Embed Size (px)
Citation preview

IN VIVO AND IN VITRO TRANSFORMATIONS OF MOUSE TISSUES
FROM A MURINE LYMPHOSARCOMA
DISSERTATION
Presented to the Graduate Council of the
North Texas State University in Partial
Fulfillment of the Requirements
For the Degree of
DOCTOR OF PHILOSOPHY
By
James E. Carnes, M.A.
Denton, Texas
August, 1972

Carnes, James E., In Vivo and In Vitro Transformations
of Mouse Tissues from a Murine Lymphosarcoma. Doctor of
Philosophy (Biology), August, 1972, 79 pp., 6 tables,
14 figures, bibliography, 101 titles.
The problem with which this investigation is concerned
is that of determining the nature of events leading to the
change. of normal cells into malignant cells. The design
of the study is multi-phasic: (A) to establish the presence
or absence of an oncogenic virion, (B) to demonstrate by
use of the electron microscopy any ultracellular alteration
in malignant or transformed tissues, (C) to investigate
the nature of the transforming agent in the murine lympho-
sarcoma, and (D) to employ various methods to demonstrate
cellular transformations in vivo and in vitro.
The tumor line was produced by subdermal transplants
of a 20-methylcholanthrene-induced lymphosarcoma tumor
into isologous DBA/IJ mice. The tumor has been maintained
through more than four hundred passages. The mean survival
time for the tumor-implanted mice was approximately eleven
days.
Tumor, liver, spleen, and kidney tissues from the
tumor-bearing mice were prepared for electron microscopy,

thin-sectioned, and observed for ultracellular alterations
and for virus particles. Sections of the tissues were also
prepared for light microscopy.
Single-cell suspensions of tumor, tumor liver, and
tumor spleen were prepared. The cell suspensions were used
in tissue culture preparations, diffusion chamber, and
parabiotic chamber experiments for cellular transformations.
Nuclei and mitochondria were isolated from the cell
suspensions and implanted into DBA/lJ mice.
The tissues observed by the electron microscope
showed the mitochondria to be swollen and structurally
deficient: fewer irregular or parallel tubular cristae,
myelin-like figures, and pleomorphic. Some of the mito-
chondria had inclusions of dense and irregular bodies.
The nucleus of the tumor tissue repeatedly had
invaginations of the nuclear membrane into the nucleoplasm,
and in the older tumor tissue the nuclear membrane had
become separated. In the necrotic area of the tumor,
the nucleus was last to undergo alterations. The nucleolus
in the tumor became enlarged as the tumor progressed.
In the tumor cells there was a noticeable lack of
ergastoplasmic lamellae in the cytoplasm. Free ribosomes

and polysomal formations were abundant. The swollen
cristae were disrupted into various sized vesicles.
The tumor could be massaged into the isologous DBA/lJ
mice by subdermal implantation of tumor liver, tumor spleen,
tumor kidney, and single-cell suspension from each of these
tissues. The single-cell preparations, when placed in
diffusion chambers with twenty-five millimicron filters
and implanted subdermally into mice, induced the formation
of a lymphosarcoma. In parabiotic chambers, with normal
cells in one chamber and tumor cells in the other chamber,
separated by a filter, the normal cells became transformed
and induced a lymphosarcoma in mice after implant.
When the isolated nuclei from tumor tissue were implanted
into mice, a tumor developed. Treatment of the nuclei with
nucleases and then implantation in mice cause a tumor only
when the nuclei were treated with deoxyribonuclease.
It is concluded that the transforming and tumor-
inducing agent in this' investigation was not a virion,
but an infectious ribonucleic acid genome or a segment of
a viral genome which had become integrated into the
genome of the mouse cells. The vision has lost its
ability to form a protein coat; therefore it is not

demonstrable as a virion. But the ribonucleic acid is
able to infect other cells and transform them from normal
to neoplastic tissues.

