Embed Size (px)
Citation preview

Inactivation of Cytochrome P450 2E1 by BenzylIsothiocyanate
Rosa L. Moreno, Ute M. Kent, Kimberly Hodge,† and Paul F. Hollenberg*
Department of Pharmacology, The University of Michigan, Ann Arbor, Michigan 48109
Received January 12, 1999
The cytochrome P450 enzymes constitute a family of phase I enzymes that play a prominentrole in the metabolism of a great variety of endogenous and xenobiotic compounds. In thisstudy, the kinetics for the inactivation of cytochrome P450 2E1 by benzyl isothiocyanate (BITC)were elucidated. BITC is a naturally occurring compound found in cruciferous vegetables suchas broccoli. BITC inhibited the 7-ethoxy-4-(trifluoromethyl)coumarin (7-EFC) O-deethylationactivity of purified and reconstituted P450 2E1 in a time- and concentration-dependent manner.The concentration of inactivator needed for half-maximal inactivation (KI) was 13 µM, andthe maximum rate of inactivation at saturation (kinact) was 0.09 min-1. The partition ratio forthe inactivation of P450 2E1 by BITC was found to have an approximate value of 27.Inactivation of P450 2E1 by BITC was dependent on the presence of NADPH. Followingincubation for 5 min with BITC, a 65% loss in enzymatic activity was observed, whileapproximately 74% of the spectrally detectable enzyme remained. 7-Ethoxycoumarin (7-EC),a substrate of P450 2E1, protected P450 2E1 from BITC inactivation, reducing the loss in7-EFC O-deethylation activity from 50 to 18% when a 1:20 molar ratio of BITC:7-EC was used.Inactivation of P450 2E1 by BITC was irreversible, and no activity was regained after extensivewashes to remove BITC. Addition of cytochrome b5 to the reconstituted system did not affectthe rate of inactivation. Reductase activity was unaffected by BITC. The results reported hereindicate that BITC is a mechanism-based inactivator of cytochrome P450 2E1 and that theinactivation was primarily due to a modification of the apoprotein by BITC.
Introduction
The family of cytochrome P450 (P450)1 enzymes playsa prominent role in the metabolism of a great variety ofendogenous and xenobiotic compounds (1, 2). The basiccatalytic steps that are shared by all the isozymes in thisfamily consist of a two-electron reduction of molecularoxygen to form water and a reactive oxygen species. Thecatalytic diversity of these enzymes has been well estab-lished by studies in which either liver microsomalpreparations or purified proteins were used in thereconstituted system (3). The crystal structures of varioussoluble bacterial P450 enzymes such as P450 101, 102,and 108 have been elucidated (4-6). These crystalstructures have helped to predict regions of the membrane-bound, mammalian P450 proteins that comprise theactive site and that may be critical in the process ofcatalysis. Mammalian P450 enzymes have not beencrystallized to date. Therefore, most of our knowledgeabout the structure of the active site and the relevanceof specific amino acids in catalysis has been derived from
various approaches such as sequence alignments withbacterial P450 enzymes (7-9), site-directed mutagenesis(10, 11), and chemical modifications (12-14). Chemicalmodification of P450 enzymes can be achieved by the useof mechanism-based inactivators (15). These compoundsare substrates for P450 enzymes and form reactiveintermediates in the process of catalysis. These reactivespecies react rapidly with amino acid side chains or theP450 heme (15). Labeling of amino acids in the activesite by these reactive intermediates has been useful inunderstanding the importance of particular residues incatalysis (16, 17).
The inhibition of P450 enzymes has been implicatedas a factor involved in the chemopreventive action of theisothiocyanates (18-20). Isothiocyanates are naturallyoccurring compounds found in cruciferous vegetablessuch as cabbage and broccoli and are hydrolyzed bymyrosinase from glucosinolated precursors (21). Thesecompounds have been extensively studied to determinetheir role as chemopreventive agents and have beenshown to inhibit tumorigenesis associated with exposureto many types of nitrosamines (20, 22, 23). Isothiocyan-ates exhibit their chemopreventive activity when admin-istered either prior to or during exposure to carcinogens.Their chemopreventive activity has been demonstratedin numerous tissues, including rat lung, esophagus, liver,colon, and bladder (23). Two main mechanisms for thechemopreventive action of isothiocyanates have beenidentified: (a) induction of phase II enzymes such asglutathione S-transferase and quinone reductase and (b)inhibition of phase I enzymes such as the P450 enzymes
* To whom correspondence should be addressed: Department ofPharmacology, Medical Science Research Building III, 1150 W. MedicalCenter Dr., Ann Arbor, MI 48109-0632. E-mail: [email protected].
† K.H. was from Meharry Medical College and a recipient of aCharles Ross Fellowship Award.
1 Abbreviations: P450, cytochrome P450; BITC, benzyl isothiocy-anate; reductase, NADPH-cytochrome P450 reductase; DLPC, dilau-royl-L-R-phosphatidylcholine; BSA, bovine serum albumin; 7-EC,7-ethoxycoumarin; 7-EFC, 7-ethoxy-4-(trifluoromethyl)coumarin; 7-HFC,7-hydroxy-4-(trifluoromethyl)coumarin; p-NP, p-nitrophenol; PEITC,phenethyl isothiocyanate; NDMA, N-nitrosodimethylamine; NNN,N-nitrosonornicotine; NNK, 4-(methylnitrosamino)-1-(3-pyridyl)-1-bu-tanone.
582 Chem. Res. Toxicol. 1999, 12, 582-587
10.1021/tx9900019 CCC: $18.00 © 1999 American Chemical SocietyPublished on Web 06/09/1999

(19, 24, 25). Three main mechanisms have been identifiedthat are responsible for the inhibition or inactivation ofP450 enzymes (26, 27). Isothiocyanates inhibit or inac-tivate P450 enzymes by direct interaction of the parentcompound with nucleophilic residues in the enzyme orby the interaction of the isocyanate metabolite withavailable nucleophiles in the active site. In addition,inactivation could also result from a covalent modificationof the enzyme by atomic sulfur produced through oxida-tive desulfuration (26).
