Embed Size (px)
Citation preview

[CANCER RESEARCH 61, 5248–5254, July 1, 2001]
Influence of b1 Integrins on Epidermal Squamous Cell Carcinoma Formation in aTransgenic Mouse Model:a3b1, but not a2b1, Suppresses Malignant Conversion1
David M. Owens and Fiona M. Watt2
Imperial Cancer Research Fund, Keratinocyte Laboratory, London WC2A 3PX, United Kingdom
ABSTRACT
Although aberrant integrin expression has been documented in manyepithelial tumors, little is known about how integrins influence neoplasticprogression. To examine this issue, transgenic mice in which thea2b1 ora3b1 integrin was expressed in the suprabasal epidermal layers via theinvolucrin promoter were subjected to skin carcinogenesis. Equal num-bers of benign squamous papillomas were observed in transgenic andwild-type animals. However, the frequency of conversion of papillomas tomalignant squamous cell carcinomas was much lower ina3b1 transgenicthan in a2b1 transgenic and wild-type mice. No differences were observedin apoptosis or in the expression of endogenous integrins in transgenic andwild-type papillomas. However,a3b1 transgenic papillomas displayed adiminished proliferative capacity and were more highly differentiated asjudged by BrdUrd incorporation and keratin 10 expression, respectively,than a2b1 transgenic and wild-type papillomas. Two proteins that asso-ciate with a3b1 and not a2b1 are extracellular matrix metalloproteinaseinducer and CD81. Extracellular matrix metalloproteinase inducer ex-pression correlated inversely with the degree of differentiation in normalepidermis and in transgenic and wild-type papillomas. Up-regulation ofCD81 was observed in 100% of wild-type and 88% ofa2b1 transgenicpapillomas but in only 25% of a3b1 transgenic papillomas. CD81 wasundetectable in untreated epidermis and strongly expressed in all trans-genic and wild-type squamous cell carcinomas. Our results demonstratethat the a3b1 integrin can suppress malignant conversion, and that themechanism may involve CD81.
INTRODUCTION
Integrins are cell surface receptors that mediate cell-extracellularmatrix and cell-cell adhesion events and are capable of transducingsignals that are implicated in a wide variety of cellular responses (1,2). In the epidermis, the major keratinocyte integrins area2b1, a3b1,and a6b4 (3). In addition to mediating keratinocyte adhesion toextracellular matrix proteins,b1 integrins regulate epidermal prolif-eration and terminal differentiation (4–8).
Integrin expression is normally restricted to keratinocytes in thebasal layer of the epidermis. However, integrin expression is alteredwhen the epidermis is damaged and in benign and neoplastic diseases.Suprabasal integrin expression has been observed during wound heal-ing and in psoriatic epidermis (9, 10). SCCs3 exhibit variable patternsof integrin expression, including normal expression, loss of expres-sion, and overexpression, and variation is observed both within andbetween individual tumors. In squamous papillomas and SCC, supra-basal expression ofa6b4, as well as increased basala6b4 expression,is associated with tumor progression and invasion (11–13). Highly
undifferentiated spindle cell carcinomas exhibit up-regulated expres-sion of thea5b1 integrin (14). Increased expression ofa2b1 anda3b1 integrin has been observed in basal cell carcinomas (3, 15).Complete or focal loss of expression ofa2b1, a3b1, anda6b4 hasalso been observed in SCC (16, 17). Hence, both loss of expressionand overexpression of keratinocyte integrins have been observedin SCC.
Various in vitro models have been established to examine thesignificance of alterations in keratinocyte integrin expression in SCC(13, 18, 19). The behavior of SCC lines has been analyzed before andafter introduction of specific integrin subunits. Although such studiesclearly demonstrate that abnormal integrin expression contributes tothe failure of SCC cells to differentiate normally, they suffer from thelimitation that it is not possible to study the role of integrins in tumordevelopment. To overcome this difficulty, we have made use of atransgenic mouse model that targets the expression of human integrinsubunits to the suprabasal layers of the epidermis via the involucrinpromoter (20, 21). Mice expressing suprabasala2b1 or a3b1 exhibitsporadic epidermal hyperplasia and skin inflammation, which areindicative of the benign hyperproliferative disease psoriasis (20, 21).However, no spontaneous epidermal tumors have been observed inthese lines. To determine whether suprabasal integrin expression canalter the sensitivity of keratinocytes to malignant conversion, we havesubjected transgenic mouse skin expressinga2b1 or a3b1 to achemical carcinogenesis protocol that allows evaluation of the devel-opment of both benign (papillomas) and malignant (SCC) tumors(22, 23).
MATERIALS AND METHODS
Reagents.DMBA was purchased from Argos Chemicals. TPA was pur-chased from LC Laboratories. Acetone was purchased from BDH Chemicals.BrdUrd was purchased from Sigma Chemical Co. Citra Plus antigen retrievalsolution was purchased from BioGenex (San Ramon, CA). Automation bufferwas purchased from Biomeda. Fluorescein apoptosis detection system waspurchased from Promega.
Animals. All transgenic and wt mice were generated and maintained in theImperial Cancer Research Fund Animal Containment unit. Animals were kepton a 12 h light/dark cycle and fed R20 rodent chow (Special Dietary Services,Essex, United Kingdom) and waterad libidum. a2 (founder line 1070; 49copies/cell),a3 (founder line 1120C; 20 copies/cell), andb1 (founder line0840; 42 copies/cell) transgenic mouse lines previously on an F1 hybrid(CBA 3 C57Bl/6) genetic background (20, 21) were backcrossed six gener-ations onto a homogeneous C57Bl/6 background.a2b1 and a3b1 doubletransgenic mice were then generated by crossinga2 anda3 single transgenicsto b1 single transgenics.
Tumor Studies. Femalea2b1 anda3b1 integrin transgenic and wt litter-mate mice 7 weeks of age (25 animals/group) were shaved once on the dorsalsurface with electric clippers. One week later all animals that did not showsigns of hair regrowth received one topical application of 100 nmol (25mg)DMBA in 200 ml of acetone or 200ml of acetone alone. One week later, micereceived thrice-weekly applications of 6 nmol (3.7mg) TPA in 200ml acetoneor 200 ml acetone alone. C57Bl/6 mice have been shown previously to beresistant to twice-weekly applications of TPA but do develop tumors with athrice-weekly protocol (22, 23). Benign and malignant skin tumors wererecorded once a week for a period up to 52 weeks after the start of promotion.Student’st test was conducted to statistically compare papilloma and malig-nancy formation betweena2b1 anda3b1 transgenic mice and wt mice. The
Received 1/8/01; accepted 4/25/01.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 This research was supported by the Imperial Cancer Research Fund and by NationalResearch Service Award CA75638 from the National Cancer Institute (to D. M. O.).
2 To whom requests for reprints should be addressed, at Imperial Cancer ResearchFund, Keratinocyte Laboratory, 44 Lincoln’s Inn Fields, London WC2A 3PX, UnitedKingdom.
