s were i ncorporated in the NH2 -termina l pos ition of
peptid e s when prote in synthe s i s was initiated . Wettstein and
Noll ( 1 8 7 ) have employed rad ioac tive tRNA to determine that
2 or 3 AAtRNA ' s were bound per r ibosome dur ing protein
synthe s i s . Igara shi and Ka j i ( 17 1 ) have obta ined a s imilar
result with saturation bind ing curves . They a l so showed that
the aminoacyl s ite of 7 0 S r ibosome s wa s more read i ly occupied
than the peptidyl s ite by phe-tRNA in the absence of GTP and
enzyme s . Matthaei and Mi lberg ( 1 7 0 ) obta ined simi lar results
by studying the kinetic s of AAtRNA bind i ng .
Ar l i nghau s , Schaef fer and Schweet ( 1 8 8 ) have demons trated
the exi s tence of two types of AAtRNA binding to rabbit
reticulocyte r ibosome s . One mode is non-enzymatic , non-GTP
dependent , whi le the other requires the presence of GTP and
enzyme s . The exi s tence of an enzymatic and GTP-dependent
bind ing of an AAtRNA other than N-F-met- tRNA or N-acetyl-phe
tRNA ( 18 9 ) ha s been demonstrated in E . coli extrac ts , very
recently ( 1 9 0 , 1 9 1 , 1 9 2 ) . Previous fai lures to do so may
ref lect d i f f erenc e s in cha in initiation or may be the result
of inadequate r ibosome pur if ication technique s .
Moldave et . a l . , ( 1 9 3 , 1 9 4 , 1 9 5 ) have isolated complexe s
of the supernatant enzyme , Transferase I , from rat l iver
preparations and showed that thi s enzyme bound GTP and AAtRNA .
They sugge sted that thi s complex wa s involved in the binding
of AAtRNA ' s to r ibosome s . Al lende , Monro and Lippman ( 1 9 6 )
have reported the i solation from E . col i of 2 or 3 prote ins ,
Tu , Ts and G , from the non-r ibosoma l ( supernatant) por tion of

5 2
extracts which are d i stinct from the components o f the AAtRNA
synthe s i z ing system and which are necessary for peptide bond
synthe s i s . Allende and Wei s sbach ( 1 9 7 ) identif ied a complex
s imi lar to that reported by Moldave between GTP and E . coli
initiation fac tor preparations . GDP wa s able to replace
GTP in thi s complexation but ATP , CTP , UTP AND GMP were not
able to compete with GTP . The heat instabi lity and the
chromatographic behavior of the protein invo lved in the
f ormation of the complex suggested that the binding wa s with
the supernatant enzyme T . Al lende , Seed s , Conway and
Wei s sbach ( 1 9 8 ) have now reported that the or igina l initiation
fac tor preparations conta ined trace s of T and that the removal
of this protein caused a los s of bind ing activity . Separately
i so lated T enzyme wa s shown to pos s e s s the origina l
comp lexation a b i l i ty . They i s o lated a T - GTP complex on
cellulose ester f i lters .
Ravel , Shorey and Shive ( 1 9 9 ) i ndependently observed
s imi lar complexation in mixtures of GTP , phe-tRNA and super
natant factor preparations . They showed tha t GTP wa s initially
hydrolyz ed to GDP by binding to the protein and that following
this s tep , GDP wa s further bound in a complex with AAtRNA .
Gordon ( 2 0 0 ) wa s able to i sola te a complex of GTP which
wa s formed by T in the pre s ence of AAtRNA . Later , he
propo sed a two s tep bind ing scheme of GTP to T, fol lowed by
f ormation of a ternary complex with AAtRNA ( 2 0 1 ) . Lucas-Lenard
and Haenni ( 1 9 1 ) have isolated thi s same ternary complex and
report that it is capable of binding to the ami noacyl s i te

of 7 0 S r ibosome s . When highly pur if ied r ibosome s are
emp loyed , bind ing of thi s complex to r ibosome s is much
greater than bind ing of AAtRNA a lone . GDP i s not able to
subs titute for GTP i n r ibosoma l bind ing , a lthough it forms
a ternary complex with T and AAtRNA . They have also shown
that AAtRNA bound to the aminoacyl s i te in the presence of
T a nd GTP i s able to form a d ipeptide if the peptidyl s i te
ha s previou sly been occupied .
Step 5 . - Formation of the peptide bond .
53
The forma tion of the peptide bond between peptidyl-tRNA
and the cx2 -amine of the AAtRNA i s shown a s g iven by Watson
( 67 ) in F ig . V I I . Monro and Marcker ( 2 0 2 ) and Monro ( 2 0 3 )
have recently shown that N-F-met i s bound to the 5 0 S
subunit when i t i s e ster if ied with the termina l hexanuc leotide
fragment of tRNAf . They demonstrated that N-F-met bound to
5 0 S subunits i n this way i s capable of forming the model
d ipeptide , N-F-met-Pm . This Pm reaction i s not enhanced by
GTP and the b i nd i ng must be carr ied out in ethanolic solution .
The 5 0 S subunits u t i l i zed were as free of initiation and
supernatant enzymes as it is pre sently pos sible to prepare
( 4 X NH4 C l wa shed ) and the Pm reacb�dn cata lyzed by the se
subunits const i tutes good evidence that the peptide synthetase
enzyme is a part of the r ibosome .
Luc a s -Lenard and Haenni ( 1 9 1 ) report that d ipeptide but
not tripeptide formation occurs on 7 0 S r ibosomes ( 4 X NH4C l
wa shed ) between prebound N-Acetyl-phe-tRNA and added phe-tRNA .
Forma tion of thi s d ipeptide i s not enhanced by the supernatant

enzyme G . I n thi s study phe- tRNA wa s added a s the ternary
complex between phe- tRNA , GTP and supernatant enzyme T .
S tep 6 . - Pul sation (Movement ) of the r ibosome .
Wettstein and Noll ( 1 8 7 ) have proposed that their data
indicated that tRNA from the peptidyl s i te wa s transf erred
to a loo sely attached " ex i t " s ite a f ter formation of a
peptide bond . The ir proposal wa s based on the number of
5 4
tRNA molecules found a s socia ted with the r ibosome dur ing
prote in synthe s i s . Fur ther evidence concerning the exi stence
of such a s i te ha s not been pre sented to date . I ndeed ,
evidence that a spec if ic tRNA leaves the r i bosome i s not
ava i lable and probably w i l l not be ava i lable until very highly
pur if ied preparations of tRNA are obtained . Our pre sent
picture of the se steps is ba sed on the proposa l s of Wa tson
( 6 7 ) •
L ipmann e t . a l . , ( 2 0 4 , 2 0 5 ) have proposed tha t the mo st
l ikely func tion of their ribosome-dependent GTPa se , G, is to
cause the pulsation or contractile movement of the r ibosome .
They have shown that one mo lecule of GTP i s hydrolyzed for
each pept ide bond formed ( 2 0 3 ) and Luca s -Lenard and Haenni
( 1 9 1 ) have shown that this GTP hydro lysis occur s af ter
d ipeptide formation .
Step 8 . - Cha i n termina tion and relea se .
I t i s we l l known that cel l-free systems d irected by
natura l mRNA ' s release the ir peptide s into the superna tant
( 7 0 ) whi le systems d irec ted by synthetic homopo lyrner s retain
the produc t peptide on the r ibosome ( 7 1 ) . I t ha s been shown

that nonsense codon s produced in bac teriophage mutants cause
the release of incomplete proteins ( 2 0 6 ) . These codons , UAA
( ochre ) , UAG ( ambre ) , and UGA , lead to termination of peptide
cha in growth in sys tems d irec ted by mRNA ' s from the mutant
s trains or phage s . Synthetic mRNA ' s which contain these
codons a l s o terminate prote in synthe s i s ( 2 0 6 ) . Last et . a l . ,
( 2 0 7 ) have shown that the mRNA AUG-UUU- (A) n synthe s i zes the
peptide met-phe-polylys i ne whi le AUG-UUU-UAA- (A) n which i s
phased t o read the cha in termination codon UAA , d irec ts the
synthe s i s of polylys ine only . S imilar results were obtained
with o ther synthetic mRNA ' s containing termination codons .
These worker s proposed that the absence of met-phe- tRNA in
the supernatant in the exper iment u t i l i z ing AUG-UUU-UAA- (A) n
indicated that hydrolysis of peptidyl-tRNA occurred on the
r ibosome . That i s , the cha i n termination codon , UAA , not
only terminated peptide synthes i s but caused r ibosoma l
hydroly s i s of the completed peptide . Unl ike AAtRNA ' s ( 2 0 8 ) ,
peptidyl -tRNA ' s , e . g . polylysyl-tRNA ( 2 0 9 ) , are re latively
stable in the supernatant of cel l-free systems . Therefore ,
5 5
i t s eems l ikely that hydro lys is o f the ester may occur o n the
r ibosome . Kur iki and Kaj i ( 2 1 0 ) have shown tha t tRNA obta ined
by hydr o ly s i s of peptidyl-tRNA is no longer capable of
accepting new ami no ac id .
Capecchi ( 2 1 1 ) ha s i solated an R 1 7 phage mutant whose
RNA terminates peptide f ormation in its coat protein premature ly .
By contr o l li ng the supply of amino acids to a cel l-free sys tem
d irec ted by thi s mRNA he i so lated a protein , R fac tor , which

5 6
i s neces sary f o r the rej ec tion o f the peptide from r ibosomes .
This R fac tor does not catalyze hydroly s i s of peptidyl -tRNA
i n the absence of r ibosome s . The exi s tence of an R factor
has not been shown in other preparat ions and it is not known
whether it i s of general importance . Bretscher ( 2 1 2 ) has
used Capecchi ' s system to search for a nonsense tRNA . Such
a tRNA i s one which would be c apable of interacting with
the nonsense codon and thereby cause cha in termination . He
demons trated that cha in termination occur s in systems which
are uti l i z ing highly pur i f i ed tRNA ' s and proposes that i t is
very unl ikely that such a nonsense tRNA exists .
B . 6 . The Effect of Tetracyc l ines on C e l l-Free Protein
Synthes i z i ng Sys tems
In an ear ly s tudy of the mode of chloramphenicol ac tion
Rendi and Ochoa ( 2 1 3 ) mentioned , without suppor ting data ,
that oxytetracyc l ine i nhibited amino acid incorporation in
their cell-free system . They were u s i ng a c e l l - free system
directed by synthetic polyr ibonuc leotide s . Franklin ( 2 1 4 )
reported exper iments which showed that chlortetracyc l ine
i nhibited leuc ine incorporation in rat l iver and in E . coli
cell-free systems . Inhibition of the system derived from
E . coli wa s greater than that observed with the sys tem from
rat l iver . C hlortetracyc l ine caused a 9 0 % inhibition of
leuc i ne incorporation in the E . coli system at a concentration
of 4 X l o - 4� . Chlortetracycline d id not affect the formation
of leu-tRNA by e i ther extract but did prevent incorporation
of preformed leu-tRNA into protein . S imilar results were

obtained with oxytetracyc l i ne and tetracyc l ine . Laskin and
Chan ( 2 1 5 ) have shown that c e l l-free systems d irected by
5 7
poly U were i nhibi ted equa l l y b y tetracyc l i ne , chlortetra
cyc l ine , oxytetracyc l ine and 6 -demethy l - 7 -chlortetracyc l ine
but that 4 -dedimethylamino - 5 -hydroxytetracyc l i ne was only
a bout 1/1 0 as potent . They a l so reported that tetracyc l ine
d i d not affect the formation of phe-tRNA . Suarez and Nathans
( 2 1 6 ) and Day ( 2 17 ) have repor ted s imi lar i nhibi tions of
phe-tRNA i ncorporation . Suarez and Nathans ( 2 1 6 ) reported
that inhibition wa s rever sed by the addi tion of r ibosome s -
poly U but not by the add i tion of phe-tRNA or supernatant
extract . Hierowski ( 2 18 ) has shown that lys ine , proline and
phenylalanine i ncorporation i s inhibited in systems d irec ted
by synthetic polyr ibonuc leotide s . Okamoto and Mi zuno ( 2 1 9 )
and Maxwe l l ( 2 2 0 ) have reported inhibition of cel l-free
sys tems in which incorporation of a lgal protein hydrolyzate
wa s d irec ted by endogenous mRNA . Rif i no e t . a l . , ( 2 2 1 ) have
reported that s ever a l tetracyc l i ne der ivatives are ac tive
inhibitor s of poly U d irected phenylalanine incorporation but
noted that i sochlortetracyc l i ne , an inactive antibacter ial ,
wa s an inhibitor of endog enous mRNA d irected incorporation .
Because of thes e report s i t has been as sumed that tetra
cycl ines cause a genera l inhibition of amino acid i ncorporation
by c e l l - free systems , Tur ley , Thomas and Sne l l ( 2 2 2 ) have
recently i nvestigated the incorporation of 2 0 d i ff erent amino
acids i nto protei n by a c e l l -free system from E . coli d irected
by endogenous mRNA . They found that i ncorporation of 18 of

these amino acids was inhibited at low leve ls of oxytetracy
c l ine . Cys te ine and tryptophan incorporation were not
inhibited at low leve l s of oxytetracycl ine and the ir
58
incorporation was s t imul ated at high leve l s of oxytetracyc line .
The authors postulated that this s timulation may be due to
induced codon misreading or that the se amino ac ids might be
incorporated into prote in by a non-ribosomal system .
B . 6 . a . E f fects o f Tetracyc l ines in terms o f the mode l
o f Ce l l-Free Protein Synthe s i s
I nterfe rence with Step l . - B inding o f a 3 0 S subuni t
t o mRNA .
Hierowsk i ( 2 1 8 ) has reported that the inhibition by
tetracycl ines of the incorporation of AAtRNA ' s into prote in
is more pronounced in sys tems in which polyribonuc leotides
serve as the me ssenger than i n sys tems directed by endogenous
mRNA . She noted that tetracyc l ine did not change the Mg ion
optima of these system . Polypeptide synthe s i s directed by
poly C in the presence of chlortetracycl ine ( 4 5� �) was inhibited
by 7 5 % . Les s inhibition was observed when poly U ( 7 0 % ) , poly A
( 5 6 % ) , poly U§A1 ( 5 0 % ) or poly UC ( 2 5 % ) were used as me s sengers .
I t was mentioned that the binding of poly U to ribosomes was
not a ffected by tetracy c l ine . Suare z and Nathans ( 2 1 6 )
f · d h · 1 · · · 1 4c 1 con l rme t l S resu t ln an experlment us lng -po y U .
Connamacher and Mandel ( 2 3 3 ) have also reported that tetracycl ine
did not prevent the formation of ribosome - codon complexe s .
They showed that tetracycl ine was , however , ab le to bind to
poly U in EDTA smlution s . Day ( 2 1 7 ) has reported binding to

5 9
poly U and poly A i n the absence o f Mg ion a s wel l a s a t
severa l Mg ion concentrations . Kohn ( 2 2 4 ) has reported that
metal ions are nec e s sary for binding to DNA . Maxwel l ( 2 2 5 )
ha s recently shown that cultures o f B . megater ium inhibited
by tetracyc l ine are able to form polysome s . The se results
constitute a body o f evidence that tetracyc lines do not inter
f ere with the attachment of mRNA ' s to r ibosome s , but that the
par ticular mRNA used in a c e l l-free system does af fect the
degree of inhibition produced by tetracyc l ine s .
I nter f erence with Step 2 . - Formation of the initiation
complex .
Exper iments des igned to test the hypothes i s that
tetracyc l ine inhibits the f ormation of the initiation complex
have not been performed . However , exper iments have been
performed which sugge s t that tetracyc line does not interfere
with binding to the peptidyl s ite on 7 0 S r ibosomes .
Gotte sman ( 2 2 6 ) ha s recently stud ied the binding of polylysyl-
tRNA to r ibosome s . I t i s be l i eved that thi s peptidyl- tRNA
e ither b i nd s d irectly to or i s rapidly trans located to the
peptidyl s ite on 7 0 S r ibosome s . Ribosoma l bound polylysyl
tRNA i s c apable of reacting with puromyc in ( Pm ) to form
polylysyl-Pm ( 2 2 6 , 2 2 7 ) and thi s produc t i s released from the
r ibosome under certain c ircumstance s . Chlortetracyc l ine does
not prevent the bind i ng of this peptidyl- tRNA tm r ibosome s
( 2 2 6 , 2 2 8 , 2 2 9 ) nor does i t prevent the reac tion or release
of bound polylysyl -tRNA with puromyc in . I f r ibosomes with
prebound polylysyl-tRNA are mixed with lys-tRNA in the absence

6 0
o f GTP and superna tant enzyme s , a s ing le lys ine is added to
the polylysyl on the r ibosome ( 1 9 1 ) . Tetracyc l ine prevents
thi s add i tion whi le chloramphenicol does not prevent thi s
s ing le peptide bond formation . S imi larly , r ibosome s containing
prebound N•acetyl-phe-tRNA ( a presumed initiator of poly u d irected systems ) react with Pm and release N-acetyl-phe-Pm
into the supernatant . Chlortetracyc l ine doe s not prevent
e i ther the reac tion o£ the relea s e . Chlortetracyc l ine ,
however , does par t i a l ly prevent the addi tion of phe-tRNA to
r ibosomes conta ining prebound N-acetyl-phe-tRNA . The phe- tRNA
which i s bound to the ribosomes i s able to f orm a d ipeptide
with the N-acetyl-phe-tRNA .
The above results are consi s tent with the general idea
that tetr acyc l ines do not af fect AAtRNA bind i ng to the peptidyl
s ite but rather they inhibi t bind i ng of AAtRNA ' s to the
aminoacyl s ite . D irect evidence for this type of inhibition
is pres ented in the f o l lowing sections .
Interf erence with S tep 3 . - Attachment of 5 0 S subunits
to the Initiation complex .
Thi s s tep ha s not been studied direc tly , but Day ( 2 17 )
ha s shown that i so lated 3 0 S subunits containing bound
tetracyc l ine are able to f orm 7 0 S r ibosomes upon the add i tion
of 5 0 S subunit s .
Suzuka , Kaj i and Kaj i ( 1 6 9 ) and Vazquez and Monro ( 2 3 0 )
have shown that tetracyc l ine prevents the binding o f AAtRNA ' s
to 3 0 S subunit s - codon and a l so prevents the s timulation
of bind i ng observed when 5 0 S subunits are added . Binding of

6 1
AAtRNA ' s t o 3 0 S subuni t s - codon ha s now been shown t o be
non-functiona l unless the AAtRNA i s N-F-met-tRNAf . I t is
gowever , generally agreed that the stimulat ion of binding
observed when 5 0 S subunits are added i s due to the creation
of the ami noacy l s ite .
I t seems unlikely that tetracyc l ines interfere with
step 3 if i ndeed such a s tep is i nvolved in prote in synthesis .
I nterference with Step 4 . - Bind i ng to the aminoacyl s i te
of r ibosome s .
Hierowski ( 2 18 ) u s ed sucrose den s i ty gradient centr ifuga
t ion to show that binding of phe-tRNA to 70 S r ibosomes-codon
i s partially inhibited ( 6 0 % ) by chlortetracyc l ine . The se
stud i e s were conduc ted at 1 2 mM Mg ion and it is probable that
phe-tRNA i s c apable of enter ing the peptidyl si te as wel l a s
the ami noacyl s ite under the se c ircums tances . Suare z and
Nathans ( 2 1 6 ) have shown that the bind ing of N-acetyl-phe-
tRNA at 2 0 mM Mg ion is inhibited by chlortetracyc l ine . The
maximum inhibition obta i nable at very high tetracycline
concentrat ions wa s 5 0 % . C lark and Chang ( 2 3 1 ) observed s imi lar
i nhibi t ion ( 4 0 % ) of AAtRNA binding to rabbit reticulocyte
r ibosomes i n the presence of the transfer enzyme s . They a l so
showed that tetracyc l ine d id not i nterfere with the puromyc in
reaction in systems synthes i z ing protein . Vazque z and Monro
( 2 3 ) have noted inhibi t ion of poly A directed lys- tRNA ( 5 1 to
6 1 % ) a s wel l a s poly U d irected phe-tRNA binding ( 3 8 to 5 2 % ) .

Because of these reports , i t ha s been genera l ly as sumed
that tetracyc l ine s prevent binding of AAtRNA ' s to only one
s ite on the r ibosome - codon . S i nce these experiments were
carr ied out under condi tions which favor binding to the
ami noacyl s ite , these i nve stigators have proposed that this
6 2
i s the s ite of tetracyc l ine action . The recent reports of
Gottesman ( 2 2 6 ) and Lucas-Lenard and Haenni ( 1 9 1 ) , mentioned
ear l ier , have substantiated thi s conc lus ion . I t should be
noted , however , that Wolfe and Hahn ( 2 3 2 ) were only able to
demons trate a very sma l l inhibition of phe-tRNA binding to
r ibosome s - poly U by sucrese dens i ty gradient centrifugation .
S imilar ly , La skin ( 1 6 ) ha s s tated that he wa s only able to
demonstrate a sma l l inhibition ( 3 0 % ) of thi s binding in
exper iments util i z i ng the technique of c e l lulose ester
f i ltration . He emphas i zed the poor quantitative nature of
these experiments . I t would seem , that the conc lusive
experiment ha s not as yet been performed .
I nterference with Step 5 . - Formation of the peptide
bond .
I n the pre sence of tetracyc l ine , the abi l i ty of prebound
polylysyl-tRNA to react with Pm has been demonstrated by
Gotte sman ( 2 2 6 ) . Thi s result and the abi l i ty 0f prebound
N-acety l-phe-tRNA to react with either Pm or bound phe- tRNA
in the presence of tetracyc l i ne , demonstrated by Luca s-Lenard
and Haenni ( 1 9 1 ) , i nd icates that s tep 5 is not inhibited .

6 3
I nterference with S tep 6 . - Pul sation (Movement) of the
Ribosome .
S i nce i t i s thought that release of Pm - peptides occurs
by norma l ribosomal pul s ation and thi s is not inhibited by
te tracy c l ine , i t is unl ikely that te tracyclines inhibi t this
s tep in protei n synthe s i s .
I nterference with Step 8 . - Chain termination and release .
E f fects o f tetracycl ine on thi s step have not been studied .
B . 6 . b . Binding o f Tetracycl ines to Ribosomes . -
The pre sumed mode of tetracyc line action es tablished by
these exper iments i s that of a d i rect competition with
AAtRNA ' s for the aminoacyl b inding s i te on 70 S ribosomes . I t
i s unl ikely that tetracy c l ine inhibi tion i s due to a n indirect
e f fect caused by binding of tetracycline to some other ribo
s omal s ite , s ince a l l other ribosomal functions appear normal .
Direct binding to the aminoacyl s i te has , of course , not
been demons trated . Connamacher and Mandel ( 2 2 3 ) have , however ,
shown by equ i l ibrium dialys is and by sucrose dens i ty gradient
centri fugation , that tetracycl ine is bound to 70 S E . coli
and B . cereus ribosome s . When experiments were conducted at
low Mg ion concentrations , the tetracycl ine was largely
associated with the 3 0 S subuni t . Ribosome s prepared from
preincubated extracts , so that endogenous mRNA activity was
low , showed a reduced abi l i ty to bind tetracyc l ine . Las t ,
I z ak i and Sne l l ( 2 3 3 ) were unable to show any binding of
tetracycl i ne to a ribosome sediment produced by centri fugation
in sucrose . Thi s failure may be due to a l ack of endogenous

6 4
mRNA in their preparations or to the ir sed imentation technique
which may require s tronger binding than bind i ng demonstrated
by sucro se density grad ient c entr ifugation . Day ( 2 3 4 ) ha s
substantiated the f i nd ing s of Connamacher and Mandel ( 2 2 3 )
that tetracyc l ine i s a s sociated with the 7 0 S r ibosomes
dur i ng sucrose density gradient centr ifugation . He showed
that tetracyc l ine can bind to e ither 3 0 S or 5 0 S subunits .
Tetracyc l i ne , that i s bound to e ither of the se subunits i s
able t o d i s soc iate and rebind t o the other subunit if i t i s
added . A l l o f the sucros e den s i ty gradient centrifugations
repor ted by Day �ere conducted on r i bosome - tetracyc l ine
migtures that had been subj ected to extensive d ialysis pr ior
to centr ifugation . Day calculated that approximately 1 mole
of tetracyc l i ne is a s sociated with 1 mole of 70 S ribosome s
and that 1 mole of tetracyc l ine i s a s sociated with 2 mo les
of e i ther 30 S or 50 S subunits . He a l so showed that ribosome s
contai ning bound tetracyc l ine were not able to bind phe-tRNA
or incorporate phe-tRNA i nto prote in af ter addition of
supernatant enzyme s . Day ( 2 3 5 ) later showed that the cellu lose
ester f i l tration technique a l so indicated that tetracyc l i ne
bound to r�bosome s .
Recently , Maxwel l ( 2 3 6 ) has shown that large quantities
of tetracyc l ine are found a s sociated with the sediment of
und i a ly z ed r ibosome - tetracycl ine mixtures a f ter high speed
centr i fuga tion . He found about 3 0 0 molecules of rever sibly
a s s oc iated tetracyc l ine per 70 S r ibosome . I t i s d i f f icult

to say if the tetracycl ine a s soc iated with the r ibosomes
under these c ircums tanc e s is the result of binding or of
entrapment . However , if the se sediments were subj ected to
dialysis or an addi tional centr ifugation through sucrose
6 5
and then binding s tudied by sucrose dens ity grad ient
centr ifugation , a sma l l amount of tetracycl ine remained with
the polysome s or r ibosome s . Sedimentation at low Mg ion
concentrations revea led that mos t of the tetracyc l ine was
a s sociated with the 3 0 S subunits .
In the l ight of these experiments , i t seems l ikely that
tetracyc l ine does bind to r i bosome s , probably the 3 0 S
subuni t , and that such binding i s capable of preventing
attachment of AAtRNA ' s to the aminoacyl s ite .

