Embed Size (px)
Citation preview

Increased bone loss and amountof osteoclasts in kinin B1receptor knockout miceGoncalves-Zillo TO, Pugliese LS, Sales VMT, da Silva Mori MA, Squaiella-BaptistaoCC, Longo-Maugeri IM, Lopes JD, de Oliveira SM, Monteiro AC, Pesquero JB.Increased bone loss and amount of osteoclasts in kinin B1 receptor knockout mice.J Clin Periodontol 2013; 40: 653–660. doi: 10.1111/jcpe.12097.
AbstractAim: The pathophysiology of periodontal diseases involves aspects of immunityand bone remodelling. Considering the role of the kinin B1 receptor (Bdkrb1) ininflammation and healing, the purpose of this study was to evaluate the contribu-tion of Bdkrb1 to the pathogenesis of periodontitis.Material and Methods: We used a model of ligature-induced experimental peri-odontitis (LIEP) in mice lacking Bdkrb1 (Bdkrb1�/�) to test the role of thisreceptor in bone loss and cytokine secretion by lymph nodes cells. Angiotensin-converting enzyme inhibitor (ACEi) was used as a pharmacological strategy tosupport the genetic model. Also, autonomous effect of Bdkrb1 deletion wasevaluated in osteoclasts precursors from bone marrow.Results: Bdkrb1�/� mice exhibit increased bone loss and IL-17 secretion inresponse to LIEP when compared to wild type. LIEP does not modify TNF-a,IFN-c and IL-10 levels in Bdkrb1�/� mice after 21 days. Bone marrow cells fromBdkrb1�/� displayed increased differentiation into functional osteoclasts withconsistent artificial calcium phosphate degradation. Furthermore, treatment ofmice with ACEi prevented bone destruction.Conclusion: Bdkrb1 participates in the pathogenesis of LIEP bone loss possiblythrough mechanisms that involve modulation of the TH17 response, therebydemonstrating its role in the development of periodontitis.
Thais Oliveira Goncalves-Zillo1, LıviaSouza Pugliese2, Vicencia Micheline
Toledo Sales1, Marcelo Alves daSilva Mori1, Carla Cristina
Squaiella-Baptistao3,Ieda Maria Longo-Maugeri2,
Jose Daniel Lopes2, Suzana Macedode Oliveira1, Ana Carolina
Monteiro4 and Joao BoscoPesquero1
1Departamento de Biofısica, Universidade
Federal de Sao Paulo, Sao Paulo, Brazil;2Disciplina de Imunologia, Departamento de
Microbiologia Imunologia e Parasitologia,
Universidade Federal de Sao Paulo, Sao
Paulo, Brazil; 3Laborat�orio de Imunoquımica,
Instituto Butantan, Sao Paulo, Brazil;4Instituto Nacional de Cancer CPQ, MEDEX,
Rio de Janeiro, Brazil
Key words: Bdkrb1; bone loss; kinin;
osteoclast; periodontitis
Accepted for publication 11 February 2013
The pharmacologically active peptidesof the kinin group are primarymediators of inflammation. They arereleased in tissue and blood as a con-
sequence of kininogen cleavage bykallikrein and are processed by pep-tidases (Leeb-Lundberg et al. 2005).Kinins exert their broad action by theactivation of the two transmembraneG protein-coupled receptors, namedkinin B1 (Bdkrb1) and B2 (Bdkrb2)receptors (Pesquero et al. 1996). Inmice, bradykinin (BK) binds toBdkrb2 while des-Arg9BK, a productof subsequent cleavage of BK, actsvia Bdkrb1. On the other hand,angiotensin-converting enzyme(ACE) degrades kinins into inactivefragments, decreasing kinin availabil-
ity in circulation (Regoli & Barabe1980).
Bdkrb2 is constitutivelyexpressed and widely distributed intissues, including gingival fibroblastsand human dental pulp (Bruniuset al. 2005), mediating the majorityof the physiological effects ofkinins. In turn, Bdkrb1 is shown tobe almost absent in most cells andtissues, but is up-regulated inresponse to injury and inflammation(Leeb-Lundberg et al. 2005). Con-stitutive expression of Bdkrb1 recep-tor has been demonstrated in
Conflict of interest and source offunding statement
This study was supported by ConselhoNacional de Desenvolvimento Cientıficoe Tecnol�ogico (CNPq) and Fundacaode Amparo �a Pesquisa do Estado deSao Paulo (FAPESP – Proc. 2008/06676-8). The authors declare that theyhave no conflict of interests.
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd 653
J Clin Periodontol 2013; 40: 653–660 doi: 10.1111/jcpe.12097

neutrophils (Ara�ujo et al. 2001) andosteoblasts MC3T3-E1 (Ljunggren& Lerner 1990).
Among the processes in whichBdkrb1 is up-regulated, it is worthmentioning the autoimmune dis-eases. In this case, the expression ofBdkrb1 mRNA is increased inT lymphocytes (Prat et al. 1999).It was also demonstrated that bothBdkrb1 and Bdkrb2 are up-regulatedin human osteoblast-lineage cellsMG-63, human fibroblasts and miceosteoblasts by IL-1b or TNF-a(Brechter et al. 2008).
