Embed Size (px)
Citation preview

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 1/11
doi:10.1152/ajpregu.00134.2004
287:R749-R758, 2004. First published 6 May 2004; Am J Physiol Regul Integr Comp PhysiolMorganHarriet Andersén, Jan Tornell, Claes Ohlsson, Mike Snaith and David G. A.Annika Åstrand, Mohammad Bohlooly-Y, Sara Larsdotter, Margit Mahlapuu,altered autonomic activity1 demonstrate increased heart rate associated withMice lacking melanin-concentrating hormone receptor
You might find this additional info useful...
46 articles, 26 of which can be accessed free at:This article cites
http://ajpregu.physiology.org/content/287/4/R749.full.html#ref-list-1
4 other HighWire hosted articlesThis article has been cited by
[PDF][Full Text][Abstract]
2006; 55 (2): 428-434. DiabetesPissios and Eleftheria Maratos-FlierJustin Y. Jeon, Richard L. Bradley, Efi G. Kokkotou, Francis E. Marino, Xiaomei Wang, PavlosResistance
Mice Are Resistant to Aging-Associated Increases in Body Weight and Insulin− / −MCH
[PDF][Full Text][Abstract]2006; 55 (3): 725-733. Diabetes
Oscarsson, Michael Snaith, Jan Törnell and Mohammad Bohlooly-YMikael Bjursell, Anna-Karin Gerdin, Karolina Ploj, David Svensson, Lennart Svensson, JanObese Leptin-Deficient Mice Without Affecting Body WeightMelanin-Concentrating Hormone Receptor 1 Deficiency Increases Insulin Sensitivity in
[PDF][Full Text][Abstract]2007; 56 (3): 583-593. Diabetes
Anna-Lena Berg, Jan Oscarsson and Daniel LindénRhedin, Xiao-Rong Peng, Karolina Ploj, Anna-Karin Gerdin, Gunnel Arnerup, Anders Elmgren,Mikael Bjursell, Andrea Ahnmark, Mohammad Bohlooly-Y, Lena William-Olsson, MagdalenaOpposing Effects of Adiponectin Receptors 1 and 2 on Energy Metabolism
[PDF][Full Text][Abstract]
, January , 2011; 300 (1): R47-R55. Am J Physiol Regul Integr C omp PhysiolMahesh Karnani and Denis BurdakovMultiple hypothalamic circuits sense and regulate glucose levels
including high resolution figures, can be found at:Updated information and services
http://ajpregu.physiology.org/content/287/4/R749.full.html
can be found at: and Comparative Physiology American Journal of Physiology - Regulatory, IntegrativeaboutAdditional material and information
http://www.the-aps.org/publications/ajpregu
This information is current as of May 2, 2012.
ISSN: 0363-6119, ESSN: 1522-1490. Visit our website at http://www.the-aps.org/.Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2004 by the American Physiological Society.
ranging from molecules to humans, including clinical investigations. It is published 12 times a year (monthly) by the Americanilluminate normal or abnormal regulation and integration of physiological mechanisms at all levels of biological organization,
publishes original investigations that American Journal of Physiology - Regulatory, Integrative and Comparative Physiology

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 2/11
Mice lacking melanin-concentrating hormone receptor 1 demonstrate
increased heart rate associated with altered autonomic activity
Annika Astrand,1 Mohammad Bohlooly-Y,2 Sara Larsdotter,1 Margit Mahlapuu,2
Harriet Andersen,2 Jan Tornell,2 Claes Ohlsson,3 Mike Snaith,2 and David G. A. Morgan1
1 Department of Integrative Pharmacology and 2 AstraZeneca Transgenic and ComparativeGenomics Centre, AstraZeneca Research and Development, SE-431 83 Molndal; and 3 Department of Internal Medicine, Sahlgrenska University Hospital, SE-413 45 Gothenburg Sweden
Submitted 27 February 2004; accepted in final form 22 April 2004
Astrand, Annika, Mohammad Bohlooly-Y, Sara Larsdotter,Margit Mahlapuu, Harriet Andersen, Jan Tornell, Claes Ohlsson,Mike Snaith, and David G. A. Morgan. Mice lacking melanin-concentrating hormone receptor 1 demonstrate increased heart rateassociated with altered autonomic activity. Am J Physiol Regul Integr
Comp Physiol 287: R749–R758, 2004. First published May 6, 2004;10.1152/ajpregu.00134.2004.—Melanin-concentrating hormone (MCH)plays an important role in energy balance. The current studies were
carried out on a new line of mice lacking the rodent MCH receptor(MCHR1 / mice). These mice confirmed the previously reported leanphenotype characterized by increased energy expenditure and modestlyincreased caloric intake. Because MCH is expressed in the lateral hypo-thalamic area, which also has an important role in the regulation of theautonomic nervous system, heart rate and blood pressure were measuredby a telemetric method to investigate whether the increased energyexpenditure in these mice might be due to altered autonomic nervoussystem activity. Male MCHR1 / mice demonstrated a significantlyincreased heart rate [24-h period: wild type 495 4 vs. MCHR1 /
561 8 beats/min (P 0.001); dark phase: wild type 506 8 vs.MCHR1 / 582 9 beats/min (P 0.001); light phase: wild type484 13 vs. MCHR1 / 539 9 beats/min (P 0.005)] with nosignificant difference in mean arterial pressure [wild type 110 0.3 vs.MCHR1 / 113 0.4 mmHg (P 0.05)]. Locomotor activity and core
body temperature were higher in the MCHR1 / mice during the dark phase only and thus temporally dissociated from heart rate differences.On fasting, wild-type animals rapidly downregulated body temperatureand heart rate. MCHR1 / mice displayed a distinct delay in the onset of this downregulation. To investigate the mechanism underlying thesedifferences, autonomic blockade experiments were carried out. Admin-istration of the adrenergic antagonist metoprolol completely reversed thetachycardia seen in MCHR1 / mice, suggesting an increased sympa-thetic tone.
melanin-concentrating hormone; sympathetic; fasting
MELANIN-CONCENTRATING HORMONE (MCH) is a peptide synthe-sized exclusively in the lateral hypothalamic area and zonaincerta (6, 40) that has been implicated in the hypothalamicregulation of energy balance. Levels of MCH mRNA areregulated by altered energy balance in animal models such asfasting and in models with impaired leptin signaling (30).Intracerebroventricular (30) and intrahypothalamic (1) injec-tion of MCH increases food intake, and mice overexpressingMCH are hyperphagic and obese (22). Conversely, mice lack-ing a functioning MCH gene are hypophagic and lean (37).
Two receptors for MCH have been identified. The first,MCHR1, was originally characterized as an orphan receptor,SLC-1 (15). MCH was subsequently identified as the cognateligand by a number of groups (5, 10, 21, 35, 38). It isdistributed extensively in the central nervous system, with adistribution consistent with a role in energy balance (34). Itdemonstrates high sequence homology among species investi-gated so far. The second receptor, MCHR2, was identified by
a number of groups using low-stringency homology searchingor 5 /3-rapid amplification of cDNA ends (3, 15, 24, 31, 33,44). This receptor is expressed as a functional receptor in dogs,nonhuman primates, and humans but is expressed as a pseu-dogene in rabbits and guinea pigs and is absent in rodentspecies (42).
Recently two groups have reported the phenotype of micelacking the only known rodent receptor for MCH (11, 23).These have a similar phenotype to those lacking MCH itself.However, one striking difference is the lack of hypophagia inMCHR1 null mice; instead these mice eat slightly more thantheir wild-type littermates, suggesting that the lean phenotypeof these animals is, to a large degree, brought about by anincrease in energy expenditure. Indeed, a significantly raised24-h energy expenditure has been recorded in these animalsusing indirect calorimetry.
Increasing circumstantial evidence suggests that MCH mayalter energy balance not only by increasing food intake but alsoby decreasing energy expenditure consistent with an alteredlevel of autonomic nervous system activity. Thus a recentreport suggests that intracerebroventricular MCH might de-crease body temperature and brown fat uncoupling protein(UCP-1) (16), whereas ob / ob mice lacking MCH showed anincreased level of brown fat UCP-1 and an increased bodytemperature compared with control ob / ob animals (36). Inaddition, sympathetic projections to a number of organs havebeen retrogradely traced to the lateral hypothalamic area, and
projections to the brown fat have even been traced to MCH-containing cells (26).
In this study we investigate diurnal variations in energyexpenditure of MCHR1 / mice and compare it to variationsin heart rate and blood pressure activity. We have also inves-tigated the autonomic mechanisms underlying differences be-tween MCHR1 / mice and wild-type controls, using phar-macological blockade of sympathetic and parasympathetic ner-vous systems.
Address for reprint requests and other correspondence: D. G. A. Morgan,Dept. of Integrative Pharmacology, AstraZeneca R&D Molndal, S-431 83Molndal, Sweden (E-mail [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement ”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Regul Integr Comp Physiol 287: R749–R758, 2004.First published May 6, 2004; 10.1152/ajpregu.00134.2004.
0363-6119/04 $5.00 Copyright © 2004 the American Physiological Societyhttp://www.ajpregu.org R749

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 3/11
Fasting at an ambient temperature below thermoneutralityresults in large transient depressions in heart rate, mean arterialpressure, and oxygen consumption in mice (46). This responseis thought to be mediated in part via a reduction in sympatheticnervous system activity, and so changes in this response canindicate changes in sympathetic nervous system output. Toinvestigate this we have compared the cardiovascular re-
sponses to fasting in MCHR1 / mice and wild-type controls.