TABLE OF CONTENTS
PageLIST OF TABLES . . . ...........iv
LIST OF ILLUSTRATIONS. . . . . . . . . . . . . . . . v
Chapter
I. INTRODUCTION . . . . . . . . . . . . . . . ..
Viral CarcinogenesisTransformation by VirusesTumor Cell Fine Structure
II. MATERIALS AND METHODS. . . . . . . . . . . . . 17
Electron MicroscopyTissue TransplantsCell Suspension PreparationsPreparation of Cell-Free ExtractPreparation of MitochondriaPreparation of NucleiChamber ImplantsMaintenance of Tumor Cells in Tissue CultureParabiotic Chamber
III. RESULTS AND DISCUSSION....-..-..............30
IV. SUMMARY. .....-.............. . 68
LITERATURE CITED..-........-............-...........69
iii

LIST OF TABLES
TablePage
I. Incidence of Lymphosarcoma Tumor ResultingTransplants of Tumor in DBA/IJ Mice
.II. Incidence of Lymphosarcomas Resulting FromTransplant of Liver Tissue From MiceReceiving Lymphosarcoma Implants. -
III. Incidence of Lymphosarcoma Resulting FromTransplant of Spleen Tissue from MiceReceiving Lymphosarcoma Implants. .
IV. Incidence of Lymphosarcoma Resulting FromTransplant of Kidney Tissue From MiceReceiving Lymphosarcoma Implants.. -
from0 . ." 46
- . . 48
" . . 52
. 56
V. Tumors Produced by Cell Suspension in DBA/IJMice from a Trypsinized Tumor . - . .
IV. Development of Tumor in DBA/IJ Mice fromDiffusion Chambers ContainingLymphosarcoma Cell Suspensions............61
iv
. ."
. . 57

LIST OF ILLUSTRATIONS
Figure Page
1. Photographs of DBA/IJ mice bearing lymphosarcomatransplant.-...... ... . . . . . . . . . 31
2. Photographs of DBA/lJ mice bearing lymphosarcomatransplant.........-.-......... . 32
3. An electron micrograph of tumor tissue fourdays after implant . . . . . . . . . . . . . . 35
4. An electron micorgraph of tumor tissue six daysafter implant.................. .36
5. An electron micrograph of tumor tissue eight daysafter implant-......... . . . . . . . . . 37
6. An electron micrograph of tumor tissue ten daysafter implant..-.....-..- . ........ .38
7. An electron micrograph of tumor tissue twelvedays after implant................ . 39
8. An electron micrograph of the central necroticarea of tumor tissue.-....... ..... . 40
9. An electron micrograph of a tumor cell from asingle cell suspension . . . . . . . . . . 44
10. An electron micrograph of a tumor cell grown intissue culture...-.............. .45
11. An electron micrograph of tumor liver tissueshowing presence of tumor cell . . . . ..... 49
12. An electron micrograph of tumor cell packet intumor liver tissue ..-....-.-..-.. . . . . 50
v

Figure
Page13 An electron micrograph of tumor cell packetin t mor plee tis ue. - . . . . . - . . - . 5314. An electron micrograph of a Sternberg-Reedcell found in tumor spleen tissue ,. -. 55
vi

CHAPTER I
INTRODUCTION
This research, based on experiments with a murine
lymphosarcoma, examines the nature of events leading to
the change of normal cells into malignant cells. In the
mouse, a system has been developed in which the malignant
changes occur rapidly and fairly synchronously in both
a homogeneous and a heterogeneous population of cells.
Through this system, inquiry, employing both in vivo and
in vitro experiments, has revealed that the presence of
a virion is not necessary for cellular transformation.
Further, ultrastructure of the tumor and fine structural
alterations in the transformed tissue have been studied,
and the results of the studies are reported.
Although the etiology of cancer has spawned numerous
hypotheses, only those which have received further support
from new studies have escaped obscurity. In the recent
past, two main theories have guided investigators in
their efforts to discover the key to the puzzling nature
1

