Embed Size (px)
Citation preview

Induced resistance and control of charcoal rot inCicer arietinum (chickpea) by Pseudomonasfluorescens
Alok K. Srivastava, Tanuja Singh, T.K. Jana, and Dilip K. Arora
Abstract: Pseudomonas fluorescensisolate 4-92 induced systemic resistance against charcoal rot disease in chickpea(Cicer arietinumL.) caused byMacrophomina phaseolina(Tassi) Goidanich. Time-course accumulation ofpathogenesis-related (PR) proteins (chitinases and glucanases) in chickpea plants inoculated withP. fluorescenswassignificantly (P = 0.05) higher than in control plants. The level of chitinases and glucanases increased by 6.6- to 7-foldup to 4 days postinoculation; thereafter, little decrease in the activity of PR proteins was observed. Root-colonizingpopulations ofP. fluorescenswere at a maximum 2 days after transplantation at different inoculum concentrations, anddecreased over time. Inoculation of root tips of chickpea byP. fluorescens, 2,6-dichloroisonicotinic acid, ando-acetylsalicylic acid induced systemic resistance against charcoal rot. Disease was 33 to 55.5% higher in control plantsthan in plants inoculated with chemical inducers orP. fluorescens. Single treatment of plants withP. fluorescensin-creased disease resistance by 33%, whereas combined application ofP. fluorescenswith either of the chemical inducerswas most effective in inducing the resistance by 2- to 2.25-fold. The time-course study shows that an interval of atleast 2 days was required between induction treatment and challenge inoculation. Biocontrol efficacy ofP. fluorescensagainst charcoal rot disease in chickpea was demonstrated under greenhouse conditions.
Key words:biological control, induced resistance,Macrophomina phaseolina, Pseudomonas fluorescens.
Résumé: L’isolat 4-92 duPseudomonas fluorescensinduit une résistance systémique contre la pourriture charbonneusecausée par leMacrophomina phaseolina(Tassi) Goidanich chez le pois chiche (Cicer arietinumL.). L’accumulationdans le temps de protéines PR (chitinases et glucanases) chez des plants de pois chiche inoculés avec leP. fluorescensest significativement plus élevée que chez les plants témoins. Les teneurs en chitinases et glucanases augmentent de 6,6à 7 fois jusqu’à 4 jours après l’inoculation; par après, on observe peu de diminution dans l’activité des protéines PR.Les populations duP. fluorescenscolonisant les racines atteignent un maximum 2 jours après la transplantation en pré-sence de différentes concentrations d’inoculum, et diminuent avec le temps. L’inoculation des apex racinaires de poischiches avec leP. fluorescens, l’acide 2,6-dichloroisonicotinic et l’acideo-acétylsalicylique induit une résistance systé-mique contre la pourriture charbonneuse. Chez les plantes témoins, la maladie est de 33 à 55,5% plus élevée que chezles plantes inoculées avec les inducteurs chimiques ou leP. fluorescens. Un seul traitement avec leP. fluorescensaug-mente la résistance à la maladie de 33%, alors qu’une application combinée duP. fluorescensavec un des deux induc-teurs chimiques est la plus efficace, augmentant la résistance induite de 2 à 2,25 fois. L’étude dans le temps montrequ’il faut une intervalle d’au moins 2 jours entre le traitement d’induction et l’inoculation avec le pathogène. On aainsi démontré l’efficacité duP. fluorescens, en serre, comme agent de lutte biologique contre la pourriture charbon-neuse chez le pois chiche.
Mots clés: lutte biologique, résistance induite,Macrophomina phaseolina, Pseudomonas fluorescens.
[Traduit par la Rédaction] Srivastava et al. 795
Introduction
Chickpea (Cicer arietinumL.) is the world’s third mostimportant pulse crop after beans and peas, with India ac-counting for approximately 75% of the world production(FAO 1993). Chickpea productivity, however, has remainedvirtually stagnant over recent decades because of its intoler-
ance to insect pests such asHelicoverpa armigera,suscepti-bility to diseases such as wilt (Fusarium oxysporumf.sp.ciceri), charcoal rot (Macrophomina phaseolina(Tassi)Goidanich), and blight (Ascochyta rabiei). The soilbornefungusM. phaseolinais endemic throughout the temperateand tropical regions of the world and has been isolated fromover 400 plant species (Mihail and Taylor 1995). Charcoal
Can. J. Bot.79: 787–795 (2001) © 2001 NRC Canada
787
DOI: 10.1139/cjb-79-7-787
Received August 29, 2000. Published on the NRC Research Press Web site at http://cjb.nrc.ca on June 26, 2001.
A.K. Srivastava. PG Department of Botany, SMM Town PG College, Ballia 277 001, India.T. Singh and D.K. Arora.1 Laboratory of Applied Mycology, Centre of Advanced Study in Botany, Banaras Hindu University,Varanasi 221 005, India.T.K. Jana. National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute, New Delhi 110012, India.
1Corresponding author (e-mail: [email protected]).
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:05 AM
Color profile: DisabledComposite Default screen

rot of chickpea is one of the most common diseases in Indiaand severely limits chickpea production (ICRISAT AnnualReport 1988; Pandey and Singh 1990). The colonization ofseeds, soil and root surfaces, distribution and hosts, longev-ity in soil, and competitive saprophytic ability of this patho-gen have been studied extensively (Mihail and Alcorn 1987;Olanya and Campbell 1989; Collins et al. 1991; Olaya et al.1996).
Owing to a lack of resistance in plants to most root dis-eases as well as ecological and economical restrictions onfungicide use for control of soilborne diseases, biologicalcontrol and other ecofriendly disease control measures havegained increasing interest. The common feature in many in-compatible interactions between plants and pathogens is thedevelopment of the hypersensitive response (HR). The HRcoincides with many metabolic changes in the affected plantcells, such as strengthening of structural barriers, synthesisof lytic enzymes, or production of phytoalexins (Kuc 1987;Van Peer et al. 1991). Furthermore, many plants that haveundergone the HR show increased resistance against subse-quent microbial attack. This nonlocalized and long-lastinginduced protection, which is active against a broad spectrumof pathogens, is known as induced systemic resistance (ISR).Treatment of plants with nonpathogenic rhizosphere micro-organisms or chemical inducers (e.g., isonicotinic acid orsalicylic acid) enhances plant resistance to subsequent chal-lenge infection by a variety of viral, bacterial, and fungalpathogens (Raupach and Kloepper 1997; Sticher et al. 1997;Van Loon et al. 1998; Meyer et al. 1999; Tuzun and Bent1999; Chen et al. 1999; Yedidia et al. 1999; Enebak andCarey 2000).
