Embed Size (px)
Citation preview

Induction of systemic resistance to anthracnose in cucumber by natural components of Allium vegetables and shiitake mushrooms
H. Inagaki1,, A. Yamaguchi1, K. Kato1,, C. Kageyama1 and H. Iyozumi1 1 Shizuoka prefectural research institute of agriculture and forestry, Shizuoka, Japan
Systemic resistance (SR) against microbial pathogens—the introduction of broad-spectrum and long-lasting immunity in non-infected tissues by induction of the plant’s own defense mechanisms—has gained considerable attention in crop production. Here, we report that extracts of Allium vegetables, such as rakkyo (Allium chinense) and garlic (A. sativum), and shiitake mushrooms induce SR in cucumber. Anthracnose symptoms were less severe in cucumber plants treated with these plant extracts than in those treated with a water control. Further, the expression of the peroxidase (POD) gene—a putative SR marker—was higher in cucumber plants treated with the plant extracts. Similarly, we found that addition of waste from cultures of the shiitake mushroom Lentinus edodes to the soil induced SR in cucumber plants, with particular efficacy against microbial pathogens in nursery cucumber production.
Keywords systemic resistance; anthracnose; natural products; crop production
1. Introduction
1.1 Systemic resistance (SR)
Chemical pesticides are often used for pathogen control in plants. However, the negative environmental impact of pesticides has raised the need for urgent development of alternative compounds. Moreover, the use of chemical pesticides encourages the growth of pesticide-resistant pathogens. In this scenario, plant activator-induced systemic resistance (SR) has gained considerable attention in crop production as a novel next-generation pesticide [1–4]. In SR, the plant’s own defense mechanisms are induced in order to achieve broad-spectrum and long-lasting immunity in non-infected tissues [5-7]. Plant activators are novel substances that protect plants by enhancing their inherent disease-resistance mechanisms [8-10]. At present, several plant activators, including probenazole, acibenzolar-S-methyl, carpropamid, and tiadinil, are used in commercially available chemical agents [11-15]. Plant activators do not act directly on the pathogen, but rather activate the defense mechanisms of the plant, ultimately activating disease pathogen control. Therefore, it is expected that plant activators do not favor the growth of resistant pathogens. On the other hand, they may contribute to the reduction of negative environmental impact [16].
1.2 SR induced by natural products
Several reports have documented the antifungal and insecticidal activities of natural products. In contrast, studies on natural product-induced SR are limited. Yet, it has been shown that yeast extracts, such as AgrevoEX® (Agrevo Co.,Kawasaki, Japan) and Housakumonogatari® (Asahi Food and Healthcare Co., Tokyo, Japan), induce SR in plants [17-19]. In addition, it is known that extracts of plants that contain abundant amounts of oxalic acid, such as rhubarb and spinach, induce SR to other plants [20]. Moreover, formulated extracts of giant knotweed (Reynoutria sachalinenesis) are part of the commercially available plant activator, Milsana® (KHH BioSci, Inc. Raleigh, USA) [21-25]. Here, we show that natural components extracted from Allium vegetables and shiitake mushrooms induce SR in cucumber plants.
2. Material and methods
2.1 Cucumber anthracnose bioassay
Colletorichum orbiculare was maintained on pseudomonas-selective agar (PSA) in the dark at 25 C. Conidial suspensions were prepared from 6-d cultures. Three pregerminated seeds of cucumber (hanfushinari) were sown in a pot of 10.5 cm of diameter. Two weeks after seedling, a conidial suspension of C. orbiculare (106 spores/ml) containing 0.001% Tween 20 was inoculated onto the surface of the cotyledon and the first leaf of each plant, using a hand sprayer (2 ml per plant). The inoculated plants were placed in moistened humid containers that were sealed and maintained at 25 C. After 24 h, the containers were opened and the plants were returned to the chambers at 25 C under long-day photoperiod conditions (16 h, 3000 lux). The plants were observed daily for the presence of C. orbiculare disease
728 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

symptoms. Four days after inoculation, the disease incidence was determined as the percentage of infected area in the total area of the inoculated leaves (cotyledon) and upper leaves (first true leaf). Three pots (9 cucumber plants) per treatment were used in each experiment.
2.2 Expression of the POD gene in cucumber plants
The first leaf of each cucumber plant was sampled after 2 weeks of seedling. Isolation of total RNA from the cucumber leaf was performed using the Agilent Plant RNA Isolation Mini Kit (Agilent Technologies Inc., California, USA) with an on-column DNase treatment (Qiagen Sciences, Maryland, USA). Complementary DNAs were synthesized from 500 ng of total RNA primed with random hexamers using the Perfect Real Time™ RT–PCR kit (Takara Bio, Tokyo, Japan) according to the manufacturer’s protocol. To quantify Acidic Peroxidase 3UTR (POD) gene [26] expression, a 40-fold dilution of cDNA mixture was used as template with the SYBR® Premix Ex Taq™ (Takara Bio) in a quantitative real-time PCR assay performed on the Mx3000P Real-Time PCR System (Stratagene). Reverse-transcribed mRNAs of the POD gene were amplified with a specific primer pair (5-tcaggatgggaaatctcgac-3 and 5-cgtggccaactcatacacac-3; 188-bp amplicon). The expression levels of the POD gene were normalized against the expression of an actin gene (5-tgctggtcgtgaccttactg-3 and 5-gaatctctcagctccgatgg-3; 226-bp amplicon) [27]. We confirmed that actin expression was stable in all treatments. The cycling conditions were 10 s of polymerase activation at 95 °C and 40 cycles at 95 °C for 5 s and 64 °C for 30 s. Each assay included (in duplicate): a standard curve of 8 serial dilution points of the water-treated sample for the internal control gene or plant activator (acibenzolar-S-methyl)-treated sample for the target gene (ranging from 50 ng to 640 ng), a no-template control for each primer set, and each test cDNA. All assays were performed in triplicate.
