Embed Size (px)
Citation preview

of May 29, 2018.This information is current as
Osteoarthritis in Humans and MiceElevated MMP-13 Expression in Increase in ALK1/ALK5 Ratio as a Cause for
KraanGoumans, Wim B. van den Berg and Peter M. van der Marie-JoseVitters, Henk M. van Beuningen, Arjen B. Blom,
Esmeralda N. Blaney Davidson, Dennis F. G. Remst, Elly L.
http://www.jimmunol.org/content/182/12/7937doi: 10.4049/jimmunol.0803991
2009; 182:7937-7945; ;J Immunol
Referenceshttp://www.jimmunol.org/content/182/12/7937.full#ref-list-1
, 11 of which you can access for free at: cites 39 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Errata
/content/185/4/2629.full.pdfor:
next pageAn erratum has been published regarding this article. Please see
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Increase in ALK1/ALK5 Ratio as a Cause for ElevatedMMP-13 Expression in Osteoarthritis in Humans and Mice1
Esmeralda N. Blaney Davidson,2* Dennis F. G. Remst,* Elly L. Vitters,* Henk M. van Beuningen,*Arjen B. Blom,* Marie-Jose Goumans,† Wim B. van den Berg,* and Peter M. van der Kraan*
During osteoarthritis (OA) chondrocytes show deviant behavior resembling terminal differentiation of growth-plate chondrocytes,characterized by elevated MMP-13 expression. The latter is also a hallmark for OA. TGF-� is generally thought to be a protectivefactor for cartilage, but it has also displayed deleterious effects in some studies. Recently, it was shown that besides signaling viathe ALK5 (activin-like kinase 5) receptor, TGF-� can also signal via ALK1, thereby activating Smad1/5/8 instead of Smad2/3. TheSmad1/5/8 route can induce chondrocyte terminal differentiation. Murine chondrocytes stimulated with TGF-� activated theALK5 receptor/Smad2/3 route as well as the ALK1/Smad1/5/8 route. In cartilage of mouse models for aging and OA, ALK5expression decreased much more than ALK1. Thus, the ALK1/ALK5 ratio increased, which was associated with changes in therespective downstream markers: an increased Id-1 (inhibitor of DNA binding-1)/PAI-1 (plasminogen activator inhibitor-1)ratio. Transfection of chondrocytes with adenovirus overexpressing constitutive active ALK1 increased MMP-13 expression,while small interfering RNA against ALK1 decreased MMP-13 expression to nondetectable levels. Adenovirus overexpress-ing constitutive active ALK5 transfection increased aggrecan expression, whereas small interfering RNA against ALK5resulted in increased MMP-13 expression. Moreover, in human OA cartilage ALK1 was highly correlated with MMP-13expression, whereas ALK5 correlated with aggrecan and collagen type II expression, important for healthy cartilage. Col-lectively, we show an age-related shift in ALK1/ALK5 ratio in murine cartilage and a strong correlation between ALK1 andMMP-13 expression in human cartilage. A change in balance between ALK5 and ALK1 receptors in chondrocytes causedchanges in MMP-13 expression, thereby causing an OA-like phenotype. Our data suggest that dominant ALK1 signalingresults in deviant chondrocyte behavior, thereby contributing to age-related cartilage destruction and OA. The Journal ofImmunology, 2009, 182: 7937–7945.
O steoarthritis (OA)3 is the most common joint disease.Patients with OA suffer from cartilage degeneration re-sulting in loss of joint function. The major risk factor for
OA is age. Cartilage cells, chondrocytes, maintain the homeostasisof normal articular cartilage. However, in OA, chondrocytes havea distorted phenotype and produce MMP-13. This enzyme is themost potent type II collagen-degrading protein. Thus, in OA, thechondrocytes themselves induce destruction of articular cartilage.The articular chondrocytes in OA cartilage behave analogous tohypertrophic growth plate chondrocytes that have undergone ter-minal differentiation and start to produce MMP-13 in high amounts(1, 2). Until now it is not clear why chondrocytes in OA articularcartilage behave this way.
Chondrocyte terminal differentiation is repressed by the Smad2/3pathway (3, 4). This pathway is activated upon binding of TGF-�to its type II receptor, which then recruits the type I receptor ALK5(activin-like kinase 5). This complex in turn phosphorylates theintracellular effectors Smad2 and Smad3 (5–9). TGF-� signalinghas been found to be important for maintenance and protection ofhealthy cartilage (4).
However, besides being a potent protective factor for cartilage,TGF-� also has the ability to induce OA under certain conditions(10–14). Until now this has been a puzzling, unexplained TGF-�property.
It has recently been shown that TGF-� also has the ability tosignal via the alternative receptor ALK1 instead of the well-char-acterized ALK5 route. Activation of ALK1 has been found to re-sult in activation of the Smad1/5/8 pathway (15), a pathway that isknown to induce chondrocyte terminal differentiation (16). Thebalance between signaling via either ALK5 or ALK1 can deter-mine the response of cells to TGF-� stimulation, which can betotally opposite (17, 18). For instance, in endothelial cells, ALK5inhibits migration, whereas ALK1 stimulates migration and pro-liferation (19).
Our group has previously demonstrated that with age ALK5expression and Smad2 phosphorylation strongly decrease in mu-rine cartilage (20). Additionally, signaling via ALK5 drasticallydecreased with development of OA (21).
We hypothesize that with this decrease in ALK5 signaling,a shift toward a dominant TGF-� signaling via ALK1 occurs,thereby inducing a chondrocyte phenotype resembling hypertro-phic chondrocytes expressing elevated MMP-13 levels. Thiswould explain the contradictory results that have been found until
*Rheumatology Research and Advanced Therapeutics, Radboud University NijmegenMedical Center, Nijmegen, the Netherlands; and †Department of Molecular Biology,University Medical Center Leiden, Leiden, the Netherlands
Received for publication December 1, 2008. Accepted for publication April 14, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This research was supported by the Dutch Arthritis Association “NationalReumafonds”.2 Address correspondence and reprint requests to Dr. Esmeralda N. Blaney Davidson,Rheumatology Research and Advanced Therapeutics, Radboud University NijmegenMedical Center, Geert Grooteplein 28, 6525 GA Nijmegen, the Netherlands. E-mailaddress: [email protected] Abbreviations used in this paper: OA, osteoarthritis; Ad-caALK, adenovirusoverexpressing constitutive active ALK; ALK, activin-like kinase; DMM, desta-bilization of the medial meniscus; Id-1, inhibitor of DNA binding-1; PAI-1, plas-minogen activator inhibitor-1; shRNA, short hairpin RNA.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0803991
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

now. Therefore, we have studied ALK1 and ALK5 expression inarticular cartilage in aging mice, experimental models of OA, andhuman OA. Moreover, we have examined the functional conse-quences of dominant ALK1 or ALK5 signaling in chondrocytes,revealing that a shift in balance toward ALK1 induces an OAphenotypic change, thereby providing a new view on TGF-� sig-naling during aging and OA.
Materials and MethodsCell culture
Chondrocytes were obtained from mouse cartilage by digestion with collage-nase. They were immortalized by transduction with SV40 large T Ag andsubsequently cloned by minimal dilution as previously described (22). Allchondrocyte cell lines had both ALK5 and ALK1 expression. The chondrocytecell line (H1) was cultured in 105 cells per well in 24-well plates in DMEM/HAM’s F12 (1:1) with 10% FCS and penicillin and streptomycin.
The H1 chondrocytes were stimulated with TGF-� (1 ng/ml) under se-rum-free conditions to evaluate functional TGF-� signaling via ALK5 orALK1. This experiment was repeated in bovine primary chondrocytes toensure that the chondrocyte cell line responded similarly to primary cells.The primary bovine chondrocytes were isolated from bovine metacarpo-phalangeal joint as previously described (23).
Additionally, H1 chondrocytes were transduced with either adenovirusoverexpressing constitutive active ALK1 (Ad-caALK1) or Ad-caALK5 orAd-LacZ control (multiplicity of infection of 50) for 2 h and cultured for48 h thereafter to evaluate whether active ALK1 resulted in Smad1/5/8Pand active ALK5 resulted in Smad2P or to investigate expression patternson the mRNA level.