2
of tumors: (A) the multi-etiological theory that cancer
can be caused by many extremely varied factors such as
chemical substances (23), physical factors (101), and
biological agents (72); and (B) the virus theory (38, 42,
72, 94).
This study explores the virus theory; and the fact
that so much work has been done with the theory suggests
its importance. The following discussion notes significant
background information relevant to viral carcinogenesis
and viral transformation.
Viral Carcinogenesis
The concept of the possible viral origin of tumors
was first stated by Borrel (14). The first experimental
evidence of the capacity of viruses to induce neoplastic
processes was presented by Ellermann and Bang (29), who
demonstrated the viral nature of fowl leukosis, and by
Rouss (77), who discovered the virus capable of producing
a chicken sarcoma.
Further development of the viral concept was slow.
The most important contributions were the discovery of
the Shope rabbit papilloma virus (85) and the Bittner

3
virus (11). Shope was the first to demonstrate the
masked viral genome.
Interest in the role of viruses in the etiology of
cancer has risen sharply, and many investigations have
been devoted to the study of the problem (24, 37, 70, 85).
Most investigators studying the tumor viruses considered
that the action of the virion was similar to viruses in-
ducing clinical infectious diseases, and that the
acceleration of cell proliferation, as found in the
neoplastic processes, was caused by the continuous
presence of the virus in the affected tissue (71).
Zilbur (100) proposed the theory that the virus does
not act as an infectious agent, but as a transforming one.
The neoplastic process initiated by viruses comprised two
phases, with the virus playing quite a different role in
each of them. The first phase was a hereditary transformation
of normal cells to tumor cells. The second phase was the
reproduction of tumor cells, in which the virus did not
play an important role, and this phase led to the clinical-
ly apparent disease.
Fraenkel-Conrat (33) and Gierer and Schramm (35)
published almost simultaneously experimental data showing

4
that ribonucleic acid (RNA) of tobacco mosaic virus could
cause disease in a certain percentage of trials. This was
the first report of an infectious nucleic acid. Numerous
experiments have shown that RNA of varying purity, obtained
from the virus of Western equine encephalitis, poliomyelitis,
West Nile encephalitis, West tick-borne encephalitis,
stomatitis aphthosea, and mouse encephalitis can cause the
corresponding diseases (4, 15, 16, 34, 47, 98). In these
experiments, the extracted nucleic acids did not cause the
infection, but induced the synthesis of virions within
the cell, and that caused the pathological condition.
Hays, et al. (42) reported that the inoculation of salt
suspensions of deoxyribonucleic acid (DNA) and RNA, from
leukemic and non-leukemic organs of mice, into various
strains of mice caused leukemia to develop. Latarjet, et al.
(54) reported that it is possible to induce leukemia in AKR
and C3H mice by means of nucleic acids (DNA and RNA) obtained
from leukemic tissues of AKR mice. Lacour, et al. (50)
observed similar results with RNA from Ehrlich's ascites
tumor. Bielka and Graffi (10) prepared RNA from leukemia
myeloid tissue and induced leukemia in mice. But efforts

5
to induce tumor with RNA from filtrates of Rous sarcoma
proved negative (49).
Di Mayorca,et al. (22) reported that DNA obtained
from mouse embryonic tissue infected with polyoma virus
had infective properties. Neither transformation nor
cytopathic effects were observed if deoxyribonuclease had
been added.
Graffi and Fritz (39) prepared DNA and RNA from
polyoma tumors and showed a cytopathic effect on cultures
of embryonic mouse tissue, and the medium from the culture
induced multiple tumors in new born animals. Treatment of
the samples with ribonuclease did not eliminate the
activity of the RNA preparation.
Dmochowski,et al. (25) found that cell-free extracts
of tissue culture, inoculated with polyoma virus and phenol
treated for deproteinization, caused parotid gland tumors
in mice and subcutaneous sarcoma in hamsters. The
nucleic acid preparation also caused cytopathic lesions
in embryonic mouse tissue, and the material from the latter
was carcinogenic.

