Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 276, No. 1, January, pp. 259-264,199O
Induction of the Enzyme Aldose Reductase in a Lens Epithelial Cell Line from a Transgenic Mouse
Paul Russell,’ Takahiko Yamada, and Lorenzo 0. Merola National Eye Institute, National Institutes of Health Bethesda, Maryland 20892
Received July 13, 1989, and in revised form September 13, 1989
A lens epithelial cell line established from a trans- genie mouse synthesizes high levels of the enzyme al- dose reductase which converts sugars to polyols. This enzyme has been implicated in the formation of sugar cataracts in animals and with diabetic complications in man. The mouse aldose reductase has been character- ized and by sodium dodecyl sulfate-polyacrylamide gel electrophoresis has an apparent molecular mass of 38,000, similar to the enzyme in rat and man. The cel- lular enzyme is inhibited by two aldose reductase inhib- itors: Sorbinil (I&,, = 1.8 X lo-’ M) and Alcon 1576 (ICE0 = 7.8 X 10-‘M). The amount and the specific activ- ity of the aldose reductase can be further increased in the cells by raising the osmolarity of the medium to 500 mOSM. Although the amount of aldose reductase is in- creased approximately sevenfold under these condi- tions, a-crystallin, one of the main lens specific pro- teins, remained at about the same concentration. No detectable increase in sorbitol was found within the cells, in contrast to published reports on renal cells in which this polyol increases under similar hyperosmotic conditions; however, in the lens cells there was a five- fold increase in the inositol content, suggesting that this polyol rather than sorbitol may be used to compensate for some of the changes in the osmolarity. The induction of the enzyme aldose reductase without the apparent accumulation of its product suggests a complex mecha- nism for OSXUOregUlatiOU in the lens CellS. cr mm Academic
Press, Inc.
The enzyme aldose reductase (AR)’ is a member of the aldo-ketoreductases which catalyze the reduction of sugars to polyols (1). Recently, AR has been shown to be
’ To whom correspondence should be addressed at Bldg. 6, Rm. 228, National Institutes of Health, Bethesda, MD 20892.
’ Abbreviations used: AR, aldose reductase; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; IEF, isoelectric fo- cusing.
000%9861/90 $3.00 Copyright 0 1990 by Academic Press, Inc. All rights of reproduction in any form reserved.
present in cultured rabbit renal inner medullary cells and this enzyme can be induced with an increase in os- molarity in culture medium (2, 3). One hypothesis that has been suggested is that osmoregulation in the kidney cells is a result of the change in the AR enzyme and re- sulting fluxes of sorbitol within the cell (4). It is not cer- tain at present if the induction of the enzyme is charac- teristic of certain types of cells or whether it represents a more general reaction to major changes in osmolarity.
For several years, AR has been implicated in causing sugar cataracts in animals and may be related to galac- tose cataracts in man (5). This enzyme is found in many of the tissues of the eye (5-8) and is present in the lens epithelium in apparently higher concentration than in the differentiated cells of the deeper lens (9). One of the main reasons for the interest in AR beside its role in cat- aract formation has been the hypothesis that this en- zyme is related to some of the complications found in diabetes. Galactose cataracts in rats can be prevented by inhibitors of AR (lo), and the basement membrane thickening of retinal vessels that is found as a complica- tion in diabetes is also lessened when animals are given inhibitors of this enzyme (11). The relationship of this enzyme to diabetes in man is not as clear, although there is an increase in the amount of sorbitol, the polyol which is derived from glucose, in endoneurial tissue (12).
An epithelial cell line, aTN4, from the lens of a mouse was established from a transgenic animal that had the T-antigen from SV40 virus linked to the aA-crystallin promoter (13). The cell line was found to synthesize CY- crystallin, a protein which constitutes 30% of the mouse lens. In addition to the olA-crystallin the cells also syn- thesize (YB- and aInsert-crystallins. The level of the cy- crystallin produced was found to be 1.6% of the total sol- uble protein in this cell line (T. Yamada, and P. Russell, submitted for publication). The a-crystallin present in the cell line shared similar properties with a-crystallin isolated from the mouse lens in that it aggregated into very large molecular weight components and the sub- units appeared to undergo the same type of post-transla-
259

260 RUSSELL. YAMADA, AND MEROLA
SDS IEF
PH A
FIG. 1. SDS-PAGE and IEF gels (A) and Western blots (B) of the rat lens (1) and mouse lens epithelial cell line (2) soluble proteins. The Western blots were incubated with antibody to rat lens aldose reductase. The positions of the molecular mass standards are shown to the left of the SDS gel and the pH gradient is indicated by the IEF gel. The immunoreactive AR has an approximate molecular mass of 38,000 and for the mouse, the main band of AR had a p1 of 5.1.
tional modification found in the lens in vim (14, 15). This cell line may be very important to the understand- ing of the development of the eye since the crystallins in general (16, 17) and cu-crystallin in specific (18) have been implicated in the abnormal development of the eyes of certain animals.