For blocking experiments, short hairpin RNA (shRNA) GFP-taggedplasmids containing shRNA for ALK1 or ALK5 or a negative control(SABiosciences) were used. H1 chondrocytes were plated in 75-cm2 flasks(106 cells/flask), and 5.8 �g of shRNA was transfected per flask usingLipofectamine 2000 reagent (Invitrogen). After 2 days the GFP-positivecells were separated from the nonpositive cells using FACS sorting (EpicsElite; Beckman Coulter).
Quantitative PCR
Quantitative PCR was performed as previously described on an ABI Prism7000 sequence detection system (Applied Biosystems) (24). Efficiencies (E)for all primer sets were determined (Table I) using a standard curve of fiveserial cDNA dilutions in water in duplicate. Primers were accepted if the deviationfrom the slope of the standard curve was �0.3 compared with the slope of theGAPDH standard curve and if the melting curve showed only one product.
Western blot
For Western blot analyses each sample was measured for protein concen-tration, after which 20 �g of each protein sample was loaded on a SDS7.5% polyacrylamide gel. Proteins were transferred onto a nitrocellulosemembrane (Hybond P; Amersham Pharmacia Biotech) using the iBlot sys-tem (Invitrogen). The membrane was incubated with Abs against Smad1/5/8P (1/1000) and Smad2P (1/1000) (Cell Signaling Technology) over-night at 4°C. The secondary HRP-conjugated Ab was goat-anti-rabbit (1/
1500) (Dako), and the membrane was developed using the ECL PlusWestern blotting detection system (GE Healthcare).
Animals
Male C57BL/6N mice aged 1 year (n � 7) and 2 years (n � 10) were usedfor comparison of young vs old mice.
C57BL/6N mice aged 10 wk were used to induce OA by transaction ofthe medial meniscus (n � 10) (destabilization of the medial meniscus(DMM) model). In this surgical mouse model the anterior attachment of themedial meniscus to the tibia is transected, as previously described by Glas-son et al. (25). Mice were sacrificed 8 wk after surgery.
STR/ort mice aged 3 mo (n � 10), 6 mo (n � 20), 9 mo (n � 6), and1 year (n � 10) were used as a spontaneous model of OA. The histologicallesions seen in this model resemble those seen in humans (26, 27).
Knee joints of sacrificed mice were isolated for histology. OA was con-firmed by histological evaluation. Animals were considered to have hyalinecartilage degeneration, the signature pathologic feature of OA, if they hadcartilage matrix degradation and/or regeneration/repair, chondrocyte death,chondrocyte replication, chondrocyte proliferation, cartilage lesions, bonelesion soft tissue laxity, or a combination of lesions (28).
Animals were kept in filter top cages with woodchip bedding understandard pathogen-free conditions. They were fed a standard diet with tapwater ad libitum. The Animal Ethics Committee of the Radboud UniversityNijmegen approved all animal procedures.
Histology
Knee joints were fixed in phosphate-buffered formalin for 7 days. Theywere dehydrated using an automated tissue-processing apparatus (Tissue-Tek VIP; Sakura) and embedded in paraffin. Tissue sections of 7 �m wereprepared.
Immunohistochemistry was performed as previously described (21).Specific primary Abs against ALK1 (1/100), ALK5 (1/100), Id-1 (inhibitorof DNA binding-1) (1/1000), and PAI-1 (plasminogen activator inhibi-tor-1) (1/100) were incubated overnight at 4°C (purchased from Santa CruzBiotechnology). As a negative control, the primary Ab was replaced withgoat or rabbit IgGs. A biotin-streptavidin detection system was used ac-cording to the manufacturer’s protocol (Vector Laboratories). Bound com-plexes were visualized via reaction with 3�,3�-diaminobenzidine (Sigma-Aldrich) and H2O2 resulting in a brown precipitate. Sections werecounterstained with heamatoxylin and mounted with Permount.
Image analysis
For the different Ags, the number of positive articular chondrocytes inthe tibia was determined by a blinded observer using the Qwin imageanalysis system (Leica Imaging Systems) and a Leica DC 300F digitalcamera as previously described (20). Briefly, a line with a defined widththat corresponds to the width of the noncalcified cartilage was drawnacross the cartilage in the image. The computer software defined thepositive particles according to preset values of immunopositivity. Foreach knee joint, at least three tissue sections were measured and thereafteraveraged per joint. The number of positive cells was corrected for theaverage number of chondrocytes in healthy joints determined in hematox-ylin-stained sections.
Table I. Primers used for quantitative PCR analysis
Gene R2 Ea Forward Primer (5�33�) Reverse Primer (5�33�)
MouseGAPDH 0.997 2.05 GGCAAATTCAACGGCACA GTTAGTGGGGTCTCGCTCCTGALK1 0.999 1.90 ACCATCGTGAATGGCATCGT GGTCATTGGGCACCACATCALK5 0.999 2.02 AGCGGTCTTGCCCATCTTC AGCAATGGCTGGCTTTCCTCollagen II 0.992 2.15 TTCCACTTCAGCTATGGCGA GACGTTAGCGGTGTTGGGAGAggrecan 0.992 2.15 TCTACCCCAACCAAACCGG AGGCATGGTGCTTTGACAGTGMMP-13 0.992 1.93 ACCTTGTGTTTGCAGAGCACTAACTT CTTCAGGATTCCCGCAAGAGT
HumanGAPDH 0.999 1.92 ATCTTCTTTTGCGTCGCCAG TTCCCCATGGTGTCTGAGCALK1 0.997 2.05 GACTCAAGAGCCGCAATGTG GGTCGGCGATGCAACACALK5 0.993 2.05 CGACGGCGTTACAGTGTTTCT CCCATCTGTCACACAAGTAAAATTGCollagen II 0.995 2.00 CACGTACACTGCCCTGAAGGA CGATAACAGTCTTGCCCCACTTAggrecan 0.997 2.01 GCCTGCGCTCCAATGACT ATGGAACACGATGCCTTTCACMMP-13 0.998 1.95 ATTAAGGAGCATGGCGACTTCT CCCAGGAGGAAAAGCATGAG
a Efficiency.
7938 ALTERATIONS IN TGF-� SIGNALING DURING OSTEOARTHRITIS
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Human cartilage samples
Samples of human cartilage were obtained from patients undergoing kneeor hip replacement surgery. The cartilage was instantly frozen in liquid
nitrogen and crushed using a dismembrator. The crushed material was dis-solved in RLT buffer supplied with the RNeasy Mini kit (Qiagen). Sampleswere treated with proteinase K, and subsequently RNA was isolated furtherwith the RNeasy Mini kit according to the manufacturer’s protocol, afterwhich an RT-PCR was performed followed by a quantitative PCR.
The Medical Ethics Committee of Radboud University Nijmegen Med-ical Centre approved the study protocol.
Statistical analysis
Results were analyzed with Student’s t test and considered significant if thep value was �0.05. Correlations were tested with Pearson’s correlation andconsidered significant if the p value was �0.05.