This study is the first report where the kinetics for theinactivation of the ethanol-inducible enzyme P450 2E1by benzyl isothiocyanate (BITC) in the reconstitutedsystem were elucidated. Inactivation of P450 2E1 byBITC was NADPH-dependent and proceeded in a time-and concentration-dependent manner. The results re-ported here indicate that BITC is acting as a mechanism-based inactivator of P450 2E1 and that the loss inenzymatic activity is the result of an apoprotein modifi-cation by a reactive intermediate formed during BITCmetabolism.
Experimental Procedures
Materials. BITC and 7-ethoxycoumarin (7-EC) were obtainedfrom Aldrich Chemical Co. (Milwaukee, WI). 7-Ethoxy-4-(tri-fluoromethyl)coumarin (7-EFC) was from Molecular Probes(Eugene, OR). Catalase, purified from bovine liver, NADPH,p-nitrophenol (p-NP), 4-nitrocatechol, dilauroyl-L-R-phosphati-dylcholine (DLPC), Tergitol NP-10, and poly(vinylpyrolidone)were from Sigma Chemical Co. (St. Louis, MO). Topp3 Escheri-chia coli cells were obtained from Stratagene (La Jolla, CA).HPLC grade acetonitrile was from Fisher Scientific (Pittsburgh,PA). [14C]BITC labeled at the R carbon was provided by F. L.Chung (American Health Foundation, Valhalla, NY).
Enzymes. The cDNA for rabbit P450 2E1 (provided by M. J.Coon, The University of Michigan) was expressed in E. coli cells.Expression and purification of the protein were carried outaccording to published methods (28). The cDNA for rat NADPH-P450 reductase within the expression plasmid pOR263 (29) wasexpressed in E. coli Topp3 cells. Expression and purification ofthe protein were as described by Hanna et al. (30).
Inactivation of P450 2E1 by BITC. Purified rabbit P4502E1 (0.5 nmol) was reconstituted with reductase (0.5 nmol) andlipid (300 µg/mL) at 4 °C for 45 min. After reconstitution,catalase (17 000 units/mL) and 0.5 M potassium phosphate (pH7.4) were added to final concentrations of 1800 units/mL and50 mM, respectively. Primary reaction mixtures contained 1 µMP450 2E1, 1 µM reductase, and various concentrations of BITC(5, 8, 10, 20, 40, and 50 µM in a final concentration of 1%methanol). Methanol was added to the control samples insteadof BITC. Reactions were carried out at 30 °C and were initiatedby adding NADPH to a final concentration of 1.2 mM. The lossof activity after 0, 1, 2, and 4 min following the addition ofNADPH was determined spectrofluorometrically by measuringthe extent of O-deethylation of 7-EFC to HFC (31, 32) on anSLM-Aminco model SPF-500C spectrofluorometer with excita-tion at 410 nm (slit width of 5 nm) and emission at 510 nm (slitwidth of 5 nm). For this assay, 20 pmol aliquots of P450 2E1were removed from the primary reaction mixture and added tosecondary reaction mixtures containing 0.1 mM 7-EFC, and 0.2mM NADPH in 50 mM potassium phosphate (pH 7.4) with 0.04mg/mL BSA in a final volume of 1 mL. Reactions were allowedto proceed for 15 min and were terminated by adding 0.33 mLof ice-cold acetonitrile.
Effect of BITC on P450 2E1 Activity and Content.Reconstitution and reaction conditions were as described above.The activity remaining at time zero and after incubation for 5min with 100 µM BITC was determined using the 7-EFC
O-deethylation assay. The P450 content was measured by thereduced carbon monoxide difference spectrum (33).
P450 2E1 Heme Analysis. Reconstitution of P450 2E1 (1.1nmol) and reductase (1.1 nmol) was as described previously.Primary reaction mixtures contained 2 µM P450 2E1, 2 µMreductase, and 200 µM BITC. Reactions were carried out at 30°C, and inactivation proceeded for 20 min. The loss of activitywas determined by measuring the extent of O-deethylation of7-EFC. To determine the changes in P450 2E1 heme content,samples were analyzed by HPLC on a 4.6 mm × 250 mm VydacC4 column with a solvent system consisting of buffer A (0.1%H2O/TFA) and buffer B (95% CH3CN/0.1% TFA). The hemecontent was determined by measuring the absorbance at 405nm. The heme eluted at 4 min using a gradient of 40 to 50% Bover the course of 15 min at a flow rate of 1.0 mL/min.
Effect of Cytochrome b5 on the Inactivation of P450 2E1by BITC. Cytochrome b5 was reconstituted together with P4502E1 and reductase at a molar concentration of 1 µM each.Samples were inactivated with BITC and assayed for residual7-EFC O-deethylation activity as described above.
Partition Ratio. The partition ratio for the inactivation ofP450 2E1 by BITC was determined by incubating the enzymewith increasing concentrations of BITC and allowing theinactivation to go to completion. The reconstitution was carriedout as described above. Following reconstitution, catalase andpotassium phosphate (pH 7.4) were added to final concentrationsof 1800 units/mL and 50 mM, respectively. Primary reactionmixtures contained 1 µM P450 2E1 and 1 µM reductase.Reactions were carried out for 10 min at 30 °C, and the activityremaining after inactivation was determined by measuring theextent of 7-EFC O-deethylation activity as previously described.The percent activity remaining was plotted as a function of themolar ratio of BITC to P450 2E1. The partition ratio wasobtained from the intercept between the line obtained by linearregression with the lower BITC concentrations (0-16 µM) andthe straight line obtained with higher BITC concentrations (40-100 µM) (34).