In recent years, plant growth promoting rhizobacteria(PGPR), such as strains ofPseudomonasspecies, have beenused extensively for plant growth promotion and diseasecontrol because of many properties: efficient colonization ofunderground plant organs, utilization of a large number oforganic substrates commonly found in root exudates, andproduction of a variety of secondary metabolites toxic tofungi and bacteria. They are also inexpensive to grow andcan be introduced into the rhizosphere by seed treatments oras granules. It has become apparent that PGPR-mediatedinduced resistance is associated with marked metabolicchanges, culminating in a number of physical and biochemi-cal responses designed to limit pathogen penetration anddevelopment in host tissues, either indirectly (reinforcementof plant cell walls) or directly (antimicrobial activity)(Hammerschmidt and Kuc 1995; Barka et al. 2000). ISR inPseudomonas-mediated disease suppression is relatively newand has recently gained considerable importance; however,several researchers (Leeman et al. 1995; Liu et al. 1995;Hoffland et al. 1996; Kataria et al. 1997) found that ISR isassociated with the production of pathogenesis-related (PR)proteins, siderophores (Lam and Gaffney 1993), increasedlevels of salicylic acid (Chen et al. 1999), and metabolism ofpolyunsaturated fatty acids in plants (Delanay et al. 1994;Duijff et al. 1994; Pieterse et al. 1996; Daayf et al. 1997; DeMeyer and Höfte 1997; Ishi et al. 1999). Some studies indi-cated that salicylic acid may not play a role in translocationof signals for ISR (Vernooij et al. 1994; Pieterse et al. 1996).Hoffland et al. (1996) reported that a PGPR strain of
P. fluorescens, WCS417, induced systemic resistanceagainstFusariumspp. without inducing the accumulation ofPR proteins.
Control of charcoal root rot of chickpea is achieved mainlywith the use of several fungicides, alone or integrated withother strategies (Mihail and Alcorn 1984; Bhattacharya et al.1985; Dubey 1992; Lodha 1995; Majumdar et al. 1996;Cardoso et al. 1997). No concerted effort has been made toinvestigate the possibility of controlling this disease withinducible host defense responses. We have reported earlierthat P. fluorescensisolate 4-92 effectively colonized andreduced the germination of sclerotia ofM. phaseolinaandmay act as a potential biocontrol agent against charcoal rot(Srivastava et al. 2001). In the present investigation, the roleof P. fluorescens4-92 in induction of systemic resistanceagainst charcoal rot of chickpea was evaluated. The accumu-lation of PR proteins as a prerequisite to ISR and the time-course required for the induction of resistance were alsostudied. The efficacy ofP. fluorescenswas compared with thatof known chemical inducers (isonicotinic acid ando-salicylicacid). Suppression of charcoal rot byP. fluorescens4-92through ISR and root colonization biocontrol assay was ex-amined under greenhouse conditions.
Materials and methods
Bacterial strains and pathogenPseudomonas fluorescensisolate 4-92, originally isolated from
the rhizosphere ofCicer arietinumcv. Radhey (Srivastava et al.1996), was grown on King’s B broth (King et al. 1954) at28 ± 2°C. For preparation of inoculum, exponential phase cells(A500 = 0.15–0.18) were collected from 24-h-old cultures by centri-fugation (5000 × g; 10 min), washed twice with 15.6 g/LNaH2PO4·H2O and 9 g/L NaCl phosphate buffer saline (PBS; 0.1M; pH 7) and resuspended in PBS at a concentration of 108
cells/mL. In all experiments, bacterial cell suspensions grown inexponential phase were used.Macrophomina phaseolinaisolateCH-15, which was previously isolated from diseased chickpeaplants, was obtained from the culture collection in the laboratory ofApplied Mycology, Banaras Hindu University (Srivastava et al.1996). The fungus was grown and maintained on synthetic media(Okon et al. 1973; 0.9 g/L K2HPO4, 0.2 g/L MgSO4, 0.2 g/L KCl,1.0 g/L NH4NO3, 0.002 g/L Fe2+, 0.002 g/L Mn2+, 0.002 g/L Zn2+,0.0001 g/L thiamine hydrochloride, pH 6.8) with 0.5% glucose.
Delivery system for the bacterial inoculumSeeds ofC. arietinum cv. Radhey susceptible to charcoal rot
were surface sterilized for 2 min with 70% ethanol, followed by2% NaOCl and soaked in sterilized distilled water for 12 h. Theseeds were transferred to sterilized moist filter papers in sterilizedpolycarbonate boxes (25 × 25 × 10 cm) and incubated for 48–72 hat 28 ± 2°C. Seedlings (3–3.5 cm long) were transferred to plasticpots (20 cm diam.; 2 seedlings/pot) that were half-filled with steril-ized, acid-washed sand (400 g/pot). Three methods of bacterialinoculation were used to induce resistance in chickpea seedlings:(i) inoculation of seedlings byP. fluorescens4-92 mixed with tal-cum as a carrier (E-Merck India Ltd., Bombay, India; 1:1 v/w; ca.108 cfu/g talcum); (ii ) drenching of sand with cell suspensions ofP. fluorescens; (iii ) mixing of P. fluorescenswith sand. After inocu-lation, the seedlings were kept on moist filter papers for 2 h toestablish the bacterial cells and then transplanted to pots containingsterilized sand. Drenching of the sterilized sand in pots was doneby a cell suspension ofP. fluorescens(ca. 108 cfu/mL PBS;
© 2001 NRC Canada
788 Can. J. Bot. Vol. 79, 2001
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:06 AM
Color profile: DisabledComposite Default screen

30 mL/pot) immediately before transplanting seedlings. In the thirdset of treatments, the cell suspension was mixed directly with ster-ilized sand (ca. 108 cells/g sand) before filling the pots. Seedlingswere transplanted to the pots either drenched with bacteria or sandmixed with bacteria. Control treatments consisted of (i) noninoculatedseedlings, (ii) noninoculated seedlings sand drenched withPBS, and(iii ) noninoculated seedlings mixed with sterilized sand, respectiveto their treatment. In all the transplantations, cotyledons were keptabove the surface of the sand. The pots were watered, and theupper surface of the sand was covered by a 3 cm thick layer ofsterilized perlite and kept in the greenhouse (28–30°C:24°C,light:dark) under cool fluorescent light in a complete randomizedblock design. The pots were watered every day, and once a weekthey received Hoagland and Knop’s plant nutrient solution(40 mL/pot; Dhingra and Sinclair 1985).
Protein extraction and assay for PR proteinsSeedlings were uprooted at a regular interval of 24 h after emer-
gence until 120 h (10 seedlings per sample) and washed thoroughlywith sterilized distilled water. The roots were weighed and groundin liquid nitrogen. The powder was homogenized in cold PBS (1:2w/v; 2 min at 4°C). The homogenate was centrifuged twice(10 000 ×g; 20 min at 4°C), and the supernatant was collected andstored at –20°C. The supernatant served as crude enzyme prepara-tion (CEP). PR proteins, i.e., chitinases andβ-1,3-glucanases, wereassayed by colorimetric methods. Assay of chitinases was based ondetermination ofp-nitriphenyl cleaved from a chitin analogoussubstratep-nitrophenyl-β-D-N-N ′-diacetylchitobiose (PNP) (Rob-erts and Selitrennikoff 1988). The CEP (40µL) and 10µL of PNPstock solution (2 mg/mL) was mixed with 50 mM of acetate buffer(pH 5) to a final volume of 0.5 mL. The tubes were incubated at40°C in a water bath for 50 min, and the reaction was terminatedby adding 0.5 mL of 0.2 M Na2CO3. The amount of cleavedp-nitriphenyl was determined by measuring the absorbance at410 nm in a UV-visible spectrophotometer.β-1,3-Glucanase activ-ity was estimated using the procedure of Koga et al. (1988). Theassay mixture contained 0.1 mL of crude enzyme preparation and0.4 mL of 1% laminarin solution in acetate buffer (100 mM; pH 5).The mixture was incubated for 50 min in a water bath at 40°C andthen terminated by incubation in boiling water for 5 min. Glucostat(2.5 mL mixture), consisting of PGO enzymes (0.5 mL; 100–500IU; Sigma) ando-dianisidine dihydrochloride (2.5 mg/mL; Sigma),was added to each tube and the reducing sugars released were mea-sured at 450 nm. Chitinase and glucanase activity were expressedas millimoles of PNP or milligrams of glucose (cleaved by en-zymes) obtained per gram fresh weight of roots. One unit ofchitinases andβ-1,3-glucanase was defined as the amount of en-zyme that produced 10 mM of PNP or 1 mg of glucose per gramfresh tissue per hour, respectively, under assay conditions. The as-says contained 20 replicates and were repeated twice.