6
Transformation by Viruses
The transforming capacity of a number of tumor viruses
has been studied (93). Changes in the morphology of cells, in
rate of cell metabolism and generation time, in antigenic
composition, or in karyotype have been most often used as in-
dices of transformation. Some of these changes may be accom-
panied by acquisition of neoplastic potential. It is these
cells that in recent years have undergone intensive scrutiny,
in efforts to study the mechanism underlying the conversion
of non-oncogenic cells to those with oncogenic properties.
Some of the viruses found to illustrate a transforming
capacity are SV40, Adenovirus, polyoma, and Rous sarcoma. Eddy,
et al. (28), and Girardi, et al. (37) were first to report that
SV40 could induce tumors when inoculated into newborn hamsters.
These studies, as well as those reported later (6, 12), revealed
the neoplastic lines to be fibrosarcomas from which viruses
could be recovered only occasionally, and then only in small
quantities. Subcutaneous inoculation of newborn hamsters with
phenol-extracted deoxyribonucleic acid from SV40 virus has
produced tumors (27), although the latent period was longer
than that usually observed after inoculation of the complete
virus. Treatment of the extract with DNAase rendered it non-
oncogenic.

7
Hamster cells transformed by SV40 in vivo or' in vitro
were oncogenic when transplanted into young hamsters (74).
Fewer than 100 cells were found necessary to produce tumors
in 50 per cent of the animals. Simian cells have been
transformed by SV40 (6, 31). The transformed cultures
eventually stopped producing infectious viruses, but
continued synthesizing an intranuclear tumor antigen,which
indicated viral genome presence. Diderholm,' et' al. (21)
reported on the transformation of rat and guinea pig
kidney cells with SV40. Transformation was accompanied by
appearance of predominantly epithelioid cells and the
synthesis of SV40 tumor antigen within the cells. Intro-
duction of larger numbers of the transformed cells (103
cells) into autologous, X-irradiated rats resulted in tumors,
but untreated animals did not produce tumors.
Trentin, et al. (96) first reported that adenoviruses
have oncogenic properties. They were concerned with
pulmonary tumors induced in newborn hamsters following
inoculation of adenovirus type 12. Trentin's results were
confirmed by Huebner, et al. (46) and later by McLeod and Ham
(61), who further demonstrated that adenovirus type 18 was

8
oncogenic for newborn hamsters. Although a large number of
viruses were subsequently examined, only adenoviruses type
7 (36, 51), type 31 (74), and type 3 (44) were found to be
oncogenic when inoculated into newborn hamsters.
McBride and Wiener (60) reported the only successful
transformation of cells in vitro by adenoviruses. The cells
were derived from newborn hamster kidneys. As with the H-50
line of SV40-transformed cells (74), infectious virus
could not be recovered, but cells did contain adenovirus-
related antigens. Their tumor potential was not reported.
Latarjet (52) postulated that only a fraction of a
viral genome was necessary for transformation and that
irradiation of the virus would separate the tumorigenic
property from the infective property, the former being
more radiation-resistant than the latter. His theory has
been proved correct for the polyoma virus (7, 8), for which
approximately 50 per cent of the viral genome is necessary
to transform cells of the young hamster kidney cell line
(BHK 21).
Experiments with polyoma virus added to cultures of
hamster and mouse embryo tissue showed that the transformation

9
of normal cells to tumor cells proceeds very quickly, and
the formation of foci of transformed cells can be observed
for several weeks. The cells transformed in vitro are
identical to tumor cells obtained when animals are infected
with polyoma virus (79, 97). The transformed culture cells
change morphology and form foci of multilayered growth. If
culture cells are implanted into the corresponding animals,
a tumor is induced.
Carcinogenesis caused by the Rous sarcoma virus has
been achieved by a number of investigators (40, 87, 90,
98). Temin (92), while working with Rous sarcoma virus,
concluded that the production of the virus is not essential
for carcinogenesis, and that conversion and carcinogenesis
are not conditioned by the viral genome.
The neoplastic transformation of cells by Rous sarcoma
virus was successfully reproduced in rat fibroblast culture.
Rats, after having been inoculated with the transformed
culture, developed tumors. These tumors were then trans-
planted into chicks by the cells, but not by the filtrates
(87).
As a result of observations based on the widespread
occurrence of RNA tumor viruses, two hypotheses have been