MATERIALS AND METHODS
Animal care. Knockout animals were generated using R1 ES cells(derived from 129/SvJ). All animals used in these studies had beenbackcrossed for a minimum of three generations to C57Bl/6. Controlanimals used in the experiments were always the wild-type littermatesof the MCHR1 / mice. Mice were maintained under a standard 12-hlight cycle regime, the relative humidity was between 45 and 55%, thetemperature was kept at 20°C, and the animals had ad libitum accessto chow and water, with environmental enrichment (in the form of eggcartons, wooden tongue depressors, and cotton nesting pads) presentin home cages unless otherwise stated. The study was performed afterapproval from the local ethical committee for animal experimentation.
Generation of MCHR1 null mice. The mouse MCHR1 locus wassubcloned from a positive BAC identified by screening a 129/SvJgenomic library. The targeting construct was generated by replacingthe entire MCHR1 coding sequence with a neomycin phosphotrans-ferase (Neo) cassette driven off the phosphoglycerate kinase (PGK)promoter. After linearization, the targeting construct was electropo-rated into R1 ES cells and neomycin-resistant clones were selected inG-418-containing media (300 g/ml). Of 250 G418-resistant clonesscreened, four targeted clones were identified using a Southern screen-ing strategy with an EcoR1- BamHI fragment 5 to the targetedsequence as a probe. One positive clone was expanded and injectedinto C57Bl/6 blastocysts to generate chimeric mice. Chimeric maleswere crossed to C57Bl/6 females, and genotyping of the offspring wasperformed from tail biopsies by triplex PCR using a common (for-ward) primer located at the 3-end of the short arm (5-cattgagaacag-
tacccctggctt-3), a reverse primer specific for the exon 2 of thewild-type allele (5-ctagatccacctgttactcagagga-3), and a reverseprimer in the PGK promoter specific for the targeted allele (5-agcgcatgctccagactgcctt-3).
To verify that the deletion of MCHR1 coding sequences resulted ina null mutation, total RNA was prepared from the brains of 4-wk-oldhomozygous, heterozygous, and control mice using the RNASTAT-60 Kit according to the manufacturer’s instructions (Tel-Test,Friendswood, TX). cDNA was synthesized using Superscript TM IIRNase H-Reverse Transcriptase and random hexamer primers (LifeTechnologies, Frederick, MD). Real-time semiquantitative PCR wasperformed in the ABI PRISM 7700 Sequence Detector System (Ap-plied Biosystems, Warrington, UK) using SYBR Green. All sampleswere run in triplicate, and data were normalized using the mouseacidic ribosomal phosphoprotein PO (M36B4) as an internal control.
The primers for real-time PCR specific for the MCHR1 were derivedfrom rat SLC-1 sequences (20): 5-tcagcttgggctatgctaacag-3 (for-ward) and 5-caacaccaagcgttttcgaa-3 (reverse).
Measurement of body weight and food intake. Mice used forweight-gain studies were housed three to four per cage and given freeaccess to standard mouse chow containing 4% of total energy as fat(R-34; Lactamin AB, Stockholm, Sweden) or to cafeteria diet, an adlibitum choice of full-fat cheese (38% fat), almond paste, milk chocolate, and pastry (based on vegetarian butter, sugar, rolled oats,cocoa, and coconut shreds). From 4 wk of age, mice were weighedweekly to the nearest 0.1 g on an electronic scale.
Food intake was measured in 6-wk-old single-housed mice for 6consecutive days. The mice were housed individually 3 wk beforefood consumption was measured. Mice were fed standard mouse
chow (see above), which had a total energy content of 3.0 kcal/g or ahigh-fat diet. Cafeteria diet consisted of a number of human food-stuffs, which rapidly became contaminated and dispersed in the cagesof animals fed this diet. Therefore, we turned to an alternative high-fatdiet for measurement of food intake. This contained 45% of totalenergy as fat (D12451; Research Diets, New Brunswick, NJ; a totalenergy content of 4.7 kcal/g). Cages were carefully monitored forspillage of food, which was undetectable.
Measurements of the body composition. The total fat content of 16-wk-old mice fed standard chow diet was measured by dual-energyX-ray absorptiometry (DEXA) as previously described (39) using theNorland pDEXA Sabre (Fort Atkinson, WI).
Indirect calorimetry. Oxygen consumption (VO2), carbon dioxideproduction (VCO2), and activity were measured using an open-circuitcalorimetry system (Oxymax, Columbus Instruments International,Columbus, OH). The animals were placed in calorimeter chamberswith ad libitum access to normal lab chow and water for 72 h.Environmental enrichment was removed. An air sample was with-drawn for 75 s every 20 min, and the O2 and CO2 content weremeasured by a paramagnetic oxygen sensor and a spectrophotometricCO2 sensor. These values were used to calculate VO2 and VCO2. Datafrom the first 24 h were not used in analysis of results to allowacclimatization to the novel environment. Data from corresponding
hours during the second and third 24-h periods were combined in 1-hbins. After the 3-day period in the Oxymax system, the body compo-sition of each animal was measured by DEXA, and the lean body masswas calculated. Metabolic rate (kcal/h) was calculated using a rear-rangement of the Weir equation (45) as supplied by ColumbusInstruments: (3.815 1.232RER) VO2, where RER is the respira-tory exchange ratio [volume of CO2 produced per volume of O2
consumed (both ml kg1min1)] and VO2 is the volume of O2
consumed per hour. Data for the last two 24-h periods of the exper-iment were combined and analyzed by two-way ANOVA. An eight-channel activity monitor, forming part of the Oxymax system, pro-vided ambulatory counts for each channel.
Remote measurement of temperature, heart rate, and blood pres-
sure. Core body temperature, heart rate, mean arterial pressure, andlocomotor activity were continuously monitored using remote telem-
etry transmitters (DSI, St. Paul, MN) on individually housed 8-wk-oldmale mice at room temperature. Transmitters were implanted intra-abdominally under isoflurane (Forene) anesthesia. For measurementof blood pressure, a catheter was inserted in the carotid artery. Micewere allowed to recover for at least 14 days postsurgery. All mea-surements were carried out in the home-cage environment, and envi-ronmental enrichment was available in all studies except those involv-ing fasting.
In one experiment, knockout (n 7) or wild-type (n 8) malemice were implanted with PA-C20 transmitters (DSI). Data on heartrate and mean arterial pressure were collected every 30 s and averagedin 10-min bins over a 72-h period. The average for each bin from thesame point over 3 days was calculated, and these values were used toestimate 24-h diurnal rhythms. A further experiment was carried outusing knockout (n 8) or wild-type (n 8) male mice implanted
with ETA-F20 transmitters (DSI) to monitor baseline body tempera-ture. As mean arterial pressure showed only small variations, ETA-F20 transmitters, which deliver ECG-data as well as body tempera-ture, were used for further experiments described below.
Another set of experiments was designed to compare autonomiccontrol of heart rate in MCHR1 / and wild-type mice. Animals wereimplanted with ETA-F20 telemetry transmitters and allowed to re-cover as above before undergoing four treatments, each separated bya recovery period of at least 3 days. On each treatment day animalswere randomly assigned to one of four treatment groups and receivedintraperitoneal administration of either 0.9% saline vehicle (10 ml/ kg), methylscopolamine (0.5 mg/kg), metoprolol (5 mg/kg), or acombination of methylscopolamine (0.5 mg/kg) and metoprolol (5mg/kg), so that each animal received all four treatments in random
R750 INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org
8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 4/11
order. The doses of metoprolol and methylscopolamine were chosenafter consultation with the literature (e.g., Refs. 17–19) and in-housedose-finding experiments in wild-type mice from the same supplier.Data were collected from each animal for a 2-h baseline period andthen for a further 2 h after intraperitoneal injection of the autonomicblocker. Data were averaged over 15-min bins, and the data for eachbin were compared by one-way ANOVA and post hoc Tukey ’s test.
In the final experiment, data were collected for 72 h from wild-type(n 7) and MCHR1 / (n 8) mice implanted with ETA-F20telemetry transmitters. Recording was started at 1600 on day 1, andbaseline measurements were recorded for 24 h, during which timeanimals had ad libitum access to food and water. Food was removedat 1600 on the second day, and animals were fasted for 24 h. Accessto water was ad libitum during this period. Their recovery from thefast was monitored for 24 h, starting at 1600 on day 3. Average heartrate for each animal was compared under fasted and nonfastedconditions. Episodes of reduced heart rate and body temperature weredefined as 5-min bins in which the average value was 3 SDs fromthe mean value seen during the dark period on the previous nonfastedday. The total number of such bins and the time to onset of the firstepisode were calculated for each animal, and the average values forwild-type and knockout animals were compared by t -test.