According to Monteiro et al.(2006), endogenous kinins released ininflammatory sites are capable ofmodulating innate and adaptiveimmunity through cooperationbetween toll-like receptors (TLR)2and Bdkrb2. Data from Bdkrb2�/�
mice suggest a down-regulation of Thelper (TH)1 response (IFN-c produc-tion), associated with up-regulation ofTH17 response (IL-17 production) insplenocytes of Trypanosoma cruzi-infected mice lacking Bdkrb2 (Monte-iro et al. 2007). Bdkrb1, on the otherhand, has been shown to be involvedin the regulation of the central ner-vous system (CNS) inflammation(Schulze-Topphoff et al. 2009). Asreported, activation of Bdkrb1 limitsencephalitogenic T lymphocyte infil-tration into the CNS, leading tomilder symptoms of encephalomyeli-tis, whereas blocking of Bdkrb1 leadsto accelerated disease onset.
Although the knowledge aboutthe role of kinins in the pathogenesisof periodontal disease is still sparse,it has been gradually assembled.Gustafson & Lerner (1984) publishedthe very first strong evidence of therole of kinins in bone metabolism.In this study, BK was able to stimu-late mineral mobilization and releaseof lysosomal-enzymes in culturedmouse calvaria. It was demonstratedthat the concomitant generation ofthrombin and BK in areas of traumaand inflammation may induceresorption of bone tissue neighbour.Different kinins were demonstratedto have the same effect in culturedbone reinforcing the theory thatkinins can stimulate bone resorption(Lerner, 1994).
Periodontal attachment lossoccurs due to bacterially derivedfactors and antigens that stimulatea local inflammatory reaction and
activate the innate and acquiredimmune system. This situation leadsto the activation of osteoclasts andconsequently to periodontal attach-ment loss (Graves & Cochran 2003,Teng 2006). Osteoclast precursorcells proliferation and survival aredependent on the growth factorM-CSF (Pixley & Stanley 2004). Inturn, receptor activator of nuclearfactor kB-ligand (RANKL) isrequired to induce expression ofgenes that typify the osteoclast line-age, including those encodingtartrate-resistant acid phosphatase(TRAP), cathepsin K (CATK), calci-tonin receptor and the b3-integrin,leading to the development ofmature osteoclasts (Lacey et al.1998, Boyle et al. 2003). Both M-CSF and RANKL demonstrated tobe produced by fibroblasts, osteo-blasts, macrophages and T cells
(Felix et al. 1990, Simonet et al.1997, Kostenuik & Shalhoub 2001,Hodge et al. 2007) and are involvedin periodontal inflammation process.
Taken together, these indicativedata led us to propose the hypothesisthat the kallikrein-kinin system partici-pates in osteoclastogenesis and in theimmune modulation of periodontaldisease, therefore contributing to itspathogenesis. To assess this hypothe-sis, we developed a method of ligature-induced experimental periodontitis(LIEP) in mice to test the progressionof the disease in mice lacking theBdkrb1 and combined it with theassessment of osteoclastogenesis inresponse to Bdkrb1 abrogation. Thisled us to demonstrate that the Bdkrb1plays a protective role in the pathogen-esis of experimental periodontitisinhibiting TH17 response, bone lossand osteoclastogenesis.
(a)
(b)
Fig. 1. Ligature placement procedure and its impact on periodontal bone loss.(a) Representative images of histological slides showing the furcation region haemat-oxylin/eosin (H&E stain, original amplification magnitude of 1009.) D, indicating rootdentine; PDL, periodontal ligament and AB, alveolar bone. In ligature-inducedexperimental periodontitis (LIEP) groups, inflammatory tissues, IT is represented.(b) Bone loss area. Values are from one representative experiment with five mice/group. Statistics was performed by the Student’s t-test (**p = 0.005).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
654 Gonçalves-Zillo et al.

Material and Methods
Animals
For the experiments, twelve-week-oldmale kinin B1 receptor homozygousknockout mice (Bdkrb1�/�) were used.These mice were backcrossed for tengenerations with C57Bl6 mice in theCEDEME, Unifesp. Wild-type (Wt)mice of the same age and gender wereused as control. Twelve-week-old maleWistar rats were used for the comple-mentary pharmacological assessment.The animals were kept at a light/darkcycle of 12/12 h, under controlledtemperature (21–23°C) and ad libitumaccess to food and water. Genotypingwas carried out by identifying the pres-ence of the neomycin resistance geneand the absence of the Bdkrb1 gene inknockout mice. The primer sequencesare as follows: Neomycin - 5′-CTTGGG TGG AGA GGC TAT TC-3′,5′-AGG TGA GAT GAC AGG AGAAGA TC-3′; Bdkrb1 - 5′-CCA GCAACC TGT AGC GGT CC-3′, 5′-CGGAAG CCT GGG ATC TGC TG-3′.Animal care and experimental proce-dures were approved by the EthicsCommittee of the Universidade Fed-eral de Sao Paulo (Protocol 1796/06).
LIEP and bone loss measurements
Cotton ligatures (Cotton Black, 4-0/45 cm Strand, non needle; Brasuture,S~ao Sebasti~ao da Grama, SP, Brazil)were placed around one of the firstmolars (Johnson 1975) of the mouse toallow bacteria colonization andincreased plaque accumulation. Con-tralateral molars were left without liga-tures for control. Twenty-one dayslater mice were killed and lymph nodeswere dissected and kept in saline solu-tion for evaluation of cytokine secre-tion. Furthermore, jaws were dissectedand fixed in 10% buffered formalin forhistological analysis. Slides werestained with haematoxylin/eosin (H&Estain) and the furcation region wasused for bone loss histometric mea-surements. For each animal, bone lossarea was obtained subtracting the ori-ginal periodontal ligament area of con-tralateral molar without ligature fromthe total bone loss area on furcationregion of the ligature-treated molar.
