Embed Size (px)
Citation preview

Inference of Complex Genealogical Histories In Populations and
Application in Mapping Complex Traits
Yufeng WuDept. of Computer Science and Engineering
University of Connecticut
DIMACS 2008

Genealogy: Evolutionary History of Genomic Sequences
• Tells how sequences in a population are related
• Helps to explain diseases: disease mutations occur on branches and all descendents carry the mutations
• Genealogy: unknown. Only have SNP haplotypes (binary sequences).
• Problem: Inference of genealogy for “unrelated” haplotypes
• Not easy: partly due to recombination
Sequences in current population
Diseased (case)
Healthy (control)
Disease mutation
2

3
Recombination
• One of the principle genetic forces shaping sequence variations within species
• Two equal length sequences generate a third new equal length sequence in genealogy• Spatial order is important: different parts of genome inherit
from different ancestors.
110001111111001
000110000001111
Prefix
Suffix
Breakpoint
1100 00000001111

Ancestral Recombination Graph (ARG)
10 01 00
S1 = 00S2 = 01S3 = 10S4 = 10
Mutations
S1 = 00S2 = 01S3 = 10S4 = 11
10 01 0011
Recombination
Assumption:
At most one mutation per site
1 0 0 1
1 1
00
10
5
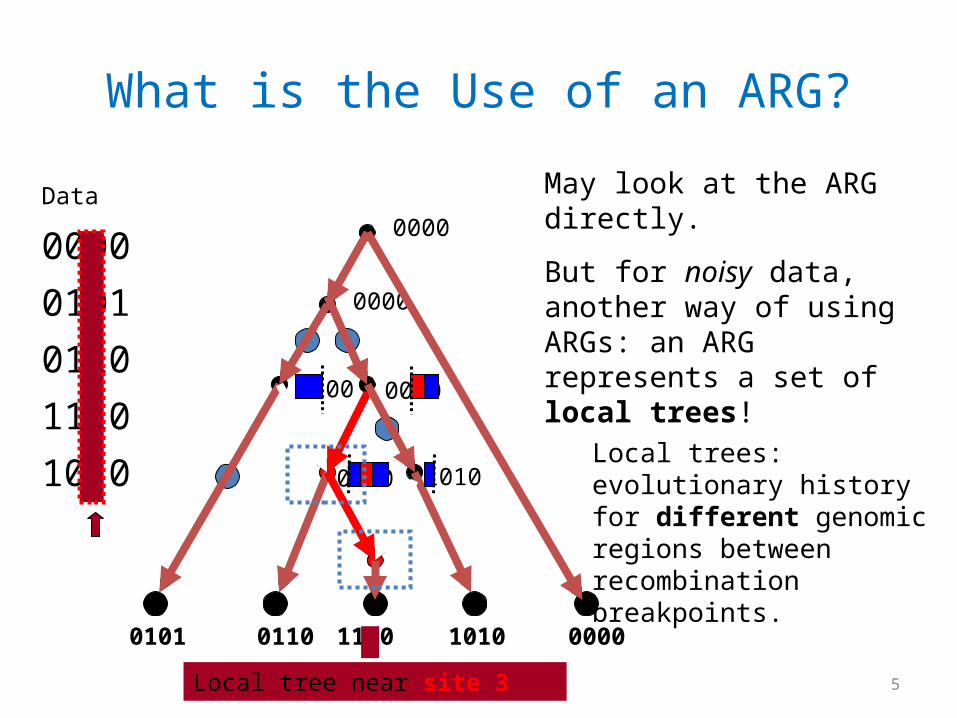
What is the Use of an ARG?
0101 1010 00000110
0100
0000
0000
0010
Local trees: evolutionary history for different genomic regions between recombination breakpoints.
0110 1010
1110
Data
0000
0101
0110
1110
1010
May look at the ARG directly.
But for noisy data, another way of using ARGs: an ARG represents a set of local trees!
Local tree near site 3

6
At which Local Tree Did Disease Mutations Occur?
• Clear separation of cases/controls: not expected for complex diseases
Case Control
Possible Disease mutation

7
How to infer ARGs?
• But we do not know the true ARG! • Goal: infer ARGs from haplotypes
• First practical ARG association mapping method (Minichiello and Durbin, 2006)– Use plausible ARGs: heuristic– Less complex disease model: implicitly assume
one disease mutation with major effects.
• My results (Wu, RECOMB 2007)– Generate ARGs with a provable property, and
works on a well-defined complex disease model– Focus on parsimonious history

8
Simulation Results (Wu, 2007)
Comparison: TMARG (minARGs), TMARG (near minARGs), LATAG (Z. P.), MARGARITA (M. D.).
TMARG (my program) and MARGRITA are much faster than LATAG.
50 ARGs per data
0
0.05
0.1
0.15
0.2
0.25
minARG Near minARG LATAG MARGARITA
• TMARG/MARGARITA: sample ARGs, decompose to local trees and look for association signals.
• LATAG: infer local trees at focal points.
• Average mapping error for 50 simulated datasets from Zollner and Pritchard

Preliminary Results: GAW16 Data
Caution: more investigation needed.
GAW16 data from the North American Rheumatoid Arthritis Consortium (NARAC), 868 cases and 1194 controls. Chromosome one: 40929 SNPs.
Running TMARG on large-scale data
• Break into non-overlapping windows
• Run fastPHASE (Scheet and Stephens 06) to obtain haplotypes
• Run TMARG with Chi-square mode
?
SNP rs2476601 reported in Begovich et al., 2004 and Carlton et al., 2005