Efficacy of chemical inducers and inoculum density ofP. fluorescens
Two known inducers of systemic resistance (2,6-dichloroisonicotinic acid (INA; 25% water soluble powder) ando-acetylsalicylic acid (SA; Sigma)) and different concentrations ofP. fluorescens(104–109 cells/mL) were used to test the induction ofPR proteins. The chemicals were applied on root tips of seedlings bydipping the tips (1.5 cm) in a suspension containing either 0.25 MINA or 2 M SA. Different concentrations (104–109 cells/mL) ofP. fluorescenswere also applied on the tips of seedlings in a simi-lar manner as described earlier. After chemical treatment or inocu-lation with bacteria, the seedlings were transplanted into pots (twoper pot) containing sterilized sand and covered by a 3 cm thickperlite layer. Control treatments consisted of seedlings dipped in
PBS. The pots were kept in the greenhouse under the same condi-tions as described and sampled daily for PR protein until 8 days.
Challenge inoculation and induced resistanceFor challenge inoculation, pots were prepared as described for
induction treatment. The pots were placed in a greenhouse underthe same conditions as described above. To study the interaction ofP. fluorescensand chemical inducers on disease incidence, the roottips of seedlings were first dipped in the suspension of 0.25 M INAor 2 M SA, allowed to stabilize for 30 min, and then dipped in a106 cells/mL cell suspension ofP. fluorescens.Inoculated andchemically induced or combined treatment seedlings were trans-planted into pots (two per pot). The growing seedlings were steminoculated withM. phaseolina2, 4, 6, and 8 days after transplanta-tion. Macrophomina phaseolinawas grown on cellophane mem-branes spread over synthetic media in Petri plates. The cellophanemembranes with hyphal masses were taken from 4-day-old platesincubated at 28 ± 2°C and cut into strips (6 × 1.5 cm). The stripswere wrapped at the base of stems just above the perlite layer andtied with parafilm. To detect possible competition betweenM. phaseolinaandP. fluorescens, the presence ofP. fluorescensatthe inoculation site on the stem was checked both at the time of in-oculation and at the end of the experiment. Stem segments (2 cmlong) were taken just above the stem base and incubated on KBmedia. The number of diseased plants was scored 4 weeks after in-oculation. A plant was considered to be diseased with the appear-ance of the first local symptom of brown-black spots at the collarregion of the stem.
Greenhouse assay for biological controlThe biocontrol ability ofP. fluorescenswas evaluated in green-
house experiments.Macrophomina phaseolinawas cultured onsand–maize meal (9:1 w/w). After 7 days of incubation at 28 ±2°C, the inoculum was mixed with sand to a final inoculum densityof approximately 103 cfu/g sand. A cell suspension ofP. fluorescens(ca. 108 cells/mL PBS), harvested from exponential-phase KB cultures, was mixed in pathogen-infested sand (ca. 108
cells/g sand). The inoculated sand mixtures were transferred intopots (400 g/pot). Surface-sterilized chickpea seeds (two per pot)were sown and the pots were maintained in a greenhouse as de-scribed earlier for induction treatment. Growing plants were sam-pled for disease incidence and root colonization byP. fluorescensat regular intervals of 7 days up to 28 days after emergence. Dis-ease was graded using the following scale: 1, healthy plants, nosymptoms; 2, browning of collar region; 3, dark brown to blackspots on collar as well as on primary roots; 4, weak and stuntedplants with rotting of roots; 5, plants dead. Lesions on the entireroot system and the disease severity index were calculated as de-scribed by Bull et al. (1991). To determine the root-colonizing pop-ulation of P. fluorescenswe cut a sample of roots into segments;2 g of root segments (1 cm long) were macerated and vortexed for2–3 min in 10 mL of sterilized PBS. Aliquots (0.2 mL) of dilutedsuspensions (10–4 and 10–5) were plated on KB medium. After 48–72 h, colonies ofP. fluorescenswere counted and expressed ascfu/g root.
All experiments were repeated twice with 20 replicates for eachtreatment. In greenhouse tests, plants were arranged using a ran-domized complete block design. Each plant constituted an experi-mental unit and each unit was replicated 20 times within eachtreatment. The time course assays of enzymatic activity and rootcolonization study (Figs. 1 and 2) were subjected to standard devi-ation. Enzymatic activity at different concentrations ofP. fluorescens(Table 1), root colonization and biological control byP. fluorescensunder greenhouse conditions (Table 2), and induced-resistance bioassay (Fig. 3) were subjected to analysis of variance,
© 2001 NRC Canada
Srivastava et al. 789
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:06 AM
Color profile: DisabledComposite Default screen
and differences between means were detected using Duncan’s mul-tiple range test.
