Embed Size (px)
Citation preview

Development 104, 669-681 (1988)Printed in Great Britain © The Company of Biologists Limited 1988
669
Induction by acid load of the maturation of prestalk cells in Dictyostelium
discoideum
KEIINOUYE
Department of Botany, Faculty of Science, Kyoto University, Kyoto 606, Japan
Summary
During the process of fruiting body construction in thecellular slime mould Dictyostelium discoideum,prestalk cells become mature stalk cells in a well-controlled manner. To identify the natural inducer ofstalk cell maturation, substances known to inducestalk cell differentiation under in vitro conditions, andsome other related compounds, were examined fortheir effects in vivo on migrating slugs, the precursorstructures of the fruiting bodies.
Among these substances, addition of weak acidssuch as CO2, and addition followed by removal ofweak bases such as NH3, strikingly induced thematuration of prestalk cells in situ in slugs. On theother hand, inhibitors of the plasma membrane protonpump did not efficiently induce the maturation ofprestalk cells in intact slugs. Differentiation inducingfactor (DIF), an endogenous inducer of prestalk differ-
entiation, seemed to be an even poorer inducer of stalkcell maturation when applied to intact slugs.
The activities of these substances in inducing stalkcell maturation showed a good correlation with theireffects on the cytoplasmic pH (pH,) of prestalk cells;the larger the pH| drop, the stronger the induction ofstalk cell maturation, suggesting a requirement for apH| decrease for the maturation of prestalk cells.Based on these results, it was proposed that stalk celldifferentiation, which is induced by DIF, is blockedhalfway during normal development by (an) agent(s)that prevent(s) the decrease in pl^.
Key words: cytoplasmic pH, cell differentiation,morphogenesis, Dictyostelium discoideum, cellular slimemould.
Introduction
The fruiting body of the cellular slime mould Dictyo-stelium discoideum consists of a spherical mass ofspores and a supporting stalk, the latter being com-posed of vacuolated cells with cell walls. The fruitingbody is formed from a mass of amoeboid cells (thepseudoplasmodium, or 'slug') in which the precursorcell types of spores and stalk cells are discernible,namely prespore cells and prestalk cells (Bonner,1967).
During the process of fruiting body construction,prestalk cells first turn into stalk cells at the apicalregion of the slug, and the stalk elongates by continu-ously adding new stalk cells to its tip (Bonner, 1952).In other words, prestalk cells become mature stalkcells in an orderly manner, suggesting the presence ofa mechanism that controls stalk cell maturation.
Since the discovery that starved D. discoideum cellscan be induced by cAMP to become mature stalk cells
(Bonner, 1970), this nucleotide has been suggested bysome workers to be the natural inducer of stalk celldifferentiation. Ammonia, on the other hand, in-hibits stalk cell differentiation (Gross et al. 1981,1983) and also prevents fruiting body formation(Schindler & Sussman, 1977). In a model proposed bySussman & Schindler (1978), antagonism betweencAMP and NH3 was supposed to play the key role inthe control of stalk cell differentiation. The findingthat ammonia inhibits the intracellular accumulationof cAMP (Schindler & Sussman, 1979) was taken asevidence for this hypothesis. However, the recentfinding by Berks & Kay (1988) that cAMP inhibitssome step in stalk cell differentiation threw doubt onits originally claimed role in stalk cell maturation.
Other than inhibiting the accumulation of cAMP,ammonia has the effect of elevating cytoplasmic pH(pH,, Inouye, 1988) and the pH of intracellular acidicvesicles (pHv, Yamamoto & Takeuchi, 1983), sugges-ting the possibility that pH, and pHv may play, in

670 K. Inouye
addition to their proposed role in the choice betweenspore and stalk pathways (Gross et al. 1983, 1988;Inouye, 1985), an important role in the control ofstalk cell maturation. To examine this possibility, Iinvestigated in this study the effects of acid load onmigrating slugs of D. discoideum. Carbon dioxide andammonia proved to be particularly useful for investi-gating the effects of weak acid and weak base,respectively, on intact migrating slugs, because theypermeate efficiently into the tissue without the needof physically disturbing the slug structure. The effectsof other weak acids and bases, as well as of proton-pump inhibitors and DIF, were also investigated. Theresults obtained clearly indicate that cytoplasmicacidification induces the maturation of prestalk cellsto become stalk cells. Based on these and otherresults, a possible mechanism for the control of stalkcell maturation during normal development will bediscussed.
Materials and methods
Organisms and cultureDictyostelium discoideum, V12M2 and NC4 (wild-typestrains), were used. They were grown on Klebsiella aero-genes using a modified SM medium (4-4g KH2PO4, 2-0 gNa2HPO4, 1-0 gMgSO4-7H2O, 7-5 g glucose, 10-0gBactopeptone (Difco), l-0g yeast extract (Difco), 15 g agar, 11H2O). Cells about to clear the bacterial lawn were collectedand washed in 20 mM-potassium phosphate buffer, pH6-0,(PB) by repeated centrifugation.
Application of CO2
A thick suspension of washed vegetative cells (approx.1-6X108 cells ml"1 in PB supplemented with 2mM-MgSO4)was deposited on agar substrata (3 ml of 1-5 % bacto agar,Difco, USA, in H2O) made in 50 ml tissue culture flasks(Becton Dickinson, USA). Each flask received a spot of10/d cell suspension in the centre. After the cells hadsettled on the agar substrata, excess fluid was removed. Theflasks were then capped and incubated at 21°C in the dark.After approx. 24 h, when slugs had moved away from theoriginal spot, the cap of each flask was replaced with asilicon double stopper, and the CO2 concentration in theflask was adjusted in such a way that the O2 concentrationremained unchanged at 21 %. Usually, x % CO2 (initialconcentration) was prepared by withdrawing 18-5 ml airfrom the flask, followed by an addition of (15-0-75*) ml air,O-l&rml O2 and 0-6xml CO2. The gasses had been satu-rated with H2O. The air space of the flask with 3 ml agarand a double stopper was 60-0 ml. Plastic syringes wereused for withdrawing and adding the gases. The withdrawalof air caused decreases in the pressure (-0-25 atm) and inthe air-space volume (—4-5 ml) of the flasks, which weretaken into consideration in calculating the volumes of thegasses to be added. Using the solubility of CO2 in water at21 °C (0-8541CO2 in 11 H2O at 1 atm CO2), the actual CO2
concentration was calculated to be 0-957 x [initial concen-
tration]. In the text, CO2 concentrations are given as theinitial concentrations. In some experiments, CO2 wasremoved by perfusing approx. 5 litres air using an aspiratorafter the indicated periods of exposure.
Removal of ammoniaSlugs (1-day-old) migrating on plain agar plates (4 ml of1-5% water agar in 5 cm plastic dishes) were exposed toNH3 by placing the plates (after removing the lids) in plasticboxes (3061) containing 0-51 ammonia source solution.The boxes were then tightly sealed. The ammonia sourcewas made by adding 0-7 ml of 1 M-NH4CI to 0-51 of 0-5 M-NaOH. The partial pressures of ammonia in the air spacewere calculated from the solubility constant of NH3 at 21 °C(Denmead et al. 1982) and the concentrations of NH3 in theammonia source solutions. The latter were measured usingan ammonia electrode (Horiba, Japan). After an exposureto NH3 for the indicated times, the plates were transferredto a box containing 1-51 solution of neutralized bromothy-mol blue. Decrease in the NH3 concentration of theammonia source solution during this procedure was negli-gible. The same method was used for trimethylamine byreplacing NRjCl with trimethylamine chloride.
Observation and scoring of the resultant structuresSlugs treated with CO2 or NH3 were further incubated for24-40h at 21 °C in the dark. Resultant structures formedwere then observed using a dissecting microscope (Olym-pus, Japan). They were classified into several groupsaccording to their morphology, as indicated in the figurelegends, and the number of individuals of each group wasscored. Whole-mount specimens and squashed specimensof representative samples of each group have been exam-ined at higher magnifications using phase-contrast andNomarski microscopy. Sections of fixed samples of somegroups have also been examined. For visualizing the cellwall, specimens were immersed in 0-1% Calcofluor whiteST (American Cyanamide, USA) solution in PB andobserved using a fluorescence microscope (excitation405nm, emission >470nm, Nikon, Japan; Harrington &Raper, 1968).