3 The abbreviations used are: SCC, squamous cell carcinoma; DMBA, 7,12-dimeth-ylbenz[a]anthracene; TPA, 12-O-tetradecanoylphorbol-13-acetate; BrdUrd, bromode-oxyuridine; wt, wild-type; EMMPRIN, extracellular MMP inducer; TM4SF, transmem-brane-4 superfamily; TUNEL, terminal deoxynucleotidyl transferase-mediated nick endlabeling.
5248
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from

experiment was repeated with a second group of animals, and the resultsobtained in the first and second experiments were the same.
Short-Term TPA Treatment. Femalea2b1 anda3b1 transgenic and wtlittermate mice 7 weeks of age (five animals/group) were shaved once on thedorsal surface with electric clippers and 1 week later received either 1, 4, 8, or12 applications of 6 nmol TPA in 200ml acetone or 200ml acetone alone ata frequency of three applications per week. All skin sections were harvested24 h after the final TPA treatment.
Tissue Harvesting. One h before sacrifice, all mice received a single i.p.injection of 100 mg/kg BrdUrd. Skin tumors and dorsal skin sections wereharvested, and portions of each tumor or skin section were fixed in 10%neutral-buffered formalin or frozen in liquid nitrogen-cooled isopentane afterembedding in OCT medium. All formalin-fixed tumor sections were stainedwith H&E and graded as either a squamous papilloma or a SCC (24).
BrdUrd Labeling. To examine BrdUrd incorporation, formalin-fixed skinand tumor sections were processed as described previously (25). Briefly, tissuesections were deparaffinized and incubated in 2M HCl for 30 min at 37°C,dipped in borate buffer for 3 min, and then digested in 0.01% trypsin for 3 minat 37°C. Sections were blocked in 10% normal goat serum for 20 min and thenprobed with antibodies for BrdUrd (Becton Dickinson) for 1 h. Sections werethen incubated with a species-specific Alexa 488-conjugated (MolecularProbes) secondary antibody, after which sections were counterstained with 1mg/ml propidium iodide. Fluorescence was observed using a Zeiss Axiophotfluorescent microscope.
Immunofluorescence.For mouse keratin 10 staining, formalin-fixed tu-mor sections were deparaffinized and microwaved in Citra Plus antigen re-trieval solution for 2 min 30 s and allowed to cool in the antigen retrievalsolution for an additional 15 min. Sections were blocked in 10% normal goatserum for 20 min and then probed with antibodies for mouse keratin 10(Babco) for 1 h. Sections were then incubated with a species-specific Alexa488-conjugated (Molecular Probes) secondary antibody, and fluorescence wasobserved using a Zeiss Axiophot fluorescent microscope.
Frozen skin and tumor sections were fixed in acetone at220°C for 10 minprior to a 10% normal goat serum block and then probed with antibodies formousea6 integrin (CD49f; Serotec), mouseb1 integrin (MB1.2; courtesy ofBosco M. Chan, University of Western Ontario, London, Ontario, Canada;Ref. 26), rat EMMPRIN (CE9; courtesy of James Bartles, NorthwesternUniversity Medical School, Chicago, IL; Ref. 27), mouse CD81 (Eat2; cour-tesy of Shoshana Levy, Stanford University Medical Center, Stanford, CA;Ref. 28), mouse involucrin (29), humana3 integrin (VM-2; Ref. 21), or humana2 integrin (HAS4; Ref. 20). Tissue sections were then incubated with aspecies-specific Alexa 488-conjugated (Molecular Probes) secondary antibodyfor a6 integrin, EMMPRIN, andb1 integrin, or species-specific Alexa 594-conjugated (Molecular Probes) secondary antibodies fora2 anda3 integrinand involucrin, or FITC-conjugated antihamster (PharMingen) for CD81. Aftera 20 min secondary antibody incubation, sections were washed and mounted inGelvatol. Immunofluorescence was visualized using a Zeiss Axiophot fluores-cent microscope.
TUNEL Assay. Mouse papilloma and skin sections were processed fordetection of apoptotic cells according to the manufacturer’s protocol. Depar-affinized formalin-fixed papilloma and skin sections were incubated in 0.85%NaCl followed by PBS and then fixed in 4% paraformaldehyde, permeabilizedwith 20 mg/ml Proteinase K for 10 min, and fixed again in 4% paraformalde-hyde. Sections were incubated with terminal deoxynucleotidyl transferase inthe presence of fluorescein-12-dUTP at 37°C for 1 h. Sections were counter-stained with 1 mg/ml propidium iodide and examined under a Zeiss Axiophotfluorescent microscope.
RESULTS
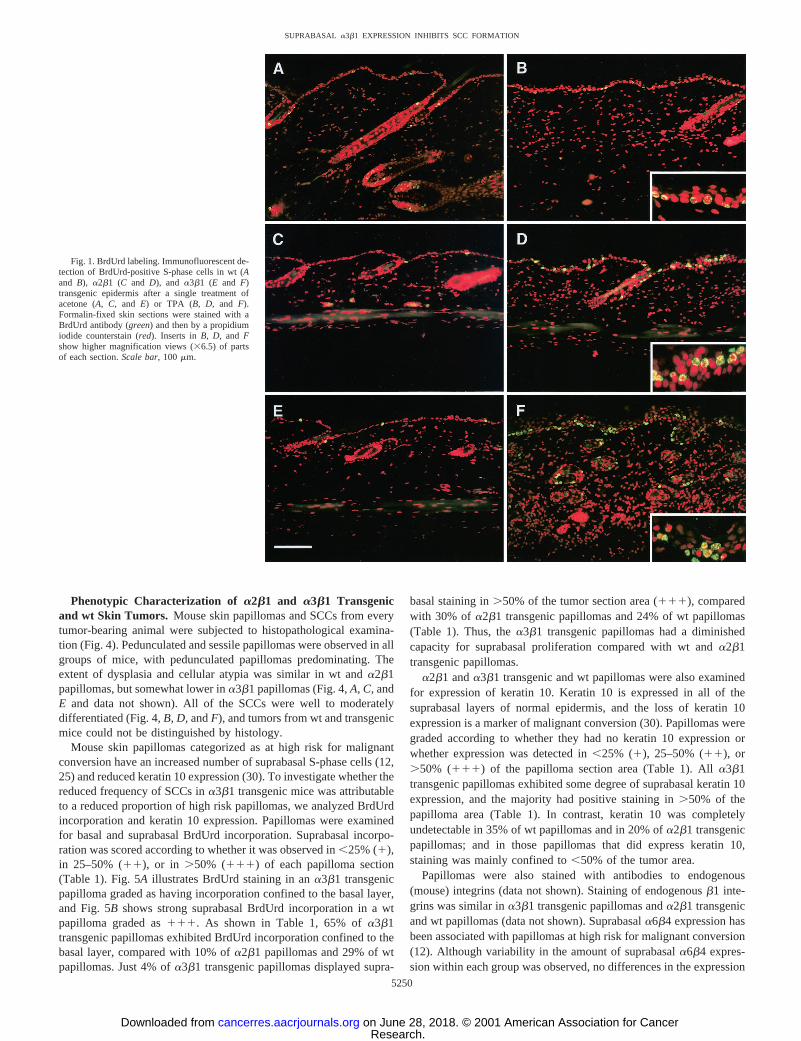
Suprabasal Keratinocyte Proliferation in a2b1 and a3b1Transgenic Mouse Skin.The previously reported sporadic hyper-proliferative and inflammatory phenotype ina2b1 anda3b1 trans-genic mouse skin is not observed in young animals either in theoriginal F1 background (CBA3 C57Bl/6) or after backcrossing ontoa homogeneous C57Bl/6 background (20, 21). As shown in Fig. 1panels A, C,andE, acetone vehicle-treateda2b1 anda3b1 transgenicmouse skin displayed a similar labeling index to wt mouse skin, with
all BrdUrd-positive S-phase cells residing in the basal layer. However,the transgenics showed a markedly different response to TPA from wtmice.a2b1 anda3b1 transgenic and wt mouse skin was treated with1, 4, 8, or 12 applications of 6 nmol TPA, and skin sections wereanalyzed for BrdUrd incorporation. After only a single application ofTPA, before the onset of significant hyperplasia, botha2b1 anda3b1transgenics exhibited BrdUrd incorporation in the suprabasal layers ofthe epidermis (Fig. 1,D and F), whereas no BrdUrd-positive kera-tinocytes were observed in the suprabasal layers of wt epidermis(Fig. 1B). The number of BrdUrd-positive keratinocytes in the basallayer did not differ betweena2b1 anda3b1 transgenic and wt mouseskin after any of the TPA treatment regimens. The suprabasal BrdUrdlabeling in transgenic epidermis was also present after 4, 8, and 12applications of TPA, whereas only basal BrdUrd incorporation wasobserved in wt mice (data not shown).