2.3 Systemic resistance in cucumber plants induced by extracts of Allium vegetables
Leafs of Allium species were cut into 1-cm pieces and soaked in distilled water (2 g of wet weight/10 ml). Then, samples were homogenized by using a Physcotron (Microtec Co., Funabashi, Japan). For embrocation, 1 ml of plant extract was applied onto the surface of the cotyledon of 0.5-leaf stage cucumber plants using a cotton bud (Fig. 1). For soil application, 10 ml of plant extract was applied to the soil. The treated cucumber plants were placed in chambers at 25 °C under long-day photoperiod conditions (16 h, 3000 lux). Two days after treatment, conidial suspensions of C. orbiculare were prepared as described above. Distilled water was used as negative control for each treatment. As a positive control, 10 ml of 50 ppm of acibenzolar-S-methyl were used. It has been previously suggested that acibenzolar-S-methyl induces SR in cucumber plants [28].
Fig. 1 Schematic representation of the bioassay for induction of SR by embrocation.
2.4 Systemic resistance in cucumber plants induced by waste from Lentinus edodes cultures
Nurse Houdo soil (Houchi Agrico Co., Ltd., Omaezaki, Japan) was used as control. Culture waste was autoclaved 1 h at 120 C. Cracked and spawned substrate used for L. edodes culture and the waste from the later were mixed with equal volumes of control. Three imbibed cucumber seeds were sown in a pot, with a diameter 10.5 cm, containing mixed compost. Two weeks after sowing, conidial suspensions of C. orbiculare were prepared as described above. Controls were the same as above. Three plants per treatment were used in each experiment, and each experimental set was performed in duplicate.
Four days after inoculation, the disease incidence was assessed as a percentage of leaf infected area of the first true leaf, and the percentage of distilled water control was calculated.
Two days after treatment, conidial suspensions of C. orbiculare (10-6 spores ml-1) were inoculated to the surface of cotyledon and first true leaf of plants.
Plant extract was applied to the surface of cotyledon of cucumber plants.
Four days after inoculation, the disease incidence was assessed as a percentage of leaf infected area of the first true leaf, and the percentage of distilled water control was calculated.
Two days after treatment, conidial suspensions of C. orbiculare (10-6 spores ml-1) were inoculated to the surface of cotyledon and first true leaf of plants.
Plant extract was applied to the surface of cotyledon of cucumber plants.
729©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________
3. Systemic resistance is induced in cucumber plants by extracts of Allium vegetables
3.1 Effect of extract of Allium vegetables on anthracnose symptoms
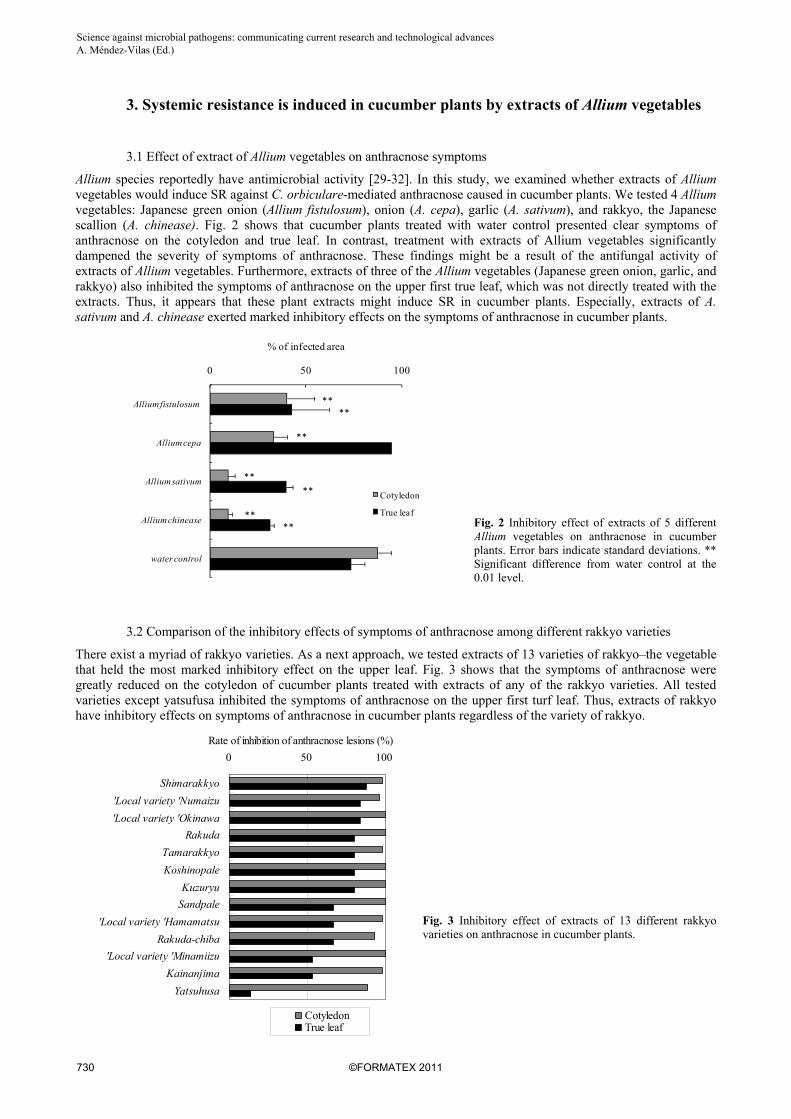