Lymph node cells cytokine profile
To assess the role of T lymphocytes inthe disease onset, total submandible
lymph nodes cells from 10 mice/groupwere carefully obtained in Petri dishusing 16 cm2 sterilized voile fabricimmersed in RPMI-mercaptoethanol(5 lM) containing foetal calf serum(10%), penicillin (100 U/ml), strepto-mycin (1%), pH 7.2 and furtherstrained over a 70 lm cell strainer (BDFalconTM, San Jose, CA, USA). Cellswere seeded at a density of 3 9 105
cells/well and incubated for 72 h with1 lg/ml of anti-mouse CD3e antibody(BD PharmingenTM, Mountain View,CA, USA). Levels of IL-17, IL-10,IFN-c and TNF-a in supernatantswere determined by enzyme-linkedimmunosorbent assay (ELISA), usingcommercial anti-cytokine antibodypairs (R&D Systems, Minneapolis,MN, USA) according to the manufac-turer’s protocols. This method alloweda sensitivity range of 15–1000 pg/ml.
ACE inhibitor treatment
To corroborate the data obtainedusing the knockout model, rats andmice were submitted to ligatureprocedure and treated with the ACEienalapril maleate in the drinkingwater. Blood pressure was measuredin conscious rats at 30°C by tailcuffplethysmography using a pressure
transducer (PE-300; Narco Biosys-tems, Austin, TX, USA) connected toa polygraph recorder (Gould Recor-der, Cleveland, OH, USA) as previ-ously described (Santos et al. 2009).Once the dose of enalapril maleatethat does not alter blood pressure inrats was established, mice were treatedwith the optimal dose and histologicalanalysis was performed.
Primary osteoclast culture
Osteoclast precursor enrichment
Osteoclast precursors were obtainedfrom femurs of C57Bl/6 andBdkrb1�/� mice by washing the me-dullar cavity with a-MEM (Sigma,St Louis, MO, USA). The collectedcells were stained with Fluoresceinisothiocyanate (FITC)-conjugatedanti-mouse CD11b (BD Pharmingen)and Phycoerythrin (PE)-conjugatedanti-mouse F4/80 for flow cytometryanalysis. To obtain enriched cultureavoiding B and T cells, which areRANKL sources (Kawai et al.2006), two Petri dishes were coatedwith anti-mouse IgM antibody (BDPharmingen) and two Petri disheswere coated with anti-mouse CD3eantibody (BD Pharmingen) at a finalconcentration of 10 lg/ml. Cells
Fig. 2. Cytokine profile (IL-17, IL-10, IFN-c and TNF-a concentration) was deter-mined by ELISA. Cells from submandibular lymph nodes of wild type (Wt) and kininB1 knockout (Bdkrb1�/�) mice were stimulated with anti-CD3 and 72 h after thesupernatant was obtained to determine cytokine levels. Data represent themean � SEM of 10 mice/group. Statistics were performed by ANOVA and pair-wisecomparisons were done by the Tukey test (*p < 0.05, **p < 0.01 and ***p < 0.001).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
Bone loss in mice lacking B1 receptor 655

were sequentially incubated for15 min in each plate aiming todeplete IgM-expressing and CD3-expressing cells through adsorbedantibodies binding. Cells werediluted in a-MEM and layered overFicoll-Hypaque 1077 (density:1077 g/ml Sigma) for separation ofmononuclear cells and newlyevaluated by flow cytometry. This pro-tocol enriched from 31.78% and36.83% of the CD11b+ cells in Wtand Bdkrb1�/�, respectively, to68.00% (Wt) and 70.63% (Bdkrb1�/
�). No statistical difference wasdetected in enriched culture from Wtor Bdkrb1�/� cells (data not shown).
Osteoclast culture
After enrichment, 104 cells per well werelet to adhere for 1 h in a 96-well plate orin a BD BioCoatTM OsteologicTM16-wellMultitest Slides (BD Bioscience, Bed-ford, MA, USA) for calcium phosphatedemineralization assay. Adherent cellswere then incubated for 48 h in a-MEMpH 7.2 supplemented with foetal calfserum (10%), penicillin (100 U/ml),streptomycin (1%) and 25 ng/ml of M-CSF (Sigma), and then maintained for7 days in the presence of M-CSF(25 ng/ml) and RANKL (50 ng/ml)(Sigma). Themediumwas changed every72 h. On day 9, cells were stained usingthe Acid Phosphatase Leukocyte(TRAP) Kit (Sigma). Cells with three ormore nuclei were considered multinucle-ated (Mn); Mn-TRAP+ cells werecounted and cell area was determined.DNA was stained with Hoechst stain(Invitrogen Life Technologies, Carlsbad,CA, USA) and actin was stained withphalloidin–rhodamine (diluted 1:500;Sigma) for nuclei quantitation. On day14, the ability of cells to demineralize cal-cium phosphate – themain bonemineralcomponent – was assessed in vitro afterperforming the culture onto transparentquartz substrates covered with a thinartificial calcium phosphate film, BDBioCoatTM OsteologicTM 16-well Multit-est Slides (BD Bioscience). Cells wereremoved withMilli-Q water followed by5 min incubation with bleach solution(6% NaOCl, 5.2% NaCl). Three cyclesof vigorous washing and aspiration stepswere performed to complete removal ofthe bleach solution. The slides weremicroscopically examined to assure thatall remaining cells were removed andvon Kossa staining was performed to
allow visualization of the demineraliza-tion lacunae (Pugliese et al. 2012).