I I I . EXPERIMENTAL
A . Mater i a l s and Equipment
A . 1 . Tetracyc l i ne Der ivativ e s
6 6
The antibiotic s used i n this study were the generous
g if t s of Dr . James H . Boothe , Leder le Laborator ies , Pear l
River , N . Y . , and Dr . Char les R . Stephens , Charles Pfi zer
Laborator i e s , Groton , Conn . . They have been identif ied by
their ultraviolet absorption spectra . The wave lengths of
several absorption maxima and the absorptiv i ty at these
maxima are g iven i n Table I . The spectra were obtained from
solutions buf f ered a t pH 6 . 6 5 with phosphate buffer ( 0 . 2 M) .
Solutions of the samples of 7 -amino - 6-demethyl-6 -deoxy
tetracyc l ine and 9-nitro- 6 -demethy l - 6-deoxy-tetracyc line had
" shoulder s " in their absorption spectra at approximately
4 0 0 mp . Thes e solutions were separable into two spots by
thin layer s i l ica gel chr omatography . The solvent system for
this chromatography was a mixture ( 2 : 2 : 3 : 3 ) of phosphate
buf f er ( 0 . 2 �' pH 6 . 6 5 ) , ethanol , butanol and ethyl acetate .
It seems l i kely that the se samples contain trace quantities
of 5a ( 6 ) -anhydrote tracyc l i ne s , however , they were used without
further puri f ication .
A . 2 . Escherichia coli
E scher ichia c o l i W wa s obtai ned as a frozen pa ste . 2 I t
was cultured over several growth cyc les in broth and on agar
plate s . An i solate from a s ing le colony was used to inoculate
2Genera l B iochemica ls , Chagrin Fal l s , Ohio .

Tab le I . - The Tetracyc l ines .
Tetracyc l ine Ana loga pKa Va luesb Ultraviolet Abs orpt ion Spectrac
max .'1 m}l Abs orpt ivity x lo-4
7 -N itro-dm-do-tetracyc l ine 360 , 2 6 5 l . 3 , 1 . 6
7 -Chloro-dm-tetracyc line 3 . 3 , 7 . 3 , 9 . 3 ( 2 3 7 ) 3 6 7 , 2 7 5 1 . 4 , 1 . 5
7 -Chlorotetracyc l ine 3 . 3 , 7 . 4 , 9 . 3 ( 2 3 7 ) 368 , 2 7 5 l . l , 1 . 4
Tetracycl ine 3 . 3 , 7 . 7 , 9 . 5 ( 2 3 7 ) 3 58 , 2 7 5 1 . 5 , 1 . 5
5 -Hydroxytetracyc line 3 . 6 , 7 . 4 , 9 . 1 ( 2 38 ) 3 5 5 , 2 7 5 1 . 4 , 1 . 5
7 -Amino-dm-do-tetracyc l ine 400 , 340 , 2 7 8 0 . 4 , l . l ,
9-Amino-dm-do-tetracycl ine 344 , 2 8 2 l . 3 , 1 . 2
drn-do-tetracyc l ine 3 . 5 , 7 . 9 , 9 . 7 ( 17 ) 348 , 2 7 3 l . 3 , 1 . 7
a . dm-do- is 6-demethy l-6-deoxy - , while dm- is 6-demthyl-6-epihydroxy- . b . Numbers in parenthes is indicate l iterature referenc e . c . Values g iven were obta ined in 0 . 2 M phosphate bu f fer , pH 6 . 6 5 �
a Mode l DK�2 Ratio Recording Double Beam Spectrophotometer , Beckman Instruments , Palo Alto , Cal i f . was used .
1 . 4
0'\ -...)

Tab le I . - ( c ent . )
Tetracyc l ine Ana log pKa Va lues
7 -Brorno-drn-do-tetracyc l ine
9-Nitro-drn-do-tetracyc l ine
4 -Dedirnethy l arnino-tetracyc l ine 5 . 97 , 8 . 56 ( 1 3 )
9 -D irnethy larnino-drn-do-tetracyc l ine
5a ( 6 ) -Anhydrotetracyc l ine
1 2 a-Deoxytetracyc l ine
Tetracyc l ine rnethiod ide 3 . 56 , 7 . 80 ( 1 3 )
5a ( l la ) -Dehydroc nlorotetracyc l ine
7 -Chloro- i s otetracyc l ine 3 . 9 , 6 . 7 , 7 . 9 ( 2 3 8 )
2 -Cyano- 2 -decarboxarnidotetracJClme
Ultraviolet Absorpt ion Spectra
max . rnu Absorpt ivity x lo-4
3 50 , 2 7 2 0 . 7 ' 1 . 4
4 1 5 , 3 54 ' 2 8 5 0 . 4 , 0 . 7 , 1 . 1
360 , 2 7 2 l . l , 1 . 3
34 6 , 280 0 . 8 , 0 . 7
4 2 5 , 2 68 1 . 2 , 5 . 0
4 5 0 , 3 3 0 , 2 6 5 0 . 2 , l . 3 , 1 . 7
3 5 0 , 300 , 2 7 5 l . 6 , 1 . 4 , 1 . 6
408 , 2 7 0 1 . 1 , 2 . 1
344 , 288 0 . 3 , 1 . 7
360 , 2 7 5 0 . 9 , 1 . 0
0"1 (X)

a nutrient agar s lant . This s l ant was s tored at 3 ° C and
maintained by monthly trans fers . I t served as the source
of the tes t organ i s m . For experiments in dextrose-salts
broth another s lant was prepared from organisms which had
grown through several cycles in this broth .
A . 3 . Culture Broths
6 9
The culture broth used for activity determinations was
a high peptone media containing in a volume of l L . , 40 g .
casein hydrolysate ( GB I # 2 0 ) 2 , 1 5 g . yeast autolysate ( GBI # 1 2 0 ) 2 ,
3 g . K2 H P043 and 0 . 7 g . KH2 Po4 . 3 The pH o f thi s broth was
constant throughout the experiments at 6 . 6 5 . The divalent
cation content of the broth was estimated to be equivalent
to about l o - 3 M calcium ion by means of a divalent cation
e lectrode . 4 For exper iments at pH 6 . 2 or 7 . 4 , the broth
contained in addition to the peptones either 0 . 5 g . K2HP04 and
2 . 5 g . KH2Po4 or 2 1 . 8 5 g . K2 HP04 and 1 . 7 g . KH2P04 , respective ly .
S imilar divalent cation concentrations were observed in the se broths .
The dextrose-salts broth ( pH 6 . 7 ) contained in each l iter ,
1 0 g . dextrose , 3 g . (NH4 ) 2 so4 , 6 g . K2HP04 and 8 g . KH2P04 .
A l l broths were f i l tered through ce l lulose ester membranes 5
( 0 . 2 2 p dia . pores ) and autoclaved prior to use .
3Al l unspeci fied chemicals were analytical reagent grade .
4orion Re search I nc . , Cambridge , Mas s . 0 2 1 3 9
5Mi l l ipore Corporation , Bed ford , Mas s .

A . 4 . Cul ture Turbidity and Colorimetry
A d i f f raction grating color imeter6 equipped with a red
sensitive phototube in conj unction with matched l/2 inch
square cuvettes was used . Culture turbidity was measured
at 6 5 0 mp . Absorbance o f orcinol reaction mixture s was
measured at 6 5 0 mp whi l e that of Folin-Lowry reactions was
measured at 7 5 0 mp .
A . 5 . Total Ce l l Counts
An electronic particle counter7 was used . The counter
was equipped with a 3 0 p orifice and two threshold pulse
70
height analy zers . Ins trument s ettings whi ch gave satis factory
results were :
1/ Aperture Current , 0 . 7 0 7 ; 1 /Amp l i fi c ation , l/4 ;
Matching Switch , 3 2 H ; Gain , 1 0 0 ; Lower Thre shold , 1 0 and
Upper Thre shold , o f f . These settings were es tab l ished by s i ze
frequency analy s i s of latex suspens ions o f known particle s i ze
as wel l as suspensions of E . col i W .
Cel l s were counted in a s a l ine - formal dehyde solution
which had been f i l tered to remove particulate contamination .
Background counts in thi s fluid were 2 0 0 to 6 0 0 partic les
per 50 l ambda . The formaldehyde concentration in the counting
fluid was e i ther 0 . 2 or 1 % . The h i gher concentration gave
constant values for total ce l l s over a 2 4 hour period whi l e
6 spectronic 2 0 , Bausch and Lomb , Roches te r , N . Y .
7coulter Counter Mode l B , Coulter E lectronics , Inc . Hialeah , F l a .

the lower concentration a l lowed some growth ( 1 to 5 % ) in
the f ir s t ten minutes a f ter samples were removed from the
culture .
B . Methods
B . 1 . Escherichia c o l i Growth
7 1
A broth culture was a l lowed t o grow for 1 2 hour s at
3 7 . 5 °C . A d i lution of this cul ture , whose growth was
measured turbidimetr ica l ly , wa s a l lowed to grow into the log
phase . When a concentration of 1 x 1 0 8 organisms/ml . wa s
obtai ned , 2 ml . samples were used to i noculate nine replicate
1 0 0 ml . volume s of broth conta ined in loosely capped 5 0 0 ml .
f la sks . The f la s k s were sfuaken i n a constant temperature
water bath8 at 3 7 . 5 ° C . Freshly prepared antibiotic solutions
were added i n 1 ml . volumes to 8 of the 9 repl icate cultures
af ter 2 , 7 0 0 s econds of growth .
Exper iments d e s igned to s tudy time of mnset of inhibition
were performed with culture s of e ither 2 0 0 ml . per 1 L . f la sk
or 3 5 0 ml . per 2 L . f la s k .
B . 2 . Viable Count Method
One ml . sampl e s of the cultures were obta ined at 9 0 0 or
1 , 8 0 0 s econd i nterva l s . A minimum of 8 samples was obtained
from each culture . Sample s were d iluted with ster i l e 0 . 9 %
sal ine so lution so that 1 0 0 to 2 0 0 organisms/ml . would result .
One ml . of this aaspension wa s pipetted onto each of 5 agar
plate s . Agar plates had previou sly been made by add i ng
8American Instrument Company , S i lver Spring , Md .

approximately 1 5 ml . of s terile mel ted Peptone-Casein Agar
U . S . P . 9 to s terile plastic petri dishes ( 1 0 0 x 15 mm . ) .
After spreading the bacterial suspension over the surface of
the plate , approximately 5 ml . o f warm agar was added . When
the agar had solidified the p l ates were incubated at 3 7 . 5 ° C .
The colonies which were visible a fter 3 6 to 4 8 hours of
incubation were counted with the aid o f an e lectronic
reg i s ter . 1 0 The mean number of colonies v i s ible on the
5 plate s from e ach s ample was used to calculate the number
o f v i ab le organisms in the original culture .
B . 3 . Total Count Method
One ml . culture s amples were obtained at the same time
7 2
a s the s ample s ased for viab le c e l l counts . They were placed
in· bottles containing appropriate volume s of the s al ine -
formaldehyde counting f luid so that a concentration o f 1 0 , 0 0 0
to 3 0 , 0 0 0 organisms per 5 0 l ambda was obtained . The mean o f
.4 counts per s amp l e was corrected for background noise , but
not coincidence and used to calculate the total number of cel l s
i n the original culture .
B . 4 . Nuc leic Acid and the - P rotein Determinations
Appropriate volumes of cul ture ( 0 . 5 to 25 ml . ) were f i l tered
under vacuum through 25 mm . diameter cel lulose e s ter membranes
( 0 . 2 2 )1 di a . pores ) which had previous ly been treated to
extract detergent . Culture s amples were obtained at times
s imi lar to those at which viable and total cel l counts were made .
9 Fisher Scienti fic , Fairlawn , N . J . l O 0 0 1 C I t t 0 0 0 f f 1 N Y Amerl can Optlca ompany , ns rumen DlVlS lon , Bu a o , . .

7 3
I t wa s necessary t o r emove the detergent impregnated
in these membranes ( 2 3 9 ) s ince i t reacted with the orc i nol
reagent a nd gave high blank value s , if not removed . Detergent
extra c t ion was accomp l i shed by suspendi ng f i lters in cold
d i s t i l led water ( 4 L . per 1 0 0 f i lter s ) and then boi l i ng the
suspens ion for 1 5 m i nute s . Thi s proc e s s was repeated 3 t imes
w i th f r e sh cold water . F i l ters treated i n thi s manner were
s tored i n d i s t il led water for t ime s no longer than 4 8 hour s .
These f il ter s were able to r e t a i n E . coli qua n t i ta tive ly and
gave m i n imal a nd constant blank reactions with the orc inol
reagent .
Af ter culture samples were collec ted on the f ilter s ,
the r e t a i ned bacter i a wer e wa shed with 0 . 9 % s a l i ne solution
under v acuum . The washed s amples were prec ipita ted by
layer i ng , without v acuum , 2 ml . of t richloracetic acid (TCA)
over the f il ter s for 5 minute s . TCA soluble material was
r emoved by app l ic a tion of vacuum a nd 2 ml . of 9 5 % ethanol
l ayered over the f i lters for 5 m i nu te s . After remova l of the
ethanolic extrac t , the f i lters with retained nuc leic acids
and proteins wer e placed i n screw cap test tubes . Two ml . of
5 % TCA was added a nd the tubes were placed i n a boi l i ng water
bath for twe nty minutes .
Nuc leic Ac id DeterminatieftB
One ml . a liquot s of the hot-TCA soluble mater ial were
r emoved from the cooled tube s . Three ml . of the orc i no l
reagent wer e added a nd after heat i ng i n a boi li ng water bath
for 2 0 m i nutes , the absorption at 6 5 0 � of the cooled solution

was mea sured . This absorption wa s corrected for that
produced when an equal volume of s teri le broth wa s appl ied
7 4
to the f ilter s instead o f culture . I t was neces sary to prepare
separate blank reactions for each lot of f il ter s and each lot
of broth . The orc i no l reagent wa s pur if ied , mixed and used
accordi ng to the method s of Schneider ( 2 4 0 ) . It i s a
solution of recrysta l l i z ed orc inol ( 0 . 6 6 % ) , FeC l 3 ( 0 . 3 3 % ) and
5% TCA in concentrated hydrochloric acid .
The quantity reported a s " Nuc leic Ac id s " i s the corrected
absorbance due to a 1 2 . 5 ml . culture sampl e in an orc inol
reac tion volume of 4 ml . S ince only the logari thm of a
quantity proportional to nuc leic a c id concentration was needed
to calculate g eneration rate constants the absorbance was
used directly rather than a proportiona l quantity obtained
by r eference to a s ta ndard curve . A s tandard curve for the
a s say procedure was prepared by f i lter i ng var iou s volume s of
cultur e . An example is shown in F ig . VI I I . I t and other
standard curves prepared with E . c o l i extr ac ts were l inear
throughout the tested rang e s . Standard curve s were also
prepared with highly polymer i zed yea s t RNAl l and also with
s a lmon sperm DNAl l . They were found to be l inear throughout
the tested conc entration range (as high as 6 0 mcg . /ml . of
e ither RNA or DNA) . The RNA had an absorptivity of 0 . 0 2 3 em .
2/mcg . and a negative i ntercept of 0 . 0 4 2 whi le the DNA had
an absorptivity of 0 . 0 0 4 cm . 2/mcg . Mixture s of DNA and RNA
produced absorbanc es equivalent to those ca lculated from the
l lcalbiochem , Box 5 4 2 8 2 , Los Angel e s , California

7 5
g., 0 0 . 6-Lf) r--
0 0 . 5 -
g., 0 0 . 4 -Lf) 1.0
• 0 . 3 -
+l cU
Q) 0 . 2- • •NUCLE IC AC IDS•
() !=! 0 •PROTEI N S • cU ..0 D . l-)...! 0 Ul ..0 �
l 2 3 4 5
Volume o f C e l l s F i l tered , ml .
F igure VI I I . Cal ibration Curves for " Nuc l e ic Ac id s "
and " Prote i n " Determinations . The amounts o f
"Nucleic Ac ids " o r " Protein" found b y the a s says
are l inear ly related to the amount of culture
f i l tered .

7 6
sum o f the two Beer ' s law equations .
Maa �e and K j eldgaard ( 2 4 0 ) have estimated the DNA : RNA
ratio i n c e l l s a s a function of growth rate . They f ind this
ratio to be about 1 : 1 0 in rapidly dividing c e l l s . While in
s lowly d ividing c e l l s the ratio of DNA : RNA was reduced to
about 1 : 4 . U s ing the se f igur e s and the absorptivities for
yeas t RNA and salmon sperm DNA g iven above one can ca lculate
that the contribution of DNA to the quantity cal led " Nuc leic
Acids " in thi s work rang e s from about 2 to 5 % . S ince thi s
i s the case it might have been more proper t o report the
results a s " Ribonuc leic Acid s " rather than " Nuc leic Ac ids "
but s ince there i s a contribution to thi s quantity by DNA in
thes e exper iments the more g eneral term wa s preferred .
Another reason for preferring this term i s that in tetracyc l ine
i nhibited c e l l s the DNA : RNA ratio might be dif ferent than
that found in norma l c e l l s . v � The results of Cerny and
Habermann ( 3 3 ) , however , indicate that thi s ratio does not
change very muc h .
Prote in Determinations
The TCA solution was completely removed from the tube s
containing the f ilter s , by vacuum aspiration . The f i lter s
with retained protein precipitate were covered with 1 . 5 ml�
of a trypsin solution- ( 1 8 ) 10 mcg . /ml in 0 . 0 0 5 � NH40H and
0 . 0 5 � (NH4 ) 2C03 - . The suspended f i l ter s were stirred
vigorou s l y and a llowed to digest for 2 hours ( the same results
were obtained af ter a s much a s 48 hour s of digestion) . I t wa s

7 7
nece s sary t o utili z e tryps in digestion rather than s trong
a lka l i , because the c e llulose e s ter membrane s were hydro
l y z ed and gave very high b la nk reactions in the pre sence of
s trong alka l i . The resulting solution wa s u sed a s a sample
for protein determination by the Folin-Lowry method . The
r eagents were added directly to the tubes and the absorbance
of the colored solutions mea sured at 7 5 0 rnp . B lank reac tions
were prepared by f i l ter ing s teri le broth and u sed to correct
the absorbance of the samp le s . The Fol in-Lowry reagents were
prepared , standardi z ed and used according to the methods of
Layne ( 24 2 ) . A solution of cupric sulfate ( 0 . 0 1% ) , sodium
tartrate ( 0 . 0 2 % ) and sodium carbonate in 0 . 1 N NaOH was added
to the prote i n solution ( 5 ml . per tube ) and a f ter 10 minutes ,
0 . 5 m l . of 1 . 0 N Folin-C iocal teu phenol reagentl 2 was added .
The quantity reported as " Prote i n " i s the corrected
absorbance due to a 2 5 ml . sample of culture in a f inal
reaction volume of 6 . 5 m l . A s in the previous ly mentioned
case of " Nuc leic Ac ids " absorbance wa s used directly rather
than a proportional quant i ty obtained from a calibration
curve . A standard curve for the a s say procedure wa s prepared
by f il ter ing various volnmes of E . coli culture s . An example
is shown in F ig . V I I I . I t and other s tandard curves prepared
with E . coli extract s were linear throughout the tested range s .
I t i s , however , we l l known ( 2 4 2 ) that the Folin-Lowry reaction
1 2wi l l Scientif ic I nc . , Baltimor e , Md . 2 1 2 2 4

7 8
-�oe s not a lway s fol low Beer ' s law . Standard curves were also
prepared with bovi ne serum albuminl l and pancreatic trypsin . ll
Adherence to Beer ' s law wa s observed i n the case of the a lbumin
i n the tested concentration range (as high as 45 mcg . /ml . )
whi l e Beer ' s law was only followed for tryp s in concentrations
of l e s s than 2 5 mcg . /ml . The a lbumin had an absorptivity of
0 . 0 0 8 cm . 2/mcg . whi l e the initial absorptivi ty of the trypsin
was 0 . 03 4 cm . 2jmcg . The dec i s ion to uti l i z e the absorbance
of Folin-Lowry reaction mixtures d irectly in the calculation
of the term cal l ed " Protein" wa s based , therefore , f ir stly
on the fact that the avai lable standard curves with E . coli
extracted by the exper imental method were l inear and secondly
on the rea l i zation that conversion of absorbance to concentra
tion of some s tandard protei n would be val id only if the
s tandard protei n were the same , in respect to non-linear i ty of
absorbance and absorptiv i ty , as the experimental ly measured
proteins . Such standard proteins were not avai lable .

IV . RESULTS AND DISCUSS ION .
A . E ffect o f Tetracy c l ines on Rate Cons tants for Ce l l
Divis ion .
I t has previou s l y been s hown ( 2 2 ) that the e f fect o f
7 9
low concentrations o f tetracy c l ine on an exponentially dividing
culture is to cause a reduction i n the generation rate constant ,
that i s :
[ N ] Eq . ( 1 )
When the t ime , t , i s greater than 3 0 minute s after ant ibiotic
add i t ion and where [N° ] is the concentration o f cells in the
culture 3 0 minutes a fter antibiotic add i tion , N is the con
centration of ce l l s at any later time , ked is the generation
rate cons tant for ce l l d iv i s ion in the presence of antibiotic
and k�d is the generation rate cons tant i n the absence o f
antibiot i c . Generation rate con s tants f o r c e l l divis ion ,
ked ' o f cultures i nh i b i ted by 1 8 tetracycl ines have been
measured by v i able , kv , and by tota l , kt , c e l l count methods .
Typical results o f the s e exper iments are shown in Fig . I X and
F i g . X . These exper iments can a l l be quanti tative ly de scr ibed
by Eq . ( 1 ) . Generation rate constants obtained from the f i t
o f t h i s exper imental data t o Eq . ( 1 ) as we l l as data t o be
presented l ater are given in Tables I I and I I I . Te sts for
the f i t of e xperimental data to Eq . ( 1 ) were found to be highly
s igni f icant in cases where data obtained at time s greater than
30 minutes but l e s s than 4 hours after antibiotic addit ion
were used .

8 0
'J, -
6 -.--l E -......
\D 5 - • I 0 .--l
:><: 4 -UJ E UJ • +
-� 3 - �ICH,Ia s:: 0-(lj � co� tJ> 0 0 )..j 0
2 - AN TIBIOTIC COliC . •
Q) W/L X 10 o.oo 0 s::
...:I 2.34 • 1 - 4.87 •
7.01 ... • . 34 •
l 2 3 4
T ime , hr .
F igure I X . An Example of Viable Cell Generation Rate
Curve s . The logarithm of the number of viable cells a s
a func tion of time of growth in culture s with var ious
concentrations of 9-Nitro-dm-do-tetracyc l ine . The
s lope s , kv , are g iven in Table I I I .