Allium species reportedly have antimicrobial activity [29-32]. In this study, we examined whether extracts of Allium vegetables would induce SR against C. orbiculare-mediated anthracnose caused in cucumber plants. We tested 4 Allium vegetables: Japanese green onion (Allium fistulosum), onion (A. cepa), garlic (A. sativum), and rakkyo, the Japanese scallion (A. chinease). Fig. 2 shows that cucumber plants treated with water control presented clear symptoms of anthracnose on the cotyledon and true leaf. In contrast, treatment with extracts of Allium vegetables significantly dampened the severity of symptoms of anthracnose. These findings might be a result of the antifungal activity of extracts of Allium vegetables. Furthermore, extracts of three of the Allium vegetables (Japanese green onion, garlic, and rakkyo) also inhibited the symptoms of anthracnose on the upper first true leaf, which was not directly treated with the extracts. Thus, it appears that these plant extracts might induce SR in cucumber plants. Especially, extracts of A. sativum and A. chinease exerted marked inhibitory effects on the symptoms of anthracnose in cucumber plants.
Fig. 2 Inhibitory effect of extracts of 5 different Allium vegetables on anthracnose in cucumber plants. Error bars indicate standard deviations. ** Significant difference from water control at the 0.01 level.
3.2 Comparison of the inhibitory effects of symptoms of anthracnose among different rakkyo varieties
There exist a myriad of rakkyo varieties. As a next approach, we tested extracts of 13 varieties of rakkyo–the vegetable that held the most marked inhibitory effect on the upper leaf. Fig. 3 shows that the symptoms of anthracnose were greatly reduced on the cotyledon of cucumber plants treated with extracts of any of the rakkyo varieties. All tested varieties except yatsufusa inhibited the symptoms of anthracnose on the upper first turf leaf. Thus, extracts of rakkyo have inhibitory effects on symptoms of anthracnose in cucumber plants regardless of the variety of rakkyo.
Fig. 3 Inhibitory effect of extracts of 13 different rakkyo varieties on anthracnose in cucumber plants.
0 50 100
Allium fistulosum
Allium cepa
Allium sativum
Allium chinease
water control
% of infected area
Cotyledon
True leaf
**
**
**
**
**
****
0 50 100
Shimarakkyo
'Local variety 'Numaizu
'Local variety 'Okinawa
Rakuda
Tamarakkyo
Koshinopale
Kuzuryu
Sandpale
'Local variety 'Hamamatsu
Rakuda-chiba
'Local variety 'Minamiizu
Kainanjima
Yatsuhusa
Rate of inhibition of anthracnose lesions (%)
CotyledonTrue leaf
730 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________
3.3 Comparison between treatments with extracts of rakkyo
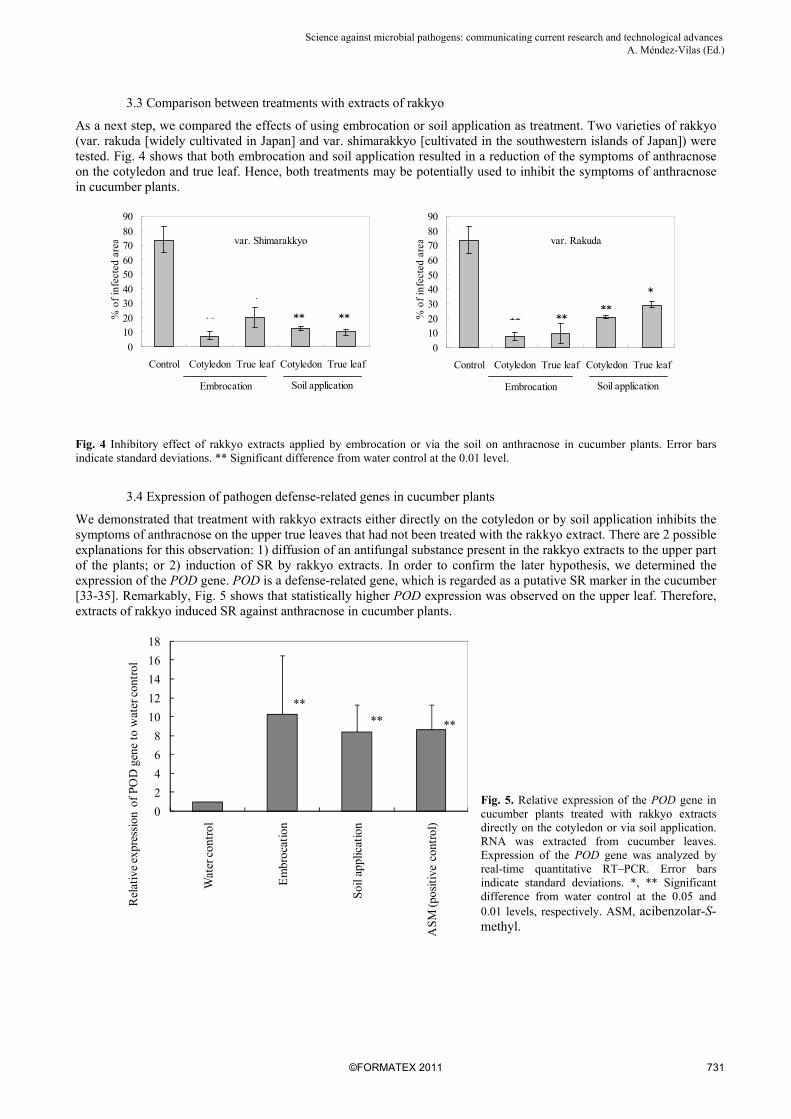