Effect of 7-EC on the Rates of Inactivation of P450 2E1by BITC. P450 2E1 and reductase were reconstituted asdescribed above. Following reconstitution, catalase and potas-sium phosphate (pH 6.8) were added to final concentrations of232 units/mL and 78 mM, respectively. Primary reactionmixtures containing 0.4 µM P450 2E1, 0.8 µM reductase, 10 µMBITC, and increasing concentrations of 7-EC (20-200 µM) wereincubated at 30 °C for 0, 2, 4, 6, and 8 min. The activityremaining was determined electrochemically by measuring theextent of formation of 4-nitrocatechol from p-NP as previouslydescribed (31, 35). Secondary reaction mixtures contained 5 mMMgCl2, 0.2 mM p-NP, 0.5 mM NADPH, and 40 µg/mL BSA in100 mM potassium phosphate (pH 6.8).
Irreversibility of P450 2E1 Inactivation by BITC. P4502E1 and reductase were reconstituted as described previously.Following inactivation with 50 µM BITC for 5 min, the loss inactivity was measured with the 7-EFC O-deethylation assay.The P450 content was determined by diluting an aliquot of thereaction mixture 1:10 in 50 mM potassium phosphate (pH 7.7)containing 40% glycerol and 0.6% Tergitol. The remainder ofthe primary reaction mixtures was loaded into Microcon 30microconcentrators (Amicon, Inc., Beverly, MA) that had beenpretreated overnight with 0.5% poly(vinylpyrolidone) in 100 mMacetic acid. Samples were concentrated by centrifuging at 8000gfollowed by three washes (200 µL each) with 50 mM potassiumphosphate (pH 7.4), containing 20% glycerol. Concentratedproteins were reconstituted with 50 µg of DLPC at 4 °C for 20min and diluted to their original volume with 50 mM potassiumphosphate (pH 7.4). Activity was measured as described previ-ously and corrected for P450 2E1 recovery. The P450 2E1concentration was determined from the reduced carbon mon-oxide difference spectrum.
Binding Spectra and Binding Constant. P450 2E1 (1nmol) was reconstituted with 100 µg of lipid for 45 min at 4 °C.The reconstituted protein was diluted to 0.5 µM with 50 mM
Inactivation of P450 2E1 by Benzyl Isothiocyanate Chem. Res. Toxicol., Vol. 12, No. 7, 1999 583
potassium phosphate (pH 7.4). The sample was divided, andequal volumes were added to a reference and a sample cuvette.Samples were scanned from 350 to 500 nm on a DW2-UV/visspectrophotometer (SLM Aminco, Urbana, IL) equipped withan OLIS spectroscopy operating system (On-Line InstrumentSystems, Inc., Bogart, GA). Following the addition of 1 µL (2.1nmol) aliquots of BITC dissolved in methanol to the samplecuvette and 1 µL aliquots of methanol to the reference cuvette,scans were recorded for BITC concentrations of 2.1-12.6 µM.The binding constant, Ks, was calculated from a plot of theinverse of the change in absorption versus the inverse of theBITC concentration. The Ks was obtained from the x-interceptof the linear regression line (36).
Covalent Binding. Reconstitution was carried out as de-scribed above with 1.65 nmol of P450 2E1, 1.65 nmol ofreductase, and 100 µg of lipid. Primary reaction mixturescontaining P450 2E1 and reductase at a concentration of 2 µMeach were treated either with 45 µM [14C]BITC and NADPH orwith [14C]BITC alone. Samples were incubated for 20 min at30 °C and then dialyzed against 50 mM potassium phosphatebuffer (pH 7.4) containing 20% glycerol, 0.1 M EDTA, and 0.4%cholate. Following dialysis, 40 pmol aliquots of P450 2E1 fromeach sample were denatured in the presence or absence ofâ-mercaptoethanol and were loaded onto a 10% polyacrylamidegel. Proteins were separated by SDS-PAGE and transferredto a nitrocellulose membrane. Autoradiography was performedby exposing the membrane to Biomax MR film supplementedwith a Biomax intensifying screen (Eastman Kodak Co.,Rochester, NY) for 2 weeks at -80 °C.
Results
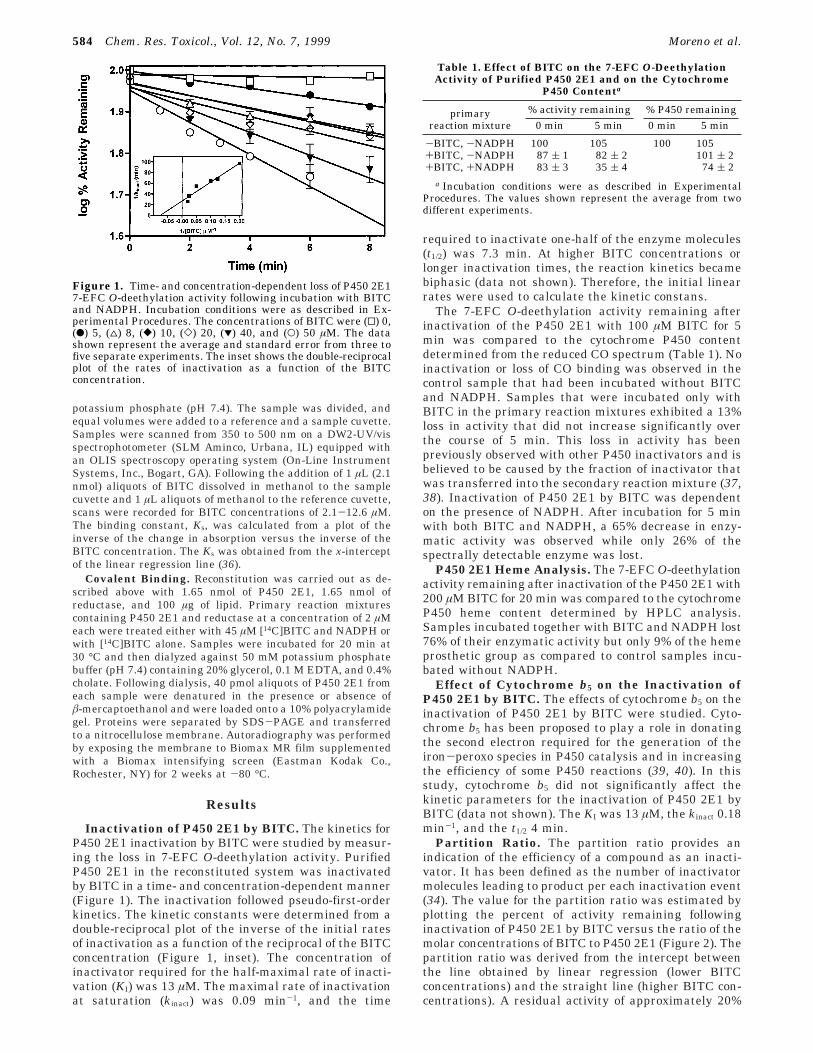
Inactivation of P450 2E1 by BITC. The kinetics forP450 2E1 inactivation by BITC were studied by measur-ing the loss in 7-EFC O-deethylation activity. PurifiedP450 2E1 in the reconstituted system was inactivatedby BITC in a time- and concentration-dependent manner(Figure 1). The inactivation followed pseudo-first-orderkinetics. The kinetic constants were determined from adouble-reciprocal plot of the inverse of the initial ratesof inactivation as a function of the reciprocal of the BITCconcentration (Figure 1, inset). The concentration ofinactivator required for the half-maximal rate of inacti-vation (KI) was 13 µM. The maximal rate of inactivationat saturation (kinact) was 0.09 min-1, and the time
required to inactivate one-half of the enzyme molecules(t1/2) was 7.3 min. At higher BITC concentrations orlonger inactivation times, the reaction kinetics becamebiphasic (data not shown). Therefore, the initial linearrates were used to calculate the kinetic constans.