ResultsChondrocytes signal both via ALK5 and ALK1
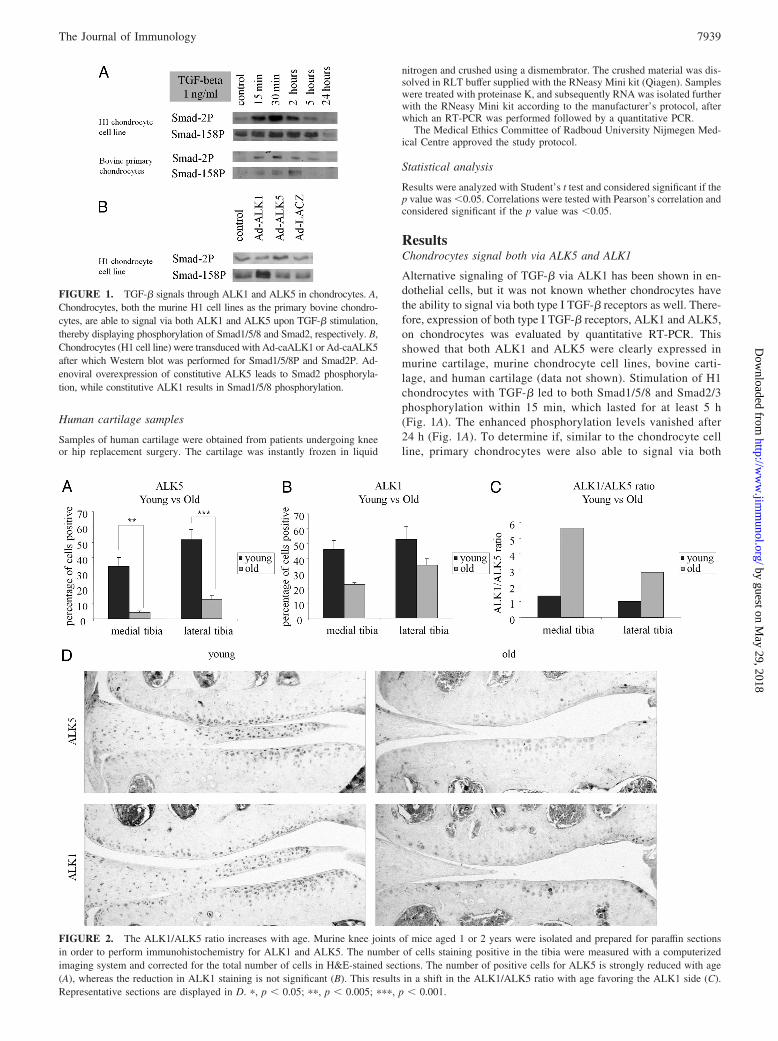
Alternative signaling of TGF-� via ALK1 has been shown in en-dothelial cells, but it was not known whether chondrocytes havethe ability to signal via both type I TGF-� receptors as well. There-fore, expression of both type I TGF-� receptors, ALK1 and ALK5,on chondrocytes was evaluated by quantitative RT-PCR. Thisshowed that both ALK1 and ALK5 were clearly expressed inmurine cartilage, murine chondrocyte cell lines, bovine carti-lage, and human cartilage (data not shown). Stimulation of H1chondrocytes with TGF-� led to both Smad1/5/8 and Smad2/3phosphorylation within 15 min, which lasted for at least 5 h(Fig. 1A). The enhanced phosphorylation levels vanished after24 h (Fig. 1A). To determine if, similar to the chondrocyte cellline, primary chondrocytes were also able to signal via both
FIGURE 1. TGF-� signals through ALK1 and ALK5 in chondrocytes. A,Chondrocytes, both the murine H1 cell lines as the primary bovine chondro-cytes, are able to signal via both ALK1 and ALK5 upon TGF-� stimulation,thereby displaying phosphorylation of Smad1/5/8 and Smad2, respectively. B,Chondrocytes (H1 cell line) were transduced with Ad-caALK1 or Ad-caALK5after which Western blot was performed for Smad1/5/8P and Smad2P. Ad-enoviral overexpression of constitutive ALK5 leads to Smad2 phosphoryla-tion, while constitutive ALK1 results in Smad1/5/8 phosphorylation.
FIGURE 2. The ALK1/ALK5 ratio increases with age. Murine knee joints of mice aged 1 or 2 years were isolated and prepared for paraffin sectionsin order to perform immunohistochemistry for ALK1 and ALK5. The number of cells staining positive in the tibia were measured with a computerizedimaging system and corrected for the total number of cells in H&E-stained sections. The number of positive cells for ALK5 is strongly reduced with age(A), whereas the reduction in ALK1 staining is not significant (B). This results in a shift in the ALK1/ALK5 ratio with age favoring the ALK1 side (C).Representative sections are displayed in D. �, p � 0.05; ��, p � 0.005; ���, p � 0.001.
7939The Journal of Immunology
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
routes in response to TGF-�, the experiment was repeated withprimary bovine chondrocytes. These chondrocytes also responded toTGF-� by phosphorylation of Smad1/5/8 and Smad2. The bovineprimary chondrocytes had a slower response on Smad1/5/8 phosphor-ylation (Fig. 1A). These data show that TGF-� is not only able tosignal via the Smad2/3 pathway, but also via the Smad1/5/8 route inchondrocytes.
To assess whether activation of the ALK1 pathway leads tothe expected Smad1/5/8 phosphorylation and ALK5 to Smad2/3phosphorylation, we transduced the H1 chondrocytes with Ad-caALK1, Ad-caALK5, or Ad-LacZ as a control. The ��CT val-ues of transfected samples vs nontransfected controls was 10.99for ALK1 and 11.3 for ALK5, and thus transfection led to acomparable increase in mRNA expression levels. It was clearthat transfection with Ad-ALK1 specifically led to phosphory-lation of Smad1/5/8P, whereas ALK5 resulted in Smad2 phos-phorylation (Fig. 1B).
Reduction in ALK5 expression is much stronger than reductionin ALK1 in aging mice
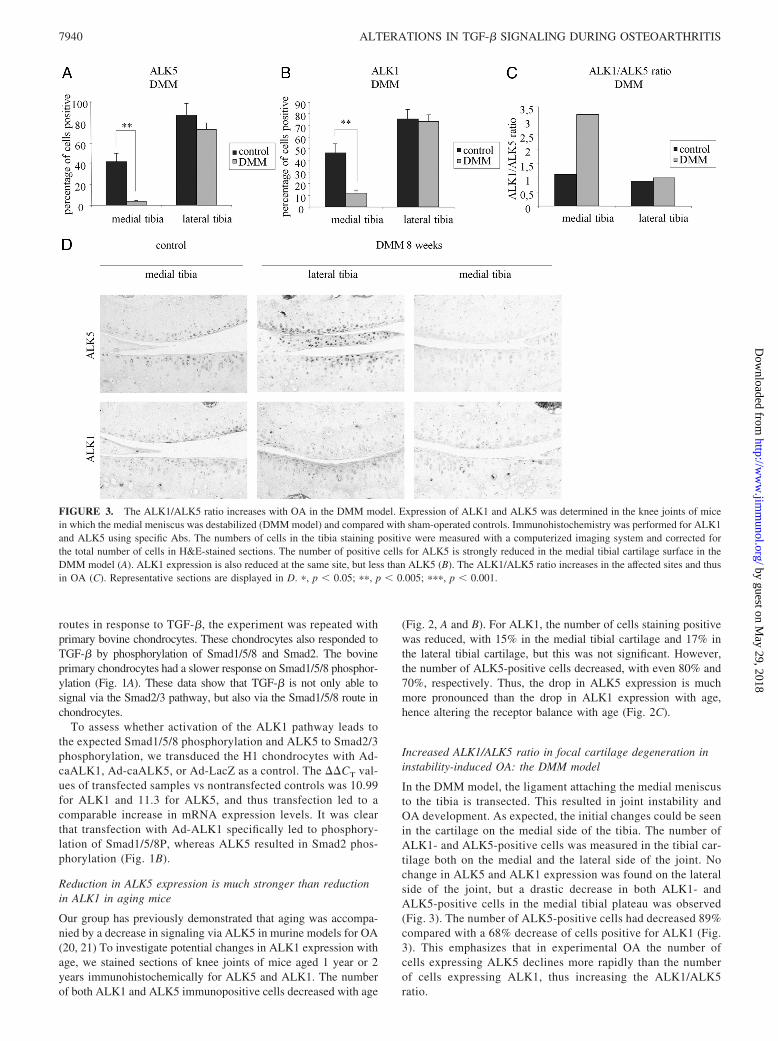
Our group has previously demonstrated that aging was accompa-nied by a decrease in signaling via ALK5 in murine models for OA(20, 21) To investigate potential changes in ALK1 expression withage, we stained sections of knee joints of mice aged 1 year or 2years immunohistochemically for ALK5 and ALK1. The numberof both ALK1 and ALK5 immunopositive cells decreased with age
(Fig. 2, A and B). For ALK1, the number of cells staining positivewas reduced, with 15% in the medial tibial cartilage and 17% inthe lateral tibial cartilage, but this was not significant. However,the number of ALK5-positive cells decreased, with even 80% and70%, respectively. Thus, the drop in ALK5 expression is muchmore pronounced than the drop in ALK1 expression with age,hence altering the receptor balance with age (Fig. 2C).
Increased ALK1/ALK5 ratio in focal cartilage degeneration ininstability-induced OA: the DMM model
In the DMM model, the ligament attaching the medial meniscusto the tibia is transected. This resulted in joint instability andOA development. As expected, the initial changes could be seenin the cartilage on the medial side of the tibia. The number ofALK1- and ALK5-positive cells was measured in the tibial car-tilage both on the medial and the lateral side of the joint. Nochange in ALK5 and ALK1 expression was found on the lateralside of the joint, but a drastic decrease in both ALK1- andALK5-positive cells in the medial tibial plateau was observed(Fig. 3). The number of ALK5-positive cells had decreased 89%compared with a 68% decrease of cells positive for ALK1 (Fig.3). This emphasizes that in experimental OA the number ofcells expressing ALK5 declines more rapidly than the numberof cells expressing ALK1, thus increasing the ALK1/ALK5ratio.
