Embed Size (px)
Citation preview

REGULAR PAPER
Influence of germination date on Dioon edule (Zamiaceae) seedlingtolerance to water stress
Laura Yanez-Espinosa • Joel Flores •
Paulina S. Rodrıguez Millan • Gabriel Rubio Mendez
Received: 22 August 2013 / Accepted: 18 December 2013
� The Botanical Society of Japan and Springer Japan 2014
Abstract Dioon edule seedling mortality is mostly
attributed to dehydration by prolonged drought, even when
they present xeromorphic characteristics like the adult
plants. The effect of germination date (GD) and soil water
deficit on seedling tolerance to water stress was assessed.
The seedlings germinated and grown from mature seeds
every month from December to April GD were selected to
evaluate the leaf area, photosynthetic pigment content,
crassulacean acid metabolism (CAM) activity, stomatal
conductance (gs) and leaflet anatomy at soil water potential
(Ws) of 0.0 MPa (day 1), -0.1 MPa (day 40), -1.0 MPa
(day 90), -1.5 MPa (day 130), and a control (0.0 MPa at
day 130) to recognize differences due to leaf development.
The seedlings shifted from C3 to CAM cycling when
exposed to water stress at Ws of -1.0 MPa, like adult
plants. The March–April GD seedlings with undeveloped
sclerified hypodermis and stomata, presented reduced leaf
area, lower Chlorophyll a/b ratio, higher CAM activity and
midday partial stomatal closure when reached Ws of
-1.0 MPa. These have higher probability of dehydration
during severe drought (February–April) than those of the
December–February GD with similar Ws. Plants used for
restoration purposes must have full leaf development to
increase the survival.
Keywords CAM cycling � Cycads � Drought � Leaflet
anatomy � Mexico � Seedling mortality
Introduction
The endangered cycad Dioon edule Lindl. (Zamiaceae) is a
dioecious gymnosperm with a relatively limited production
of seeds and high mortality rate at the seed stage, up to
90 % under natural conditions (Vovides 1990). Seed
mortality has been attributed mainly to pre-germination
predation by both insects (Coleoptera) (Mora et al. 2013)
and the field mouse Peromyscus mexicanus (Rodentia:
Muridae) (Vovides et al. 2003).
Nevertheless, the high germination capacity of viable
seeds of D. edule, up to 75–98 % (Vovides 1990), com-
pensates for the aforementioned disadvantage. Although D.
edule seeds germinate in both shadow and light under
appropriate temperature and moisture in nursery condi-
tions, shadow increased 55.5 % germinability along
three months (Lopez-Ovando and Trevino-Garza 2008).
Therefore, there must be some ecological advantages for
seedlings provided by seed germination under shadow in
the field, as Dioon species most likely require low light
conditions during the initial growth stages, which are
L. Yanez-Espinosa (&)
Instituto de Investigacion de Zonas Deserticas, Universidad
Autonoma de San Luis Potosı, Altair 200, Col. del Llano,
San Luis Potosı, S.L.P 78377, Mexico
e-mail: [email protected]
L. Yanez-Espinosa � P. S. Rodrıguez Millan � G. Rubio Mendez
Programas Multidisciplinarios de Posgrado en Ciencias
Ambientales, Universidad Autonoma de San Luis Potosı, Av. Dr.
Manuel Nava 8, Zona Universitaria Poniente, San Luis Potosı,
S.L.P 78290, Mexico
P. S. Rodrıguez Millan
e-mail: [email protected]
G. Rubio Mendez
e-mail: [email protected]
J. Flores
Division de Ciencias Ambientales, Instituto Potosino de
Investigacion Cientıfica y Tecnologica, A.C. Camino a la Presa
San Jose 2055, Col. Lomas 4a Seccion, San Luis Potosı,
S.L.P 78216, Mexico
e-mail: [email protected]
123
J Plant Res
DOI 10.1007/s10265-014-0624-4

provided by filtered sunlight in forest canopies or maternal
plants (Alvarez-Yepiz et al. 2011).
The seedling mortality rate observed in some popula-
tions reaches up to 40 % for D. edule (Pavon-Saure, pers.
comm.) attributed mainly to the effect of water deficit due
to high temperatures and prolonged dry season during the
first year (Vovides 1990; Vovides et al. 2002). However,
why does dehydration mortally affect seedlings during the
first year?
Considering that adult plants of D. edule have the typ-
ical cycad xeromorphic traits that could allow them to
survive prolonged drought, including leaves with a coria-
ceous texture, thick cuticle, heavy cutinization, sunken
stomata and the presence of mucilage (Medeghini Bonatti
and Baroni Fornasiero 1990), relatively low net CO2
assimilation and stomatal conductance to H2O, such as in
succulents and evergreen conifers (Marler and Willis
1997), and C3 metabolism when the available water supply
is sufficient, they can shift to a crassulacean acid metabo-
lism (CAM) photosynthetic pathway in response to
increased CO2 assimilation and water use efficiency under
water stress, triggered at a soil water potential below
-1.0 MPa (Silvera et al. 2010; Vovides et al. 2002). Has
also been observed that seedlings of D. edule begin to
absorb large volumes of water in a short period of time
when the soil water potential reaches -0.1 MPa (unpub-
lished data).
Here, we explored why not all D. edule seedlings sur-
vive the drought if they have the same xeromorphic traits
than adult plants that allow them to tolerate water stress,
addressing the following questions: (a) How the germina-
tion date (GD) and the water stress would affect the leaf
development? and (b) How soil water potential would
affect the seedling tolerance to water stress? To answer
these questions we examined leaf area, photosynthetic
pigments, CAM activity (DH?), stomatal conductance (gs)
and leaflet anatomical characters, at soil water potential
(Ws) of 0.0 MPa (day 1), -0.1 MPa (day 40), -1.0 MPa
(day 90), -1.5 MPa (day 130), and a control (0.0 MPa at
day 130) to recognize differences due to leaf development.
Materials and methods
Plant material
In Dioon spp. the embryos are completely developed
immediately after the seed is released from the cone and
can germinate (Calonje et al. 2011). The seeds were col-
lected from mature strobilus ready to release them in
October 2007, in a population located in the southern
region of the Sierra Madre Oriental in the state of San Luis
Potosi, Mexico, at 1,167 m above the sea level (a.s.l.)
along a 33� slope with N exposure, clay loam shallow
(40–45 cm) soil rich in Ca (103.8 meq 100 g-1). The mean
temperature varies from 28.4 �C in summer to 14.1 �C in
winter. The mean total precipitation is 684.5 mm, with a
prolonged dry season from November to May. The soil
water potential (Ws) during the driest period (February–
April) is -1.26 MPa (±0.318) and during the wettest
month (September) is -0.01 MPa (±0.01). The D. edule
population is distributed in the oak forest–submontane
scrub ecotone associated with Quercus polymorpha
Schltdl. and Cham., Q. laeta Liebm. (Fagaceae), Flouren-
sia laurifolia DC. (Asteraceae), Randia laetevirens Standl.