Statistical analysis
All data have been presented asmean � standard error. Statistics
were performed by ANOVA andpair-wise comparisons were per-formed by the Tukey tests (a = 5%) orby unpaired two-tailed Student’s t-test(a = 5%). Statistical analyses of bloodpressure data were performed bypaired Student’s t-test (a = 5%).
(a)
(b)
(c)
(d)
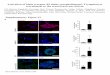
Fig. 3. Osteoclastogenesis. (a) Representative micrographs taken after nine days ofculture and consecutive TRAP staining under the conditions: M-CSF (25 ng/ml) orM-CSF (25 ng/ml) and RANKL (50 ng/ml), original amplification magnitude of1009. (b) Number of tartrate-resistant acid phosphatase-positive multinucleated cells(Mn-TRAP+) on culture plates from control (Wt) and Bdkb1r�/� mice under thesame culture conditions: M-CSF (25 ng/ml) and RANKL (50 ng/ml) and cell area. (c)Representative micrographs taken after 9 days of culture stained with Hoechst andphalloidin under the conditions: M-CSF (25 ng/ml) or M-CSF (25 ng/ml) andRANKL (50 ng/ml), original amplification magnitude of 1009. (d) Number of nucleiper cell. Statistics were performed by the Student’s t-test (*p = 0.03; ***p = 0.0001).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
656 Gonçalves-Zillo et al.

Results
Increased bone loss and IL-17 secretion
during LIEP
Periodontal inflammation wasinduced by placing a ligature aroundthe mandible first molar of mice inorder to verify whether the lack ofBdkrb1 would modify the develop-ment of experimental periodontitis.Histomorphometric quantificationrevealed increased bone loss in thefurcation area of Bdkrb1�/� mice21 days after the ligature (Fig. 1).As evidenced, cotton ligature pro-moted plaque accumulation and con-sequent local inflammation, whichled to a more pronounced alveolarbone loss in Bdkrb1�/�. To confirmactivation of T lymphocytes by liga-ture plaque, cells from submandiblelymph nodes were activated ex vivowith or without anti-mouse CD3antibody and cytokine secretion wasmeasured by ELISA (Fig. 2). With-out anti-mouse CD3 antibody, theamount of cytokines was low, belowthe range of the ELISA test sensitiv-ity. LIEP resulted in increased IL-17secretion by Wt and Bdkrb1�/�
animals. However, this increase wasmore evident in Bdkrb1�/� mice. Incontrast, IL-10 secretion wasreduced in Wt mice 21 days after theligature, while no change in IL-10secretion was observed in Bdkrb1�/�
mice in response to the LIEP. TNF-a was increased in Wt mice as a con-sequence of the LIEP, but no effectof the procedure was observed inBdkrb1�/�. Interestingly, basal levelsof IL-10 and TNF-a were lower inBdkrb1�/� compared to Wt mice.IFN-c remained unaltered in all con-ditions.
Bdkrb1−/− primary osteoclasts are more
numerous
After demonstrating increased boneloss in Bdkrb1�/� mice and the partic-ipation of adaptive immunity as asource of pro-resorptive IL-17 in thesemice, the autonomous effect of thelack of Bdkrb1 in osteoclasts wasinvestigated. Bone marrow cells fromBdkrb1�/� and Wt mice were differ-entiated into osteoclasts using M-CSFand RANKL. At the end of the differ-entiation protocol, the cells fromBdkrb1�/� mice were larger and more
numerous, with a higher number ofnuclei if compared to Wt (Fig. 3).
As the larger cells could havefunctional impairment, we evaluatedthem by calcium phosphate deminer-alization assay. Wt and Bdkrb1�/�
osteoclasts were functionally active;however, Bdkrb1 abrogation resultedin increased formation of lacunae ofdemineralization when compared toWt cells (Fig. 4), which is consistentwith the higher number of osteo-clasts in Bdkrb1�/� (Fig. 3).
Enalapril maleate treatment reduces LIEP
bone loss
To corroborate the results obtainedwith the knockout model, enalaprilmaleate, an inhibitor of ACE, wasadministered in rodents via drinkingwater. Under these conditions, the invivo availability of kinins is increasedand their integrity is preserved by pre-venting degradation by ACE (Molina-ro et al. 2002). However, hypotensionis a confounding consequence of ACEitreatment that denotes the inhibitionof the renin–angiotensin system. Thus,to dissociate the direct effect of ACEinhibition on osteoclastogenesis of itseffect on blood pressure, rats were firstsubmitted to LIEP and treated withhypotensive (60 mg/l) or normotensive(12 mg/l) doses of enalapril. Bloodpressure was assessed at day 0 andafter 21 days. As expected, enalaprilmaleate was able to reduce blood pres-sure in rats receiving the dose of60 mg/l, whereas the dose of 12 mg/ldid not change blood pressure(Fig. 5a). However, treatment withACEi significantly reduced bone loss21 days after the LIEP regardless ofthe dose (Fig. 5b and c). Once theoptimal dose was obtained, Wt micewere treated with 12 mg/l enalaprilmaleate via drinking water. Consistentwith our observations with the knock-out and the rat models, enalapril male-ate also resulted in lower bone loss inmice 21 days of LIEP (Fig. 6a and b),reinforcing the role of kinin in thepathophysiology of periodontitis.