Agar-sandwich method1-day-old slugs were transferred onto agar plates containing20 mM-potassium phosphate buffer, at the indicated pH,and the substances to be tested. About half of the slugswere dissected lengthwise at a position about one fifth fromthe tip, and each fragment was dissociated separately with ahair loop. Then the slugs, both intact and dissected, werecovered with coverslips. The plates were incubated at 21 °Cin the dark. After approx. 24 h incubation, the plates wereexamined using a phase-contrast microscope. The cover-slips were then carefully lifted off the agar surface (almostall the cells adhered to the coverslips), placed on a glassslide carrying a drop of 0-1% Calcofluor and observed asdescribed above.
Agar-plate method1-day-old slugs were transferred onto agar plates containingthe indicated amounts of the test substances. The plateswere then incubated in unidirectional light at 21CC. Indue-

Stalk cell maturation in D. discoideum 671
tion of fruiting by each added substance was estimated bycounting the numbers of fruiting bodies and migrating slugs20 h later.
Other methodsFor neutral red staining, washed vegetative cells wereincubated in 0-005% neutral red solution in PB for lOminat 0°C. Tetramethylrhodamine staining and grafting ofslugs were carried out as described previously (Akiyama &Inouye, 1987). The effects of CO2 and NH3 on the pH of theagar substratum were assessed by measuring the pH of a20mM-KCl solution using a glass pH electrode underexactly the same conditions as in the experimentals.
ChemicalsDIF-1 (natural and synthetic, giving the same results) waskindly provided by Dr R. Kay (MRC, Cambridge, Eng-land). DES and miconazole were purchased from Sigma (StLouis, USA), and zearalenone from Mikor Chemicals(Jerusalem, Israel). CO2 (over 99-9% purity) and otherchemicals were obtained from local suppliers).
Results
Effects of gasses(1) Effects of CO2
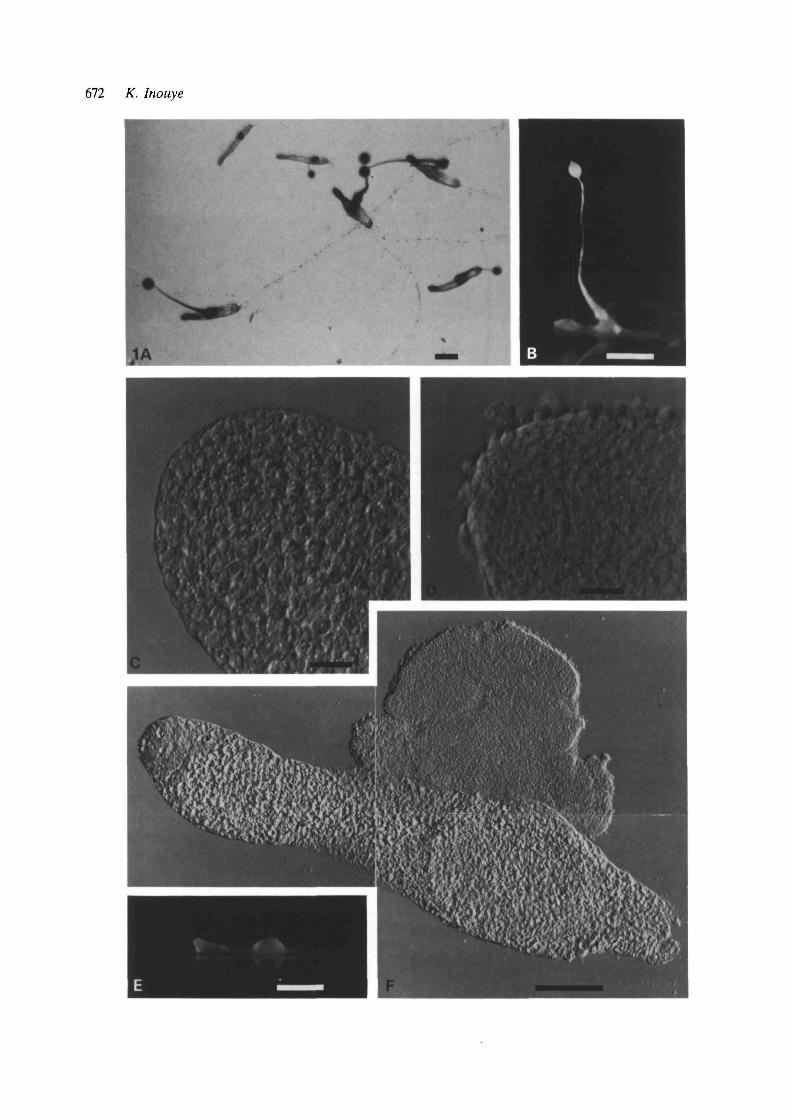
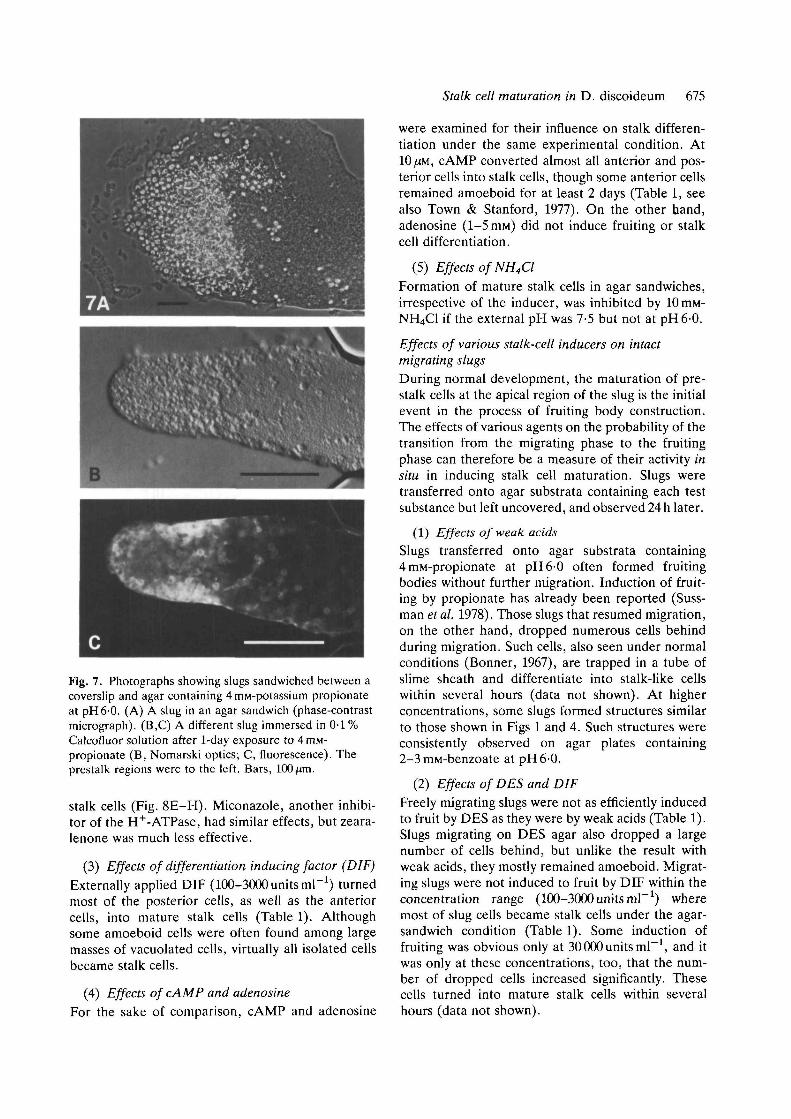
When migrating slugs were exposed to 10% CO2,they immediately stopped migrating and all the pre-stalk cells, and the majority of prestalk-like cells inthe posterior region (see below), became maturestalk cells. (In this paper, no distinction will be madebetween stalk cell and basal disk cell.) Meanwhile,the prespore cells of each slug underwent regulationto make one or more normal fruiting bodies, consist-ing of mature spores and stalk cells, within 24 h(Fig. 1A-D). The fates of both precursor cell typesafter a CO2 exposure were revealed by prestalk-specific staining with neutral red (Bonner, 1952;Sternfeld & David, 1981); while many of the maturestalk cells formed within the original slug had largered granules, most cells in the stalk of the fruitingbody structures did not (Fig. 2). Experiments usingchimeric slugs with a rhodamine-labelled prestalkregion also indicated that only a small fraction of theoriginal prestalk cells contributed to the stalk of thefruiting body structures (data not shown). Time-lapsefilms of the entire process revealed that the cells in theposterior region resumed mobility within about 12 hand formed a fruiting body, while the anterior cellsremained immobile throughout.