6 -
,.., 5 -E1 ""-.
\!) I 0 ,.., 4 -X rn + E1 rn 3 -
���·
·rl c
OHHa Cll 01 0 0
lo-l 0 2 - AH TIBIOTIC CONC. ___.
M/L )( 1) 6 Q) 0.00 0
c H 2.34 ...
l- 4.87 e 7.01 A 8. 34 •
l 2 3 4
Time , hr .
F igure X . An Example of Tota l C e l l Generat ion Rate
Curve s . The logar i thm of the total number of c e l l s a s
a func tion o f time of growth in cultur e s with var ious
concentrations of 9 -N itro-dm-do-tetracyc l i ne . The
s lopes , kt , are g iven in Table I I I .
8 1

Tab le I I . - F irst Order Generat ion Rate C onstants ,
- 1 4 sec . x 10 , for E . c o l i W Cu ltures Inh ibited by
var ious Tetracyc l ine Ant ib iotic s , M x 106 .
M
Tetracyc l ine
0 . 000 5 . 57 5 . 8 5
0 . 599 3 . 80 4 . 4 6
0 . 898 3 . 2 3 2 . 97
1 . 198 2 . 27 1 . 96
1 . 4 9 7 1 . 4 9 1 . 7 6
1 . 7 96 1 . 10 1 . 2 5
7 -Chlorotetracyc l ine
0 . 000 5 . 4 1 5 . 94
0 . 24 6 4 . 8 2 4 . 5 1
0 . 4 9 2 3 . 48 3 . 3 3
0 . 7 88 2 . 3 3 2 . 17
0 . 98 5 2 . 16 1 . 84
1 . 2 3 1 0 . 5 2 0 . 9 3
8 2
5 . 07
3 . 8 5
2 . 89
2 . 10
1 . 4 5
1 . 2 7
5 . 80
4 . 97
3 . 5 3
2 . 14
2 . 16
0 . 5 1

Tab le I I . - ( c ont . ) 8 3
M kv kp kna -, 5 -Hydroxy tetracyc l ine
0 . 000 5 . 7 5a 5 . 7 9 5 . 97
0 . 5 3 7 4 . 7 1 3 . 4 9 3 . 7 5
0 . 87 8 4 . 04 3 . 4 7
0 . 87 8 3 . 7 5b
1 . 07 3 2 . 2 5 2 . 24 2 . 14
1 . 3 7 6 2 . 2 3 2 . 24 2 . 4 0
1 . 3 7 6 2 . 2 0
7 -Chloro-dm-tetracyc l inec
0 . 000 5 . 4 2 5 . 0 2 5 . 90
0 . 2 2 1 4 . 99 4 . 4 1 5 . 2 6
0 . 44 1 3 . 37 3 . 0 5 3 . 68
0 . 7 06 1 . 4 6 1 . 1 3 1 . s o
0 . 88 2 0 . 5 5 0 . 68 1 . 06
dm-do-tetracyc lined
0 . 000 5 . 89 5 . 0 5 5 . 2 5
0 . 97 8 4 . 58 4 . 5 5 4 . 7 7
1 . 9 5 5 3 . 6 5 3 . 4 2 3 . 50
2 . 97 2 2 . 7 6 2 . 57 2 . 4 7
3 . 9 10 1 . 4 6 1 . 5 7 1 . 6 5
4 . 888 0 . 84 1 . 07 0 . 97
a . An average va lue , determined on severa l d i fferent days .
b . Values of kt which were used in tne c a lcu lat ion of the v iab le inh ibitory rate c onstant , k� .
c . dm- is 6-demethy l-6-ep ihydroxy- . d . dm-do- is 6-demethy l-6-deoxy- .

Tab le I I . - (cont . )
M e kp kt k n a
7 -Bromo-dm-do-tetracyc l ined
0 . 000 5 . 8 9 5 . 4 1 5 . 8 9
2 . 4 68 4 . 2 9 3 . 7 8 3 . 5 2
4 . 9 38 2 . 7 0 2 . 59 2 . 2 9
7 . 406 l . 26 l . s o l . s o
9 . 87 6 0 . 8 5 0 . 5 5
7 -Ch loro- i s otetracyc l ine
0 . 000 5 . 7 1 5 . 2 6 5 . 40
16 3 . 4 9 5 . 8 1 4 . 74 5 . 3 9
3 2 2 . 30 5 . 7 1 4 . 86 5 . 5 3
l 2a-Deoxytetracyc l ine
0 . 000 5 . 7 1 5 . 26 5 . 40
7 9 . 14 3 . 90 3 . 6 9 3 . 59
1 5 6 . 0 3 l . 59 l . 57 0 . 9 5
Sa ( l la ) -Dehydrochlortetracyc l ine
0 . 000 5 . 7 1 5 . 26 5 . 40
1 5 0 . 68 5 . 2 3 4 . 26 4 . 5 5
2 97 . 06 4 . 64 4 . 0 3 4 . 66
2 -Cyano-2 -decarboxam ido-tetracyc l ine
0 . 000 5 . 7 1 5 . 2 6 5 . 40
3 3 . 16 5 . 94 4 . 92 5 . 44
1 28 . 88 5 . 88 4 . 6 3 4 . 7 2
e . Va lue s o f kt were obta ined with 0 . 2 % HCHO in the d i lut ing s olut ion , see Experimenta l .
8 4

Table I II . - F irst Order Generat ion Rate Constants , sec- 1 x 104 , for E . c o l i w .
Cu ltures Inhibited by Tetracyc l ines , M X 106 , and the Fract ion of Viable to
Tota l C e l l s Present at the Start , el , and at the End , e5 of the Experiments .
kv ka
kp k b b M t na e l e5
7 -Amino-dm-do-tetracyc l inec
0 . 000 5 . 84 5 . 87 5 . 4 6 5 . 9 3 0 . 84 0 . 8 1
0 . 4 9 3 4 . 9 3 4 . 80 4 . 3 2 4 . 7 2 0 . 80 0 . 96
0 . 98 5 3 . 7 1 3 . 8 3 3 . 36 3 . 66 0 . 7 5 0 . 6 3
1 . 4 7 8 2 . 98 3 . 0 5 2 . 74 2 . 7 7 0 . 68 0 . 62
l . 97 1 2 . 4 1 2 . 4 1 l . 96 2 . 02 0 . 7 2 0 . 7 1
a . Va lues of kt · we re obta ined with 0 . 2 % HCHO in the d i luting s olut ion , see Exper imenta l .
b . e was c alculated from Eq . ( l ) us ing the va lues ked and N° , obtained by regress ion ana lys is of the orig ina l data .
c . dm-do- is 6-demethy l-6-deoxy- .
co lJl

Tab le I I I . - ( c on t . )
M kv ka t
9-Amino-dm-do-tetracyc l ineC
0 . 000 5 . 84 5 . 87
0 . 474 5 . 27 5 . 27
0 . 94 8 4 . 06 4 . 59
1 . 4 2 1 3 . 7 9 3 . 7 3
1 . 89 5 3 . 06 -
5a ( 6 ) -Anhydrotetracyc l ine
0 . 000 5 . 82 5 . 6 2
4 . 4 7 7 5 . 00 5 . 4 6
8 . 9 5 3 4 . 38 4 . 67
1 3 . 4 30 4 . 12 3 . 88
17 . 907 - 3 . 32
kp kn a
5 . 4 6 5 . 9 3
4 . 6 1 4 . 86
3 . 9 1 4 . 08
3 . 38 3 . 44
3 . 02 3 . 17
5 . 40 5 . 90
4 . 70 4 . 90
3 . 7 9 3 . 87
3 . 2 5 3 . 4 5
2 . 56 2 . 7 7
e£
0 . 84
0 . 74
0 . 87
0 . 7 2
0 . 9 1
0 . 69
0 . 7 9
0 . 64
eg
0 . 8 1
0 . 7 4
0 . 40
0 . 80
1 . 2 1
0 . 3 5
0 . 52
0 . 90
OJ 0'\

Table I II . - ( c ont . )
M kv k� 7 -Nitro-dm-do-tetracyc l inec
0 . 000 5 . 8 2 5 . 62
0 . 1 1 1 5 . 68 5 . 50
0 . 2 2 2 4 . 20 4 . 7 5
0 . 3 3 3 3 . 17 3 . 8 2
0 . 444 2 . 4 5 2 . 8 2
9 -Nitro-dm-do-tetracyc l inec
0 . 000 6 . 10 5 . 86
2 . 3 3 7 4 . 7 7 5 . 37
4 . 67 4 3 . 9 3 4 . 11
7 . 0 1 1 2 . 9 3 2 . 96
9 . 348 l . 9 5 2 . 02
kp kna
5 . 4 0 5 . 90
4 . 86 5 . 44
3 . 88 4 . 32
3 . 1 1 3 . 5 2
2 . 19 2 . 59
5 . 8 1 6 . 14
5 . 07 5 . 1 5
3 . 48 3 . 54
2 . 8 2 2 . 55
l . 9 2 1 . 8 5
e�
0 . 9 1
0 . 67
0 . 74
0 . 7 2
1 . 00
0 . 74
0 . 97
0 . 68
0 . 8 2
0 . 69
e�
l . 2 1
0 . 8 7
0 . 3 3
0 . 28
0 . 6 1
1 . 0 5
0 . 4 1
0 . 5 3
0 . 7 7
0 . 62
00 -..J

Table I I I . - ( c ont . )
M kv It� kp kna �f .eb 5
4 - Dedimethy lamino-tetracyc l ine
0 . 000 6 . 10 5 . 86 5 . 8 1 6 . 14 0 . 74 1 . 0 5
3 . 0 30 5 . 2 7 5 . 39 4 . 6 3 5 . 0 1 0 . 7 9 0 . 67
6 . 060 4 . 3 2 4 . 50 3 . 96 4 . 3 0 0 . 8 0 0 . 53
9 . 091 3 . 4 3 3 . 80 3 . 58 3 . 5 1 0 . 86 0 . 50
1 2 . 12 1 3 . 17 3 . 12 2 . 97 3 . 04 0 . 6 5 0 . 7 0
9-Dimethylamino-dm-do-tetracyc l inec
0 . 000 6 . 2 5 6 . 09 5 . 4 5 5 . 4 0 0 . 66 0 . 8 3
3 . 4 6 7 5 . 60 5 . 7 1 5 . 0 1 5 . 00 0 . 87 0 . 74
6 . 9 3 5 4 . 32 4 . 69 3 . 92 3 . 99 0 . 7 0 0 . 4 1
10 . 40 2 3 . 34 3 . 4 7 3 . 0 9 3 . 5 1 0 . 60 0 . 50
1 3 . 87 0 2 . 8 3 2 . 94 2 . 4 0 2 . 56 0 . 74 0 . 6 3
Tetracyc l ine methiod ide
0 . 00 6 . 2 5 6 . 09 5 . 4 5 5 . 4 0 0 . 66 0 . 8 3
6 3 . 4 8 4 . 87 5 . 14 4 . 0 3 4 . 2 6 0 . 8 3 0 . 56
1 2 6 . 9 5 3 . 02 3 . 39 2 . 7 8 2 . 9 1 0 . I l 0 . 4 2 OJ OJ

Generation rate constants for cell divis ion have been �
obtained by two d i f ferent methods , by viable counts , kv , and
by tetal c e l l counts , kt . I t can be seen in Table I I I that
rate constants for mos t cultures obtained by the two methods
are apparently identical , within experimental error .
However , i n these cultures the concentrations o f cel l s
a t any time as determined b y the two methods were quite
d i f fe rent . This i s shown in Fig . XI for cultures inhibi ted
by 7 - amino - 6 - demethyl - 6 -deoxy-te tracycl ine . S imilar results
were obtained with the other tetracycl ines tes ted by both
methods . The fraction of viable ce l l s to total cel l s , e ,
8 9
would be 1 i f there was no d i f ference between the concentration
of c e l l s measured by the two methods . This f raction , � ' has
been calculated f rom the regre s s ion f i t of the experimental
data to Eq . ( 1 ) for the tetracy c l ines tested by the two
method s . Values o f e at two cul ture growth time s : e1 , 9 0 0
s econds a f ter antibiotic addi tion and 85 , 1 5 , 3 0 0 seconds
a fter antibiotic addition , are given in Table I I I . I t
should be noted that values o f e calculated in this manner
are very s en s i tive to sma l l d i f ference s in generation rate
constants . Despite thi s sens i t ivity , € remains relatively
cons tant in mos t cultures throughout the time o f antibiotic
i nhibi t ion . I n those cases where i t does vary , i t doe s not
appear to be con s i s tently related to antibiotic concentration .
The average value of e in these cultures i s about 0 . 6 . At
higher antibiotic concentrations when generation rate cons tants
obtained by the viable count method are negative , thi s fraction

1
6
..-; 8 ......_ I.D I 0 ..-;
� en 8 en ·ri � cU 0> � 2 0 ......,
Q) � ...:<
F igure XI .
+
��· ANTBOTIC CONe. COUIT METHOD
lot/L X 106 TOTAL VIABLE o.oo 0 ... o.ee e A 1 .9 7 • 0
Time , hr .
Examples of the Difference Between Total
and Viable C e l l Generation Rate Curve s . The logarithm
9 0
o f either the total number o r the number of v iable cells
a s a func tion of t ime of growth in culture s with var ious
concentrations of 7 -Amino-dm-do-tetracyc l i ne . The
s lope s , kt or kv , are g iven in Table I I I .

9 1
has value s o f 0 . 1 o r 0 . 0 1 . I t i s not pos s ible in the present
work to conclude w i th certainty that the tetracycl ines are
acting solely a s growth inhibitors and not as both
bacteriostatic and bactericidal agents . However , i f
there i s a n antibiotic induced cell death , i t is not large .
Rather i t seems mos t l ikely that the d i f ference in cell
concentrations obtained by the two me thods i s caused by the
format ion o f s ingle colonies by more than one viable cell .
I nclusion o f a sur factant in the viable cell di luting solution
did not , howeve r , greatly a ffect this fraction . The use of
the higher formaldehyde concentration in the di luting fluid
for total counts made thi s fraction , e , more cons tant .
Garrett and Brown ( 2 2 ) and Garrett , Mi ller and Brown
( 2 4 3 ) have observed that generation rate constants for cell
divi s ion obtained in the presence o f tetracycl ine obey the
re lationship :
i f f kcQ>O Eq . ( 2 )
I whe re ked may be e i ther kv or kt , ked i s the inhibitory rate
cons tant for ce l l divis ion and [ Te l is the antib iotic
concentration . This equation was obtained by simpl i f ication
of a more general relationship :
Eq . ( 3 )
where K i s a cons tant . The der ivation o f the se equations
will be pre sented in a later section of this the s is .
The generation rate cons tants obtained in the pre sent
work can be described by Eq . ( 2 ) but not by Eq . ( 3 ) . Typ ical
' , ,

examples o f the f i t o f experimental data to thi s expre s s ion _ ,
are shown in Figures XI I , X I I I , XIV and XV . Values o f the
I inhibi tory rate con s tan t , ked ' obtained by app l ication o f
regres s ion analy s i s t o the experimentally determined
generation rate con s tants and the antibiotic concentrations
in the cultures are given in Table IV . Thes e values are
e s timates of the activi ty of the tetracycl ines which should
be sui table for use in s tructure- activity relationships .
B . E f fect o f Tetracyc l ines on Rate Constants for
Protein and Nuc l e i c Aci d Synthe s i s .
I t was mentioned i n the l i terature survey that many
inves tigators have observed that tetracyclines cause an
inhibition of protein synthe s i s in bacterial culture s . The
results of two typical experiments in which the rates of
9 2
prote i n synthe s i s i n E . col i cultures inhibited by tetracyclines
were s tudied a re shown in Fig . XVI and XVI I . The se results
can be described by an equation s imilar to Eq . ( 1 ) :
[ P ] = [ P O ] ekPt , Eq . ( 4 )
when the time , t , i s greater than 3 0 minutes a fter antibiotic
addition and where [ P0 ] i s the concentration o f prote in in
the ce lls 30 minutes a fter antibiotic addi tion , [ P ] i s the
protein concentration in the cel l s at any later time and k p
i s the generation rate cons tant for prote i n synthes i s in
presence of antib iotic . The generation rate cons tant for
prote in synthe s i s in the absence of antibiotic , k� , i s
identical with the generation rate constant f o r cel l division
in the absence o f antib iotic , k�d · Value s o f kp are g iven in

<:!' 0 .--l :><
.--l I
() (j) Ul
� Ul -!-) � m -!-) Ul � 0 u
(j) -!-) m 0::
� 0
· ri -!-) m H (j) � (j) 0
9 3
+ 6 - �D'h o-
COt.Ha 5 - OH O H O 0
4 - 0 VIABLE Ci"NUCLEIC ACIDS"
3 - 4"PROTEIN"
2 -
1 -
1 . 0 2 . 0
Tetracyc line , M/L X 1 0 6
F igure X I I . The Linear Dependence of Generation Rate
Constants , kv , kp a nd kna , upon Tetracyc l ine Concentra
t ions . The line shown i s the average inhibitory
rate constant , kivg ' it and va lues of k� , k� and k�a
are g iven in Table IV .

""' 0 .--i X
.--i I
() <lJ UJ
UJ 4-l c: rcl +l UJ c: 0 u
<lJ +l rcl p::;
c: 0
·ri +l rcl )...! <lJ c: <lJ �
6 -
5 -
4 -
3 -
2 -
1 -
5 . 0
+ om�·-o-
0/f COMi OH 0 HO 0 1
• TOTAL c V I �BLE
0 -NucLEIC ACIDS• A �ROTE IN•
1 0 . 0
9 -Nitro-dm-do-tetracyc l ine , M/L x 1 0 6
F igure X I I I . The Linear Dependence o f Generation Rate
9 4
Constants , kv , kt , kp and kna , upon 9-Nitro-dm-do
tetracyc l i ne concentration . The slope of the line shown
i s the average inhibitory rate constant , klvg , i t and
values o f k� , kf_ , k� and kAa. are g iven in Table IV .

""' 0 .--i �
.--i I
u (J) (/)
(/) +J s:: m +J (/) s:: 0 u
(J) +J m �
s:: 0
·rl +J m H (J) s:: (J) (.9
6 -
5 -
4 -
3 -
2 -
1 -
+ NHICt\l1
C H, �o-" :-... H N CON ct( ai 0 H O
Ha
• TO TA L c VI ABLE
:0 •N UC L E IC A C I D S
•
A •
P R O T E I N•
5 . 0 1 0 . 0
9-Dimethylamino-dm-do-tetracyc l ine , M/L x 1 0 6
F igure XIV . The L inear Dependence of Generation Rate
Constant s , kv , kt , kp and kna , upon Concentration of
9-Dimethylamino-dm-do-tetracyc l ine . The s lope of the
l ine s hown is the average inhibitory rate constant ,
k�vg ' i t and values of k� , kf , k� and k�a are g iven
in Table IV .
9 5

""' 0 .-I X
.-I I
u Q) Ul
Ul .jJ c ctl .jJ Ul c 0 u Q) .jJ ctl p:: c 0 ·ri .jJ ctl � Q) c Q) t9
6- · CONH2 HO 0
R, Ra Ra 5 - • H H NOa " PROTEIN"
0 H H H VIABLE 4 - 0 H ltia H TOTAL
A OH CL "NUCLEICAOOS'
3 -
2 -
1 -
5 . 0 1 0 . 0
Antibiotic Concentration , M/L x 10 6
F igure XV . The Linear Dependence of Generation Rate
Con s tants , kv , kt , kp and kna ' upon Concentration of
s everal Tetracyc l ine s . The s lopes of the l ine s shown
are the average inhibitory rate constants , k�vg ' they
and va lue s of k3 , k£ , k� and k�a for each antibiotic
are l i s ted in Table IV .
96

Table IV . - Inhibitory Rate Cons tants , 1./M- sec . , for Inhibition o f E . coli W by
Various Tetracyc l ine s .
Tetracycl ine Analoga -ki -ki -ki -ki -ki b v t p na avg
7 -Ni tro-dm-do-tetracyc l ine 8 3 3 . 5 1 6 5 6 . 7 6 7 3 5 . 6 8 7 6 9 . 1 9 7 4 8 . 7 8 * *
7 -Chloro-dm-te tracyc l ine 5 9 1 . 8 6 - 4 6 0 . 2 5 5 3 6 . 4 8 5 1 7 . 2 1
7-Ch lorotetracyc l ine 3 8 7 . 7 7 - 3 9 6 . 1 4 4 2 0 . 2 0 4 0 1 . 3 7
Tetracyc l ine 2 6 7 . 8 4 - 2 7 9 . 9 7 2 6 7 . 9 5 2 7 1 . 5 0
5 -Hydroxytetracy c l ine 2 7 5 . 1 9 - 2 5 4 . 7 9 2 5 1 . 8 3 2 5 7 . 1 5
7-Amino-dm-do- tetracycl ine 1 7 9 . 0 1 1 7 5 . 9 0 1 7 4 . 0 8 1 9 7 . 9 5 1 8 1 . 7 3 * *
a . dm-do- i s 6 -demethy l - 6 -deoxy- , whi le dm- i s 6 -demethyl - 6 -epihydroxy- . b . Tested by ana lys is of covariance to determine i f s ign i f icant d i f ferences
exi sted between the inhib itory rate constant determined by us ing a l l values of generation rate constants col lective ly , ki
v , and ind ividual inhibitory rate constants determined separately . The symbo l � �ndicates sign i f i cant d i fferences at P) 0 . 9 5 , whi l e * * indicates s igni f icant a t P> 0 . 9 7 5 . S e e f o r example , S tee l , P. . and Torrie , 0 . , " Principles and P rocedure s of S tatistic s " , McGraw H i l l Book Co . , I nc . , New York , N . Y . , ( 1 9 6 0 ) pp . 1 7 4 .
1.0 -...)

Tab l e I V . - ( cont . )
Tetracyc l ine a n a l o g
9 - Amino-dm-do-te tracyc l ine
dm- do- tetracy c l ine
7 - Bromo- dm-do - t e tracyc l ine
- k i v
1 5 5 . 5 3
1 0 3 . 6 2
9 - N i tro - dm-do- tetracy c l ine 4 3 . 4 0
4 - Dedime thy l amino - te tracyc l ine 2 5 . 3 9
9 - D imethy l amino-dm-do- t e tracyc l ine 2 6 . 2 2
S a (6 ) -Anhydrote t r a cyc l ine 1 2 . 7 5
1 2 a - Deoxytetracyc l ine
Te tracyc l ine me thiodide 2 . 5 4
S a ( l l a ) -Dehydrochlorote tracyc l ine
7 - Ch loro- i s ot e t racyc l ine
2 - Cyano - 2 -decarboxamido - te tracyc l i ne
- k i t
1 5 1 . 2 3
6 2 . 6 8
4 3 . 1 5
2 3 . 2 9
2 4 . 6 2
l 3 . 81
2 . 6 3
2 . 1 2
0 . 3 6
- k i p
1 2 8 . 82
86 . 7 0
4 6 . 0 9
4 2 . 9 1
2 2 . 2 4
2 3 . l l
1 5 . 9 1
3 . 85
l . 9 6
0 . 2 5
- k i n a
1 4 6 . 1 3
9 2 . 9 3
5 1 . 4 2
4 7 . 86
2 5 . 4 5
2 0 . 7 3
1 7 . 2 3
2 . 3 7
2 . 1 0
0 . 4 2
no a c t iv i ty obs e rved
no a c t ivi ty obs e rved
- k i avg
b
1 4 5 . 0 3 *
9 4 . 4 2 *
5 1 . 7 9
4 4 . 3 3
2 4 . 0 9
2 3 . 6 7 * *
1 5 . 4 8* *
2 . 6 2 c
2 . 1 8C
0 . 3 4 c
c . I n s u f f i c i ent d a t a ava i l ab l e for t e s tinq by a n a l y s i s o f cova r i ance .
I.D co

9 9
+ 2 -
+ 1 -
.:: . .., 0 -Q) +l 0 H P< +
,. , � 1 - w���· .......
Q) O.N CONHa .:: 0 H
- 2 - AN TIBIOTIC CONe. MIL X 10
6
0.00 0 2 .34 •
- 3 - 4.67 Q 7.01 A 9 .34 •
l 2 3 4
Time , hr .
F igure XVI . An Example of Generation Rate Curves for
Protein Synthe s i s . The logar ithm o f the " Protein"
f ound in E . coli cultures a s a function of time of
growth i n the pre sence of various concentrations of
9 -N i tro-dm-do-tetracycl ine . The s lope s , kp , are g iven
in Table I I I .

