Embed Size (px)
Citation preview

NeuroToxicology xxx (2010) xxx–xxx
G Model
NEUTOX-1176; No. of Pages 10
Inflammation-like glial response in rat brain induced by prenatalPFOS exposure
Huai-cai Zeng a,b,1, Ling Zhang a,1, Yuan-yuan Li a, Yan-jian Wang a, Wei Xia a,Yi Lin a, Jie Wei a, Shun-qing Xu a,*a Ministry of Education Key Laboratory of Environment and Health, School of Public Health, Tongji Medical College,
Huazhong University of Science and Technology, Wuhan 430030, Chinab School of Public Health, University of South China, Hengyang 421001, China
A R T I C L E I N F O
Article history:
Received 17 June 2010
Accepted 5 October 2010
Available online xxx
Keywords:
PFOS
Inflammatory
Glia
Cytokines
A B S T R A C T
Numerous studies have indicated the neurotoxicity of perfluorooctane sulfonate (PFOS), a persistent and
bioaccumulative compound, particularly during developmental stages of higher organisms. To explore
the pro-inflammatory effect in the developmental neurotoxicity, effects of prenatal exposure to PFOS on
glial activation in hippocampus and cortex were examined in offspring rats. Dams received 0.1, 0.6 and
2.0 mg/kg bw PFOS by gavage from gestational day 2 (GD2) to GD21. Astrocyte activation markers, glial
fibrillary acidic protein (GFAP) and S100 calcium binding protein B (S-100b) in hippocampus and cortex
were both upregulated on postnatal day 0 (PND0) or PND21. In addition, the astrocyte activation was
accompanied with the elevation of pro-inflammatory cytokines interleukin (IL-1b) and tumor necrosis
factor (TNF)-a. The mRNA levels of pro-inflammatory transcription factors, including activation protein-
1 (AP-1), nuclear factor-kB (NF-kB), and cAMP response element-binding protein (CREB) were also
increased, at least in the 2.0 mg/kg group. In addition to the inflammatory response, two synaptic
proteins, synapsin 1 (Syn1) and synaptophysin (Syp) were reduced in cortex on PND0 and PND21. In
hippocampus, the Syn1 were also reduced, while the Syp were increased in cortex on either PND0 or
PND21. Obtained results indicated chronic glial activation with coexisting inflammatory and synapse
injury features as a new mechanism of PFOS developmental neurotoxicity, and enhanced expression of
AP-1, NF-kB and CREB may contributed to the adverse effect.
� 2010 Elsevier Inc. All rights reserved.
Contents lists available at ScienceDirect
NeuroToxicology
1. Induction
Perfluorooctane sulfonate (PFOS), which is a kind of fluorine-saturated eight-carbon compounds and belongs to the degrada-tion product of many perfluorinated compounds, has been used ina variety of commercial and industrial applications, such aswetting agents, fire retardants, stain resistant treatments forleather and carpets, food packaging bags, cosmetics, componentsof pharmaceuticals and insecticides (Giesy and Kannan, 2002;Renner, 2001; Sohlenius et al., 1994). PFOS is believed to be anemerging persistent organic pollutant because of its worldwidedistribution, extreme stability and bioaccumulative nature(Renner, 2001). The accumulation of PFOS in mammals is mainlyin the liver and serum, as well as in the brain up to as high as 32%of the serum concentration (Chang et al., 2009). The develop-mental neurotoxictiy induced by PFOS has attracted more
* Corresponding author. Tel.: +86 27 83657705; fax: +86 27 83657781.
E-mail address: [email protected] (S.-q. Xu).1 Both authors contributed equally to this work.
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
0161-813X/$ – see front matter � 2010 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuro.2010.10.001
attention because that PFOS could cross the placenta barrier(Apelberg et al., 2007; Olsen et al., 2008) and blood–brain barrier(Austin et al., 2003), and also presented in milk (Karrman et al.,2009; Tao et al., 2008a,b; von Ehrenstein et al., 2009). Thus, thedeveloping fetus and infants were at high risk for PFOS exposure.Exposure to PFOS in uterus had been known to cause a strongereffect on development than lactation or adult exposure in rat ormice (Lau et al., 2003; Luebker et al., 2005). Recently, Fuenteset al. (2007) evaluated behavioural effects of PFOS on adult mice,and found that administration PFOS to adult mice resulted in adeficit in spatial memory. Johansson et al. (2008) found that muchlower developmental exposures of mice to PFOS and PFOAproduced abnormal cognitive function and behavioural defectspersisting into adulthood. Butenhoff et al. (2009) reported thatgestational and lactation exposure to PFOS in rat resulted inincreased motor activity and impaired cognitive function onPND17. In short, increased motor activity, reduced habitation andimpaired cognitive function have been noted in developmentalneurotoxicological evaluations in PFOS exposed rats and mice(Butenhoff et al., 2009; Fuentes et al., 2007; Johansson et al.,2008; Liu et al., 2009).
glial response in rat brain induced by prenatal PFOS exposure.

H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx2
G Model
NEUTOX-1176; No. of Pages 10
Pro-inflammatory effects of PFOS have been observed in severaltissues and organs. For example, administration of PFOS to adult ratsfor 28 days resulted in focal hemorrhage, erythrocytic transudation,and focal hepatocytic degeneration accompanying by inflammatorycellular infiltration in liver (Cui et al., 2009). In addition, PFOSexposure resulted in the brain injury, including focal hyperplasia ofcerebral gliocytes, dilatation and congestion in inferior caval veins ofcerebral arachnoid mater accompanied by slight focal hemorrhage,broadened gap around veins and neurocytes, and focal demyelin-ation of nerve fibers (Cui et al., 2009). Short-term exposure of PFOSwith higher concentration to mice increased the expression ofinflammatory factors in peritoneal macrophages, such as TNF-a andIL-6 (Qazi et al., 2009). In addition, PFOS exposure enhanced theinflammatory responses of macrophages to lipopolysaccharide andaltered the immunological function (Qazi et al., 2009). SubchronicPFOS exposure for adult male C57BL/6 mice elevated the expressionof pro-inflammatory cytokines TNF-alpha, IL-1beta, IL-6, in spleen(Dong et al., in press). Therefore, we speculated that pro-inflammatory effects contributed to the developmental neurotox-icological process.
Inflammation in the CNS is driven by the activation of residentmicroglia, astrocytes and infiltrating peripheral macrophages(Whitney et al., 2009). Both activated astroglia and microgliamay generate and/or maintain the inflammatory reaction in brainby producing cytokines (Graeber and Streit, 2009; Sofroniew andVinters, 2009), and enhance expression of GFAP (Sofroniew andVinters, 2009), which is a marker of astrocyte activation. In thehealthy CNS, the expression levels of cytokines in the CNS are verylow or undetectable due to accurate expression control. However,under pathological conditions, such as injury, infection, autoim-mune disease, and exposure to toxicant, the expression levels ofvarious cytokines show a significant increase. Some of thesecytokines are involved in neuronal cell damage, while others play aneuroprotective role. It is postulated inflammatory factors releasedby glial such as tumor necrosis factor-a (TNF-a), IL-1b and IL-6contribute to the destructive processes resulting in neuronal celldamage. These inflammatory cytokines contribute to pathophysi-ological changes associated with several neurodegenerativedisorders, including Alzheimer’s, Parkinson’s, and multiple sclero-sis (Graeber and Streit, 2009; Sofroniew and Vinters, 2009).
In the present study, we examined whether the prenatalexposure of PFOS would induce proneuroinflammatory in off-spring. The expressions of GFAP, inflammatory factors, pro-inflammatory transcription factors, and synaptic marker inoffspring brain were focused.
2. Materials and methods
2.1. Chemicals and animals
PFOS (potassium salt, purity �98%) was purchased from Fluka(Sigma–Aldrich, Switzerland). The vehicle was 0.5% Tween 80(Sigma–Aldrich, Switzerland) in deionized water. The PFOS wasdissolved in the vehicle and diluted to 0.025, 0.15, and 0.5 mg/ml(not adjusted for purity).