When CO2 (10%) was removed after a 10-minexposure, no stalk cell mass was formed. However, a20-min exposure to 10 % CO2 was sufficient for stalkcell induction. In such a case, the anterior region ofthe slugs became a mass of mature stalk cells, just asin the case of a continuous exposure to CO2. On theother hand, prespore cells, instead of making fruiting
body structures, reorganized themselves to form anew slug which resumed migration. A similar situ-ation occurred with some of the slugs that had beencontinuously exposed to 5-10% CO2. At lowerconcentrations of CO2 (1-5%), only the normalfruiting body formation was induced. When the CO2
concentration exceeded 10%, some slugs failed toform the fruiting body structures, giving rise to slug-shaped masses of stalk cells with some amoeboid cellshidden inside (Fig. 1E-F). When the CO2 concen-tration was very high, some cells, especially those inthe tip region, lysed. The effects of various concen-trations of CO2 on migrating slugs are summarized inFig. 3.
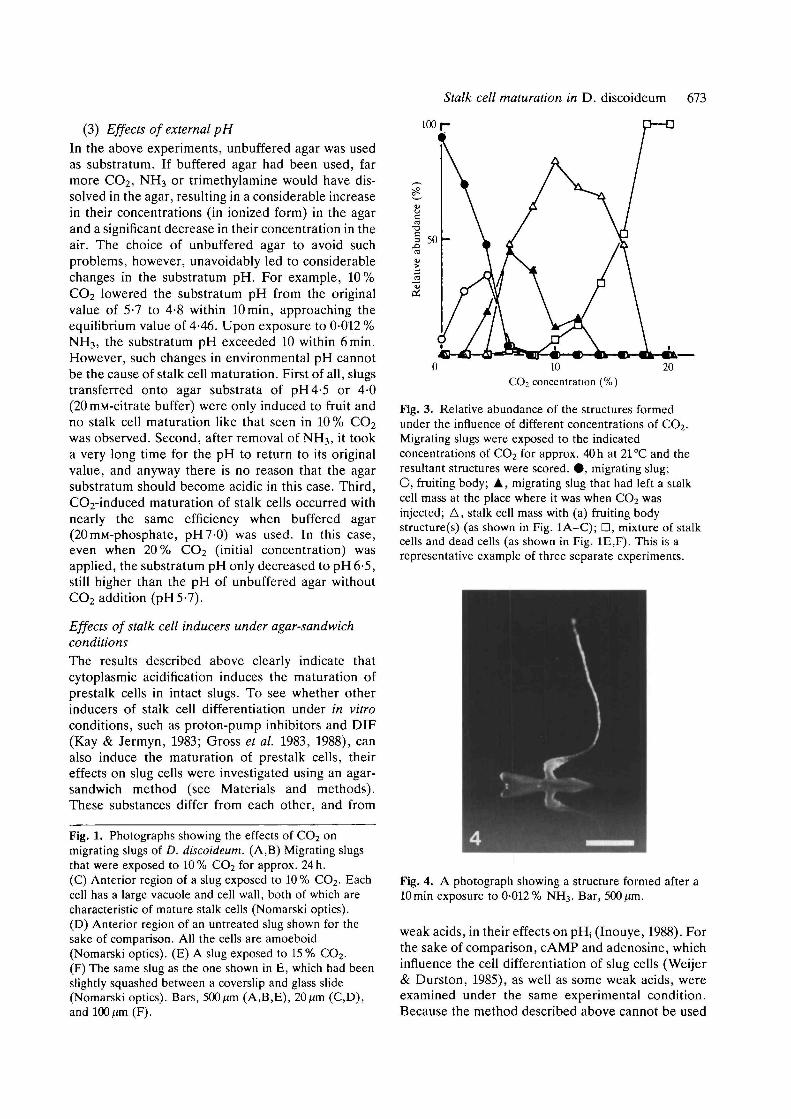
(2) Effects of ammonia and trimethylamineAn exposure to CO2 causes a transient decrease in thepHj of slug cells (Inouye, 1988), which raises thepossibility that cytoplasmic acidification induces thematuration of prestalk cells to become true stalk cells.Supporting this is the observation that simultaneouslyadded ammonia nullified the effects of CO2 (data notshown). To examine further this possibility, migratingslugs were first exposed to NH3 and then transferredinto an atmosphere without NH3. (pHj can also bedecreased by removing weak base, Inouye, 1988).Meanwhile, the relative humidity was kept at 100%.Upon removal of NH3, the slugs stopped migration.Subsequently, if the NH3 concentration and thelength of the exposure were appropriate, prestalkcells became mature stalk cells while prespore cellsformed fruiting bodies, giving rise to exactly the samestructures as those formed in 10% CO2. Fig. 4 showsan example of the structure formed after a lOminexposure to 0-012% NH3. At higher concentrationsof NH3, many of the cells in the slug lysed, whereas atlower concentrations (0-002-0-005%), normal fruit-ing body formation was induced (Fig. 5).
If cytoplasmic acidification is the cause of stalk cellmaturation, the effect of NH3 removal is expected tovary depending on the length of the exposure to NH3,because the degree of acid load by NH3 removalshould become larger if the length of the exposure isprolonged (Roos & Boron, 1981). Indeed, increasingthe period of exposure to the same concentration ofNH3 resulted in correspondingly more pronouncedeffects on slugs (Fig. 6). If NH3 was not removed butpresent continuously, all the slug cells lysed even at aconcentration of 0-002 %.
Addition followed by removal of trimethylamine,another weak base, also induced the formation offruiting bodies and the maturation of prestalk cells insitu. As in the case of NH3 removal, the magnitude ofits effects was dependent on the length of the ex-posure period (data not shown).
672 K. Inouye
Stalk cell maturation in D. discoideum 673
(3) Effects of external pH
In the above experiments, unbuffered agar was usedas substratum. If buffered agar had been used, farmore CO2, NH3 or trimethylamine would have dis-solved in the agar, resulting in a considerable increasein their concentrations (in ionized form) in the agarand a significant decrease in their concentration in theair. The choice of unbuffered agar to avoid suchproblems, however, unavoidably led to considerablechanges in the substratum pH. For example, 10%CO2 lowered the substratum pH from the originalvalue of 5-7 to 4-8 within lOmin, approaching theequilibrium value of 4-46. Upon exposure to 0-012 %NH3, the substratum pH exceeded 10 within 6min.However, such changes in environmental pH cannotbe the cause of stalk cell maturation. First of all, slugstransferred onto agar substrata of pH4-5 or 4-0(20 mM-citrate buffer) were only induced to fruit andno stalk cell maturation like that seen in 10 % CO2
was observed. Second, after removal of NH3, it tooka very long time for the pH to return to its originalvalue, and anyway there is no reason that the agarsubstratum should become acidic in this case. Third,CO2-induced maturation of stalk cells occurred withnearly the same efficiency when buffered agar(20mM-phosphate, pH7-0) was used. In this case,even when 20% CO2 (initial concentration) wasapplied, the substratum pH only decreased to pH6-5,still higher than the pH of unbuffered agar withoutCO2 addition (pH5-7).