RESULTS
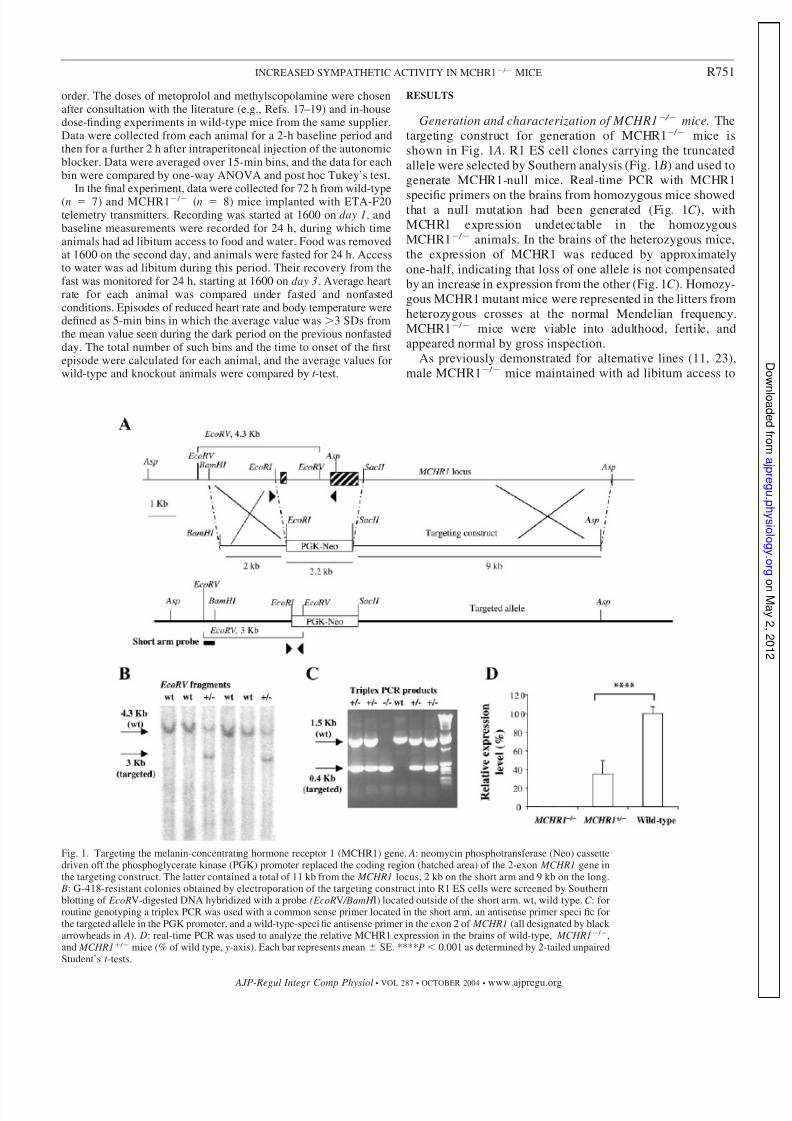
Generation and characterization of MCHR1 / mice. The
targeting construct for generation of MCHR1 / mice is
shown in Fig. 1 A. R1 ES cell clones carrying the truncated
allele were selected by Southern analysis (Fig. 1 B) and used to
generate MCHR1-null mice. Real-time PCR with MCHR1
specific primers on the brains from homozygous mice showedthat a null mutation had been generated (Fig. 1C ), with
MCHR1 expression undetectable in the homozygous
MCHR1 / animals. In the brains of the heterozygous mice,
the expression of MCHR1 was reduced by approximately
one-half, indicating that loss of one allele is not compensated
by an increase in expression from the other (Fig. 1C ). Homozy-
gous MCHR1 mutant mice were represented in the litters from
heterozygous crosses at the normal Mendelian frequency.
MCHR1 / mice were viable into adulthood, fertile, and
appeared normal by gross inspection.
As previously demonstrated for alternative lines (11, 23),
male MCHR1 / mice maintained with ad libitum access to
Fig. 1. Targeting the melanin-concentrating hormone receptor 1 (MCHR1) gene. A: neomycin phosphotransferase (Neo) cassettedriven off the phosphoglycerate kinase (PGK) promoter replaced the coding region (hatched area) of the 2-exon MCHR1 gene inthe targeting construct. The latter contained a total of 11 kb from the MCHR1 locus, 2 kb on the short arm and 9 kb on the long.
B: G-418-resistant colonies obtained by electroporation of the targeting construct into R1 ES cells were screened by Southernblotting of EcoRV-digested DNA hybridized with a probe (EcoRV /BamH I) located outside of the short arm. wt, wild type. C : forroutine genotyping a triplex PCR was used with a common sense primer located in the short arm, an antisense primer speci fic forthe targeted allele in the PGK promoter, and a wild-type-speci fic antisense primer in the exon 2 of MCHR1 (all designated by black arrowheads in A). D: real-time PCR was used to analyze the relative MCHR1 expression in the brains of wild-type, MCHR1 / ,and MCHR1 / mice (% of wild type, y-axis). Each bar represents mean SE. ****P 0.001 as determined by 2-tailed unpairedStudent’s t -tests.
R751INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org
8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 5/11
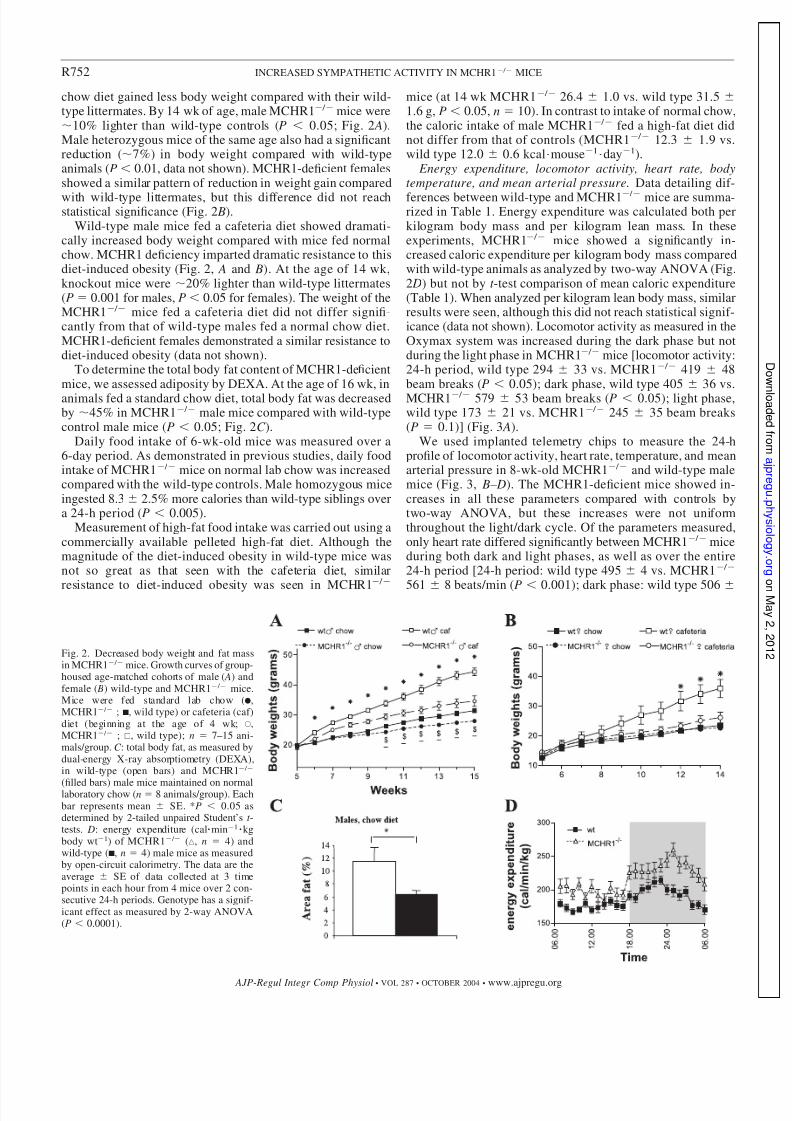
chow diet gained less body weight compared with their wild-type littermates. By 14 wk of age, male MCHR1 / mice were10% lighter than wild-type controls (P 0.05; Fig. 2 A).Male heterozygous mice of the same age also had a significantreduction (7%) in body weight compared with wild-typeanimals (P 0.01, data not shown). MCHR1-deficient femalesshowed a similar pattern of reduction in weight gain compared
with wild-type littermates, but this difference did not reachstatistical significance (Fig. 2 B).
Wild-type male mice fed a cafeteria diet showed dramati-cally increased body weight compared with mice fed normalchow. MCHR1 deficiency imparted dramatic resistance to thisdiet-induced obesity (Fig. 2, A and B). At the age of 14 wk,knockout mice were 20% lighter than wild-type littermates(P 0.001 for males, P 0.05 for females). The weight of theMCHR1 / mice fed a cafeteria diet did not differ signifi-cantly from that of wild-type males fed a normal chow diet.MCHR1-deficient females demonstrated a similar resistance todiet-induced obesity (data not shown).
To determine the total body fat content of MCHR1-deficientmice, we assessed adiposity by DEXA. At the age of 16 wk, inanimals fed a standard chow diet, total body fat was decreasedby 45% in MCHR1 / male mice compared with wild-typecontrol male mice (P 0.05; Fig. 2C ).
Daily food intake of 6-wk-old mice was measured over a6-day period. As demonstrated in previous studies, daily foodintake of MCHR1 / mice on normal lab chow was increasedcompared with the wild-type controls. Male homozygous miceingested 8.3 2.5% more calories than wild-type siblings overa 24-h period (P 0.005).