As a next step, we compared the effects of using embrocation or soil application as treatment. Two varieties of rakkyo (var. rakuda [widely cultivated in Japan] and var. shimarakkyo [cultivated in the southwestern islands of Japan]) were tested. Fig. 4 shows that both embrocation and soil application resulted in a reduction of the symptoms of anthracnose on the cotyledon and true leaf. Hence, both treatments may be potentially used to inhibit the symptoms of anthracnose in cucumber plants.
Fig. 4 Inhibitory effect of rakkyo extracts applied by embrocation or via the soil on anthracnose in cucumber plants. Error bars indicate standard deviations. ** Significant difference from water control at the 0.01 level.
3.4 Expression of pathogen defense-related genes in cucumber plants
We demonstrated that treatment with rakkyo extracts either directly on the cotyledon or by soil application inhibits the symptoms of anthracnose on the upper true leaves that had not been treated with the rakkyo extract. There are 2 possible explanations for this observation: 1) diffusion of an antifungal substance present in the rakkyo extracts to the upper part of the plants; or 2) induction of SR by rakkyo extracts. In order to confirm the later hypothesis, we determined the expression of the POD gene. POD is a defense-related gene, which is regarded as a putative SR marker in the cucumber [33-35]. Remarkably, Fig. 5 shows that statistically higher POD expression was observed on the upper leaf. Therefore, extracts of rakkyo induced SR against anthracnose in cucumber plants.
Fig. 5. Relative expression of the POD gene in cucumber plants treated with rakkyo extracts directly on the cotyledon or via soil application. RNA was extracted from cucumber leaves. Expression of the POD gene was analyzed by real-time quantitative RT–PCR. Error bars indicate standard deviations. *, ** Significant difference from water control at the 0.05 and 0.01 levels, respectively. ASM, acibenzolar-S-methyl.
0
2
4
6
8
10
12
14
16
18
Wat
er c
ontr
ol
Em
broc
atio
n
Soi
l app
licat
ion
AS
M (p
osit
ive
cont
rol)
** **
Rel
ativ
e ex
pres
sion
of
PO
D g
ene
to w
ater
con
trol
**
var. Shimarakkyo
0
1020
3040
5060
7080
90
Control Cotyledon True leaf Cotyledon True leaf
% o
f in
fect
ed a
rea
Embrocation Soil application
**
*
** **
var. Rakuda
0
1020
30
4050
60
7080
90
Control Cotyledon True leaf Cotyledon True leaf
% o
f in
fect
ed a
rea
Embrocation Soil application
** ****
*
731©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________
3.5 Garlic extract-induced priming
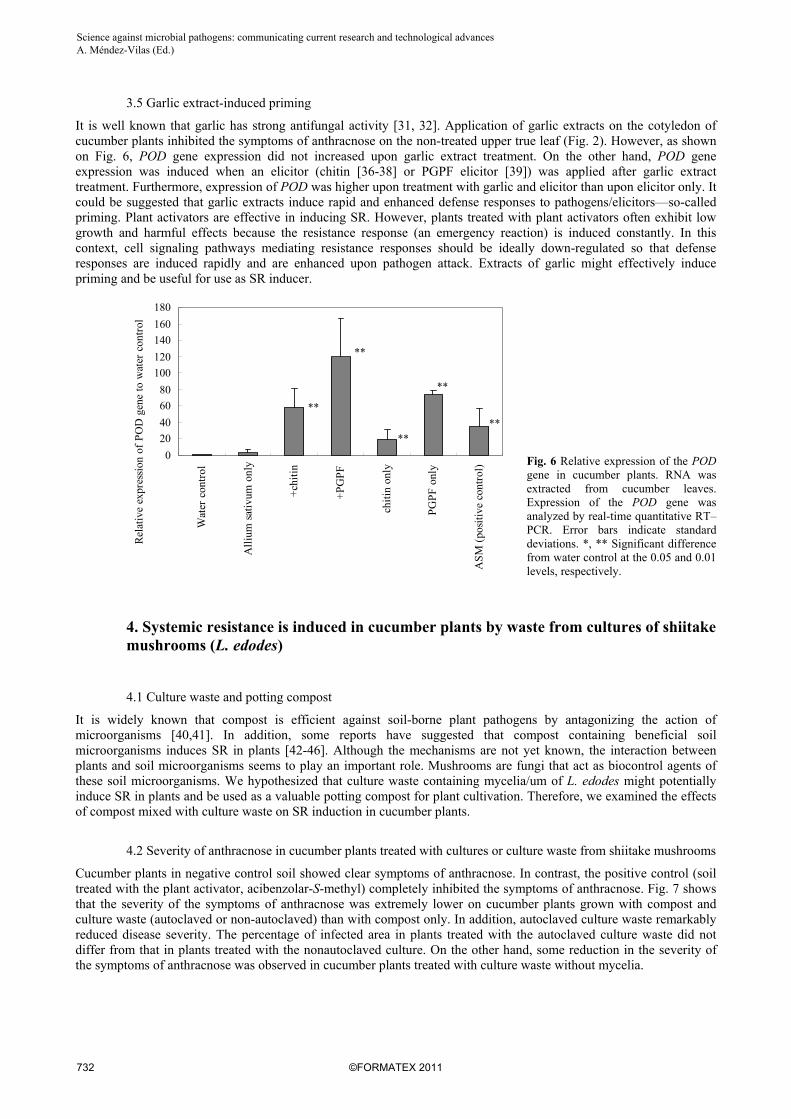
It is well known that garlic has strong antifungal activity [31, 32]. Application of garlic extracts on the cotyledon of cucumber plants inhibited the symptoms of anthracnose on the non-treated upper true leaf (Fig. 2). However, as shown on Fig. 6, POD gene expression did not increased upon garlic extract treatment. On the other hand, POD gene expression was induced when an elicitor (chitin [36-38] or PGPF elicitor [39]) was applied after garlic extract