Effects of stalk cell inducers under agar-sandwichconditions
The results described above clearly indicate thatcytoplasmic acidification induces the maturation ofprestalk cells in intact slugs. To see whether otherinducers of stalk cell differentiation under in vitroconditions, such as proton-pump inhibitors and DIF(Kay & Jermyn, 1983; Gross et al. 1983, 1988), canalso induce the maturation of prestalk cells, theireffects on slug cells were investigated using an agar-sandwich method (see Materials and methods).These substances differ from each other, and from
Fig. 1. Photographs showing the effects of CO2 onmigrating slugs of D. discoideum. (A,B) Migrating slugsthat were exposed to 10% CO2 for approx. 24 h.(C) Anterior region of a slug exposed to 10 % CO2. Eachcell has a large vacuole and cell wall, both of which arecharacteristic of mature stalk cells (Nomarski optics).(D) Anterior region of an untreated slug shown for thesake of comparison. All the cells are amoeboid(Nomarski optics). (E) A slug exposed to 15 % CO2.(F) The same slug as the one shown in E, which had beenslightly squashed between a coverslip and glass slide(Nomarski optics). Bars, 500^m (A,B,E), 20/an (C,D),and 100 f<m (F).
100
20CO2 concentration (%)
Fig. 3. Relative abundance of the structures formedunder the influence of different concentrations of CO2.Migrating slugs were exposed to the indicatedconcentrations of CO2 for approx. 40 h at 21 °C and theresultant structures were scored. # , migrating slug;O, fruiting body; A, migrating slug that had left a stalkcell mass at the place where it was when CO2 wasinjected; A, stalk cell mass with (a) fruiting bodystructure(s) (as shown in Fig. 1A-C); D, mixture of stalkcells and dead cells (as shown in Fig. 1E,F). This is arepresentative example of three separate experiments.
Fig. 4. A photograph showing a structure formed after alOmin exposure to 0-012% NH3. Bar, 500/an.
weak acids, in their effects on pH; (Inouye, 1988). Forthe sake of comparison, cAMP and adenosine, whichinfluence the cell differentiation of slug cells (Weijer& Durston, 1985), as well as some weak acids, wereexamined under the same experimental condition.Because the method described above cannot be used
674 K. Inouye
100
0-005 0-010 0-015NH3 concentration (%)
Fig. 5. Relative abundance of the structures formed aftera lOmin exposure to different concentrations of NH3 gas.1-day-old slugs were exposed to the indicatedconcentrations of NH3 for lOmin, then incubated forapprox. 40 h at 21 °C, and the resultant structures werescored. Since, in this case, most slugs that were exposedto 0 % NH3 formed fruiting bodies during the incubationperiod after NH3 removal, the following grouping wasadopted; # , migrating slug that was not induced to fruitimmediately after the removal of NH3; O, migrating slugthat was induced to fruit upon removal of NH3;T, abnormal fruiting body; A, stalk cell mass with (a)fruiting body structure(s) (as shown in Fig. 4); D, mixtureof stalk cells and dead cells, with increasing ratio of deadcells as NH3 concentration rose. In the transition zone(0-004%-0-008%), the distinction between normalfruiting body, abnormal fruiting body and stalk cell masswith fruiting body structures was not always very clear.This is a representative example of three separateexperiments.
for these nonvolatile substances, they were includedin the agar substrata.
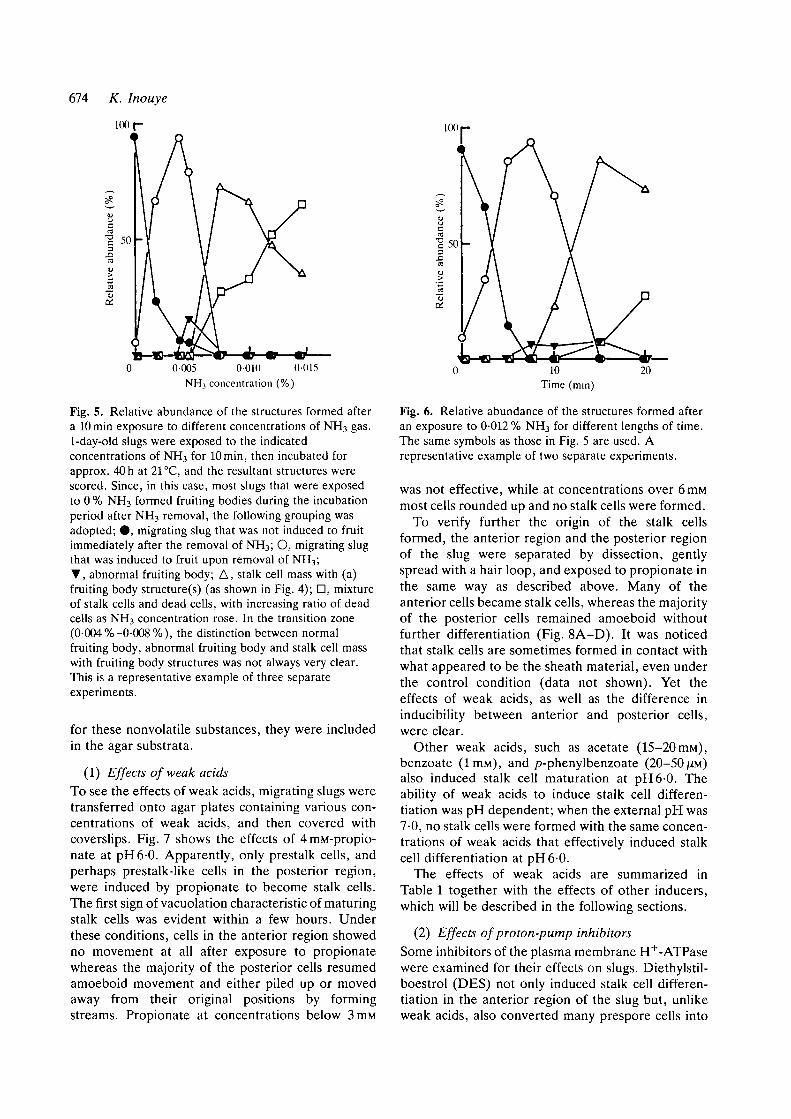
(1) Effects of weak acidsTo see the effects of weak acids, migrating slugs weretransferred onto agar plates containing various con-centrations of weak acids, and then covered withcoverslips. Fig. 7 shows the effects of 4mM-propio-nate at pH6-0. Apparently, only prestalk cells, andperhaps prestalk-like cells in the posterior region,were induced by propionate to become stalk cells.The first sign of vacuolation characteristic of maturingstalk cells was evident within a few hours. Underthese conditions, cells in the anterior region showedno movement at all after exposure to propionatewhereas the majority of the posterior cells resumedamoeboid movement and either piled up or movedaway from their original positions by formingstreams. Propionate at concentrations below 3mM
10Time (min)
20
Fig. 6. Relative abundance of the structures formed afteran exposure to 0-012 % NH3 for different lengths of time.The same symbols as those in Fig. 5 are used. Arepresentative example of two separate experiments.
was not effective, while at concentrations over 6mMmost cells rounded up and no stalk cells were formed.
To verify further the origin of the stalk cellsformed, the anterior region and the posterior regionof the slug were separated by dissection, gentlyspread with a hair loop, and exposed to propionate inthe same way as described above. Many of theanterior cells became stalk cells, whereas the majorityof the posterior cells remained amoeboid withoutfurther differentiation (Fig. 8A-D). It was noticedthat stalk cells are sometimes formed in contact withwhat appeared to be the sheath material, even underthe control condition (data not shown). Yet theeffects of weak acids, as well as the difference ininducibility between anterior and posterior cells,were clear.
Other weak acids, such as acetate (15-2benzoate (IITIM), and p-phenylbenzoate (20-5also induced stalk cell maturation at pH6-0. Theability of weak acids to induce stalk cell differen-tiation was pH dependent; when the external pH was7-0, no stalk cells were formed with the same concen-trations of weak acids that effectively induced stalkcell differentiation at pH6-0.
The effects of weak acids are summarized inTable 1 together with the effects of other inducers,which will be described in the following sections.