Measurement of high-fat food intake was carried out using acommercially available pelleted high-fat diet. Although themagnitude of the diet-induced obesity in wild-type mice wasnot so great as that seen with the cafeteria diet, similar
resistance to diet-induced obesity was seen in MCHR1 /
mice (at 14 wk MCHR1 / 26.4 1.0 vs. wild type 31.5 1.6 g, P 0.05, n 10). In contrast to intake of normal chow,the caloric intake of male MCHR1 / fed a high-fat diet didnot differ from that of controls (MCHR1 / 12.3 1.9 vs.wild type 12.0 0.6 kcal mouse1
day1). Energy expenditure, locomotor activity, heart rate, body
temperature, and mean arterial pressure. Data detailing dif-
ferences between wild-type and MCHR1 /
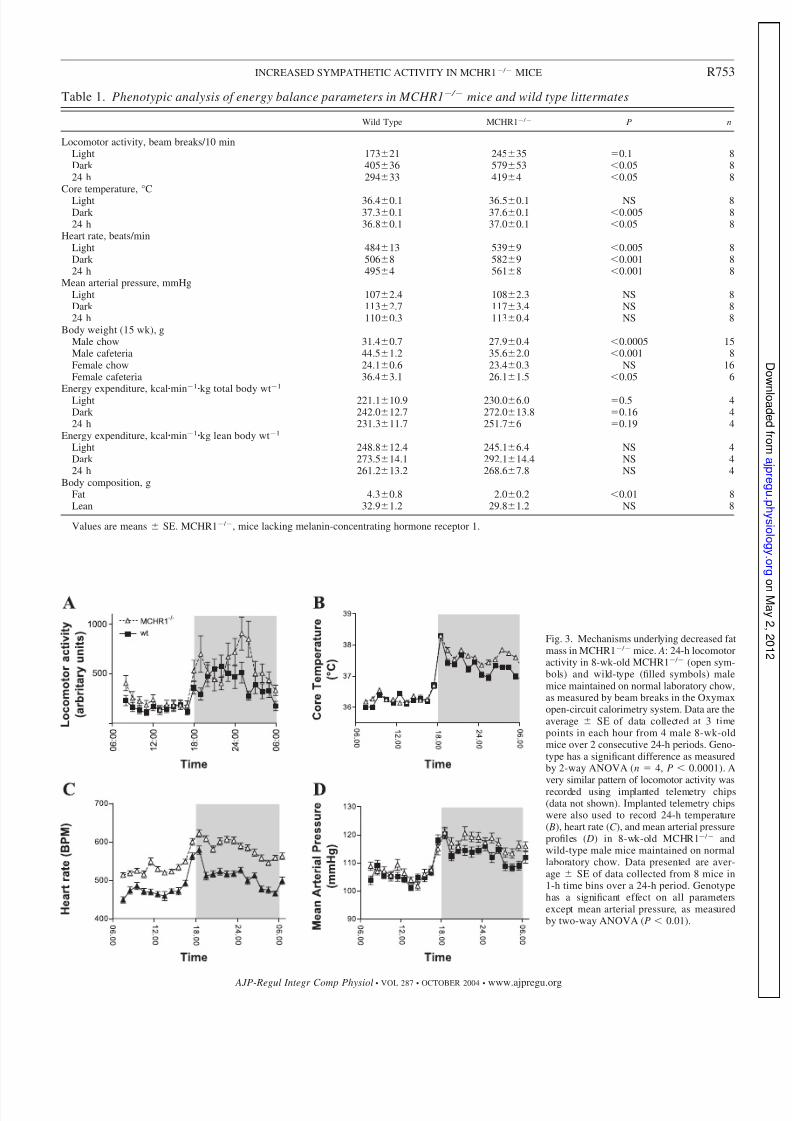
mice are summa-rized in Table 1. Energy expenditure was calculated both perkilogram body mass and per kilogram lean mass. In theseexperiments, MCHR1 / mice showed a significantly in-creased caloric expenditure per kilogram body mass comparedwith wild-type animals as analyzed by two-way ANOVA (Fig.2 D) but not by t -test comparison of mean caloric expenditure(Table 1). When analyzed per kilogram lean body mass, similarresults were seen, although this did not reach statistical signif-icance (data not shown). Locomotor activity as measured in theOxymax system was increased during the dark phase but notduring the light phase in MCHR1 / mice [locomotor activity:24-h period, wild type 294 33 vs. MCHR1 / 419 48beam breaks (P 0.05); dark phase, wild type 405 36 vs.MCHR1 / 579 53 beam breaks (P 0.05); light phase,wild type 173 21 vs. MCHR1 / 245 35 beam breaks(P 0.1)] (Fig. 3 A).
We used implanted telemetry chips to measure the 24-hprofile of locomotor activity, heart rate, temperature, and meanarterial pressure in 8-wk-old MCHR1 / and wild-type malemice (Fig. 3, B – D). The MCHR1-deficient mice showed in-creases in all these parameters compared with controls bytwo-way ANOVA, but these increases were not uniformthroughout the light/dark cycle. Of the parameters measured,only heart rate differed significantly between MCHR1 / miceduring both dark and light phases, as well as over the entire24-h period [24-h period: wild type 495 4 vs. MCHR1 /
561 8 beats/min (P 0.001); dark phase: wild type 506
Fig. 2. Decreased body weight and fat massin MCHR1 / mice. Growth curves of group-housed age-matched cohorts of male ( A) andfemale ( B) wild-type and MCHR1 / mice.Mice were fed standard lab chow (F,MCHR1 / ; s, wild type) or cafeteria (caf)diet (beginning at the age of 4 wk; E,MCHR1 / ; , wild type); n 7–15 ani-mals/group. C : total body fat, as measured bydual-energy X-ray absorptiometry (DEXA),in wild-type (open bars) and MCHR1 /
(filled bars) male mice maintained on normallaboratory chow (n 8 animals/group). Eachbar represents mean SE. *P 0.05 asdetermined by 2-tailed unpaired Student’s t -tests. D: energy expenditure (cal min1
kgbody wt1) of MCHR1 / (‚, n 4) andwild-type (s, n 4) male mice as measuredby open-circuit calorimetry. The data are theaverage SE of data collected at 3 timepoints in each hour from 4 mice over 2 con-secutive 24-h periods. Genotype has a signif-icant effect as measured by 2-way ANOVA(P 0.0001).
R752 INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org
8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 6/11
Fig. 3. Mechanisms underlying decreased fatmass in MCHR1 / mice. A: 24-h locomotoractivity in 8-wk-old MCHR1 / (open sym-bols) and wild-type (filled symbols) malemice maintained on normal laboratory chow,as measured by beam breaks in the Oxymaxopen-circuit calorimetry system. Data are theaverage SE of data collected at 3 timepoints in each hour from 4 male 8-wk-oldmice over 2 consecutive 24-h periods. Geno-type has a significant difference as measuredby 2-way ANOVA (n 4, P 0.0001). Avery similar pattern of locomotor activity wasrecorded using implanted telemetry chips(data not shown). Implanted telemetry chipswere also used to record 24-h temperature( B), heart rate (C ), and mean arterial pressureprofiles ( D) in 8-wk-old MCHR1 / andwild-type male mice maintained on normallaboratory chow. Data presented are aver-age SE of data collected from 8 mice in1-h time bins over a 24-h period. Genotypehas a significant effect on all parametersexcept mean arterial pressure, as measuredby two-way ANOVA (P 0.01).
Table 1. Phenotypic analysis of energy balance parameters in MCHR1 / mice and wild type littermates
Wild Type MCHR1 / P n
Locomotor activity, beam breaks/10 minLight 17321 24535 0.1 8Dark 40536 57953 0.05 824 h 29433 4194 0.05 8
Core temperature, °CLight 36.40.1 36.50.1 NS 8Dark 37.30.1 37.60.1 0.005 824 h 36.80.1 37.00.1 0.05 8
Heart rate, beats/minLight 48413 5399 0.005 8Dark 5068 5829 0.001 824 h 4954 5618 0.001 8
Mean arterial pressure, mmHgLight 1072.4 1082.3 NS 8Dark 1132.7 1173.4 NS 824 h 1100.3 1130.4 NS 8
Body weight (15 wk), gMale chow 31.40.7 27.90.4 0.0005 15Male cafeteria 44.51.2 35.62.0 0.001 8Female chow 24.10.6 23.40.3 NS 16Female cafeteria 36.43.1 26.11.5 0.05 6
Energy expenditure, kcal
min1
kg total body wt1
Light 221.110.9 230.06.0 0.5 4Dark 242.012.7 272.013.8 0.16 424 h 231.311.7 251.76 0.19 4
Energy expenditure, kcalmin1kg lean body wt1
Light 248.812.4 245.16.4 NS 4Dark 273.514.1 292.114.4 NS 424 h 261.213.2 268.67.8 NS 4
Body composition, gFat 4.30.8 2.00.2 0.01 8Lean 32.91.2 29.81.2 NS 8
Values are means SE. MCHR1 / , mice lacking melanin-concentrating hormone receptor 1.