Eight-week-old SD rats were obtained from the Center forAnimal of Tongji Medical College, Huazhong University of Scienceand Technology (Wuhan, China), and housed individually in plasticcages in a room with an ambient temperature of 22 � 2 8C, a normal12/12-h cycle of light and dark, and a normal humidity of 50 � 10%.After acclimatization for 2 weeks, each female rat was paired with onemale rat at 06:00 p.m. Vaginal smear was performed to find sperm at08:00 a.m. in the next morning. The day when sperm was found wasdefined GD0 (gestation day 0).
A total of 40 pregnant rats were weighted and randomly dividedinto four groups (n = 10 each group), including control group (0.5%
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
Tween 80) and three dosage groups (0.1, 0.6 and 2.0 mg/kg bw/dayPFOS). Every 1–3 days, dams were weighed and the PFOS dosageswere adjusted by body weight. The test and vehicle controlsolutions were administered orally by gavage at a volume of 4 ml/kg bw once daily from GD2 to GD21. On GD21, dams weremonitored for delivery at hourly intervals. The day of birth wasdesigned postnatal day 0 (PND0). After delivery, five pupsrandomly selected from each litter were sacrificed by decapitationwithin 2–3 h after birth, and trunk blood, cortex, and hippocampuswere collected, and stored frozen at �80 8C. The remaining pupswere randomly redistributed to the nursing dams withintheir respective dosage groups, with litter size kept at 6 pups.On PND21, all offspring were sacrificed by decapitation, and trunkblood, cortex and hippocampus were collected, and stored frozenat �80 8C.
2.2. Determination of PFOS concentrations in hippocampus, cortex
and serum
Hippocampus samples, cortex samples and serum samples(pooled by three pups from litter on PND0, and individual offspringon PND21) for PFOS analysis were obtained from pups on PND0and PND21. Due to limited sample volume and size, serum andhippocampus from three PND0 pups were pooled. PFOS concen-trations were determined by liquid chromatography tandem–massspectrometry (LC–MS/MS) according to the method reported byLau et al. (Tan et al., 2008). The procedures of pretreatments ofhippocampus tissues, cortex tissues and serum samples were inaccordance to previous reports with some minor modifications(Kannan et al., 2001; Olivero-Verbel et al., 2006). Briefly, eachsample was extracted by 1 ml of 0.5 M tetrabutylammoniumhydrogen sulfate solution and 2 ml of 0.25 M sodium carbonatebuffer in a 15-ml polypropylene tube. After being thoroughlymixed, 5 ml of methyl tert-butyl ether (MTBE) was added to thesolution, and the mixture was shaken for 20 min. After beingseparated by centrifugation, 4 ml of the supernatant was removedfrom the solution, and the process was repeated twice. The solventwas allowed to evaporate under nitrogen before being recon-stituted in 1 ml of methanol. The sample was vortexed and passedthrough a 0.2 mm nylon filter into an autosampler vial. Analysiswas then performed using LC/MS/MS (Agilent 1100 LC-MSD-trap-XCT, Agilent 1100 Series LC/MS Ion-Trap, Agilent, Palo Alto, CA)with a sample volume of 5 ml. Separation was achieved on anInertsil ODS-3 column (2.1 mm � 150 mm, 5 mm), being carriedout using a mobile phase of 1.0 mM ammonium acetate/methanol(vol/vol) at a flow rate of 0.2 ml/min. The gradient profile was asfollows: linear increase from 40% to 75% methanol solution for 5–12 min, then hold at 75% for 3 min. PFOS was quantitativelyanalyzed by single mass mode using characteristic ions at m/z498.9 (Martin et al., 2004; Tseng et al., 2006).
2.3. Immunohistochemical analysis of GFAP
After dewaxing, brain sections were boiled (in an 850-Wmicrowave oven) for 15 min in citrate buffer. Endogenousperoxidase was inhibited by 1% hydrogen peroxide in puremethanol for 15 min. Sections were incubated with 10% normalpig serum in Tris-buffered saline to block nonspecific binding ofimmunoglobulins and then with the mouse monoclonal anti-bodies: glial fibrillary acidic protein (GFAP) (SANTA, 1:200)overnight at 4 8C, washed three times with PBS for 15 min, andincubated the horseradish peroxidase-conjugated goat anti-mouseIgG (1:500; Beyotime, Shanghai, China) for 1 h. The sections werethen washed four times and counterstained with Maier’shemalum. As negative controls, the primary antibodies wereomitted. After immunostaining, brain sections were examined
glial response in rat brain induced by prenatal PFOS exposure.

H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx 3
G Model
NEUTOX-1176; No. of Pages 10
under light microscopy. Immunoreactivity of GFAP was evaluatedin the cortex and hippocampus. The numbers of GFAP-positive ofeach rat brain section were counted in three nonoverlapping high-power fields (HPFs, 400� magnification) for each section, andsections of three rats per group were analyzed. The HPFs wereselected from cortex and hippocampus areas that had a maximumof positive cells. In each field studied, only positive cells with thenucleus at the focal plane were counted.
2.4. RNA extraction and cDNA synthesis
Hippocampus and cortex samples were collected on PND0 andPND21. Total RNA was isolated from the individual hippocampusand cortex with SV Total RNA Isolation System bought fromPromega (Madison, USA) according to the manufacture. The ratioof the optical densities from the RNA samples measured at 260 and280 nm was used to evaluate nucleic acid purity, and the total RNAconcentrations were determined by absorbance at 260 nm. Thequality of total RNA was further estimated with the integrity of 28Sand 18S RNA by 1% agarose gel electrophoresis. Reversetranscription was performed using the Fermentas RevertAidTM
First Strand cDNA Synthesis Kit (Ontario, Canada) according to themanufacture’s protocol. Briefly, 1 mg total RNA was mixed with1 ml random hexamer primer, added DEPC-treated water to 12 ml.The mixture was mixed gently, centrifuged briefly and incubate at65 8C for 5 min, chilled on ice, and then, added the followingcomponents: including 5� reaction buffer, RNase inhibitor, dNTPmix, RevertAidTM M-Mulv reverse transcriptase. The reversetranscription mixture was incubated at 25 8C for 5 min followedby at 42 8C for 60 min, and the reaction stopped by heating at 70 8Cfor 5 min. The cDNA was stored at �80 8C until analysis.
2.5. QPCR
QPCR was carried out using the SYBR TaqTM kit bought from ABIcompany (Warrington, UK). mRNA sequence for target genes wereobtained from the GeneBank database (http://www.ncbi.nlm.nih.-gov), and gene specific primers were designed by Shanghai SangonCompany (Shanghai, China). The identities and the correspondingprimer sequences used in the study were listed in Table 1. The QPCRreaction was performed on an Applied Biosystems model 7900HTFast Real-Time PCR System in triplicate using 25 ng cDNA andSybgreen Universal PCR Master Mix in a total volume of 25 ml. Theprogram consisted of an initial denaturation at 95 8C for 10 min,followed by 40 cycles: 95 8C for 15 s (denaturation) and 60 8C for1 min (annealing/extension). Each sample was run in triplicate forthe target and reference genes along with a single NRT (no reversetranscriptase) negative control in a 384 well plate format. Thereporter dye signal (SYBR green) was normalized using the passivereference dye, ROX, to eliminate volume-handling error. GAPDH,which was uniformly expressed among all samples (cycle thresholdstandard deviation less than 0.5), was used as an endogenousreference gene. Fold change of the target genes was calculated usingthe 2�
DDCT method (Livak and Schmittgen, 2001).
Table 1Primer sequence for QPCR in this study.