(2) Effects of proton-pump inhibitorsSome inhibitors of the plasma membrane H+-ATPasewere examined for their effects on slugs. Diethylstil-boestrol (DES) not only induced stalk cell differen-tiation in the anterior region of the slug but, unlikeweak acids, also converted many prespore cells into

Fig. 2. Photographs showing the origin of the stalk cells constituting the stalk of the fruiting body structures formed in10 % CO2. Cells were vitally stained with neutral red and allowed to form slugs. Slugs that had been migrating for 2days were exposed to 10% CO2. (A) Part of a slug exposed to 10% CO2 for approx. 24 h, slightly squashed between acoverslip and glass slide. The lower part of the stalk is seen in the upper left extruding from the slug body. Most cellsseen in this picture actually have large vacuoles and a cell wall, although neutral red proved to be slightly inhibitory forstalk cell differentiation (data not shown). (B) Anterior region of a slug exposed to CO2. (C) Part of the stalk of thefruiting body structure formed from the slug shown in B. Also shown, for the sake of comparison, are (D) part of thestalk of a fruiting body formed under normal conditions; and (E) part of the stalk of a fruiting body formed from aposterior isolate of a slug that had been depleted of anterior-like cells by means of two successive cuts as described inSternfeld & David (1982). All samples were derived from the same batch of vitally stained cells. Whereas the stalk cellsshown in D most likely derived from prestalk cells which contained neutral red granules, the stalk cells shown in Eoriginated from initially prespore cells that had undergone transdifferentiation after the removal of the prestalk region.Vitally stained prespore cells, although still possessing the dye, usually do not exhibit red granules even when they haveconverted to prestalk or stalk cells. Bar, 100 jan for A and 50/on for B-E.
Stalk cell maturation in D. discoideum 675
Fig. 7. Photographs showing slugs sandwiched between acoverslip and agar containing 4 miu-potassium propionateat pH6-0. (A) A slug in an agar sandwich (phase-contrastmicrograph). (B,C) A different slug immersed in 0-1%Calcofluor solution after 1-day exposure to 4mM-propionate (B, Nomarski optics; C, fluorescence). Theprestalk regions were to the left. Bars, 100 ^m.
stalk cells (Fig. 8E-H) . Miconazole, another inhibi-tor of the H+-ATPase, had similar effects, but zeara-lenone was much less effective.
(3) Effects of differentiation inducing factor (DIF)Externally applied DIF (100-3000 units ml"1) turnedmost of the posterior cells, as well as the anteriorcells, into mature stalk cells (Table 1). Althoughsome amoeboid cells were often found among largemasses of vacuolated cells, virtually all isolated cellsbecame stalk cells.
(4) Effects of cAMP and adenosine
For the sake of comparison, cAMP and adenosine
were examined for their influence on stalk differen-tiation under the same experimental condition. At10 (XM, cAMP converted almost all anterior and pos-terior cells into stalk cells, though some anterior cellsremained amoeboid for at least 2 days (Table 1, seealso Town & Stanford, 1977). On the other hand,adenosine (1-5 ITIM) did not induce fruiting or stalkcell differentiation.
(5) Effects of NH4Cl
Formation of mature stalk cells in agar sandwiches,irrespective of the inducer, was inhibited by 10 ITIM-NH4CI if the external pH was 7-5 but not at pH6-0.
Effects of various stalk-cell inducers on intactmigrating slugs
During normal development, the maturation of pre-stalk cells at the apical region of the slug is the initialevent in the process of fruiting body construction.The effects of various agents on the probability of thetransition from the migrating phase to the fruitingphase can therefore be a measure of their activity insitu in inducing stalk cell maturation. Slugs weretransferred onto agar substrata containing each testsubstance but left uncovered, and observed 24 h later.
(1) Effects of weak acids
Slugs transferred onto agar substrata containing4 mM-propionate at pH6-0 often formed fruitingbodies without further migration. Induction of fruit-ing by propionate has already been reported (Suss-man et al. 1978). Those slugs that resumed migration,on the other hand, dropped numerous cells behindduring migration. Such cells, also seen under normalconditions (Bonner, 1967), are trapped in a tube ofslime sheath and differentiate into stalk-like cellswithin several hours (data not shown). At higherconcentrations, some slugs formed structures similarto those shown in Figs 1 and 4. Such structures wereconsistently observed on agar plates containing2-3mM-benzoate at pH6-0.
(2) Effects of DES and DIF
Freely migrating slugs were not as efficiently inducedto fruit by DES as they were by weak acids (Table 1).Slugs migrating on DES agar also dropped a largenumber of cells behind, but unlike the result withweak acids, they mostly remained amoeboid. Migrat-ing slugs were not induced to fruit by DIF within theconcentration range (100-3000 units ml"1) wheremost of slug cells became stalk cells under the agar-sandwich condition (Table 1). Some induction offruiting was obvious only at 30 000 units m P ' , and itwas only at these concentrations, too, that the num-ber of dropped cells increased significantly. Thesecells turned into mature stalk cells within severalhours (data not shown).
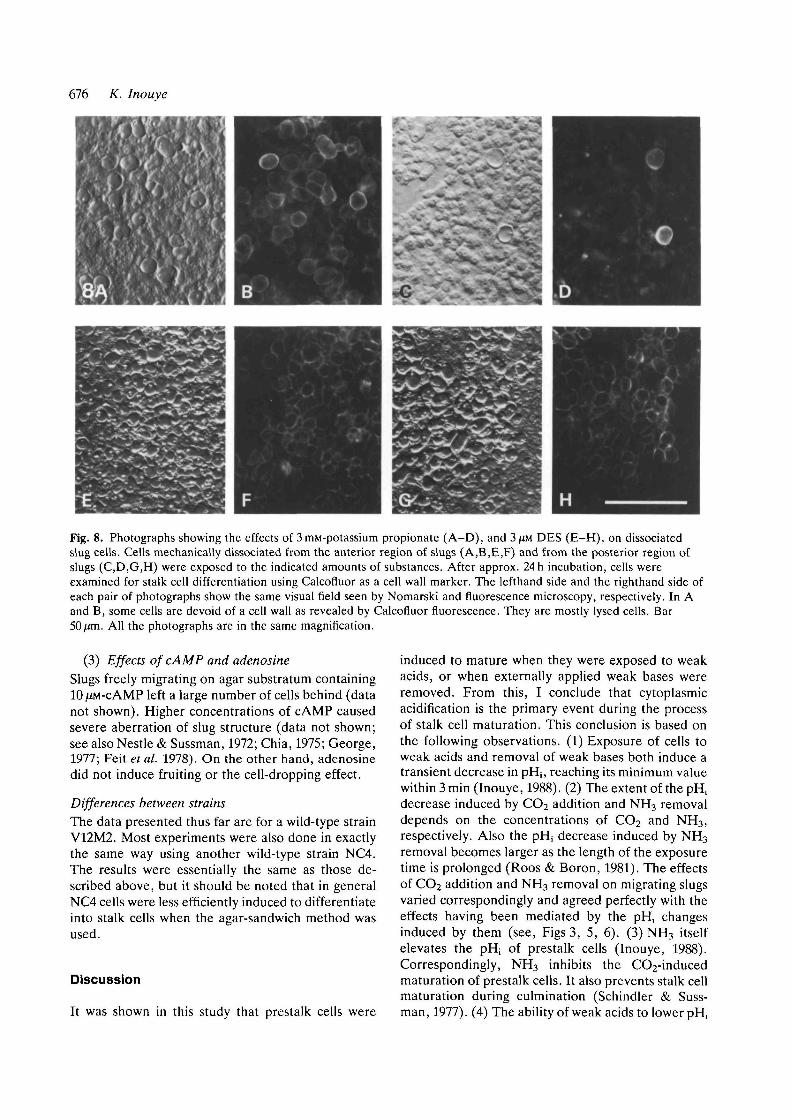
676 K. Inouye
Fig. 8. Photographs showing the effects of 3mM-potassium propionate (A-D), and 3JJM DES (E-H), on dissociatedslug cells. Cells mechanically dissociated from the anterior region of slugs (A,B.E,F) and from the posterior region ofslugs (C,D,G,H) were exposed to the indicated amounts of substances. After approx. 24h incubation, cells wereexamined for stalk cell differentiation using Calcofluor as a cell wall marker. The lefthand side and the righthand side ofeach pair of photographs show the same visual field seen by Nomarski and fluorescence microscopy, respectively. In Aand B, some cells are devoid of a cell wall as revealed by Calcofluor fluorescence. They are mostly lysed cells. Bar
All the photographs are in the same magnification.