Q) s::
H
+3
+ I
0
-I
·2
·3
0:/� ��
2
OH 0 HO 0 ANTIBIOTIC COHC.
MIL XI08 0.00 0 0. 1 1 • 0.22 Q 0.33 A 0.44 •
3
T ime , hr .
4 4
F igure XVI I . An Example of Generation Rate Curves for
Protein Synthe s i s . The logarithm of the " Protein "
f ound i n E . c o l i cultures a s a func tion o f time of
growth i n the pr esence of var iou s concentrations of
7 -Nitro-dm-do-tetracyc l ine . The slope s , kp , are g iven
in Table I I I .
1 0 0

Tab le s I I and I I I for the 1 8 antibiotics s tudied . I t can be
seen that the se value s are e i ther identical with or are
very close to the values of kv or kt observed in any given
culture .
1 0 1
�eneration r ate cons tants for prote in synthe s i s obtained
by regres s ion analy s i s from Eq . ( 4 ) are in agreement with an
equation s imilar to Eq . ( 2 ) :
Eq . ( 5 )
where k� i s the inhibi tory rate constant for protein synthe s i s .
Values o f kp do not f i t an equation s imilar to Eq . ( 3 ) . Fig .
XII , XI I I , XIV and XV are examples o f the fit of values o f kp
to Eq . ( 5 ) . Values o f the inhibi tory rate cons tants for
protein synthe s i s are given in Tab le I V .
The results o f two typical experiments i n which the rate s
o f nuc leic acid synthe s i s in culture s inhibited by tetracycl ines
were s tudied are shown in Figures XVI I I and XI X . The se
experiments did not show an increased rate o f nucleic acid
synthes i s in inhibi ted cul tures as sugges ted by several
inve stigators ( 1 8 , 3 0 , 3 6 ) but rather showed a decreased
amount o f nucleic acids corre lated to the observed inhib ition
of cel l divis ion . The data from these exper iments a l so f i t
a n expre s s ion s imilar t o Eq . ( l ) :
[ NA] = [ NA0 ] eknat , i f f kga>kna� Eq . ( 6 )
when the time , t , is greater than 3 0 minutes after antibiotic
addition and where [NA0 ] i s the concentration o f nucleic ac ids
in the cells 30 minutes after antibiotic addition , [NA] i s
the nuc leic a c i d concentration in the ce l l s a t any later time

1 0 2
+ 2 -
•
= + 1 -Ul
'0 · rl u � 0-u · rl Q) r-i + u - 1 - NHk;H,J1 ::l
���NHa z
--' H 0
Q) - 2 - A N TlBIOTIC COHC. c
...:l W,I\. X 6 10
0.00 0
- 3 - 2 .34 ... 4.67 8 7 .0i A 9 .34 •
1 2 3 4
T ime , hr .
F igure XVI I I . An Example of Generation Rate Curves for
Nuc leic Ac id Synthe s i s . The logar i thm o f " Nuc l e ic Ac ids "
f ound in E . c o l i culture s a s a function of time of
growth in the pre sence of various concentrations of
9 -N i tro-dm-do-tetracyc l ine . The slope s , kna , are g iven
in Table I I I .

= Ul
'"d ·ri u � u
·ri (J)
� u ::l z
+3
+ I
0
-I
-2
0
2
• +
W
N�
� 0
ANllBIOTIC CONC.
Mil.. . )CI06 0.00 .-.- 0
J
0.98 - · 1 .96 � e 2 .9 7 · A 3.9 1 Q 4 -89 � .
T ime , hr .
•
F igure XIX . An Example of Generation Rate Curves for
1 0 3
Nuc leic Ac id Synthe s i s . The logar i thm o f "Nuc leic Ac ids "
found i n E . c o l i culture s a s a function of time o f
growth i n the pre sence of various concentratQons o f
diD-do-tetracyc l ine . The s lopes , kna ' are g iven in
Table I I I .

and kna i s the generation rate cons tant for nucleic acid in
the presence o f antibiotic . The generation rate constant
1 0 4
for nucleic ac id s ynthes i s i n the absence of antibiotic , kga ,
i s identical to the generation rate cons tant for ce ll divis ion
in the absence of antibiotic , k�d · Generation rate constants
obtained from the f i t of experimental data to Eq . ( 6 ) are
given in Tables I I and I I I . I t can be seen that these values
are e i ther i dentical to or very s imi lar to the values o f
kv , kt o r kp observed in any given culture .
Generation rate cons tants obtained from Eq . ( 6 ) are
in excellent agreement with an equation s imi lar to Eq . ( 2 } :
Eq . ( 7 )
whe re k i i s the inhibito ry rate constant for nucleic acid na
synthe s i s . Figure s XI I , XI I I , XIV and XV are examples of the
f i t o f va lue s o f kna to Eq . ( 7 ) . Values of the inhibitory
rate cons tants derived from Eq . ( 7 ) are l i s ted in Table I V .
Value s o f kna do n o t f i t a n equation s imilar t o Eq . ( 3 ) .
As mentioned above , the values o f kv , kt , kp and kna
for any given culture are e i ther identical or very s imi l ar .
There fore , values of the respective inhibitory rate constants ,
I I I I d b . ' 1 � h kv , k t , k or kna ' would be expecte to e slml ar . n e ave p
tes ted the hypothe s i s that the se constants are experimentally
ind i s tingui shable and find no s tatistically s igni f i cant
dif ferences between the individual values and an average
inhibitory rate constant , k�vg ' for 7 of the antibioti cs .
Stati s tically s igni ficant d i f ferences were found , Table I V ,
for the 6 o the r antibiotics tes ted . Suff icient data was not

l O S
ava i lable t o test the other antibiotics i n this fashion . This
s imple analy s i s of covari ance te s t did not consider the error
invo lved in the determination of the individual generation
rate constants . I t seems l ikely that within present experimental
accuracy the 4 generation rate cons tants , viable and total cell
divis ion , protein and nuc l e i c acid synthe s i s , are the same
for a l l antibiotics . That i s a one-to-one relationship , balanced
growth , is found among the se parameters not only in contro l
cul tures but also in antibiotic inhibited culture s .
I f balanced growth i s indeed maintained in tetracycline
inhibited cultures then the average inhibito ry rate constant ,
k�vg in Table I V , would be the bes t measure o f tetracycline
activity to use in s tructure-activi ty relationships .
C . Time o f Onset o f Tetracyc l ine Inhibition of Whole
Cell Culture s .
The fact that inhibited cultures exhibit paral lel rates
o f cell divis ion , prote in and nuc l e i c acid synthe s i s is not
in accord w i th the interpretation of previous experiments
( 1 8 , 3 0 , 3 4 , 3 6 ) . These experiments were e i ther s ingle
point measurements or kinetic experiments which were carried
out only during t ime s immediately preceeding antibiotic
addition . The se expe riments reported e i ther decreased
protei n/ce l l and increased nuc l e i c acid/ce l l or inhibition
of rate o f prote in synthes i s and s timulation o f rate o f
nucleic acid s ynthe s i s . V /
However , Cerny and Habermann ( 3 3 )
in a kinetic experiment covering several generations showed
parallel rates o f protein and nucleic acid synthe s i s with
different t ime s of onset of action . The i r work was done at

antibiotic concentrations 1 5 times higher than those
presently employed , which caused complete inhibition and
undoubtedly cell death .
The agreement of exper imental data with Eq . ( L ) , Eq .
1 0 6
( $ ) o r Eq . ( ¢ ) wa s only observed for data obta ined approxima tely
3 0 minutes af ter antibiotic addi tion . Dur ing thi s initial
t ime per iod cultures appeared to maintain growth rates s imi lar
to tho s e observed in control s . Thi s can be observed in the
exper iments shown in F igures IX , X , XI , XVI , XVI I , XVI I I and
XIX . S imi lar observations were made in the other experiments
not shown . Figure XX shows an experiment des igned to document
this observa tion . I t can ea s i ly be seen that the onset of
inhibition for a l l parameter s is cons iderably beyond the time
of antibiotic addi tion . Viable plate counts , not shown ,
par a l l e led tho se obta ined by the total count method . Thi s
exper iment ha s been reproduced at a dif ferent oxytetracyc l ine
conc entration with s imi lar results . In addi tion , Fig . XX
shows that the time of onset of recovery from inhibition
upon d i lution of cultures into antibiotic free media is not
immed iate e i ther , but a l so exhibits a lag time . I t was
thought that both inhibition of and recovery of protein syn
the s i s occurred s l ightly before the other parameters are aff ected .
In order to test the observation on time of onset of inhibition
an exper iment wa s performed in a dextrose-salts broth where
the rate of c e l l d iv i s ion in control cultures wa s 1/2 that in
the high peptone broth . Thi s s ing le exper iment , Fig . XXI ,
sugg e s t s that prote in synthe s i s measured by the Fo lin-Lowry
technique and a l so c e l l ma ss as measured by turbidity might be

1 0 7
F igure XX . T ime o f O n s e t o f and Recovery F r om Oxytetracy
c l ine Inhibition i n Peptone Broth C u l tur e s . Genera t i o n rate
curves for tota l c e l l s , nuc l e i c a c id synthe s i s a nd pro t e i n
synthe s i s i n t h e pre s ence o f 1 . 3 8 x l o - 6 � 5 -Hydroxytetra
cycl i ne and a one - to - e l even d i lu t i on i n to a n t i b i o t i c -
free med ia of the i nh i b i ted c u l tur e . The gener a t i on r a te
curve f o r pro t e i n synthe s i s has been transpo s ed by sub
trac t i ng 0 . 5 f r om a c tu a l v a l u e s of Lne " Pr o t e i n . " The
genera tion rate c o n s ta n t s , sec . - 1 x 1 0 4 , f o r the und i luted
c u l tur e s are g iven i n Table II exc ept f or v a lu e s obta i ned by
measur i ng cu lture tur b i d i ty ( no t s hown ) whi c h were 5 . 6 3 a nd
2 . 1 9 before and a f ter a n t i b i o t i c add i t i on , r e spec t iv e ly . The
values of the generation r a t e c o n s t a n t s in the d i luted c u l ture
wer e k t 5 . 4 3 ; kp = 5 . 0 9 ; kna = 4 . 6 1 ; kv ( not s hown ) = 5 . 3 3
and for c u l ture turb i d i ty = 4 . 7 6 .
The t ime o f a n t i b i o t i c add i t i o n wa s 4 , 4 7 0 sec . and the
t ime o f d i lution wa s 1 0 , 8 5 0 sec . The t im e s o f onset o f
inh i b i t io n and the t im e s o f o n s e t o f r ecovery c a lc u l ated a s
the inter s e c t i on o f the regr e s s ion l i ne s , r e spec t i v e l y i n
sec . , wer e : f or protein synthe s i s , 6 , 2 0 0 a n d 1 1 , 8 0 0 ; f or
turbid ity , 6 , 4 0 0 and 1 1 , 5 0 0 ; f o r nuc l e i c a c i d synthe s i s ,
6 , 4 0 0 and 1 2 , 4 0 0 ; and for c e l l d iv i s ion - tota l 7 , 1 0 0 a nd
1 2 , 5 0 0 and viable , 6 , 5 0 0 a nd 1 2 , 2 0 0 .

l O B
DILUTION
+4
/ r /'
r OXYTETD�yt - LINE 1.38X 106 MIL
!
l
0 TOTAL A "NUCLEJCACICS" e "PROTEIN"
-4
T ime , hr .
F igure XX .

F igure XXI . Time of Onset of Oxytetracyc line Inhibition of
Cultures in Dextrose-Sa lts Broth . Generation rate curves
1 0 9
for nuc leic acid synthe si s , protein synthes i s and culture
turbidity in the presence of 1 . 1 6 x 1 0- 6� 5-Hydroxytetra
cycline . The generation rate curve for nuc leic acid
synthesis has been transposed by adding 2 . 0 to the actual
value of lne "Nucleic Ac ids . " Values of generation rate
constants , sec . -1 x 10 4 , in the absence and presence of
antibiotic respectively , were : kna = 2 . 1 3 and 0 . 5 6 ; kp = 2 . 6 2
and 0 . 3 7 ; kt ( not shown ) = 2 . 4 4 and 0 . 1 2 ; and for turbidity
2 . 5 4 and 0 . 3 7 .
The time of antibiotic addition was 8 , 9 0 0 sec . The
times of onset of inhibition , sec . , ca lculated a s the inter
cept of the regre s sion lines were : for protein synthesi s ,
9 , 9 0 0 , for turbidity , 1 0 , 3 0 0 ; for cell division , 1 0 , 7 5 0 ; and
for nuc leic acid synthesis , 1 4 , 2 0 0 .

1 1 0
+3
+t
+I
s:: 0 . o ·.-i .j.) 1\l )...! .j.) s:: Q) u s:: 0 u �
Q) s:: ..:I A "Nucc.£Jc /lCD/I' • "Fitol'EIN"
e lUt8lliTY
- ·
2 4 8 10
Time , hr .
F igure XXI .

1 1 1
inhibited sooner than nuc leic acid synthe s i s . Cell divis ion ,
not shown , was inhibited at an intermediate time . The exis tence
of the s e d i f ferent l ag times explains the dis crepancy between
parallel equ i l ibrium inhibition rate s observed in thi s work
and non-parallel amounts of nucleic acid and prote in per cell
reported by o thers ( 1 8 , 3 0 , 3 4 , 3 6 ) . Experiments per formed
in cul ture med i a whi ch do not al low rapid rates of cell
divis i on would be expecte d to accentuate thi s d i f ference in
times o f onse t o f inhibi tion .
Thi s o rder o f inhibition and recovery is cons i stent with
the hypothe s i zed primary inhibi tion o f prote in synthes i s
fol lowed b y a secondary , indirect inhibition of cell divis ion
and nucleic acid synthe s i s due to a lack of prote in .
D . E f fe c t o f Broth pH Changes on Tetracyc line-Inhibited
Culture s .
Jones and Morris son ( 2 0 ) and Benbough and Horris son ( 1 7 )
have reported that when the pH o f culture media o f known free
metal ion concentration i s varied , the inhibition produced
is proportional only to the " mo lecular " , presumab ly zwitterionic ,
species o f several tetracyclines . Connamache r , Mande l and
Hahn ( 6 6 ) have recently reported data on the uptake o f
tetracycline in B . cereus cultures at pH 6 . 7 and 7 . 3 and
at two Mg ion concentrations which are in qual itative agree
ment with the hypothe s i s o f Jones and Horr i s son .
Because the s l ow growth rate characteristics o f E . coli
w in dextro s e -s a lts broth made it unlikely that the
tetracyc l ines would be suffic iently stable it was not
pos sible to ver i fy these results for other te tracyclines in

1 1 2
a media o f known free metal ion concentration . However ,
experiments in peptone broth are con s i s tent with these results
at two broth pH values where free metal ion concentration would
be expected to be constant . Figure XXI I shows a plot o f
Eq . ( 2 ) f o r experiments at p H 6 . 2 and 6 . 6 5 . The concentration
o f tetracycl ine is adj us ted to reflect the concentration of
zwitterionic form rather than the total concentration .
Experiments at a broth pH o f 7 . 4 , where a much lower free
me tal ion concentration would be expected because of the
higher HP04- concentration , do not f i t the s ingle l ine
plotted in Fig . XXI I . Activities obtained with 5 a ( 6 )
anhydrotetracycline ( pH 6 . 2 and pH 6 . 6 5 ) and 4 - dedimethyl
amino-tetracycline ( pH 6 . 6 5 and pH 7 . 4 ) do not f i t such a
s imple concept e i ther , Table V . The se tetracycl ines have ,
however , unusual pKa ' s .
I n addition to the growth-rate dependent d i fference in
time of onset o f inhibi tion o f the various culture parameters ,
there i s a lag time observed before any parameter is inhibited ,
Fig . XXI . Two general explanations are pos s ible for thi s
l a g time . Firs t , a finite time may b e required t o es tablish
equ i l ibrium concentrations within the cell s . Second , antibiotic
may permeate the cell and arrive at the receptor s i te but
require finite time to exert a measurable e f fect .
Bes ides causing a change in the degree o f equ i l ibrium
inhibition produced by a given total antibiotic concentration ,
broth pH changes are also capable o f changing the growth-rate
independent lag time for onset o f inhibition . Figure XXI I I
shows an experiment identical to that pre sented previously ,

\0 0 H X
..-il u
u Q) Ul
Ul +l .:: m +l Ul .:: 0 u Q) +l m p::; .:: 0 ·rl +l m H Q) .:: Q) (j
6 -
3 -
4 -
3 -
2 -
1 -
l .,H 6 .65 .,H 6.1S
... A •NUCLEIC ACIDS•
�� ' • 0 •PROTE IN•
• VIABLE
c TOTA L oe D
0 . 5 1 . 0
[Chlortetracyc l ine ] , M/L x 1 0 6
1 + Ka/ [ H+ ]
F igure XXI I . The Linear Dependence of Generation Rate
1 1 3
Constants upon Concentration o f Zwitter ionic Chlortetra-
cyc l ine . Constants were obta ined at two broth pH value s ,
6 . 1 5 and 6 . 6 5 . Values of generation rate constants
obta ined at a higher broth pH value , 7 . 4 , and , therefore ,
a higher HP04 = concentration do not f it thi s s ingle line .
The s lope of the line shown i s 4 7 0 L/M- sec .

1 1 4
Table V . - F irst Order Generat ion Rate Cons tant s , sec- 1 x 104 ,
and Inhibitory Rate C onstant s , L/M-s ec . , f or E . c o l i w in
Various Broth C u l tures Inhib ited by Several Tetracyc l ines .
7 -Chlorotetracyc l ine , peptone broth , pH 6 . 2
0 . 000 5 . 56 5 . 50 4 . 88
0 .. ll.92 4 . 9 5 4 . 55 4 . 66
0 . 384 4 . 06 3 . 50 3 . 38
0 . 57 5 3 . 0 1 2 . 69 2 . 7 1
0 . 7 6 7 2 . 10 1 . 78 1 . 86
-4 6 1 . 1 1 -484 . 15 -4 17 . 52 -4 54 . 28**
Sa ( 6 ) -Anhydrotetracyc l ine , peptone brot h , pH 6 . 2
0 . 00 5 . 56 5 . 50 4 . 88
4 . 67 4 . 7 1 4 . 68 3 . 90
9 . 34 3 . 94 3 . 2 3 3 . 4 5
14 . 0 1 3 . 35 2 . 57 2 . 92
18 . 68 1 . 7 3 1 . 99 2 . 3 5
- 1 9 . 27 - 19 . 54 - 1 2 . 9 5 - 1 7 . 2 5 **
* * S ig n i f icant d i f ferences between inh ib itory rate constants determ ined by analy s is of c ovar ianc e as expla ined in Tab le IV .

Table v . - ( cont . )
106 M kt kp kna k i avg
4 -Ded imethy l amino-tetracyc l in e , peptone broth , pH 7 . 4
0 . 00 4 . 57 5 . 2 2 4 . 7 6
3 . 34 3 . 87 5 . 0 1 3 . 96
6 . 6 5 3 . 3 3 3 . 7 3 3 . 48
9 . 96 2 . 7 1 3 . 0 3 3 . 00
1 3 . 24 2 . 0 9 2 . 1 3 2 . 54
k i - 1 8 . 48 - 24 . 7 0 - 16 . 3 2 - 1 9 . 8 3 **
7 -Chlorotetracyc l in e , pH 6 . 7 , dextrose-sa lts broth
0 . 000 2 . 39 2 . 30 2 . 18
0 . 16 3 2 . 07 1 . 92 2 . 0 1
0 . 32 7 0 . 97 0 . 84 0 . 7 9
0 . 4 90 0 . 39 0 . 51 0 . 50
0 . 6 5 5 0 . 2 2 0 . 38 0 . 37
k i - 368 . 34 - 3 1 5 . 8 1 - 3 12 . 59 - 3 3 2 . 2 5
* * S ign i f icant d i f ferences between inhib itory rate constants dete rmined by ana lys is of c ovar iance as exp la ined in Table IV .
1 1 5

F igure XXI I I . The E f f e c t of Broth pH on Time o f O n s e t of
1 1 6
Oxytetracy c l ine I nhib i t i on . Genera t ion r a t e curve s , done on
s epara te days a t two peptone broth pH v a l ue s , f o r c e l l
d i v i s ion- b y the to t a l c ount me thod ana f or c u l ture
turb i d i ty in the presence of two d i f f er e n t c oncentr a t ions o f
5 - Hydroxy t e tr a c yc l i ne . The g ener a t i on r a t e curve s f o r c e l l
d iv i s ion have been transposed b y subtr a c t i ng 3 . 0 f r om the
a c tual value o f Lne ( orga n i sms/ml . x l o - 6 ) . The g ener a t i on
rate constant s , sec . - 1 x 1 0 4 , i n the absence and pr e senc e
of antibiot i c , re spec tively , f o r c e l l d i v i s io n , were 5 . 1 6
and 2 . 2 0 a t pH 6 . 6 5 and 4 . 7 5 and 2 . 3 0 a t pH 7 . 4 , and f o r
turb i d i ty wer e , 5 . 6 3 and 2 . 1 9 a t pH 6 . 6 5 a n d 5 . 5 6 a n d 2 . 2 1
a t ph 7 . 4 .
The t ime s of a n t i b i o t i c add i t ion were 4 , 4 7 0 s ec . a t ph
6 . 6 5 and 5 , 4 0 5 sec . at pH 7 . 4 . The t im e s o f o n s e t of
inhibi tion , sec . , c a l c u la ted a s the inte r s ec t i o n o f the
reg r e s s ion l i nes were f o r c e l l d iv i s ion 7 , 1 0 0 and 6 , 2 0 0 and
for turbid i ty 6 , 4 0 0 and 5 , 9 0 0 a t pH 6 . 6 5 a nd 7 . 4 , r e s pec t i v e ly .

1 1 7
+ 3 -
+ 2 -
+ 1 -
s:: 0 0 -·rl .jJ C1l H - 1 -.jJ s:: Q) u - 2 -s:: 0 u ---'
Q) - 3 -
s:: ...:l - 4 - ,.H 6.65 ,H 7.4
0 • TOTAL
"' • TURBIDtTY - 5 -
- 6 -
1 2 3 4
T ime , hr .
F igure XXI I I .

1 1 8
Fig . XX , except that the pH o f the broth was 7 . 4 instead o f
6 . 7 . The lag for onset o f inhibition o f protein synthesis
( me as ured a s cel l mas s ) and ce l l divis ion are seen to be
shortened by two-thirds . The equil ibrium amount o f inhibition
achieved by two d i f fe rent total oxytetracycl ine concentrations
was the s ame i n both experiments . I f the hypothes i s o f Jones
and Mor r i s son ( 2 0 ) i s correct , then the concentration of
zwitterioni c species present in the broth mus t be the same in
both experiments . The nature o f the initial inhibi tory process
mus t be quantitative ly d i fferent i n these experiments . I t
seems mos t l ikely that the proces s o f permeation into the
cel l s is a f fecte d by the pH change and that thi s is the proces s
respons ible for the growth-rate i ndependent lag time observed
in the s e experiments .
Arima and I z ak i ( 6 4 , 2 4 4 , 2 4 5 ) have s tudied permeation
o f oxytetracy c line in sens i tive and res i s tant cells at high
antibiotic concentrations and found that i t depends upon an
energy requ iring proce s s . Franklin ( 6 5 ) has veri fied a
glucose-dependent uptake at low concentrations o f tetracycline
and chlortetracy c l ine . The condition o f the cells in these
cultures i s so d i f ferent from the cel l s in the present work
that it is hard to make meaningful comparisons . However , i t
i s c l e a r that the abi li ty to permeate the ce l l s is a n important
factor in antibiotic activity , whether thi s proces s is an
inactive or an active di ffus ion . E i ther proces s might be
expected to be sensitive to the broth pH .

E . K inetic Mode l s for Inh ibition o f Bacterial Cultures
by Tetracyc l i ne s .
S tudi e s of the k inetics o f inhibi tion o f ce l l growth
are capabl e of provi ding in formation of a type other than
1 1 9
the estimate s o f rel ative potencies o r the estimates o f order
of onset of inhibition which have been mentioned above .
Kinetic s tudies can provide information which is useful in
testing proposed mode l s of antibiotic activity . Any proposed
mode l of tetracy c line activity should be consistent with
several results of the present study .
The se are :
a . Exponential growth i s maintained in cultures inhibited
by l ow concentrations o f tetracyc l ines ; see Eq . ( 1 ) , ( 4 ) and
( 6 ) • b . The generation rate constants for ce l l divi s ion ,
prote in and nuc leic acid synthes i s are the same in any given
tetracyc line-inh i b i ted culture , kcd�O .
c , Rate cons tants for inhibited cultures are l inearly
dependent upon the f i r s t power of the antibiotic concentration ;
see Eq . ( 2 ) , ( 5 ) and ( 7 ) .
d . Ant ib io t i c inhibition i s reversed by di lution into
antibiotic free med i a , Fig . XX .
I n addi tion , it seemed l ikely that :
1 . P rote in synthes i s i s inhibi ted be fore ce l l divis ion or
nucl e i c ac id synthe s i s .
2 . Permeation o f antibiotic into ce l l s i s important and
might be responsible for the l ag in time of onset of inhibition .