(Rubiaceae), Dodonaea viscosa Jacq. (Sapindaceae) and
Acacia angustissima (Mill.) Kuntze (Fabaceae).
To assess whether seeds were viable a float test in water
was applied (Calonje et al. 2011). Full seeds were placed in
plastic containers using a commercial germination mix
(peat, perlite and limestone, 3:1:1), with available soil
moisture maintained at -0.02 MPa (±0.0001), and
exposed to light under a 60 % black shade cloth at
144.1 lmol m-2 s-1 (±8.9) in a glasshouse. Any seed
with at least 2 mm of radicle growth was considered to
have germinated, and every ten days were recorded and
tagged until the end of the experiment.
Four weeks after the last seed germinated, the seedlings
tagged with monthly GD from December to April were
transplanted to black polyethylene bags containing 0.8 kg
of a mix of clay and coarse sand (3:1), housed under
glasshouse conditions and irrigated to field capacity.
Four weeks after the seedlings were transplanted, the
substrate was irrigated with 330 cm3 of water per plant,
and each bag was covered with plastic film to avoid
evaporation. Daily measurements of photosynthetic photon
flux density (PPDF), temperature, and relative humidity
were registered with a quantum light sensor and with a
temperature/relative humidity sensor data logger (Watch-
Dog 450, Spectrum Technologies, IL, USA). The mean
PPFD, air temperature and relative humidity ± SE mea-
sured at noon every day from June to September 2008 were
171.5 ± 2.2 lmol m-2 s-1, 33.5 ± 0.6 �C and 35.9 ±
1.7 %, respectively. The light intensity conditions simu-
lated those of a forest understory with a low PPFD
(100–200 lmol m-2 s-1).
The soil moisture was calculated gravimetrically using
the equation [(wet soil weight ? bag weight) - (soil dry
weight ? bag weight)/(soil dry weight)] 9 soil density.
The soil water potential (Ws) was calculated with a soil
water characteristic curve generated in the laboratory for
the substrate with a digital soil moisture tester (KS-D1,
Delmhorst Instrument Co., NJ, USA).
Seedlings tagged with monthly GD ranging from
December to April were assessed in the beginning of
the experiment (Ws = 0.0 MPa), 40 days later (Ws =
J Plant Res
123

-0.1 MPa), 90 days later (Ws = -1.0 MPa), 130 days
after (Ws = -1.5 MPa) and a control 130 days later
(Ws = 0.0 MPa) in order to compare hydrated versus
dehydrated seedlings at the end of the experiment and
exclude differences attributable to development of leaves.
The experimental design was completely randomized, with
three seedlings per GD and Ws.
Leaf area
Leaf area was measured by applying the WinFOLIA
(Regent Instruments Inc., Canada) image analysis software
program to scanned images of the leaves taken before
sampling.
Photosynthetic pigments
Chlorophyll a (Chl a), chlorophyll b (Chl b) and carote-
noids (Car) concentrations were calculated according to the
equation proposed by Lichtenthaler and Welburn (1983).
The chlorophyll a/chlorophyll b (Chl a/b) and chlorophyll
a ? b/carotenoids (Chl a ? b/Car) ratios were also cal-
culated. The average of three readings from leaf extracts
was analyzed with a visible range spectrophotometer
(Genesys 20 Model 4001/4, Thermo Spectronic, NY, USA)
at a wavelength of 470 nm for Chl a, 647 nm for Chl b and
663 nm for Car.
CAM activity
CAM activity was measured as DH? by calculating the
total titratable acids content according to the method pro-
posed by Vovides et al. (2002) for D. edule plants, which
involves collecting leaf samples at 6:00 a.m. (dark) and
7:00 p.m. (light) to calculate the titratable acidity of mac-
erated tissue with NaOH 0.01 N at pH 8. The mean of three
titrations per individual was thus determined.
Stomatal conductance (gs)
The gs measurements were performed with a leaf porometer
(SC-1, Decagon Devices, WA, USA) every 3 h over a 24 h
period. Leaflets were narrower than the diameter of the
cuvette; thus, two to three leaflets were inserted side by side
with no gap or overlap (Marler and Willis 1997). The stomatal
conductance of April GD plants with a Ws of 0.0 MPa was not
measured because the soft texture of the leaves would have
been damaged by the porometer sensor head.
Leaflet anatomy
One section of a leaflet was sampled in the middle leaf
region of December–April GD plants with Ws of 0.0 MPa,
fixed in 3 % glutaraldehyde solution and embedded in
glycol-methacrylate (Technovit� 7100, Heraeus Kulzer,
DE, USA). Transverse sections measuring 2 lm thick were
cut with a rotatory microtome (RM2125RT, Leica Micro-
systems, DE, USA), stained with 0.6 % brilliant cresyl blue
(Ruzin 1999) and observed under a light microscope (DM
2000, Leica Microsystems, DE, USA). Sections from the
same sample were slowly dried in a vacuum desiccator at
room temperature. The samples were mounted on double-
sided carbon tape on metal pins and sputtered with gold.
Stomata were observed under a scanning electron micro-
scope (Quanta 200, FEI, OR, USA).
Stomatal density was measured observing in five fields
of an epidermal impression of the leaflet adaxial surface
under a light microscope. The impression was made by
spreading a thin layer of nail varnish and removed after dry
by attaching clear sticky tape to it, peeling it from the
leaflet surface and sticking it to the slide.
Statistical analyses
Data analyses were performed using a two-way analysis of
variance (ANOVA) to examine the effects of GD, Ws and
their interaction on leaf area, DH? and the photosynthetic
pigment content (Chl a, Chl b, Car) and ratios (Chl a/b, Chl
a ? b/Car) for different Ws (0.0, -1.5 MPa). Leaflet sto-
matal density data analysis was performed with one-way
ANOVA to study the influence of GD. Differences among
the means were evaluated using Tukey’s test (P \ 0.05).
Linear regressions were performed with Ws as a predictor
of dark total H?, and stomatal density as a predictor of
stomatal conductance (gs) at 13:00, when they presented
the highest value throughout the day, and all variables were
previously transformed to their natural logarithm forms
except Ws.
The gs data analysis was performed with a two-way
repeated measures ANOVA, and because the assumption
of sphericity was violated, the degrees of freedom were
corrected with the Huynd–Feldt correction. Differences
among means were evaluated using the Bonferroni test
(P \ 0.05). All statistical analyses were performed with
XLSTAT software (2013.01.1 v, Addinsoft, NY, USA).