FIGURE 3. The ALK1/ALK5 ratio increases with OA in the DMM model. Expression of ALK1 and ALK5 was determined in the knee joints of micein which the medial meniscus was destabilized (DMM model) and compared with sham-operated controls. Immunohistochemistry was performed for ALK1and ALK5 using specific Abs. The numbers of cells in the tibia staining positive were measured with a computerized imaging system and corrected forthe total number of cells in H&E-stained sections. The number of positive cells for ALK5 is strongly reduced in the medial tibial cartilage surface in theDMM model (A). ALK1 expression is also reduced at the same site, but less than ALK5 (B). The ALK1/ALK5 ratio increases in the affected sites and thusin OA (C). Representative sections are displayed in D. �, p � 0.05; ��, p � 0.005; ���, p � 0.001.
7940 ALTERATIONS IN TGF-� SIGNALING DURING OSTEOARTHRITIS
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
The ALK1/ALK5 ratio increases with OA progression inspontaneous OA: STR/ort mice
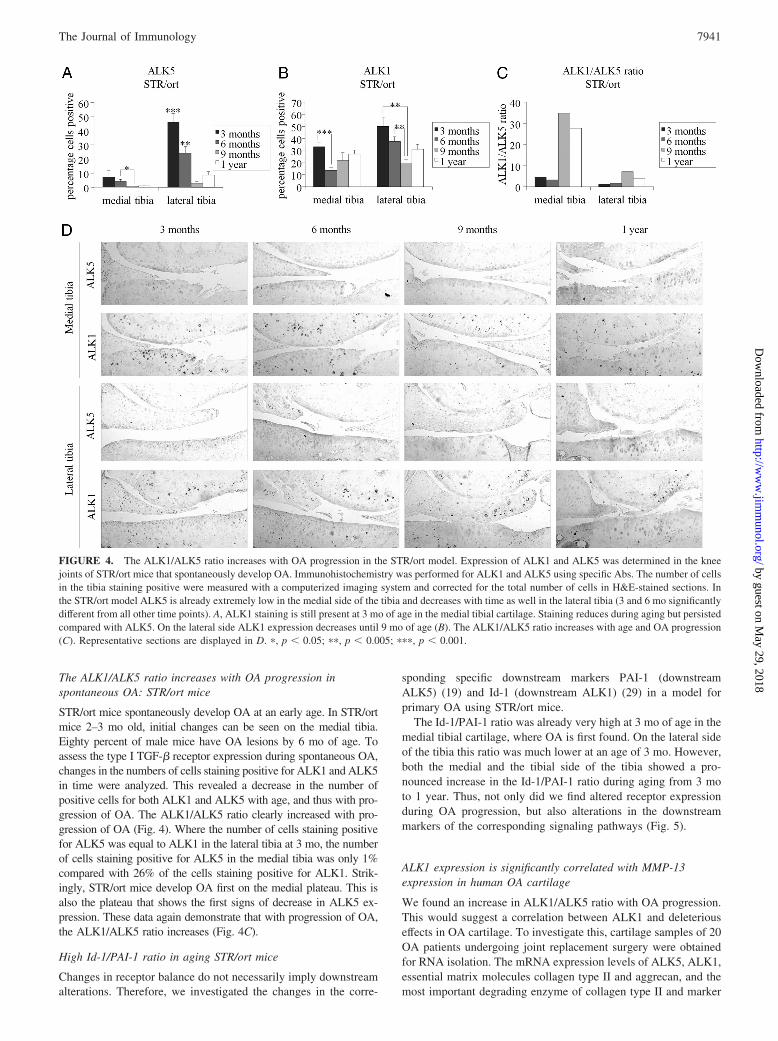
STR/ort mice spontaneously develop OA at an early age. In STR/ortmice 2–3 mo old, initial changes can be seen on the medial tibia.Eighty percent of male mice have OA lesions by 6 mo of age. Toassess the type I TGF-� receptor expression during spontaneous OA,changes in the numbers of cells staining positive for ALK1 and ALK5in time were analyzed. This revealed a decrease in the number ofpositive cells for both ALK1 and ALK5 with age, and thus with pro-gression of OA. The ALK1/ALK5 ratio clearly increased with pro-gression of OA (Fig. 4). Where the number of cells staining positivefor ALK5 was equal to ALK1 in the lateral tibia at 3 mo, the numberof cells staining positive for ALK5 in the medial tibia was only 1%compared with 26% of the cells staining positive for ALK1. Strik-ingly, STR/ort mice develop OA first on the medial plateau. This isalso the plateau that shows the first signs of decrease in ALK5 ex-pression. These data again demonstrate that with progression of OA,the ALK1/ALK5 ratio increases (Fig. 4C).
High Id-1/PAI-1 ratio in aging STR/ort mice
Changes in receptor balance do not necessarily imply downstreamalterations. Therefore, we investigated the changes in the corre-
sponding specific downstream markers PAI-1 (downstreamALK5) (19) and Id-1 (downstream ALK1) (29) in a model forprimary OA using STR/ort mice.
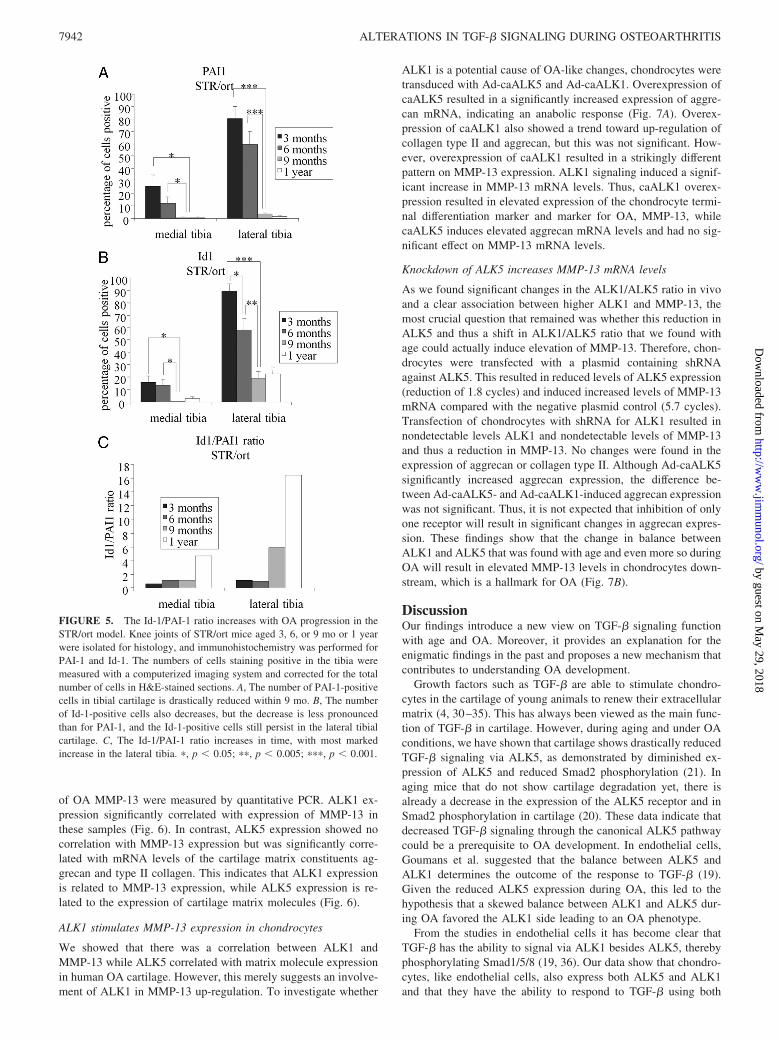
The Id-1/PAI-1 ratio was already very high at 3 mo of age in themedial tibial cartilage, where OA is first found. On the lateral sideof the tibia this ratio was much lower at an age of 3 mo. However,both the medial and the tibial side of the tibia showed a pro-nounced increase in the Id-1/PAI-1 ratio during aging from 3 moto 1 year. Thus, not only did we find altered receptor expressionduring OA progression, but also alterations in the downstreammarkers of the corresponding signaling pathways (Fig. 5).