Discussion
In in vitro or ex vivo systems, it hasbeen demonstrated that the activationof Bdkrb1or Bdkrb2 receptors resultsin increased prostaglandin-inducedbiosynthesis of IL-1b and TNF-a andhence increased expression of
(a)
(b)
Fig. 4. Pit formation assay. (a) Representative micrographs taken after 9 days ofculture and consecutive Von-Kossa staining of BD BioCoatTM OsteologicTM 16-wellMultitest Slides under the conditions: M-CSF (25 ng/ml) or M-CSF (25 ng/ml) andRANKL (50 ng/ml). (b) Mean grey value, which is inversely correlated to pit forma-tion, in control (Wt) and Bdkb1r�/� under the same culture conditions. Statistics wereperformed by the Student’s t-test (*p = 0.01).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
Bone loss in mice lacking B1 receptor 657

RANKL (Schenkein 2006, Brechter &Lerner 2007), which suggests a possi-ble role of the kallikrein-kinins systemin the pathogenesis of periodontal dis-ease. Here, we take in vivo and ex vivoapproaches to consider the complexnetwork involving the various playersthat participate in the onset of peri-odontitis, including the innate andadaptive immune responses that affectbone metabolism itself. Based on thein vitro studies, one of the hypotheseswe raised was that the absence of ki-nin receptors or its pharmacologicalblockade would result in a protectiveeffect against periodontitis, whichcould include reduced availability ofinflammatory mediators and pro-oste-
oclastogenic molecules. However, dif-ferently from what we expected, herewe show increased bone loss inducedby ligature in mice lacking Bdkrb1.Consistent with this notion, morerecent reports tend to characterize thekallikrein-kinin system as a rathercomplex system, which may playboth pro- or anti-inflammatory rolesdepending on the disease and the siteof action. The experiments performedby Monteiro et al. (2007) showincreased expression of IL-17 anddecreased expression of IFN-c insplenocytes from Bdkrb2�/� miceafter infection with T. cruzi, suggest-ing a protective role of the Bdkrb2 inthis model. Furthermore, treatment
with Bdkrb1 agonist attenuated theonset and progression of experimentalencephalomyelitis (Schulze-Topphoffet al. 2009). Also, genetic deletion orpharmacological blockade of theBdkrb1 results in earlier onset of thisdisease and more severe clinicalsymptoms in mice. Taken together,these data highlight the importance ofthe Bdkrb1 in the modulation of theimmune system, which goes beyond apro-inflammatory role. It is notewor-thy in the experimental encephalomy-elitis model that the TH17 populationis increased in Bdkrb1�/� mice,whereas no difference is observed inthe proportions of either INF-c-producing cells or FoxP3+ CD25+
T cells (Schulze-Topphoff et al. 2009).Similarly, here we show that the lackof Bdkrb1 leads to increased secretionof IL-17 by cells from submandiblelymph node after 21 days of LIEPwithout changes in INF-c, TNF-aand IL-10. It is important to note thatexperimental periodontitis in micedoes not interfere in IFN-c secretionby lymph node cells, despite all sug-gestive data of the involvement of thiscytokine in bone metabolism. On theother hand, IL-10 is shown here to bedown-regulated whereas TNF-a wasup-regulated by LIEP in Wt mice,which is in accordance to anotherstudy (Kinane et al. 2011). The factthat bone loss was more pronouncedin Bdkrb1�/� is related neither toTNF-a increase nor to IL-10 decreaseonce these cytokine physiological lev-els were already low in the knockoutanimals and were not modified afterLIEP. Therefore, it seems clear thatIL-17 is involved in the pathogenesisof this experimental disease.
This study is a pioneer in investi-gating T cell response to LIEP andin relating IL-17 to bone loss in thismodel. It is known, however, thatthe RANK/RANKL/OPG axis ismodulated by IL-17 in rheumatoidarthritis (Daoussis et al. 2010). IL-17is capable of stimulating the produc-tion of RANKL by osteoblasts andT lymphocytes. TH17 cells are alsosources of RANKL themselves (Satoet al. 2006). Thus, we suggest thatBdkrb1 could be an important mod-ulator of T cells, participating in thepathogenesis of periodontitis bystimulating the production ofRANKL via IL-17.