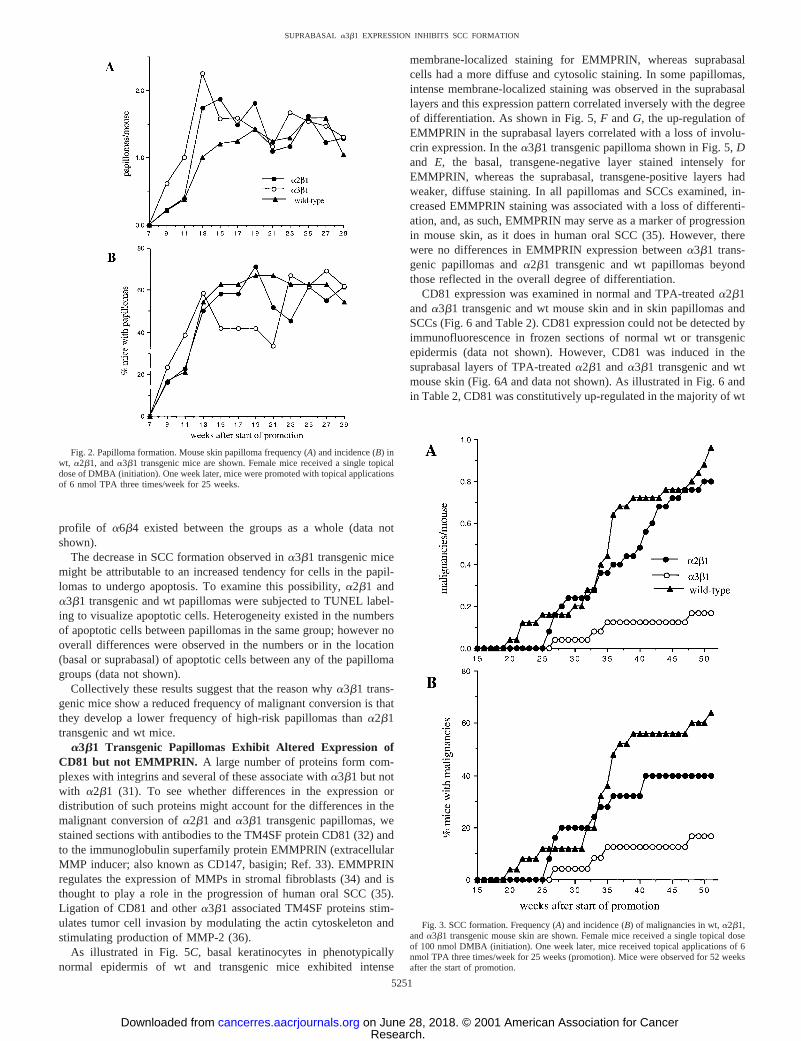
a2b1 and a3b1 Transgenic Mice and wt Mice Are EquallySensitive to Papilloma Formation.To determine whether the acutephenotypic response to short-term TPA treatment observed in trans-genic mouse skin (Fig. 1) could be extended to tumor formation, wecompared the responses of transgenic and wt mouse skin to chemicalcarcinogenesis.a2b1 anda3b1 transgenic and wt littermate femalemice were initiated with a single dose of 100 nmol DMBA. One weeklater, mice were treated with 6 nmol TPA three times/week for25 weeks. As shown in Fig. 2A, papillomas first emerged in all groups8 weeks after the start of TPA promotion, and maximum papillomaresponses reached a plateau by 20 weeks. No significant differences inpapilloma frequency were observed betweena2b1 (1.9 papillomas/mouse) ora3b1 transgenic mice (2.3 papillomas/mouse) and wt mice(1.6 papillomas/mouse; Fig. 2A; P 5 0.637 for a2b1 versuswt;P 5 0.352 fora3b1 versuswt). In addition, there was no differencein the incidence of papillomas (percentage of mice with papillomas) ina2b1 anda3b1 transgenic mice and wt mice (Fig. 2B). No papillomaswere observed in transgenic or wt mice that were initiated withDMBA and promoted with acetone vehicle or that were initiated withacetone vehicle and promoted with TPA (data not shown). In a repeatexperiment, no significant differences in papilloma incidence or fre-quency were observed betweena2b1 (1.88 papillomas/mouse) ora3b1 transgenic mice (2.13 papillomas/mouse) and wt mice (3.12papillomas/mouse;P 5 0.128 for a2b1 versuswt; P 5 0.164 fora3b1 versuswt).
a3b1 Transgenic Mice Are Resistant to SCC Formation.Todetermine whether aberrant integrin expression could influence ma-lignant conversion,a2b1 and a3b1 transgenic and wt mice weremaintained under weekly observation for SCC formation for a periodof 52 weeks after the start of TPA promotion. As shown in Fig. 3A,malignancies first emerged in wt mice 20 weeks after the start ofpromotion, whereas the first malignancies did not arise ina2b1 anda3b1 transgenic mice until 26 and 27 weeks, respectively. No differ-ence in the average number malignancies/mouse was observed be-tweena2b1 transgenic and wt mice (P5 0.583; Fig. 3A). However,a3b1 transgenic mice developed 5–6 fold fewer malignancies than wtmice (P 5 0.0005), and by 52 weeks, just three malignancies wereobserved in the entire group (Fig. 3A). As shown in Fig. 3B,a3b1transgenic mice also displayed a profound decrease in SCC incidencecompared witha2b1 transgenic and wt mice. H&E-stained histolog-ical sections from all malignancies were examined and graded to beSCCs in all groups (24). Similar decreases in SCC formation ina3b1transgenic (0.30 malignancies/mouse;P 5 0.020) but not ina2b1(0.61 malignancies/mouse;P 5 0.231) transgenic mice comparedwith wt (0.96 malignancies/mouse) mice were obtained in a repeatexperiment (data not shown).