Gene name Forward primer Reverse pr
GFAP GCGGGAGTCGGCGAGTTA CAGGAAT
S-100b AGCACCCTAACTCACGACCGA CACTCACA
IL-1b GTGTTTCCCTCCCTGCCTCT CAATGCTG
TNF-a CTACTGAACTTCGGGGTGATCG TGGGCTAC
NF-kB CCGCCCCTTCTAAAACTCTTATG GGCTGCCT
CREB ACCATTGCCCCTGGAGTTG GCTTCCCT
AP-1 ACCCGGTGGGCAATCTGA GGGTCGG
GAPDH ACAGCAACAGGGTGGTGGAC TTTGAGGG
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
2.6. Western blot analysis
Hippocampus or cortex tissue (individual pups or offspring)was thawed on ice, washed with PBS and suspended at 4 8C in RIPAlysis buffer (Beyotime, Shanghai, China). Tissues was disrupted bysonication (3� for 10 s) and centrifuged for 30 min at 10,000 � g
(4 8C). Protein concentrations in the supernatant were determinedby Bradford Protein Assay Kit (Beyotime, Shanghai, China).Proteins were electrophoretically resolved on 12% SDS-polyacryl-amide gels (25 mg per lane), and then transferred onto nitrocellu-lose membranes (Bio-Rad, Hercules, CA, USA). Nonspecific bindingwas reduced by incubating the membrane in blocking buffer(Beyotime, Shanghai, China) for 1 h. Membranes were incubatedwith specific primary antibodies [anti-Syn1 (abcam, 1:1000); anti-Syp (Sigma, 1:500); anti-GFAP (SANTA, 1:400); anti-GAPDH(Promab, 1:1000); anti-S100b (abcam, 1:500)] overnight at 4 8C,washed three times with PBS for 15 min, and incubated with thehorseradish peroxidase-conjugated goat anti-rabbit IgG (1:1000;Beyotime, Shanghai, China) or the horseradish peroxidase-conjugated goat anti-mouse IgG (1:1000; Beyotime, Shanghai,China) for 1 h. The membranes were then washed four times anddeveloped with BeyoECL Plus (Beyotime, Shanghai, China). GAPDHwas used as the internal control.
2.7. Statistical analysis
All data were presented as mean � S.D., and statistical analysiswas performed with one-way ANOVA followed by LSD and Dunnett’sT3 test using SPSS 12.0. P-Value < 0.05 was deemed statisticallysignificant in all experiments.
3. Results
3.1. PFOS concentration in hippocampus, cortex and serum
The PFOS concentrations in the hippocampus, cortex and serumwere presented in Table 2. The hippocampus, cortex and serum ofpups were collected on PND0 and PND21. PFOS content of controlgroup was very low (not detected) on either PND0 or PND21. Withthe administrated-dose increasing, the PFOS concentration in-creased quickly in cortex, hippocampus and serum, and showed adose-dependent manner (P < 0.05). In addition, a time-relateddecrease of PFOS concentration was observed in the presentresults. The PFOS concentrations of offspring were higher at PND0than that at PND21 in the same group (P < 0.05).
3.2. Astrocytic activation in hippocampus and cortex
One of the most common characters of astrocytes is theirreaction to CNS damage with reactive gliosis, which is associatedwith morphological changes inside the cell and an increasedsynthesis of many proteins (Sofroniew, 2009). Classically, GFAP isused as a marker of astrocyte activation. Immunohistochemistryexperiments were conducted to evaluate the numbers of GFAP-
imer Product (bp) Accession no.
GGTGATGCGGTTT 188 NM_017009
TTATGGGCAGCAGA 168 NM_013191
CCTCGTGACCC 108 NM_031512
GGGCTTGTCA 134 NM_012675
GGATCACTTCAA 119 XM_342346
GTTCTTCATTAGACG 107 NM_031017
TGTAGTGGTGATGTG 150 NM_021835
TGCAGCGAACTT 252 NM_017008
glial response in rat brain induced by prenatal PFOS exposure.
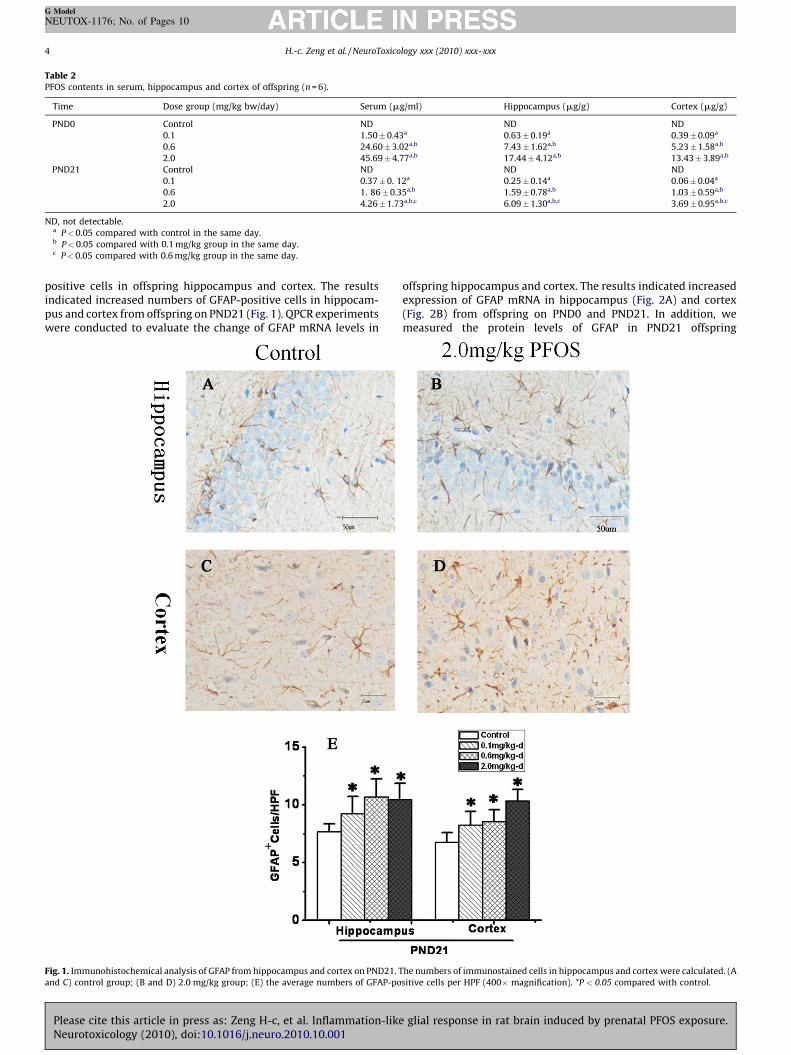
Table 2PFOS contents in serum, hippocampus and cortex of offspring (n = 6).
Time Dose group (mg/kg bw/day) Serum (mg/ml) Hippocampus (mg/g) Cortex (mg/g)
PND0 Control ND ND ND
0.1 1.50�0.43a 0.63�0.19a 0.39�0.09a
0.6 24.60�3.02a,b 7.43�1.62a,b 5.23�1.58a,b
2.0 45.69�4.77a,b 17.44�4.12a,b 13.43�3.89a,b
PND21 Control ND ND ND
0.1 0.37�0. 12a 0.25�0.14a 0.06�0.04a
0.6 1. 86�0.35a,b 1.59�0.78a,b 1.03�0.59a,b
2.0 4.26�1.73a,b,c 6.09�1.30a,b,c 3.69� 0.95a,b,c
ND, not detectable.a P<0.05 compared with control in the same day.b P<0.05 compared with 0.1 mg/kg group in the same day.c P<0.05 compared with 0.6 mg/kg group in the same day.
H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx4
G Model
NEUTOX-1176; No. of Pages 10
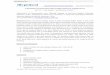
positive cells in offspring hippocampus and cortex. The resultsindicated increased numbers of GFAP-positive cells in hippocam-pus and cortex from offspring on PND21 (Fig. 1). QPCR experimentswere conducted to evaluate the change of GFAP mRNA levels in
Fig. 1. Immunohistochemical analysis of GFAP from hippocampus and cortex on PND21. T
and C) control group; (B and D) 2.0 mg/kg group; (E) the average numbers of GFAP-po
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
offspring hippocampus and cortex. The results indicated increasedexpression of GFAP mRNA in hippocampus (Fig. 2A) and cortex(Fig. 2B) from offspring on PND0 and PND21. In addition, wemeasured the protein levels of GFAP in PND21 offspring
he numbers of immunostained cells in hippocampus and cortex were calculated. (A
sitive cells per HPF (400� magnification). *P < 0.05 compared with control.
glial response in rat brain induced by prenatal PFOS exposure.