induced to mature when they were exposed to weakacids, or when externally applied weak bases wereremoved. From this, I conclude that cytoplasmicacidification is the primary event during the processof stalk cell maturation. This conclusion is based onthe following observations. (1) Exposure of cells toweak acids and removal of weak bases both induce atransient decrease in pH,, reaching its minimum valuewithin 3 min (Inouye, 1988). (2) The extent of the pH,decrease induced by CO2 addition and NH3 removaldepends on the concentrations of CO2 and NH3,respectively. Also the pHj decrease induced by NH3
removal becomes larger as the length of the exposuretime is prolonged (Roos & Boron, 1981). The effectsof CO2 addition and NH3 removal on migrating slugsvaried correspondingly and agreed perfectly with theeffects having been mediated by the pH, changesinduced by them (see, Figs 3, 5, 6). (3) NH3 itselfelevates the pHj of prestalk cells (Inouye, 1988).Correspondingly, NH3 inhibits the CO2-inducedmaturation of prestalk cells. It also prevents stalk cellmaturation during culmination (Schindler & Suss-man, 1977). (4) The ability of weak acids to lower pH,
(3) Effects of cAMP and adenosineSlugs freely migrating on agar substratum containingIOJIM-CAMP left a large number of cells behind (datanot shown). Higher concentrations of cAMP causedsevere aberration of slug structure (data not shown;see also Nestle & Sussman, 1972; Chia, 1975; George,1977; Feit et al. 1978). On the other hand, adenosinedid not induce fruiting or the cell-dropping effect.
Differences between strainsThe data presented thus far are for a wild-type strainV12M2. Most experiments were also done in exactlythe same way using another wild-type strain NC4.The results were essentially the same as those de-scribed above, but it should be noted that in generalNC4 cells were less efficiently induced to differentiateinto stalk cells when the agar-sandwich method wasused.
Discussion
It was shown in this study that prestalk cells were
Stalk cell maturation in D. discoideum 677
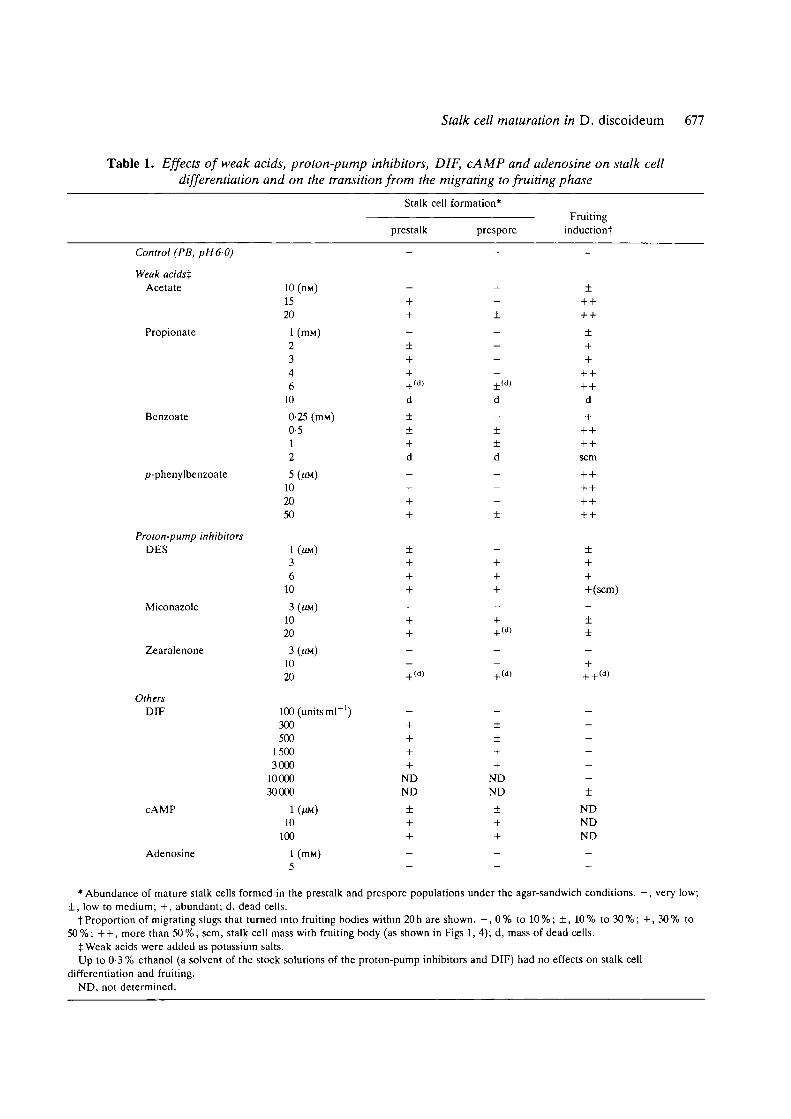
Table 1. Effects of weak acids, proton-pump inhibitors, DIF, cAMP and adenosine on stalk celldifferentiation and on the transition from the migrating to fruiting phase
Stalk cell formation*
prestalk prespore
Fruitinginductiont
Control (PB, pH6-0)
Weak acidstAcetate
Propionate
Benzoate
p-phenylbenzoate
Proton-pump inhibitorsDES
Miconazole
Zearalenone
OthersDIF
cAMP
Adenosine
10 (nM)1520
1 (ITIM)
2346
10
0-25 (mM)0-512
5 (MM)
102050
1O036
10
3 (^M)
1020
3 (MM)
1020
100 (units m P 1
300500
15003000
1000030000
1(MM)10
100
1 (mM)5
d
±±
++
±+++
++
NDND
d
±±d
+
k(d)
±±
NDND
(scm)
±±
±NDNDND
•Abundance of mature stalk cells formed in the prestalk and prespore populations under the agar-sandwich conditions. —, very low;± , low to medium; + , abundant; d, dead cells.
t Proportion of migrating slugs that turned into fruiting bodies within 20 h are shown. - , 0 % to 10 %; ± , 10 % to 30 %; + , 30 % to50%; + + , more than 50%; scm, stalk cell mass with fruiting body (as shown in Figs 1, 4); d, mass of dead cells.
tWeak acids were added as potassium salts.Up to 0-3 % ethanol (a solvent of the stock solutions of the proton-pump inhibitors and DIF) had no effects on stalk cell
differentiation and fruiting.ND, not determined.

678 K. Inouye
and their ability to induce stalk cell maturation areboth dependent on the external pH, being moreeffective at lower external pHs.
Williams et al. (1984) have shown that weak acidsprevent the secretion of cAMP whereas ammoniasuppresses its production, thus having opposite ef-fects on the intracellular level of cAMP. It thereforeseems possible that the effects of weak acids andweak bases may be mediated by changes not in pHjbut in the intracellular level of cAMP, which may playthe key role in the control of stalk cell differentiation.Recent observations that 8-bromo cAMP, a mem-brane-permeable derivative of cAMP, induces thematuration of prestalk cells (Kwong et al. 1988;Maeda, 1988) seems to support this possibility. How-ever, it has not been demonstrated that NH3 removal,which was also shown to induce stalk cell maturation,elevates the intracellular level of cAMP. In any case,the rapid changes in pHj induced by weak acids andweak bases, being consequences of simple physico-chemical reactions, are likely to precede changes incAMP-release and -production rates and, in fact, it isquite possible that the former are the cause of thelatter.
Other weak acids and substances known to inducestalk cell differentiation, such as diethylstilboestrol(DES), differentiation inducing factor (DEF) andcAMP, were also examined for their effects onmigrating slugs (Table 1). To analyse the mode ofaction of these substances, it is convenient to dividethe process of stalk cell differentiation into two steps;the differentiation of aggregated cells into prestalkcells (step 1) and the maturation of prestalk cells tobecome true stalk cells (step 2).