Our work is also the first toreport the direct involvement of the
(a)
(b)
(c)
Fig. 5. (a) Mean systolic blood pressure of adult rats in baseline and after 21 daystreatment or not with enalapril maleate in drinking water. Data represent themean � SEM of seven rats. Statistics was done by paired t-test (*p = 0.05). (b)Representative images of histological slides of furcation region of all groups used formeasurements haematoxylin/eosin (H&E stain, original amplification magnitude of209). Histology of furcation region of rats, indicating root dentine, D; alveolar bone,AB and inflammatory tissue, IT. (c) Bone loss area. Data represent the mean � SEMof seven rats. Statistics were performed by ANOVA and pair-wise comparisons weredone by the Tukey test (**p = 0.04).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
658 Gonçalves-Zillo et al.

kallikrein-kinin system with osteocl-astogenesis. However, the mecha-nism whereby deletion of the kininB1 receptor increases osteoclastogen-esis is not fully understood. Lowernitric oxide inhibitory signal toosteoclastogenesis may be involved.Nitric oxide (NO) is produced bythe oxidation of one of the guanidi-no nitrogens of L-arginine by a fam-ily of NO synthase (NOS) isoforms.NO is described as an autocrine neg-ative feedback signal to limit osteo-clastogenesis concurrently stimulatedby RANKL (van’t Hof & Ralston1997, Zheng et al. 2006). In addi-tion, NO is a known suppressor ofthe proliferation and function ofpolarized murine and human TH17cells (Niedbala et al. 2011). In a pre-vious work, targeted deletion ofBdkrb1 gene in mice mesentericarterioles resulted in decreased NObioavailability, accompanied bysurprising enhanced NOS activity(Loiola et al. 2011). In addition tothe cell autonomous effects ofBdkrb1 absence in osteoclast precur-sors, it is likely that the increased os-teoclastogenesis in mice lackingBdkrb1 is associated with increasedexpression of IL-17 by T lympho-cytes, which also contributes toincreased bone loss under experimen-tal periodontitis. Further, our experi-ments do not demonstrate changes inIFN-c, a potential NO inducer, atleast by lymph node cells.
The pharmacological confirmationof kinin receptors role in experimentalperiodontitis was performed indirectlyin our study. Injection of Bdkrb1 ago-nist requires daily peritoneal puncture,which results in peritonitis and activa-tion of T lymphocytes (data notshown), confusing the analysis of thepathophysiology of periodontitis.Hence, we tested the effect of ACEi inthe mice in order to increase the effectof kinins in our system. Enalaprilmaleate, an ACEi, was administeredvia drinking water, eliminating theneed of puncture and consequentlyinflammation. As the ACE is commonto kallikrein-kinin system and renin–angiotensin system, the optimal dosethat does not cause hypotension wasused. In rats, 60 mg/l enalapril male-ate prevents bone loss and promoteshypotension, while administration of12 mg/l enalapril maleate preventsbone loss with no effect on bloodpressure. These results confirm theimportance of kinins for properinflammatory modulation and peri-odontitis in response to LIEP. This isthe first evidence of the role of the kal-likrein-kinin system in periodontalhomeostasis.
In conclusion, we found thatBdkrb1 may contribute to modulationof inflammatory response and ex vivocytokine production by lymph nodecells, which affect bone loss in amodel of experimental periodontitisand also modulates in vitro osteoclas-
togenesis. In addition, we could showthat a frequently used anti-hyperten-sive drug – namely an ACE inhibitor– could also be used to decreaseperiodontitis bone loss in this model,suggesting the role of kinins inthis disease. Therefore, the resultsdescribed here demonstrate an impor-tant role for the Bdkrb1 in the patho-genesis of periodontitis. Moreover,based on our data, we can suggest theuse of ACE inhibition as an adjuvantto periodontal treatment.
Acknowledgements
The authors thank Elton Dias daSilva for his technical assistance.
References
Ara�ujo, R. C., Kettritz, R., Fichtner, I., Paiva, A.C., Pesquero, J. B. & Bader, M. (2001) Alteredneutrophil homeostasis in kinin B1 receptor-deficient mice. Biological Chemistry 382, 91–95.
Boyle, W. J., Simonet, W. S. & Lacey, D. L.(2003) Osteoclast differentiation and activation.Nature 423, 337–342.
Brechter, A. B. & Lerner, U. H. (2007) Bradyki-nin potentiates cytokine-induced prostaglandinbiosynthesis in osteoblasts by enhanced expres-sion of cyclooxygenase 2, resulting in increasedRANKL expression. Arthritis & Rheumatism56, 910–923.
Brechter, A. B., Persson, E., Lundgren, I. & Lerner,U. H. (2008) Kinin B1 and B2 receptor expres-sion in osteoblasts and fibroblasts is enhancedby interleukin-1 and tumour necrosis factor-alpha. Effects dependent on activation of NF-kappaB and MAP kinases. Bone 43, 72–83.
Brunius, G., Domeij, H., Gustavsson, A. & Yucel-Lindberg, T. (2005) Bradykinin upregulates IL-8production in human gingival fibroblasts stimu-lated by interleukin-1beta and tumor necrosisfactor alpha. Regulatory Peptides 126, 183–188.
Daoussis, D., Andonopoulos, A. P. & Liossis, S. N.(2010) Wnt pathway and IL-17: novel regulatorsof joint remodeling in rheumatic diseases. Look-ing beyond the RANK-RANKL-OPG axis. Sem-inars in Arthritis and Rheumatism 39, 369–383.
Felix, R., Cecchini, M. G. & Fleisch, H. (1990)Macrophage colony stimulating factor restoresin vivo bone resorption in the op/op osteopet-rotic mouse. Endocrinology 127, 2592–2594.
Graves, D. T. & Cochran, D. (2003) The contri-bution of interleukin-1 and tumor necrosis fac-tor to periodontal tissue destruction. Journal ofPeriodontology 74, 391–401.