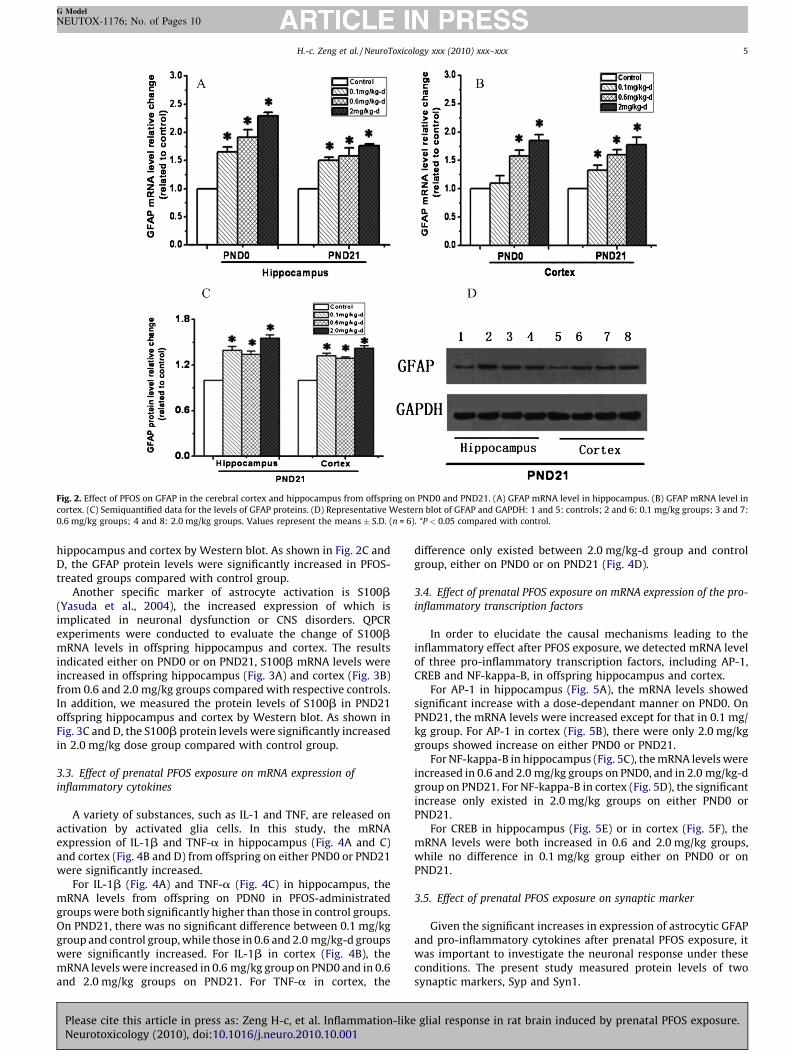
Fig. 2. Effect of PFOS on GFAP in the cerebral cortex and hippocampus from offspring on PND0 and PND21. (A) GFAP mRNA level in hippocampus. (B) GFAP mRNA level in
cortex. (C) Semiquantified data for the levels of GFAP proteins. (D) Representative Western blot of GFAP and GAPDH: 1 and 5: controls; 2 and 6: 0.1 mg/kg groups; 3 and 7:
0.6 mg/kg groups; 4 and 8: 2.0 mg/kg groups. Values represent the means � S.D. (n = 6). *P < 0.05 compared with control.
H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx 5
G Model
NEUTOX-1176; No. of Pages 10
hippocampus and cortex by Western blot. As shown in Fig. 2C andD, the GFAP protein levels were significantly increased in PFOS-treated groups compared with control group.
Another specific marker of astrocyte activation is S100b(Yasuda et al., 2004), the increased expression of which isimplicated in neuronal dysfunction or CNS disorders. QPCRexperiments were conducted to evaluate the change of S100bmRNA levels in offspring hippocampus and cortex. The resultsindicated either on PND0 or on PND21, S100b mRNA levels wereincreased in offspring hippocampus (Fig. 3A) and cortex (Fig. 3B)from 0.6 and 2.0 mg/kg groups compared with respective controls.In addition, we measured the protein levels of S100b in PND21offspring hippocampus and cortex by Western blot. As shown inFig. 3C and D, the S100b protein levels were significantly increasedin 2.0 mg/kg dose group compared with control group.
3.3. Effect of prenatal PFOS exposure on mRNA expression of
inflammatory cytokines
A variety of substances, such as IL-1 and TNF, are released onactivation by activated glia cells. In this study, the mRNAexpression of IL-1b and TNF-a in hippocampus (Fig. 4A and C)and cortex (Fig. 4B and D) from offspring on either PND0 or PND21were significantly increased.
For IL-1b (Fig. 4A) and TNF-a (Fig. 4C) in hippocampus, themRNA levels from offspring on PDN0 in PFOS-administratedgroups were both significantly higher than those in control groups.On PND21, there was no significant difference between 0.1 mg/kggroup and control group, while those in 0.6 and 2.0 mg/kg-d groupswere significantly increased. For IL-1b in cortex (Fig. 4B), themRNA levels were increased in 0.6 mg/kg group on PND0 and in 0.6and 2.0 mg/kg groups on PND21. For TNF-a in cortex, the
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
difference only existed between 2.0 mg/kg-d group and controlgroup, either on PND0 or on PND21 (Fig. 4D).
3.4. Effect of prenatal PFOS exposure on mRNA expression of the pro-
inflammatory transcription factors
In order to elucidate the causal mechanisms leading to theinflammatory effect after PFOS exposure, we detected mRNA levelof three pro-inflammatory transcription factors, including AP-1,CREB and NF-kappa-B, in offspring hippocampus and cortex.
For AP-1 in hippocampus (Fig. 5A), the mRNA levels showedsignificant increase with a dose-dependant manner on PND0. OnPND21, the mRNA levels were increased except for that in 0.1 mg/kg group. For AP-1 in cortex (Fig. 5B), there were only 2.0 mg/kggroups showed increase on either PND0 or PND21.
For NF-kappa-B in hippocampus (Fig. 5C), the mRNA levels wereincreased in 0.6 and 2.0 mg/kg groups on PND0, and in 2.0 mg/kg-dgroup on PND21. For NF-kappa-B in cortex (Fig. 5D), the significantincrease only existed in 2.0 mg/kg groups on either PND0 orPND21.
For CREB in hippocampus (Fig. 5E) or in cortex (Fig. 5F), themRNA levels were both increased in 0.6 and 2.0 mg/kg groups,while no difference in 0.1 mg/kg group either on PND0 or onPND21.
3.5. Effect of prenatal PFOS exposure on synaptic marker
Given the significant increases in expression of astrocytic GFAPand pro-inflammatory cytokines after prenatal PFOS exposure, itwas important to investigate the neuronal response under theseconditions. The present study measured protein levels of twosynaptic markers, Syp and Syn1.
glial response in rat brain induced by prenatal PFOS exposure.
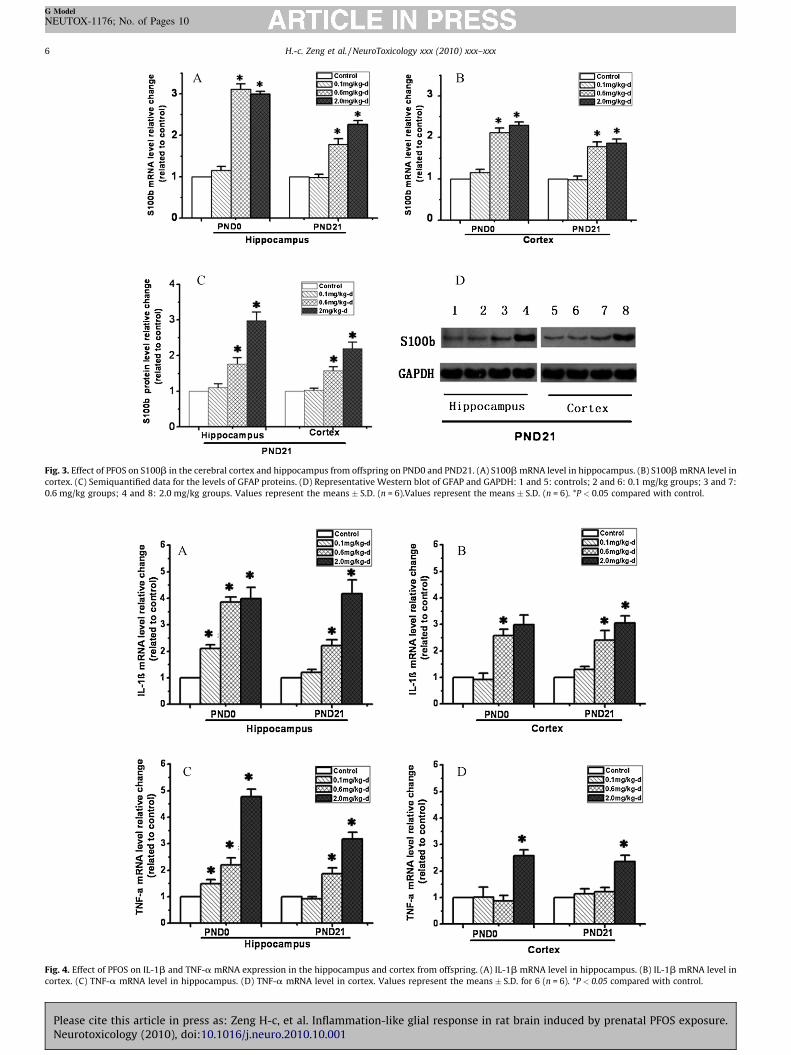
Fig. 3. Effect of PFOS on S100b in the cerebral cortex and hippocampus from offspring on PND0 and PND21. (A) S100b mRNA level in hippocampus. (B) S100b mRNA level in
cortex. (C) Semiquantified data for the levels of GFAP proteins. (D) Representative Western blot of GFAP and GAPDH: 1 and 5: controls; 2 and 6: 0.1 mg/kg groups; 3 and 7:
0.6 mg/kg groups; 4 and 8: 2.0 mg/kg groups. Values represent the means � S.D. (n = 6).Values represent the means � S.D. (n = 6). *P < 0.05 compared with control.