R753INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org
8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 7/11
8 vs. MCHR1 / 582 9 beats/min (P 0.001); light phase:wild type 484 13 vs. MCHR1 / 539 9 beats/min (P 0.005)]. The telemetry chip system was a much less sensitivemeasure of locomotor activity than the Oxymax system, with anumber of bins during the light phase recording no locomotoractivity and no significant difference (NS) in the averagenumber of movements between genotypes [24-h period: wild
type 3.57 0.18 vs. MCHR1 /
5.06 0.22 (P NS); dark phase: wild type 5.33 0.28 vs. MCHR1 / 7.82 0.34 (PNS); light phase: wild type 1.83 0.20 vs. MCHR1 / 2.33 0.25 (P NS)]. Nevertheless, a very similar overall patternwas recorded, with genotype giving a significant effect bytwo-way ANOVA, and with locomotor activity increased dur-ing the dark phase to a greater extent than during the lightphase in MCHR1 / mice. Body temperature was increasedduring the dark phase but not the light phase in MCHR1 /
mice [24-h period: wild type 36.8 0.1 vs. MCHR1 /
37.0 0.1°C (P 0.05); dark phase: wild type 37.3 0.1 vs.MCHR1 / 37.6 0.1°C (P 0.005); light phase: wild type36.4 0.1 vs. MCHR1 / 36.5 0.1°C (P NS)]. Average
mean arterial pressure showed no significant difference be-tween MCHR1 / mice and wild-type mice [24-h period: wildtype 110 0.3 vs. MCHR1 / 113 0.4 mmHg (P 0.05)].
Thus there appeared to be temporal differentiation betweenthe effects on temperature, mean arterial pressure, and loco-motor activity, where the difference was more marked duringthe dark phase, and the effect on heart rate, which was apparent
throughout the 24-h period. As the effect on mean arterialpressure was relatively minor, telemetry chips measuring heartrate but not mean arterial pressure were used for experimentson the intrinsic heart rate and fasting responses of MCHR1 /
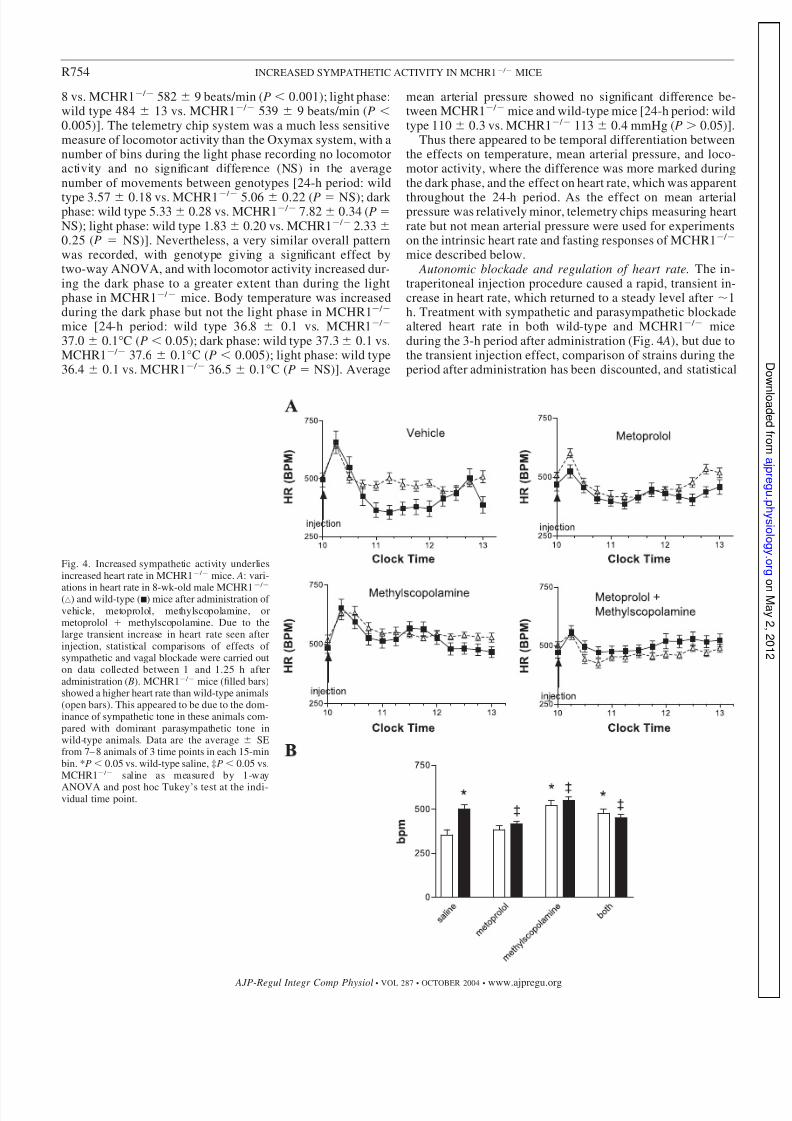
mice described below. Autonomic blockade and regulation of heart rate. The in-
traperitoneal injection procedure caused a rapid, transient in-crease in heart rate, which returned to a steady level after 1h. Treatment with sympathetic and parasympathetic blockadealtered heart rate in both wild-type and MCHR1 / miceduring the 3-h period after administration (Fig. 4 A), but due tothe transient injection effect, comparison of strains during theperiod after administration has been discounted, and statistical
Fig. 4. Increased sympathetic activity underliesincreased heart rate in MCHR1 / mice. A: vari-
ations in heart rate in 8-wk-old male MCHR1 /
(‚) and wild-type (s) mice after administration of vehicle, metoprolol, methylscopolamine, ormetoprolol methylscopolamine. Due to thelarge transient increase in heart rate seen afterinjection, statistical comparisons of effects of sympathetic and vagal blockade were carried outon data collected between 1 and 1.25 h afteradministration ( B). MCHR1 / mice (filled bars)showed a higher heart rate than wild-type animals(open bars). This appeared to be due to the dom-inance of sympathetic tone in these animals com-pared with dominant parasympathetic tone inwild-type animals. Data are the average SEfrom 7– 8 animals of 3 time points in each 15-minbin. *P 0.05 vs. wild-type saline, ‡P 0.05 vs.MCHR1 / saline as measured by 1-way
ANOVA and post hoc Tukey’s test at the indi-vidual time point.
R754 INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org
8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 8/11
analysis was carried out on data gathered between 1 and 1.25 hpostinjection (Fig. 4 B). During this period, intraperitonealvehicle-treated male MCHR1 / mice had a significantlyhigher heart rate than wild-type animals receiving a similartreatment [wild type 355 30 vs. MCHR1 / 502 24beats/min (P 0.001)]. Treatment with the sympatheticblocker metoprolol had no effect on the heart rate of wild-type
animals but significantly reduced the tachycardia seen inMCHR1 / mice [wild type 385 30 beats/min (P 0.05 vs.wild type saline); MCHR1 / 417 15 beats/min (P 0.01vs. knockout saline; P 0.05 vs wild type metoprolol)].Treatment with methylscopolamine significantly increased theheart rate of wild-type animals and also caused a significantincrease in the knockout animals [wild type 522 28 beats/ min (P 0.001 vs. wild type saline); MCHR1 / 550 22beats/min (P 0.01 vs. MCHR1 / saline; P 0.05 vs. wildtype methylscopolamine)]. Treatment with a combination of methylscopolamine and metoprolol showed that there was nosignificant difference in intrinsic heart rate between wild-typeand MCHR1 / mice and that this was intermediate betweenthe basal heart rates of the two mouse strains [wild type 477 23 beats/min (P 0.001 vs. wild type saline); MCHR1 /
451 21 beats/min (P 0.01 vs. MCHR1 / saline; P 0.05 vs. wild type methylscopolamine metoprolol)].