Gustafson, G. T. & Lerner, U. (1984) Bradykininstimulates bone resorption and lysosomal-enzyme release in cultured mouse calvaria. Bio-chemical Journal 219, 329–332.
Hodge, J. M., Kirkland, M. A. & Nicholson, G.C. (2007) Multiple roles of M-CSF in humanosteoclastogenesis. Journal of Cellular Biochem-istry 102, 759–768.
van’t Hof, R. J. & Ralston, S. H. (1997) Cytokine-induced nitric oxide inhibits bone resorption byinducing apoptosis of osteoclast progenitors andsuppressing osteoclast activity. Journal of Boneand Mineral Research 12, 1797–1804.
(a)
(b)
Fig. 6. (a) Representative images of histological slides of furcation region from micetreated or not with 12 mg/l enalapril maleate haematoxylin/eosin (H&E stain, originalamplification magnitude of 1009.) indicating root dentine, D; alveolar bone, AB andinflammatory tissue, IT. (b) Bone loss area. Data represent the mean � SEM of eightmice. Statistics were performed by the Student’s t-test (***p = 0.001).
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
Bone loss in mice lacking B1 receptor 659

Johnson, I. H. (1975) Effects of local irritationand dextran sulphate administration on theperiodontium of the rat. Journal of PeriodontalResearch 10, 332–345.
Kawai, T., Matsuyama, T., Hosokawa, Y., Makih-ira, S., Seki, M., Karimbux, N. Y., Goncalves, R.B., Valverde, P., Dibart, S., Li, Y. P., Miranda, L.A., Ernst, C. W., Izumi, Y. & Taubman, M. A.(2006) B and T lymphocytes are the primarysources of RANKL in the bone resorptive lesionof periodontal disease. American Journal ofPathology 169, 987–998.
Kinane, D. F., Preshaw, P. M. & Loos, B. G.(2011) Working Group 2 of Seventh EuropeanWorkshop on Periodontology. Host-response:understanding the cellular and molecular mech-anisms of host-microbial interactions–consensusof the Seventh European Workshop on Peri-odontology. Journal of Clinical Periodontology38, 44–48.
Kostenuik, P. J. & Shalhoub, V. (2001) Osteopro-tegerin: a physiological and pharmacologicalinhibitor of bone resorption. Current Pharma-ceutical Design 7, 613–635.
Lacey, D. L., Timms, E., Tan, H. L., Kelley, M.J., Dunstan, C. R., Burgess, T., Elliott, R.,Colombero, A., Elliott, G., Scully, S., Hsu, H.,Sullivan, J., Hawkins, N., Davy, E., Capparelli,C., Eli, A., Qian, Y. X., Kaufman, S., Sarosi,I., Shalhoub, V., Senaldi, G., Guo, J., Delaney,J. & Boyle, W. J. (1998) Osteoprotegerin ligandis a cytokine that regulates osteoclast differenti-ation and activation. Cell 93, 165–176.
Leeb-Lundberg, L. M. F., Marceau, F., Uller-Esterl, W. M., Pettibone, D. J. & Zuraw, B. L.(2005) Classification of the kinin receptor fam-ily: from molecular mechanisms to pathophysi-ological consequences. Pharmacological Reviews57, 27–77.
Lerner, U. H. (1994) Regulation of bone metabo-lism by the kallikrein-kinin system, the coagu-lation cascade, and the acute-phase reactants.Oral Surgery, Oral Medicine and Oral Pathol-ogy 78, 481–493.
Ljunggren, €O. & Lerner, U. H. (1990) Evidencefor B1 bradykinin-receptor mediated prosta-glandin formation in osteoblasts and subse-quent enhancement of bone resorption. BritishJournal of Pharmacology 101, 382–386.
Loiola, R. A., Reis, F. C., Kawamoto, E. M.,Scavone, C., Abdalla, D. S., Fernandes, L. &Pesquero, J. B. (2011) Role of vascular KininB1 and B2 receptors in endothelial nitric oxidemetabolism. Peptides 32, 1700–1705.
Molinaro, G., Cugno, M., Perez, M., Lepage, Y.,Gervais, N., Agostoni, A. & Adam, A. (2002)Angiotensin-converting enzyme inhibitor-asso-
ciated angioedema is characterized by a slowerdegradation of des-arginine(9)-bradykinin.Journal of Pharmacology and ExperimentalTherapeutics 303, 232–237.
Monteiro, A. C., Schmitz, V., Morrot, A., deArruda, L. B., Nagajyothi, F., Granato, A.,Pesquero, J. B., M€uller-Esterl, W., Tanowitz,H. B. & Scharfstein, J. (2007) Bradykinin B2Receptors of dendritic cells, acting as sensorsof kinins proteolytically released by Trypanoso-ma cruzi, are critical for the development ofprotective type-1 responses. PLoS Pathogens 3,1730–1744.
Monteiro, A. C., Schmitz, V., Svensjo, E., Gazzi-nelli, R. T., Almeida, I. C., Todorov, A., deArruda, L. B., Torrecilhas, A. C., Pesquero, J.B., Morrot, A., Bouskela, E., Bonomo, A.,Lima, A. P., M€uller-Esterl, W. & Scharfstein,J. (2006) Cooperative activation of TLR2 andbradykinin B2 receptor is required for induc-tion of type 1 immunity in a mouse model ofsubcutaneous infection by Trypanosoma cruzi.The Journal of Immunology 177, 6325–6335.