5249
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from
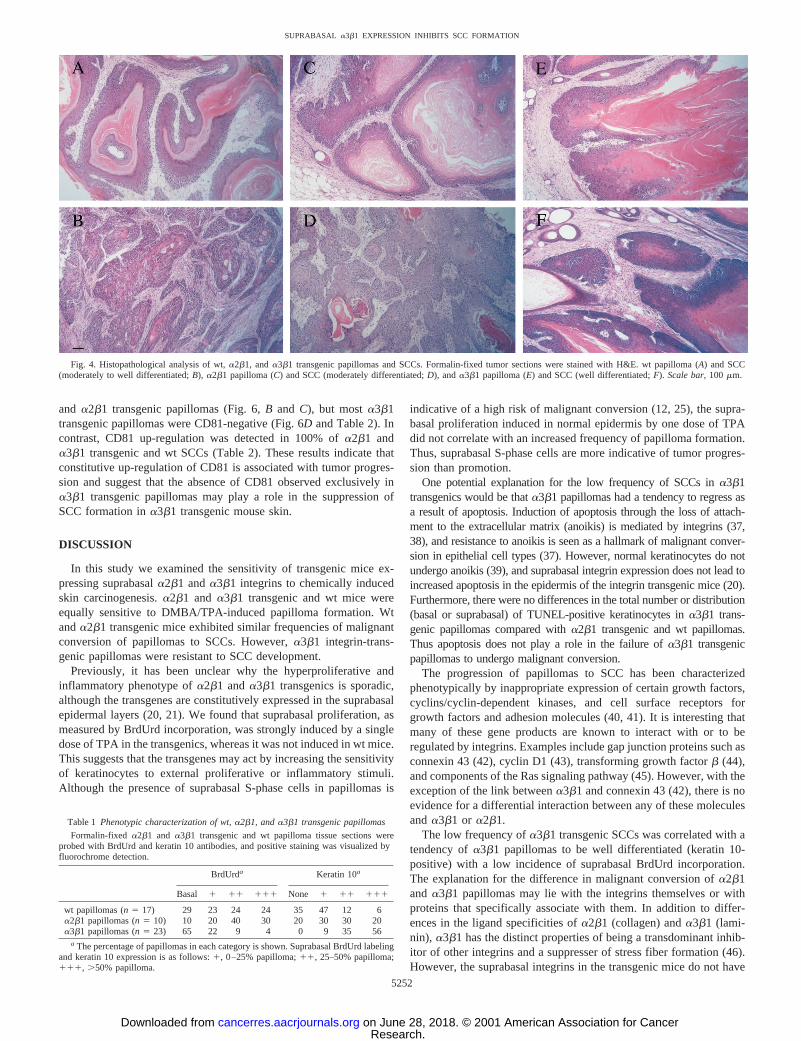
Phenotypic Characterization of a2b1 and a3b1 Transgenicand wt Skin Tumors. Mouse skin papillomas and SCCs from everytumor-bearing animal were subjected to histopathological examina-tion (Fig. 4). Pedunculated and sessile papillomas were observed in allgroups of mice, with pedunculated papillomas predominating. Theextent of dysplasia and cellular atypia was similar in wt anda2b1papillomas, but somewhat lower ina3b1 papillomas (Fig. 4,A, C,andE and data not shown). All of the SCCs were well to moderatelydifferentiated (Fig. 4,B, D,andF), and tumors from wt and transgenicmice could not be distinguished by histology.
Mouse skin papillomas categorized as at high risk for malignantconversion have an increased number of suprabasal S-phase cells (12,25) and reduced keratin 10 expression (30). To investigate whether thereduced frequency of SCCs ina3b1 transgenic mice was attributableto a reduced proportion of high risk papillomas, we analyzed BrdUrdincorporation and keratin 10 expression. Papillomas were examinedfor basal and suprabasal BrdUrd incorporation. Suprabasal incorpo-ration was scored according to whether it was observed in,25% (1),in 25–50% (11), or in.50% (111) of each papilloma section(Table 1). Fig. 5Aillustrates BrdUrd staining in ana3b1 transgenicpapilloma graded as having incorporation confined to the basal layer,and Fig. 5Bshows strong suprabasal BrdUrd incorporation in a wtpapilloma graded as111. As shown in Table 1, 65% ofa3b1transgenic papillomas exhibited BrdUrd incorporation confined to thebasal layer, compared with 10% ofa2b1 papillomas and 29% of wtpapillomas. Just 4% ofa3b1 transgenic papillomas displayed supra-
basal staining in.50% of the tumor section area (111), comparedwith 30% of a2b1 transgenic papillomas and 24% of wt papillomas(Table 1). Thus, thea3b1 transgenic papillomas had a diminishedcapacity for suprabasal proliferation compared with wt anda2b1transgenic papillomas.
a2b1 anda3b1 transgenic and wt papillomas were also examinedfor expression of keratin 10. Keratin 10 is expressed in all of thesuprabasal layers of normal epidermis, and the loss of keratin 10expression is a marker of malignant conversion (30). Papillomas weregraded according to whether they had no keratin 10 expression orwhether expression was detected in,25% (1), 25–50% (11), or.50% (111) of the papilloma section area (Table 1). Alla3b1transgenic papillomas exhibited some degree of suprabasal keratin 10expression, and the majority had positive staining in.50% of thepapilloma area (Table 1). In contrast, keratin 10 was completelyundetectable in 35% of wt papillomas and in 20% ofa2b1 transgenicpapillomas; and in those papillomas that did express keratin 10,staining was mainly confined to,50% of the tumor area.
Papillomas were also stained with antibodies to endogenous(mouse) integrins (data not shown). Staining of endogenousb1 inte-grins was similar ina3b1 transgenic papillomas anda2b1 transgenicand wt papillomas (data not shown). Suprabasala6b4 expression hasbeen associated with papillomas at high risk for malignant conversion(12). Although variability in the amount of suprabasala6b4 expres-sion within each group was observed, no differences in the expression
Fig. 1. BrdUrd labeling. Immunofluorescent de-tection of BrdUrd-positive S-phase cells in wt (Aand B), a2b1 (C and D), and a3b1 (E and F)transgenic epidermis after a single treatment ofacetone (A, C,and E) or TPA (B, D, and F).Formalin-fixed skin sections were stained with aBrdUrd antibody (green) and then by a propidiumiodide counterstain (red). Inserts inB, D, and Fshow higher magnification views (36.5) of partsof each section.Scale bar,100 mm.