Results
Leaf area
No significance was indicated when analyzing the inter-
action between both of the factors for leaf area (n = 75,
d.f. = 16, F = 0.36, P [ 0.05). Although there was a
tendency for the December and January GD seedlings to
increase in leaf area, even though the soil water potential
J Plant Res
123
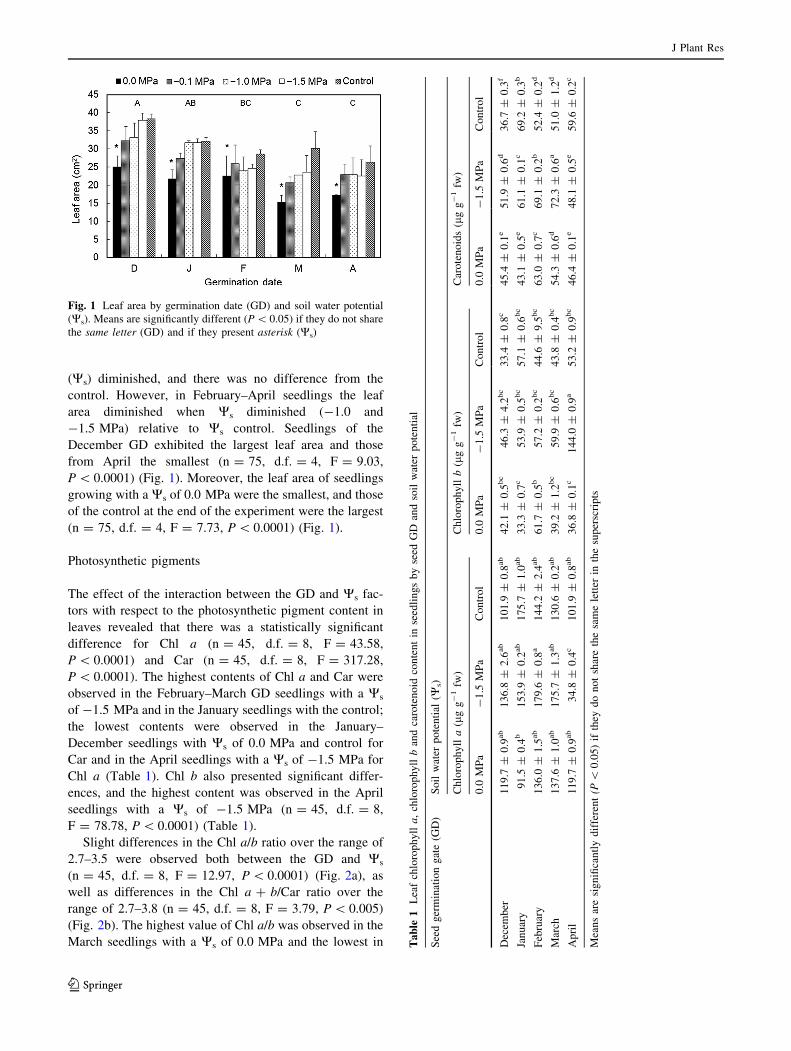
(Ws) diminished, and there was no difference from the
control. However, in February–April seedlings the leaf
area diminished when Ws diminished (-1.0 and
-1.5 MPa) relative to Ws control. Seedlings of the
December GD exhibited the largest leaf area and those
from April the smallest (n = 75, d.f. = 4, F = 9.03,
P \ 0.0001) (Fig. 1). Moreover, the leaf area of seedlings
growing with a Ws of 0.0 MPa were the smallest, and those
of the control at the end of the experiment were the largest
(n = 75, d.f. = 4, F = 7.73, P \ 0.0001) (Fig. 1).
Photosynthetic pigments
The effect of the interaction between the GD and Ws fac-
tors with respect to the photosynthetic pigment content in
leaves revealed that there was a statistically significant
difference for Chl a (n = 45, d.f. = 8, F = 43.58,
P \ 0.0001) and Car (n = 45, d.f. = 8, F = 317.28,
P \ 0.0001). The highest contents of Chl a and Car were
observed in the February–March GD seedlings with a Ws
of -1.5 MPa and in the January seedlings with the control;
the lowest contents were observed in the January–
December seedlings with Ws of 0.0 MPa and control for
Car and in the April seedlings with a Ws of -1.5 MPa for
Chl a (Table 1). Chl b also presented significant differ-
ences, and the highest content was observed in the April
seedlings with a Ws of -1.5 MPa (n = 45, d.f. = 8,
F = 78.78, P \ 0.0001) (Table 1).
Slight differences in the Chl a/b ratio over the range of
2.7–3.5 were observed both between the GD and Ws
(n = 45, d.f. = 8, F = 12.97, P \ 0.0001) (Fig. 2a), as
well as differences in the Chl a ? b/Car ratio over the
range of 2.7–3.8 (n = 45, d.f. = 8, F = 3.79, P \ 0.005)
(Fig. 2b). The highest value of Chl a/b was observed in the
March seedlings with a Ws of 0.0 MPa and the lowest in
Fig. 1 Leaf area by germination date (GD) and soil water potential
(Ws). Means are significantly different (P \ 0.05) if they do not share
the same letter (GD) and if they present asterisk (Ws)
Ta
ble
1L
eaf
chlo
rop
hy
lla
,ch
loro
ph
yll
ban
dca
rote
no
idco
nte
nt
inse
edli
ng
sb
yse
edG
Dan
dso
ilw
ater
po
ten
tial
See
dg
erm
inat
ion
gat
e(G
D)
So
ilw
ater
po
ten
tial
(Ws)
Ch
loro
ph
yll
a(l
gg
-1
fw)
Ch
loro
ph
yll
b(l
gg
-1
fw)
Car
ote
no
ids
(lg
g-
1fw
)
0.0
MP
a-
1.5
MP
aC
on
tro
l0
.0M
Pa
-1