Fig. 4. Effect of PFOS on IL-1b and TNF-a mRNA expression in the hippocampus and cortex from offspring. (A) IL-1b mRNA level in hippocampus. (B) IL-1b mRNA level in
cortex. (C) TNF-a mRNA level in hippocampus. (D) TNF-a mRNA level in cortex. Values represent the means � S.D. for 6 (n = 6). *P < 0.05 compared with control.
H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx6
G Model
NEUTOX-1176; No. of Pages 10
Please cite this article in press as: Zeng H-c, et al. Inflammation-like glial response in rat brain induced by prenatal PFOS exposure.Neurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
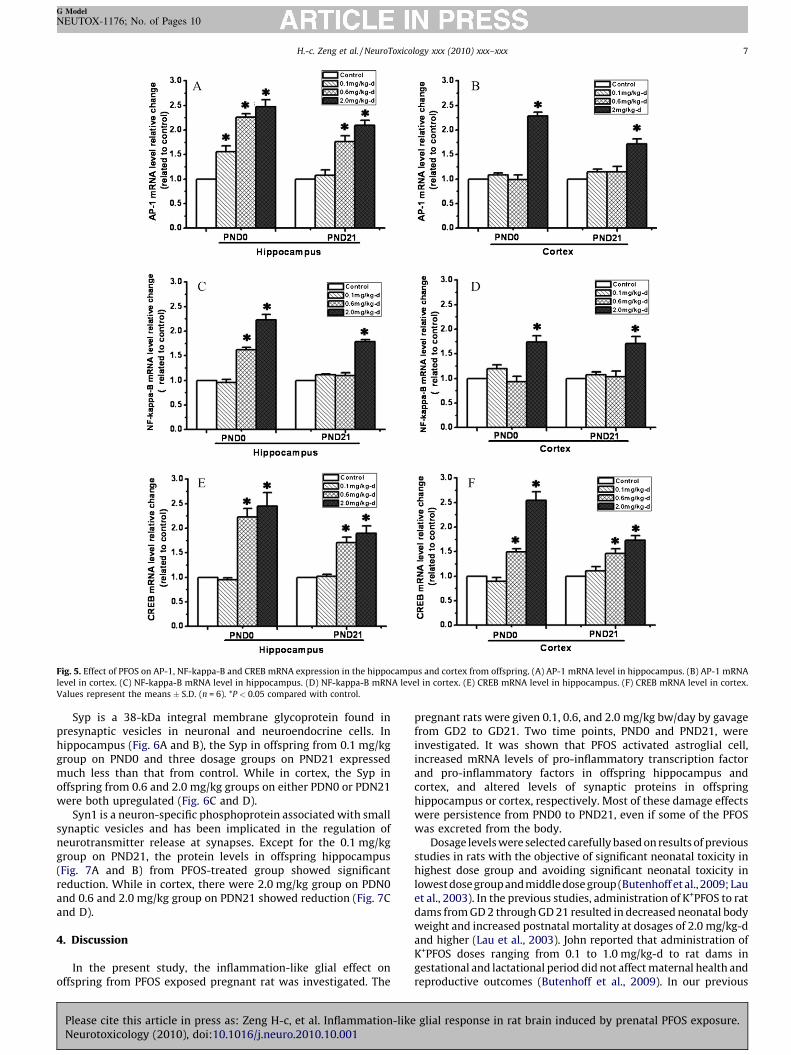
Fig. 5. Effect of PFOS on AP-1, NF-kappa-B and CREB mRNA expression in the hippocampus and cortex from offspring. (A) AP-1 mRNA level in hippocampus. (B) AP-1 mRNA
level in cortex. (C) NF-kappa-B mRNA level in hippocampus. (D) NF-kappa-B mRNA level in cortex. (E) CREB mRNA level in hippocampus. (F) CREB mRNA level in cortex.
Values represent the means � S.D. (n = 6). *P < 0.05 compared with control.
H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx 7
G Model
NEUTOX-1176; No. of Pages 10
Syp is a 38-kDa integral membrane glycoprotein found inpresynaptic vesicles in neuronal and neuroendocrine cells. Inhippocampus (Fig. 6A and B), the Syp in offspring from 0.1 mg/kggroup on PND0 and three dosage groups on PND21 expressedmuch less than that from control. While in cortex, the Syp inoffspring from 0.6 and 2.0 mg/kg groups on either PDN0 or PDN21were both upregulated (Fig. 6C and D).
Syn1 is a neuron-specific phosphoprotein associated with smallsynaptic vesicles and has been implicated in the regulation ofneurotransmitter release at synapses. Except for the 0.1 mg/kggroup on PND21, the protein levels in offspring hippocampus(Fig. 7A and B) from PFOS-treated group showed significantreduction. While in cortex, there were 2.0 mg/kg group on PDN0and 0.6 and 2.0 mg/kg group on PDN21 showed reduction (Fig. 7Cand D).
4. Discussion
In the present study, the inflammation-like glial effect onoffspring from PFOS exposed pregnant rat was investigated. The
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
pregnant rats were given 0.1, 0.6, and 2.0 mg/kg bw/day by gavagefrom GD2 to GD21. Two time points, PND0 and PND21, wereinvestigated. It was shown that PFOS activated astroglial cell,increased mRNA levels of pro-inflammatory transcription factorand pro-inflammatory factors in offspring hippocampus andcortex, and altered levels of synaptic proteins in offspringhippocampus or cortex, respectively. Most of these damage effectswere persistence from PND0 to PND21, even if some of the PFOSwas excreted from the body.
Dosage levels were selected carefully based on results of previousstudies in rats with the objective of significant neonatal toxicity inhighest dose group and avoiding significant neonatal toxicity inlowest dose group and middle dose group (Butenhoff et al., 2009; Lauet al., 2003). In the previous studies, administration of K+PFOS to ratdams from GD 2 through GD 21 resulted in decreased neonatal bodyweight and increased postnatal mortality at dosages of 2.0 mg/kg-dand higher (Lau et al., 2003). John reported that administration ofK+PFOS doses ranging from 0.1 to 1.0 mg/kg-d to rat dams ingestational and lactational period did not affect maternal health andreproductive outcomes (Butenhoff et al., 2009). In our previous
glial response in rat brain induced by prenatal PFOS exposure.