10
proposed: (A) the oncogene hypothesis (47) and (B) the
protovirus hypothesis (94). The oncogene hypothesis states
that the genetic composition of all vertebrates contains
the DNA provirus for an RNA tumor virus. The provirus,
in development or in carcinogenesis, is activated to make
virus-specific products and virions. The provirus hypothesis
states that in the germ-line of chordates there are con-
tained regions of DNA which can evolve in various directions
through DNA to RNA to DNA transfers in the somatic cells.
This display of evolution is normally a part of embryonic
differentiation, but an abnormal evolution might lead to
the formation of the RNA tumor virus genome.
Tumor Cell Fine Structure
The fine structure of tumor cells remains as variable
as that found in their normal homologues, and no specific
or universal pattern of cancer has been found in electron
microscope investigations (9). In routine diagnoses of
tumors, histologidal, rather than cytological or ultra-
structural, criteria prevail. The cancer cell should be
referred to in the plural because of its polymorphism.
In spite of the prodigious polymorphism, the characteristic

11
lesions encountered in tumor cells have been observed and
interpreted.
The interphase nucleus of a tumor cell usually
exhibited an enlarged and irregular shape. The nucleo-
cytoplasmic ratio was increased, and the increase is
attributed to the ploidy of the chromosome set, frequently
encountered in tumor cells (18). The enlargement was
caused by swelling under unphysiological conditions
(nuclear edema) (9) .
The nuclear membrane, inasmuch as fine structure is
concerned, did not indicate any difference between normal
and malignant cells (18). The only unusual feature
repeatedly found in the membrane was reported by Bernhard
(9) as deep invaginations of the membrane into the nucleo-
plasm. These invaginations were frequently in contact with
the nucleolus. The other malformation of the nuclear
membrane led to infolding or to cytoplasmic projections
called "nuclear blebs" (3).
With the exception of the nucleolus, the nucleoplasm
has been defined as the nuclear contents, including hetero-
chromatic, euchromatin, and the interchromatic matrix

12
of the interphase nucleus. An increase or an irregular
distribution of heterochromatin has been observed in many
tumor cells (8). Margination of the chromatin has been
shown to be accentuated and very pronounced in the nuclei
of dying cells (18). In addition, perichromatin granules
have been observed in' increased numbers in the nucleoplasm
of many tumors (67).
The increased size of the cancer cell nucleolus has
been a rule with rare exceptions in classical pathology.
The fine structural variations have been reported as
numerous and extreme: dense, compact granules in nucleolar
bodies, or a nucleolonema fibrillar structure without
granules. Vacuoles, lipid inclusions, or inclusions of
unknown composition have been shown in the nucleolus.
Some of the altered nucleoli in tumor cells resembled
those observed after protein deficiency in the rat liver
nucleolus (9).
The mitochondrial laterations have been shown to be
the most recurrent and striking anomalies found in the
fine structural changes within tumor cells (9). But it
has been reported that many tumors do have mitochondria

13
with number, shape, an.d ultrastructure that cannot be
distinguished from mitochondria found in homologous
normal cells (44). More often, the tumor cell has swollen
and structurally deficient mitochondria: fewer irregular
or parallel tubular cristae (55), concentric cristae (81),
altered matrix, myelin-like figures, pleomorphic mitochondrial
body shape or cup-shaped mitochondria (88). Tumor cell
mitochondria at times exhibit various kinds of inclusions:
glycogen (89), -protein crystals (44), and dense, irregular
bodies (88).
Cancer cells grow rapidly,and therefore need to
produce greater quantities of proteins in comparison with
embryonic cells, but a constant and specific feature for the
ultrastructural substrate of protein synthesis has not been
detected. The nucleolar apparatus, producing ribosomal
precursors, displays hypertrophy, and polysomal formations
have been seen more often than in normal homologues.
Dalton (17) has shown that in benign adenoma, and in cancer
cells, the cytoplasm may be packed with functional ergasto-
plasm. The same condition may also appear in milk-producing
mouse mammary tumor, thyroid, and myeloma cells (17, 59).