.5M
Pa
Co
ntr
ol
0.0
MP
a-
1.5
MP
aC
on
tro
l
Dec
emb
er1
19
.7±
0.9
ab
13
6.8
±2
.6ab
10
1.9
±0
.8ab
42
.1±
0.5
bc
46
.3±
4.2
bc
33
.4±
0.8
c4
5.4
±0
.1e
51
.9±
0.6
d3
6.7
±0
.3f
Jan
uar
y9
1.5
±0
.4b
15
3.9
±0
.2ab
17
5.7
±1
.0ab
33
.3±
0.7
c5
3.9
±0
.5bc
57
.1±
0.6
bc
43
.1±
0.5
e6
1.1
±0
.1c
69
.2±
0.3
b
Feb
ruar
y1
36
.0±
1.5
ab
17
9.6
±0
.8a
14
4.2
±2
.4ab
61
.7±
0.5
b5
7.2
±0
.2bc
44
.6±
9.5
bc
63
.0±
0.7
c6
9.1
±0
.2b
52
.4±
0.2
d
Mar
ch1
37
.6±
1.0
ab
17
5.7
±1
.3ab
13
0.6
±0
.2ab
39
.2±
1.2
bc
59
.9±
0.6
bc
43
.8±
0.4
bc
54
.3±
0.6
d7
2.3
±0
.6a
51
.0±
1.2
d
Ap
ril
11
9.7
±0
.9ab
34
.8±
0.4
c1
01
.9±
0.8
ab
36
.8±
0.1
c1
44
.0±
0.9
a5
3.2
±0
.9bc
46
.4±
0.1
e4
8.1
±0
.5e
59
.6±
0.2
c
Mea
ns
are
sig
nifi
can
tly
dif
fere
nt
(P\
0.0
5)
ifth
eyd
on
ot
shar
eth
esa
me
lett
erin
the
sup
ersc
rip
ts
J Plant Res
123
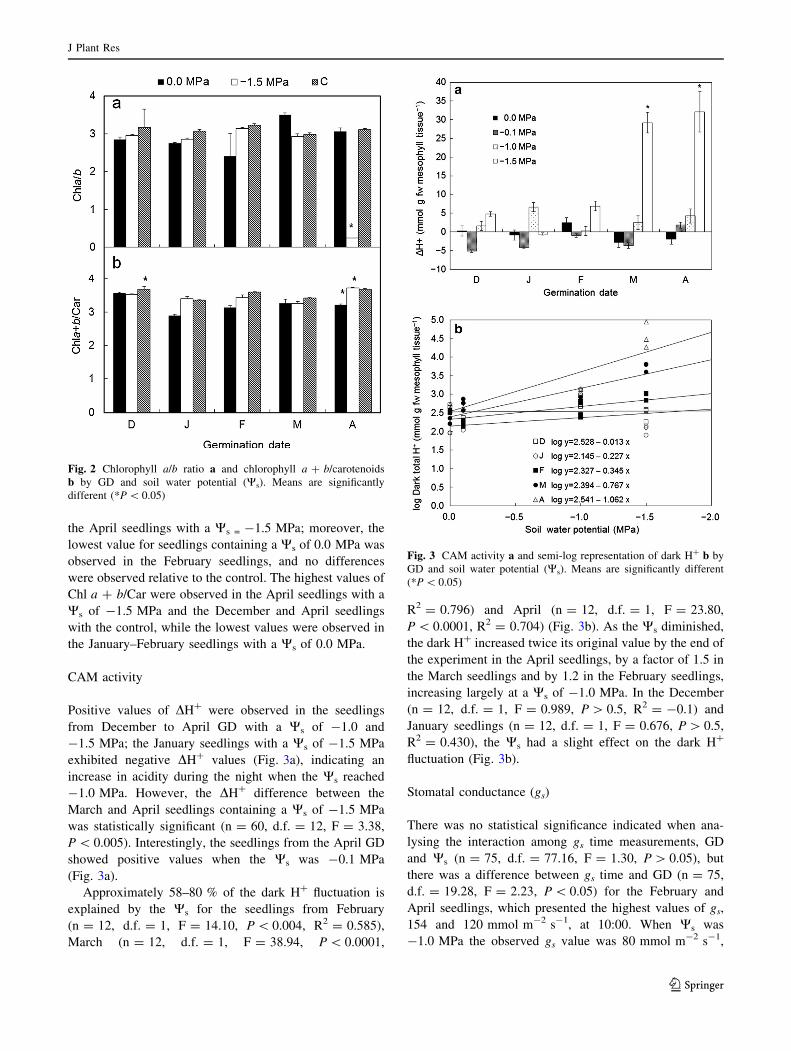
the April seedlings with a Ws = -1.5 MPa; moreover, the
lowest value for seedlings containing a Ws of 0.0 MPa was
observed in the February seedlings, and no differences
were observed relative to the control. The highest values of
Chl a ? b/Car were observed in the April seedlings with a
Ws of -1.5 MPa and the December and April seedlings
with the control, while the lowest values were observed in
the January–February seedlings with a Ws of 0.0 MPa.
CAM activity
Positive values of DH? were observed in the seedlings
from December to April GD with a Ws of -1.0 and
-1.5 MPa; the January seedlings with a Ws of -1.5 MPa
exhibited negative DH? values (Fig. 3a), indicating an
increase in acidity during the night when the Ws reached
-1.0 MPa. However, the DH? difference between the
March and April seedlings containing a Ws of -1.5 MPa
was statistically significant (n = 60, d.f. = 12, F = 3.38,
P \ 0.005). Interestingly, the seedlings from the April GD
showed positive values when the Ws was -0.1 MPa
(Fig. 3a).
Approximately 58–80 % of the dark H? fluctuation is
explained by the Ws for the seedlings from February
(n = 12, d.f. = 1, F = 14.10, P \ 0.004, R2 = 0.585),
March (n = 12, d.f. = 1, F = 38.94, P \ 0.0001,
R2 = 0.796) and April (n = 12, d.f. = 1, F = 23.80,
P \ 0.0001, R2 = 0.704) (Fig. 3b). As the Ws diminished,
the dark H? increased twice its original value by the end of
the experiment in the April seedlings, by a factor of 1.5 in
the March seedlings and by 1.2 in the February seedlings,
increasing largely at a Ws of -1.0 MPa. In the December
(n = 12, d.f. = 1, F = 0.989, P [ 0.5, R2 = -0.1) and
January seedlings (n = 12, d.f. = 1, F = 0.676, P [ 0.5,
R2 = 0.430), the Ws had a slight effect on the dark H?
fluctuation (Fig. 3b).