To test whether the induction of the AR enzyme might be common to cell lines other than renal cells, the mouse cell line was chosen because not only is this enzyme im- portant to lens metabolism but also the cell line offered a readily reproducible system to study AR. In addition, the cell line had substantial synthesis of a lens specific protein that could be measured during manipulation of the culture conditions and could be used as a marker for general protein accumulation. Since sorbitol is a product of the AR enzyme reaction but its accumulation is thought to be detrimental to the lens, the changes in this sugar alcohol under hyperosmotic conditions were also measured.
MATERIAL AND METHODS
The mouse lens epithelial cell line, LuTN~, was maintained in Dul- becco’s MEM medium with 4500 mg/liter of glucose and 10% heat- inactivated fetal bovine serum. The culture medium had an osmolarity of 330 mOSM. To increase osmolarity of the culture medium, NaCl was added to bring the osmolarity of the medium to 500 mOSM. When galactose was added to the culture media, the final concentration of the sugar was 30 mM. Incubation under experimental conditions was for 3 days after which the cells were collected by scraping the flasks. The cells were washed three times in phosphate-buffered saline. The cells could then be sonicated or homogenized in H20 and centrifuged at 14,000g for 10 min to separated the soluble fraction from the insolu- ble pellet. Protein determinations were done according to the method of Bradford (19). SDSPAGE and isoelectric focusing (IEF) were done on the Phastsystem using the manufacturer’s suggested protocol (Pharmacia LKB, Piscataway, NJ). Western blots were performed us- ing an antibody dilution of 1:500 for the rat lens AR antibody and a dilution of 1:lOO for the u-crystallin antibody. Incubation of the nitro- cellulose blots with the primary antibody was for 12 h at 4°C. Peroxi- dase-labeled second antibody was used with the AR antibody, whereas biotinylated second antibody and avidin-biotinylated peroxidase was
used for the a-crystallin blots (Vector Labs, Burlingame, CA). The blots were incubated in the secondary antibody for 1 h at 37°C. The immunoreactive bands were visualized with 4-chloro-l-naphthol.
Slot blot analyses were done by adding 50 ~1 of sample to each well of a slot blot apparatus. The wells were washed and then processed similar to Western blots. A minimum of four dilutions was done with each sample and three separate sets of experiments were done. Quanti- tation of the blots was done with a laser densitometer (Pharmacia LKB) and the data, analyzed by Gelscan LC software, were normal- ized for each experiment by setting the a-crystallin content of the con- trol sample and the AR content of the high osmolarity sample at 100%.
The enzyme activities of AR in the samples were determined as pre- viously described (6, 8, 9). The assay measured the decrease in the absorbance of NADPH over time. For determination of the IC,, con- centrations of inhibitors of AR, DL-glyceraldehyde was used as the substrate. Results of the enzyme assays are reported as the change in optical density at 340 nm per one minute per milligram of soluble pro- tein. Enzyme activity measurements were done three separate times for each substrate using different cell homogenates.
Cells were prepared for gas chromatography by homogenization in a equal volumes of ZnS04 and Ba(OH),, (8). The supernatants were lyophilized and derivatized by silylation (20) using N,O-bis(trimethyl- silyl)trifluoroacetamide with 1% trimethylchlorosilane. An aliquot was injected using pyridine as a solvent into a Varian 3700 gas chro- matograph equipped with a SE-52 column (Supelco, Inc., Bellefonte, PA). Results are expressed as nanomoles per milligram protein.