A s imple mode l which des cribes the kinetics o f ce ll
div$s i on observed in E . coli culture s inhibited by low
tetracycl ine concentrations has been proposed by Garrett ,
Mil ler and Brown ( 2 4 3 ) . I t assumes equ i l ibrium ( K )
between free E . co l i ( Ef ) and tetracycline ( T c ) t o form
inhibited E . coli ( Ei ) :
1 2 0
E f + T c .J<..t: Ei Eqj ( 8 )
and i t a l s o a s s ume s that the rate o f generation o f total
E . coli ( E t ) i s proportional to the concentration of free
E . c o l i 'f
Eq . ( 9 )
When the fraction of inhibited E . coli ( � ) i s calculated
from Eq . ( 8 ) :
� E · K [ T e l l Eq . Et 1 + K [ Te l
and when s ub s t i tuted i n Eq . ( 9 } , one obtains :
d ( Et ) = k�d ( l - � ) Et = k�d ( l - �K:..:........::...[ T:._c:....;l:..__ ) l + K [Te l a t
S ince it i s found experimental ly that ,
d E t Eq . ked
( Et ) d t
by comb ining Eq . ( ll } and Eq . ( 1 2 ) ' one obtains :
ked kgd l - K [TcJ Eq .
l + K [Te l
f E ( 3 ) When K [Tc ] i s much which is a rearrangement o q . ·
( 1 0 )
Eq . ( l l }
( 1 2 )
( 1 3 )
less than 1 , a " l imiting case " , then Eq . ( 1 3 ) i s equivalent to
Eq . ( 2 ) , where k; . d . = k� . d .
K .

Thi s simp l e kinetic mode l is con s istent with re sults
outlined as a, c arrl d.. I t does not consider the observations
outl ined in b , 1 and 2 . Consequently , i t does not lead to
1 2 1
Eq . ( 2 ) i n a biochemi cally meaningful fashion . Garrett et . al .
( 2 4 3 ) have tried to exp l ain why this l in;iting case of Eq , ( 1 3 )
i s obs erved during tetracyc l ine inhibi tion by postulating
that there are d i f ferences between rates of protein synthes i s
and the rates o f c e l l divis ion in inhibited cultures . The
present report has shown that this is mos t probab ly not true ,
result b , and i t would be helpful i f another hypothes is were
advanced to explain thi s adherence to the l imiting case o f
E q . ( 1 3 ) .
A model cons i stent with the results outl ined above has
been sought w i thin the proposed b i ochemical mode of action .
This mode l a s s umes that tetracyclines bind to the aminoacyl
site of ribosome s and thereby prevent the binding of AAtRNA .
The evidence for this mode of action has been presented in
the l i te rature survey . I f the usual type o f enzyme- inhibitor
kinetics is f o l l owed it would be expect�d that thi s mode o f
action would generate a n inverse re lationship between rate o f
protein synthe s i s and inhibitor concentration . However , a
growing E . coli culture i s not the normal in vitro test systero .
Such a culture w i l l not have constant concentrations o f s ubs trate
or enzyme s . It was thought , therefore , that a mode l which
a ll owed the se concentrations to vary with the ti�e of growth
and a l s o the inhibitor concentration , might lead to results
s imi l ar to that observed in the experimental cultures .

1 2 2
Conditions have not been found such that the proposed
model generates outputs which agree with the exper imental
data . The model wi l l be pre sented , however , s i nce it or a
s imple var iation of i t may be con s i s tent with the exper imenta l
observations when other conditions are tr ied .
The model , F ig . XXIV , i s pre sented i n terms of some
of the reactions i nvo lved in protein synthes i s in a bacter ial
culture . The s implest model invo lving the lea s t number of
reactions ha s been pos tulated as a starting point . Reac tants
are expres s ed in terms o f concentration i n bhe tota l culture .
The model s tates that there i s a rever s ible pa s sage of
tetracyc l ines from out s ide the c e l l s ( Tc ' ) into the c e l l s
( Tc ) , Eq . ( 14 ) . T h i s s tep ha s been ignored i n initial model s .
Tetracyc l ine s inside the c e l l s bind to r ibosome-RNA ( R ) and
form a complex ( R • Tc ) which i s not capable o f synthe s i z ing
protein , Eq . ( 1 5 ) . Norma l r i bo some s-mRNA complexes are , of
cour se , c apable of bind i ng sub s trate ( S ) , AAtRNA and forming
a peptide bond ( P ) on the r ibosoma l sur face , Eq . ( 1 6 ) .
Peptide bond f orma tion frees the aminoacyl s ite of the
r ibosome-mRNA complex and a l lows either tetracyc l i ne or AAtRNA
to bind .
Two steps , Eq . ( 17 ) and ( 1 8 ) , have been i nc orporated
i nto the mode l with the expectation that they might lead to
results d i f fe�ent from ordinary enzyme kinetic s . The f ir s t
o f these steps i s that substrate i s synthe s i z ed from substrate
precur sors ( S ' ) i n a reaction which ha s been pre sumed to
proceed at a rate proportiona l to the concentration of prote in

1 2 3
Tc ' K1 < Tc
Ka R + Tc � R ·Tc �
R + s & R · S k , , � p + R I k 8 .. s · C,P,
s
p Ca \ R
P;. * 'b ( R + R · S + R ·Tc ) + � p
F igure XXIV . Propo s ed Mode l o f Tetracyc l i ne Inhib i t ion of Pro-
t e i n S y n th e s i s i n Bacter i a l Cu l tur e s . Tc ' is the c oncentr a t i on
o f t e t r a c y c l ine i n the cu l ture bro t h , Tc i s the c oncentrat ion
w i th i n the c e l l s , R is r i bo s om e s - mRNA - peptidy l tRNA , S i s
AAtRNA , P i s t h e p e p t i d e bond c o n c e n tr a t i o n , S ' a r e AAtRNA pre-
cur s or s , S ' are AAtRNA d egradation produc t s , P t is the tot a l
pro t e i n c onc e ntra t i o n w i t h i n the c e l l s , q wa s 0 . 4 and oC wa s 2 0 .

1 2 4
i n the cul ture . That i s , the rate o f this synthes i s step has
been pre s umed to be proportional to P . The proportionality
constant uti l i zec , c 1 , i s a pseudorate cons tant and contains
in addi tion to the concentrations of the precursors , the
fraction of the total number of peptide bonds in the culture
whi ch are part of the enzymes cataly z ing this reaction , i . e .
aminoacyl synthetas es . S ubs trate i s also lost , non-enzymatically ,
in thi s mode l by hydrolys i s , Eq . ( 17) . The second d i f ferentiating
feature of thi s mode l is that it postulates that a fraction
o f the product peptide bonds are ribosomal precursor proteins
and that these prote ins give rise to new ribosomes , Eq . ( 1 8 ) .
The constant , c 2 , uti l i zed in thi s step i s also a pseudo-rate
constant which contains the concentration of rRNA in the
culture as we l l as the fraction of total peptide bonds which
are ribo s oma l peptides . However , the as sumption that c 1 or c2 are cons tants , which has been employed , is probably
not valid . The concentration of tRNA or rRNA per ce l l mis;ht
be re lative ly cons tant , but the concentration of the se species
in the culture is undoubtedly not constant . They are probab ly
increas ing at the s ame rate as the number of ce l l s in the
cul ture . Perh aps there are o f f setting change s in other subs trate
or ribosomal precur sors and there fore , thi s approach , because
of i ts s imp l i c i ty , was tr ied f i rs t .
The concentration o f tota l prote in ( Pt
) present i n the
culture has been a s s umed to be the sum of the ribosoma l
and non-r ibosomal prote in , Eq . ( 1 9 ) . For the se calculat ions
it was a s s umed that ribosomes were 4 0 % protein ( q ) and that

1 2 5
there are 2 0 peptide bonds in the average E . coli protein pc) .
The di f ferential equations , Eq . ( 2 0 ) through ( 2 6 ) ,
derived from thi s mode l are g iven in Table VI . They are too
complex to be solved by ordinary integration . They do
however , lead to a s imple express ion for the rate of total
prote in synthe s i s :
0/oe q ) [ P ] Eq . ( 2 7 ) d t
Thi s equation s tre s s e s the important role played by oc and
q in determining the nature of the Pt output in the model .
The above mode l i s based upon the concentration of the
various reactants in the total culture . I f one were to write
a s imi lar mode l based upon the concentrations per ce ll one
might be j us t i f ied in a s s uming a s teady-state in R . S 1 in
which case :
[ R · s I l [ S I l Eq . ( 2 8 )
+ [ S I l
where the s uperscript ( I ) i s used to de signate concentrations
per individual ce ll rather than in the total culture and Rt is the concentration of a l l forms of r ibosome s within the cel l .
While thi s equation cannot b e substituted into Eq . ( 2 7 ) , i t
ind icates that the model might b e tes ted under two general
conditions , when s is greater than Km and when S is less than
S ince ordinary integration methods are not use ful , the
model ha s been te s ted by analog s imulation . An unsealed
an�log computer program is given in Fig . XXV . The mode l is
too big ( too many integrations ) for the avai lable analog

Tab le V I . - The Different ial Equ a t i on s from a Mode l of
Tetracyc l in e Inhibit ion o f P rote in Synthes is in
Bacter ial Cu lture s .
d Tc 1 d t
d Tc d t
d R · Tc d t
d R d t
d s d t
d R •S d t
d p d t
k_ 1 (Tc )
k l (TC I � k_ 2 [R · Tc )
k 2 ( R ] (TC )
k_2 (R · TC ]
k_ 3 [R · S ]
k 3 [R ] [ S ]
k1
(Tc 1 )
k_ 1 (Tc ) - k2 (R ] (Tc ] +
k_ 2 [R · TC )
k2 [ R ] [TC ] + k4 [R · S ]
k 3 [R ] [ S ] + c 2 [P ]
+
1 2 6
Eq . ( 2 0 )
Eq . ( 2 1)
Eq . ( 2 2 )
Eq . ( 2 3 )
Eq . ( 24 )
Eq . ( 2 5 )
Eq . ( 2 6 )
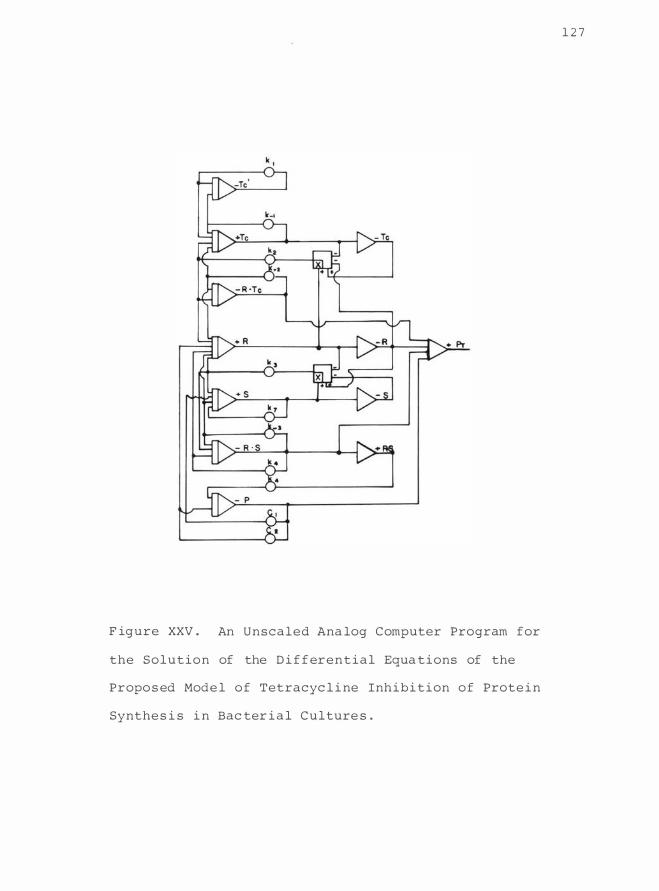
F igure XXV . An Unsealed Ana log Computer Program for
the Solution of the D i f f erential Equa tions of the
Propos ed Mode l of Tetracyc l i ne Inhibition of Protein
Synthes i s i n Bac ter ial Culture s .
1 2 7

computer1 3 and could only be part i a l ly tes ted with this
ins trument . I n add i tion , mos t analog computers are only
able to perform accurate calculations over about 2 log
cycle s . The s e d i f f iculties were overcome by us ing the
I nternational Bus iness Machines 1 1 3 0 digital computerl 4
with associ ated Continuous Sys tems Mode ling Package . The
actual program uti l i zed i s given , in F i g . XXVI , in ordinary
1 2 8
analog computer symbo l s . E i ghty analog e lements were avai lable
for use wi th thi s package and the output i s accurate over
several log cycle s .
S ince the value s for initial concentrations o f reactants
and rate constants which were tried did not lead to outputs
which agreed with the e xperimental data , the va lues will
not be de s cribed in detai l . The value s tried are l i s ted in
Table VI I . When Km was greater than S , no conditions could
be found where the output was s imilar to experimental data ,
even when Tc ' and Tc were zero . Whi l e i t was pos s ible to
obtain an exponential increase o f total prote in synthe s i s ,
ba lanced growth was not maintained . That i s , the ratio
of ( S ) to ( R) was not constant . These experiments were
unfortunately performed before the need to uti l i z e a value
o f � larger than l was rea l i zed . Inspection o f the results
reveals that thi s would not have changed the results o f the se
experiments , however , other condi tions might be found where
1 3 . TR 2 0 , Elec tron ic Assoc iates Inc . , Long Branch , N . J .
1 4 . We are grateful to Mrs . L i l l i an Kornhaber o f the Department o f B i ometry , Medical Col lege of Virginia , for performing the se calculations .

F igure XXVI . The Continuous Systems Modeling Package
Prog ram Used in the Search for Solutions to the
D i f f erential Equations from the Proposed Mode l of
Tetracyc l ine Inhibition of Prote in Synthe s i s in
Bac ter i a l Cultur e s .
1 2 9

Table VII . - C omp i l a t ion of C ond i t i on s Which Were Emp loyed to Test the Mode l of
Tetracyc l ine Inhib it ion of Prot e in Synthe s is in Bacter i a l C u lture s .
Ident ity # so Ro Po t k3 k_ 3 k4 � cl c 2 k
7
A 3 . 7 50 0 . 07 5 0 . 0 3 0 . 02 0 . 004 1 . 2 6 0 . 20 1 . 4 1 . 0 0 . 008
B 3 . 7 50 0 . 07 5 0 . 0 3 0 . 02 0 . 004 1 . 2 6 0 . 2 0 1 . 4 1 . 1 0 . 008
c 0 . 37 5 0 . 07 5 0 . 0 3 0 . 02 0 . 004 1 . 2 60 . 20 1 . 4 1 . 0 0 . 008
D 0 . 37 5 0 . 07 5 0 . 0 3 0 . 10 0 . 02 0 6 . 0 60 . 20 7 . 0 5 . 0 0 . 04 0
E 3 . 7 50 0 . 7 50 0 . 30 0 . 10 0 . 02 0 6 . 0 60 . 20 7 . 0 5 . 0 0 . 04 0
F 0 . 37 5 0 . 07 5 0 . 0 3 1 . 00 0 . 02 0 6 . 0 6 . 0 2 7 . 0 5 . 0 0 . 040
G 1 0 . 37 5 0 . 07 5 0 . 0 3 1 . 00 0 . 200 6 . 0 6 . 20 7 . 0 5 . 0 , 0 . 04 0 2 5 . 5 , 3 6 . 0 , 4 6 . 5
H 1 0 . 37 5 0 . 07 5 0 . 0 3 1 . 00 0 . 200 6 . 0 6 . [20 10 . 0 , 6 . 0 0 . 040 2 1 5 . 0
I 1 0 . 37 5 0 . 07 5 0 . 0 3 0 . 10 0 . 02 0 6 . 0 60 . 20 1 . 6 5 , 0 . 5 0 . 040 2 1 . 7 5 , 3 1 . 8 5
J 1 0 . 37 5 0 . 07 5 0 . 0 3 100 . 00 20 . 000 6 . 0 0 . 26 2 . 0 , 0 . 5 0 . 004 2 2 . 4 , 3 2 . 5 , f-'
w 4 3 . 0 0

Tab l e V I I . - ( cont . )
I . D . # so Ro R • S0 Po pg k 3 k_ 3 k4 � c l c 2 k7
K 1 0 . 37 5 0 . 07 5 - - 0 . 0 3 0 100 . 0 2 0 . 0 6 . 0 0 . 26 12 . 0 2 . 5 0 . 02 0
K 2 0 . 1 5 0 0 . 300 0 . 02 2 5 0 . 0 2 5 0 . 04 6 100 . 0 2 0 . 0 6 . 0 0 . 26 1 2 . 0 2 . 5 0 . 02 0
L 1 0 . 1 50 0 . 0 30 0 . 02 2 5 0 . 02 5 0 . 04 6 100 . 0 2 0 . 0 6 . 0 0 . 26 1 2 . 0 2 . 6 , 0 . 02 0 2 2 o 7 1 3 2 . 8 , 4 3 . 0 , 5 3 . 2 5 , 6 3 . 5
M l 0 . 1 50 0 . 0 30 0 . 0 2 2 5 0 . 0 2 5 0 . 04 6 100 . 0 2 0 . 0 6 . 0 0 . 26 1 2 . 0 3 . 5 0 . 2 50 , 2 0 . 500 , 3 1 . 000 , 4 2 . 500 , 5 5 . 000
N 0 . 1 3 5 0 . 0 30 0 . 017 5 0 . 0 5 5 0 . 074 100 . 0 2 0 . 0 6 . 0 0 . 2 6 2 . 5 0 . 5 0 . 004
0 0 . 4 0 5 0 . 090 0 . 0 5 2 5 0 . 0 5 5 0 . 1 1 2 100 . 0 20 . 0 6 . 0 0 . 2 6 2 . 5 0 . 5 0 . 004
p 0 . 100 0 . 0 28 0 . 1 100 0 . 0 55 0 . 0 18 100 . 0 2 0 . 0 6 . 0 0 . 2 6 2 . 5 0 . 5 0 . 004
Q 0 . 244 0 . 07 1 0 . 06 3 5 0 . 2 3 9 0 . 066 100 . 0 20 . 0 6 . 0 0 . 2 6 2 . 5 0 . 5 0 . 004
a . For exper iments A to Q inc lus ive va lues of kl , k_ 1 , k2 , k_2 , Tc ' , and Tc were not
used . For experiment s A t o 0 inc lus ive , was programmed as 1 . I-' w I-'

Tab l e V I I . - ( c on t . )
I dent ity # Tc 0 k2 k_2 Tc ' O kl
k_ l
Q 2 l 1000 . 00 2 0 0 . 000
Q 3 l , 2 , 3 100 . 00 2 0 . 00 0
Q 4 3 10 . 00 2 . 000
Q 5 3 1 . 00 0 . 2 0 0
Q 6 2 0 0 . 10 0 . 02 0
Q 7 20 , 80 , 240 0 . 0 1 0 . 00 2
Q 8 240 0 . 0 2 , 0 . 00 5 0 . 00 2
Q 9 240 0 . 0 1 0 . 00 1 , 0 . 004 , 0 . 0 10 , 0 . 020
Q 10 24 0 0 . 0 1 , 0 . 10
Q l l - 0 . 10 0 . 020 50 , 100 , l 0 . 1 150 , 200
b . For exper iment s Q 2 t o Q l l , inc l u s ive , the values of so , Ro , R · S0 , Po , 0 Pt , k 3 , k_ 3 , k4 , kffi , C 1 , C 2 , and k7 wh ich were u s ed were the s ame as those g iven above under Ident ity # Q .
f--' w N

1 3 3
thi s would not be true .
Conditions which allowed an exponential increase o f
total prote in and where ( S ) and ( R ) increased exponential ly
at the same rate were found when ( S ) was greater than K and m
Tc ' and Tc were equal to zero . However , when Tc ' or Tc were
di f ferent from zero , it was not poss ible to f ind conditions
o f Eq . ( 14 ) and/or Eq . ( 1 5 ) such that prote in synthes i s was
maintained at a reduced exponential rate . Rather the rate
of prote in synthe s i s a lways decreased initial ly and then
incre ased to the rate occurring in the ab sence of antibiot ic .
I t was not poss ible to f ind conditions which maintained the
i n i t i a l reduced rate of prote in synthe s i s for time s as long
as those seen experimental ly . I n addition , the rate constants
evaluated from the initial inhib ited protein synthe s i s , Tab le
VI I , did not have a l inear dependence upon the total antibiotic
concentration .
Several variations o f the mode l have been sugges ted
by the present work . When ( S ) is less than �m , value s of
rate cons tants were rej ected because the rate of prote in
synthe s i s was not f irst-order . The se non-exponential re sults
might have been caused by the value of oC uti l i zed . These value s
o f rate constants might lead to exponential protein synthes i s
and ( S ) and ( F ) might be produced in proportionate amounts i f
a larger number were uti l i zed for oC
When ( S ) i s greater than Km , the mode l as presently
s tated appe ars to be inherently balanced , i . e . after e s tab l ish
ment o f equi l ibria between antibiotic and ribosome s it re turns

1 3 4
to the rate o f prote in synthes i s preva i l ing be fore addition
o f antibiotic . Perha lt t 1 ps a erna e va ue s of rate cons tants
can be found which would prolong the time of establ i shment of
thi s equi l ibria . The s tatement o f the equi l ibria between
Tc ' and Tc is not exact as expre ssed in Eq . ( 1 4 ) . It has
only been used in a very l imited manner . Expre s s ion of thi s
equi l ibria in terms o f the concentration of tetracycl ine in
the broth ( Tc ' ) and the concentration within an individual
c e l l (Tci ), might be a de s i rable way of a l tering the present
model such that attainment o f equi l ibria between antib iotic
and ribosomes is not achieved .
I f the s e changes are not sufficient , then the e f fect
o f variation of the pseudo-rate cons tants , c1 and C 2 ,
\vi th time and w i th tetracycline concentration should be
cons i dered . The concentrations of e i ther tffiJA or rRNA in
the cul ture whi ch are contained in these " cons tants "
undoubtedly change during culture growth . I t seems l ikely
that tetracy c l ines would indirectly af fect these " constants "
by v irtue of the i r secondary inhibi t ion of nucleic acid
synthe s i s . The present work has uti l i zed the se parameters
as cons tants solely because this was the s imp l e s t and easiest
procedure . The introduction of such a mod i f i cation would
nece s s i tate a very large number o f integrations and is pro-
bab l y not practical . I t would require a great deal more
knowledge about metabol i c events within the inhibited bacterial
c e l l s than i s presently known in a quantitative manner . I t
should b e rea l i zed that the present work as we l l a s others

( 1 8 , 3 0 , 3 3 and 3 4 ) has measured e f fects on synthe s i s of
und i f ferenti ated nuc leic acids and not on the individual
species of ribonuc leic acids .
Another a l ternative to the present mode l would be to
formulate a s imi lar mode l on the bas i s of concentrations per
ce l l and then sum the se concentrations over the number o f
1 3 5
ce l l s which exi s t a t any given time . I n this type of a mode l
i t should be pos s ible to make certain s impl i fying assumptions :
such a s s teady- s tate concentrations o f various intermediates ,
perhaps R · S , tRNA and rRNA . The summation o f values over
indices which change with time and inhibitor concentration
doe s , however , present unusual problems in analog s imulation
and has not been uti l i zed in thi s s tudy .