Stomatal conductance (gs)
There was no statistical significance indicated when ana-
lysing the interaction among gs time measurements, GD
and Ws (n = 75, d.f. = 77.16, F = 1.30, P [ 0.05), but
there was a difference between gs time and GD (n = 75,
d.f. = 19.28, F = 2.23, P \ 0.05) for the February and
April seedlings, which presented the highest values of gs,
154 and 120 mmol m-2 s-1, at 10:00. When Ws was
-1.0 MPa the observed gs value was 80 mmol m-2 s-1,
Fig. 2 Chlorophyll a/b ratio a and chlorophyll a ? b/carotenoids
b by GD and soil water potential (Ws). Means are significantly
different (*P \ 0.05)
Fig. 3 CAM activity a and semi-log representation of dark H? b by
GD and soil water potential (Ws). Means are significantly different
(*P \ 0.05)
J Plant Res
123
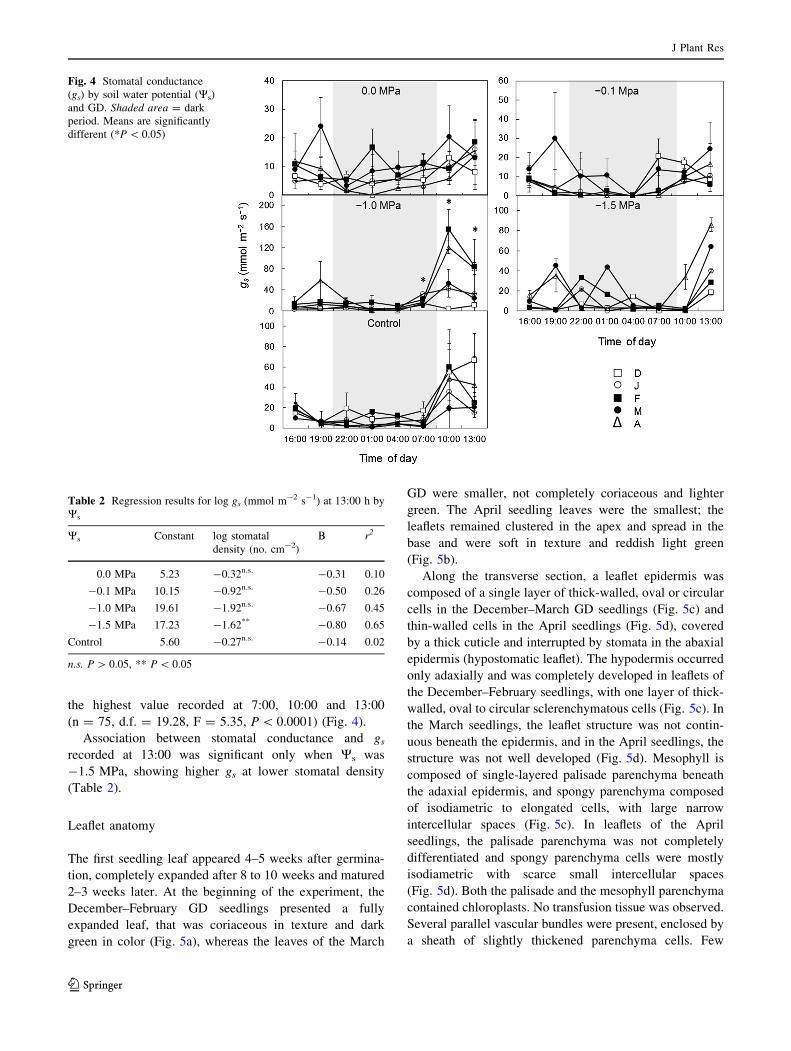
the highest value recorded at 7:00, 10:00 and 13:00
(n = 75, d.f. = 19.28, F = 5.35, P \ 0.0001) (Fig. 4).
Association between stomatal conductance and gs
recorded at 13:00 was significant only when Ws was
-1.5 MPa, showing higher gs at lower stomatal density
(Table 2).
Leaflet anatomy
The first seedling leaf appeared 4–5 weeks after germina-
tion, completely expanded after 8 to 10 weeks and matured
2–3 weeks later. At the beginning of the experiment, the
December–February GD seedlings presented a fully
expanded leaf, that was coriaceous in texture and dark
green in color (Fig. 5a), whereas the leaves of the March
GD were smaller, not completely coriaceous and lighter
green. The April seedling leaves were the smallest; the
leaflets remained clustered in the apex and spread in the
base and were soft in texture and reddish light green
(Fig. 5b).
Along the transverse section, a leaflet epidermis was
composed of a single layer of thick-walled, oval or circular
cells in the December–March GD seedlings (Fig. 5c) and
thin-walled cells in the April seedlings (Fig. 5d), covered
by a thick cuticle and interrupted by stomata in the abaxial
epidermis (hypostomatic leaflet). The hypodermis occurred
only adaxially and was completely developed in leaflets of
the December–February seedlings, with one layer of thick-
walled, oval to circular sclerenchymatous cells (Fig. 5c). In
the March seedlings, the leaflet structure was not contin-
uous beneath the epidermis, and in the April seedlings, the
structure was not well developed (Fig. 5d). Mesophyll is
composed of single-layered palisade parenchyma beneath
the adaxial epidermis, and spongy parenchyma composed
of isodiametric to elongated cells, with large narrow
intercellular spaces (Fig. 5c). In leaflets of the April
seedlings, the palisade parenchyma was not completely
differentiated and spongy parenchyma cells were mostly
isodiametric with scarce small intercellular spaces
(Fig. 5d). Both the palisade and the mesophyll parenchyma
contained chloroplasts. No transfusion tissue was observed.
Several parallel vascular bundles were present, enclosed by
a sheath of slightly thickened parenchyma cells. Few
Table 2 Regression results for log gs (mmol m-2 s-1) at 13:00 h by
Ws
Ws Constant log stomatal
density (no. cm-2)
B r2
0.0 MPa 5.23 -0.32n.s. -0.31 0.10
-0.1 MPa 10.15 -0.92n.s. -0.50 0.26
-1.0 MPa 19.61 -1.92n.s. -0.67 0.45
-1.5 MPa 17.23 -1.62** -0.80 0.65
Control 5.60 -0.27n.s. -0.14 0.02
n.s. P [ 0.05, ** P \ 0.05
Fig. 4 Stomatal conductance
(gs) by soil water potential (Ws)
and GD. Shaded area = dark
period. Means are significantly
different (*P \ 0.05)
J Plant Res
123
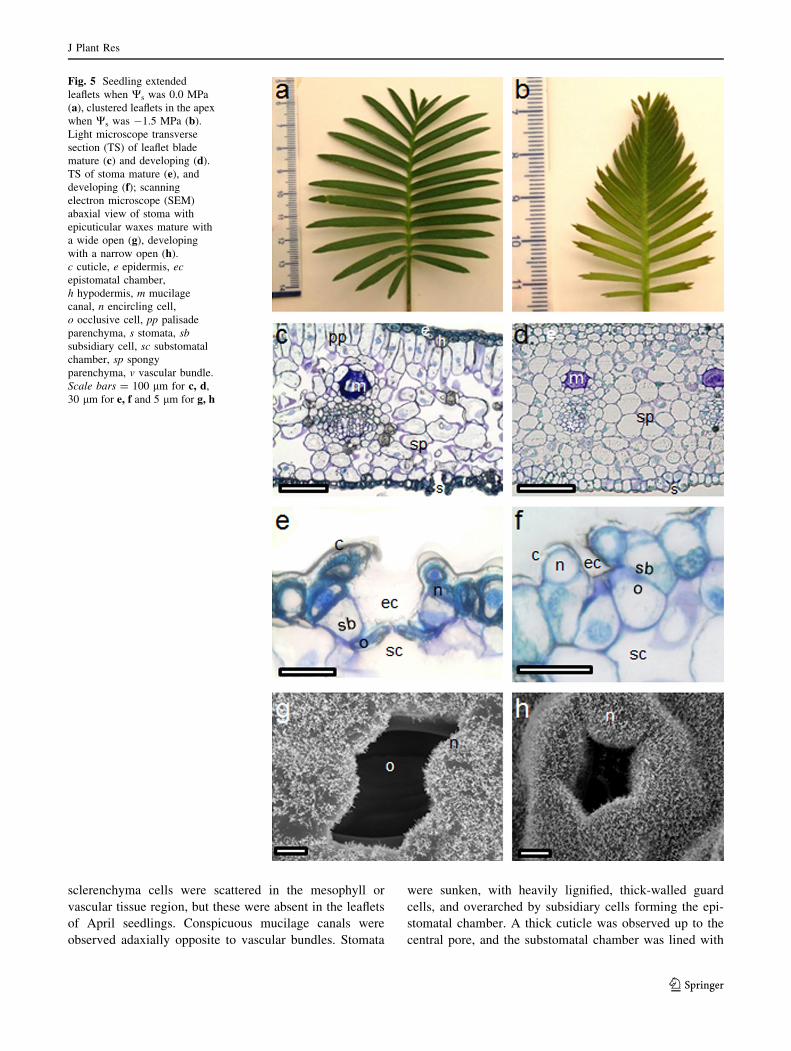
sclerenchyma cells were scattered in the mesophyll or
vascular tissue region, but these were absent in the leaflets
of April seedlings. Conspicuous mucilage canals were
observed adaxially opposite to vascular bundles. Stomata
were sunken, with heavily lignified, thick-walled guard
cells, and overarched by subsidiary cells forming the epi-
stomatal chamber. A thick cuticle was observed up to the
central pore, and the substomatal chamber was lined with
Fig. 5 Seedling extended
leaflets when Ws was 0.0 MPa
(a), clustered leaflets in the apex
when Ws was -1.5 MPa (b).