RESULTS
The lens epithelial cell line synthesized the AR en- zyme with an apparent molecular mass of 38,000 by SDS-PAGE. This value was similar to the value in rat lens (Fig. 1). No immunoreactive AR was observed in the homogenates of young mouse lenses, indicating that the level of the enzyme in the cell line was higher than that in the whole lens which included the central enucleated fiber cells. The isoelectric focusing pattern of the sample from the cell line showed that the main band of immuno- reactivity was more basic than the main band found with the rat lens. The pI of the mouse AR was 5.1 compared to the value of around 4.8 for the rat lens. Minor bands were present in the mouse sample at around pI 5.3 and 5.7 and a minor band was also present in the rat sample at pI 5.0. The enzyme activity measurements of the AR from the cultured cells were normally done with 100 pg of total soluble protein in the l-ml reaction mixture (Ta- ble I). For reference, AR from a weanling rat lens, one of
TABLE I
Aldose Reductase Activity in Mouse Lens Epithelial Cell Line
Substrate
Dl,-Glyceraldehyde DL-Glyceraldehyde + LiSO, L-Gulono-y-lactone II-Glucuronate D-XylOSe
oL,-Glyceraldehyde (cells at 500 mOSM)
Specific activity ( AOD/min/mg protein)
0.022 f 0.003 0.032 + 0.004
Not detectable 0.006 f 0.001 0.010 f 0.001
0.073 f 0.007
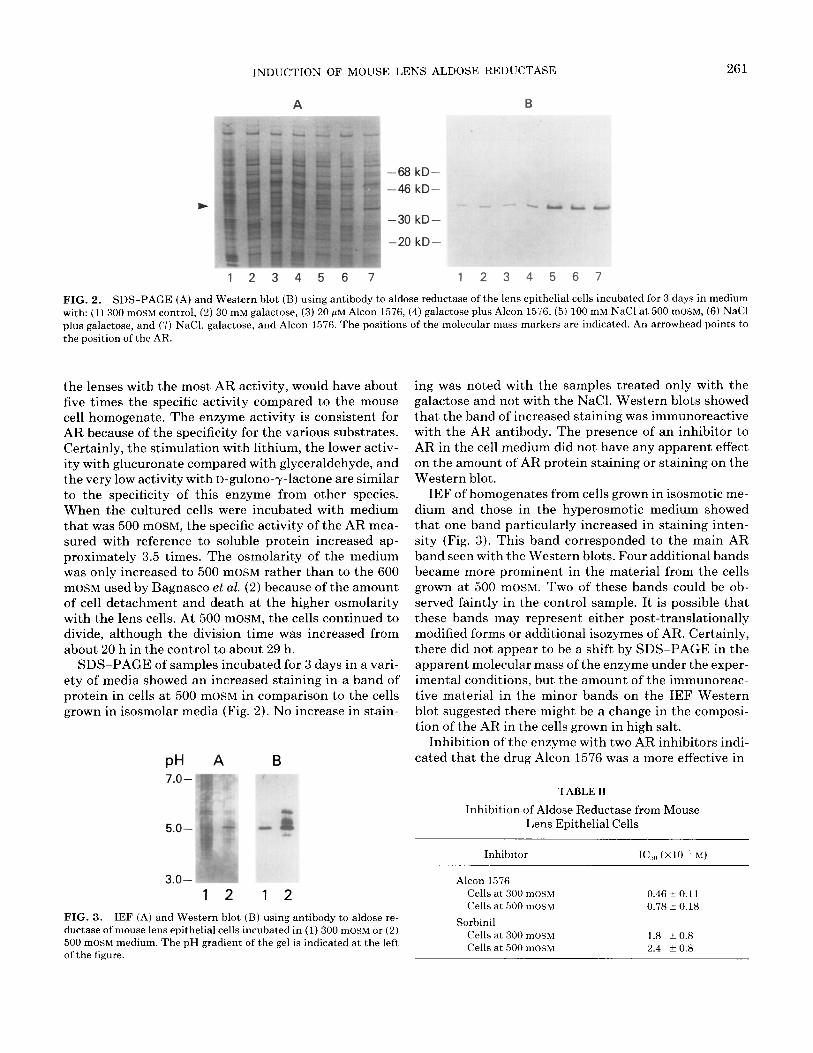
INDUCTION OF MOUSE LENS ALDOSE REDUCTASE 261
1234567 1234567
FIG. 2. SDS-PAGE (A) and Western blot (B) using antibody to aldose reductase of the lens epithelial cells incubated for 3 days in medium with: (1) 300 mOSM control, (2) 30 mM galactose, (3) 20 pM Alcon 1576, (4) galactose plus Alcon 1576, (5) 100 mM NaCl at 500 mOSM, (6) NaCl plus galactose, and (7) NaCl, galactose, and Alcon 1576. The positions of the molecular mass markers are indicated. An arrowhead points to the position of the AR.