14
Cell lines of mesenchymal tumors that have been
maintained during many passages in vitro may have a large
amount of ergastoplasmic lamellae; for example, Rous sarcoma
cells and methylcholanthrene-induced tumor cell lines.
Adenovirus 12-induced tumor, cultivated in vitro, has a
decreased ergastoplasm, but many free ribosomes in the
cytoplasm. The viral genome may influence the architecture
of the tumor cell. For instance, the RAV virus maintained
many ergastoplasmic lamellae, and the Rous virus induced
partial disorganization (9).
The Golgi apparatus appears well developed in many
tumors: exocrine organ tumors, mouse mammary tumors in which
virus particles may appear, spontaneous or experimentally
induced hepatomas, and Rous sarcoma. The multivesicular
bodies in association with the Golgi apparatus have been
found extremely hypertrophic in reticulosarcoma cells and
myelomas (9, 69).
Abercrombie and Ambrose (1) have shown that certain
strains of tumor cells display a loss of contact inhibition
in tissue culture. They indicated that a generalized
diminution of adhesiveness might be an important point in

15
malignant transformations. They also observed an increase
in the negative charge in some tumor cell strains, and linked
it with mucopolysaccharide production at the cell surface.
Defendi and Gasic (19) have shown that hamster fibroblast
transformed in vitro with polyoma virus produced considerable
sialic acid. This observation has been confirmed by
histochemistry and electron microscopy.
Oberling and Bernhard (70) have demonstrated that
anaplasia is characterized by a low degree of differentiation
in rapidly growing tumors. Malignant tissues gradually
lost the architecture of the normal homologous tissue and
their growth form tended toward the chaotic. Ultrastruc-
turally, the cytoplasm was much less organized than usual.
The mitochondria number varied, but had the tendency to
decrease. Ergastoplasmic lamellae were rare or completely
absent, as they were in the Golgi apparatus. The cytoplasm
had a hydropic appearance similar to the cellular organi-
zation found in tumor regression. The nucleus had strong
pleomorphic outlines.
Bernhard (9) posed a question: Do rapidly dividing
cells have the time to build a complete cellular organization

16
which would enable the tumor cell to function normally?
Even normal tissues', when ' grown in tissue 'culture and
induced to rapid cell divisions, have exhibited anaplasia
(dedifferentiation). Slowly growing tumors have been shown
to be histologically complex, but cytologically organized.
Thus the growth rate alone does not explain the total
phenomenon of anaplasia.
The design of this study was multi-phasic: (A) to
establish the presence or absence of an oncogenic virion,
(B) to demonstrate by use of the electron microscope any
ultracellular alterations in malignant or transformed
tissues, (C) to investigate the nature of the transforming
agent in the murine lymphosarcoma, and (D) to employ various
methods to demonstrate cellular transformations in vivo
and in vitro. The information reported in this investigation
gives an insight into the nature of the causative transforming
"factor" found to be present in the murine lymphosarcoma
used in the study.

CHAPTER II
MATERIALS AND METHODS
DBA/IJ pure-line male mice , four to six weeks old,
obtained from Jackson Memorial Laboratory, Bar Harbor,
Maine, were used to maintain the transplantable urine
lymphosarcoma tumor used in this study. The original
tumor was induced in the mice by painting on the dorsal
unepilated interscapular skin a solution containing 0.6
per cent 20-methylcholanthrene in reagent grade benzene.
The solution was applied with a number four camel's
hair brush five times a week for nine weeks. After one
week, epilation occurred at the painted site. By the
sixth week, papillomas appeared. During the ninth week
after the primary painting, squamous-cell carcinomas
appeared (82).
The tumor line was produced by subdermally trans-
planting into an isologous DBA/lJ mouse a lymphoid tumor
which emerged in the 20-methylcholanthrene-painted mice.
The tumor was maintained through one hundred ninety-two
17