The 7-EFC O-deethylation activity remaining afterinactivation of the P450 2E1 with 100 µM BITC for 5min was compared to the cytochrome P450 contentdetermined from the reduced CO spectrum (Table 1). Noinactivation or loss of CO binding was observed in thecontrol sample that had been incubated without BITCand NADPH. Samples that were incubated only withBITC in the primary reaction mixtures exhibited a 13%loss in activity that did not increase significantly overthe course of 5 min. This loss in activity has beenpreviously observed with other P450 inactivators and isbelieved to be caused by the fraction of inactivator thatwas transferred into the secondary reaction mixture (37,38). Inactivation of P450 2E1 by BITC was dependenton the presence of NADPH. After incubation for 5 minwith both BITC and NADPH, a 65% decrease in enzy-matic activity was observed while only 26% of thespectrally detectable enzyme was lost.
P450 2E1 Heme Analysis. The 7-EFC O-deethylationactivity remaining after inactivation of the P450 2E1 with200 µM BITC for 20 min was compared to the cytochromeP450 heme content determined by HPLC analysis.Samples incubated together with BITC and NADPH lost76% of their enzymatic activity but only 9% of the hemeprosthetic group as compared to control samples incu-bated without NADPH.
Effect of Cytochrome b5 on the Inactivation ofP450 2E1 by BITC. The effects of cytochrome b5 on theinactivation of P450 2E1 by BITC were studied. Cyto-chrome b5 has been proposed to play a role in donatingthe second electron required for the generation of theiron-peroxo species in P450 catalysis and in increasingthe efficiency of some P450 reactions (39, 40). In thisstudy, cytochrome b5 did not significantly affect thekinetic parameters for the inactivation of P450 2E1 byBITC (data not shown). The KI was 13 µM, the kinact 0.18min-1, and the t1/2 4 min.
Partition Ratio. The partition ratio provides anindication of the efficiency of a compound as an inacti-vator. It has been defined as the number of inactivatormolecules leading to product per each inactivation event(34). The value for the partition ratio was estimated byplotting the percent of activity remaining followinginactivation of P450 2E1 by BITC versus the ratio of themolar concentrations of BITC to P450 2E1 (Figure 2). Thepartition ratio was derived from the intercept betweenthe line obtained by linear regression (lower BITCconcentrations) and the straight line (higher BITC con-centrations). A residual activity of approximately 20%
Figure 1. Time- and concentration-dependent loss of P450 2E17-EFC O-deethylation activity following incubation with BITCand NADPH. Incubation conditions were as described in Ex-perimental Procedures. The concentrations of BITC were (0) 0,(b) 5, (4) 8, ([) 10, (]) 20, (1) 40, and (O) 50 µM. The datashown represent the average and standard error from three tofive separate experiments. The inset shows the double-reciprocalplot of the rates of inactivation as a function of the BITCconcentration.
Table 1. Effect of BITC on the 7-EFC O-DeethylationActivity of Purified P450 2E1 and on the Cytochrome
P450 Contenta
% activity remaining % P450 remainingprimaryreaction mixture 0 min 5 min 0 min 5 min
-BITC, -NADPH 100 105 100 105+BITC, -NADPH 87 ( 1 82 ( 2 101 ( 2+BITC, +NADPH 83 ( 3 35 ( 4 74 ( 2
a Incubation conditions were as described in ExperimentalProcedures. The values shown represent the average from twodifferent experiments.