1 3 6
V . Summary
S ixteen of the 1 8 tetracycl ine s under inve stigation
have been shown to inhibi t c e l l divis ion , prote in synthesis
and nuc l e i c ac id synthe s i s in cultures of exponenti a l ly
dividing E . c o l i W . The number o f cell s i n inhibited cultures
has been obtained as a function of time of growth by viable
and by total ce l l count methods . Sma l l diffe rences found between
the total number of c e l l s and the number of viable ce l l s are
not b e l i eved to be related to the e ffect of the tetracyclines
in these culture s . Rather the se d i fferences are felt to be
c aused by aggregation o f viable ce l l s during the assay for
v i ab l e c e l l s . A f i l te r technique us ing detergent-extracted
c e l lulose e s ter membranes was devel oped to isolate TCA-ethanol
precipitates of the E . col i ce l l s as a function of the time
o f growth . The hot TCA soluble extract o f these precipitate s
was used to measure nucleic ac ids by the orcinol reaction .
The hot TCA insolub l e material remaining on the fi lter was
tryps in-dige s ted and the protein content e s t imated by the
Fol in-Lowry a s s ay .
At l ow concentrations o f these tetracycl ines , inhibition
is produced after an initial time l ag during which no e ffect
on the te s ted culture parameters was seen . After thi s l ag
time , a l l tes ted culture parameters could be de scribed by
f i r s t order k inetic expre s s ions , such as ; Eq . ( 1 ) , ( 4 ) and
( 6 ) . Generation rate cons tants for te tracy c l ine- inhibi ted
cultures , kv , kt , kp , kna 1 have been obtained from the

1 3 7
l e a s t s quares f i t o f the exper imental data t o these equations .
I t s eems mos t l ike ly that generation rate constants obtained
for a l l culture parame ters are identical for any given
culture ; that i s , balanced growth is mainta ined . Thi s result ,
a lthough not s tr i ctly in agreement with previous reports i s
exp lained b y the f a c t that previous exper iments were e i ther
only s ingle-point measurements or were not determined over
a suffi c ient number of generations such that equi l ibrium
inhibitionwwas not observed by these workers .
Generation rate cons tants for a l l cul ture parameters
were found to be l inearly related to the concentration of
the tetracyc l ine ana log in the culture , Equations ( 2 ) , ( 5 )
and ( 7 ) . The proportional i ty constant s , k� , kf , k� , k�a ' o f
the s e re l ationships have been determined . When the data for
a l l culture parameters were treated together a s ingle
proportional ity cons tant , k�vg ' was obtained . Thi s value
i s a measure o f the antibacte r i a l potency o f the tetracycl ine
analog . These potencies are thought to be sui tab le for use
in s tructure-activi ty relationships s ince they are kinetic
cons tants determined by several d i f ferent a s s ay me thods .
The s imple mode l o f Garrett , Mi l ler and Brown ( 2 4 3 )
which attempts to expl a in the l inear re lationship o f
generation rate constants t o antibiotic concentration has
been pre sented . The i nadequacy o f the kinetic exp l anation
proposed by thi s mode l has been noted and attempts have been
made to derive a more satis fac tory mode l of te tracycl ine
inhib i tion . A mode l based on the b iochemical mode o f action

1 3 8
o f the tetracy c l ines was proposed . T h i s model was investigated
to see i f i t might l e ad directly to the l inear re lationship
o f generation rate constants to antibiotic concentration .
I t was fe l t that thi s might result from the inclusion of
reactions leading to the growth o f " subs trate " and " enzyme "
concentrations in add i tion to the usual enzyme-kinetic
reactions . Cond i t i on s such that the mode l generated output
cons i s tent with the experimenta l l y observed type of inhibition
were not found . O f course , a l l condi tions were not tes ted .
Several mod if i cations o f the mode l have been sugges ted such
that the experimental results might be exp la ined by thi s
type o f mode l but because o f the l arge number o f differential
equations which need to be s olved the se have not been te sted .
The time o f onset o f inhibi tion was approximately 3 0
minutes ( l l/2 generation s ) in these culture s . Thi s time
and the t ime o f recovery from inhibition upon di lution into
antibiotic- free med i a was observed to be shorter for the
inhibi tion of prote i n s ynthe s i s than for the inhibition of
cell divi s i on or nucl e i c acid synthe s i s . This difference in
time o f onset o f inhibi tion was espe c i a l ly pronounced when
the broth was changed such that the rate of growth was
reduced . Under the se condi t ions the time of onset o f
inhibi tion o f prote in synthe s i s d i d not vary whi le the time
o f onset o f inhibition o f c e l l divis ion and nuc leic acid
synthes i s were increased . An increase in broth pH vailiue ,
achieved by changing total phosphate as we l l as phosphate

ion ration , decreased the time of onset of inhibition of
c e l l mas s and c e l l div i s ion . These exper iments can be
mos t ea s i ly expla ined if one assumes that inhibition of
nuc leic ac id synthes i s and inhibi tion of cell divis ion are
the result of a pr ior inhibition of prote�n synthes i s . In
addi tion there i s a f inite lag time , which is broth pH
sensitive , before prote in synthe s i s is inhibited .
1 3 9
Thi s interpretation i s cons i s tent with , and these
resu l t s are further evidence tha t , the pr imary mode of ac tion
of tetracyc lines is to cause an inhibition of protein synthes i s .
S inc e a l l tetracyc l ine s behaved qua l i tatively the same it
seems mo st l ikely that they are a l l acting by the same
mechani sm . However , this cannot be said with certa inty
s ince the d i fferences between time of onset of inhibi tion
of protein synthes i s and the times of onset of inhibition of
c e l l d iv i s ion and nucleic acid synthe s i s were only measured
for culture s inhibited by oxytetracyc line . Indeed it should
be emphas i zed that these conc lus ions with regard to the mode
of act ion of oxytetracycl i ne are based on a s ing le exper iment ,
F ig . XXI .

1 4 0
Bib l iography .
l . Sne l l , J . F . and Cheng , L . , " Stud ies on Modes of Act ion
o f Tetracyc l ine ( II ) " , Deve lop. Ind . Microb ial . , 1 , 107
( 19 6 1 ) .
2 . Gale , E . F . , "Mechan isms of Ant ib iot ic Ac t ion " ,
Pharmaca l . Rev . , �' 4 8 1 ( 1 9 6 3 ) .
3 . Frank l in , T . J . , "Mode of Ac t ion of the Tetracyc l ine s " ,
Syrnpos . Soc . Gen . Microbial . XVI , B iochemic a l Studies of
An t imicrob i a l Drugs , 1966 .
4 . Parth ier , B . , "Wirksungsmechan ismen und Wirksungspektren
der An t ib iotica im Protein und Nuc le in Saurestoffwechse l " ,
Pharma z ie , lQ, 4 6 5 ( 19 6 5 ) .
5 . Barrett , G . c . , " Synthes is of Tetracyc line Analogs " ,
J . Pha rm . Sc i . , 21, 309 ( 19 6 3 ) .
6 . Boothe , J . H . , "Chem is try and B iolog ic a l Act iv i t ie s of
the Tetracyc l in e s " , in "An t imicrob i a l Agents and
Chemotherapy " , Sylvester , J . C . , Ed . , ( 19 6 2 ) pp . 2 1 3 .
7 . P lakunov , V . K . , "Correlat ion of Structure and Mechan i sm
of Act iv ity in the Series of Tetracyc l ine Ant ib iot i cs " ,
Dok l ady Akademi i Nauk SSSR , 1 5 1 , 1207 ( 1 9 6 3 )
8 . Hochstein , F . A . , Stephens , C . R . , Conove r , L . H . ,
Regn a , P . P . , Pasternack , R . , Bru n ings , K . J . and

Woodwar d , R . B . , " Terramyc in VI I . The Structure of
Terramyc in " , J . Am . Chern . Soc . , 74 , 3708 ( 1952 ) .
1 4 1
9 . Donohue , J . , Dun itz , J . D . , Trueblood , K . N . and Web ster ,
M . S . , " The C rystal Structure of Aureomyc in
(Chlorotetracyc l ine) Hydrochlor ide . Con j ugat ion ,
Bond D is tances and C onformat ion " , J . Am . Chern . Soc . ,
8 5 ' 8 5 1 ( 1 9 6 3 ) .
10 . Remmers , E . G . , S ieger , G . M . and Doerschuk , A . P . ,
" S ome Obs e rvat ions on the Kinet ic s of the C - 4
Epimerizat ion o f Tetracyc l ine Ant ib iot ic s " , J . Pharm .
Sc i . , 2£, 7 52 ( 196 3 ) .
l l . C o l a izz i , J . L . , Kneve l , A . M . and Mart in , A . N . ,
" B iophys ic a l Study of the Mode of Act ion of the
Tetracyc l ine An t ib iot ic s " , J . Pharm . Sc i . , 54 , 14 2 5 ( 196 5 ) .
1 2 . Albert , A . , " Av id ity of Terramyc in and Aureomyc in for
M6!t a l l ic Cat ions " , Nature , 17 2 , 2 0 1 ( 19 5 3 ) .
1 3 . Stephens , C . R . , Mura i , K . , Brun ings , K . J . and
Woodward , R . B . , "Ac id ity C on stants of the Tetracyc line
An t ib iot ic s " , J . Am. Chern . Soc . , 2§_, 4 1 55 ( 1 9 5 6 ) .
14 . Lee s on , L . J . , Krueger , J . E . and Nash , R . A . ,
"Concern ing the Structura l As s ignment of the Second
and Th ird Ac id ity C onstants of the Tetracyc l ine
Ant ib io t ic s " , Tetrahedron Letters , 18 , 1155 ( 19 6 3 )
1 5 . R ig ler , N . E . , Saswat i , P . B . , Leyden , D . E . , Sudme ir ,
J . L . and Re i l ley , c . N . , "Determ inat ion of a Protonat ion

1 4 2
Scheme of Tetracyc l ine Us ing Nuc lear Magnetic Resonance " ,
Ana l . Chern . , 22 , 8 7 2 ( 19 6 5 ) .
16 . Ke s s e l r ing , U . W . and Benet , L . Z . , "Determinat ion of a
Protonat ion Scheme for Isoch lortetracyc l ine " , Abstracts
of the Am. Pharm . As s oc . , Acad . Pharm . Sc i . Meet ing ,
Miami Beach , F la . , May , 1968 .
17 . Lask in , A . I . , " Tetracyc l ines " , in "Ant ibiot ic s ,
Mechan i sms of Act ion " , va l . l , Got t l ieb , D . and Shaw ,
P . D . , Ed . , Spr inger-Verlag Inc . , New York , N . Y . ( 1967 )
pp . 3 3 1 .
18 . Benbough , J . and Morr ison , G . A . , "Bacter iostat ic Act ions
of Some Tetracyc l ines " , J. Pharm . Pharmacal . , 1.2. ,
4 0 9 ( 19 6 5 ) .
1 9 . Has h , J . H . , Wishn ick , M . and M i l ler , P . A . , "On the
Mode of Ac t ion of the Tetracyc l ine Ant ibiot ic s in
Staphylococcus aureus " , J . Biol . Chern . , 2 3 9 , 2070 ( 1964 )
20 . Garrett , E . R . , Person a l Commun icat ion ; and Personal
Observat ion of Unpub l ished Data .
2 1 . Jones , J . G . and Morrison , G . A . , " The Bacteriosta t ic
Ac t ions of Tetracyc l ine and Oxytetracyc l ine " , J . Pharm .
Pharmac a l . , 1± , 808 ( 19 6 2 ) .
2 2 . c iak , J . and Hahn , F . E . , "Mechan isms o f Act ion of
Ant ib i ot ic s I . Addit ive Ac t i on of Chloramphenicol and
Tetracyc l ines on the Growth of Escherichia c o l i " ,
J . Bacterial . , J..2, 1 2 5 ( 19 58 ) .

1 4 3
2 3 . Garrett , E . R . and Brown , M . R . w . , "The Act ion of
Tetracyc l ine and Chloramphen icol Alone and in Admixture
on the Growth of Escher ichia coli " , J . Pharm . Pharmacal . ,
.1:2 , l8 5T ( l96 3 ) .
24 . Eag le , H . and Saz , A . K . , "An t ib iot ics " , Ann . Rev .
Microb i a l . , ,9 , l7 3 ( 1 9 5 5 ) .
2 5 . Cheng , L . and Sne l l , J . F . , " Studies in Metabol ic
Spec t ra IV . Ef fects of Tetracyc l ines , Some of The ir
Der ivat ives and Chloramphen icol on Accumu lat ion of
G lutamic Ac id in Escherichia c o l i " , J . Bacterial . ,
8 3 , 7 1 1 ( 1962 ) .
2 6 . C rema , A . and Baro l i , F . , "Az ione De lla Terramic ina
S u l l a Respirazione Di Alcu n i Germ i E Di Alcuni Tessut i " ,
Bu l l . S oc . Ita l . Biol . Sper . , 2 9 , 2 9 9 ( 19 5 3 ) .
2 7 . Wong , D . T . o . , Barban , S . and Aj l , S . , " Inhibit ion of
Respiration of Chlortetracyc l ine and Oxytetracyc line" ,
Ant ib io . and Chemotherapy, 1, 3 0 7 ( 19 5 3 ) .
2 8 . G a le , E . F . and Pa ine , T . F . , "Effect of Inh ib itors and
Ant ib iot ic s on G lutam ic Ac id Accumu lat ion and on Prote in
Synthe s i s in Staphylococcus aureus " , Biochem . J . , 4 7 ,
XXVI ( 1 9 5 0 ) .
2 9 . Hahn , F . E . and Wis seman , c . L . , " Inh ib i t i on of Adapt ive
Enzyme Forma t ion by Ant im ic rob ia l Agents " , B iochem . J . ,
2_l, 4 9 3 ( 19 5 3 ) .
30 . Gale , E . F . and Folke s , J . P . , " The Ass im i lat ion of

Am ino Ac ids by Bacteria 1 5 . Ac t ions of Ant ibiot ic s on
Nuc l e ic Ac id and Protein Synthes is in staphylococcus
aureus " , Biochem . J . , �� 4 9 3 ( 19 5 3 ) .
3 1 . Park, J . T . , Inhib it ion of C e l l-Wa l l Synthes is in
S t aphylococcus aureus by Chemicals Wh ich Cause
Acc umu lat ion of Wa l l Precursors " , B iochem . J . , J.!}_ ,
2 P ( 1 9 5 8 ) .
3 2 . Sazyk in , Yu . 0 . and Bor isova , G . N . , "The Effect of
3 3 .
Bacter ios tat ic Ant ib iot ic s on Synthes is of Prote in
and Nuc l e ic Ac ids in the C e l l s of Staphylococcus
aureus " , Ant ib iotik i , ]_ , 1 1 ( 1962) thru Chern . Abstr .
6 1 ; 86 5 2 e .
v' / Cerny , R . and Habermann , V . , " On the Effects of
Tetracyc l ine on the B iosynthe s i s of Proteins and
Nuc l e ic Ac ids with Escherichia c o l i and Bac i l lus
c e reus " , C ol l . C zech . Chern . Commun . , �� 1 3 2 6 ( 1964 ) .
34 . Holmes , I . A . and W i ld , D . G . , " The Synthes is of
R ibonucleic Ac id Dur ing Inhib it ion of Escher ichia
c o l i by Chlortetracyc l i ne " , Biochem . J . , !}2, 2 7 7 ( 196 5 )
3 5 . Holme s , I . A . and W i ld , D . G . , "Con sequences of
Inh ib i t ion of Escher ichia c o l i by Tetracyc l ine
Ant ib i o t ic s " , Nature , 2 10 , 104 7 ( 1966) .
36 . Holme s , I . A . and Wild , D . G . , " Inhibit ion of the
Growth of Esche richia c o l i by Chlortetracyc l ine " ,
B iochem . J . , 104 , 6 7 9 ( 1967 ) .
1 4 4

3 7 . Grunberger , D . , Skoda , J . and Sorm , F . , "Mechan isms o f
Ant ib iot ic Act ion V . Effeet o f Chloramphen ic ol ,
Ch lortetracyc l ine and Oxytetracyc l ine on the Synthe s is
of G l ut amic Ac id Decarboxy lase in E scher ichia coli
1 4 5
and of Tyros ine Decarboxylase i n Streptomyc in faeca lis " .
Chern . L i s ty , 48 , 1 7 1 1 ( 19 54 ) thru Chern . Abstr . , 4 9 : 4 78 3d .
3 8 . Loom i s , W . F . , " On the Mechan i sm o f Act ion of
Aureomyc in " , Sc ienc e , l l l , 4 7 4 ( 19 50 ) .
3 9 . Brody , T . M . and Ba in , J . A . , " The Effect of Aureomyc in
and Terramyc in on Ox idat ive Phosphory lat ion " , J .
Pharmac a l . Expt l . Therap. , 10 3 , 388 ( 19 5 1 ) .
4 0 . Van Meter , J . C . , Spec tor , A . , O l e s on , J . J . and
Wi l l iams , J . H . , " In vitro Ac t ion of Aureomycin on
Oxidat ive Phosphory lat ion in An ima l T i s sues " , Proc .
Soc . Expt l . B io l . and Med . , 81 , 2 1 5 ( 19 5 2 ) .
4 1 . Brody , T . M . , Hu rwitz , R . and B a in , J . A . , "Magnes ium
and the Effect of the Tet racyc l ine Ant ib iot ic s on
Oxida t ive Proc e s s e s in Mitochondria " , Ant ib i o . and
Chemotherapy, i · 8 64 ( 19 64 ) .
4 2 . Miura , Y . , Nakamura , Y . , Matsua i ra , H . and Kone i j i , T . ,
"The Mode of Ac t ion of oxytetracyc l ine " , Ant ib i o . and
Chemotherapy , i · 864 ( 19 54 ) .
4 3 . Brody , T . M . , " Uncoupl ing of Ox idat ive Phosphorylat ion " ,
Pharmacal . Rev . , 2 · 3 3 5 ( 19 5 5 ) .
44 . Hunter , F . E . and Lowry , 0 . H . , " Ef f ects of Drugs on

1 4 6
Enzyme Systems " , Pharmacal . Rev . ,_ .§_ , 8 9 ( 1 9 5 6 ) .
4 5 . Goma zkov , 0 . A . , " The Mechanism o f Action of Tetracyc line
Antibiotics " , Vopr . Hed . Khim . , 2_ , 3 9 3 ( 1 9 6 3 ) thru Fed .
Proc . ( Trans . Suppl . ) � ' T 8 7 6 ( 1 9 6 4 ) .
4 6 . Santi , R . , " Drugs with Selective Action on Oxidative
Phosphoryl ation " , Arch . Ital . S c i . Farmacol . , g , 5 ( 19 6 2 )
thru Chern . Abs tr . , � : 7 2 5 8c .
4 7 . Morikawa , F . , Sakata , S . , Shikano , H . , Kuriyama , K . and
Fuj i , Y . , " E f fect o f Some Antibioti c s on Oxidative
Phosphorylation in L iver Homogenate " , Kyoto Furitsu Ika
Daigaku Z a s s hi " , �' 2 2 5 ( 1 9 5 9 ) thru Chern . Abs tr . � : 4 0 4 6 a .
4 8 . Saz , A . K . and Maramur , J . , " The Inhibition o f O rganic
N i tro-Eeductase by Chlorte tracyc l ine in C e l l - Free Extracts " ,
Proc . Soc . Exptl . B iol . and Med . , � ' 7 8 3 ( 1 9 5 3 ) .
4 9 . S a z , A . K . and S l i e , R . B . , " t-�angane se Rever s a l o f
Chlortetracycl ine Inhibi tion o f Bacterial C e l l -Free
N i tro-Reducta se " , J . Am. Chern . Soc . , 7...2. , 4 6 2 6 ( 1 9 5 3 ) .
5 0 . S a z , l L K . and S l ie , E . B . , " Reversal o f Ch lortetracyc l ine
I nhib i t ion of Bacterial C e l l- Free N i tro-Reductase by
Mangane s e " , J . Biol . Chern . , 2 1 0 , 4 0 7 ( 1 9 5 4 ) .
5 1 . S a z , A . K . and S l i e , R . B . , " The I nhibition o f Organic Nitro
Reductase by Chlortetracycl ine in C e l l-Free Extracts I I .
Cofactor Requirements for the N itro-Reductase Enzyme Complex " ,

5 2 .
Archiv . Biochem . B iophys . , 21 . 5 ( 1 9 54 ) .
S a z , A . K . and S l ie , R . B . , " Inhib it ion of Organ ic
Nitro-Reduc tase by Chlortetracyc l ine in Bacte r i a l
C e l l-Free Extracts " , Antib iot ics Ann . , 3 0 3 ( 1 9 5 3 ) .
5 3 . Sa z , A . K . , Browne l l , L . W . and S l ie , R . B . ,
"Aureomyc in-Res istant C e l l-Free Nitro-Reductase from
Aureomyc in-Re s is tant Escher ich ia col i " , J . Bacter ia l . ,
n . 4 2 1 ( 19 56 ) .
54 . Saz , A . K . and Mart inez , L . M . , "En zymat ic Bas is of
Res is t ance to Chlortetracyc l ine I . Difference Between
F lavoprote in in Nitro-Reductases of Sens it ive and
Res i s tant Escher ich ia coli " , J . B iol . Chern . , 2 2 3 ,
2 8 5 ( 19 5 6 ) .
1 4 7
5 5 . P l akunov , V . K . , "B iochemical Changes i n Metabo l ism of
S taphylococcus aureu s 209P Connected with Re s i s tance to
Tetracyc l ines " , Doklady Akademi i Nauk SSR, 1 5 1 , 952 ( 19 6 3 )
56 . Yag i , K . , Okuda , J . , Ozawa , T . and Okada , K . , "Mechan ism
of Inh ib i t ion of D-Amino Ac id ox idase " , Biochim . B iophys .
Acta . , 34 , 3 7 2 ( 1 9 5 9 ) .
57 . saz , A . K . and Mart inez , L . M . , "En zymat ic Bas is of
Res is tance to Chlortetracyc l ine I I . Inhib i t ion of
E lectron Transport in Escher ic hia coli by Aureomyc in " .
J . Biol . Chern . , 222 · 1020 ( 1 9 58) .
58 . saz , A . K . and Mart ine z , L . M . , " En zymat ic Bas is of
Res i s tanc e to chlortetracyc l ine I I I . Inh ib it ion by

Aureomyc in of Prote in - S t imu lated Elec tron Tran sport " ,
J . Bacter ial . , J.:i, 527 ( 1 960) .
1 4 8
5 9 . V i l lanueva , J . R . , "Attempt s t o Show Inhibition of
60 .
6 1 .
6 2 .
Nitro-Reduc tase by Ch lortetracyc l ine in C e l l-Free
Ext rac t s and Isolat ion of an Extrac t f rom Chlortetracyc line-
Re s is tant Nocardia V " , Nature , 1 9 3 , 95 ( 196 2 ) .
!,...- ./ Kremery , V . and Brunettova , H . , "Mechanism of the
Act ivity of Tetracyc l ine Ant ibiotics V . Effect of
Oxytetracyc l ine on the Reduct ion of Tr iphenyltetrazol ium
Chlor ide by Staphylococcu s and Some Other Organ isms " ,
Fo l ia Microb ial . , 2, 2 2 2 ( 1 964 ) thru Chern . Abs t r . , �: 15068 .
\.;- ./ Kremery , V . and Ke llen , J . , "Changes in Some Enzymes of
Bacteria l E lec tron Tran sport Accompanying Deve lopment
of Res is tance to Oxytetracyc l ine " , J. Bacter ial . , � ,
1 2 64 ( 19 6 6 ) .
v ,. Krcmery , V . , "Mechan i sms of Act ion of Tetracyc line
Ant ib i ot ic s V I I I . Absence of Te l lur ium Reductase in
Oxyte tracyc l ine-Res istant Var iants of Escherichia
c o l i and Staphylococcus aureus " , Cezk . Epidemol .
Mikrobiol . Imunol . , l2 , 2 14 ( 1966 ) thru Chern . Abs tr .
6 5 : l 90 2 6 f .
3 v _,
v d Ke l len J "The Mechan ism of Ac t ion of 6 . Krcmery , . an , . ,
Tetracyc l ine Ant ib iot ics IX . Inh ib it ion of Cytochrome
ox idase by Oxytetracyc l ine " , J . Hyg . Epidemiol .
Mic�obiol . Immunol . ( Praque ) , 10 , 3 3 9 ( 1 966 ) thru Chern .

1 4 9
Abst r . , 66 : 7 3 507 t .
64 . Izak i , K . , Ar ima , K . and K iuch i , K . , " Spec if ic ity and
Mecha n i sm of Tetracyc l ine Res istance in a Mu ltip le Drug
Res is tant Stra in of Escher ichia c o l i " , J . Bacter ial . ,
2.!_, 6 2 8 { 1966 ) .
6 5 . Fra nk l i n , T . J . and Godfrey , A . , " Re s istance of
Escher ichia c o l i to Tetracyc l ines " , Biochem . J . , 94 ,
54 ( 1 9 6 5 ) .
66 . C onnamacher , R . H . , Mande l , H . G . and Hahn , F . E . ,
"Adapta t i on of Popu lat ions of Bac i l lus cereus to
Tetracyc line " , Mol . Pharmaca l . , l_ , 586 ( 1967 ) .
6 7 . Wat s on , J . D . , " The Synthe s is of Proteins Upon R ibosomes " ,
Bu l l . S oc . Chim . Biol . , 4 6 , 1 3 9 9 { 1964 ) .
6 8 . Moldave , K . , "Nuc le ic Ac ids and Protein Biosynthes is " ,
Ann . Rev . Biochem . , ]i, 4 19 ( 19 6 5 ) .
6 9 . Schweet , R . and He intz , R . , " Prot e in Synthes i s " , ib id ,
12_ , 7 2 3 ( 1966 ) .
7 0 . Att a rd i , G . , " The Mechanism of Prote in Synthes i s " , Ann .
Rev . Microb i a l . , 11, 383 { 1967 ) .
7 1 . G i lbert , w . , " Po lypept ide Synthes i s in E . Col i , I .
R ibosomes and t he Ac t ive Complex " , J . Mol . Biol . , .§_,
3 7 4 ( 19 6 3 ) .
7 2 . Hurwit z , c . and Rosano , C . L . , " The Intrace l lu lar
c oncent rat ion of Bound and Unbound Magnes ium Ions in
Escher ichia c o l i " , J . Biol . Chern . , 24 2 , 3 7 1 9 ( 1967 ) .