treatment. Furthermore, expression of POD was higher upon treatment with garlic and elicitor than upon elicitor only. It could be suggested that garlic extracts induce rapid and enhanced defense responses to pathogens/elicitors—so-called priming. Plant activators are effective in inducing SR. However, plants treated with plant activators often exhibit low growth and harmful effects because the resistance response (an emergency reaction) is induced constantly. In this context, cell signaling pathways mediating resistance responses should be ideally down-regulated so that defense responses are induced rapidly and are enhanced upon pathogen attack. Extracts of garlic might effectively induce priming and be useful for use as SR inducer.
Fig. 6 Relative expression of the POD gene in cucumber plants. RNA was extracted from cucumber leaves. Expression of the POD gene was analyzed by real-time quantitative RT–PCR. Error bars indicate standard deviations. *, ** Significant difference from water control at the 0.05 and 0.01 levels, respectively.
4. Systemic resistance is induced in cucumber plants by waste from cultures of shiitake mushrooms (L. edodes)
4.1 Culture waste and potting compost
It is widely known that compost is efficient against soil-borne plant pathogens by antagonizing the action of microorganisms [40,41]. In addition, some reports have suggested that compost containing beneficial soil microorganisms induces SR in plants [42-46]. Although the mechanisms are not yet known, the interaction between plants and soil microorganisms seems to play an important role. Mushrooms are fungi that act as biocontrol agents of these soil microorganisms. We hypothesized that culture waste containing mycelia/um of L. edodes might potentially induce SR in plants and be used as a valuable potting compost for plant cultivation. Therefore, we examined the effects of compost mixed with culture waste on SR induction in cucumber plants.
4.2 Severity of anthracnose in cucumber plants treated with cultures or culture waste from shiitake mushrooms
Cucumber plants in negative control soil showed clear symptoms of anthracnose. In contrast, the positive control (soil treated with the plant activator, acibenzolar-S-methyl) completely inhibited the symptoms of anthracnose. Fig. 7 shows that the severity of the symptoms of anthracnose was extremely lower on cucumber plants grown with compost and culture waste (autoclaved or non-autoclaved) than with compost only. In addition, autoclaved culture waste remarkably reduced disease severity. The percentage of infected area in plants treated with the autoclaved culture waste did not differ from that in plants treated with the nonautoclaved culture. On the other hand, some reduction in the severity of the symptoms of anthracnose was observed in cucumber plants treated with culture waste without mycelia.