5250
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from
profile of a6b4 existed between the groups as a whole (data notshown).
The decrease in SCC formation observed ina3b1 transgenic micemight be attributable to an increased tendency for cells in the papil-lomas to undergo apoptosis. To examine this possibility,a2b1 anda3b1 transgenic and wt papillomas were subjected to TUNEL label-ing to visualize apoptotic cells. Heterogeneity existed in the numbersof apoptotic cells between papillomas in the same group; however nooverall differences were observed in the numbers or in the location(basal or suprabasal) of apoptotic cells between any of the papillomagroups (data not shown).
Collectively these results suggest that the reason whya3b1 trans-genic mice show a reduced frequency of malignant conversion is thatthey develop a lower frequency of high-risk papillomas thana2b1transgenic and wt mice.
a3b1 Transgenic Papillomas Exhibit Altered Expression ofCD81 but not EMMPRIN. A large number of proteins form com-plexes with integrins and several of these associate witha3b1 but notwith a2b1 (31). To see whether differences in the expression ordistribution of such proteins might account for the differences in themalignant conversion ofa2b1 anda3b1 transgenic papillomas, westained sections with antibodies to the TM4SF protein CD81 (32) andto the immunoglobulin superfamily protein EMMPRIN (extracellularMMP inducer; also known as CD147, basigin; Ref. 33). EMMPRINregulates the expression of MMPs in stromal fibroblasts (34) and isthought to play a role in the progression of human oral SCC (35).Ligation of CD81 and othera3b1 associated TM4SF proteins stim-ulates tumor cell invasion by modulating the actin cytoskeleton andstimulating production of MMP-2 (36).
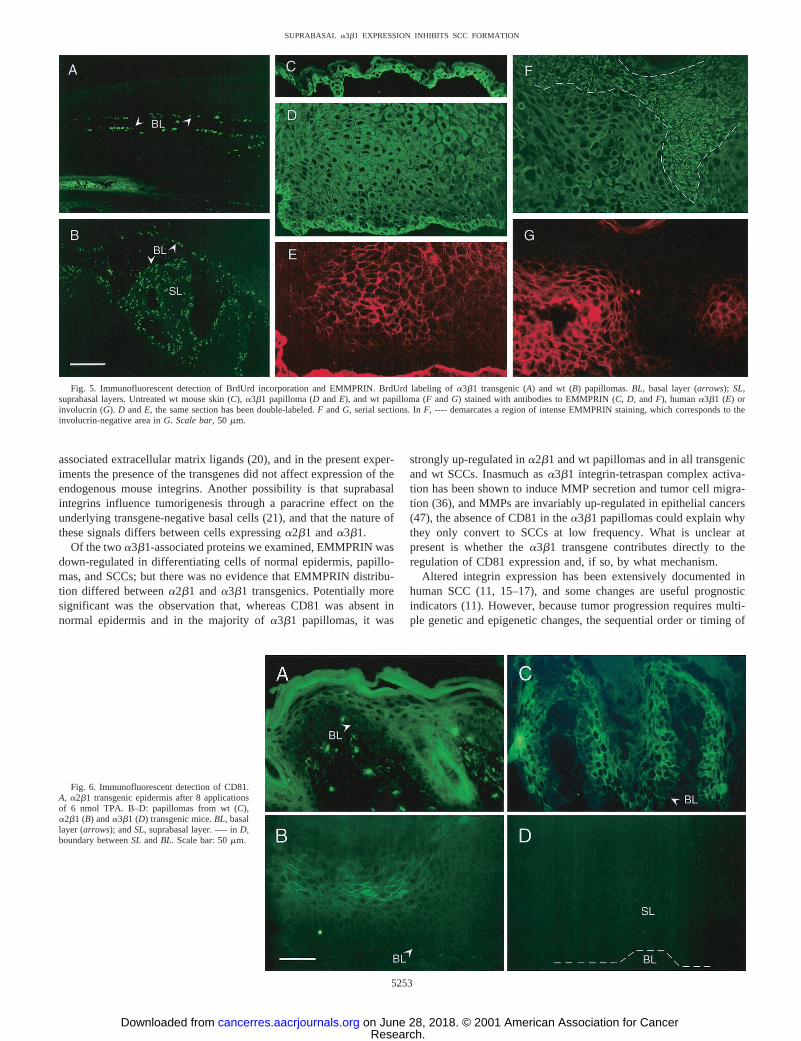
As illustrated in Fig. 5C, basal keratinocytes in phenotypicallynormal epidermis of wt and transgenic mice exhibited intense
membrane-localized staining for EMMPRIN, whereas suprabasalcells had a more diffuse and cytosolic staining. In some papillomas,intense membrane-localized staining was observed in the suprabasallayers and this expression pattern correlated inversely with the degreeof differentiation. As shown in Fig. 5,F andG, the up-regulation ofEMMPRIN in the suprabasal layers correlated with a loss of involu-crin expression. In thea3b1 transgenic papilloma shown in Fig. 5,Dand E, the basal, transgene-negative layer stained intensely forEMMPRIN, whereas the suprabasal, transgene-positive layers hadweaker, diffuse staining. In all papillomas and SCCs examined, in-creased EMMPRIN staining was associated with a loss of differenti-ation, and, as such, EMMPRIN may serve as a marker of progressionin mouse skin, as it does in human oral SCC (35). However, therewere no differences in EMMPRIN expression betweena3b1 trans-genic papillomas anda2b1 transgenic and wt papillomas beyondthose reflected in the overall degree of differentiation.
CD81 expression was examined in normal and TPA-treateda2b1anda3b1 transgenic and wt mouse skin and in skin papillomas andSCCs (Fig. 6 and Table 2). CD81 expression could not be detected byimmunofluorescence in frozen sections of normal wt or transgenicepidermis (data not shown). However, CD81 was induced in thesuprabasal layers of TPA-treateda2b1 anda3b1 transgenic and wtmouse skin (Fig. 6Aand data not shown). As illustrated in Fig. 6 andin Table 2, CD81 was constitutively up-regulated in the majority of wt
Fig. 3. SCC formation. Frequency (A) and incidence (B) of malignancies in wt,a2b1,anda3b1 transgenic mouse skin are shown. Female mice received a single topical doseof 100 nmol DMBA (initiation). One week later, mice received topical applications of 6nmol TPA three times/week for 25 weeks (promotion). Mice were observed for 52 weeksafter the start of promotion.
Fig. 2. Papilloma formation. Mouse skin papilloma frequency (A) and incidence (B) inwt, a2b1, anda3b1 transgenic mice are shown. Female mice received a single topicaldose of DMBA (initiation). One week later, mice were promoted with topical applicationsof 6 nmol TPA three times/week for 25 weeks.