1 5 0
7 3 . Miyazawa , T . and Tamok i , T . , " Revers ible Dissoc iat ion
of Escher ichia c o l i R ibosomes by p-Chloromercuribenzoate "
J . Mol . Biol . , 24 , 4 8 5 ( 1967 ) .
74 . Petermann , M . L . and Pavlovec , A . , " The Effect of
Temperature on the Magne s ium B ind ing and Ult racentrifuga l
Proper t ie s of R a t L iver R ibosome s " , Biochem . , .§_, 2 9 50
( 1967 ) .
7 5 . Moore , P . B . , " S tudies on the Mechan i sm of Messenger
R ibonuc leic Ac id Attachment to R ibosome s " , J. Mol .
B i ol . , 2 2 , 14 5 ( 1966 ) .
7 6 . Geste land , R . F . and S taehe l in , T . , " Elect rophoret ic
Ana lys i s of Proteins from Norma l and Ces ium Ch lor ide
Treated Esche r ichia c o l i R ibosome s " , ib id , 24 , 14 9 ( 1967 ) .
7 7 . Trau t , R . R . , "Acry lam ide Ge l E lectrophores is of
Rad ioact ive R ibosomal Prot e in " , ib id , £.!_, 57 1 ( 1966)
78 . Ross et , R . , Mon ier , R . and Ju l ien , J . , " Les R ibosomes
D ' Esche r ich ia col i . I . - Mise En Ev idence D ' Un RNA
R ibosom ique De F a ible P o ids Molecula ire " , Bu ll . Soc .
Chim . B io l . , 4 6 , 87 ( 1 964 ) .
7 9 . Suthe rland , B . M . , C a r r ie r , W . L . and Setlow , R . B . ,
"Nuc leot ide Sequence of KB C e l l 5 S RNA " , Sc ienc e , 1 58 ,
169 5 ( 1967 ) .
80 . Brown lee , G . G . and ·sanger , F . , "Nuc leot ide Sequences
from the Low Mo lec ular We ight R ibosoma l RNA from
Escher ichia c ol i " , J . Mol . Biol . , n, 3 3 7 ( 19 6 7 ) .
,

8 1 . Brown lee , G . G . , Sanger , F . and Barrel l , B . G . ,
" Nuc leot ide Sequence of 5-8-Ribosomal RNA from
Escher ichia c o l i " , Nature , 2 1 5 , 7 3 5 ( 1967 ) .
82 . C a ntor , C . R . , " The Extent of Base Pa ir ing in 5 s
R ibosomal RNA " , P roc. Nat . Acad . Sc i . u . s . , _22,
4 7 8 ( 1968 )
8 3 . Takanam i , M . , "Nuc leot ide Sequences at the 5 ' -termin i i
of Escher ichia c o l i R ibosomal RNA " , J . Mol . B io l . , � �
3 2 3 ( 1 967 ) .
1 5 1
84 . Sug iura , M . and Takanam i , M . , "An a ly s i s o f the 5 ' -termina l
Nuc leotide Sequences of Ribonuc leic Ac ids , II .
C ompa r is on of the 5 ' -terminal Nuc leot ide Sequences of
R ibosoma l RNA ' s from Different Organ isms " , Proc . Nat .
Acad . Sc i . U . S . , 58 , 1 5 9 5 ( 1967 ) .
8 5 . Felsenfeld , G . and Miles , T . H . , " The Phys ic a l and
Chemica l Properties of Nuc le ic Ac ids " , Ann . Rev .
B iochem . , �, 407 ( 1 9 6 7 ) .
86 . Wal ler , J . P . and Harr is , J . I . , " S tudies on the
C ompos it ion of the Prote in from Escherichia c o l i
R ibosome s " , Proc . Nat . Acad . Sc i . U . S . , 4 7 , 18 ( 1961 )
87 . M�l ler , W . and Chambach , A . , " Phys ica l Heterogene i ty
o f the R ibosomal P roteins from Esche r ichia c o l i " ,
J . Mol . B io l . , fl, 3 7 7 ( 1967 ) .
88 . Sp i r in , A . s . , " The Structure of R ibos omes " , Abhand l .

Deu t . Akad . Wis s . Ber l in , Kl . Med . , 16 3 , ( 1964 ) thru
Chern . Abs t r . , g: l4 94 6 f .
1 5 2
8 9 . Atsman , A . , Spinik-Els on , P . and Elson , D . , "Characterizat ion
of the Particu late and Free Proteins Obta ined After
Treatment of Ribosomes with 2 �-Lithium Chlor ide " ,
J . Mol . Biol . , £2, 1 6 1 { 1967 ) .
90 . Traub , P . , Nomura , M . and Tu , L . , " Phys ica l and
Func t iona l Heterogene ity of R ibosoma l Prote ins " ,
ibid , 1 9 , 2 15 ( 1966 ) .
9 1 . Foge l , S . and Sypherd , P . S . , "Chemica l Bas is for
Heterogeneity of R ibos omal Prote ins " , Proc . Nat . Acad .
Sc i . , U . S . , 5 9 , 1 3 2 9 { 1968) .
9 2 . Mo l ler , w . and Widdowson , J . , "Fract ionation Studies
of the R ibos oma l P rote ins from Escher ichia coli " , J .
Mo l . B io l . , li' 367 { 1967 ) .
9 3 . Raska s , H . J . and Staeh l in , T . , "Messenger and sRNA
B ind ing by Ribosoma l Subunits and Rec on s t ituted
R ibosome s " , ib id , ll' 8 9 { 196 7 ) .
94 . Traub , P . , Hosokawa , K . , Craven , G . R . and Nomura , M . ,
" Structure a nd Funct ion of Escher ichia c o l i R ibosomes IV .
Isolat ion and Characterizat ion o f Function a l ly Ac t ive
R ibosoma l Prote in s " , Proc . Nat . Acad . Sc i . U . S . , ��
24 30 ( 1967 ) .
9 5 . Traub , P . and Nomura , M . , "Structure and Funct ion of
Escher ichia c o l i R ibos omes V . Rec on s t itut ion of

Functional l y Active 3 0 S P ibosomal Particles from RNA
and Proteins " , ibid , 21, 7 7 7 ( 1 9 6 8 ) .
1 5 3
9 6 . Staeh l in , T . and Me s e l son , M . , " De termination o f S treptomyc in
S e n s i t ivity by a Subunit of the 3 0 S Ribosome of
E s cherichia col i " , J . Hol . Biol . , �' 2 0 7 ( 1 9 6 6 ) .
9 7 . Traub , P . , Hosokawa , K . and Nomura , '1 . , " S treptomycin
Sens i tivity and the S tructural Components of the 30 S
Ribosome o f E s cherichia col i ' ' , ibid , � ' 2 1 1 ( 1 9 6 6 ) .
9 8 . Marinkovic , D . , " S treptomycin Res i s tance Mutation in
E . co l i : Alte red Ribosomal Prote in '' , Sc ience , 1 6 0 ,
1 9 8 ( 1 9 6 8 ) .
9 9 . Suzuka , I . , " E f fect o f r:thylenediaminetetraacetate on
E . co l i B Ribosome s-Binding o f Phenyl alanyl sRNA to 5 0
S Partic l e s Converted from 7 0 S Ribosome s " , B iochem .
B i ophys . Res . Commun . , � , 6 6 7 ( 1 9 6 7 ) .
1 0 0 . Otaka , E . , I toh , T . and Osawa , S . , " P rotein Components
in the 4 0 S Ribonuc leoprotein Particles in E . col i " ,
1 0 1 .
Sc ience , 1 5 7 , 1 4 5 2 ( 1 9 6 7 ) .
Gol dber<J , A . , " Hagnes ium B inding by E . c o l i Ribosome s ' ,
J . Mol . Biol . , !2• 6 6 3 ( 1 9 6 6 ) .
1 0 2 . Cho i , Y . S . and Carr , C . W . , " Ion-bind ing Studies of
Hibonucl e i c Ac id and E s cherichia col i Ribosome s " ,
ibid , �' 3 3 1 ( 1 9 6 7 ) .
1 0 3 . Geste l and , H . F . , " Unfo lding o f Escherichia col i
Hibosor:1e s by Hemoval o f Magne s ium'' , ibid , !..§_, 3 5 6 ( 1 9 6 6 ) .

104 . C onwa y , T . W . , "On the Role of Ammon ium or Potass ium
Ions in Amino Ac id Polymer izat ion " . Proc . Nat . Acad .
Sc i . U . S . , 2_1, 1 2 16 ( 1964 ) .
10 5 . Spyr ides , G . J . , "The Ef fec ts of Un iva lent Cat ions on
the B ind ing of sRNA to the Template-Ribosome comp lex " ,
ib id , 2_1 , 1 2 2 0 ( 1964 ) .
106 . Reve l , M . and Hiatt , H . H . , "Magnes ium Requ irement for
the Format ion of an Act ive Mes senger RNA-Ribosome-sRNA
C omplex " , J . Mol . Biol . , Q, 467 ( 196 5}.
107 . Gordon , J . and Lipmann , F . , "Role of Diva lent Ion s in
Poly U-Directed Phenyla lan ine Polymer izat ion " . ibid ,
n_ , 2 3 ( 1 967 ) .
1 5 4
108 . Jacob , F . and Monad , J . , " Genetic Regu latory Mechan isms
in the Synthes is of Prot e in s " , ibid , l_ , 3 18 ( 196 1 ) .
109 . S impson , M . V . , " Prote in B iosynthes is " , Ann . Rev .
B ioc hem . , l.l , 3 3 3 ( 19 6 � .
1 1 0 . S inger , M . F . and Leder , P . , "Messenger RNA : An Eva lua t ion " ,
ib id , � . 1 9 5 ( 1966�
1 1 1 . Kuff , E . L . and Roberts , N . E . , " In vivo Labe l l ing
Patterns of Free Polyribosomes : Re lat ionsh ip to the Tape
Theory of Mes s enger R ibonuc le ic Ac id Funct ion " , J. Mol .
B i ol . , l.§_ , 2 11 ( 19 67 ) .
1 1 2 . Nathans , D . , Notan i , G . , Schwartz , J . H . and Z inder ,
N . D . , "B iosynthes is of the Coat Proteins of Col iphage
f 2 by E . c o l i Extracts " , Proc . Nat . Acad . Sc i . U . S . ,

1 5 5
4 8 , 14 24 ( 1962 ) .
1 1 3 . Nirenberg , M . W . and Matthae i , J . H . , "The Dependence
of C e l l-Free Prote in Synthe s i s in E . coli upon Natura lly
Occurr ing or Synthet ic Polyr ibonuc leot ides " , ib id , 4 7 ,
1 588 ( 19 6 1 ) .
1 14 . N i renberg , M . W . and Leder , P . , "RNA C odewords and
1 1 5 .
1 1 6 .
Prote in Synthe s i s . The Effects of Tr inuc leot ides upon
the B inding of sRNA to Ribosomes " , Sc ience , 14 5 , 1 3 9 9 ( 1964 )
" . S o l l , D . , Cheray 1 l , J . D . and Bock , R . M . , "Studies on
Polynuc leot ides LXXV . Spec i f ic ity of tRNA for Codon
Rec ogn i t ion a s Studied by the R ibosoma l B inding Techn ique " ,
J . Mol . B iol . , �, 97 ( 1 96 7 ) .
s(;l l , D . and Ra j Bhandary , U . L . , " Studies on Polynuc leot ides
LXXVI . Spec i f ic ity of Transfer RNA for Codon Recogn i t ion
as S tud ied by Amino Ac id Incorporation " , ib id , 2 9 , 1 1 3 ( 1967 )
1 1 7 . We i sb lum , B . , Gonan o , F . , von Ehren s t e in , G . and Benzer ,
s . , "A Demons t rat ion of C od ing Degeneracy for Leuc ine
in the Synthe s is of Prote in " , Proc . Nat . Acad . Sc i .
u . s . , 21, 328 ( 196 5 ) .
1 18 . Ke l log , D . A . , Doctor , B . P . , Loebe l , J . E . and Nirenberg ,
M . w . , "RNA C odon s and Prote in Synthe s i s IX , Synonym
c od on Recogn it ion by Mu l t iple Spec ies of Va l ine- ,
Alan ine- , Methionine-sRNA " , ib id , �, 9 12 ( 1966 ) .
1 19 . c r ick , F . H . c . , "Codon-An t icodon Pa ir ing : The Wobble
Hypothe s is " , J . Mol . B iol . , J:.2, 548 ( 1966 ) .

1 5 6
1 20 . Ghos h , H . P . , Sol l , D . and Khorana , H . G . , " Studies on
Polynuc leot ides LXXVI I . In i t ia t i on of Prote in Synthe s is
in v itro a s Studied by Us ing Ribopolynuc leot ides with
Repeat ing Nuc leot ide Sequences as Messengers " , ib id , �, 2 7 5 ( 1967 ) .
12 1 . McE lroy , W . D . , Deluca , M . and Trav is , J . , "Molecu lar
Un iform ity in B iolog ica l Cata lyses " , Sc ience , 157 , 150
( 19 6 7 ) .
1 2 2 . Nove l l i , G . D . , "Am ino Ac id Act ivat ion for Protein
Synthes is " , Ann . Rev . B ioc hem . , �, 44 9 ( 1967 ) .
12 3 . Chapev i l le , F . , Lipmann , F . , von Ehrenstein , G . ,
We isb lum , B . , Ray , W . J . Jr . and Benzer , s . , " On the
Role of S olub le R ibonuc leic Ac id in Cod ing for Amino
Ac ids " , Proc . Nat . Acad . Sc i . U. S . , 48 , 1086 ( 196 2 ) .
124 . Holley , R . W . , Apg a r , J . , Everett , G . A . , Mad ison , J . T . ,
Marqu isee , M . , Merri l l , S . H . , Penswick , J . R . and
1 2 5 .
Zam i r , A . , " S tructure of a R ibonuc le ic Ac id " . Sc ience ,
14 7 , 14 62 ( 1 9 6 5 ) .
zachau , H . , Dutt ing , D . and Fe ldman , H . , "Nuc leotide
Sequences of Two Serine-Spec i f ic Trans fer R ibonuc le ic
Ac ids " , Angew . Chern . , J..!i, 3 9 2 ( 1966) , thru Chern . Abstr . ,
6 5 : 2 5 l2b .
1 2 6 . Mad i s on , J . T . , Everett , G . A . and Kung , H . , "Nuc leot ide
Sequence of a Yeas t Tyros ine Trans fer RNA " , Sc ience ,
1 5 3 , 5 3 1 ( 1966 ) .

1 5 7
1 2 7 . Raj Bhandary , R . M . and Khorana , H . � . , ' S tudie s on
Po lynucleotides LXXVI I I . The P r imary S tructure o f Yeast
Phenylalanine Trans fer RNA '' , Proc . Nat . Acad . S c i .
u . s . , � . 7 5 1 ( 1 9 6 7 ) .
1 2 8 . Cannon , M . , " The Ribosomal B inding S i te for Peptidyl
Tran s fer-Ribonuc l e i c Acid " , B iochem . J . , 1 0 4 , 9 3 4 ( 1 9 6 7 ) .
1 2 9 . Ful ler , W . and Hodgson , A . , " Conformation of the
Anticodon Loop in t-RNA" , Nature , 2 1 5 , 8 1 7 ( 1 9 6 7 ) .
1 3 0 . Okamoto , T . and Takanami , M . , " Interaction o f Hibosome s
and Natural Polyribonucleotide s " , Bioch i rn . Biophys .
Acta , 7.i._ , 2 6 6 ( 1 9 6 3 ) .
1 3 1 . Moore , P . B . , " Polynucleotides Attachment to Ribosome s ' ,
J . Mol . B i ol . , 1 8 , 8 ( 1 9 6 6 ) .
1 3 2 . Moore , P . B . and Asano , K . , " Divalent Cation-dependent
Binding of Me s senger- to Ribosoma l - RNA" , ibid , !§_ ,
2 1 ( 1 9 6 6 ) .
1 3 3 . S zer , �\1 . and Nowak , L . , " Interaction o f Polyur idyl i c
A c i d Analogue s with E s cherichia col i Ribosome s " , ibid ,
� . 3 3 3 ( 1 9 6 7 ) .
1 3 4 . Wal ler , J . P . , " The NH2 -Terminal Res idues o f the
Proteins from C e ll -Free Extracts of E . col i " , ibid ,
7_, 4 8 3 ( 1 9 6 3 ) .
1 3 5 . Marcker , K . and Sanger , F . , " N-F'orrnyl-Methionyl· - sRNA. ' ,
ibid , � . 8 3 5 ( 1 9 6 4 ) .
1 3 6 . Marcker , K . , " The Formation o f N-Formy l-�ethionyl - s RNA ' · ,
ibid , .!_i , 6 3 ( 1 9 6 5 ) .

1 5 8
1 3 7 . C la rk , B . F . C . and Marcker , K . , "Cod ing Response of
N-Formy l -Methiony l-sRNA to UUG " , Nature , 207 , 10 38 ( 19 6 5 ) .
1 38 . C lark , B . F . C . and Marcker , K . , "The Role of N
Formyl-Meth iony l-sRNA in Protein B iosynthesis " , J. Mol .
B io l . , Q, 3 94 ( 1966 ) .
1 3 9 . Adams , J . M . and C apecchi , M . R . , "N-Formylmethiony l
s RNA a s the In it iator of Protein Synthe s is " , Proc .
Nat . Aaad . Sc i . u . s . , 22, 14 7 ( 1 966 ) .
140 . Web ster , R . E . , Enge lhardt , D . L . and Z inder , N . D . ,
" In vitro Prot e in Synthe s is : Cha in Init iat ion " , ib id ,
22 , 1 5 5 ( 1966 ) .
14 1 . C apecch i , M . R . , " In i t iat ion of E . c o l i Prote ins " , ib id ,
2,.? , 1 5 17 ( 1966 ) .
14 2 . We i s sbach , H . and Red f ie ld , B . , "De formylation of
N-Formy l Methion ine by Escher ichia coli Extracts " ,
B i ochem . Biophys . Res . C omm . , 22 , 7 ( 19 6 7 ) .
14 3 . Lugay , J . C . and Aron s on , J . N . , "Am ino Ac id Acy lases :
Proposed Re lat ion to Metabolic Recyc l ing of Methion ine
dur ing Prot e in B iosynthe s is " , B iochem . B iophys . Res .
Commun . , £1 , 4 3 7 ( 1967 ) .
144 . Vi;u e l a , E . , Salas , M . and Ochoa , S . , ' 'Tran s lat ion of
the Genet ic Me ssage , I I I . Formylmethion ine as In it iator
of Prote ins Programed by Polyc is tron ic Mes s enger RNA " ,
Proc . Nat . Acad . Sc i . U . S . , 22, 7 2 9 ( 1967 ) .

14 5 .
146 .
1 5 9
Sundarara j an , T . and Thach , R . , " Role o f Formylmethion ine
C odon AUG in Phas ing Trans lat ion of Synthet ic mRNA " ,
J . Mol . B io l . , 1 9 , 74 ( 1966 ) .
Thach , R . E . , Dewey , K . F . , Brown , J c and D t p • • 0 y , . '
" Formy lmethion ine Codon AUG as an In itiator of Polypept ide
Synthes is " , Sc ienc e , 1 5 3 , 4 16 ( 1966 ) .
14 7 . Lede r , P . a nd Burs ztyn , H . , " In it iat ion of Protein
Synthe s i s , I . E f fec t of Formylat ion of Methiony l-tRNA
on C odon Rec ogn i t i on " , Proc . Nat . Ac ad . Sc i . u . s . , 5 6 ,
1 5 7 9 ( 1966 ) .
148 . Nomura , M . , Lowry , C . V . and Guthr ie , C . , "The In it iat ion
of Prote in Synthes is : Join ing of the 50 S Ribosoma l
Subun i t t o the In i t iat ion Comp lex " , Proc . Nat . Acad .
Sc i . u . S . , .2.§_ , 1487 ( 1967 ) .
14 9 . Nakamoto , T . and Kolakofsky , D . , "A Pos s ib le Mechan ism
for In i t iat ion of Prote in Synthes is " , ib id , 2_2, 606 ( 1966)
1 50 . Kolakofsky , D . and Nakamot o , T . , "The In it iat ion of
Vira l P rote in Synthes is in Escher ichia c o l i Extracts " ,
ibid , �. 1786 ( 1966 ) .
1 5 1 . c lark , B . F . c . and Marcker , K . A . , "N-Formyl-Meth ionyl-
s - R ibon u c l e i c Ac id and Cha in In i t iat ion in Protein
B iosynthe s i s " , Natu re , 2 1 1 , 3 7 8 ( 1966 ) .
1 5 2 . Sa las , M . , M i l l e r , M . , Wahba , A . and Ochoa , S . ,
" Tran s lat ion of the Genet ic Message , V . Effect of
Magne s ium Ion s and Formy lat ion of Methion ine in Prot e in

1 6 0
Syn thes is " , Proc . Nat . Acad . Sc i . U . s . , 21._, 186 5 ( 1967 )
1 5 3 . Economou , A . E . and Nakamoto , T . , "Further Stud ies on
the Init iat ion of Prot e in Synthes is with N-Formy l
Met ion ine in E . coli Extracts " , ib id , 58 , 10 3 3 ( 1967 ) .
1 54 . S tanley , W . M . , Sa las , M . , Wahba , A . J . and Ochoa , s . ,
"Tran s lat i on of the Genet ic Message : Factors Involved
in the In it iat ion of Prot e in Synthes is " , ib id , ��
2 90 ( 1966 ) .
1 5 5 . S a las , M . , H i l le , M . , Las t , J . A . , Wahba , A . and
Ochoa , S . , "Trans lat ion of the Genet ic Mes s age , I I .
Effec t of In itiat ion Fact ors on the B inding of
Formyl-Methionyl-tRNA to R ibosomes " , ib id , 21._ , 387 ( 1967 ) .
156 . Matthae i , H . and Vo igt , H . , "Mechan i sm in Protein
Syn t he s i s , IV . Further Ev idence for Two Di fferent
R ibosomal S ites , One B ind ing Formy lmeth ionyl-tRNA ,
the other Meth ionyl- and other Am inoacyl-tRNA ' s " ,
B iochem . B iophys . Res . C ommun . , 2 8 , 7 30 ( 1967 ) .
1 5 7 . Bre t s c her , M . S . and Marcker , K . A . , " Polypept idy l
sRNA and Aminoacyl-sRibonucleic Ac id Bind ing S ites on
R ibosome s " , Natu re , 2 1 1 , 380 ( 1966 ) .
1 5 8 . .Zam ir , A . , Leder , P . and E l s on , D . , "A Ribos ome-Cata lyzed
Reac t i on Between N-Formy lmethionyl- tRNA and Pu romyc in " ,
Proc . Nat . Acad . Sc i . U . S . , 56 , 1 7 94 ( 1966) .
1 5 9 . Hershey , J . W . B . and Thach , R . E . , "Role of Guanos ine-
5 ' -Triphospnate in the Init iat ion of Pept ide Synthes is ,

I . Synthe s is of Formylmethionyl -Puromyc in " , ib id , !32, 7 59 ( 1967 ) .
1 6 1
160 . Anders en , J . S . , Bret scher , M . s . , C lark , B . F . c . and
Marcker , K. A . , "A GITP Requ irement for Binding In it iator
tRNA t o Ribosomes " , Nature , 2 1 5 , 490 ( 1967 ) .
16 1 . Thach , R . E . , Dewey , K . F . and Mykolaj ewycz , N . , "Role
of Guanos ine - 5 ' -Tr iphosphate in the Initiat ion of
Pept ide Syntne s is , II . Syntne s is of Dipept ides " . Proc .
Nat . Acad . Sc i . U . S . , !32, 1 10 3 ( 19 6 7 ) .
1 6 2 . Leder , P . and Nau , M . M . , " In it iat ion of Protein Synthe s is ,
I I I . Fac tor - GTP - Codon-dependent Bind ing of Formyl
Methiony l -tRNA to R ibosomes " , ibid , 58 , 7 74 ( 1967 ) .
1 6 3 . E isenstadt , J . M . and Brawerman , C . , "A Fac tor from
Esche r ichia c o l i Concerned wit h the S t imu lat ion of C e l l
Free Polypept ide Synbhe s i s by Exogenous Ribonuc leic
Ac id , I . Ev idence for t he Occurrence of a S t imu lat ion
Factor " , Biochem . , 2, 2 7 77 ( 1966 ) .
164 . Brawerman , G . and Eisen s tadt , J . M . , "A Factor from
Esche r ich ia c o l i Concerned with the S t imu lat ion of C e l l
Free Polypept ide Synthes i s by Exogenous Ribonuc leic
Ac id , II . Charac terist ic s of the React ion Promoted by
the S t imulat ion Factor " , ib id , 2 , 2 7 84 ( 1966 ) .
1 6 5 . Reve l , M . and Gros , F . , "A Factor from E . c o l i Requ ired
for the Trans lat ion of Natura l Messenger RNA " , B iochem .
Biophys . Res . C ommun . , £2, 124 ( 1966) .