Results
Delivery system for the bacterial inoculum andaccumulation of PR proteins
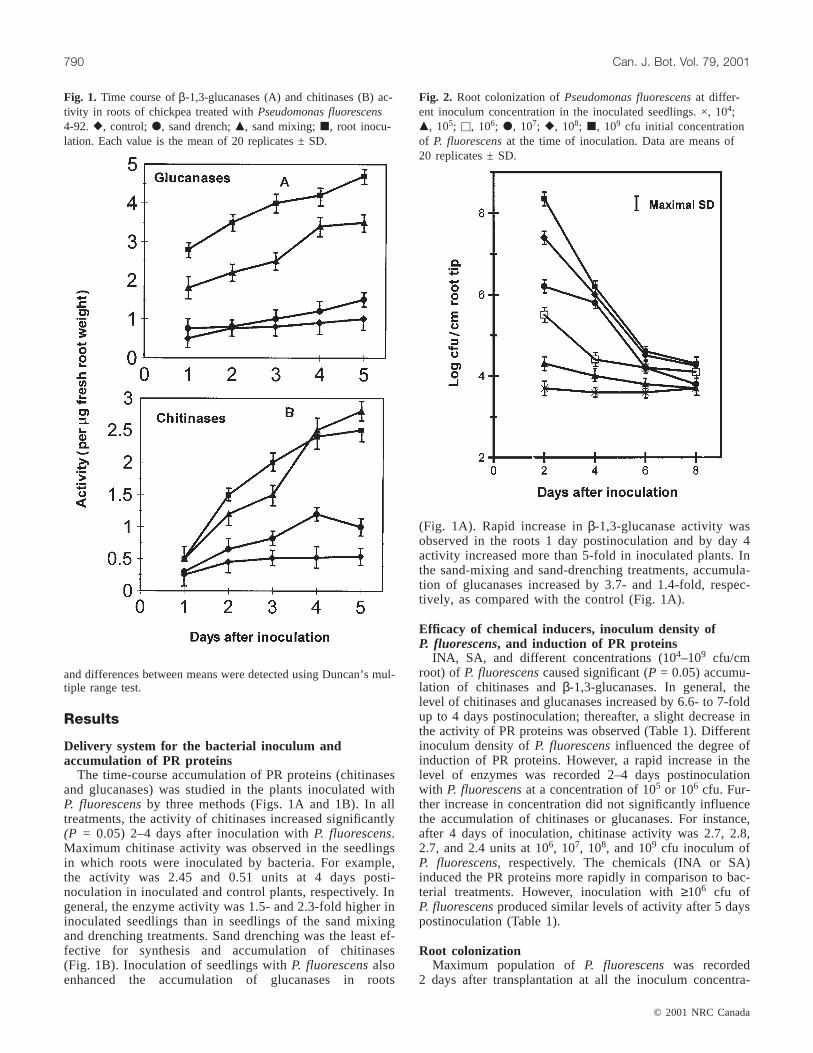
The time-course accumulation of PR proteins (chitinasesand glucanases) was studied in the plants inoculated withP. fluorescensby three methods (Figs. 1A and 1B). In alltreatments, the activity of chitinases increased significantly(P = 0.05) 2–4 days after inoculation withP. fluorescens.Maximum chitinase activity was observed in the seedlingsin which roots were inoculated by bacteria. For example,the activity was 2.45 and 0.51 units at 4 days posti-noculation in inoculated and control plants, respectively. Ingeneral, the enzyme activity was 1.5- and 2.3-fold higher ininoculated seedlings than in seedlings of the sand mixingand drenching treatments. Sand drenching was the least ef-fective for synthesis and accumulation of chitinases(Fig. 1B). Inoculation of seedlings withP. fluorescensalsoenhanced the accumulation of glucanases in roots
(Fig. 1A). Rapid increase inβ-1,3-glucanase activity wasobserved in the roots 1 day postinoculation and by day 4activity increased more than 5-fold in inoculated plants. Inthe sand-mixing and sand-drenching treatments, accumula-tion of glucanases increased by 3.7- and 1.4-fold, respec-tively, as compared with the control (Fig. 1A).
Efficacy of chemical inducers, inoculum density ofP. fluorescens, and induction of PR proteins
INA, SA, and different concentrations (104–109 cfu/cmroot) of P. fluorescenscaused significant (P = 0.05) accumu-lation of chitinases andβ-1,3-glucanases. In general, thelevel of chitinases and glucanases increased by 6.6- to 7-foldup to 4 days postinoculation; thereafter, a slight decrease inthe activity of PR proteins was observed (Table 1). Differentinoculum density ofP. fluorescensinfluenced the degree ofinduction of PR proteins. However, a rapid increase in thelevel of enzymes was recorded 2–4 days postinoculationwith P. fluorescensat a concentration of 105 or 106 cfu. Fur-ther increase in concentration did not significantly influencethe accumulation of chitinases or glucanases. For instance,after 4 days of inoculation, chitinase activity was 2.7, 2.8,2.7, and 2.4 units at 106, 107, 108, and 109 cfu inoculum ofP. fluorescens, respectively. The chemicals (INA or SA)induced the PR proteins more rapidly in comparison to bac-terial treatments. However, inoculation with≥106 cfu ofP. fluorescensproduced similar levels of activity after 5 dayspostinoculation (Table 1).
Root colonizationMaximum population of P. fluorescenswas recorded
2 days after transplantation at all the inoculum concentra-
© 2001 NRC Canada
790 Can. J. Bot. Vol. 79, 2001
Fig. 1. Time course ofβ-1,3-glucanases (A) and chitinases (B) ac-tivity in roots of chickpea treated withPseudomonas fluorescens4-92. r, control; d, sand drench;m, sand mixing;j, root inocu-lation. Each value is the mean of 20 replicates ± SD.
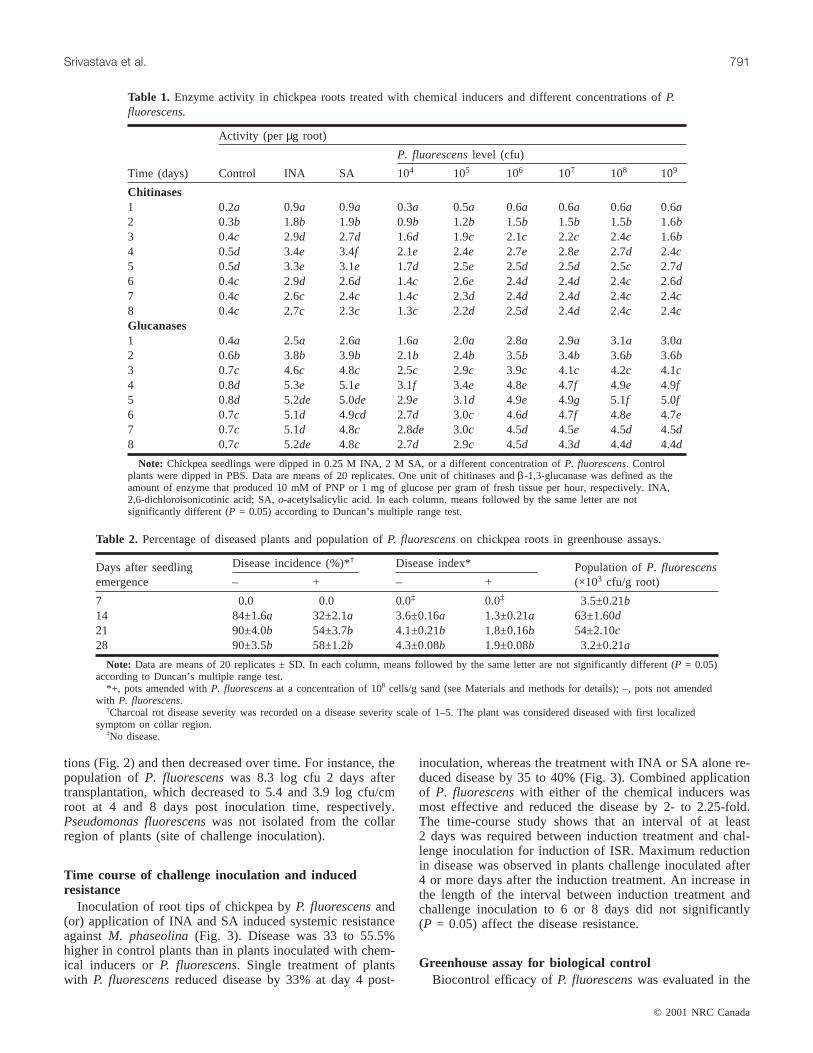
Fig. 2. Root colonization ofPseudomonas fluorescensat differ-ent inoculum concentration in the inoculated seedlings. ×, 104;m, 105; u, 106; d, 107; r, 108; j, 109 cfu initial concentrationof P. fluorescensat the time of inoculation. Data are means of20 replicates ± SD.