584 Chem. Res. Toxicol., Vol. 12, No. 7, 1999 Moreno et al.
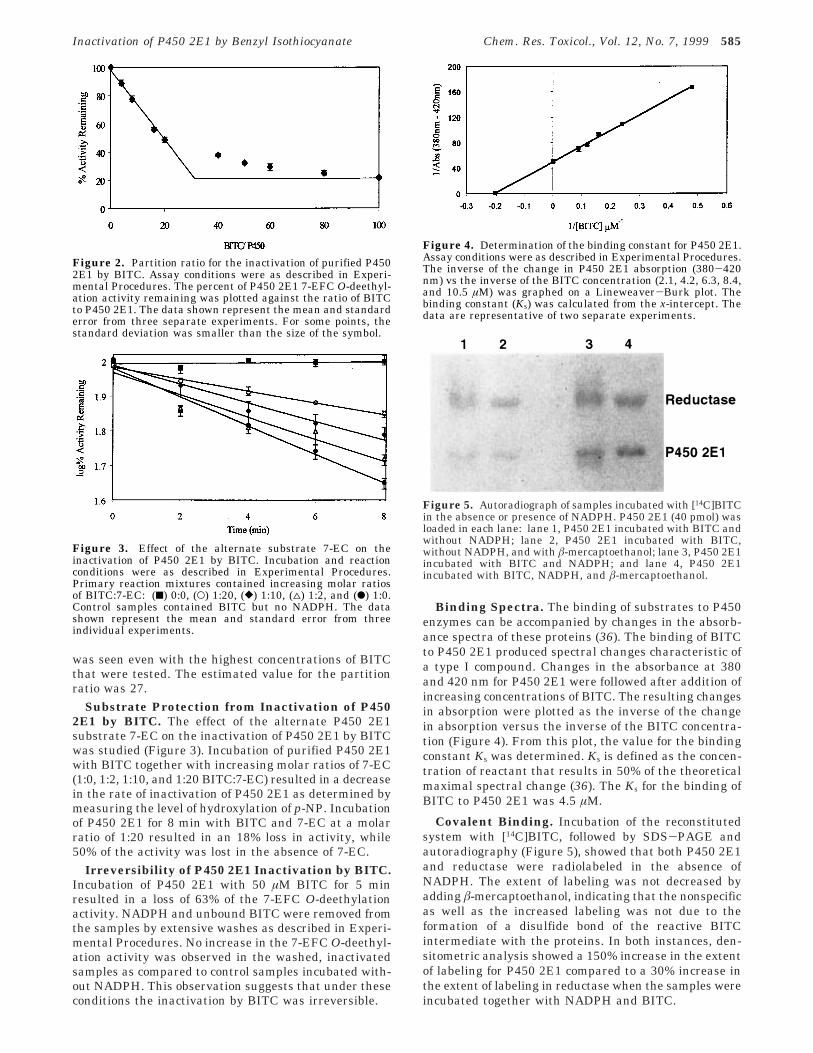
was seen even with the highest concentrations of BITCthat were tested. The estimated value for the partitionratio was 27.
Substrate Protection from Inactivation of P4502E1 by BITC. The effect of the alternate P450 2E1substrate 7-EC on the inactivation of P450 2E1 by BITCwas studied (Figure 3). Incubation of purified P450 2E1with BITC together with increasing molar ratios of 7-EC(1:0, 1:2, 1:10, and 1:20 BITC:7-EC) resulted in a decreasein the rate of inactivation of P450 2E1 as determined bymeasuring the level of hydroxylation of p-NP. Incubationof P450 2E1 for 8 min with BITC and 7-EC at a molarratio of 1:20 resulted in an 18% loss in activity, while50% of the activity was lost in the absence of 7-EC.
Irreversibility of P450 2E1 Inactivation by BITC.Incubation of P450 2E1 with 50 µM BITC for 5 minresulted in a loss of 63% of the 7-EFC O-deethylationactivity. NADPH and unbound BITC were removed fromthe samples by extensive washes as described in Experi-mental Procedures. No increase in the 7-EFC O-deethyl-ation activity was observed in the washed, inactivatedsamples as compared to control samples incubated with-out NADPH. This observation suggests that under theseconditions the inactivation by BITC was irreversible.
Binding Spectra. The binding of substrates to P450enzymes can be accompanied by changes in the absorb-ance spectra of these proteins (36). The binding of BITCto P450 2E1 produced spectral changes characteristic ofa type I compound. Changes in the absorbance at 380and 420 nm for P450 2E1 were followed after addition ofincreasing concentrations of BITC. The resulting changesin absorption were plotted as the inverse of the changein absorption versus the inverse of the BITC concentra-tion (Figure 4). From this plot, the value for the bindingconstant Ks was determined. Ks is defined as the concen-tration of reactant that results in 50% of the theoreticalmaximal spectral change (36). The Ks for the binding ofBITC to P450 2E1 was 4.5 µM.
Covalent Binding. Incubation of the reconstitutedsystem with [14C]BITC, followed by SDS-PAGE andautoradiography (Figure 5), showed that both P450 2E1and reductase were radiolabeled in the absence ofNADPH. The extent of labeling was not decreased byadding â-mercaptoethanol, indicating that the nonspecificas well as the increased labeling was not due to theformation of a disulfide bond of the reactive BITCintermediate with the proteins. In both instances, den-sitometric analysis showed a 150% increase in the extentof labeling for P450 2E1 compared to a 30% increase inthe extent of labeling in reductase when the samples wereincubated together with NADPH and BITC.
Figure 2. Partition ratio for the inactivation of purified P4502E1 by BITC. Assay conditions were as described in Experi-mental Procedures. The percent of P450 2E1 7-EFC O-deethyl-ation activity remaining was plotted against the ratio of BITCto P450 2E1. The data shown represent the mean and standarderror from three separate experiments. For some points, thestandard deviation was smaller than the size of the symbol.
Figure 3. Effect of the alternate substrate 7-EC on theinactivation of P450 2E1 by BITC. Incubation and reactionconditions were as described in Experimental Procedures.Primary reaction mixtures contained increasing molar ratiosof BITC:7-EC: (9) 0:0, (O) 1:20, ([) 1:10, (4) 1:2, and (b) 1:0.Control samples contained BITC but no NADPH. The datashown represent the mean and standard error from threeindividual experiments.
Figure 4. Determination of the binding constant for P450 2E1.Assay conditions were as described in Experimental Procedures.The inverse of the change in P450 2E1 absorption (380-420nm) vs the inverse of the BITC concentration (2.1, 4.2, 6.3, 8.4,and 10.5 µM) was graphed on a Lineweaver-Burk plot. Thebinding constant (Ks) was calculated from the x-intercept. Thedata are representative of two separate experiments.
Figure 5. Autoradiograph of samples incubated with [14C]BITCin the absence or presence of NADPH. P450 2E1 (40 pmol) wasloaded in each lane: lane 1, P450 2E1 incubated with BITC andwithout NADPH; lane 2, P450 2E1 incubated with BITC,without NADPH, and with â-mercaptoethanol; lane 3, P450 2E1incubated with BITC and NADPH; and lane 4, P450 2E1incubated with BITC, NADPH, and â-mercaptoethanol.