ALK1 expression is significantly correlated with MMP-13expression in human OA cartilage
We found an increase in ALK1/ALK5 ratio with OA progression.This would suggest a correlation between ALK1 and deleteriouseffects in OA cartilage. To investigate this, cartilage samples of 20OA patients undergoing joint replacement surgery were obtainedfor RNA isolation. The mRNA expression levels of ALK5, ALK1,essential matrix molecules collagen type II and aggrecan, and themost important degrading enzyme of collagen type II and marker
FIGURE 4. The ALK1/ALK5 ratio increases with OA progression in the STR/ort model. Expression of ALK1 and ALK5 was determined in the kneejoints of STR/ort mice that spontaneously develop OA. Immunohistochemistry was performed for ALK1 and ALK5 using specific Abs. The number of cellsin the tibia staining positive were measured with a computerized imaging system and corrected for the total number of cells in H&E-stained sections. Inthe STR/ort model ALK5 is already extremely low in the medial side of the tibia and decreases with time as well in the lateral tibia (3 and 6 mo significantlydifferent from all other time points). A, ALK1 staining is still present at 3 mo of age in the medial tibial cartilage. Staining reduces during aging but persistedcompared with ALK5. On the lateral side ALK1 expression decreases until 9 mo of age (B). The ALK1/ALK5 ratio increases with age and OA progression(C). Representative sections are displayed in D. �, p � 0.05; ��, p � 0.005; ���, p � 0.001.
7941The Journal of Immunology
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
of OA MMP-13 were measured by quantitative PCR. ALK1 ex-pression significantly correlated with expression of MMP-13 inthese samples (Fig. 6). In contrast, ALK5 expression showed nocorrelation with MMP-13 expression but was significantly corre-lated with mRNA levels of the cartilage matrix constituents ag-grecan and type II collagen. This indicates that ALK1 expressionis related to MMP-13 expression, while ALK5 expression is re-lated to the expression of cartilage matrix molecules (Fig. 6).
ALK1 stimulates MMP-13 expression in chondrocytes
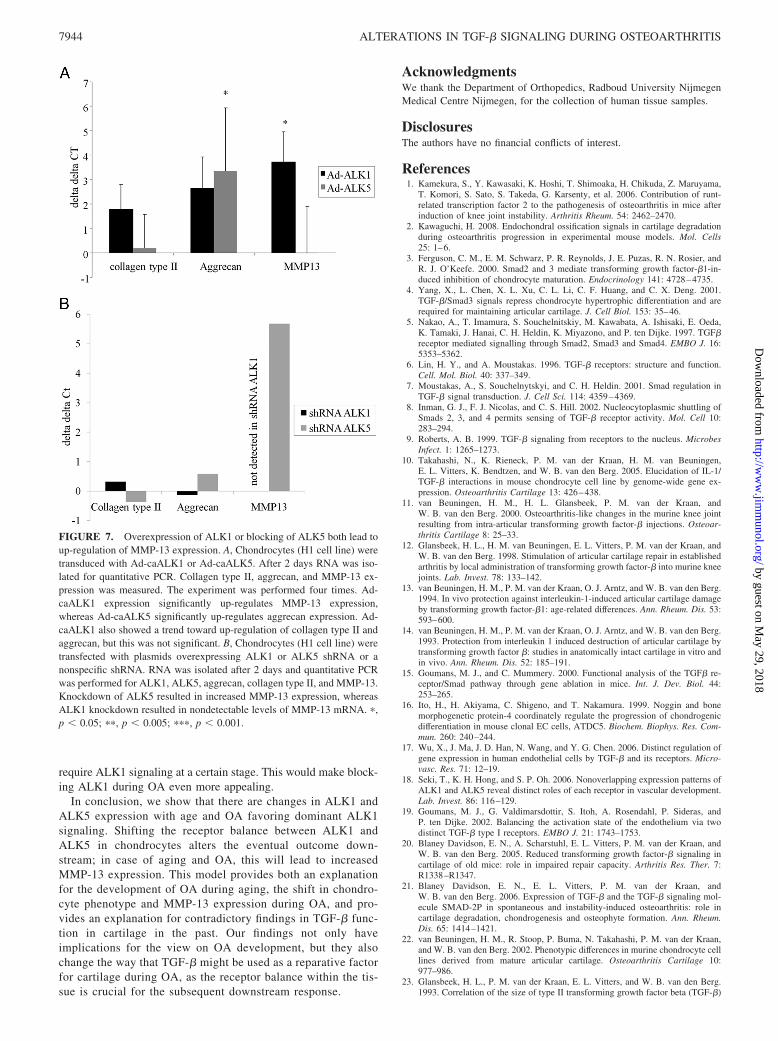
We showed that there was a correlation between ALK1 andMMP-13 while ALK5 correlated with matrix molecule expressionin human OA cartilage. However, this merely suggests an involve-ment of ALK1 in MMP-13 up-regulation. To investigate whether
ALK1 is a potential cause of OA-like changes, chondrocytes weretransduced with Ad-caALK5 and Ad-caALK1. Overexpression ofcaALK5 resulted in a significantly increased expression of aggre-can mRNA, indicating an anabolic response (Fig. 7A). Overex-pression of caALK1 also showed a trend toward up-regulation ofcollagen type II and aggrecan, but this was not significant. How-ever, overexpression of caALK1 resulted in a strikingly differentpattern on MMP-13 expression. ALK1 signaling induced a signif-icant increase in MMP-13 mRNA levels. Thus, caALK1 overex-pression resulted in elevated expression of the chondrocyte termi-nal differentiation marker and marker for OA, MMP-13, whilecaALK5 induces elevated aggrecan mRNA levels and had no sig-nificant effect on MMP-13 mRNA levels.
Knockdown of ALK5 increases MMP-13 mRNA levels
As we found significant changes in the ALK1/ALK5 ratio in vivoand a clear association between higher ALK1 and MMP-13, themost crucial question that remained was whether this reduction inALK5 and thus a shift in ALK1/ALK5 ratio that we found withage could actually induce elevation of MMP-13. Therefore, chon-drocytes were transfected with a plasmid containing shRNAagainst ALK5. This resulted in reduced levels of ALK5 expression(reduction of 1.8 cycles) and induced increased levels of MMP-13mRNA compared with the negative plasmid control (5.7 cycles).Transfection of chondrocytes with shRNA for ALK1 resulted innondetectable levels ALK1 and nondetectable levels of MMP-13and thus a reduction in MMP-13. No changes were found in theexpression of aggrecan or collagen type II. Although Ad-caALK5significantly increased aggrecan expression, the difference be-tween Ad-caALK5- and Ad-caALK1-induced aggrecan expressionwas not significant. Thus, it is not expected that inhibition of onlyone receptor will result in significant changes in aggrecan expres-sion. These findings show that the change in balance betweenALK1 and ALK5 that was found with age and even more so duringOA will result in elevated MMP-13 levels in chondrocytes down-stream, which is a hallmark for OA (Fig. 7B).
DiscussionOur findings introduce a new view on TGF-� signaling functionwith age and OA. Moreover, it provides an explanation for theenigmatic findings in the past and proposes a new mechanism thatcontributes to understanding OA development.
Growth factors such as TGF-� are able to stimulate chondro-cytes in the cartilage of young animals to renew their extracellularmatrix (4, 30–35). This has always been viewed as the main func-tion of TGF-� in cartilage. However, during aging and under OAconditions, we have shown that cartilage shows drastically reducedTGF-� signaling via ALK5, as demonstrated by diminished ex-pression of ALK5 and reduced Smad2 phosphorylation (21). Inaging mice that do not show cartilage degradation yet, there isalready a decrease in the expression of the ALK5 receptor and inSmad2 phosphorylation in cartilage (20). These data indicate thatdecreased TGF-� signaling through the canonical ALK5 pathwaycould be a prerequisite to OA development. In endothelial cells,Goumans et al. suggested that the balance between ALK5 andALK1 determines the outcome of the response to TGF-� (19).Given the reduced ALK5 expression during OA, this led to thehypothesis that a skewed balance between ALK1 and ALK5 dur-ing OA favored the ALK1 side leading to an OA phenotype.