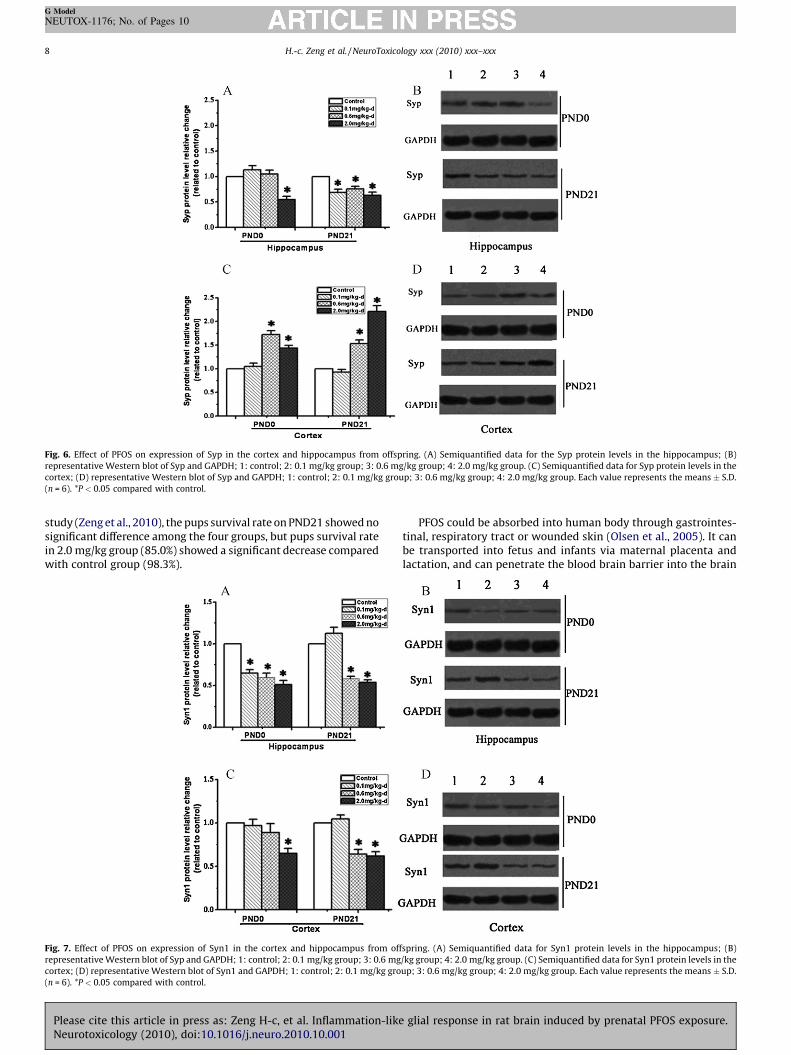
Fig. 6. Effect of PFOS on expression of Syp in the cortex and hippocampus from offspring. (A) Semiquantified data for the Syp protein levels in the hippocampus; (B)
representative Western blot of Syp and GAPDH; 1: control; 2: 0.1 mg/kg group; 3: 0.6 mg/kg group; 4: 2.0 mg/kg group. (C) Semiquantified data for Syp protein levels in the
cortex; (D) representative Western blot of Syp and GAPDH; 1: control; 2: 0.1 mg/kg group; 3: 0.6 mg/kg group; 4: 2.0 mg/kg group. Each value represents the means � S.D.
(n = 6). *P < 0.05 compared with control.
H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx8
G Model
NEUTOX-1176; No. of Pages 10
study (Zeng et al., 2010), the pups survival rate on PND21 showed nosignificant difference among the four groups, but pups survival ratein 2.0 mg/kg group (85.0%) showed a significant decrease comparedwith control group (98.3%).
Fig. 7. Effect of PFOS on expression of Syn1 in the cortex and hippocampus from off
representative Western blot of Syp and GAPDH; 1: control; 2: 0.1 mg/kg group; 3: 0.6 mg
cortex; (D) representative Western blot of Syn1 and GAPDH; 1: control; 2: 0.1 mg/kg gro
(n = 6). *P < 0.05 compared with control.
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
PFOS could be absorbed into human body through gastrointes-tinal, respiratory tract or wounded skin (Olsen et al., 2005). It canbe transported into fetus and infants via maternal placenta andlactation, and can penetrate the blood brain barrier into the brain
spring. (A) Semiquantified data for Syn1 protein levels in the hippocampus; (B)
/kg group; 4: 2.0 mg/kg group. (C) Semiquantified data for Syn1 protein levels in the
up; 3: 0.6 mg/kg group; 4: 2.0 mg/kg group. Each value represents the means � S.D.
glial response in rat brain induced by prenatal PFOS exposure.

H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx 9
G Model
NEUTOX-1176; No. of Pages 10
and produce neurotoxic effect (Apelberg et al., 2007; Olsen et al.,2008). It was shown that PFOS administration could cause long-term accumulation of PFOS in the brain and prenatal PFOSadministration caused more accumulation in pup brains than dambrains (Chang et al., 2009). The data in this study showed that PFOSexposure from GD2 to GD21 also caused a significant accumulationof PFOS in the offspring hippocampus, cortex and serum. Moreover,the results showed the positive correlation between PFOSaccumulation in the hippocampus, cortex, or serum and adminis-tration-dose of PFOS on PND0 or PND21. Meanwhile, theconcentration of PFOS showed significant decrease with a time-dependent manner in offspring serum, hippocampus and cortex,respectively. The concentrations of PFOS in offspring tissues inPND0 were higher in PND21 offspring tissue. This discrepancycould be explained by the metabolism of offspring, which the half-life of PFOS for neonates is about 14 days (Wang et al., 2010). Inaddition, the volume expansion may contribute to the lowering ofbrain PFOS concentrations seen postnatally.
The most important function of astroglial cells is maintainingthe homeostatic environment for proper functioning of neurons(Sofroniew and Vinters, 2009). During pathological conditions,including toxicant insults, a universal cellular reaction to damageof the CNS is hypertrophy of astrocytes (Qazi et al., 2009; Sofroniewand Vinters, 2009). The hallmark of this response, often termed‘reactive gliosis’, is the enhanced expression of the majorintermediate filament protein of astrocytes, glial fibrillary acidicprotein (GFAP) (Sofroniew and Vinters, 2009). The latter observa-tion suggests that increased synthesis of GFAP would occur inresponse to diverse neurotoxic insults (O’Callaghan et al., 1995). Inaddition, overexpression of S-100b protein in response to neuronaldamage has been observed (Griffin et al., 1998; Yasuda et al., 2004).In the present study concerning the toxicity induced by PFOS inimmature rat brain, we found that activation of astroglial cellsoccurred as evidence by the increased expression of GFAP and S-100b. S-100b secreted from astrocytes plays both beneficial anddetrimental functions (Donato, 2001). At low physiologicalconcentration, it acts as a neurotrophic factor to promote thebrain development and to maintain the cellular homeostasis. Incontrast, overproduction of S-100b by activated glia can lead toexacerbation of neuroinflammation and neuronal dysfunction(Koppal et al., 2001; Li et al., 2000). Therefore, S-100b acts as abiomarker for brain damage, implicate glial activation in acute andchronic CNS disorders (Van Eldik and Wainwright, 2003).
The major findings of the present study included demonstrationof the occurrence of increased pro-inflammatory factors and pro-inflammatory transcription factor production, and altered levels ofsynaptic markers companying with astrocytic activation in PFOS-treated offspring brain. The observed changes were mild in nature,suggesting that the chronic neuroinflammatory states developedslowly. Although recently studies have demonstrated that PFOScan cause the inflammation in liver and the hypertrophy ofastrocytes in brain (Cui et al., 2009), and the enhanced theinflammatory responses of macrophages to lipopolysaccharide(LPS) (Qazi et al., 2009), until now there is no evidence that PFOScauses pro-inflammatory effects in brain in vivo.
Glial cells are the major producers of inflammatory mediators,and cytotoxic activation of glial cells is linked to severalneurodegenerative diseases by producing pro-inflammation fac-tors such as IL-1b and TNF-a (von Bernhardi, 2007). In thedeveloping brain and following acute glial activation in response toinjury, S-100b acts as a neurotrophic factor and neuronal survivalprotein. In contrast, overproduction of S100B by activated glia canlead to exacerbation of neuroinflammation and neuronal dysfunc-tion. In the current study, the overproduced S-100b by activatedglia may act as a pro-inflammatory trigger, further contribution toneuronal dysfunction as evidence by altering the expression of
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
synaptic markers (Van Eldik and Wainwright, 2003). In addition, itwas observed that TNF-a and IL-b expression also increased inoffspring hippocampus and cortex, which manifested that PFOSproduced the inflammation in brain. However, it is also possible thatactivation of immune cells in the periphery induced by PFOS leads toleukocyte infiltration and subsequent activation of microglia andastroglia. This status cannot be excluded, because the blood–brainbarrier in embryonal and neonatal rats is quite immature and porous,and is not completed until PND 24 (Chang et al., 2009). Immaturity ofthe BBB lacks the protective effect and enables blood-derivedmononuclear phagocytes to easily penetrate the nervous tissue.