the lenses with the most AR activity, would have about five times the specific activity compared to the mouse cell homogenate. The enzyme activity is consistent for AR because of the specificity for the various substrates. Certainly, the stimulation with lithium, the lower activ- ity with glucuronate compared with glyceraldehyde, and the very low activity with D-gulono-y-lactone are similar to the specificity of this enzyme from other species. When the cultured cells were incubated with medium that was 500 mOSM, the specific activity of the AR mea- sured with reference to soluble protein increased ap- proximately 3.5 times. The osmolarity of the medium was only increased to 500 mOSM rather than to the 600 mOSM used by Bagnasco et al. (2) because of the amount of cell detachment and death at the higher osmolarity with the lens cells. At 500 mOSM, the cells continued to divide, although the division time was increased from about 20 h in the control to about 29 h.
SDS-PAGE of samples incubated for 3 days in a vari- ety of media showed an increased staining in a band of protein in cells at 500 mOSM in comparison to the cells grown in isosmolar media (Fig. 2). No increase in stain-
PH A B
FIG. 3. IEF (A) and Western blot (B) using antibody to aldose re- ductase of mouse lens epithelial cells incubated in (1) 300 mOSM or (2) 500 mOSM medium. The pH gradient of the gel is indicated at the left of the figure.
ing was noted with the samples treated only with the galactose and not with the NaCl. Western blots showed that the band of increased staining was immunoreactive with the AR antibody. The presence of an inhibitor to AR in the cell medium did not have any apparent effect on the amount of AR protein staining or staining on the Western blot.
IEF of homogenates from cells grown in isosmotic me- dium and those in the hyperosmotic medium showed that one band particularly increased in staining inten- sity (Fig. 3). This band corresponded to the main AR band seen with the Western blots. Four additional bands became more prominent in the material from the cells grown at 500 mOSM. Two of these bands could be ob- served faintly in the control sample. It is possible that these bands may represent either post-translationally modified forms or additional isozymes of AR. Certainly, there did not appear to be a shift by SDS-PAGE in the apparent molecular mass of the enzyme under the exper- imental conditions, but the amount of the immunoreac- tive material in the minor bands on the IEF Western blot suggested there might be a change in the composi- tion of the AR in the cells grown in high salt.
Inhibition of the enzyme with two AR inhibitors indi- cated that the drug Alcon 1576 was a more effective in-
TABLE II
Inhibition of Aldose Reductase from Mouse Lens Epithelial Cells
Inhibitor
Alcon 1576 cells at 300 mOSM
Cells at 500 mOSM
Sorbinil cells at 300 mOSM
cells at 500 mOSM
I&,, (X10 ‘M)
0.46 + 0.11 0.78 + 0.18
1.8 t 0.8 2.4 + 0.8
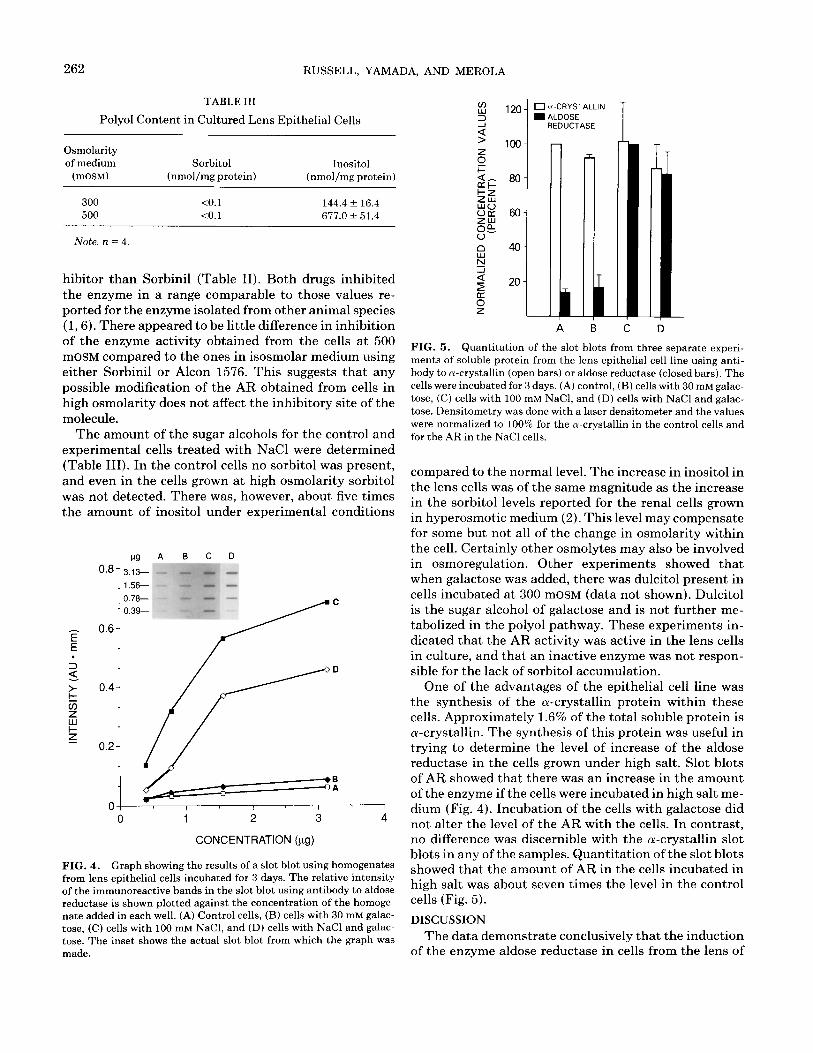