From the studies in endothelial cells it has become clear thatTGF-� has the ability to signal via ALK1 besides ALK5, therebyphosphorylating Smad1/5/8 (19, 36). Our data show that chondro-cytes, like endothelial cells, also express both ALK5 and ALK1and that they have the ability to respond to TGF-� using both
FIGURE 5. The Id-1/PAI-1 ratio increases with OA progression in theSTR/ort model. Knee joints of STR/ort mice aged 3, 6, or 9 mo or 1 yearwere isolated for histology, and immunohistochemistry was performed forPAI-1 and Id-1. The numbers of cells staining positive in the tibia weremeasured with a computerized imaging system and corrected for the totalnumber of cells in H&E-stained sections. A, The number of PAI-1-positivecells in tibial cartilage is drastically reduced within 9 mo. B, The numberof Id-1-positive cells also decreases, but the decrease is less pronouncedthan for PAI-1, and the Id-1-positive cells still persist in the lateral tibialcartilage. C, The Id-1/PAI-1 ratio increases in time, with most markedincrease in the lateral tibia. �, p � 0.05; ��, p � 0.005; ���, p � 0.001.
7942 ALTERATIONS IN TGF-� SIGNALING DURING OSTEOARTHRITIS
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

routes. We show that with age and in OA cartilage, the reductionin ALK5 is far more drastic than the mild reduction in ALK1-positive cells, thus leading toward a skewed balance when com-pared with normal cartilage. The change in the ratio of Id-1/PAI-1expression was in line with a shift from a dominant ALK5 to amore pronounced ALK1 signaling. The initial PAI-1 signaling wasrelatively high, similar to our previous findings of Smad2P expres-sion in young mice (20). Thus, with age, not only is the anabolicfunction of TGF-� signaling reduced, but the terminal-differenti-ation potentiating function that we found in vitro becomes moredominant, thereby potentially making aged cartilage prone todevelop OA.
ALK1 stimulates the Smad1/5/8 route, which is known to stim-ulate terminal differentiation and MMP-13 expression in growthplate chondrocytes. It has been frequently hypothesized that OAchondrocytes undergo differentiation toward a hypertrophic-likestate. Chondrocytes from OA cartilage, even in unaffected areas,already express markers of hypertrophy when compared with nor-mal cartilage (1, 2, 37). In human OA cartilage, ALK5 correlatedwith high aggrecan expression as well as high levels of collagentype II expression, both crucial constituents of the cartilage matrix.In contrast, samples with a high ALK1 expression also had a highMMP-13 expression, which is a hallmark of chondrocyte hyper-trophy and OA.
As in endothelial cells, also altered signaling in chondrocytesleads to a change in outcome: rather than an anabolic response ina predominant ALK5 signaling situation, a shift in receptor bal-ance favoring ALK1 signaling, either by overexpression of ALK1or by blocking ALK5, led to elevation of MMP-13. This not onlyconfirms our human cartilage data, but moreover adds a function tothe correlation that was found: changing the receptor balance be-tween ALK1 and ALK5 favoring the ALK1 side has a functionalconsequence and is able to induce an OA-like phenotype.
Therefore, we postulate that in human OA cartilage, ALK1 sig-naling apparently stimulates type II collagen degradation viaMMP-13 while ALK5 promotes anabolic pathways in chondro-cytes, thereby stimulating the synthesis of cartilage matrix mole-cules. Thus, with age and OA, ALK1 pushes the chondrocytestoward a hypertrophic-like state, expressing MMP-13.
We propose the following hypothesis for OA development andprogression. During aging the ALK1/ALK5 ratio increases and
Smad1/5/8 signaling (ALK1) becomes dominant relative toSmad2/3 signaling (ALK5). This results in stimulation of articularchondrocytes to differentiate to a chondrocyte with a hypertrophic-like phenotype, expressing high levels of MMP-13. High MMP-13expression results in degradation of the cartilage collagen network,making the cartilage matrix vulnerable to normal loading. This isthe developmental stage of cartilage degeneration in age-relatedprimary OA. When cartilage lesions have developed, a subpopu-lation of chondrocytes will show high MMP-13 expression, butanother subpopulation of chondrocytes in the affected cartilagewill show an attempted repair reaction. As a consequence, OAcartilage will be a mixture of cells with a hypertrophic-like phe-notype expressing high ALK1 and MMP-13 levels and a popula-tion of chondrocytes with high ALK5 expression and high expres-sion of matrix molecules such as aggrecan and type II collagen.The ratio of both populations will be dissimilar in different samplesof OA cartilage as a result of OA stage and location of cartilagesampling (distance to severe OA lesions).
A central role for TGF-� in OA becomes apparent. InitiallyTGF-�, signaling via ALK5 in chondrocytes, is a protective factorfor articular cartilage, blocking terminal differentiation andMMP-13 synthesis and stimulating the synthesis of matrix mole-cules. During aging the role of TGF-� changes. Due to variation inreceptor expression (ratio ALK1/ALK5), TGF-� becomes a factorstimulating terminal differentiation and MMP-13 expression viaALK1 signaling. A role for TGF-� in OA joints is confirmed bythe role of TGF-� in the formation of osteophytes and synovialfibrosis, as we have demonstrated in previous studies (31, 32, 35,38). TGF-� is protective for cartilage at a young age but is highlyinvolved in OA pathology, inducing cartilage degradation, osteo-phyte formation, and synovial fibrosis in older individuals. Thisconcept explains our enigmatic findings in earlier studies with re-gard to TGF-�, with TGF-� being both a potent protective factorfor cartilage and a factor that has the ability to induce OA undercertain conditions (10–14). Whether TGF-� induction of osteo-phytes and synovial fibrosis requires ALK5 or ALK1 as a pre-ferred route remains to be investigated. However, a role for ALK1in fibrosis has been suggested (39). Osteophytes undergo chondro-genic differentiation similar to what is observed in the growthplate. It can be anticipated that the cells forming the osteophytes
FIGURE 6. ALK1 is significantlycorrelated with MMP-13 expression,whereas ALK5 is significantly corre-lated with cartilage matrix componentsin human cartilage. Human cartilagewas obtained from patients undergoingknee or hip replacement surgery. RNAwas isolated from the cartilage and aquantitative PCR was performed forALK1, ALK5, aggrecan, collagen typeII, and MMP-13. ALK1 expression issignificantly correlated with MMP-13expression (A and D) while ALK5 ex-pression is significantly related to typeII collagen and aggrecan expression(A–C). �, p � 0.05; ��, p � 0.005; ���,p � 0.001.
7943The Journal of Immunology
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
require ALK1 signaling at a certain stage. This would make block-ing ALK1 during OA even more appealing.
In conclusion, we show that there are changes in ALK1 andALK5 expression with age and OA favoring dominant ALK1signaling. Shifting the receptor balance between ALK1 andALK5 in chondrocytes alters the eventual outcome down-stream; in case of aging and OA, this will lead to increasedMMP-13 expression. This model provides both an explanationfor the development of OA during aging, the shift in chondro-cyte phenotype and MMP-13 expression during OA, and pro-vides an explanation for contradictory findings in TGF-� func-tion in cartilage in the past. Our findings not only haveimplications for the view on OA development, but they alsochange the way that TGF-� might be used as a reparative factorfor cartilage during OA, as the receptor balance within the tis-sue is crucial for the subsequent downstream response.
AcknowledgmentsWe thank the Department of Orthopedics, Radboud University NijmegenMedical Centre Nijmegen, for the collection of human tissue samples.
DisclosuresThe authors have no financial conflicts of interest.
References1. Kamekura, S., Y. Kawasaki, K. Hoshi, T. Shimoaka, H. Chikuda, Z. Maruyama,
T. Komori, S. Sato, S. Takeda, G. Karsenty, et al. 2006. Contribution of runt-related transcription factor 2 to the pathogenesis of osteoarthritis in mice afterinduction of knee joint instability. Arthritis Rheum. 54: 2462–2470.
2. Kawaguchi, H. 2008. Endochondral ossification signals in cartilage degradationduring osteoarthritis progression in experimental mouse models. Mol. Cells25: 1–6.
3. Ferguson, C. M., E. M. Schwarz, P. R. Reynolds, J. E. Puzas, R. N. Rosier, andR. J. O’Keefe. 2000. Smad2 and 3 mediate transforming growth factor-�1-in-duced inhibition of chondrocyte maturation. Endocrinology 141: 4728–4735.