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:09 AM
Color profile: DisabledComposite Default screen
© 2001 NRC Canada
Srivastava et al. 791
tions (Fig. 2) and then decreased over time. For instance, thepopulation ofP. fluorescenswas 8.3 log cfu 2 days aftertransplantation, which decreased to 5.4 and 3.9 log cfu/cmroot at 4 and 8 days post inoculation time, respectively.Pseudomonas fluorescenswas not isolated from the collarregion of plants (site of challenge inoculation).
Time course of challenge inoculation and inducedresistance
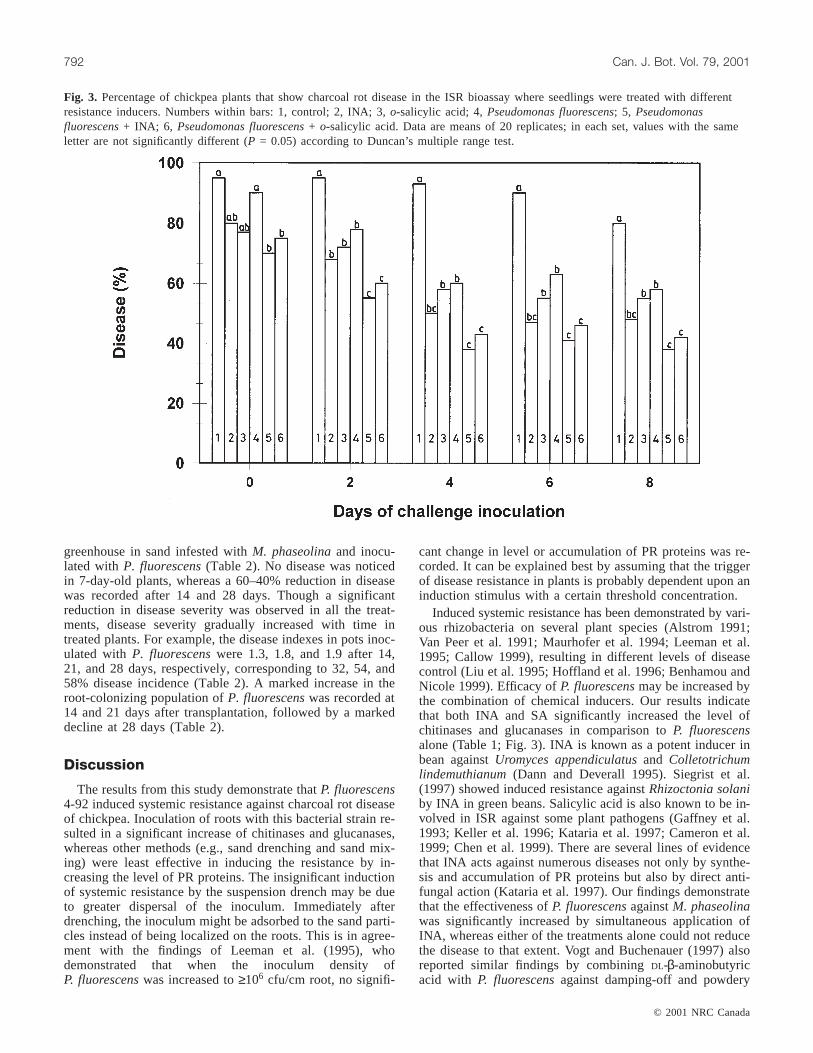
Inoculation of root tips of chickpea byP. fluorescensand(or) application of INA and SA induced systemic resistanceagainstM. phaseolina(Fig. 3). Disease was 33 to 55.5%higher in control plants than in plants inoculated with chem-ical inducers orP. fluorescens. Single treatment of plantswith P. fluorescensreduced disease by 33% at day 4 post-
inoculation, whereas the treatment with INA or SA alone re-duced disease by 35 to 40% (Fig. 3). Combined applicationof P. fluorescenswith either of the chemical inducers wasmost effective and reduced the disease by 2- to 2.25-fold.The time-course study shows that an interval of at least2 days was required between induction treatment and chal-lenge inoculation for induction of ISR. Maximum reductionin disease was observed in plants challenge inoculated after4 or more days after the induction treatment. An increase inthe length of the interval between induction treatment andchallenge inoculation to 6 or 8 days did not significantly(P = 0.05) affect the disease resistance.
Greenhouse assay for biological controlBiocontrol efficacy ofP. fluorescenswas evaluated in the
Activity (per µg root)
P. fluorescenslevel (cfu)
Time (days) Control INA SA 104 105 106 107 108 109
Chitinases1 0.2a 0.9a 0.9a 0.3a 0.5a 0.6a 0.6a 0.6a 0.6a2 0.3b 1.8b 1.9b 0.9b 1.2b 1.5b 1.5b 1.5b 1.6b3 0.4c 2.9d 2.7d 1.6d 1.9c 2.1c 2.2c 2.4c 1.6b4 0.5d 3.4e 3.4f 2.1e 2.4e 2.7e 2.8e 2.7d 2.4c5 0.5d 3.3e 3.1e 1.7d 2.5e 2.5d 2.5d 2.5c 2.7d6 0.4c 2.9d 2.6d 1.4c 2.6e 2.4d 2.4d 2.4c 2.6d7 0.4c 2.6c 2.4c 1.4c 2.3d 2.4d 2.4d 2.4c 2.4c8 0.4c 2.7c 2.3c 1.3c 2.2d 2.5d 2.4d 2.4c 2.4cGlucanases1 0.4a 2.5a 2.6a 1.6a 2.0a 2.8a 2.9a 3.1a 3.0a2 0.6b 3.8b 3.9b 2.1b 2.4b 3.5b 3.4b 3.6b 3.6b3 0.7c 4.6c 4.8c 2.5c 2.9c 3.9c 4.1c 4.2c 4.1c4 0.8d 5.3e 5.1e 3.1f 3.4e 4.8e 4.7f 4.9e 4.9f5 0.8d 5.2de 5.0de 2.9e 3.1d 4.9e 4.9g 5.1f 5.0f6 0.7c 5.1d 4.9cd 2.7d 3.0c 4.6d 4.7f 4.8e 4.7e7 0.7c 5.1d 4.8c 2.8de 3.0c 4.5d 4.5e 4.5d 4.5d8 0.7c 5.2de 4.8c 2.7d 2.9c 4.5d 4.3d 4.4d 4.4d
Note: Chickpea seedlings were dipped in 0.25 M INA, 2 M SA, or a different concentration ofP. fluorescens. Controlplants were dipped in PBS. Data are means of 20 replicates. One unit of chitinases andβ -1,3-glucanase was defined as theamount of enzyme that produced 10 mM of PNP or 1 mg of glucose per gram of fresh tissue per hour, respectively. INA,2,6-dichloroisonicotinic acid; SA,o-acetylsalicylic acid. In each column, means followed by the same letter are notsignificantly different (P = 0.05) according to Duncan’s multiple range test.
Table 1. Enzyme activity in chickpea roots treated with chemical inducers and different concentrations ofP.fluorescens.
Days after seedlingemergence
Disease incidence (%)*† Disease index* Population ofP. fluorescens(×103 cfu/g root)– + – +
7 0.0 0.0 0.0‡ 0.0‡ 3.5±0.21b14 84±1.6a 32±2.1a 3.6±0.16a 1.3±0.21a 63±1.60d21 90±4.0b 54±3.7b 4.1±0.21b 1.8±0.16b 54±2.10c28 90±3.5b 58±1.2b 4.3±0.08b 1.9±0.08b 3.2±0.21a
Note: Data are means of 20 replicates ± SD. In each column, means followed by the same letter are not significantly different (P = 0.05)according to Duncan’s multiple range test.