262 RUSSELL, YAMADA, AND MEROLA
TABLE III
Polyol Content in Cultured Lens Epithelial Cells
Osmolarity of medium
(mOSM)
Sorbitol Inositol (nmol/mg protein) (nmol/mg protein)
300 10.1 144.4 f 16.4 500 <O.l 677.Ok 51.4
Note. n = 4.
hibitor than Sorbinil (Table II). Both drugs inhibited the enzyme in a range comparable to those values re- ported for the enzyme isolated from other animal species (1,6). There appeared to be little difference in inhibition of the enzyme activity obtained from the cells at 500 mOSM compared to the ones in isosmolar medium using
either Sorbinil or Alcon 1576. This suggests that any possible modification of the AR obtained from cells in high osmolarity does not affect the inhibitory site of the molecule.
The amount of the sugar alcohols for the control and experimental cells treated with NaCl were determined (Table III). In the control cells no sorbitol was present, and even in the cells grown at high osmolarity sorbitol was not detected. There was, however, about five times the amount of inositol under experimental conditions
E .
pg A B C D
cm- 3.13- _I _ 1.5s --
0.78- * 0.3%
0 1 2 3 4
CONCENTRATION (pg)
FIG. 4. Graph showing the results of a slot blot using homogenates from lens epithelial cells incubated for 3 days. The relative intensity of the immunoreactive bands in the slot blot using antibody to aldose reductase is shown plotted against the concentration of the homoge- nate added in each well. (A) Control cells, (B) cells with 30 mM galac- tose, (C) cells with 100 mM NaCl, and (D) cells with NaCl and galac- tose. The inset shows the actual slot blot from which the graph was made.
0 u-CRYSTALLIN m ALDOSE
REDUCTASE
I A
FIG. 5. Quantitation of the slot blots from three separate experi- ments of soluble protein from the lens epithelial cell line using anti- body to n-crystallin (open bars) or aldose reductase (closed bars). The cells were incubated for 3 days. (A) control, (B) cells with 30 mM galac- tose, (C) cells with 100 mM NaCl, and (D) cells with NaCl and galac- tose. Densitometry was done with a laser densitometer and the values were normalized to 100% for the ol-crystallin in the control cells and for the AR in the NaCl cells.
compared to the normal level. The increase in inositol in the lens cells was of the same magnitude as the increase in the sorbitol levels reported for the renal cells grown in hyperosmotic medium (2). This level may compensate for some but not all of the change in osmolarity within the cell. Certainly other osmolytes may also be involved in osmoregulation. Other experiments showed that when galactose was added, there was dulcitol present in cells incubated at 300 mOSM (data not shown). Dulcitol is the sugar alcohol of galactose and is not further me- tabolized in the polyol pathway. These experiments in- dicated that the AR activity was active in the lens cells in culture, and that an inactive enzyme was not respon- sible for the lack of sorbitol accumulation.