Light microscope transverse
section (TS) of leaflet blade
mature (c) and developing (d).
TS of stoma mature (e), and
developing (f); scanning
electron microscope (SEM)
abaxial view of stoma with
epicuticular waxes mature with
a wide open (g), developing
with a narrow open (h).
c cuticle, e epidermis, ec
epistomatal chamber,
h hypodermis, m mucilage
canal, n encircling cell,
o occlusive cell, pp palisade
parenchyma, s stomata, sb
subsidiary cell, sc substomatal
chamber, sp spongy
parenchyma, v vascular bundle.
Scale bars = 100 lm for c, d,
30 lm for e, f and 5 lm for g, h
J Plant Res
123
spongy mesophyll cells (Fig. 5e). However, in March–
April seedlings, the stomata were still developing (Fig. 5d),
with thin-walled guard and subsidiary cells and a wide
lumen and a thin cuticle observed near the central pore
(Fig. 5f); the dorsal walls of guard cells were observed to
have lignified before the ventral walls. Subsidiary cells
were partially overlapped by two concentric rings of
encircling cells, which were raised to form a rim at the
same level as other epidermal cells, covered with a thick
cuticle and epicuticular wax tubules (Fig. 5g). The stoma
of the March–April seedlings showed only one ring of
raised encircling cells, but they were thin-walled and not
completely lignified, forming a narrow opening edge
(Fig. 5f, h).
Significant differences were observed among values of
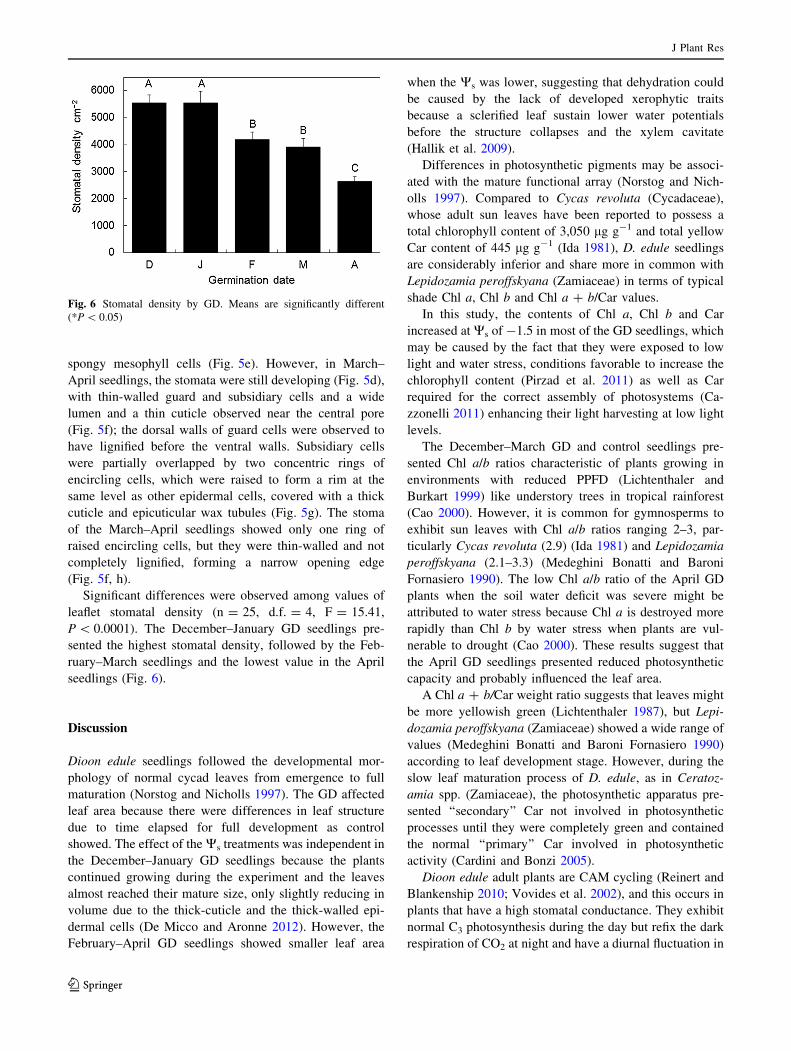
leaflet stomatal density (n = 25, d.f. = 4, F = 15.41,
P \ 0.0001). The December–January GD seedlings pre-
sented the highest stomatal density, followed by the Feb-
ruary–March seedlings and the lowest value in the April
seedlings (Fig. 6).
Discussion
Dioon edule seedlings followed the developmental mor-
phology of normal cycad leaves from emergence to full
maturation (Norstog and Nicholls 1997). The GD affected
leaf area because there were differences in leaf structure
due to time elapsed for full development as control
showed. The effect of the Ws treatments was independent in
the December–January GD seedlings because the plants
continued growing during the experiment and the leaves
almost reached their mature size, only slightly reducing in
volume due to the thick-cuticle and the thick-walled epi-
dermal cells (De Micco and Aronne 2012). However, the
February–April GD seedlings showed smaller leaf area
when the Ws was lower, suggesting that dehydration could
be caused by the lack of developed xerophytic traits
because a sclerified leaf sustain lower water potentials
before the structure collapses and the xylem cavitate
(Hallik et al. 2009).