It is evident from the above results that weak acidsinduce step 2. Induction of fruiting by weak acids orby removal of weak bases can be interpreted as aconsequence of relatively weak cytoplasmic acidifi-cation; the process of fruiting body constructionbegins with the maturation of prestalk cells in the tipregion followed by successive maturation of theadjacent prestalk cells (Raper & Fennell, 1952). It isconceivable that a relatively mild acid load wouldtrigger maturation only at the tip region (probably themost susceptible region within the slug to environ-mental changes) and, in this way, initiate the se-quence of events. Stronger acid loads, on the otherhand, would overcome the intrinsic control of stalkcell maturation (see below) and induce step 2 simul-taneously in all the prestalk cells. The stalk cellsformed in the posterior region of the slug probablyoriginated from 'anterior-like cells' (Sternfeld &David, 1981) and 'rear-guard cells' (Bonner, 1957,1967). These cell types are almost indistinguishablefrom prestalk cells, except that they are located in theposterior region (Bonner, 1967; Devine & Loomis,
1985; Kakutani & Takeuchi, 1986) and are preventedfrom maturing by the existence of real prestalk cells(Sternfeld & David, 1982). Conceivably, these cellsbecame mature stalk cells after prestalk cells in thesame slug ceased to emit the hypothetical maturation-inhibiting signal by becoming true stalk cells (seeSternfeld & David, 1982).
DES, DIF and cAMP were different from weakacids in that they not only caused the maturation ofprestalk cells but also induced the transdifferentiationof prespore cells to become mature stalk cells. DIFhas been shown to induce step 1 at the transcriptionallevel (Williams et al. 1987) and also to cause transdif-ferentiation from prespore to prestalk (Kay & Jer-myn, 1983). DES has also been shown to inducestep 1 (Kay et al. 1986). However, DIF and DES, bythemselves, do not seem sufficiently active to inducestep 2 in intact slugs. Although not conclusive, thisview seems to be favoured by the following argumentwhich is based on the idea described above that theinducer of step 2 should also induce fruiting. DIF didnot efficiently induce fruiting, nor did it cause thematuration of prestalk cells in situ within migratingslugs, even at concentrations almost two orders ofmagnitude higher than the effective concentration forinducing stalk cell maturation under the agar-sandwich condition. In the case of DES and otherproton-pump inhibitors, fruiting and stalk cell induc-tion are both induced at about the same concen-tration. This is in contrast with the case of weak acidswhere slugs were efficiently induced to fruit by lowerconcentrations of weak acids than those needed for insitu maturation of prestalk cells. These observationssuggest that, whereas weak acids induce step 2 inintact migrating slugs, DIF and DES do not. This mayappear to contradict the fact that a large number ofmature stalk cells are formed in cell monolayers inagar sandwiches, or in submerged cultures, whenDES or DIF are present. However, these results donot necessarily mean that they induce step 2. Theagar-sandwich method used in this study, and thesubmerged conditions used so far to examine theeffects of DIF and the other inducers, could requirethe inducer of step 1 for maintaining prestalk cells butcould allow them to mature without the need for theinducer of step 2. Conceivably, the action of DIF orDES is counteracted in intact migrating slugs by somefactor(s), which are diluted away under the agar-sandwich and submerged conditions. Recently,Sobolewski & Weeks (1988) showed that the tran-sition from prestalk cells to stalk cells is restrained inthe slug until the culmination stage but unrestrainedin vitro, supporting the above interpretation.
The effects of cAMP, under the agar-sandwichcondition, could at least partly be interpreted as aconsequence of the stimulation of DIF production by

Stalk cell maturation in D. discoideum 679
cAMP (Brookman et al. 1982). In this connection, itmay be worth noting that prespore cells were moreefficiently induced by cAMP to become stalk cellsthan prestalk cells (see Results). This could beexplained if prespore cells are more efficient DIFproducers. In fact, the prespore region of the slug hasbeen shown to contain at least twice as much DIF asthe prestalk region (Brookman et al. 1987), and thisdifference might reflect a difference between the twocell types in rate of DIF production. The effects ofcAMP on freely migrating slugs will be furthercomplicated by disturbance of the chemotactic signal-ling system, which is thought to be operating also inthe slug.
It will be interesting to compare the differencesbetween weak acids, DES and DIF with the differ-ence in their influences on the pH, of slug cells;whereas weak acids, as noted above, elicit an abruptdecrease in pHj, which is followed by a gradualrecovery, DES induces a slow and steady decline ofpHj. DIF, on the other hand, caused only a slight andtransient drop of pH, (Inouye, 1988). It thereforeseems possible that these pH, changes induced byweak acids, DES and DIF would be reflected, re-spectively, in the strong, weak and negligible induc-tion of step 2. This, together with the fact thatammonia inhibits both the pH; decrease and stalk cellmaturation, suggests that for prestalk cells to maturetheir pHj is required to drop sufficiently. However, itremains to be elucidated whether the pH; decreasedirectly induces the maturation process or whether itseffect is further mediated by other factors, such as adecrease in pHv as has been proposed to be the casein prestalk differentiation (Gross et al. 1988) or anincrease in the intracellular level of cAMP as men-tioned above (Williams et al. 1984). These possi-bilities are not mutually exclusive. If pHv proved tobe decisive, a small pHv drop would induce differen-tiation into prestalk cells (step 1) while a larger pHv
drop would be required for their maturation (step 2).Based on the above considerations, the following
hypothesis can be offered about the construction ofthe fruiting body (Sobolewski & Weeks, 1988 give asimilar discussion). While DIF induces stalk celldifferentiation, it is somehow blocked at step 2 duringslug migration. Ammonia is one of the candidates forthe blocker, because it is continuously produced bycells (Gregg et al. 1954; Schindler & Sussman, 1977;Feit & Sollitto, 1987), and because exogenouslyapplied ammonia blocks step 2. Alternatively, thecAMP signalling system may be responsible for theinhibition of DIF action, as cAMP has recently beenshown to interfere with DIF induction of stalk celldifferentiation (Berks & Kay, 1988) and also to raisethe pH, of aggregative cells (Aerts et al. 1987). If theformer is the case, loss of NH3 by evaporation from
the slug tip would relieve the block of step 2 andinitiate the maturation of prestalk cells. This is similarto the model proposed by Sussman & Schindler(1978), but, in their model, ammonia inhibits theaccumulation of cAMP which is postulated to be theinducer of stalk cell maturation.
However, there is some doubt as to whetherpreferential loss of ammonia by evaporation from thetip would be sufficient for restricting the maturationof prestalk cells within the limited region of thegrowing end of the stalk. The finding of this study thatweak acids induce stalk cell maturation raises anotherpossibility which is worthy of consideration. It hasbeen shown that stalk cells in the process of matu-ration degrade proteins and RNAs at a high ratereleasing a significant amount of ammonia (Gregg etal. 1954; Wilson & Rutherford, 1978). While am-monia (with its very high permeability through thecell membrane) will diffuse quickly through thetissue, other degradation products such as organicacids will diffuse relatively slowly. Therefore, it isreasonable to assume that some organic acid, whichare weak acids, accumulate near the apical end of thegrowing stalk, causing cytoplasmic acidification (andconsequently the maturation of nearby prestalkcells), while fast-diffusing ammonia would suppressacidification in the rest of the prestalk region. Thistype of regulation mechanism based on a short-rangeactivation signal and long-range inhibition has beenthought to be important in a variety of pattern-forming systems (Turing, 1952; for a review, seeMeinhardt, 1982). In order to know whether such amechanism is at work during the construction of thefruiting body in the cellular slime moulds, furtherbiochemical and cytological studies will be necessary.
I wish to thank Dr Julian Gross for discussion and forsuggestions for improvement of the manuscript, Dr MichaelHanna for comments on an earlier draft of the manuscriptand Dr Rob Kay for suggesting the ammonia-removalexperiments and for kindly providing DIF.
References
AERTS, R. J., DE WIT, R. J. W. & VAN LOOKEREN-CAMPAGNE, M. M. (1987). Cyclic AMP induces atransient alkalinization in Dictyostelium discoideum.FEBS Lett. 200, 366-370.
AKIYAMA, Y. & INOUYE, K. (1987). Cell-type conversionin normally proportioned and prestalk-enrichedpopulations of slug cells in Dictyostelium discoideum.Differentiation 35, 83-87.
BERKS, M. & KAY, R. R. (1988). Cyclic AMP is aninhibitor of stalk cell differentiation in Dictyosteliumdiscoideum. Devi Biol. 126, 108-114.
BONNER, J. T. (1952). The pattern of differentiation inamoeboid slime molds. Amer. Nat. 86, 79-89.

680 K. Inouye
BONNER, J. T. (1957). A theory of the control ofdifferentiation in the cellular slime molds. Q. Rev.Biol. 32, 232-246.