4. Yang, X., L. Chen, X. L. Xu, C. L. Li, C. F. Huang, and C. X. Deng. 2001.TGF-�/Smad3 signals repress chondrocyte hypertrophic differentiation and arerequired for maintaining articular cartilage. J. Cell Biol. 153: 35–46.
5. Nakao, A., T. Imamura, S. Souchelnitskiy, M. Kawabata, A. Ishisaki, E. Oeda,K. Tamaki, J. Hanai, C. H. Heldin, K. Miyazono, and P. ten Dijke. 1997. TGF�receptor mediated signalling through Smad2, Smad3 and Smad4. EMBO J. 16:5353–5362.
6. Lin, H. Y., and A. Moustakas. 1996. TGF-� receptors: structure and function.Cell. Mol. Biol. 40: 337–349.
7. Moustakas, A., S. Souchelnytskyi, and C. H. Heldin. 2001. Smad regulation inTGF-� signal transduction. J. Cell Sci. 114: 4359–4369.
8. Inman, G. J., F. J. Nicolas, and C. S. Hill. 2002. Nucleocytoplasmic shuttling ofSmads 2, 3, and 4 permits sensing of TGF-� receptor activity. Mol. Cell 10:283–294.
9. Roberts, A. B. 1999. TGF-� signaling from receptors to the nucleus. MicrobesInfect. 1: 1265–1273.
10. Takahashi, N., K. Rieneck, P. M. van der Kraan, H. M. van Beuningen,E. L. Vitters, K. Bendtzen, and W. B. van den Berg. 2005. Elucidation of IL-1/TGF-� interactions in mouse chondrocyte cell line by genome-wide gene ex-pression. Osteoarthritis Cartilage 13: 426–438.
11. van Beuningen, H. M., H. L. Glansbeek, P. M. van der Kraan, andW. B. van den Berg. 2000. Osteoarthritis-like changes in the murine knee jointresulting from intra-articular transforming growth factor-� injections. Osteoar-thritis Cartilage 8: 25–33.
12. Glansbeek, H. L., H. M. van Beuningen, E. L. Vitters, P. M. van der Kraan, andW. B. van den Berg. 1998. Stimulation of articular cartilage repair in establishedarthritis by local administration of transforming growth factor-� into murine kneejoints. Lab. Invest. 78: 133–142.
13. van Beuningen, H. M., P. M. van der Kraan, O. J. Arntz, and W. B. van den Berg.1994. In vivo protection against interleukin-1-induced articular cartilage damageby transforming growth factor-�1: age-related differences. Ann. Rheum. Dis. 53:593–600.
14. van Beuningen, H. M., P. M. van der Kraan, O. J. Arntz, and W. B. van den Berg.1993. Protection from interleukin 1 induced destruction of articular cartilage bytransforming growth factor �: studies in anatomically intact cartilage in vitro andin vivo. Ann. Rheum. Dis. 52: 185–191.
15. Goumans, M. J., and C. Mummery. 2000. Functional analysis of the TGF� re-ceptor/Smad pathway through gene ablation in mice. Int. J. Dev. Biol. 44:253–265.
16. Ito, H., H. Akiyama, C. Shigeno, and T. Nakamura. 1999. Noggin and bonemorphogenetic protein-4 coordinately regulate the progression of chondrogenicdifferentiation in mouse clonal EC cells, ATDC5. Biochem. Biophys. Res. Com-mun. 260: 240–244.
17. Wu, X., J. Ma, J. D. Han, N. Wang, and Y. G. Chen. 2006. Distinct regulation ofgene expression in human endothelial cells by TGF-� and its receptors. Micro-vasc. Res. 71: 12–19.
18. Seki, T., K. H. Hong, and S. P. Oh. 2006. Nonoverlapping expression patterns ofALK1 and ALK5 reveal distinct roles of each receptor in vascular development.Lab. Invest. 86: 116–129.
19. Goumans, M. J., G. Valdimarsdottir, S. Itoh, A. Rosendahl, P. Sideras, andP. ten Dijke. 2002. Balancing the activation state of the endothelium via twodistinct TGF-� type I receptors. EMBO J. 21: 1743–1753.
20. Blaney Davidson, E. N., A. Scharstuhl, E. L. Vitters, P. M. van der Kraan, andW. B. van den Berg. 2005. Reduced transforming growth factor-� signaling incartilage of old mice: role in impaired repair capacity. Arthritis Res. Ther. 7:R1338–R1347.
21. Blaney Davidson, E. N., E. L. Vitters, P. M. van der Kraan, andW. B. van den Berg. 2006. Expression of TGF-� and the TGF-� signaling mol-ecule SMAD-2P in spontaneous and instability-induced osteoarthritis: role incartilage degradation, chondrogenesis and osteophyte formation. Ann. Rheum.Dis. 65: 1414–1421.
22. van Beuningen, H. M., R. Stoop, P. Buma, N. Takahashi, P. M. van der Kraan,and W. B. van den Berg. 2002. Phenotypic differences in murine chondrocyte celllines derived from mature articular cartilage. Osteoarthritis Cartilage 10:977–986.
23. Glansbeek, H. L., P. M. van der Kraan, E. L. Vitters, and W. B. van den Berg.1993. Correlation of the size of type II transforming growth factor beta (TGF-�)
FIGURE 7. Overexpression of ALK1 or blocking of ALK5 both lead toup-regulation of MMP-13 expression. A, Chondrocytes (H1 cell line) weretransduced with Ad-caALK1 or Ad-caALK5. After 2 days RNA was iso-lated for quantitative PCR. Collagen type II, aggrecan, and MMP-13 ex-pression was measured. The experiment was performed four times. Ad-caALK1 expression significantly up-regulates MMP-13 expression,whereas Ad-caALK5 significantly up-regulates aggrecan expression. Ad-caALK1 also showed a trend toward up-regulation of collagen type II andaggrecan, but this was not significant. B, Chondrocytes (H1 cell line) weretransfected with plasmids overexpressing ALK1 or ALK5 shRNA or anonspecific shRNA. RNA was isolated after 2 days and quantitative PCRwas performed for ALK1, ALK5, aggrecan, collagen type II, and MMP-13.Knockdown of ALK5 resulted in increased MMP-13 expression, whereasALK1 knockdown resulted in nondetectable levels of MMP-13 mRNA. �,p � 0.05; ��, p � 0.005; ���, p � 0.001.
7944 ALTERATIONS IN TGF-� SIGNALING DURING OSTEOARTHRITIS
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

receptor with TGF-� responses of isolated bovine articular chondrocytes. Ann.Rheum. Dis. 52: 812–816.
24. Blaney Davidson, E. N., E. L. Vitters, F. M. Mooren, N. Oliver, W. B. Berg, andP. M. van der Kraan. 2006. Connective tissue growth factor/CCN2 overexpres-sion in mouse synovial lining results in transient fibrosis and cartilage damage.Arthritis Rheum. 54: 1653–1661.
25. Glasson, S. S., T. J. Blanchet, and E. A. Morris. 2007. The surgical destabiliza-tion of the medial meniscus (DMM) model of osteoarthritis in the 129/SvEvmouse. Osteoarthritis Cartilage 15: 1061–1069.
26. Das Gupta, E. P., T. J. Lyons, J. A. Hoyland, D. M. Lawton, and A. J. Freemont.1993. New histological observations in spontaneously developing osteoarthritisin the STR/ORT mouse questioning its acceptability as a model of human os-teoarthritis. Int. J. Exp. Pathol. 74: 627–634.
27. Mason, R. M., M. G. Chambers, J. Flannelly, J. D. Gaffen, J. Dudhia, andM. T. Bayliss. 2001. The STR/ort mouse and its use as a model of osteoarthritis.Osteoarthritis Cartilage 9: 85–91.
28. Pritzker, K. P., S. Gay, S. A. Jimenez, K. Ostergaard, J. P. Pelletier, P. A. Revell,D. Salter, and W. B. van den Berg. 2006. Osteoarthritis cartilage histopathology:grading and staging. Osteoarthritis Cartilage 14: 13–29.
29. Lebrin, F., M. J. Goumans, L. Jonker, R. L. Carvalho, G. Valdimarsdottir,M. Thorikay, C. Mummery, H. M. Arthur, and P. ten Dijke. 2004. Endoglinpromotes endothelial cell proliferation and TGF-�/ALK1 signal transduction.EMBO J. 23: 4018–4028.