*+, pots amended withP. fluorescensat a concentration of 108 cells/g sand (see Materials and methods for details); –, pots not amendedwith P. fluorescens.
†Charcoal rot disease severity was recorded on a disease severity scale of 1–5. The plant was considered diseased with first localizedsymptom on collar region.
‡No disease.
Table 2. Percentage of diseased plants and population ofP. fluorescenson chickpea roots in greenhouse assays.
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:10 AM
Color profile: DisabledComposite Default screen
greenhouse in sand infested withM. phaseolinaand inocu-lated withP. fluorescens(Table 2). No disease was noticedin 7-day-old plants, whereas a 60–40% reduction in diseasewas recorded after 14 and 28 days. Though a significantreduction in disease severity was observed in all the treat-ments, disease severity gradually increased with time intreated plants. For example, the disease indexes in pots inoc-ulated with P. fluorescenswere 1.3, 1.8, and 1.9 after 14,21, and 28 days, respectively, corresponding to 32, 54, and58% disease incidence (Table 2). A marked increase in theroot-colonizing population ofP. fluorescenswas recorded at14 and 21 days after transplantation, followed by a markeddecline at 28 days (Table 2).
Discussion
The results from this study demonstrate thatP. fluorescens4-92 induced systemic resistance against charcoal rot diseaseof chickpea. Inoculation of roots with this bacterial strain re-sulted in a significant increase of chitinases and glucanases,whereas other methods (e.g., sand drenching and sand mix-ing) were least effective in inducing the resistance by in-creasing the level of PR proteins. The insignificant inductionof systemic resistance by the suspension drench may be dueto greater dispersal of the inoculum. Immediately afterdrenching, the inoculum might be adsorbed to the sand parti-cles instead of being localized on the roots. This is in agree-ment with the findings of Leeman et al. (1995), whodemonstrated that when the inoculum density ofP. fluorescenswas increased to≥106 cfu/cm root, no signifi-
cant change in level or accumulation of PR proteins was re-corded. It can be explained best by assuming that the triggerof disease resistance in plants is probably dependent upon aninduction stimulus with a certain threshold concentration.
Induced systemic resistance has been demonstrated by vari-ous rhizobacteria on several plant species (Alstrom 1991;Van Peer et al. 1991; Maurhofer et al. 1994; Leeman et al.1995; Callow 1999), resulting in different levels of diseasecontrol (Liu et al. 1995; Hoffland et al. 1996; Benhamou andNicole 1999). Efficacy ofP. fluorescensmay be increased bythe combination of chemical inducers. Our results indicatethat both INA and SA significantly increased the level ofchitinases and glucanases in comparison toP. fluorescensalone (Table 1; Fig. 3). INA is known as a potent inducer inbean againstUromyces appendiculatusand Colletotrichumlindemuthianum(Dann and Deverall 1995). Siegrist et al.(1997) showed induced resistance againstRhizoctonia solaniby INA in green beans. Salicylic acid is also known to be in-volved in ISR against some plant pathogens (Gaffney et al.1993; Keller et al. 1996; Kataria et al. 1997; Cameron et al.1999; Chen et al. 1999). There are several lines of evidencethat INA acts against numerous diseases not only by synthe-sis and accumulation of PR proteins but also by direct anti-fungal action (Kataria et al. 1997). Our findings demonstratethat the effectiveness ofP. fluorescensagainstM. phaseolinawas significantly increased by simultaneous application ofINA, whereas either of the treatments alone could not reducethe disease to that extent. Vogt and Buchenauer (1997) alsoreported similar findings by combiningDL-β-aminobutyricacid with P. fluorescensagainst damping-off and powdery
© 2001 NRC Canada
792 Can. J. Bot. Vol. 79, 2001
Fig. 3. Percentage of chickpea plants that show charcoal rot disease in the ISR bioassay where seedlings were treated with differentresistance inducers. Numbers within bars: 1, control; 2, INA; 3,o-salicylic acid; 4,Pseudomonas fluorescens; 5, Pseudomonasfluorescens+ INA; 6, Pseudomonas fluorescens+ o-salicylic acid. Data are means of 20 replicates; in each set, values with the sameletter are not significantly different (P = 0.05) according to Duncan’s multiple range test.
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:12 AM
Color profile: DisabledComposite Default screen

mildew of cucumber. The enhanced induction of the resis-tance mechanism in combined treatments may be due to thestimulation of growth and activity of bacteria in the rhizo-sphere by alteration of root exudation by chemicals. Theresults of the time-course study show that at least 2 days arerequired between the initial inoculation with resistance in-ducers and challenge inoculation withM. phaseolinaforinduction of resistance (Fig. 3). The level of protectionthrough ISR increased with time, reaching maximum protec-tion at day 4 postinoculation. The disease severity in controlplants at 8 days postinoculated was relatively low, whichmay be due to unfavourable conditions required for diseasedevelopment. These data suggest that a lag period is re-quired for the putative signal to be transported from the siteof induction to other plant parts (Fig. 3). In this experiment,the population ofP. fluorescenswas not detected in the col-lar region, which indicated that the bacterium and pathogenremained spatially separated during the experiment (data notshown). Therefore, it appears that control of the diseasecould be due to ISR via synthesis and accumulation of chiti-nases andβ-1,3-glucanases. Depolymerization of cell wallsby the combined activity of chitinases and glucanases wasreported to kill fungi in vitro (Schlumbaum et al. 1986).Punja and Zhang (1993) also discussed the role of these en-zymes in disease resistance and explained that resistance re-sponse initially was an HR that may be involved in slowingdown pathogen growth and sporulation in later stages of dis-ease development. In this study, possible production of anti-fungal metabolites byP. fluorescens, its absorption by thechickpea roots, and role in induction of disease was not in-vestigated. Along with the induced resistance, the biocontrolefficacy ofP. fluorescens4-92 was also evaluated (Table 2).A significant (P = 0.05) reduction in disease incidence andseverity was observed in all the samples. Sixty percent dis-ease suppression was recorded after 14 days of incubation,which decreased to 40% by 28 days with an increase in dis-ease severity from 1.3 to 1.9 (Table 2). This may be due to adecrease in the root colonizing population ofP. fluorescenswith time (Table 2) or some other common mechanism bywhich P. fluorescensisolates act as a biocontrol agentagainst other soilborne pathogenic fungi (Loper et al. 1994).In conclusion, our findings demonstrate thatP. fluorescens4-92 can induce resistance against charcoal rot of chickpeacaused byM. phaseolina, possibly by accumulation of chiti-nases andβ-1,3-glucanases.Pseudomonas fluorescenscom-bined with chemical inducers also significantly increased theeffectiveness of disease resistance. To the best of our knowl-edge, this is the first report showing ISR against the charcoalrot pathogenM. phaseolina. These results are of particularrelevance because they provide further insight into the con-cept that certain agents of a chemical or biological natureare capable of evoking biochemical events characteristic ofthe natural plant disease resistance process from pathogeninfection.
Acknowledgements
AKS is thankful to Department of Science and Technol-ogy, New Delhi, for financial assistance (Grant SR/FTP/LS-22/2000).