Differences in photosynthetic pigments may be associ-
ated with the mature functional array (Norstog and Nich-
olls 1997). Compared to Cycas revoluta (Cycadaceae),
whose adult sun leaves have been reported to possess a
total chlorophyll content of 3,050 lg g-1 and total yellow
Car content of 445 lg g-1 (Ida 1981), D. edule seedlings
are considerably inferior and share more in common with
Lepidozamia peroffskyana (Zamiaceae) in terms of typical
shade Chl a, Chl b and Chl a ? b/Car values.
In this study, the contents of Chl a, Chl b and Car
increased at Ws of -1.5 in most of the GD seedlings, which
may be caused by the fact that they were exposed to low
light and water stress, conditions favorable to increase the
chlorophyll content (Pirzad et al. 2011) as well as Car
required for the correct assembly of photosystems (Ca-
zzonelli 2011) enhancing their light harvesting at low light
levels.
The December–March GD and control seedlings pre-
sented Chl a/b ratios characteristic of plants growing in
environments with reduced PPFD (Lichtenthaler and
Burkart 1999) like understory trees in tropical rainforest
(Cao 2000). However, it is common for gymnosperms to
exhibit sun leaves with Chl a/b ratios ranging 2–3, par-
ticularly Cycas revoluta (2.9) (Ida 1981) and Lepidozamia
peroffskyana (2.1–3.3) (Medeghini Bonatti and Baroni
Fornasiero 1990). The low Chl a/b ratio of the April GD
plants when the soil water deficit was severe might be
attributed to water stress because Chl a is destroyed more
rapidly than Chl b by water stress when plants are vul-
nerable to drought (Cao 2000). These results suggest that
the April GD seedlings presented reduced photosynthetic
capacity and probably influenced the leaf area.
A Chl a ? b/Car weight ratio suggests that leaves might
be more yellowish green (Lichtenthaler 1987), but Lepi-
dozamia peroffskyana (Zamiaceae) showed a wide range of
values (Medeghini Bonatti and Baroni Fornasiero 1990)
according to leaf development stage. However, during the
slow leaf maturation process of D. edule, as in Ceratoz-
amia spp. (Zamiaceae), the photosynthetic apparatus pre-
sented ‘‘secondary’’ Car not involved in photosynthetic
processes until they were completely green and contained
the normal ‘‘primary’’ Car involved in photosynthetic
activity (Cardini and Bonzi 2005).
Dioon edule adult plants are CAM cycling (Reinert and
Blankenship 2010; Vovides et al. 2002), and this occurs in
plants that have a high stomatal conductance. They exhibit
normal C3 photosynthesis during the day but refix the dark
respiration of CO2 at night and have a diurnal fluctuation in
Fig. 6 Stomatal density by GD. Means are significantly different
(*P \ 0.05)
J Plant Res
123

the concentration of malic acid (Lambers et al. 2008). The
December–March seedlings exhibited C3 photosynthesis,
but shifted to CAM cycling when substrate reached a Ws of
-1.0 MPa as observed in adult plants, but with higher
concentrations than those reported in previous studies
(Vovides et al. 2002). However, the April seedlings
exhibited CAM cycling at higher Ws because underdevel-
oped leaf xeromorphic traits may have caused higher stress.
Although it has been suggested that at lower temperatures,
lower water stress and lower light intensity, D. edule adult
plants do not present CAM cycling (Vovides et al. 2002),
this experiment showed that seedlings under water stress
and low light intensity presented CAM cycling.
Seedlings with leaflets exhibiting fully developed
structural xeromorphic traits (December–January GD)
showed only slight increases in dark H?, whereas leaflets
with underdeveloped xeromorphic traits (February–April
GD) showed exponential increases in dark H? when Ws
decreased. Interestingly, some CAM plants have shown
increases in different organic acid content with increases in
osmotic pressure and, as a consequence, store a large
amount of water, making them less resistant to frost and
heat (Losch 1984).
The stomatal conductance of all the GD seedlings at
higher Ws showed a single mid-day peak characteristic of
C3 plants that are well supplied with water. Meanwhile, the
March–April GD seedlings at the lowest Ws showed
morning and late-afternoon peaks and mid-day partial
stomatal closure during the time of the day when the
evaporative demand was highest, characteristic of plants
growing in a water-limited environment (Lambers et al.
2008). When the Ws was -1.0 MPa, the gs increased near
the end of the night in all seedlings, as observed in other
CAM cycling species such as Talinum (Portulacaceae) and
Peperomia camptotricha (Piperaceae), which is explained
by the accelerated rate of malic acid formation toward the
end of the night (Harris and Martin 1991).
Some studies have reported that adult plants showing
mature dark foliage exhibit higher gas exchange values than
when fully expanded but more immature light-green foliage
(Marler and Willis 1997), possibly because stomata and
mesophyll are not full developed. However, in this experiment
there were no significant differences between the mature and
the immature leaves of the seedlings when the Ws was higher,
and low values were maintained, i.e., 32 % gs on average
reported for cultivated adult plants of D. edule growing in
Florida (155 mmol m-2 s-1) (Marler and Willis 1997).
The stomatal anatomy coincides with descriptions of
Cycas circinalis (Rao 1972) and Dioon spinulosum (Tang
et al. 2004). The sunken stomata of D. edule leaflets pro-
vide an additional advantage against drought by decreasing
gs, although the rate of CO2 influx is also reduced, and by
the creation of special microclimatic conditions such as
higher humidity directly above the stomatal pore (Roth-
Nebelsick 2007). The epistomatal chamber, with a depth of
20 lm, decreased the conductance by 30 % (Roth-Nebel-
sick 2007), and D. edule seedling stomata presented
chambers with an average depth of 20.34 lm; however,
seedlings from the March–April GDs showed less devel-
oped chambers, resulting in higher values of gs when water
stressed.
Stomatal density of seedlings was lower than reported
for adult D. edule but similar to other cycad species (Vo-
vides et al. 2002). The stomatal density presented a nega-
tive association with gs only when severe drought was
present (Ws of -1.5 MPa), although there is not a clear
relationship between stomatal density and gs (Camargo and
Marenco 2011).
Our experiment revealed the effect of GD and soil water
deficit (Ws) on seedling tolerance to water stress, helping to
understand the most relevant seedling establishment limi-
tation identified for the species D. edule. Remarkably D.
edule seedlings from all GDs are able to shift from C3 to
CAM photosynthetic pathway triggered at Ws of
-1.0 MPa, like adult plants. Particularly, the March–April
GD seedlings (61 %) with less developed xeromorphic
traits during the drought season (Ws of -1.0 to -1.5 MPa)
would have higher probability of death by dehydration.
Even less developed leaves have higher positive values of
DH?, they show midday partial stomatal closure, reduced
sclerification that may not sustain lower water potentials
and decreased photosynthetic capacity.
The seedling mortality has a relevant impact on population
structure and our results suggest that during prolonged and
severe drought, a low proportion of seedlings that have
developed completely the xeromorphic traits in the leaf
increase their probability of survival to secure the cohort. For
restoration purposes it is recommended to use plants with
xeromorphic traits of the leaf completely developed and
planting after the drought season to increase survival.