5251
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from
and a2b1 transgenic papillomas (Fig. 6,B and C), but mosta3b1transgenic papillomas were CD81-negative (Fig. 6D and Table 2). Incontrast, CD81 up-regulation was detected in 100% ofa2b1 anda3b1 transgenic and wt SCCs (Table 2). These results indicate thatconstitutive up-regulation of CD81 is associated with tumor progres-sion and suggest that the absence of CD81 observed exclusively ina3b1 transgenic papillomas may play a role in the suppression ofSCC formation ina3b1 transgenic mouse skin.
DISCUSSION
In this study we examined the sensitivity of transgenic mice ex-pressing suprabasala2b1 anda3b1 integrins to chemically inducedskin carcinogenesis.a2b1 and a3b1 transgenic and wt mice wereequally sensitive to DMBA/TPA-induced papilloma formation. Wtanda2b1 transgenic mice exhibited similar frequencies of malignantconversion of papillomas to SCCs. However,a3b1 integrin-trans-genic papillomas were resistant to SCC development.
Previously, it has been unclear why the hyperproliferative andinflammatory phenotype ofa2b1 anda3b1 transgenics is sporadic,although the transgenes are constitutively expressed in the suprabasalepidermal layers (20, 21). We found that suprabasal proliferation, asmeasured by BrdUrd incorporation, was strongly induced by a singledose of TPA in the transgenics, whereas it was not induced in wt mice.This suggests that the transgenes may act by increasing the sensitivityof keratinocytes to external proliferative or inflammatory stimuli.Although the presence of suprabasal S-phase cells in papillomas is
indicative of a high risk of malignant conversion (12, 25), the supra-basal proliferation induced in normal epidermis by one dose of TPAdid not correlate with an increased frequency of papilloma formation.Thus, suprabasal S-phase cells are more indicative of tumor progres-sion than promotion.
One potential explanation for the low frequency of SCCs ina3b1transgenics would be thata3b1 papillomas had a tendency to regress asa result of apoptosis. Induction of apoptosis through the loss of attach-ment to the extracellular matrix (anoikis) is mediated by integrins (37,38), and resistance to anoikis is seen as a hallmark of malignant conver-sion in epithelial cell types (37). However, normal keratinocytes do notundergo anoikis (39), and suprabasal integrin expression does not lead toincreased apoptosis in the epidermis of the integrin transgenic mice (20).Furthermore, there were no differences in the total number or distribution(basal or suprabasal) of TUNEL-positive keratinocytes ina3b1 trans-genic papillomas compared witha2b1 transgenic and wt papillomas.Thus apoptosis does not play a role in the failure ofa3b1 transgenicpapillomas to undergo malignant conversion.
The progression of papillomas to SCC has been characterizedphenotypically by inappropriate expression of certain growth factors,cyclins/cyclin-dependent kinases, and cell surface receptors forgrowth factors and adhesion molecules (40, 41). It is interesting thatmany of these gene products are known to interact with or to beregulated by integrins. Examples include gap junction proteins such asconnexin 43 (42), cyclin D1 (43), transforming growth factorb (44),and components of the Ras signaling pathway (45). However, with theexception of the link betweena3b1 and connexin 43 (42), there is noevidence for a differential interaction between any of these moleculesanda3b1 or a2b1.
The low frequency ofa3b1 transgenic SCCs was correlated with atendency ofa3b1 papillomas to be well differentiated (keratin 10-positive) with a low incidence of suprabasal BrdUrd incorporation.The explanation for the difference in malignant conversion ofa2b1and a3b1 papillomas may lie with the integrins themselves or withproteins that specifically associate with them. In addition to differ-ences in the ligand specificities ofa2b1 (collagen) anda3b1 (lami-nin), a3b1 has the distinct properties of being a transdominant inhib-itor of other integrins and a suppresser of stress fiber formation (46).However, the suprabasal integrins in the transgenic mice do not have
Table 1 Phenotypic characterization of wt,a2b1, anda3b1 transgenic papillomas
Formalin-fixed a2b1 and a3b1 transgenic and wt papilloma tissue sections wereprobed with BrdUrd and keratin 10 antibodies, and positive staining was visualized byfluorochrome detection.
BrdUrda Keratin 10a
Basal 1 11 111 None 1 11 111
wt papillomas (n5 17) 29 23 24 24 35 47 12 6a2b1 papillomas (n5 10) 10 20 40 30 20 30 30 20a3b1 papillomas (n5 23) 65 22 9 4 0 9 35 56
a The percentage of papillomas in each category is shown. Suprabasal BrdUrd labelingand keratin 10 expression is as follows:1, 0–25% papilloma;11, 25–50% papilloma;111, .50% papilloma.
Fig. 4. Histopathological analysis of wt,a2b1, anda3b1 transgenic papillomas and SCCs. Formalin-fixed tumor sections were stained with H&E. wt papilloma (A) and SCC(moderately to well differentiated;B), a2b1 papilloma (C) and SCC (moderately differentiated;D), anda3b1 papilloma (E) and SCC (well differentiated;F). Scale bar,100 mm.
5252
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from
associated extracellular matrix ligands (20), and in the present exper-iments the presence of the transgenes did not affect expression of theendogenous mouse integrins. Another possibility is that suprabasalintegrins influence tumorigenesis through a paracrine effect on theunderlying transgene-negative basal cells (21), and that the nature ofthese signals differs between cells expressinga2b1 anda3b1.
Of the twoa3b1-associated proteins we examined, EMMPRIN wasdown-regulated in differentiating cells of normal epidermis, papillo-mas, and SCCs; but there was no evidence that EMMPRIN distribu-tion differed betweena2b1 anda3b1 transgenics. Potentially moresignificant was the observation that, whereas CD81 was absent innormal epidermis and in the majority ofa3b1 papillomas, it was
strongly up-regulated ina2b1 and wt papillomas and in all transgenicand wt SCCs. Inasmuch asa3b1 integrin-tetraspan complex activa-tion has been shown to induce MMP secretion and tumor cell migra-tion (36), and MMPs are invariably up-regulated in epithelial cancers(47), the absence of CD81 in thea3b1 papillomas could explain whythey only convert to SCCs at low frequency. What is unclear atpresent is whether thea3b1 transgene contributes directly to theregulation of CD81 expression and, if so, by what mechanism.
Altered integrin expression has been extensively documented inhuman SCC (11, 15–17), and some changes are useful prognosticindicators (11). However, because tumor progression requires multi-ple genetic and epigenetic changes, the sequential order or timing of
Fig. 5. Immunofluorescent detection of BrdUrd incorporation and EMMPRIN. BrdUrd labeling ofa3b1 transgenic (A) and wt (B) papillomas.BL, basal layer (arrows);SL,suprabasal layers. Untreated wt mouse skin (C),a3b1 papilloma (DandE), and wt papilloma (FandG) stained with antibodies to EMMPRIN (C, D,andF), humana3b1 (E) orinvolucrin (G).D andE, the same section has been double-labeled.F andG, serial sections. InF, ---- demarcates a region of intense EMMPRIN staining, which corresponds to theinvolucrin-negative area inG. Scale bar,50 mm.