1 6 6 . Reve l , M . and Gros , F . , " The St imu lat ion by Ribosomes
of DNA Transcription : Requ irement for a Trans lat ion
Factor " , ib id , rJ.. , 12 ( 1967 ) .
1 6 2
167 . Ka j i , H . , Suzuka , I . and Ka j i , A . , "Bind ing o f Spec i f ic
Solub le R ibonuc l e ic Ac id to Ribosomes " , J . B io l . Chern . ,
24 1 , 1 2 5 1 ( 19 6 6 ) .
168 . Pestka , S . and Nirenberg , M . , "Regu latory Mechan isms
and Prote in Synthes is X . Codon Rec ogn ition on 30 s
R ibosomes " , J . Mol . Biol . , �' 14 5 ( 1966 ) .
16 9 . Suzuka , I . , Kaj i , H . and Ka j i , A . , " B ind ing of a
Spec i f ic sRNA to 30 S Ribos oma l Subun its : Effect of
50 S R ibosoma l Subunits " , Proc . Nat . Acad . Sc i . U . S . ,
� ' 148 3 ( 1966 ) .
1 7 0 . Mattha e i , H . and Milberg , M . , "Mechan isms in Protein
Synthe s is V . Ev idence for Two Ribosoma l S ites from
Equ i l ibria in B ind ing of Pheny la lanyl-tRNA " , B iochem .
B i ophys . Res . C ommun . , �' 593 ( 1967 ) .
1 7 1 . Iga rashi , K . and Ka j i , A . , " On the Nature of Two
R ibosoma l S ites for Spec i f ic sRNA B ind ing " , Proc .
Nat . Acad . Sc i . U . S . , 58 , 197 1 ( 1967 ) .
1 7 2 . Nomura , M . and Lowry , c . V . , " Phage f2 RNA-D irected
B ind ing of Formy l-Methiony l-tRNA to Ribos omes and the
Role of Ribosoma l Subun its in In i t iat ion of Protein
Synthe s is " , ib id , 58 , 946 ( 1967 ) .

1 7 3 .
1 7 4 .
1 7 5 .
1 7 6 .
1 7 7 .
1 7 8 .
1 7 9 .
1 8 0 .
Nakada , D . and Kaj i , A . , " Function and Properties o f
the " Native " 3 0 S and 5 0 S Ribosomal Subunits o f
E sche r i chia col i " , ibid , 'i}__ , 1 2 8 ( 1 9 6 7 ) .
E i sens tadt , J . and Brawerman , G . , " The Role o f the
Native S ubribosomal Particles of E s cherichia coli in
Pol ypeptide Chai n Initiation " , ibid , �' 1 5 6 0 ( 19 6 7 ) .
Mangi arotti , G . and Sch l e s s inger , D . , " Polyribosome
Metabo l ism in E s cherichia co l i I . Extraction o f
Po lyribosomes and Ribosoma l Subunits from Fragi le ,
Growing E sche r i ch i a col i " , J . Mol . Biol . , �' 1 2 3 ( 1 9 6 6 )
Mangi aro tt i , G . and Sch l e s s inger , D . , ' · Polyribosome
Metabo l i sm in E scherichia c o l i I I . Formation and
L i fetime o f Me s senger RNA Molecule s , Ribosomal Subunit
Coup l e s and Polyribosome s " , ibid , �' 3 9 5 ( 1 9 6 7 ) .
Kaempfer , R . , Mes e l s on , M . and Raskas , H . , " Cy c l i c
D i s soc i a tion into S table Subunits and Re formation o f
Ribosomes During Bac terial Growth " , ibid , 2!, 2 7 7 ( 1 9 6 8 ) .
H i l le , M . B . , Mi l ler , M . J . , Iwasak i , K . and Wahba , A .
J . , " Tran s l ation o f the Genetic Me s s age , VI . The Role
of Ribosomal S ubuni t s in B inding of Formy l-Methiony l-
tRNA and i t s Reaction with Puromyc in " , Proc . Nat . Acad .
S c i . U . S . , � ' 1 6 5 2 ( 1 9 6 7 ) .
1 6 3
Ghos h , H . P . and Khorana , H . G . , " S tudi e s on Po lynuc leotides
LXXXIV . On the Ro le o f Ribosoma l Subunits in Prote in
Synthe s i s " , ibid , �' 2 4 5 5 ( 1 9 6 7 ) .
�' 3 4 4 ( 1 9 6 8 ) .
c , f . Addendum , ibid ,
Ohta , T . , Sarkar , s . and Thach , R . E . , " The Role o f
Guano s ine� S ' -Triphosphate i n I n i t i ation of Peptide
Synthe s i s , I I I . B inding of Formyl emethionyl-tRNA to

Ribosome s " , ibid , �' 1 6 3 8 ( 1 9 6 7 ) .
1 8 1 . Warne r , J . R . and Rich , A . , " The Number o f Soluble RNA
Molecules on Re ticulocyte Polyr ibosomes " , ibid , 2_!, 1 1 3 4 ( 1 9 6 4 ) .
1 8 2 . Bretsche r , M . S . , " The Chemical Nature o f the s RNA
Polypeptide Compl ex " , J . Hol . Biol . , 7_, 4 4 6 ( 1 9 6 3 ) .
1 8 3 . Marshal l , R . E . , Caskey , C . T . and Nirenberg , M . ,
1 6 4
" F ine S tructure o f RNA Codewords Recogni zed by Bacterial ,
Amphibian and Mamma lian Trans fer R.."JA" , Sc ience , 1 5 5 ,
1 8 4 .
8 2 0 ( 1 9 6 7 ) .
Sol l , D . , Jone s , D . s . , Ohtsuk a , E . , Faulkner , R . D . ,
Lohrmann , R . , Hayats u , H . , Khorana , H . G . , Cherayi l ,
J . D . , Hamp e l , A . and Bock , R . M . , " Specificity o f
s RNA f o r Recogn i tion o f Codons as Studied by the
R ibosomal Binding Technique " , J . Hol . Biol . , �' 5 5 6
( 1 9 6 6 ) .
1 8 5 . Gilbe rt , W . , " Polypeptide Synthe s i s in E . coli I I . The
Polypeptide Chain and s RNA " , ibid , .§_, 3 8 9 ( 1 9 6 3 ) .
1 8 6 . Cannon , M . , Krug , R . and Gilbert , W . , " The Binding of
sRNA by E . coli Ribosome s " , ibi d , 7_, 3 6 0 ( 1 9 6 3 ) .
1 8 7 . We tts te in , F . 0 . and Nol l , H . , " Binding of tRNA to
Ribosomes Engaged in Prote in Synthe s i s : Number and
P roperties of Ribosomal Binding S ites " , ibid , 1 1 ,
3 5 ( 1 9 6 5 ) .
1 8 8 . Ar l i nghaus , R . , Shae f fer , J . and Schweet , R . ,
" Me chani sm o f Peptide Bond Formation in Polypeptide
Synthe s i s " , Proc . Nat . Acad . U . S . , 2_! , 1 2 9 1 ( 1 9 6 4 ) .

1 8 9 .
1 9 0 .
1 9 1 .
1 9 2 .
1 9 3 .
1 9 4 .
1 9 5 .
1 9 6 .
Lucas -Lenard , J . and Lipmann , F . , " In i tiation of
Po lyphenylalanine Synthe s i s by N-Acetyl-Phenylalanine
s RNA" , i b id , 'i]_, 1 0 5 0 ( 1 9 6 7 ) .
Rave l , J . Jvl . , " Demons tration of a Guanos ine- 5 ' -
Triphosphate- dependent Enzymatic B inding of Aminoacyl
Ribonucl e i c Acid to E s cherichia coli Ribosome s " , ibid ,
'i]_ , 1 8 1 1 ( 1 9 6 7 ) .
Luc a s -Lenard , J . and Haenni , A . -L . , " Requirement of
Guanos ine- 5 ' -Triphosphate for Ribosomal Binding of
Aminoacyl s RNA " , ibid , �' 5 5 4 ( 1 9 6 8 ) .
E r te l , R . , B ra t , N . , Redfield , B . , Allende , J . E . and
We i s sbach , H . , " Binding of Guano s ine- 5 ' -Triphosphate
1 6 5
b y Sol ub l e F actors Required for Polypeptide Synthe s is " ,
ib i d , � ' 8 6 1 ( 1 9 6 8 ) .
Sutte r , R . P . and Ho ldave , K . , " The Interaction of
Arninoacyl Trans ferase I I and Ribosome s " , J . Biol . Chern . ,
2 4 1 , 1 6 9 8 ( 1 9 6 6 ) .
I buki F . Gas ior , E . and Holdave , K . , " The I nteraction ' ' ) o f Aminoacy l Soluble Ribonuc leic Aci d w i th Aminoacyl
Tran s ferase I " , ibid , 2 4 1 , 2 1 8 8 ( 1 9 6 6 ) .
Rao , P . and t-1oldave , K . , " The B inding of Aminoacyl
s RNA and GTP and T rans ferase I " , B iochern . Biophys . Res .
Commun . , � ' 9 0 9 ( 1 9 6 7 ) .
A l l ende , J . E . , Honro , R . and Lipmann , F . , " Resolution
of the E s cherichia col i Aminoacyl sRNA Transfer Factor
into Two Complementary Fractions " , Proc . Nat . Acad . Sci .
u . s . , �' 1 2 1 1 ( 1 9 6 4 ) .

1 9 7 .
1 9 8 .
1 9 9 .
2 0 0 .
2 0 1 .
2 0 2 .
2 0 3 .
Allende , J . E . and Wei s sbach , H . , " GTP Interaction
with a Protein Synthe s i s Initiation Factor Preparation
f rom E s cherichia col i " , Biochem . Biophys . Re s . Commun . ,
� ' 8 2 ( 1 9 6 7 ) .
1 6 6
Allende , J . E . , Seeds , N . W . , Conway , T . w . and
We i s sbach , H . , " Guano s ine- 5 ' -Triphosphate Interaction
with an Amino Aci d Polymeri zation Factor from E scherichia
col i " , Proc . Nat . Acad . S c i . U . S . , � ' 1 5 6 6 ( 1 9 6 7 ) .
Rave l , J . M . , Shorey , R . L . and Shive , �v . , " Evidence
for a Guanine Nucleotide -Aminoacyl-RNA Complex as an
I n termediate in the Enzymatic Transfer of Aminoacyl-
RNA to Ribosome s " , Biochem . Biophys . Res . Commun . , �'
6 8 ( 1 9 6 7 ) .
Gordon , J . , " Interaction o f Guanos ine - 5 ' -Triphosphate
w i th a Supernatant Fraction f rom E s cherichia coli and
Aminoacyl - s RNA " , Pro c . Nat . Acad . S c i . U . S . , � '
1 5 7 4 ( 1 9 6 7 ) .
Gordon , J . , " A S tepwise Reaction Yielding a Complex
B e tween a Supernatant Fraction from E . col i , Guanosine-
5 ' -Triphosphate and Aminoacyl - sRNA " , ibid , �' 1 7 9 ( 1 9 6 8 )
Monro , R . E . and Harcker , K . A . , " Ribosome-Catalyzed
Reaction o f Puromycin with a Formylmethionine-containing
O l i gonucleotide " , J . Mol . Biol . , �' 3 4 7 ( 1 9 6 7 ) .
Monro , R . E . , " Catalys i s o f Peptide Bond Formation by
5 0 s Ribosomal Subunits from E s cherichia col i " , ibid ,
2 6 , 1 4 7 ( 1 9 6 7 ) .
2 0 4 . N i sh i z uka , Y . and Lipmann , F . , " The Interrelationship
Between Guanos ine Triphosphatase and Amino Acid

1 6 7
2 0 5 .
Polyme r i z ation " , Arch . B iochem . Biophys . , 1 1 6 , 3 4 4 ( 1 9 6 6 ) .
Lucas -Lenard , J . and Lipmann , F . , " Separation of Three
Microb i a l Amino Aci d Polymeri zation Factors " , Proc . Nat .
Acad . S c i . U . S . , �' 1 5 6 2 ( 1 9 6 6 ) .
2 0 6 . Garen , A . , " S ense and Nonsense in the Genetic Code " ,
S c ience , 1 6 0 , 1 4 9 ( 1 9 6 8 ) .
2 0 7 . L as t , J . A . , S tanley , W . M . , Salas , M . , H i l le , M . ,
Wahba , A . J . and Ochoa , S . , " Trans lation o f the Genetic
Mes s age , I V . UAA as a Chain Termination Codon " , Proc .
Nat . Acad . S c i . U . S . , �' 1 0 6 2 ( 1 9 6 7 ) .
2 0 8 . P e s tka , S . , " S tudi e s on the Formation of tRNA-Ribosome
Complexe s , I I . A Pos s ible S i te on the 5 0 S Subunit
Prote cting Aminoacyl tran s fer Ribonucleic Acid f rom
Deac y l at i on " , J . Biol . Chern . , 2 4 2 , 4 9 3 9 ( 1 9 6 7 ) .
2 0 9 . Rych l ik , I . , " Re lease o f Lys ine Peptide s by Puromycin
f rom P o lylysyl-t RNA in the Presence o f Ribosomes " .
Biochim . B iophys . Acta , 1 1 4 , 4 2 5 ( 1 9 6 6 ) .
2 1 0 . Kuriki , Y . and Kaj i , A . , " Spec i f i c Binding of s RNA to
Ribos ome s during Polypepti de Synthe s i s and the Nature
o f Peptidyl sRNA " , J . Mol . Biol . , �, 4 0 7 ( 1 9 6 7 ) .
2 1 1 . Capec chi , H . R . , " Po lypeptide Chain Termination in vi tro :
I so l ation o f a Re lease Factor " , Proc . Nat . Acad . S c i .
u . s . , 2.§_, 1 1 4 4 ( 1 9 6 7 ) .
2 1 2 . Brets che r , M . S . , "Po lypeptide Chain Termination : An
2 1 3 .
Active Proce s s " , J . Mol . Biol . , l!r 1 3 1 ( 1 9 6 8 ) .
Rendi , R . and Ochoa , S . , " E f fe c t o f Chloramphenicol on
P ro te in Synthe s i s in C e l l- Free Preparations o f
E s cherichia coli " , J . Biol . Chern . , 2 3 7 , 3 7 1 1 ( 1 9 6 2 ) .

1 6 8
2 1 4 . Frank l in , T . , " The Inhib i tion of I ncorporation o f
Leucine into P ro te in o f Cel l-Free Systems from Rat
Liver e,nd E . c o l i by Chlortetracy c l in e " , B iochem . J . ,
� . 4 4 9 ( 1 9 6 3 ) .
2 1 5 . Laskin , A . I . and Chan , W . M . , " Inhibi tion by
Tetracy c l ines o f Polyuridy l i c Acid D i rected Phenylalanine
Incorporation in E . c o l i C e l l- Free Sys tems " , Biochem .
Biophys . Re s . Commun . , !! • 1 3 7 ( 1 9 6 4 ) .
2 1 6 . Suare z , G . and Nathan s , D . , " Inhibi tion o f Aminoacyl-
sRNA B inding to Ribosome s by Tetracy c l ine " , Biochem .
Biophys . Re s . Commun . , � , 7 4 3 ( 1 9 6 5 ) .
2 1 7 . Day , L . E . , " Tetracy c l ine I nhibition o f Ce l l- Free
P rotein S yn the s i s , I I . E f fe c t o f the B i nding of
Tetracy c l ine to the Components o f the System" , J .
Bacte r i a l . , � , 1 9 7 ( 1 9 6 6 ) .
2 1 8 . H i e rows k i , M . , " Inhibition o f Protein Synthe s i s by
Chlortetracyc l ine in the E . col i in vitro Sys tem" ,
Proc . Nat . Acad . S c i . u . S . , 2l• 5 9 4 ( 1 9 6 5 ) .
2 1 9 . Okamoto , S . and Mi zuno , D . , "Me chani s m o f Chloramphenicol
and Tetracy c l ine Re s i s tance in E . col i " , J . Gen .
Microbial . , 3 5 , 1 2 5 ( 1 9 6 4 ) .
2 2 0 . Maxwe l l , I . H . , " Partial Removal o f Bound Tran s fer
2 2 1 .
RNA f rom Polys omes Engaged in Prote in Synthe s i s in
vi tro a fter Add i tion of Tetracycl ine " , Biochim .
Biophy s . Acta . , 1 3 8 , 3 3 7 ( 1 9 6 7 ) .
Rif ino , C . B . , Bousque t , W . F . , Kneve l , A . M. , Be lcastro ,
P . and Martin , A . N . , " E f fe c t of Certain Tetracy c l ine
. 1 4 . b h . h . Analogs on Phenyl a l an 1ne-c I ncorporat1on y E s c er1c 1 a

1 6 9
c o l i B C e l l - Free Extracts " , J . Pharrn . s c 1· . , 5 7 _ , 3 5 1 ( 19 6 8 ) .
2 2 2 . Turley , R . , Thomas , P . E . and Snel l , J . P . , " Oxytetracycl ine
S timulate d I ncorporation of Cyste ine and Tryptophan
i nto an in vitro P rotein Synthe s i z ing Sys tem" , Biochern .
Biophys . Res . Cornmun . , �, 4 6 0 ( 1 9 6 7 ) .
2 2 3 . Connarna che r , R . and Mande l , H . G . , " Binding of
Tetracy c li n e to the 30 S Ribosome s and to Polyuridylic
Acid " , ibid , �, 98 ( 1 9 6 5 ) .
2 2 4 . Kohn , K . W . , " Mediation o f Divalent Me tal Ions in the
B inding o f Tetracy c l ine to Macromolecules " , Nature ,
1 9 1 , 1 1 5 6 ( 1 9 6 1 ) .
2 2 5 . Maxwe l l , I . H . , " E ffect of Tetracycl ine on Pulse
Label l ing o f Polysornes w i th H3 -urid ine in Intact Cel l s
o f � · iriegateriurn KM " , B iochirn . B iophys . Acta , 1 3 8 ,
3 2 9 ( 1 9 6 7 ) .
2 2 6 . Gottesman , M . E . , " Reaction o f Ribosome-Bound Peptidyl
Trans fer Ribonuc l e i c Ac i d with Arninoacyl Trans fer
Ribonucl e i c Acid or Puromy c in " , J . Biol . Chern . , 2 4 2 ,
5 5 6 4 ( 1 9 6 7 ) .
2 2 7 . Coutsogeorgopoulos , C . , " Inhibi tion o f Puromycin Re lease
by GTP " , Biochern . B iophys . Re s . Cornmun . , �, 3 5 2 ( 1 9 6 7 ) .
2 2 8 . Go ldberg , I . H . and Jl1itsug i , K . , " Sparsornycin Inhibition
o f Polypeptide Synthe s i s Promoted by Synthetic and
Natural Polynuc leotide s " , B iochern . , §_, 3 7 2 ( 1 9 6 7 ) .
2 2 9 . Gol dberg , I . H . and Mitsug i , K . , " Inhibi tion o f
Sparsornyc i n and o ther Antibiotics o f the Puromy cin- Induced
Re lease of Polypeptide from Ribosome s " , ibid , 6 , 3 8 3
( 1 9 6 7 ) .

1 7 0
2 3 0 . Vazque z , D . and Monro , R . E . , � E f fects o f Some Inhibi tors
of Protein Synthe s i s on the Binding of Arninoacyl tRNA
to Ribo s omal S ubuni t s � , Biochirn . Biophys . Acta , 1 4 2 .
1 5 5 ( 1 9 6 7 ) .
2 3 1 . C l ark , J . M . Jr . and Chang , A . Y . , " Inhibitions of the
Tran s f e r of Amino Acids f rom Arninoacyl Soluble
,Ribonucl e i c Acid to Proteins " , J . Biol . Chern . , 2 4 0 ,
4 7 3 4 ( 1 9 6 5 ) .
2 3 2 . Wol fe , A . D . and Hahn , F . E . , � In f l uence o f Inhibitors
of P ro te i n Synthes i s on the Me s senger-directed Binding
o f Arninoacyl s RNA to Ribo s ome s " , Fed . Proc . , �' 2 1 7
( 1 9 6 5 ) .
2 3 3 . Las t , J . A . , I zaki , K . and Sne l l , J . F . , " The Fai lure
of Te tracyc l ine to Bind to E scherichia coli Ribosomes " ,
Biochirn . Biophys . Acta , 1 0 3 , 5 3 2 ( 1 9 6 5 ) .
2 3 4 . Day , L . E . , � Te tracycl ine I nhibition of Ce l l-Free
P rotein Synthe s i s , I . Binding of Tetracycl ine to
Components of S y s tem" , J . Bacteria l . , � ' 1 9 1 7 ( 1 9 6 6 ) .
2 3 5 . Day , L . E . , Further Evidence o f Te tracycl ine Binding
to E . c o l i R ibosome s " , ibid , 2_?_, 1 2 6 3 ( 1 9 6 6 ) .
2 3 6 . Maxwel l , I . H . , � s tudi e s o f the Binding o f Tetracycline
to Ribosomes in vitro " , Hal . Pharmacal . , _! , 2 5 ( 1 9 6 8 ) .
2 3 7 . Bene t , L . A . and Goyan , J . E . , " Dete rminat ion of the
S tabi l i ty Constants of Tetracycl ine Complexe s '' , �
Pharrn . S ci . , � ' 9 8 3 ( 1 9 6 5 ) .
2 3 8 . Dolui s io , J . T . and Martin , A . N . , " Metal Comp lexation
o f the Tetracy c l ine Hydrochloride s " , J . Med . Chern . , §_ ,
1 6 ( 1 9 6 3 ) .

2 3 9 .
2 4 0 .
2 4 1 .
2 4 2 .
2 4 3 .
Cahn , R . D . , '' Detergents i n Membrane Fi lter s " ,
S c i ence , 1 5 5 , 1 9 5 ( 1 9 6 7 ) .
S chne i de r , W . C . , " Determinati on o f Nuc l e ic Acids in
T i ss ue s by Pentose Ana l ys i s " , i n " Methods of
1 7 1
Enzymology" , vol . 3 , Colow ick , S . P . and Kaplan , N . o . ,
Ed . , Academic Pre s s I nc . , New York , N . Y . , ( 1 9 5 7 ) pp . 6 8 0 .
Maaloe , 0 . and K j e l dgaard , N . 0 . , " Control o f Macro-molecular
Synthes i s " , W. A . Ben j ami n , I nc . , New York , N . Y . , ( 1 9 6 6 )
pp . 9 0 .
Layne , E . , ' Spectrophotometric and Turbidimetric Methods
for t1easuring Proteins " , in " Me thods o f En zymology " ,
vol . 3 , Colowick , S . P . and Kap l an , N . 0 . , Ed . ,
Academi c Pre s s I nc . , New York , N . Y . , ( 1 9 5 7 ) pp . 4 4 7 .
Garre t t , E . R . , Mi l ler , G . H . and Brown , M . R . W . ,
" Kinetics and Hechani sms o f Act ion o f Antibiotics on
Microorgani sms . V. Chloramphenicol and Tetracycl ine
Affected E s cherichia col i Generation Rates " , J . Pharm .
Sc i . , � , 5 9 3 ( 1 9 6 6 ) .
2 4 4 . Arima , K . and I zaki , K . , " Accumulation o f Oxytetracyc l ine
Re levant to i ts Bacter i c idal Act ion in the Ce l l s o f
2 4 5 .
E scheri ch i a col i " , Nature , 2 0 0 , 1 9 2 ( 1 9 6 3 ) .
I zaki , K . and Arima , K . , " E f fect o f Var ious Cond it ions
on Accumulation o f Oxytetracycl ine in E scheri chia col i " ,
J . Bacterial . , � , 1 3 3 5 ( 1 9 6 5 ) .





