30. Blaney Davidson, E. N., E. L. Vitters, W. B. van den Berg, andP. M. van der Kraan. 2006. TGF �-induced cartilage repair is maintained butfibrosis is blocked in the presence of Smad7. Arthritis Res. Ther. 8: R65.
31. van Beuningen, H. M., P. M. van der Kraan, O. J. Arntz, and W. B. van den Berg.1994. Transforming growth factor-�1 stimulates articular chondrocyte proteo-glycan synthesis and induces osteophyte formation in the murine knee joint. Lab.Invest. 71: 279–290.
32. van Beuningen, H. M., H. L. Glansbeek, P. M. van der Kraan, andW. B. van den Berg. 1998. Differential effects of local application of BMP-2 orTGF-�1 on both articular cartilage composition and osteophyte formation. Os-teoarthritis Cartilage 6: 306–317.
33. Frenkel, S. R., P. B. Saadeh, B. J. Mehrara, G. S. Chin, D. S. Steinbrech, B. Brent,G. K. Gittes, and M. T. Longaker. 2000. Transforming growth factor � super-family members: role in cartilage modeling. Plast. Reconstr. Surg. 105: 980–990.
34. Grimoud, E., D. Heymann, and F. Redini. 2002. Recent advances in TGF-�effects on chondrocyte metabolism potential potential therapeutic roles of TGF-�in cartilage disorders. Cytokine Growth Factor Rev. 13: 241–257.
35. Scharstuhl, A., H. L. Glansbeek, H. M. van Beuningen, E. L. Vitters,P. M. van der Kraan, and W. B. van den Berg. 2002. Inhibition of endogenousTGF-� during experimental osteoarthritis prevents osteophyte formation and im-pairs cartilage repair. J. Immunol. 169: 507–514.
36. Oh, S. P., T. Seki, K. A. Goss, T. Imamura, Y. Yi, P. K. Donahoe, L. Li,K. Miyazono, P. ten Dijke, S. Kim, and E. Li. 2000. Activin receptor-like kinase1 modulates transforming growth factor-�1 signaling in the regulation of angio-genesis. Proc. Natl. Acad. Sci. USA 97: 2626–2631.
37. Yang, K. G., D. B. Saris, R. E. Geuze, M. H. van Rijen, Y. J. van der Helm,A. J. Verbout, L. B. Creemers, and W. J. Dhert. 2006. Altered in vitro chondro-genic properties of chondrocytes harvested from unaffected cartilage in osteoar-thritic joints. Osteoarthritis Cartilage 14: 561–570.
38. Scharstuhl, A., E. L. Vitters, P. M. van der Kraan, and W. B. van den Berg. 2003.Reduction of osteophyte formation and synovial thickening by adenoviral over-expression of transforming growth factor �/bone morphogenetic protein inhibi-tors during experimental osteoarthritis. Arthritis Rheum. 48: 3442–3451.
39. Pannu, J., S. Nakerakanti, E. Smith, P. ten Dijke, and M. Trojanowska. 2007.Transforming growth factor-� receptor type I-dependent fibrogenic gene programis mediated via activation of Smad1 and ERK1/2 pathways. J. Biol. Chem. 282:10405–10413.
7945The Journal of Immunology
by guest on May 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Corrections
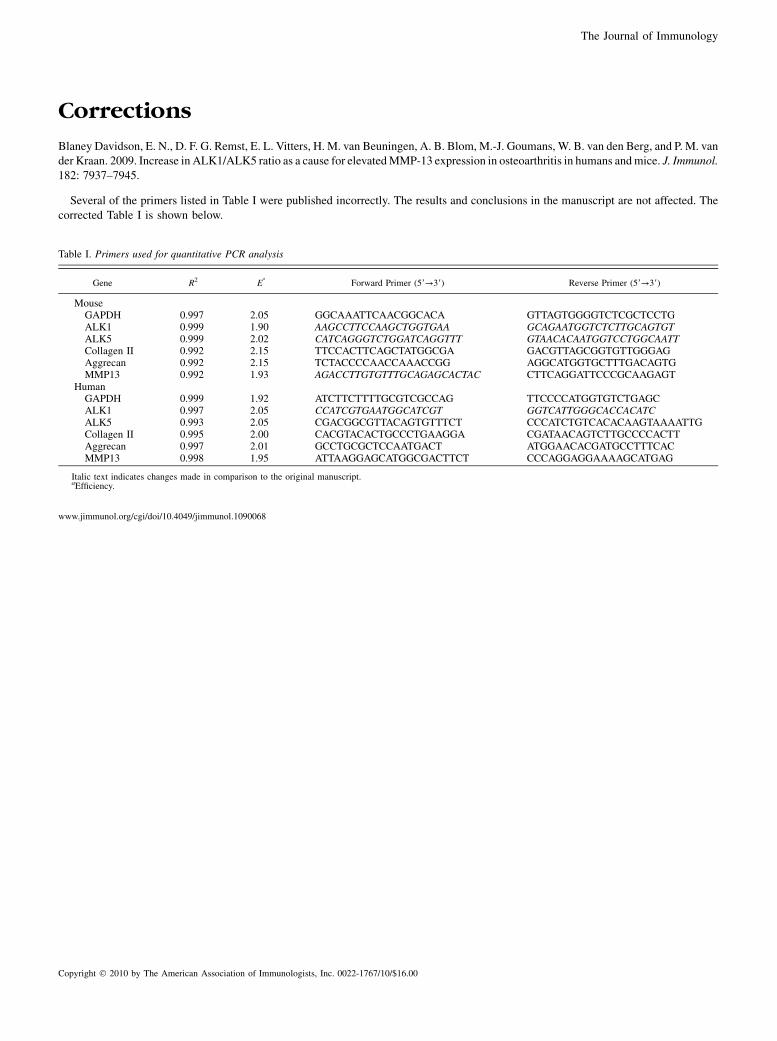
Blaney Davidson, E. N., D. F. G. Remst, E. L. Vitters, H. M. van Beuningen, A. B. Blom, M.-J. Goumans, W. B. van den Berg, and P. M. vander Kraan. 2009. Increase in ALK1/ALK5 ratio as a cause for elevatedMMP-13 expression in osteoarthritis in humans andmice. J. Immunol.182: 7937–7945.
Several of the primers listed in Table I were published incorrectly. The results and conclusions in the manuscript are not affected. Thecorrected Table I is shown below.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1090068
Table I. Primers used for quantitative PCR analysis
Gene R2 Ea
Forward Primer (59→39) Reverse Primer (59→39)
MouseGAPDH 0.997 2.05 GGCAAATTCAACGGCACA GTTAGTGGGGTCTCGCTCCTGALK1 0.999 1.90 AAGCCTTCCAAGCTGGTGAA GCAGAATGGTCTCTTGCAGTGTALK5 0.999 2.02 CATCAGGGTCTGGATCAGGTTT GTAACACAATGGTCCTGGCAATTCollagen II 0.992 2.15 TTCCACTTCAGCTATGGCGA GACGTTAGCGGTGTTGGGAGAggrecan 0.992 2.15 TCTACCCCAACCAAACCGG AGGCATGGTGCTTTGACAGTGMMP13 0.992 1.93 AGACCTTGTGTTTGCAGAGCACTAC CTTCAGGATTCCCGCAAGAGT
HumanGAPDH 0.999 1.92 ATCTTCTTTTGCGTCGCCAG TTCCCCATGGTGTCTGAGCALK1 0.997 2.05 CCATCGTGAATGGCATCGT GGTCATTGGGCACCACATCALK5 0.993 2.05 CGACGGCGTTACAGTGTTTCT CCCATCTGTCACACAAGTAAAATTGCollagen II 0.995 2.00 CACGTACACTGCCCTGAAGGA CGATAACAGTCTTGCCCCACTTAggrecan 0.997 2.01 GCCTGCGCTCCAATGACT ATGGAACACGATGCCTTTCACMMP13 0.998 1.95 ATTAAGGAGCATGGCGACTTCT CCCAGGAGGAAAAGCATGAG
Italic text indicates changes made in comparison to the original manuscript.aEfficiency.
Copyright � 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
The Journal of Immunology





