Fig. 6. Immunofluorescent detection of CD81.A, a2b1 transgenic epidermis after 8 applicationsof 6 nmol TPA. B–D: papillomas from wt (C),a2b1 (B) anda3b1 (D) transgenic mice.BL, basallayer (arrows); andSL,suprabasal layer. ---- inD,boundary betweenSL andBL. Scale bar: 50mm.
5253
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from

alterations in integrin expression may be critical, and early changes inintegrin expression may have more impact on the course of the diseasethan the changes that characterize a mature tumor. The value of ourtransgenic mouse model is that it can be used to discover how earlychanges in integrin expression affect later stages in the disease.
ACKNOWLEDGMENTS
We thank Robert Rudling, Barbara Cross, and Toni Hill for their experttechnical assistance. We are grateful to James Bartles for providing the CE9antibody, to Shoshana Levy for the Eat2 antibody, and to Bosco Chan for theMB1.2 antibody.
REFERENCES
1. Hynes, R. O. Integrins: versatility, modulation, and signaling in cell adhesion. Cell,69: 11–25, 1992.
2. Giancotti, F. G., and Ruoslahti, E. Integrin signaling. Science (Wash. DC),285:1028–1032, 1999.
3. Peltonen, J., Larjava, H., Jaakkola, S., Gralnick, H., Akiyama, S. K., Yamada, S. S.,Yamada, K. M., and Uitto, J. Localization of integrin receptors for fibronectin,collagen, and laminin in human skin. J. Clin. Investig.,84: 1916–1923, 1989.
4. Jones, P. H., Harper, S., and Watt, F. M. Stem cell patterning and fate in humanepidermis. Cell,80: 83–93, 1995.
5. Zhu, A. J., Haase, I., and Watt, F. M. Signaling viab1 integrins and mitogen-activatedprotein kinase determines human epidermal stem cell fatein vitro. Proc. Natl. Acad.Sci. USA,96: 6728–6733, 1999.
6. Levy, L., Broad, S., Diekmann, D., Evans, R. D., and Watt, F. M.b1 integrinsregulate keratinocyte adhesion and differentiation by distinct mechanisms. Mol. Biol.Cell, 11: 453–466, 2000.
7. Brakebusch, C., Grose, R., Quondamatteo, F., Ramirez, A., Jorcano, J. L., Pirro, A.,Svensson, M., Herken, R., Sasaki, T., Timpl, R., Werner, S., and Fassler, R. Skin andhair follicle integrity is crucially dependent onb1 integrin expression on keratino-cytes. EMBO J.,19: 3990–4003, 2000.
8. Raghavan, S., Bauer, C., Mundschau, G., Li, Q., and Fuchs, E. Conditional ablationof b1 integrin in skin. Severe defects in epidermal proliferation, basement membraneformation, and hair follicle invagination. J. Cell Biol.,150: 1149–1160, 2000.
9. Hertle, M. D., Kubler, M-D., Leigh, I. M., and Watt, F. M. Aberrant integrinexpression during epidermal wound healing and in psoriatic epidermis. J. Clin.Investig.,89: 1892–1901, 1992.
10. Haapasalmi, K., Zhang, K., Tonnesen, M., Olerud, J., Sheppard, D., Salo, T., Kramer,R., Clark, R. A. F., Uitto, V-J., and Larjava, H. Keratinocytes in human woundsexpressavb6 integrin. J. Investig. Dermatol.,106: 42–48, 1996.
11. Wolf, G. T., Carey, T. E., Schmaltz, S. P., McClatchey, K. D., Poore, J., Glaser, L.,Hayashida, D. J. S., and Hsu, S. Altered antigen expression predicts outcome in squamouscell carcinoma of the head and neck. J. Natl. Cancer Inst.,82: 1566–1572, 1990.
12. Tennenbaum, T., Weiner, A. K., Belanger, A. J., Glick, A. B., Hennings, H., andYuspa, S. H. The suprabasal expression ofa6b4 integrin is associated with a high riskfor malignant progression in mouse skin carcinogenesis. Cancer Res.,53: 4803–4810, 1993.
13. Shaw, L. M., Rabinovitz, I., Wang, H. H., Toker, A., and Mercurio, A. M. Activationof phosphoinositide 3-OH kinase by thea6b4 integrin promotes carcinoma invasion.Cell, 91: 949–960, 1997.
14. Gomez, M., and Cano, A. Expression ofb1 integrin receptors in transformed mouseepidermal keratinocytes: upregulation ofa5b1 in spindle carcinoma cells. Mol.Carcinog.,12: 153–165, 1995.
15. Stamp, G. W. H., and Pignatelli, M. Distribution ofb1, a1, a2, anda3 integrin chainsin basal cell carcinomas. J. Pathol.,163: 307–313, 1991.
16. Downer, C. S., Watt, F. M., and Speight, P. M. Loss ofa6 andb4 subunits coincideswith loss of basement membrane components in oral squamous cell carcinomas.J. Pathol.,171: 183–190, 1993.
17. Jones, J., Sugiyama, M., Watt, F. M., and Speight, P. M. Integrin expression innormal, hyperplastic, dysplastic, and malignant oral epithelium. J. Pathol.,169:235–243, 1993.
18. Jones, J., Sugiyama, M., Speight, P. M., and Watt, F. M. Restoration ofavb5 integrinexpression in neoplastic keratinocytes results in increased capacity for terminaldifferentiation and suppression of anchorage-independent growth. Oncogene,12:119–126, 1996.
19. Jones, J., Sugiyama, M., Giancotti, F., Speight, P. M., and Watt, F. M. Transfectionof b4 integrin subunit into a neoplastic cell line fails to restore terminal differentiationcapacity or influence proliferation. Cell Adhes. Comm.,4: 307–316, 1996.
20. Carroll, J. M., Romero, M. R., and Watt, F. M. Suprabasal integrin expression in theepidermis of transgenic mice results in developmental defects and a phenotyperesembling psoriasis. Cell,83: 957–968, 1995.
21. Romero, M. R., Carroll, J. M., and Watt, F. M. Analysis of cultured keratinocytesfrom a transgenic mouse model of psoriasis: effects of suprabasal integrin expressionon keratinocyte adhesion, proliferation and terminal differentiation. Exp. Dermatol.,8: 53–67, 1999.
22. Fischer, S. M., Jasheway, D. W., Klann, R. C., Butler, A. P., Patrick, K. E., Baldwin,J. K., and Cameron, G. S. Correlation of phorbol ester promotion in the resistantC57Bl/6J mouse with sustained hyperplasia but not ornithine decarboxylase or proteinkinase C. Cancer Res.,49: 6693–6699, 1989.
23. Yuspa, S. H. The pathogenesis of squamous cell cancer: lessons learned from studiesof skin carcinogenesis—Thirty-third G. H. A. Clowes Memorial Award Lecture.Cancer Res.,54: 1178–1189, 1994.
24. Bogovski, P. Tumours of the skin.In: V. Turusov and U. Mohr (eds.), Pathology ofTumours in Laboratory Animals, Vol. 2, pp.1–45. Lyon, France: IARC, 1994.