Niedbala, W., Alves-Filho, J. C., Fukada, S. Y.,Vieira, S. M., Mitani, A., Sonego, F., Mirchan-dani, A., Nascimento, D. C., Cunha, F. Q. &Liew, F. Y. (2011) Regulation of type 17 helperT-cell function by nitric oxide during inflamma-tion. Proceedings of National Academy ofSciences of the United States of America 108,9220–9225.
Pesquero, J. B., Pesquero, J. L., Oliveira, S. M.,Roscher, A. A., Metzger, R., Ganten, D. &Bader, M. (1996) Molecular cloning and func-tional characterization of a mouse bradykininB1 receptor gene. Biochemical and BiophysicalResearch Communications 220, 219–225.
Pixley, F. J. & Stanley, E. R. (2004) CSF-1 regula-tion of the wandering macrophage: complexityin action. Trends in Cell Biology 14, 628–638.
Prat, A., Weinrib, L., Becher, B., Poirier, J.,Duquette, P., Couture, R. & Antel, J. P. (1999)Bradykinin B1 receptor expression and func-tion on T lymphocytes in active multiple sclero-sis. Neurology 53, 2087–2092.
Pugliese, L. S., Goncalves, T. O., Popi, A. F.,Mariano, M., Pesquero, J. B. & Lopes, J. D.(2012) B-1 lymphocytes differentiate into func-tional osteoclast-like cells. Immunobiology 217,336–344.
Regoli, D. & Barabe, J. (1980) Pharmacology ofbradykinin and related kinins. PharmacologicalReviews 32, 1–46.
Santos, E. L., de Picoli Souza, K., da Silva, E.D., Batista, E. C., Martins, P. J., D’Almeida,V. & Pesquero, J. B. (2009) Long term treat-ment with ACE inhibitor enalapril decreases
body weight gain and increases life span inrats. Biochemical Pharmacology 78, 951–958.
Sato, K., Suematsu, A., Okamoto, K., Yamaguchi,A., Morishita, Y., Kadono, Y., Tanaka, S.,Kodama, T., Akira, S., Iwakura, Y., Cua, D. J.& Takayanagi, H. (2006) Th17 functions as anosteoclastogenic helper T cell subset that links Tcell activation and bone destruction. The Journalof Experimental Medicine 203, 2673–2682.
Schenkein, H. A. (2006) Host response in maintain-ing periodontal health and determining peri-odontal disease. Periodontology 2000 40, 77–93.
Schulze-Topphoff, U., Prat, A., Prozorovski, T.,Siffrin, V., Paterka, M., Herz, J., Bendix, I., If-ergan, I., Schadock, I., Mori, M. A., VanHorssen, J., Schr€oter, F., Smorodchenko, A.,Han, M. H., Bader, M., Steinman, L., Aktas,O. & Zipp, F. (2009) Activation of kinin recep-tor B1limits encephalitogenic T lymphocyterecruitment to the central nervous system.Nature Medicine 15, 788–793.
Simonet, W. S., Lacey, D. L., Dunstan, C. R.,Kelley, M., Chang, M. S., L€uthy, R., Nguyen,H. Q., Wooden, S., Bennett, L., Boone, T.,Shimamoto, G., DeRose, M., Elliott, R., Co-lombero, A., Tan, H. L., Trail, G., Sullivan, J.,Davy, E., Bucay, N., Renshaw-Gegg, L.,Hughes, T. M., Hill, D., Pattison, W., Campbell,P., Sander, S., Van, G., Tarpley, J., Derby, P.,Lee, R. & Boyle, W. J. (1997) Osteoprotegerin: anovel secreted protein involved in the regulationof bone density. Cell 89, 309–319.
Teng, Y. A. (2006) Protective and destructiveimmunity in periodontium: part 2 – T-cell-med-iated Immunity in the periodontium. Journal ofDental Research 85, 209–219.
Zheng, H., Yu, X., Collin-Osdoby, P. & Osdoby,P. (2006) RANKL stimulates inducible nitric-oxide synthase expression and nitric oxide pro-duction in developing osteoclasts. An autocrinenegative feedback mechanism triggered byRANKL-induced interferon-beta via NF-kappaB that restrains osteoclastogenesis andbone resorption. Journal of Biological Chemis-try 281, 15809–15820.
Address:Joao Bosco PesqueroUniversidade Federal de Sao PauloRua Pedro de Toledo, 669, 9º andar, VilaClementino, Sao Paulo-SP 04039-032BrazilE-mail: [email protected]
Clinical Relevance
Scientific rationale for the study:The knowledge of the inflamma-tory mediators and their effects inbone loss, including kinins andtheir receptors led us to hypothe-size that kinin B1 receptor activa-tion results in modulation of
osteolytic processes involved in peri-odontal attachment loss.Principal findings: In this study wedemonstrated that the absence of B1receptor resulted in increased boneloss. On the other hand, the block-ade of kinins degradation by angio-tensin-converting enzyme inhibitionprevented bone loss in a periodonti-
tis rodent model, which alsoallowed to demonstrate the invol-vement of T lymphocytes in thisprocess.Practical implications: We believethat the use of angiotensin-convert-ing enzyme inhibitors can beemployed as an adjuvant to con-ventional periodontal therapy.
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
660 Gonçalves-Zillo et al.