Microglia are a type of glial cells that are the quiescentmacrophages of the brain and spinal cord, which represent a sourceof many cytokines, and thus act as the first and main form of activeimmune defense in the central nervous system (CNS) (Graeber andStreit, 2009). IL-1b is one of the most potent inducers of reactiveastrogliosis, which produce both by activated microglial cells andby activated astroglial cells (Giulian et al., 1986; Herx and Yong,2001). IL-1b is a specific inflammatory factor involved inintercellular communication between microglia and astroglia ininjured brain and plays a vial role in the regulation of inflammatoryprocess (Kim et al., 2006). Thus, the enhanced expression of IL-1bby PFOS exposure involved the induction of mechanisms leading tothe inflammatory cascade and indicated the potential pro-inflammatory action. In addition, the expression level of TNF-a,which was another pro-inflammatory factor, increased in brain.Compared to the cortex, the GFAP and the cytokines appeared to beaffected to a greater extent in the hippocampus, which was amicroglia-rich structure. Thus, these data suggested that microgliamight be activated by PFOS, Syn1 is a synaptic vesicle-associatedprotein that acts as an anchoring protein by linking the vesiclephospholipid layer and the cytoskeleton in neuronal presynapticterminals (Akbergenova and Bykhovskaia, 2007; Santos et al.,2009). It plays a vial role in neurotransmitter release. Syp is a majorintegral membrane protein of small synaptic vesicles (Greengardet al., 1993), which is required for vesicle fusion and release ofneurotransmitter to occur. An imbalance in the Syp level mightaffect the long-term development of the brain in ways that couldaffect the process of LTP (long-term potentiation). New brainsynapse forms when a postsynaptic structure, the dendritic spine,interacts with a presynaptic terminal. Brain synapses and dendriticspines, membrane-rich structures, are depleted in mental disease(Wurtman et al., 2009). In the present study, PFOS exposure causeda significant decrease of Syn1, especially in hippocampus, and wellcorrelated with the mRNA level of pro-inflammatory factors.Protein level of Syp also was significantly decreased in hippocam-pus. However, the expression of Syp was significantly increased incortex, of which reason was not clear. The unbalanced expressionof pre-synaptic markers might inhibit synaptogenesis in the brain(Liao et al., 2008). The inhibition of synapses induced by theprenatal PFOS exposure was supposed to relate to the euro-behavioural defects. In adult mice which were exposed to PFOS inearly life, the susceptibility of the cholinergic system was altered(Johansson et al., 2008), whereas the PFOS exposure in adult miceinduced only slight behavioural effects (Fuentes et al., 2007).
Action of AP-1, CREB and NF-kB is considered to be a part of thegeneral of a number of pro-inflammatory gene expressions inresponse to various extracellular stimuli, and play an importantrole in the expression of pro-inflammatory mediators (Guha et al.,2000; Lee et al., 2005). However, there are little information aboutthe effects of PFOS on the three pro-inflammatory transcriptionfactors mRNA levels in brain. In the present study, we observed anincreased mRNA level of these pro-inflammatory transcriptionfactors, which were well correlated with the pro-inflammatoryfactors. These results indicated that PFOS might alter thesetranscription factor expression levels, which might be responsible
glial response in rat brain induced by prenatal PFOS exposure.

H.-c. Zeng et al. / NeuroToxicology xxx (2010) xxx–xxx10
G Model
NEUTOX-1176; No. of Pages 10
for PFOS induced overexpression of pro-inflammatory factor inhippocampus and cortex induced by PFOS.
In conclusion, this study shows that in uterus exposure to PFOScauses a chronic glial activation with coexisting inflammation andneurodegenerative features in the rat developing brain. Theenhanced pro-inflammatory transcription factors may contributeto the effect of inflammation-like glial response.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgements
This project was supported by the National Natural ScienceFoundation of China (81030051, 20807017), the R&D Special Fundfor Public Welfare Industry (Environment) (200909102), the KeyProjects in the National Science and Technology Pillar Program(grant number 2007BAC16B07), and the Fundamental ResearchFunds for the Central Universities (2010MS091).
References
Akbergenova Y, Bykhovskaia M. Synapsin maintains the reserve vesicle pool and spatialsegregation of the recycling pool in Drosophila presynaptic boutons. Brain Res2007;1178:52–64.
Apelberg BJ, Goldman LR, Calafat AM, Herbstman JB, Kuklenyik Z, Heidler J, et al.Determinants of fetal exposure to polyfluoroalkyl compounds in Baltimore, Mary-land. Environ Sci Technol 2007;41(11):3891–7.
Austin ME, Kasturi BS, Barber M, Kannan K, MohanKumar PS, MohanKumar SM.Neuroendocrine effects of perfluorooctane sulfonate in rats. Environ Health Per-spect 2003;111(12):1485–9.
Butenhoff JL, Ehresman DJ, Chang SC, Parker GA, Stump DG. Gestational and lactationalexposure to potassium perfluorooctanesulfonate (K + PFOS) in rats: developmentalneurotoxicity. Reprod Toxicol (Elmsford NY) 2009;27(3–4):319–30.
Chang SC, Ehresman DJ, Bjork JA, Wallace KB, Parker GA, Stump DG, et al. Gestationaland lactational exposure to potassium perfluorooctanesulfonate (K + PFOS) in rats:toxicokinetics, thyroid hormone status, and related gene expression. ReprodToxicol (Elmsford NY) 2009;27(3–4):387–99.
Cui L, Zhou QF, Liao CY, Fu JJ, Jiang GB. Studies on the toxicological effects of PFOA andPFOS on rats using histological observation and chemical analysis. Arch EnvironContam Toxicol 2009;56(2):338–49.
Donato R. S100: a multigenic family of calcium-modulated proteins of the EF-handtype with intracellular and extracellular functional roles. Int J Biochem Cell Biol2001;33(7):637–68.
Dong GH, Zhang YH, Zheng L, Liang ZF, Jin YH, He QC. Subchronic effects of perfluor-ooctanesulfonate exposure on inflammation in adult male C57BL/6 mice. Environ-mental toxicology [PMID: 20737580].
Fuentes S, Vicens P, Colomina MT, Domingo JL. Behavioral effects in adult mice exposedto perfluorooctane sulfonate (PFOS). Toxicology 2007;242(1–3):123–9.
Giesy JP, Kannan K. Perfluorochemical surfactants in the environment. Environ SciTechnol 2002;36(7):146A–52A.
Giulian D, Baker TJ, Shih LC, Lachman LB. Interleukin 1 of the central nervous system isproduced by ameboid microglia. J Exp Med 1986;164(2):594–604.
Graeber MB, Streit WJ. Microglia: biology and pathology. Acta Neuropathol2009;119(1):89–105.
Greengard P, Valtorta F, Czernik AJ, Benfenati F. Synaptic vesicle phosphoproteins andregulation of synaptic function. Science 1993;259(5096):780–5.
Griffin WS, Sheng JG, Royston MC, Gentleman SM, McKenzie JE, Graham DI, et al. Glial-neuronal interactions in Alzheimer’s disease: the potential role of a ‘cytokine cycle’in disease progression.. Brain Pathol (Zurich Switzerland) 1998;8(1):65–72.
Guha M, Bai W, Nadler JL, Natarajan R. Molecular mechanisms of tumor necrosis factoralpha gene expression in monocytic cells via hyperglycemia-induced oxidantstress-dependent and -independent pathways. J Biol Chem 2000;275(23):17728–39.
Herx LM, Yong VW. Interleukin-1 beta is required for the early evolution of reactiveastrogliosis following CNS lesion. J Neuropathol Exp Neurol 2001;60(10):961–71.
Johansson N, Fredriksson A, Eriksson P. Neonatal exposure to perfluorooctane sulfonate(PFOS) and perfluorooctanoic acid (PFOA) causes neurobehavioural defects in adultmice. Neurotoxicology 2008;29(1):160–9.