25. Owens, D. M., Wei, S-J. C., and Smart, R. C. A multihit, multistage model ofchemical carcinogenesis. Carcinogenesis (Lond.),20: 1837–1844, 1999.
26. Von Ballestrem, C. G., Uniyal, S., McCormick, J. I., Chau, T., Singh, B., and Chan,B. M. VLA- b1 integrin subunit-specific monoclonal antibodies MB1.1 and MB1.2:binding to epitopes not dependent on thymocyte development or regulated by phorbolester and divalent cations. Hybridoma,15: 125–132, 1996.
27. Petruszak, J. A., Nehme, C. L., and Bartles, J. R. Endoproteolytic cleavage in theextracellular domain of the integral plasma membrane protein CE9 precedes itsredistribution from the posterior to the anterior tail of the rat spermatozoon duringepididymal maturation. J. Cell. Biol.,114: 917–927, 1991.
28. Maecker, H. T., Todd, S. C., Kim, E. C., and Levy, S. Differential expression ofmurine CD81 highlighted by new anti-mouse CD81 monoclonal antibodies. Hybri-doma,19: 15–22, 2000.
29. Li, E. R., Owens, D. M., Djian, P., and Watt, F. M. Expression of involucrin innormal, hyperproliferative and neoplastic mouse keratinocytes. Exp. Dermatol.,9:431–438, 2000.
30. Roop, D. R. Keratin expression in mouse epidermal tumors.In: C. J. Conti, T. J.Slaga, and A. J. P. Klein-Szanto (eds.), Skin Tumors: Experimental and ClinicalAspects pp. 257–271. New York: Raven Press Ltd., 1989.
31. Hemler, M. E. Integrin associated proteins. Curr. Opin. Cell Biol.,10: 578–585, 1998.32. Berditchevski, F., Zutter, M. M., and Hemler, M. E. Characterization of novel
complexes on the cell surface between integrins and proteins with 4 transmembranedomains (TM4 proteins). Mol. Biol. Cell,7: 193–207, 1996.
33. Berditchevski, F., Chang, S., Bodorova, J., and Hemler, M. E. Generation of mono-clonal antibodies to integrin-associated proteins. Evidence thata3b1 complexes withEMMPRIN/basigin/OX47/M6. J. Biol. Chem.,272: 29174–29180, 1997.
34. Guo, G., Zucker, S., Gordon, M., Toole, B., and Biswas, C. Stimulation of matrixmetalloproteinase production by recombinant extracellular matrix metalloproteinase in-ducer from transfected Chinese hamster ovary cells. J. Biol. Chem.,272: 24–27, 1997.
35. Bordador, L. C., Li, X., Toole, B., Chen, B., Regezi, J., Zardi, L., Hu, Y., and Ramos,D. M. Expression of emmprin by oral squamous cell carcinoma. Int. J. Cancer,85:347–352, 2000.
36. Sugiura, T., and Berditchevski, F. Function ofa3b1-tetraspanin protein complexes intumor cell invasion. Evidence for the role of the complexes in production of matrixmetalloproteinase 2 (MMP-2). J. Cell Biol.,146: 1375–1389, 1999.
37. Varner, J. A., and Cheresh, D. A. Integrins and cancer. Curr. Opin. Cell Biol.,8:724–730, 1996.
38. Frisch, S. M., and Ruoslahti, E. Integrins and anoikis. Curr. Opin. Cell Biol.,9:701–701, 1997.
39. Gandarillas, A., Goldsmith, L. A., Gschmeissner, S., Leigh, I. M., and Watt, F. M.Evidence that apoptosis and terminal differentiation of epidermal keratinocytes aredistinct processes. Exp. Dermatol.,8: 71–79, 1999.
40. Hennings, H., Glick, A. B., Greenhalgh, D. A., Morgan, D. L., Strickland, J. E.,Tennenbaum, T., and Yuspa, S. H. Critical aspects of initiation, promotion, andprogression in multistage epidermal carcinogenesis. Proc. Soc. Exp. Biol. Med.,202:1–8, 1993.
41. Frame, S., and Balmain, A. Integration of positive and negative growth signals duringras pathway activationin vivo. Curr. Opin. Genet. Dev.,10: 106–113, 2000.
42. Lampe, P. D., Nguyen, B. P., Gil, S., Usui, M., Olerud, J., Takada, Y., and Carter,W. G. Cellular interaction of integrina3b1 with laminin 5 promotes gap junctionalcommunication. J. Cell Biol.,143: 1735–1747, 1998.
43. Roovers, K., Davey, G., Zhu, X., Bottazzi, M. E., and Assoian, R. K.a5b1 integrincontrols cyclin D1 expression by sustaining mitogen-activated protein kinase activityin growth factor-treated cells. Mol. Biol. Cell,10: 3197–3204, 1999.
44. Dalton, S. L., Scharf, E., Davey, G., and Assoian, R. K. Transforming growth factor-boverrides the adhesion requirement for surface expression ofa5b1 in normal ratkidney fibroblasts. A necessary effect for induction of anchorage-independent growth.J. Biol. Chem.,274: 30139–30145, 1999.
45. Parise, L. V., Lee, J. W., and Juliano, R. L. New aspects of integrin signalling incancer. Cancer Biol.,10: 407–414, 2000.
46. Hodivala-Dilke, K. M., DiPersio, C. M., Kreidberg, J. A., and Hynes, R. O. Novelroles for a3b1 integrin as a regulator of cytoskeletal assembly and as a trans-dominant inhibitor of integrin receptor function in mouse keratinocytes. J. Cell Biol.,142: 1357–1369, 1998.
47. Sternlicht, M. D., Bissell, M. J., and Werb, Z. The matrix metalloproteinasestromelysin-1 acts as a natural mammary tumor promoter. Oncogene,19: 1102–1113, 2000.
Table 2 Incidence of CD81 expression in papillomas and SCCs
Acetone-fixeda2b1 anda3b1 transgenic and wt papilloma and SCC frozen sectionswere probed with an antimouse CD81 antibody, and positive staining was visualized byFITC detection.
% CD81 positive tumors
Papilloma SCC
wt 100 (7/7) 100 (7/7)a2b1 transgenic 88 (7/8) 100 (3/3)a3b1 transgenic 25 (2/8) 100 (6/6)
5254
SUPRABASAL a3b1 EXPRESSION INHIBITS SCC FORMATION
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from

2001;61:5248-5254. Cancer Res David M. Owens and Fiona M. Watt
1, Suppresses Malignant Conversionβ2αnot 1, butβ3αCarcinoma Formation in a Transgenic Mouse Model:
1 Integrins on Epidermal Squamous CellβInfluence of
Updated version
http://cancerres.aacrjournals.org/content/61/13/5248
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/61/13/5248.full#ref-list-1
This article cites 42 articles, 17 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/61/13/5248.full#related-urls
This article has been cited by 12 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/61/13/5248To request permission to re-use all or part of this article, use this link
Research. on June 28, 2018. © 2001 American Association for Cancercancerres.aacrjournals.org Downloaded from