References
Alstrom, S. 1991. Induction of disease resistance in common beansusceptible to halo blight bacterial pathogen after seed bacte-rization with rhizosphere pseudomonads. J. Gen. Appl.Microbiol. 37: 495–501.
Barka, E.A., Belarbi, A., Hachet, C., Nowak, J., and Audran, J.2000. Enhancement of in vitro growth and resistance to graymould of Vitis vinifera co-cultured with plant growth promotingrhizobacteria. FEMS Microbiol. Lett.186: 91–95.
Benhamou, N., and Nicole, M. 1999. Cell biology of plant immu-nization against microbial infection: the potential of induced re-sistance in controlling plant diseases. Plant Physiol. Biochem.37: 703–719.
Bhattacharya, D., Basu, S., Chattapadhyay, J.P., and Bose, S.K. 1985.Biocontrol ofMacrophominaroot/rot disease of jute by an antago-nistic organism,Aspergillus vesicolor. Plant Soil,87: 435–438.
Bull, C.T., Weller, D.M., and Thomashow, L.S. 1991. Relationshipbetween root colonization and suppression ofGaeumannomycesgraminis var. tritici by Pseudomonas fluorescensstrain 2–79.Phytophathology,81: 945–959.
Callow, J.A. 1999. Plant disease resistance: progress in basic un-derstanding and practical applications. Adv. Plant Pathol.30:357–389.
Cameron, R.K., Paiva, N.L., Lamb, C.J., and Dixon, R.A. 1999.Accumulation of salicylic acid and PR-1 gene transcripts in rela-tion to the systemic acquired resistance (SAR) response inducedby Pseudomonas syringaepv. tomato inArabidopsis. Physiol.Mol. Plant Pathol.55: 121–130.
Cardoso, J.E., Silva-Silvia, A.G., and Marques, E.E. 1997. Chemicaland biological control of bean root rots. Fitopatologia Brasileira,22: 39–44.
Chen, C., Belanger, R.R., Benhamou, N., and Paulitz, T.C. 1999.Role of salicylic acid in systemic resistance induced byPseudo-monasspp. againstPythium aphanidermatumin cucumber roots.Eur. J. Plant Pathol.105: 477–486.
Collins, D.J., Wyllie, T.B., and Anderson, S.H. 1991. Biological ac-tivity of Macrophomina phaseolinain soil. Soil Biol. Biochem.23: 495–496.
Daayf, F., Schmitt, A., and Belanger, R.R. 1997. Evidence of phyto-alexin in cucumber leaves infected with powdery mildew followingtreatment with leaf extracts ofReynoutria sachalinensis. PlantPhysiol.113: 719–727.
Dann, E.K., and Deverall, B.J. 1995. Effectiveness of systemic resis-tance in bean against foliar and soilborne pathogens as inducedby biological and chemical means. Plant Pathol.44: 458–466.
De Meyer, G., and Höfte, M. 1997. Salicylic acid produced by therhizobacteriumPseudomonas aeruginosa7NSK2 induce resis-tance to leaf infection byBotrytis cinerea on bean. Phyto-pathology,87: 588–593.
Delanay, T.P., Uknes, S., Vernooji, B., Freidich, L., Weymann, K.,Negrotto, D., Gaffney, T., Gut-Rella, M., Kessmann, H., Ward,E., and Ryals, J. 1994. A central role of salicylic acid in plantdisease resistance. Science (Washington D.C.),266: 1247–1250.
Dhingra, O.D., and Sinclair, J.B. 1985. Basic plant pathologymethods. CRC Press, Boca Raton, Fla.
Dubey, R.C. 1992. Effects of soil solarization on the survival ofMacrophomina phaseolinain fungicide amended and unamendedsoil. Trop. Sci.32: 225–279.
Duijff, B.J., Bakker, P.A.H.M., and Schippers, B. 1994. Suppressionof Fusariumwilt of carnation byPseudomonas putidaWCS358at different levels of disease and iron availability. Biocontrol Sci.Technol.4: 279–288.
© 2001 NRC Canada
Srivastava et al. 793
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:13 AM
Color profile: DisabledComposite Default screen

Enebak, S.A., and Carey, W.A. 2000. Evidence for induced systemicprotection to fusiform rust in loblolly pine by plant growth pro-moting rhizobacteria. Plant Dis.84: 306–308.
FAO. 1993. FAO yearbook production 1992. Vol. 46. Food and Ag-riculture Organization, Rome.
Gaffney, T., Freidrich, L., Vernooji, B., Negrotto, D., Nye, G.,Uknes, S., Ward, E., Kessmann, H., and Ryals, J. 1993. Require-ment of salicylic acid for the induction of systemic acquired re-sistance. Science (Washington D.C.),261: 754–756.
Hammerschmidt, R., and Kuc, J. 1995. Induced resistance to dis-ease in plants. Developments in plant pathology, Vol. 4. KluwerAcademic Publishers, Dordrecht, Netherlands.
Hoffland, E., Hakulinen, J., and Van Pelt, J.A. 1996. Comparisonof systemic resistance induced by avirulent and nonpathogenicPseudomonasspecies. Phytopathology,86: 757–762.
ICRISAT. 1988. Chickpea kaubuli variety ICCV6. Plant materialdescription. ICRISAT Bull. 12.
Ishi, H., Tomita, Y., Horio, T., Narusaka, Y., Nakazawa, Y.,Nishimura, K., and Iwamoto, S. 1999. Induced resistance ofacibenzolar-s-methyl (CGA 245704) to cucumber and Japanesepear diseases. Eur. J. Plant Pathol.105: 77–85.
Kataria, H.R., Wilmsmeier, B., and Buchenauer, H. 1997. Efficacyof resistance inducers, free-radical scavengers and an antagonis-tic strain ofPseudomonas fluorescensfor control ofRhizoctoniasolani AG-4 in bean and cucumber. Plant Pathol.46: 897–909.
Keller, H., Bonnet, P., Galiana, E., Pruvot, L., Freidrich, L., Ryals,J., and Ricci, P. 1996. Salicylic acid mediates elicitin-inducedsystemic acquired resistance, but not necrosis in tobacco. Mol.Plant–Microbe Interact.9: 696–703.
King, E.O., Ward, M.K., and Raney, D.E. 1954. Two simple mediafor the demonstration of pyocyanin and fluorescein. J. Lab. Clin.Med. 44: 301–307.
Koga, D., Sueshige, M., Orikono, K., Utsmi, T., Tanaka, S., Yamada,T., and Ide, A. 1988. Efficiency of chitinolytic enzymes in the for-mation of Trichoderma matsutakeprotoplasts. Agric. Biol. Chem.52: 2091–93.
Kuc, J. 1987. Plant immunization and its applicability for diseasecontrol. In Innovative approaches to plant disease control.Editedby I. Chet. John Wiley & Sons, New York. pp. 225–274.
Lam, S.T., and Gaffney, T.D. 1993. Biological activities of bacteriaused in plant pathogen control.In Biotechnology in plant dis-ease control. Wiley-Liss, Inc., New York. pp. 291–319.
Leeman, M., Van Pelt, J.A., Den Ouden, F.M., Heinsbroek, M.,Bakker, P.A.H.M., and Schippers, B. 1995. Induction of systemicresistance againstFusariumwilt of radish by lipopolysaccharidesof Pseudomonas fluorescens. Phytopathology,85: 1021–1027.