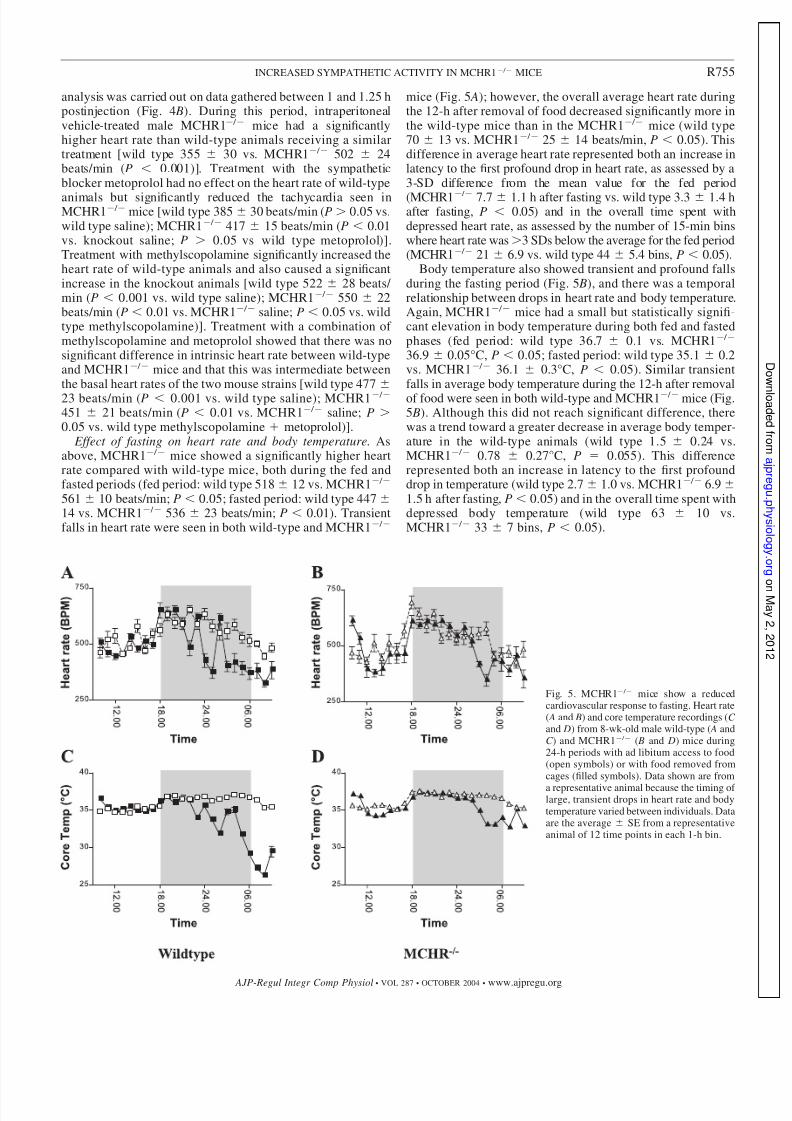
Effect of fasting on heart rate and body temperature. Asabove, MCHR1 / mice showed a significantly higher heartrate compared with wild-type mice, both during the fed andfasted periods (fed period: wild type 518 12 vs. MCHR1 /
561 10 beats/min; P 0.05; fasted period: wild type 447 14 vs. MCHR1 / 536 23 beats/min; P 0.01). Transientfalls in heart rate were seen in both wild-type and MCHR1 /
mice (Fig. 5 A); however, the overall average heart rate duringthe 12-h after removal of food decreased significantly more inthe wild-type mice than in the MCHR1 / mice (wild type70 13 vs. MCHR1 / 25 14 beats/min, P 0.05). Thisdifference in average heart rate represented both an increase inlatency to the first profound drop in heart rate, as assessed by a3-SD difference from the mean value for the fed period
(MCHR1 /
7.7 1.1 h after fasting vs. wild type 3.3 1.4 hafter fasting, P 0.05) and in the overall time spent withdepressed heart rate, as assessed by the number of 15-min binswhere heart rate was3 SDs below the average for the fed period(MCHR1 / 21 6.9 vs. wild type 44 5.4 bins, P 0.05).
Body temperature also showed transient and profound fallsduring the fasting period (Fig. 5 B), and there was a temporalrelationship between drops in heart rate and body temperature.Again, MCHR1 / mice had a small but statistically signifi-cant elevation in body temperature during both fed and fastedphases (fed period: wild type 36.7 0.1 vs. MCHR1 /
36.9 0.05°C, P 0.05; fasted period: wild type 35.1 0.2vs. MCHR1 / 36.1 0.3°C, P 0.05). Similar transientfalls in average body temperature during the 12-h after removalof food were seen in both wild-type and MCHR1 / mice (Fig.5 B). Although this did not reach significant difference, therewas a trend toward a greater decrease in average body temper-ature in the wild-type animals (wild type 1.5 0.24 vs.MCHR1 / 0.78 0.27°C, P 0.055). This differencerepresented both an increase in latency to the first profounddrop in temperature (wild type 2.7 1.0 vs. MCHR1 / 6.91.5 h after fasting, P 0.05) and in the overall time spent withdepressed body temperature (wild type 63 10 vs.MCHR1 / 33 7 bins, P 0.05).
Fig. 5. MCHR1 / mice show a reducedcardiovascular response to fasting. Heart rate( A and B) and core temperature recordings (C and D) from 8-wk-old male wild-type ( A andC ) and MCHR1 / ( B and D) mice during24-h periods with ad libitum access to food(open symbols) or with food removed fromcages (filled symbols). Data shown are from
a representative animal because the timing of large, transient drops in heart rate and bodytemperature varied between individuals. Dataare the average SE from a representativeanimal of 12 time points in each 1-h bin.
R755INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 9/11
DISCUSSION
In this series of studies we show that a third line of MCHR1 / mice has a similar phenotype to that of the
MCHR1 / mice lines previously described by Marsh et al.(23) and Chen et al. (11). All three lines have a lean phenotypeand are resistant to diet-induced obesity. In each case, in-
creased energy expenditure appears to be an important factor(11, 23) because MCHR1 / mice show a slight increase in
food consumption. We also demonstrate that the absence of afunctional MCHR1 system leads to an increased basal heartrate independent of increased locomotor activity and that this
appears to be caused by increased sympathetic and decreasedparasympathetic activity compared with wild-type animals. We
also demonstrate that under fasting conditions MCHR1 /
mice maintain heart rate and body temperature for longer thanwild-type mice before the initiation of transient but profound
drops in these two parameters, a response normally involvingreduced sympathetic activity. It thus appears that the MCHR1
system is involved in both the tonic regulation of autonomicactivity and the fast-induced autonomic suppression of heartrate and body temperature.
MCH has been considered an orexigenic peptide, with in-tracerebroventricular injection causing increased food intake
(10, 32), and MCH antagonists causing acute (41) and chronicreductions (7) of food intake. Thus it was an unexpected resultthat mice lacking the MCHR1 receptor should be hyperphagic
compared with wild-type animals. It is plausible that theincrease in food intake is due to compensatory mechanisms
acting to rectify the negative energy balance in these animals,and one might even speculate that the compensatory hyperpha-gia is somewhat impaired because MCHR1 / mice show only
a modest increase in food intake, inadequate to compensatefully and maintain a “normal” wild-type body weight. Thus it
is possible that the slight compensatory hyperphagia might bemasking an effect on food intake responsible in part for thelean phenotype in these animals. This explanation would be in
keeping with the reports of reduced food intake after admin-istration of MCHR1 antagonists (7, 41). It is interesting to notethat in the semichronic study reported by Borowsky et al. (7),
there appears to be a steady reduction in the effect of the MCHantagonist SNAP-7941 on food intake, while no such normal-
ization of body weight loss is seen.MCHR1 / mice show an increase in dark-phase locomotor
activity, accompanied by increased body temperature, in keep-
ing with previous reports (11, 23). However, energy expendi-ture, as measured by open-circuit calorimetry, is increased
during the light phase as well as the dark phase, whereas noincreased locomotor activity is seen during the light phase.This suggests that, while increased locomotor activity might be
responsible for part of the increased energy expenditure, anincrease in basal metabolic rate, or involuntary energy expen-
diture is also likely to be present. Such an increase in invol-untary energy expenditure is likely to be regulated by theautonomic nervous system, and in keeping with altered auto-
nomic regulation we also observed an increase in heart rate inMCHR1 / mice. Like the increase in energy expenditure, this
mild tachycardia was present throughout the light/dark cycle,and thus did not appear to be wholly caused by increasedlocomotor activity.
Autonomic control of heart rate. In many cases, factorsaltering food intake also have a reciprocal effect on energyexpenditure (for review, see Ref. 8). In many cases the sym-pathetic nervous system plays a major role in such changes inenergy expenditure. The sympathetic nervous system is madeup of multiple differentially regulated segments, and heart ratein particular can be regulated by a number of these, including
several that do not appear important in regulation of energybalance (see Ref. 25 for review). However, under a number of circumstances, heart rate and thermogenesis are under coordi-nate autonomic control, and specific areas of the brain, such asthe raphe pallidus, have been shown to control both (9). Thusit appears that under some circumstances heart rate and brownfat thermogenesis might be controlled by the same single unitof the sympathetic nervous system (8). Because both parame-ters appear altered in MCHR1 / mice, it seems at leastpossible that the same mechanisms underlie the increased heartrate and a proportion of the increased energy expenditure inMCHR1 / mice. We therefore investigated the autonomicmechanisms underlying the increased heart rate. To do this wemeasured the effects of sympathetic and parasympatheticblockade on heart rate in MCHR1 / mice using metoprololand methylscopolamine. This also allowed us to differentiatebetween autonomic regulation and intrinsic heart rate of MCHR1 / and wild-type mice.
Directly after injection, we observed a transient stress-induced increase in heart rate. This transient increase in heartrate disappeared within 1 h of administration, and heart ratethen returned to a steady level. We compared the effects of sympathetic and parasympathetic blockade during this latersteady period. Intrinsic heart rate, as measured in the presenceof both sympathetic and parasympathetic blockade, was thesame for both MCHR1 / and wild-type mice and lay inter-mediate to the baseline levels for the two strains.
In both MCHR1 /
and wild-type mice, heart rate wascontrolled by a balance between sympathetic and parasympa-thetic inputs. In MCHR1 / mice, it appeared that the sympa-thetic nervous system was the predominant input. Thus admin-istration of the sympathetic blocker metoprolol reduced heartrate to the level seen in wild-type mice. However, whenMCHR1 / mice were treated with both metoprolol and meth-ylscopolamine, they showed a heart rate significantly higherthan when treated with metoprolol alone, demonstrating thatparasympathetic tone was present in these animals that hadbeen unmasked by metoprolol treatment.
Conversely, in wild-type mice, it appeared that parasympa-thetic inputs predominated. Thus treatment with methylscopol-amine alone increased heart rate in wild-type mice to the same
level as seen in MCHR1 /
mice. Again, it was clear thatparasympathetic blockade unmasked a tonic sympathetic ac-tivity in wild-type mice because treatment with both metopro-lol and methylscopolamine induced a heart rate significantlylower than that seen after treatment with methylscopolaminealone. In summary, in this paradigm, ablation of the MCHR1gene leads to a state where sympathetic rather than parasym-pathetic control of heart rate predominates. This is in keepingwith the changes expected if altered autonomic regulation wereresponsible for the changes in energy expenditure in thesemice.