0
20
40
60
80
100
120
140
160
180
Wat
er c
ontr
ol
All
ium
sat
ivum
onl
y
+ch
itin
+P
GP
F
chit
in o
nly
PG
PF
onl
y
AS
M (
posi
tive
con
trol
)
**
**
Rel
ativ
e ex
pres
sion
of
PO
D g
ene
to w
ater
con
trol
**
**
**
732 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________
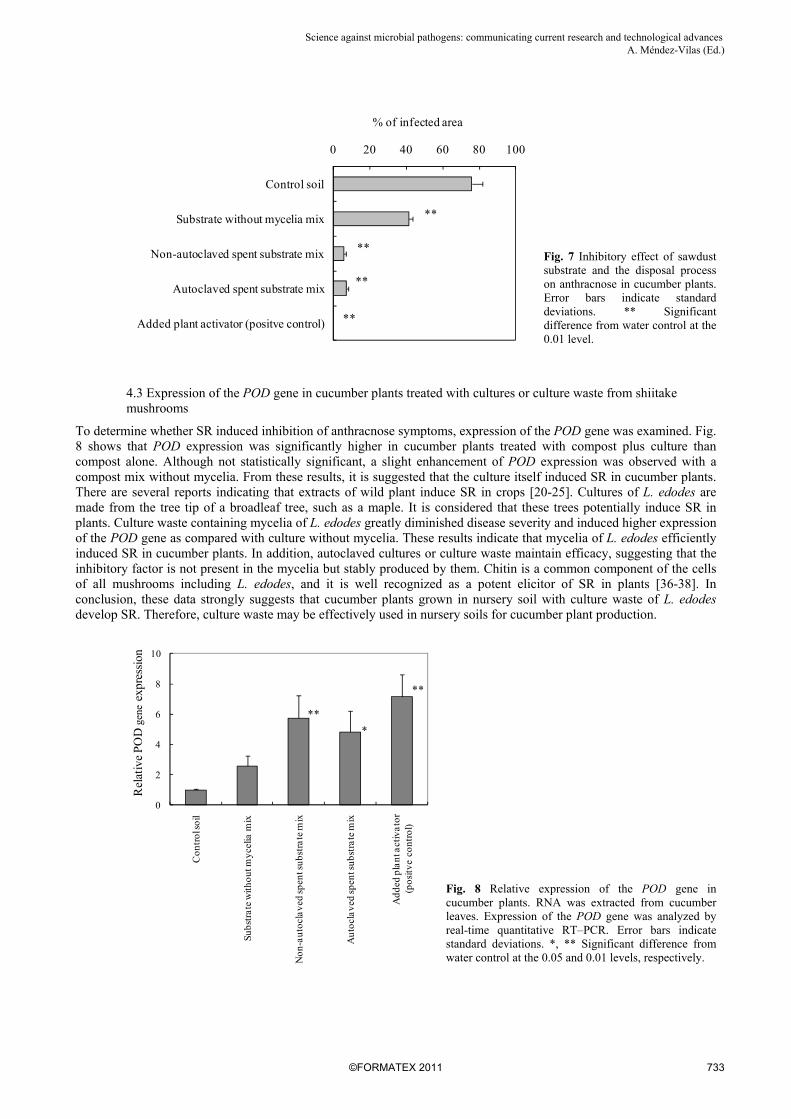
Fig. 7 Inhibitory effect of sawdust substrate and the disposal process on anthracnose in cucumber plants. Error bars indicate standard deviations. ** Significant difference from water control at the 0.01 level.
4.3 Expression of the POD gene in cucumber plants treated with cultures or culture waste from shiitake mushrooms
To determine whether SR induced inhibition of anthracnose symptoms, expression of the POD gene was examined. Fig. 8 shows that POD expression was significantly higher in cucumber plants treated with compost plus culture than compost alone. Although not statistically significant, a slight enhancement of POD expression was observed with a compost mix without mycelia. From these results, it is suggested that the culture itself induced SR in cucumber plants. There are several reports indicating that extracts of wild plant induce SR in crops [20-25]. Cultures of L. edodes are made from the tree tip of a broadleaf tree, such as a maple. It is considered that these trees potentially induce SR in plants. Culture waste containing mycelia of L. edodes greatly diminished disease severity and induced higher expression of the POD gene as compared with culture without mycelia. These results indicate that mycelia of L. edodes efficiently induced SR in cucumber plants. In addition, autoclaved cultures or culture waste maintain efficacy, suggesting that the inhibitory factor is not present in the mycelia but stably produced by them. Chitin is a common component of the cells of all mushrooms including L. edodes, and it is well recognized as a potent elicitor of SR in plants [36-38]. In conclusion, these data strongly suggests that cucumber plants grown in nursery soil with culture waste of L. edodes develop SR. Therefore, culture waste may be effectively used in nursery soils for cucumber plant production.
Fig. 8 Relative expression of the POD gene in cucumber plants. RNA was extracted from cucumber leaves. Expression of the POD gene was analyzed by real-time quantitative RT–PCR. Error bars indicate standard deviations. *, ** Significant difference from water control at the 0.05 and 0.01 levels, respectively.
0 20 40 60 80 100
Control soil
Substrate without mycelia mix
Non-autoclaved spent substrate mix
Autoclaved spent substrate mix
Added plant activator (positve control)
% of infected area
**
**
**
**
0
2
4
6
8
10
Con
trol
soil
Subs
trat
e w
ithou
t myc
elia
mix
Non
-aut
ocla
ved
spen
t sub
stra
te m
ix
Aut
ocla
ved
spen
t sub
stra
te m
ix
Add
ed p
lant
act
ivat
or(p
ositv
e co
ntro
l)
**
**
*
Rel
ativ
e P
OD
gen
eex
pres
sion
733©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________

5. Conclusion
The present work introduces 2 examples of natural products (i.e., extracts of Allium vegetables such as rakkyo and garlic, and culture waste of shiitake mushrooms [L. edodes]) that induce SR., Interestingly, these natural products greatly inhibited the severity of symptoms of anthracnose. However, the underlying mechanisms are unclear. Other SR-inducing compounds are expected to be discovered by screening in the near future.
References
[1] Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner H-Y, Hunt MD. Systemic acquired resistance. Plant Cell 1996; 8: 1809-1819.