BONNER, J. T. (1967). The Cellular Slime Molds,pp. 1-205. Princeton, N.J.: Princeton Univ. Press.
BONNER, J. T. (1970). Induction of stalk celldifferentiation by cyclic AMP in the cellular slime moldDictyostelium discoideum. Proc. natn. Acad. Sci.U.S.A. 65, 110-113.
BROOKMAN, J. J., JERMYN, K. A. & KAY, R. R. (1987).Nature and distribution of the morphogen DIF in theDictyostelium slug. Development 100, 119-124.
BROOKMAN, J. J., TOWN, C. D., JERMYN, K. A. & KAY,
R. R. (1982). Developmental regulation of a stalk celldifferentiation-inducing factor in Dictyosteliumdiscoideum. Devi Biol. 91, 191-1%.
CHIA, W. K. (1975). Induction of stalk cell differentiationby cyclic-AMP in a susceptible variant of Dictyosteliumdiscoideum. Devi Biol. 44, 239-252.
DENMEAD, O. T., FRENEY, J. R. & SIMPSON, J. R. (1982).Dynamics of ammonia volatilization during furrowirrigation of maize. Soil Sci. Soc. Am. J. 46, 149-155.
DEVINE, K. M. & LOOMIS, W. F. (1985). Molecular
characterization of anterior-like cells in Dictyosteliumdiscoideum. Devi Biol. 107, 364-372.
FEIT, I. N., FOURNIER, G. A., NEEDLEMAN, R. D. &UNDERWOOD, M. Z. (1978). Induction of stalk andspore cell differentiation by cyclic AMP in slugs ofDictyostelium discoideum. Science 200, 439—441.
FEIT, I. N. & SOLLITTO, R. B. (1987). Ammonia is the gasused for the spacing of fruiting bodies in the cellularslime mold, Dictyostelium discoideum. Differentiation33, 193-1%.
GEORGE, R. P. (1977). Disruption of multicellularorganization in the cellular slime molds by cyclic AMP.Cell Differ. 5, 293-300.
GREGG, J. H., HACKNEY, A. L. & KRIVANEK, J. O.
(1954). Nitrogen metabolism of the slime moldDictyostelium discoideum during growth andmorphogenesis. Biol. Bull. mar. biol. Lab., WoodsHole 107, 226-235.
GROSS, J. D., BRADBURY, J., KAY, R. R. & PEACEY, M. J.(1983). Intracellular pH and the control of celldifferentiation in Dictyostelium discoideum. Nature,Lond. 303, 244-245.
GROSS, J. D., PEACEY, M. J. & POGGE-VON STRANDMANN,R. (1988). Plasma membrane proton pump inhibitionand stalk cell differentiation in Dictyosteliumdiscoideum. Differentiation (in press).
GROSS, J. D., TOWN, C. D., BROOKMAN, J. J., JERMYN, K.A., PEACEY, M. J. & KAY, R. R. (1981). Cellpatterning in Dictyostelium. Phil. Trans. R. Soc. Lond.B 295, 497-508.
HARRINGTON, B. J. & RAPER, K. B. (1968). Use of afluorescent brightener to demonstrate cellulose in thecellular slime molds. Appl. Microbiol. 16, 106-113.
INOUYE, K. (1985). Measurements of intracellular pH andits relevance to cell differentiation in Dictyosteliumdiscoideum. J. Cell Sci. 76, 235-245.
INOUYE, K. (1988). Differences in cytoplasmic pH and thesensitivity to acid load between prespore cells and
prestalk cells of Dictyostelium. J. Cell Sci. (in press).KAKUTANI, T. & TAKEUCHI, I. (1986). Characterization of
anterior-like cells in Dictyostelium as analyzed by theirmovement. Devi Biol. 115, 439-445.
KAY, R. R., GADIAN, D. G. & WILLIAMS, S. R. (1986).Intracellular pH in Dictyostelium: A 3IP nuclearmagnetic resonance study of its regulation and possiblerole in controlling cell differentiation. /. Cell Sci. 83,165-179.
KAY, R. R. & JERMYN, K. A. (1983). A possiblemorphogen controlling differentiation in Dictyostelium.Nature, Lond. 303, 242-244.
KWONG, L., SOBOLEWAKI, A. & WEEKS, G. (1988). Theeffect of cAMP on differentiation inducing factor(DIF)-mediated formation of stalk cells in low-cell-density monolayers of Dictyostelium discoideum.Differentiation 37, 1-6.
MAEDA, M. (1988). Dual effects of cAMP on the stabilityof prespore vesicles and 8-bromo cAMP-enhancedmaturation of spore and stalk cells of Dictyosteliumdiscoideum. Develop. Growth & Differ, (in press).
MEINHARDT, H. (1982). Models of Biological PatternFormation, pp. 1-230. London: Academic Press.
NESTLE, M. & SUSSMAN, M. (1972). The effect of cAMPon morphogenesis and enzyme accumulation inDictyostelium discoideum. Devi Biol. 28, 545-554.
RAPER, K. B. & FENNELL, D. I. (1952). Stalk formationin Dictyostelium. Bulletin of the Torrey Botanical Club79, 25-51.
Roos, A. & BORON, W. F. (1981). Intracellular pH.Physiol. Rev. 61, 2%-434.
SCHINDLER, J. & SUSSMAN, M. (1977). Ammoniadetermines the choice of morphogenetic pathways inDictyostelium discoideum. J. molec. Biol. 116, 161-169.
SCHINDLER, J. & SUSSMAN, M. (1979). Inhibition byammonia of intracellular cAMP accumulation inDictyostelium discoideum: Its significance for theregulation of morphogenesis. Devi Genet. 1, 13-20.
SOBOLEWSKI, A. & WEEKS, G. (1988). The requirementfor DEF for prestalk and stalk cell formation inDictyostelium discoideum: A comparison of in vivo andin vitro differentiation conditions. Devi Biol. 127,2%-303.
STERNFELD, J. & DAVID, C. N. (1981). Cell sorting duringpattern formation in Dictyostelium. Differentiation 20,10-21.
STERNFELD, J. & DAVID, C. N. (1982). Fate andregulation of anterior-like cells in Dictyostelium slugs.Devi Biol. 93, 111-118.
SUSSMAN, M. & SCHINDLER, J. (1978). A possiblemechanism of morphogenetic regulation inDictyostelium discoideum. Differentiation 10, 1-5.
SUSSMAN, M., SCHINDLER, J. & KIM, H. (1978)."Sluggers", a new class of moprphogenetic mutants ofD. discoideum. Expl Cell Res. 116, 217-227.
TOWN, C. D. & STANFORD, E. (1977). Stalk celldifferentiation by cells from migrating slugs ofDictyostelium discoideum: special properties of tipcells. /. Embryol. exp. Morph. 42, 105-113.
TURING, A. M. (1952). The chemical basis ofmorphogenesis. Philos. Trans. R. Soc. Lond. B. 237,

Stalk cell maturation in D. discoideum 681
37-72.WEUER, C. J. & DURSTON, A. J. (1985). Influence of
cAMP and hydrolysis products on cell type regulationin Dictyostelium discoideum. J. Embryol. exp. Morph.86, 19-37.
WILLIAMS, G. B., ELDER, E. M. & SUSSMAN, M. (1984).
Modulation of the cAMP relay in Dictyosteliumdiscoideum by ammonia and other metabolites:possible morphogenetic consequences. Devi Biol. 105,377-388.
WILLIAMS, J. G., CECCARELLI, A., MCROBBIE, S.,
MAHBUBANI, H., KAY, R. R., EARLY, A., BERKS, M. &
JERMYN, K. A. (1987). Direct induction of
Dictyostelium prestalk gene expression by DIF providesevidence that DIF is a morphogen. Cell 49, 185-192.
WILSON, J. B. & RUTHERFORD, C. L. (1978). ATP,
trehalose, glucose and ammonium ion localization inthe two cell types of Dictyostelium discoideum. J. cell.Physiol. 94, 37-46.
YAMAMOTO, A. & TAKEUCHI, I. (1983). Vital staining ofautophagic vacuoles in differentiating cells ofDictyostelium discoideum. Differentiation 24, 83-87.
(Accepted 23 August 1988)





