Inactivation of P450 2E1 by Benzyl Isothiocyanate Chem. Res. Toxicol., Vol. 12, No. 7, 1999 585

Discussion
The interaction between purified P450 2E1 in thereconstituted system and BITC was studied for the firsttime in this report. One of the main goals of this studywas to evaluate BITC as a possible mechanism-basedinactivator of P450 2E1 for future use in labeling criticalamino acid residues in the active site. Since BITC is anaturally occurring compound in crusiferous vegetables,its potential inhibitory effect on P450 2E1 takes onadditional physiological relevance as a potential chemo-preventive agent. P450 2E1 has been suggested to playa major role in the activation of short-chain N-nitros-amines to carcinogenic species (41). In human livermicrosomes, P450 2E1 is reported to be the isozymemostly responsible for the activation of N-nitrosodi-methylamine (NDMA) and N-nitrosonornicotine (NNN)and it also contributes to the N-demethylation of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) (41).The inactivation of P450 2E1 by BITC followed pseudo-first-order kinetics, was time- and concentration-depend-ent, and required NADPH. The observed significant lossof 7-EFC O-deethylation activity did not correlate withthe minimal decrease in the level of CO binding inducedby BITC. This result indicated that inactivation occurredmainly due to a modification of the apoprotein ratherthan the heme. The observed minor decrease in CObinding does not necessarily imply that a heme modifica-tion has occurred. A similar phenomenon has beenobserved with other isozymes or inactivators where adecrease in the extent of CO binding did not correlatewith heme content determined by HPLC analysis (31, 42).In those studies, HPLC analysis of the modified proteinat 405 nm showed virtually no change in the peakcorresponding to the heme following inactivation eventhough the amount of CO-reactive heme declined signifi-cantly. The 9% loss in the ability of the BITC-modifiedP450 2E1 to bind CO did not correlate with the 76% lossin P450 2E1 O-deethylation activity. These observationssuggested that the primary site of modification by BITCoccurred on the apoprotein.
The modification by the reactive BITC intermediateappeared to be covalent since extensive washing did notlead to a recovery of enzymatic activity. The absoluterequirement of NADPH for inactivation and the protec-tion against inactivation offered by 7-EC confirmed theneed for BITC to be metabolically activated. These resultsalso suggested that the P450 2E1 active site was theplace where the inactivating event occurred. BITC didnot appear to affect the enzymatic activity of NADPH-P450 reductase. Further studies were performed to ruleout an effect of BITC on the reductase. Mainly, inactiva-tion of P450 2E1 by BITC did not affect the ability of thereductase to reduce cytochrome c (data not shown).Phenethyl isothiocyanate, another naturally occurringstructurally related isothiocyanate, was also shown toinhibit P450 2E1 without effecting reductase activitybased on cytochrome c reduction assays (43). Also,addition of new reductase to the inactivated sample didnot result in recovery of activity (data not shown). Eventhough labeling of reductase by BITC was observed inthe absence and presence of NADPH, the above resultsindicate that this modification is not contributing to theloss in P450 2E1 activity. The labeling of both P450 2E1and reductase in the absence of NADPH is not surprisingon the basis of the known reactivity of isothiocyanates
with nucleophilic residues in proteins (18), resulting inthe inhibition or inactivation of enzymes by isothiocyan-ate compounds in general.
The partition ratio was estimated by adding increasingconcentrations of BITC to the reaction mixture andletting the inactivation reaction go to completion. Thepoints derived from the higher ratios of BITC to P4502E1 deviated from the linear regression line. At thehighest concentrations tested, the maximal loss in activ-ity that could be obtained was 80%. The possibility thatthis was due to product inhibition is unlikely. Residualactivity was tested by diluting an aliquot of the primaryreaction by 50-fold into the secondary reaction. Therefore,the highest concentration of BITC carried over into thesecondary reaction was 2 µM. Further, extensive wash-ings of the BITC-inactivated sample did not restoreenzymatic activity. The extent of inactivation appears tobe dependent on the P450 isozyme and the type ofinactivator used (13-15, 31, 37, 42). One possibility thatshould be considered is that certain modifications greatlycompromise either substrate binding or catalysis but donot completely inactivate the 7-EFC O-deethylationactivity. Studies with naturally occurring or engineeredmutant enzymes have shown that single amino acidchanges can influence the rate of catalysis or the natureof the metabolites formed (10, 16, 17, and referencestherein, 30).
In preliminary HPLC studies with [14C]BITC, a stoi-chiometric binding of 0.4-1 labeled metabolite bound pernanomole of P450 2E1 was observed (data not shown).Several factors may have contributed to a <1:1 stoichio-metric binding in this system. The lability of the adductunder the acidic conditions used in the HPLC analysiscould reduce the total radioactivity associated with theprotein. Also, since one of the potential pathways forinactivation by isothiocyanates involves the covalentmodification of the protein by atomic sulfur, the involve-ment of this pathway for inactivation cannot be ruled outand could significantly decrease the stoichiometry ofbinding to less than the expected value of 1. However,the evidence presented here indicates that protein modi-fication by a metabolite of [14C]BITC was the major factorresponsible for the observed decrease in the activity.
The results of this study show that the naturallyoccurring isothiocyanate, BITC, functioned as a mecha-nism-based inactivator of cytochrome P450 2E1. Theinactivation occurred as the result of an irreversiblemodification primarily to the apoprotein. In combination,our observaions suggest that BITC will be a usefulcompound for labeling the P450 2E1 active site. Studiesthat are directed at identifying the labeled peptide andcritical amino acid(s) as well as the reactive intermediateresulting from the metabolism of BITC by P450 2E1 arein progress.
Acknowledgment. We are grateful to Dr. ElizabethS. Roberts and Jenny Juncaj for the expression andpurification of rabbit P450 2E1. We thank Dr. Minor J.Coon for providing us with the cDNA for rabbit P450 2E1.We are also grateful to Dr. Fung-Lung Chung whoprovided us with the [14C]BITC. This publication wassupported in part by NIH Grants CA 16954 (P.F.H.) andCA 46535 (Fung-Lung Chung) from the National CancerInstitute. Its contents are solely the responsibility of theauthors and do not necessarily represent the officialviews of the National Cancer Institute.