Acknowledgments This work was supported by The Cycad Soci-
ety, Inc. and PROMEP—SEP [UASLP-PTC-179]. Mexican govern-
ment authorized cone collection [SGPADGVS/00507/08]. We thank
NTBG College Biology Professors’ Fellowship for the basic knowl-
edge in cycads; Pablo Delgado provided assistance with SEM; Isaac
Jacob Chavez Acuna provided environmental data; Raymundo Mora
and Efraın Hernandez provided assistance in the field and laboratory.
References
Alvarez-Yepiz JC, Dovciak M, Burquez A (2011) Persistence of a
rare ancient cycad: effects of environment and demography. Biol
Conserv 144:122–130
Calonje M, Kay J, Griffith MP (2011) Propagation of cycad
collections from seed. Applied reproductive biology for conser-
vation. Sibbaldia 9:77–96
J Plant Res
123

Camargo MAB, Marenco RA (2011) Density, size and distribution of
stomata in 35 rainforest tree species in Central Amazonia. Acta
Amaz 41:205–212
Cao KF (2000) Leaf anatomy and chlorophyll content of 12 woody
species in contrasting light conditions in a Bornean heath forest.
Can J Bot 78:1245–1253
Cardini F, Bonzi LM (2005) Carotenoid composition and its
chemotaxonomic significance in leaves of ten species of the
genus Ceratozamia (Cycads). J Plant Physiol 162:517–528
Cazzonelli CI (2011) Carotenoids in nature: insights from plants and
beyond. Funct Plant Biol 38:833–847
De Micco V, Aronne G (2012) Morpho-anatomical traits for plant
adaptation to drought. In: Aroca R (ed) Plant responses to
drought stress, from morphological to molecular features.
Springer, Berlin, pp 37–61
Hallik L, Niinemets U, Wright IJ (2009) Are species shade and
drought tolerance reflected in leaf-level structural and functional
differentiation in Northern Hemisphere temperate woody flora?
New Phytol 184:257–274
Harris FS, Martin CE (1991) Correlation between CAM-cycling and
photosynthetic gas exchange in five species of Talinum (Port-
ulacaceae). Plant Physiol 96:1118–1124
Ida K (1981) Eco-physiological studies on the response of Taxodia-
ceous conifers to shading, with special reference to the behaviour
of leaf pigments distribution of carotenoids in green and
autumnal reddish brown leaves of gymnosperms. Bot Mag
(Tokyo) 94:41–54
Lambers H, Chapin FS, Pons TL (2008) Plant physiological ecology,
2nd edn. Springer, New York
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of
photosynthetic membranes. Methods Enzymol 148:350–382
Lichtenthaler HK, Burkart S (1999) Photosynthesis and high light
stress. Bulg J Plant Physiol 25:3–16
Lichtenthaler HK, Wellburn AR (1983) Determinations of total
carotenoids and chlorophylls a and b of leaf extracts in different
solvents. Biochem Soc T 11:591–592
Lopez-Ovando A, Trevino-Garza EJ (2008) Reproduction of chamal
(Dioon edule Lindley) by seed. Ra Ximhai 4:45–55
Losch R (1984) Species-specific responses to temperature in acid
metabolism and gas exchange performance of Macronesian
Sempervivoideae. In: Margaris NS, Arianoustou-Farragitaki M,
Oechel WC (eds) Being alive on land. Tasks for vegetation
science, vol 13. Dr. W. Junk, The Hague, pp 117–126
Marler TE, Willis LE (1997) Leaf gas-exchange characteristics of
sixteen cycad species. J Am Soc Hortic Sci 122:38–42
Medeghini Bonatti P, Baroni Fornasiero R (1990) Developmental
pattern and structural organization of leaf chloroplasts in
Lepidozamia peroffskyana. Aust J Bot 38:53–62
Mora R, Yanez-Espinosa L, Flores J, Nava-Zarate N (2013) Strobilus
and seed production of Dioon edule (Zamiaceae) in a population
with low seedlings density in San Luis Potosı, Mexico. Trop
Conserv Sci 6:268–282
Norstog KJ, Nicholls TJ (1997) The biology of the cycads. Cornell
University Press, Ithaca
Pirzad A, Reza Shakiba M, Zehtab-Salmasi S, Abolghasem Moham-
madi S, Darvishzadeh R, Samadi A (2011) Effect of water stress
on leaf relative water content, chlorophyll, proline and soluble
carbohydrates in Matricaria chamomilla L. J Med Plant Res
5:2483–2488
Rao LN (1972) Life history of Cycas circinalis. V. Seedling anatomy.
Proc Indian Acad Sci Sect B 76:47–57
Reinert F, Blankenship R (2010) Evolutionary aspects of crassulacean
acid metabolism. Oecol Aust 14:359–368
Roth-Nebelsick A (2007) Computer-based studies of diffusion
through stomata of different architecture. Ann Bot 100:23–32
Ruzin SE (1999) Plant microtechnique and microscopy. Oxford
University Press, Oxford
Silvera K, Neubig KM, Whitten WM, Williams NH, Winter K,
Cushman JC (2010) Evolution along the crassulacean acid
metabolism continuum. Funct Plant Biol 37:995–1010
Tang YJ, Liu N, Liao JP, Xie ZY, Wu QG, Chen JR (2004)
Systematics implications of pinna venation and pinna anatomy in
Zamiaceae. Acta Phytotax Sin 42:365–374
Vovides AP (1990) Spatial distribution, survival and fecundity of
Dioon edule (Zamiaceae) in a tropical deciduous forest in
Veracruz, Mexico, with notes on its habitat. Am J Bot
77:1532–1543
Vovides AP, Etherington JR, Dresser PQ, Groenhof A, Iglesias C,
Flores Ramırez J (2002) CAM-cycling in the cycad Dioon edule
Lindl. in its natural tropical deciduous forest habitat in central
Veracruz, Mexico. Bot J Linn Soc 138:155–162
Vovides AP, Perez-Farrera MA, Gonzalez-Astorga J, Gonzalez D,
Gregory T, Chemnic J, Iglesias C, Octavio-Aguilar P, Avendano
S, Barcenas C (2003) An outline of current knowledge of
Mexican cycads (Zamiaceae, Cycadales). Curr Top Plant Biol
4:159–174
J Plant Res
123











![Two Parasites of the Common Cockle {]ardium edule ... · commensal, but Patterson (1912, pp. 174-5) concluded that it lived chiefly in the kidney. Ball (1916, pp. 462-3), however,](https://img.pdfslide.us/doc/110x75/6061f16e24d3bb698c7660c1/two-parasites-of-the-common-cockle-ardium-edule-commensal-but-patterson-1912.jpg)

