[2] Hunt MD, Neuenschwander UH, Delaney TP, Weymann KB, Friedrich LB, Lawton KA, Steiner H-Y, Ryals JA. Recent advances in systemic acquired resistance research- a review. Gene 1996; 179: 89-95
[3] Sticher L, Mauch-Mani, B., Metraux JP. Systemic Acquired Resistance. Ann. Rev. Phytopathol. 1997; 35: 235-270 [4] Vallad, GE, Goodman RM. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci.
2004; 44:1920–1934. [5] Gaffney T, Friedrich L, Vernooij B, Negrotto D, Nye G, Uknes S, Ward E. Requirement of Salicylic Acid for the Induction of
Systemic Acquired Resistance. Science 1993; 261: 754-756. [6] Nakashita H, Yasuda M, Nitta T, Asami T, Fujioka S, Arai Y, Sekimata K, Takatsuto S, Yamaguchi I, Yoshida S. Brassinosteroid
functions in a broad range of disease resistance in tobacco and rice. Plant. J. 2003; 33: 887–898. [7] Mauch-Mani, B., Metraux JP. Salicylic acid and systemic acquired resistance to pathogen attack, Ann Bot 1998; 82: 535-540. [8] Kessmann H, Stub T, Hofmann C, Maetzke T, Herzog J. Induction of systemic acquired resistance in plants by chemicals. Annu
Rev Phytopathol 1994;32:439–59. [9] Tally A, Oostendrop M, Lawton K, Stub T, Bassi B. Commercial development of elicitors of induced resistance to pathogens.
In:Agrawal AA, Tuzun S, Bent E, editors. Induced plant defenses against pathogens and herbivores. Minnesota: APS Press; 1999. p. 357–69.
[10] Iwata M. Probenazole—a plant defense activator. Pesticide Outlook 2001;12:28–31. [11] Gorlach J S. Volrath GK, Hengy G, Beckhove U. Benzothiadiazole, a novel class of inducers of systemic acquired resistance,
activates gene expression and disease resistance in wheat. Plant Cell 1996; 8: 629–643. [12] Kato T, Yamaguchi Y, Harano T, Yokayama T, Uyehara Y. Unsaturated hydroxy fatty acids, the self defensive substance in rice
plants against rice blast disease. Chem. Lett. 1994; 12:409–412. [13] Rohilla R, Singh US, Singh RL. Mode of action of acibenzolar-Smethyl against sheath blight of rice, caused by Rhizoctonia
solani Ku¨hn. Pest Manage Sci 2001;58:63. [14] Sekizawa Y. Recent progress in studies on non-fungicidal controlling agent, probenazole, with reference to the induced
resistance mechanism of rice plants. Rev Plant Prot Res 1980;13:114–21. [15] Uchiyama M, Abe H, Sato R, Shimura M, Watanabe T. Fate of 3-allyloxy-1,2-benzisothiazole-1,1-dioxide (Oryzematew) in rice
plants. Agric Biol Chem 1973;37:737–45. [16] Glynn CP. Induction of systemic acquired disease resistance in plants: potential implications for desease management in
urbanforestry. Journal of Arboriculture 2001; 27: 181-192. [17] Nakamura K, Akashi T, Aoki T, Kawaguchi K, Ayabe S. Induction of isoflavonoid and retrochalcone branches of the flavonoid
pathway in cultured Glycyrrhiza echinata cells treated with yeast extract. Biosci. Biotechnol. Biochem 1999; 63:1618–1620. [18] Obara N, Mitsuhara I, Seo S, Ohashi Y, Hasegawa M, Matsuura Y. Mechanism of PR gene expression by treatment of tobacco
leaves with yeast extract (AGREVO EX). Jpn. J. Phytopathol 2007; 73: 94–101. [19] Pauw B, van Duijn B, Kijne JW, Memelink J. Activation of the oxidative burst by yeast elicitor in Catharanthus roseus cells
occurs independently of the activation of genes involved in alkaloid biosynthesis. Plant. Mol. Biol 2004; 55: 797–805. [20] Doubrava NS, Dean RA, Kuc, J. Induction of systemic resistance to anthracnose caused by Colletotrichum lagenarium in
cucumber by oxalate and extracts from spinach and rhubarb leaves. Physiological and Molecular Plant Pathology 1988; 33: 69-79.
[21] Daayf F, Schmitt A, Belanger RR. Evidence of Phytoalexins in cucumber leaves infected with powdery mildew following treatment with leaf extracts of Reynoutria sachalinensis. Plant Physiology 1997; 113: 719-727.
[22] Doltsinis KS, Markellou E, Kasselaki AM, Fanouraki MN, Koumaki CM, Schmitt A, Tsakalidis AL, Malathrakis NE. Efficacy of Milsana®, a formulated plant extract from Reynoutria sachalinensis, against powdery mildew of tomato (Leveillula taurica). BioControl 2006; 51: 375-392.
[23] Fofanab B, McNallya DJ, Labbéa C, Boulangera R, Benhamoua N, Séguinb A, Bélangera RR. Milsana-induced resistance in powdery mildew-infected cucumber plants correlates with the induction of chalcone synthase and chalcone isomerase. Physiological and Molecular Plant Pathology 2002; 61: 121-132.