Kannan K, Franson JC, Bowerman WW, Hansen KJ, Jones PD, Giesy JP. Perfluorooctanesulfonate in fish-eating water birds including bald eagles and albatrosses. EnvironSci Technol 2001;35(15):3065–70.
Karrman A, Domingo JL, Llebaria X, Nadal M, Bigas E, van Bavel B, et al. Biomonitoringperfluorinated compounds in Catalonia, Spain: concentrations and trends inhuman liver and milk samples. Environ Sci Pollut Res Int 2009;17(3):750–8.
Kim YJ, Hwang SY, Oh ES, Oh S, Han IO. IL-1beta, an immediate early protein secreted byactivated microglia, induces iNOS/NO in C6 astrocytoma cells through p38 MAPKand NF-kappaB pathways. J Neurosci Res 2006;84(5):1037–46.
Please cite this article in press as: Zeng H-c, et al. Inflammation-likeNeurotoxicology (2010), doi:10.1016/j.neuro.2010.10.001
Koppal T, Lam AG, Guo L, Van Eldik LJ. S100B proteins that lack one or both cysteineresidues can induce inflammatory responses in astrocytes and microglia. Neuro-chem Int 2001;39(5–6):401–7.
Lau C, Thibodeaux JR, Hanson RG, Rogers JM, Grey BE, Stanton ME, et al. Exposure toperfluorooctane sulfonate during pregnancy in rat and mouse. II: Postnatal evalu-ation. Toxicol Sci 2003;74(2):382–92.
Lee YW, Hirani AA, Kyprianou N, Toborek M. Human immunodeficiency virus-1 Tatprotein up-regulates interleukin-6 and interleukin-8 expression in human breastcancer cells. Inflamm Res 2005;54(9):380–9.
Li Y, Barger SW, Liu L, Mrak RE, Griffin WS. S100beta induction of the proinflammatorycytokine interleukin-6 in neurons. J Neurochem 2000;74(1):143–50.
Liao CY, Li XY, Wu B, Duan S, Jiang GB. Acute enhancement of synaptic transmission andchronic inhibition of synaptogenesis induced by perfluorooctane sulfonatethrough mediation of voltage-dependent calcium channel. Environ Sci Technol2008;42(14):5335–41.
Liu L, Jin YH, Wang L, Yu HY, Liu W, Yu QL, et al. Effects of perfluorooctane sulfonate onlearning and memory of rat pups. Zhonghua Yu Fang Yi Xue Za Zhi 2009;43(7):622–7.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-timequantitative PCR and the 2(�Delta Delta C(T)) Method. Methods (San Diego CA)2001;25(4):402–8.
Luebker DJ, Case MT, York RG, Moore JA, Hansen KJ, Butenhoff JL. Two-generationreproduction and cross-foster studies of perfluorooctanesulfonate (PFOS) in rats.Toxicology 2005;215(1–2):126–48.
Martin JW, Kannan K, Berger U, de Voogt P, Field J, Franklin J, et al. Analytical challengeshamper perfluoroalkyl research. Environ Sci Technol 2004;38(13):248A–55A.
O’Callaghan JP, Jensen KF, Miller DB. Quantitative aspects of drug and toxicant-inducedastrogliosis. Neurochem Int 1995;26(2):115–24.
Olivero-Verbel J, Tao L, Johnson-Restrepo B, Guette-Fernandez J, Baldiris-Avila R,O’Byrne-Hoyos I, et al. Perfluorooctanesulfonate and related fluorochemicals inbiological samples from the north coast of Colombia. Environ Pollut 2006;142(2):367–72.
Olsen GW, Huang HY, Helzlsouer KJ, Hansen KJ, Butenhoff JL, Mandel JH. Historicalcomparison of perfluorooctanesulfonate, perfluorooctanoate, and other fluoro-chemicals in human blood. Environ Health Perspect 2005;113(5):539–45.
Olsen GW, Mair DC, Church TR, Ellefson ME, Reagen WK, Boyd TM, et al. Decline inperfluorooctanesulfonate and other polyfluoroalkyl chemicals in American RedCross adult blood donors, 2000–2006. Environ Sci Technol 2008;42(13):4989–95.
Qazi MR, Bogdanska J, Butenhoff JL, Nelson BD, DePierre JW, Abedi-Valugerdi M. High-dose, short-term exposure of mice to perfluorooctanesulfonate (PFOS) or perfluor-ooctanoate (PFOA) affects the number of circulating neutrophils differently, butenhances the inflammatory responses of macrophages to lipopolysaccharide (LPS)in a similar fashion. Toxicology 2009;262(3):207–14.
Renner R. Growing concern over perfluorinated chemicals. Environ Sci Technol2001;35(7):154A–60A.
Santos MS, Li H, Voglmaier SM. Synaptic vesicle protein trafficking at the glutamatesynapse. Neuroscience 2009;158(1):189–203.
Sofroniew MV. Molecular dissection of reactive astrogliosis and glial scar formation.Trends Neurosci 2009;32(12):638–47.
Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta Neuropathol2009;119(1):7–35.
Sohlenius AK, Andersson K, Bergstrand A, Spydevold O, De Pierre JW. Effects ofperfluorooctanoic acid – a potent peroxisome proliferator in rat – on Morrishepatoma 7800C1 cells, a rat cell line. Biochim Biophys Acta 1994;1213(1):63–74.
Tan YM, Clewell HJ III, Andersen ME. Time dependencies in perfluorooctylacidsdisposition in rat and monkeys: a kinetic analysis. Toxicol Lett 2008;177(1):38–47.
Tao L, Kannan K, Wong CM, Arcaro KF, Butenhoff JL. Perfluorinated compounds in humanmilk from Massachusetts, U.S.A.. Environ Sci Technol 2008a;42(8):3096–101.
Tao L, Ma J, Kunisue T, Libelo EL, Tanabe S, Kannan K. Perfluorinated compounds inhuman breast milk from several Asian countries, and in infant formula and dairymilk from the United States. Environ Sci Technol 2008b;42(22):8597–602.
Tseng CL, Liu LL, Chen CM, Ding WH. Analysis of perfluorooctanesulfonate and relatedfluorochemicals in water and biological tissue samples by liquid chromatography-ion trap mass spectrometry. J Chromatogr A 2006;1105(1–2):119–26.
Van Eldik LJ, Wainwright MS. The Janus face of glial-derived S100B: beneficial anddetrimental functions in the brain. Restor Neurol Neurosci 2003;21(3–4):97–108.
von Bernhardi R. Glial cell dysregulation: a new perspective on Alzheimer disease.Neurotox Res 2007;12(4):215–32.
von Ehrenstein OS, Fenton SE, Kato K, Kuklenyik Z, Calafat AM, Hines EP. Polyfluoroalkylchemicals in the serum and milk of breastfeeding women. Reprod Toxicol (Elms-ford NY) 2009;27(3–4):239–45.
Wang F, Liu W, Jin Y, Dai J, Yu W, Liu X, et al. Transcriptional effects of prenatal andneonatal exposure to PFOS in developing rat brain. Environ Sci Technol 2010;44(5):1847–53.
Whitney NP, Eidem TM, Peng H, Huang Y, Zheng JC. Inflammation mediates varyingeffects in neurogenesis: relevance to the pathogenesis of brain injury and neuro-degenerative disorders. J Neurochem 2009;108(6):1343–59.
Wurtman RJ, Cansev M, Sakamoto T, Ulus IH. Use of phosphatide precursors to promotesynaptogenesis. Annu Rev Nutr 2009;29:59–87.
Yasuda Y, Tateishi N, Shimoda T, Satoh S, Ogitani E, Fujita S. Relationship betweenS100beta and GFAP expression in astrocytes during infarction and glial scarformation after mild transient ischemia. Brain Res 2004;1021(1):20–31.
Zeng HC, Li YY, Zhang L, Wang YJ, Chen J, Xia W, et al. Prenatal exposure to perfluor-ooctanesulfonate in rat resulted in long-lasting changes of expression of synapsinsand synaptophysin. Synapse (New York, NY) 2010, [PMID: 20687110].
glial response in rat brain induced by prenatal PFOS exposure.