It is often stated that sympathetic activity is the predominantautonomic controller of heart rate in mice (for review, see Ref.
R756 INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 10/11
18). However, contrary to this finding, we see a predominanceof parasympathetic control of heart rate in wild-type mice inour study. A careful review of the literature shows that this isnot a unique finding. Indeed there are studies showing aspectrum of autonomic control from predominantly sympa-thetic (12, 17) to a balance between sympathetic and parasym-pathetic (43) to predominantly parasympathetic control (2).
Other than the differences in autonomic control, perhaps themost pertinent observed difference between these studies is inthe resting heart rate. Not surprisingly, the higher the restingheart rate is, the larger is the sympathetic component in thecontrol of heart rate. For example, whereas we observed aresting heart rate of approximately 350– 400 beats/min inwild-type animals, Gehrmann et al. (12), using a similartelemetry system, reported resting heart rates over 700 beats/ min. A number of variables might contribute to these variationsin resting heart rate. These include the different methods of sampling heart rate data (telemetry vs. externalized catheters);the length of time between operation and measurements; ad-ministration, novel environment, and handling stress-inducedeffects; strain, age, and gender effects; and effects of theambient temperature at which the experiments were carriedout. Of these factors, ambient temperature seems a possibleunreported variant between studies. Thus in our study althoughthe room temperature was 20°C, well below the thermoneutraltemperature for most mice strains, the animals did have accessto nesting material in the cages, in keeping with local ethicalguidelines and in an attempt to make the surroundings asnormal as possible. It seems quite likely that the mice werethus able to maintain thermoneutrality and that this maintainsthe parasympathetic balance of the autonomic inputs. Nestingmaterial was removed during fasting experiments, and thisperhaps explains the smaller differences in heart rate seenbetween knockout and wild-type animals in these experiments.
Responses to fasting. Fasting induces a distinctive pattern of responses in rodents. This includes reductions in heart rate,arterial pressure, VO2, and body temperature, as well as aninitial hyperactivity followed by periods of hypoactivity (28,46, 47). A large body of indirect evidence suggests thatdecreased sympathetic activity and increased parasympatheticactivity underlie this process (4, 13, 14, 27). Given that we hadseen altered autonomic activity in MCHR1 / mice, we hy-pothesized that the MCH system might play an important rolein mediating the autonomic changes leading to altered cardio-vascular parameters during fasting. To test this we measuredheart rate and body temperature changes in MCHR1 / andwild-type mice during a 24-h fast. In this model we observeda reduced response to fasting in MCHR1 / mice.
The cardiovascular elements of the fasting response are notconstant but instead occur as transient bouts of profoundsuppression of parameters such as heart rate and body temper-ature (46). In MCHR1 / mice we observed that both thelatency to onset of these bouts and the overall time withdepressed heart rate and body temperature were reduced inMCHR1 / mice. However, we did observe that the typicalcardiovascular response to fasting was eventually seen. Thusthe data suggest that the MCHR1 system is an importantregulator of the physiological responses to fasting (as itsremoval slowed the response) but does not represent the oneand only mechanism (since the response still eventually oc-curred).
A number of hypothalamic peptide systems that are knownto alter food intake also have reciprocal effects on sympatheticnervous activity (see Ref. 8 for review), a coordinate energy-conserving response to negative energy balance. From thismodel, it would appear that MCH can be added to that number.It appears that MCH may play an important role in the balanceof autonomic nervous system activity, as in its absence there
appears to be a switch from predominantly parasympatheticcontrol to sympathetic control. It could be also be speculatedthat increases in MCH, which occur during fasting and foodrestriction, play a role in the suppression of sympatheticactivity in these states. It remains to be investigated whetherdirect intracerebroventricular injection of MCH causes a sup-pression of sympathetic activity and heart rate in rodents,although intracerebroventricular MCH has been shown to haveno effect on heart rate in sheep (29). It also remains unprovenwhether the increased heart rate seen in MCHR1 / mice isrelated to an increase in brown fat thermogenesis in theseanimals.
In summary, it appears that the MCH system may be animportant tonic regulator of sympathetic nervous activity. Thismay form part of a coordinated response to negative energybalance and may be of importance when considering modula-tion of this system as a target for body weight control.
REFERENCES
1. Abbott CR, Kennedy AR, Wren AM, Rossi M, Murphy KG, Seal LJ,Todd JF, Ghatei MA, Small CJ, and Bloom SR. Identification of hypothalamic nuclei involved in the orexigenic effect of melanin-concen-trating hormone. Endocrinology 144: 3943–3949, 2003.
2. Altman JD, Trendelenburg AU, MacMillan L, Bernstein D, LimbirdL, Starke K, Kobilka BK, and Hein L. Abnormal regulation of thesympathetic nervous system in 2A-adrenergic receptor knockout mice.
Mol Pharmacol 56: 154 –161, 1999.3. An S, Cutler G, Zhao JJ, Huang SG, Tian H, Li W, Liang L, Rich M,
Bakleh A, Du J, Chen JL, and Dai K. Identification and characterizationof a melanin-concentrating hormone receptor. Proc Natl Acad Sci USA 98:7576 –7581, 2001.
4. Avakian EV and Horvath SM. Starvation suppresses sympathoadrenalmedullary response to cold exposure in rats. Am J Physiol Endocrinol
Metab 241: E316 –E320, 1981.5. Bachner D, Kreienkamp H, Weise C, Buck F, and Richter D. Identi-
fication of melanin concentrating hormone (MCH) as the natural ligand forthe orphan somatostatin-like receptor 1 (SLC-1). FEBS Lett 457: 522–524,1999.
6. Bittencourt JC, Presse F, Arias C, Peto C, Vaughan J, Nahon JL, ValeW, and Sawchenko PE. The melanin-concentrating hormone system of the rat brain: an immuno- and hybridization histochemical characteriza-tion. J Comp Neurol 319: 218 –245, 1992.
7. Borowsky B, Durkin MM, Ogozalek K, Marzabadi MR, DeLeon J,Lagu B, Heurich R, Lichtblau H, Shaposhnik Z, Daniewska I, Black-burn TP, Branchek TA, Gerald C, Vaysse PJ, and Forray C. Antide-
pressant, anxiolytic and anorectic effects of a melanin-concentratinghormone-1 receptor antagonist. Nat Med 8: 825– 830, 2002.8. Bray GA. Reciprocal relation of food intake and sympathetic activity:
experimental observations and clinical implications. Int J Obes Relat
Metab Disord 24: S8 –S17, 2000.9. Cao WH and Morrison SF. Disinhibition of rostral raphe pallidus
neurons increases cardiac sympathetic nerve activity and heart rate. Brain
Res 980: 1–10, 2003.10. Chambers J, Ames RS, Bergsma D, Muir A, Fitzgerald LR, Hervieu
G, Dytko GM, Foley JJ, Martin J, Liu WS, Park J, Ellis C, GangulyS, Konchar S, Cluderay J, Leslie R, Wilson S, and Sarau HM.Melanin-concentrating hormone is the cognate ligand for the orphanG-protein-coupled receptor SLC-1. Nature 400: 261–265, 1999.
11. Chen Y, Hu C, Hsu CK, Zhang Q, Bi C, Asnicar M, Hsiung HM, FoxN, Slieker LJ, Yang DD, Heiman ML, and Shi Y. Targeted disruptionof the melanin-concentrating hormone receptor-1 results in hyperphagia
R757INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org

8/2/2019 Increased Sympathetic Activity in Mchr1 Ko Mice
http://slidepdf.com/reader/full/increased-sympathetic-activity-in-mchr1-ko-mice 11/11
and resistance to diet-induced obesity. Endocrinology 143: 2469 –2477,2002.
12. Gehrmann J, Hammer PE, Maguire CT, Wakimoto H, Triedman JK,and Berul CI. Phenotypic screening for heart rate variability in the mouse.
Am J Physiol Heart Circ Physiol 279: H733–H740, 2000.13. Gotoh M, Iguchi A, Tajima T, Ikari H, and Hirooka Y. Starvation
reduces norepinephrine activities in both hypothalamus and heart in rats. Brain Res 706: 351–354, 1996.
14. Grassi G, Seravalle G, Colombo M, Bolla G, Cattaneo BM, Cavagnini
F, and Mancia G. Body weight reduction, sympathetic nerve traf fic, andarterial baroreflex in obese normotensive humans. Circulation 97: 2037–2042, 1998.
15. Hill J, Duckworth M, Murdock P, Rennie G, Sabido-David C, AmesRS, Szekeres P, Wilson S, Bergsma DJ, Gloger IS, Levy DS, Cham-bers JK, and Muir AI. Molecular cloning and functional characterizationof MCH2, a novel human MCH receptor. J Biol Chem 276: 20125–20129,2001.