586 Chem. Res. Toxicol., Vol. 12, No. 7, 1999 Moreno et al.

References
(1) Gonzalez, F. J. (1998) The molecular biology of cytochrome P450s.Pharmacol. Rev. 40, 243-288.
(2) Nelson, D. R., Koymans, L., Kamataki, T., Stegeman, J. J.,Feyereisen, R., Waxman, D. J., Waterman, M. R., Gotoh, O., Coon,M. J., Eastabrook, R. W., Gunsalus, I. C., and Nebert, D. W. (1996)P450 superfamily: update on new sequences, gene mapping,accession numbers and nomenclature. Pharmacogenetics 6, 1-42.
(3) Guengerich, F. P. (1994) Analysis and characterization of en-zymes. In Principles and Methods of Toxicology (Hayes, A. W.,Ed.) pp 1259-1312, Raven Press, Ltd., New York.
(4) Poulos, T. L., Finzel, B. C., and Howard, A. J. (1987) High-resolution crystal structure of cytochrome P450cam. J. Mol. Biol.195, 687-700.
(5) Ravichandran, K. G., Boddupalli, S. S., Hasemann, C. A., Peter-son, J. A., and Deisenhofer, J. (1993) Crystal structure ofhemoprotein domain of P450 BM-3, a prototype for microsomalP450s. Science 261, 731-736.
(6) Hasemann, C. A., Ravichandran, K. G., Peterson, J. A., andDeisenhofer, J. (1994) Crystal structure and refinement of cyto-chrome P450 terp at 2.3 Å resolution. J. Mol. Biol. 236, 1169-1185.
(7) Lewis, D. F. V. (1995) Three-dimensional models of human andother mammalian microsomal P450s constructed from an align-ment with P450 102 (P450 bm3). Xenobiotica 25, 333-366.
(8) Nelson, D. R., and Strobel, H. W. (1989) Secondary structureprediction of 52 membrane-bound cytochromes P450 shows astrong structural similarity to P450cam. Biochemistry 28, 656-660.
(9) Poulos, T. L. (1991) Modeling of mammalian P450s on the basisof P450cam X-ray structure. Methods Enzymol. 206, 11-30.
(10) Halpert, J. R. (1995) Structural basis of selective cytochrome P450inhibition. Annu. Rev. Pharmacol. Toxicol. 35, 29-53.
(11) Johnson, E. F., Kronbach, T., and Hsu, M. H. (1992) Analysis ofthe catalytic specificity of cytochrome P450 enzymes through site-directed mutagenesis. FASEB J. 6, 700-705.
(12) Murray, M. (1997) Drug-mediated inactivation of cytochromeP450. Clin. Exp. Pharmacol. Physiol. 24, 465-470.
(13) Roberts, E. S., Hopkins, N. E., Alworth, W. L., and Hollenberg,P. F. (1993) Mechanism-based inactivation of cytochrome P4502B1 by 2-ethynylnaphthalene: identification of an active sitepeptide. Chem. Res. Toxicol. 6, 470-479.
(14) Kent, U. M., Bend, J. R., Chamberlin, B. A., Gage, D. A., andHollenberg, P. F. (1997) Mechanism-based inactivation of cyto-chrome P450 2B1 by N-benzyl-1-aminobenzotriazole. Chem. Res.Toxicol. 10, 600-608.
(15) Roberts, E. S., Hopkins, N. E., Zaluzec, E. J., Douglas, G. A.,Alworth, W. L., and Hollenberg, P. F. (1995) Mechanism-basedinactivation of cytochrome P450 2B1 by 9-ethynylphenanthrene.Arch. Biochem. Biophys. 323, 295-302.
(16) Roberts, E. S., Pernecky, S. J., Alworth, W. L., and Hollenberg,P. F. (1996) A role for threonine 302 in the mechanism-basedinactivation of P450 2B4 by 2-ethynylnaphthalene. Arch. Biochem.Biophys. 331, 170-176.
(17) Kent, U. M., Hanna, I. H., Szklarz, G. D., Vaz, A. D. N., Halpert,J. R., Bend, J. R., and Hollenberg, P. F. (1997) Significance ofglycine 478 in the metabolism of N-benzyl-1-aminobenzotriazoleto reactive intermediates by cytochrome P450 2B1. Biochemistry36, 11707-11716.
(18) Chung, F. L. (1992) Chemoprevention of lung carcinogenesis byaromatic isothiocyanates. In Cancer Chemoprevention (Watten-berg, L., Lipkin, M., Boone, C. W., and Kelloff, G. J., Eds.) pp227-245, CRC Press, Boca Raton, FL.
(19) Barcelo, S., Gardiner, J. M., Gescher, A., and Chipman, J. K.(1996) CYP 2E1-mediated mechanism of anti-genotoxicity of thebroccoli constituent sulforaphane. Carcinogenesis 17, 277-282.
(20) Jiao, D., Conaway, C. C., Wang, M. H., Yang, C. S., Koehl, W.,and Chung, F. L. (1996) Inhibition of N-nitrosodimethylaminedemethylase in rat and human liver microsomes by isothiocyan-ates and their glutathione L-cysteine, and N-acetyl-L-cysteineconjugates. Chem. Res. Toxicol. 9, 932-938.
(21) Conaway, C. C., Jiao, D., and Chung, F. L. (1996) Inhibition ofthe rat liver cytochrome P450 isozymes by isothiocyanates andtheir conjugates: a structure-activity relationship study. Car-cinogenesis 17, 2423-2427.
(22) Zhang, Y., and Talalay, P. (1994) Anticarcinogenic activities oforganic isothiocyanates: chemistry and mechanisms. Cancer Res.54, 76-81.
(23) Hecht, S. S. (1995) Chemoprevention by isothiocyanates. J. Cell.Biochem. 22, 195-209.