[24] Stavroula KD, Annegret S. Impact of treatment with plant extracts from Reynoutria sachalinensis (F. Schmidt) Nakai on intensity of powdery mildew severity and yield in cucumber under high disease pressure. Crop Protection 1998; 17: 649-656.
[25] Wurms K, Labbé C, Benhamou N, Bélanger RR. Effects of Milsana and Benzothiadiazole on the Ultrastructure of Powdery Mildew Haustoria on Cucumber. Phytopathology 1999; 89: 728-736.
[26]Rasmussen JB, Smith JA, Williams S, Burkhart W, Ward ER, Somerville SC, Ryals, J, Hammerschmidt R. cDNA Cloning and systemic expression of an acidic peroxidase associated with systemic acquired resistance to disease in cucumber. Physiol. Mol. Plant Pathol. 1995; 46: 389-400.
[27] Shiomi S, Inaba A. Partial cDNA sequence of Cucumis sativus (cucumber) actin gene. 1998; Journal Published Only in DataBase.
734 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

[28] Narusaka,Y, Narusaka M, Horio T, Ishii H. Comparison of local and systemic induction acquired disease resistance in cucumber [Cucumis sativus] plants treated with benzothiadiazoles or salicylic acid. Plant Cell Physiol 1999; 40: 388-395.
[29] Abdolkarim C, Fatemeh A, Hoseinhaj H. Study on antibacterial effect of some Allium species from Hamedan-Iran. International Journal of Agriculture and Biology 2007; 9: 873–876.
[30] : Amin M, Montazeri EA, Mashhadizadeh MA, Sheikh AF. Characterization of shallot, an antimicrobial extract of Allium ascalonicum. Pakistan Journal of Medical Sciences 2009; 25: 948-952.
[31] Benkeblia N. Antimicrobial activity of essential oil extracts of various onions (Allium cepa) and garlic (Allium sativum). Yeast 2004; 37: 263-268.
[32] Harris JC, Cottrell S, Plummer S, Lloyd D. Antimicrobial properties of Allium sativum (garlic) . Applied Microbiology and Biotechnology 2001; 57: 282-286.
[33] Graham TL, Graham MY. Cellular coordination of molecular responses in plant defense. Mol. Plant-Microbe Interact 1991; 4: 415-422.
[34] Rasmussen JB, Smith JA, Williams S, Burkhart W, Ward ER, Somerville SC, Ryals, J, Hammerschmidt R. cDNA Cloning and systemic expression of an acidic peroxidase associated with systemic acquired resistance to disease in cucumber. Physiol. Mol. Plant Pathol 1995; 46: 389-400.
[35] Zhang D, Han WY, Dick WA, Davis KR, Hoitink HAJ. Compost and compost water extract-induced systemic acquired resistance in cucumber and Arabidopsis. Phytopathol 1998; 88: 450-455.
[36] Felix G, Regenass M, Boller T. Specific perception of subnanomolar concentrations of chitin fragments by tomato cells: induction of extracellular alkalinization, change in protein phosphorylation, and establishment of a refractory state. Plant J 1993; 4: 307-316.
[37] Kaku H., Shibuya N, Xu P, Aryan AP, Fincher GB. N- acetylchitooligosaccharides elicit expression of a single (1→3)- β- gulucanase gene in suspension- cultured cells from barley (Hordeum vulgare), Physiol. Plantarum 1997; 100: 111-118.
[38] Yamaguchi T, Minami E, Ueki J, Shibuya N. Elicitor- induced activation of phospholipases plays an important role for the induction of defense responses in suspension- cultured rice cells. Plant Cell Physiol 2005; 46: 579-587.
[39] [23] Koike N, Hyakumachi M, Kageyama K, Tsuyumu S, Doke N. Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi: lignification and superoxide generation. Eur J Plant Pathol 2001;107:523–33.
[40]Roe NE., Stoffella PJ. Bryan, HH. Utilization of MSW compost and other organic mulches on commercial vegetable crops. Compost Sci Util 1993; 1:73-84.
[41] Hoitink HAJ, Boehm MJ. Biocontrol within the context of soil microbial communities: A substrate- dependent phenomenon. Annu. Rev. Phytopathol 1999; 37: 427-446
[42] Maurhofer M, Hase C, Meuwly P, Métraux, JP, Défago G. Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strain CHA0: Influence of the gacA gene and pyoverdine production. Phytopathol 1994; 84:139-146.
[43] Leeman M, van Pelt JA, Hendrickx MJ, Scheffer RJ, Bakker PAHM, Schippers B. Biocontrol of Fusarium wilt of radish in commercial greenhouse trials by seed treatment with Pseudomonas fluorescens WCS374. Phytopathol 1995; 85:1301-1305.
[44] Pieterse CMJ, van Wees SCM, Hoffland E, van Pelt JA, van Loon LC. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 1996; 8 :1225-1237.
[45] Zhang D, Han WY, Dick WA, Davis KR, Hoitink HAJ. Compost and compost water extract-induced systemic acquired resistance in cucumber and Arabidopsis. Phytopathol 1998; 88: 450-455.
[46] Khan J, Ooka JJ, Miller SA, Madden LV Hoitink HAJ. Systemic resistance induced by Trichoderma hamatum 382 in cucumber against phytophthora crown rot and leaf blight. Plant Disease 2004; 88: 280-286.
735©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________