16. Ito M, Gomori A, Ishihara A, Oda Z, Mashiko S, Matsushita H,
Yumoto M, Sano H, Tokita S, Moriya M, Iwaasa H, and Kanatani A.Characterization of MCH-mediated obesity in mice. Am J Physiol Endo-
crinol Metab 284: E940 –E945, 2003.17. Janssen BJ, Leenders PJ, and Smits JF. Short-term and long-term blood
pressure and heart rate variability in the mouse. Am J Physiol Regul Integr
Comp Physiol 278: R215–R225, 2000.18. Janssen BJ and Smits JF. Autonomic control of blood pressure in mice:
basic physiology and effects of genetic modification. Am J Physiol Regul Integr Comp Physiol 282: R1545–R1564, 2002.19. Johansson C, Gothe S, Forrest D, Vennstrom B, and Thoren P.
Cardiovascular phenotype and temperature control in mice lacking thyroidhormone receptor- or both 1 and . Am J Physiol Heart Circ Physiol
276: H2006 –H2012, 1999.20. Lakaye B, Minet A, Zorzi W, and Grisar T. Cloning of the rat brain
cDNA encoding for the SLC-1 G protein-coupled receptor reveals thepresence of an intron in the gene. Biochim Biophys Acta 1401: 216 –220,1998.
21. Lembo PM, Grazzini E, Cao J, Hubatsch DA, Pelletier M, Hoffert C,St-Onge S, Pou C, Labrecque J, Groblewski T, O’Donnell D, Payza K,
Ahmad S, and Walker P. The receptor for the orexigenic peptidemelanin-concentrating hormone is a G-protein-coupled receptor. Nat Cell
Biol 1: 267–271, 1999.22. Ludwig DS, Tritos NA, Mastaitis JW, Kulkarni R, Kokkotou E,
Elmquist J, Lowell B, Flier JS, and Maratos-Flier E. Melanin-concen-
trating hormone overexpression in transgenic mice leads to obesity andinsulin resistance. J Clin Invest 107: 379 –386, 2001.
23. Marsh DJ, Weingarth DT, Novi DE, Chen HY, Trumbauer ME, ChenAS, Guan XM, Jiang MM, Feng Y, Camacho RE, Shen Z, Frazier EG,Yu H, Metzger JM, Kuca SJ, Shearman LP, Gopal-Truter S, MacNeil
DJ, Strack AM, MacIntyre DE, Van der Ploeg LH, and Qian S.Melanin-concentrating hormone 1 receptor-deficient mice are lean, hyper-active, and hyperphagic and have altered metabolism. Proc Natl Acad SciUSA 99: 3240 –3245, 2002.
24. Mori M, Harada M, Terao Y, Sugo T, Watanabe T, Shimomura Y,Abe M, Shintani Y, Onda H, Nishimura O, and Fujino M. Cloning of a novel G protein-coupled receptor, SLT, a subtype of the melanin-concentrating hormone receptor. Biochem Biophys Res Commun 283:1013–1018, 2001.
25. Morrison SF. Differential control of sympathetic outflow. Am J Physiol Regul Integr Comp Physiol 281: R683–R698, 2001.
26. Oldfield BJ, Giles ME, Watson A, Anderson C, Colvill LM, and
McKinley MJ. The neurochemical characterisation of hypothalamic path-ways projecting polysynaptically to brown adipose tissue in the rat.
Neuroscience 110: 515–526, 2002.27. Overton JM, VanNess JM, and Casto RM. Food restriction reduces
sympathetic support of blood pressure in spontaneously hypertensive rats. J Nutr 127: 655– 660, 1997.
28. Overton JM, Williams TD, Chambers JB, and Rashotte ME. Centralleptin infusion attenuates the cardiovascular and metabolic effects of fasting in rats. Hypertension 37: 663– 669, 2001.
29. Parkes DG. Diuretic and natriuretic actions of melanin concentratinghormone in conscious sheep. J Neuroendocrinol 8: 57– 63, 1996.
30. Qu D, Ludwig DS, Gammeltoft S, Piper M, Pelleymounter MA,Cullen MJ, Mathes WF, Przypek R, Kanarek R, and Maratos-Flier E.
A role for melanin-concentrating hormone in the central regulation of
feeding behaviour. Nature 380: 243–247, 1996.31. Rodriguez M, Beauverger P, Naime I, Rique H, Ouvry C, Souchaud S,
Dromaint S, Nagel N, Suply T, Audinot V, Boutin JA, and Galizzi JP.
Cloning and molecular characterization of the novel human melanin-concentrating hormone receptor MCH2. Mol Pharmacol 60: 632– 639,2001.
32. Rossi M, Choi SJ, O’Shea D, Miyoshi T, Ghatei MA, and Bloom SR.
Melanin-concentrating hormone acutely stimulates feeding, but chronicadministration has no effect on body weight. Endocrinology 138: 351–355, 1997.
33. Sailer AW, Sano H, Zeng Z, McDonald TP, Pan J, Pong SS, Feighner
SD, Tan CP, Fukami T, Iwaasa H, Hreniuk DL, Morin NR, Sadowski
SJ, Ito M, Bansal A, Ky B, Figueroa DJ, Jiang Q, Austin CP, MacNeil
DJ, Ishihara A, Ihara M, Kanatani A, Van der Ploeg LH, Howard AD,
and Liu Q. Identification and characterization of a second melanin-
concentrating hormone receptor, MCH-2R. Proc Natl Acad Sci USA 98:7564 –7569, 2001.
34. Saito Y, Cheng M, Leslie FM, and Civelli O. Expression of the
melanin-concentrating hormone (MCH) receptor mRNA in the rat brain. J Comp Neurol 435: 26 – 40, 2001.
35. Saito Y, Nothacker HP, Wang Z, Lin SH, Leslie F, and Civelli O.
Molecular characterization of the melanin-concentrating-hormone recep-tor. Nature 400: 265–269, 1999.
36. Segal-Lieberman G, Bradley RL, Kokkotou E, Carlson M, Trombly
DJ, Wang X, Bates S, Myers MG Jr, Flier JS, and Maratos-Flier E.Melanin-concentrating hormone is a critical mediator of the leptin-de fi-
cient phenotype. Proc Natl Acad Sci USA 100: 10085–10090, 2003.37. Shimada M, Tritos NA, Lowell BB, Flier JS, and Maratos-Flier E.
Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature 396: 670 – 674, 1998.
38. Shimomura Y, Mori M, Sugo T, Ishibashi Y, Abe M, Kurokawa T,
Onda H, Nishimura O, Sumino Y, and Fujino M. Isolation andidentification of melanin-concentrating hormone as the endogenous ligandof the SLC-1 receptor. Biochem Biophys Res Commun 261: 622– 626,
1999.39. Sjogren K, Hellberg N, Bohlooly YM, Savendahl L, Johansson MS,
Berglindh T, Bosaeus I, and Ohlsson C. Body fat content can bepredicted in vivo in mice using a modified dual-energy X-ray absorpti-
ometry technique. J Nutr 131: 2963–2966, 2001.40. Skofitsch G, Jacobowitz DM, and Zamir N. Immunohistochemical
localization of a melanin concentrating hormone-like peptide in the ratbrain. Brain Res Bull 15: 635– 649, 1985.
41. Takekawa S, Asami A, Ishihara Y, Terauchi J, Kato K, Shimomura Y,
Mori M, Murakoshi H, Suzuki N, Nishimura O, and Fujino M.
T-226296: a novel, orally active and selective melanin-concentratinghormone receptor antagonist. Eur J Pharmacol 438: 129 –135, 2002.
42. Tan CP, Sano H, Iwaasa H, Pan J, Sailer AW, Hreniuk DL, Feighner
SD, Palyha OC, Pong SS, Figueroa DJ, Austin CP, Jiang MM, Yu H,
Ito J, Ito M, Guan XM, MacNeil DJ, Kanatani A, Van der Ploeg LH,
and Howard AD. Melanin-concentrating hormone receptor subtypes 1
and 2: species-specific gene expression. Genomics 79: 785–792, 2002.43. Uechi M, Asai K, Osaka M, Smith A, Sato N, Wagner TE, Ishikawa
Y, Hayakawa H, Vatner DE, Shannon RP, Homcy CJ, and Vatner SF.
Depressed heart rate variability and arterial baroreflex in conscious trans-
genic mice with overexpression of cardiac Gs. Circ Res 82: 416 – 423,1998.
44. Wang S, Behan J, O’Neill K, Weig B, Fried S, Laz T, Bayne M,
Gustafson E, and Hawes BE. Identification and pharmacological char-acterization of a novel human melanin-concentrating hormone receptor,mch-r2. J Biol Chem 276: 34664 –34670, 2001.
45. Weir J. New methods for calculating metabolic rate with special referenceto protein. J Physiol 109: 1, 1949.
46. Williams TD, Chambers JB, Henderson RP, Rashotte ME, and Over-
ton JM. Cardiovascular responses to caloric restriction and thermoneu-trality in C57BL/6J mice. Am J Physiol Regul Integr Comp Physiol 282:R1459 –R1467, 2002.
47. Williams TD, Chambers JB, May OL, Henderson RP, Rashotte ME,
and Overton JM. Concurrent reductions in blood pressure and metabolicrate during fasting in the unrestrained SHR. Am J Physiol Regul Integr
Comp Physiol 278: R255–R262, 2000.
R758 INCREASED SYMPATHETIC ACTIVITY IN MCHR1 / MICE
AJP-Regul Integr Comp Physiol • VOL 287 • OCTOBER 2004 • www.ajpregu.org