(24) Leonard, T. B., Popp, J. A., Graichen, M. E., and Dent, J. G. (1981)R-Naphthylisothiocyanate induced alterations in hepatic drug
metabolizing enzymes and liver morphology: implications con-cerning anticarcinogenesis. Carcinogenesis 2, 473-482.
(25) Yang, C. S., Smith, T. J., and Hong, J. Y. (1994) Cytochrome P450enzymes as targets for the chemoprevention against chemicalcarcinogenesis and toxicity: opportunities and limitations. CancerRes. 54, 1982-1986.
(26) Lee, M.-S (1996) Enzyme induction and comparative oxidativedesulfuration of isothiocyanates to isocyanates. Chem. Res. Toxi-col. 9, 1072-1078.
(27) Guo, Z., Smith, T. J., Wang, E., Eklind, K. I., Chung, F.-L., andYang, C. S. (1993) Structure-activity relationships of arylalkylisothiocyanates for the inhibition of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone metabolism and the modulation of xenobiotic-metabolizing enzymes in rats and mice. Carcinogenesis 14, 1167-1173.
(28) Larson, J. R., Coon, M. J., and Porter, T. D. (1991) Alcohol-inducible cytochrome P450 IIE1 lacking the hydrophobic NH2-terminal segment retains catalytic activity and is membrane-bound when expressed in Escherichia coli. J. Biol. Chem. 266,7321-7324.
(29) Shen, A. L., Porter, T. D., Wilson, T. E., and Kasper, C. B. (1989)Structural analysis of the FMN binding domain of NADPH-cytochrome P450 oxidoreductase by site-directed mutagenesis. J.Biol. Chem. 264, 7584-7589.
(30) Hanna, I. H., Teiber, J. F., Kokones, K. L., and Hollenberg, P. F.(1998) Role of alanine at position 363 of cytochrome P450 2B2 ininfluencing the NADPH- and hydroperoxide-supported activities.Arch. Biochem. Biophys. 350, 324-332.
(31) Kent, U. M., Roberts, E. S., Chun, J., Hodge, K., Juncaj, J., andHollenberg, P. F. (1998) Inactivation of cytochrome P450 2E1 bytert-butyl isothiocyanate. Chem. Res. Toxicol. 11, 1154-1161.
(32) Buters, J. T. M., Schiller, C. D., and Chou, R. C. (1993) A highlysensitive tool for the assay of cytochrome P450 enzyme activityin rat, dog and man. Biochem. Pharmacol. 46, 1577-1584.
(33) Omura, T., and Sato, R. (1964) The carbon monoxide-bindingpigment of liver microsomes. I. Evidence for its hemo proteinnature. J. Biol. Chem. 239, 2370-2378.
(34) Silverman, R. B. (1996) Mechanism-based enzyme inactivators.In Contemporary Enzyme Kinetics and Mechanisms (Purich, D.L., Ed.) pp 291-335, Academic Press, San Diego, CA.
(35) Mishin, V. M., Koivisto, T., and Lieber, C. S. (1996) The deter-mination of cytochrome P450 2E1-dependent p-nitrophenol hy-droxylation by high-performance liquid chromatography withelectrochemical detection. Anal. Biochem. 233, 212-215.
(36) Estabrook, R. W., and Werringloer, J. (1978) The measurementof difference spectra: application to the cytochromes of mi-crosomes. Methods Enzymol. 52, 212-220.
(37) Sharma, U., Roberts, E. S., and Hollenberg, P. F. (1996) Inactiva-tion of cytochrome P450 2B1 by the monoamine oxidase inhibitorsR-(-)-deprenyl and clorgyline. Drug Metab. Dispos. 24, 669-675.
(38) Koop, D. R. (1990) Inhibition of ethanol-inducible cytochrome P4502E1 by 3-amino-1,2,4-triazole. Chem. Res. Toxicol. 3, 377-383.
(39) Bell, L. C., and Guengerich, F. P. (1997) Oxidation kinetics ofethanol by human cytochrome P450 2E1. J. Biol. Chem. 272,29643-29651.
(40) Patten, C. J., Ishizaki, H., Aoyama, T., Lee, M., Ning, S. M.,Huang, W., Gonzalez, F. J., and Yang, C. S. (1992) Catalyticproperties of the human cytochrome P450 2E1 produced by cDNAexpression in mammalian cells. Arch. Bioch. Biophys. 299, 163-171.
(41) Yamazaki, H., Inui, Y., Yun, C. H., Guengerich, F. P., andShimada, T. (1992) Cytochrome P450 2E1 and 2A6 enzymes asmajor catalysts for metabolic activation of N-nitrosodialkylaminesand tobacco-related nitrosamines in human liver microsomes.Carcinogenesis 13, 1789-1794.
(42) He, K., Iyer, K. R., Hayes, R. N., Sinz, M. W., Woolf, T. F., andHollenberg, P. F. (1998) Inactivation of cytochrome P450 3A4 bybergamottin, a component of grapefruit juice. Chem. Res. Toxicol.11, 252-259.
(43) Ishizaki, H., Brady, J. F., Ming, S. M., and Yang, C. S. (1990)Effect of phenethyl isothiocyanate on microsomal N-nitrosodi-methyl-amine metabolism and other monooxygenase activities.Xenobiotica 20, 255-264.
TX9900019
Inactivation of P450 2E1 by Benzyl Isothiocyanate Chem. Res. Toxicol., Vol. 12, No. 7, 1999 587







![nanodiagnostics.ppt [Schreibgeschützt] [Kompatibilitätsmodus] · Sr + Benzyl Alcohol + Ti(OiPr) 4 SrTiO 3 Li + Benzyl Alcohol + Nb(OEt) 5 LiNbO 3 Ba,Sr + Benzyl Alcohol + Ti(OiPr)](https://img.pdfslide.us/doc/110x75/6059c0a096e778411b20081e/schreibgeschtzt-kompatibilittsmodus-sr-benzyl-alcohol-tioipr-4-srtio.jpg)





