Liu, L., Kloepper, J.W., and Tuzun, S. 1995. Induction of systemicresistance in cucumber againstFusariumwilt by plant growth-promoting rhizobacteria. Phytopathology,85: 695–698.
Lodha, S. 1995. Soil solarization, summer irrigation and amend-ments for the control ofFusarium oxysporumf. sp. cumini andMacrophomina phaseolinain soils. Crop Prot.14: 215–219.
Loper, J.E., Corbell, N., Kraus, J., Thompson, B.N., Henkels, M.D.,and Carneige, S. 1994. Contributions of molecular biology towardsunderstanding the mechanisms by which rhizosphere pseudo-monads effect biological control.In Improving plant productivitywith rhizosphere bacteria.Edited byM.H. Ryder, P.M. Stephens,and G.D. Bowen. CSIRO, Australia. pp. 89–86.
Majumdar, V.L., Jat, J.R., and Gour, H.N. 1996. Effects of bio-control agents on the growth ofMacrophomina phaseolina, theincitant of blight of moth bean. Ind. J. Mycol. Plant Pathol.26:202–203.
Maurhofer, M., Hase, C., Meuwly, P., Metraux, J.P., and Defago,G. 1994. Induction of systemic resistance of tobacco to tobacco
necrosis virus by the root-colonizingPseudomonas fluorescensstrain CHAO: influence of thegacA gene and of pyoverdineproduction. Phytopathology,84: 139–146.
Meyer, D. G., Audenaert, K., and Hofte, M. 1999.Pseudomonasaeruginosa7NSK2-induced systemic resistance in tobacco dependson in planta salicyclic acid accumulation but is not associated withPR1a expression. Eur. J. Plant Pathol.105: 513–517.
Mihail, J.D., and Alcorn, S.M. 1984. Effects of soil solarization onMacrophomina phaseolinaandSclerotium rolfsii.Plant Dis.68:156–159.
Mihail, J.D., and Alcorn, S.M. 1987.Macrophomina phaseolinaspatial pattern in a cultivated soil and sampling strategies. Phyto-pathology,77: 1126–1131.
Mihail, J.D., and Taylor, S.D. 1995. Interpreting variability amongisolates ofMacrophomina phaseolinain pathogenicity, pycnidiumproduction, and chlorate utilization. Can. J. Bot.73: 1596–1603.
Okon, Y., Chet, I., and Henis, Y. 1973. Effect of lactose, ethanoland cyclohexamide on translocation pattern of radiactive com-pounds and on sclerotium formation inSclerotium rolfsii. J.Gen. Microbiol.74: 23–54.
Olanya, O.M., and Campbell, C.M. 1989. Density and spatial pat-tern of propagules ofMacrophomina phaseolinain corn rhizo-spheres. Phytopathology,79: 1119–1122.
Olaya, G., Abawi, G.S., and Barnard, J. 1996. Influence of waterpotential on survival of sclerotia in soil and on colonization ofbean stem segments byMacrophomina phaseolina.Plant Dis.80: 1351–1354.
Pandey, G., and Singh, R.B. 1990. Survey of root diseases ofchickpea in Allahabad region. Curr. Nematol.1: 77–78.
Pieterse, C.M., Van Wees, S.C.M., Hoffland, E., Van Pelt, J.A., andVan Loon, L.C. 1996. Systemic resistance inArabidopsis in-duced by biocontrol bacteria is independent of salicylic acid ac-cumulation and pathogenesis related gene expression. PlantCell, 8: 1225–1237.
Punja, Z.K., and Zhang, Y.Y. 1993. Plant chitinases and their rolesin resistance to fungal diseases. J. Nematol.25: 526–540.
Raupach, G.S., and Kloepper, J.W. 1997. Integrated pest manage-ment of multiple cucumber pathogens through PGPR-mediatedinduced systemic resistance. Proceedings of the 4th InternationalWorkshop on Plant Growth-Promoting Rhizobacteria. 5–10 Oct.1996, Sapporo, Japan. pp. 281–284.
Roberts, W.K., and Selitrennikoff, C.P. 1988. Plant and bacterialchitinases differ in antifungal activity. J. Gen. Microbiol.134:169–176.
Schlumbaum, A., Mauch, F., Vogeli, U., and Boller, T. 1986. Plantchitinases are potent inhibitors of fungal growth. Nature (Lon-don), 324: 365–367.
Siegrist, J., Glenewinkel, D., Kolle, C., and Schmidtke, M. 1997.Chemically induced resistance in green bean against bacterialand fungal pathogens. J. Plant Dis. Prot.104: 599–610.
Srivastava, A.K., Arora, D.K., Gupta, S., Pandey, R.R., and Lee,M.W. 1996. Diversity of potential antagonists colonizing sclerotiaof Macrophomina phaseolinain soil, and agglutination propertiesof some potential antagonists toward sclerotia. Biol. Fertil. Soils,22: 136–140.
Srivastava, A.K., Singh, T., Jana, T.K., and Arora, D.K. 2001. Micro-bial colonization ofMacrophomina phaseolinaand suppression ofcharcoal rot of chickpea.In Microbes and plants. CommonwealthPublishers, New Delhi, India. pp. 269–319.
Sticher, L., Mauch-Mani, B., and Metraux, J-P. 1997. Systemic ac-quired resistance. Annu. Rev. Phytopathol.35: 235–270.
Tuzun, S., and Bent, E. 1999. The role of hydrolytic enzymes inmultigenic and microbially induced resistance in plants. Inducedplant defenses against pathogens and herbivores: biochemistry,
© 2001 NRC Canada
794 Can. J. Bot. Vol. 79, 2001
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:14 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
Srivastava et al. 795
ecology and agriculture. American Phytopathological SocietyPress, St. Paul, Minn. pp. 95–115.
Van Loon, L.C., Baker, P., and Pieterse, C.M.J. 1998. Systemic re-sistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol.36: 453–483.
Van Peer, R., Neimann, G.J., and Schippers, B. 1991. Induced re-sistance and phytoalexin accumulation in biological control offusarium wilt of carnation byPseudomonas fluorescensstrainWCS417r. Phytopathology,81: 728–734.
Vernooij, B., Friedrich, L., Morse, A., Reist, R., Kolditz-Jawahar, R.,Ward, E., Uknes, S., Kessmann, H., and Ryals, J. 1994. Salicylic
acid is not the translocated signal responsible for inducing sys-temic acquired resistance but is required in signal transduction.Plant Cell,6: 959–965.
Vogt, W., and Buchenauer, H. 1997. Enhancement of biological con-trol by combination of antagonistic fluorescentPseudomonasstrains and resistance inducers against damping-off and powderymildew in cucumber. J. Plant Dis. Prot.104: 272–280.
Yedidia, I., Benhamou, N., and Chet, I. 1999. Induction of defenseresponse in cucumber plants (Cucumis sativusL.) by the bio-control agentTrichoderma harzianum. Appl. Environ. Microbiol.65: 1061–1070.
J:\cjb\cjb79\cjb-07\B01-054.vpWednesday, June 20, 2001 9:47:14 AM
Color profile: DisabledComposite Default screen





