One of the advantages of the epithelial cell line was the synthesis of the a-crystallin protein within these cells. Approximately 1.6% of the total soluble protein is a-crystallin. The synthesis of this protein was useful in trying to determine the level of increase of the aldose reductase in the cells grown under high salt. Slot blots of AR showed that there was an increase in the amount of the enzyme if the cells were incubated in high salt me- dium (Fig. 4). Incubation of the cells with galactose did not alter the level of the AR with the cells. In contrast, no difference was discernible with the a-crystallin slot blots in any of the samples. Quantitation of the slot blots showed that the amount of AR in the cells incubated in high salt was about seven times the level in the control cells (Fig. 5).
DISCUSSION
The data demonstrate conclusively that the induction of the enzyme aldose reductase in cells from the lens of

INDUCTION OF MOUSE LENS ALDOSE REDUCTASE 263
the eye occurred when the osmolarity of the culture me- dium was increased. The specific activity of the enzyme increases about fourfold under conditions of high osmo- larity but this increase is most probably the result of an increase in the amount of the enzyme in the cells rather than an activation of the enzyme itself. The results are consistent with those reported from the renal medullary cells (2, 3) and suggest that a common mechanism for the induction of the AR may occur in cultured cells in response to increases in osmolarity of the cell medium. Even though the enzyme is increased in the cells incu- bated in the high salt medium, no sorbitol was detected in these cells. One possible explanation for the low level of sorbitol could be that the enzyme sorbitol dehydroge- nase which degrades sorbitol to fructose prevented the accumulation of this sugar alcohol. In the lens cells as with the renal cells (4), the level of sorbitol dehydroge- nase activity was at the low end of the detectable range. There was, however, a small increase in the amount of fructose in cells incubated in high salt medium, suggest- ing that more glucose is metabolized through the polyol pathway under hyperosmotic conditions.
In contrast to the low level of sorbitol under experi- mental conditions, the amount of the inositol increased substantially. Thus, the response of the lens cells to an increase in osmolarity may be somewhat different than that reported for the renal cells in culture. Although one hypothesis suggests that the renal cells increase sorbitol as a mechanism to compensate for the increased osmo- larity in the culture medium, the lens cells do not appear to utilize sorbitol for this purpose. The increase in inosi- to1 would not appear to be linked to AR or the polyol pathway directly, but changes in the inositol content would certainly influence metabolic pathways.
The AR from the mouse cells is similar in molecular mass to the enzyme that has been prepared from other species. This is the first characterization of the AR from mouse cells. One of the reasons that the mouse has not been used in studies of the polyol pathway has been the relatively low level of the AR in the mouse lens. At- tempts to induce a galactose cataract in mouse have been unsuccessful because of the this low activity of AR in the lens. The difhculty in getting the disease state in this species can be compared to the relative ease and repro- ducibility of getting the cataract in the rat. The enzyme L-hexonate dehydrogenase was thought to be present in the mouse lens rather than AR (21). The specificity and immunoreactivity of the enzyme in the cultured cells in- dicate that AR was indeed present in the cell line ob- tained from the mouse lens and that the mouse enzyme shares properties with AR from other species.
The mouse AR has a slightly more basic isoelectric point than the enzyme from the rat lens. The principal immunoreactive band of the mouse can be clearly seen in the IEF of the samples incubated in high osmolarity medium. Under these conditions, four additional bands
of immunoreactivity become evident in the mouse sam- ple. The relationship among the immunoreactive bands is unknown; however, all of the immunoreactive mate- rial had the same apparent molecular weight on SDS- PAGE. A possible post-translational modification of the AR has not really been well researched although multi- ple bands are reported in the IEF pattern from other spe- cies. It may be that modifications in the enzyme occur more easily under hyperosmotic conditions.
One of the obvious questions concerning the induction is what is regulating the AR. The induction of the AR in the lens cells suggests that the control of this enzyme is somehow related to the osmolarity of the culture me- dium. The reason for the induction of the AR in the mouse cells does not seem as straightforward as in the renal cells unless there is only one gene for the AR in the mouse or the gene for the AR that is present in the lens cells is the same one that is expressed in the kidney cells. Alternatively, since the enzyme is induced, new iso- zymes of AR may be regulated by the high salt condi- tions. With this cell line it may be possible to resolve these questions.