10
A Related Problem
Inference of Local Tree Topologies Directly
(Wu, 2008, Submitted)

Inference of Local Tree Topologies
11
• Recall ARG represents a set of local trees.
• Question: given SNP haplotypes, infer local tree topologies (one tree for each SNP site, ignore branch length)
– Hein (1990, 1993)
• Song and Hein (2003,2005): enumerate all possible tree topologies at each site
– Parsimony-based

Local Tree Topologies
12
• Key technical difficulty: enumerate all tree topologies– Brute-force enumeration of local tree topologies: not
feasible when number of sequences > 9
• Trivial solution: create a tree for a SNP containing the single split induced by the SNP.– Always correct (assume one mutation per site)– But not very informative: need more refined trees!
A: 0B: 0C: 1D: 0E: 1F: 0G: 1H: 0
C
E
G
AB
DF
H

How to do better? Neighboring Local Trees are Similar!
• Nearby SNP sites provide hints!– Near-by local trees are often topologically similar– Recombination often only alters small parts of the
trees
• Key idea: reconstruct local trees by combining information from multiple nearby SNPs
13

RENT: REfining Neighboring Trees
• Maintain for each SNP site a (possibly non-binary) tree topology– Initialize to a tree containing the split induced by
the SNP
• Gradually refining trees by adding new splits to the trees– Splits found by a set of rules (later)– Splits added early may be more reliable
• Stop when binary trees or enough information is recovered
14

15
0 0 0 1 0 0 0 0 1 1 0 1 0 1 1
1 2 3
abcde
M
A Little Background: Compatibility
• Two sites (columns) p, q are incompatible if columns p,q contains all four ordered pairs (gametes): 00, 01, 10, 11. Otherwise, p and q are compatible.• Easily extended to splits.• A split s is incompatible with tree T if s is incompatible with
any one split in T. Two trees are compatible if their splits are pairwise compatible.
Sites 1 and 2 are compatible, but 1 and 3 are incompatible.

Fully-Compatible Region: Simple Case
• A region of consecutive SNP sites where these SNPs are pairwise compatible.– May indicate no topology-altering recombination
occurred within the region
• Rule: for site s, add any such split to tree at s.– Compatibility: very strong property and unlikely arise
due to chance.
16

Split Propagation: More General Rule
• Three consecutive sites 1,2 and 3. Sites 1 and 2 are incompatible. Does site 3 matter for tree at site 1?– Trees at site 1 and 2 are different.– Suppose site 3 is compatible with sites 1 and 2. Then?– Site 3 may indicate a shared subtree in both trees at sites 1 and 2.
• Rule: a split propagates to both directions until reaching a incompatible tree.
17

One Subtree-Prune-Regraft (SPR) Event
• Recombination: simulated by SPR.– The rest of two trees (without pruned subtrees) remain the same
• Rule: find compatible subtree Ts in neighboring trees T1 and T2, s.t. the rest of T1 and T2 (Ts removed) are compatible. Then joint refine T1- Ts and T2- Ts before adding back Ts.
Subtree to prune
18
More complex rules possible.
?
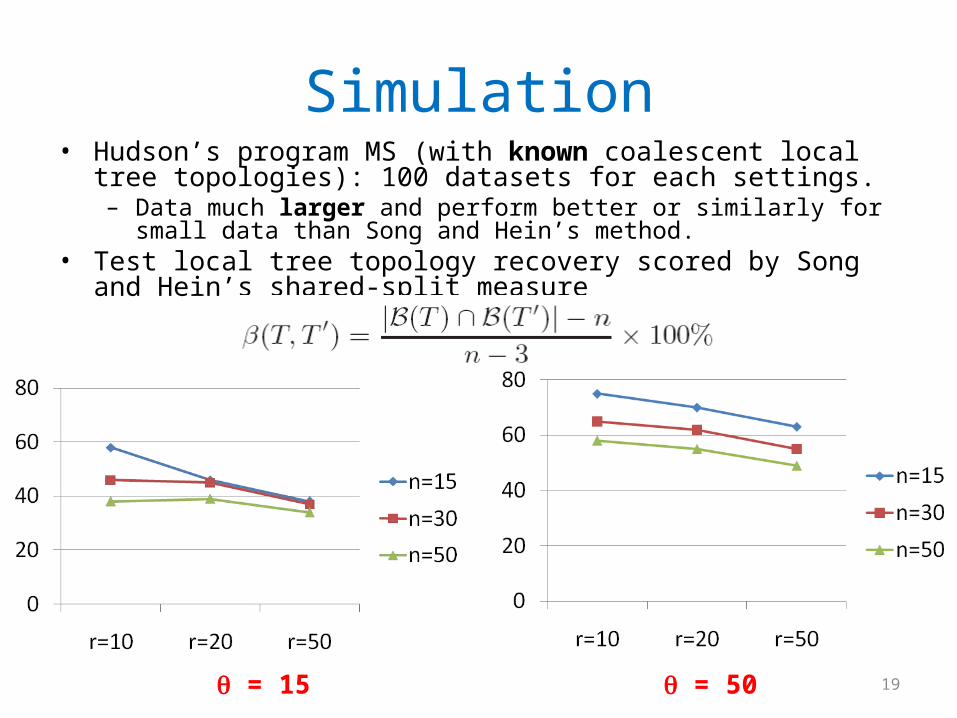
Simulation• Hudson’s program MS (with known coalescent local tree topologies):
100 datasets for each settings.– Data much larger and perform better or similarly for small data than Song
and Hein’s method.• Test local tree topology recovery scored by Song and Hein’s shared-
split measure
= 15 = 50 19

20
Acknowledgement
• More information available at: http://www.engr.uconn.edu/~ywu
• I want to thank– Dan Gusfield– Yun S. Song– Charles Langley– Dan Brown– And National Science Foundation and UConn
Research Foundation





