Although these cells should be capable of forming sor- bitol, inositol appears to be one of the osmolytes that allow the cells to compensate at least partially for the increase in osmolarity. However, the increase in the amount and specific activity of the AR would implicate this enzyme and the polyol pathway in the regulation of cellular homeostasis under hyperosmotic conditions. Explanations for the induction of AR and regulation of this induction may help in the understanding of osmo- regulation in general and may help elucidate any rela- tionship this enzyme might have with inositol.
ACKNOWLEDGMENTS
We thank Drs. Heiner Westphal and Takafumi Nakamura of the National Institute of Child Health and Human Development for the gift of the cell line, aTN4, J. Samuel Zigler, Jr., of the National Eye Institute for his antibody to a-crystallin, and Peter Kador of the Na- tional Eye Institute for his antibody to aldose reductase. We also thank Dr. Marjorie F. Lou of Alcon Laboratories, Ft. Worth, Texas, for the Alcon 1576.
REFERENCES
1. Wermuth, B. (19851 in Enzymology of Carbonyl Metabolism. 2. Aldehyde Dehydrogenase, Aldo-keto Reductase, and Alcohol De- hydrogenase (Flynn, T. G., and Weiner, H., Eds.), pp. 209-236, A. R. Liss, New York.
2. Bagnasco, S. M., Uchida, S., Balaban, R. S., Kador, P. F., and Burg, M. B. (1987) Proc. N&Z. Acad. Sci. USA 84,1718~1720.
3. Bedford, J. J., Bagnasco, S. M., Kador, P. F., Harris, H. W., Jr., and Burg, M. B. (1987) J. Biol. Chem. 262, 14,255-14,259.
4. Bagnasco, S. M., Murphy, H. R., Bedford, J. J., and Burg, M. B. (1988) Amer. J. Physiol. 254, C788-C792.
5. Kinoshita, J. H. (1965) Znuest. Ophthalmol. 4, 786-799.
6. Kador, P. F., Kinoshita, J. H., Tung, W. H., and Chylack, L. T., Jr. (1980) Znuest. Ophthalmol. Vis. Sci. 19,980-982.

264 RUSSELL, YAMADA, AND MEROLA
7. Kern, T. S., and Engerman, R. L. (1981) Exp. Eye Res. 33, 175- 182.
8. Russell, P., Merola, L. O., Yajima, Y., and Kinoshita, J. H. (1982) Exp. Eye Res. 35,331-336.
9. Kador, P. F., Carper, D., and Kinoshita, J. H. (1981) Anal. Bio- them. 114,53-58.
10. Hu, T. S., Datiles, M., and Kinoshita, J. H. (1983) Inuest. Ophthal- mol. Vis. Sci. 24,640-644.
11. Robison, Jr., W. G., Kador, P. F., Akagi, Y., Kinoshita, J. H., Gon- zalez, R., and Dvornik, D. (1986) Diabetes 35,295-299.
12. Dyck, P. J., Zimmerman, B. R., Vilen, T. H., Minnerath, S. R., Karnes, J. L., Yao, J. K., and Poduslo, J. F. (1988) New Engl. J. Med. 319,542-548.
13. Mahon, K. A., Chepelinsky, A. B., Khillan, J. S., Overbeek, P. A., Piatigorsky J., and Westphal, H. (1987) Science 235, 1622-1628.
14. Chiesa, R., Gawinowicz, M. A., Kleinman, N. J., and Spector, A. (1988) Exp. Eye Rex 46,199-208.
15. Piatigorsky, J. (1981) Differentiation 19, 134-153.
16. Breitman, M. L., Clapoff, S., Rossant, J., Tsui, L.-C., Glode, L. M., Maxwell, I. H., and Bernstein, A. (1987) Science 238,1563-1565.
17. Landel, C. P., Zhao, J., Bok, D., and Evans, G. A. (1988) Genes Deu. 2,1168-1178.
18. Hendriks, W., Leunissen, J., Nevo, E., Bloemendal, H., and de Jong, W. W. (1987) Proc. Natl. Acad. Sci. USA 84,5320-5324.
19. Bradford, M. M. (1976) Anal. Biochem. ‘72,248-254.
20. Sweeley, C. C., Bently, A., Makito, M., and Wills, W. W. (1963) J. Amer. Chem. Sot. 85,2495-2507.
21. Markus, H. B., Raducha, M., and Harris, H. (1983) Biochem. Med. 29,31-45.





