18
passages by subdermal transplantation with a 12-gauge
trocar needle of a section of tumor (2 to 3 mm in
diameter) obtained from a tumor-bearing mouse one or two
days before death. The mean survival time for the tumor-
implanted mouse was approximately eleven days.
Electron Microscopy
Immediately after the mice were sacrificed, tissues
were taken from the animals and fixed in 4 per cent
paraformaldehyde (Eastman Chemical Company) in 0.2 M
s-collidine (Sigma Chemical Company) buffer (pH 7.2),and
placed in a refrigerator for 2-24 hr (78). Some of the
tissues were fixed in 3 per cent glutaraldehyde (J.T.
Baker Chemical Company) in 0.1 M cacodylate buffer (Fisher
Scientific Company), pH 7.3, and placed in the refrigerator
for 2-24 hr (78). Following the aldehyde fixation, the
tissues were rinsed two times (5-10 min each) in 0.2 M
s-collidine or 0.1 M cacodylate buffer (pH 7.2). The
specimens were post-fixed for from one to two hours in 1.5
per cent osmium tetroxide (Fisher Scientific Company),
in 0.2 M s-collidine or 0.1 M cacodylate buffer (pH 7.2),
at 4-10 C. After the osmium post-fixation, the specimens

19
were rinsed two times (5 min each) in 0.2 M s-collidine
or 0.1 cacodylate buffer (pH 7.2), and then dehydrated in
ethanol or acetone (30, 50, 75, 95, and 100 per cent), and
then embedded in Epon 812 (56).
Thin sections were prepared with a Porter-Blum
MT-2 Ultramicrotome equipped with a diamond knife. The
thickness of the sections was determined by interference
colors (silver-gray to straw) to be 60 to 80 mp. The
thin sections were collected on 200-mesh copper grids and
stained with saturated uranyl acetate in 50 per cent
ethanol for 5-10 min, and counterstained in lead citrate
(63) for 3-5 min. The grids were coated with a thin layer
of carbon in a Mikros Vacuum Evaporator for section stabili-
zation in the electron beam. The specimens were examined
in an RCA 3-G Electron Microscope fitted with a 45-micron
objective aperture and operated at 50 Kv.
Thick sections were obtained from the same specimen
blocks from which the thin sections were taken. The
thick sections, approximately one micron thick, were
removed from the water surface of the diamond knife by
a single-bristle brush, and placed in a single drop

of water on a microscope slide. The water was evaporated
by placing the slide on a hot plate 'at approximately 200 C.
The sections were stained with Toluidine Blue 0 (Fisher
Scientific Company) for 30 sec or with Paragon Stain
(Paragon C and C Company) for 10 sec, rinsed in distilled
water, dried, cover-slipped, and examined with a light
microscope.
In each of the electron microscope specimen prepara-
tions, control tissues were processed simultaneously.
This measure was taken to prevent misinterpretations of
cellular abnormalities caused by improper fixation or
embedding. - Thus, any alteration exhibited in the tumor
tissues was not an artifact, but was induced as a result
of malignancy.
Tissue Transplants
A group of mice were implanted with tumor tissue,
and on days two, four, six, eight, and ten after implant
the mice were sacrificed by cervical dislocation. Liver,
spleen, and kidney tissues (2 to 3 mm in diameter) were
taken and transplanted subdermally into the axillary
region of isologous DBA/lJ mice. The animals were
20

21
observed daily for tumor progression. Tumor take and day
were determined by an increase in size of the transplant
tissue nodule.
Cell Suspension Preparations
Tumor tissue was taken aseptically from sacrificed
animals from eight to ten days after tumor transplantation,
and placed in a trypsinizing flask with 20 ml of the 0.25 per
cent trypsin solution in calcium- and magnesium-free phosphate-
buffered saline (CMF-PBS). A magnet stirring bar was used
for gentle agitation of the solution until the tissue
dispersed into single cells. The cell suspension was
centrifuged for 5 minutes at 1000 rpm to form a cell
pellet. The cells were washed two times in 10 ml of
CMF-PBS. Viable cell number was determined by mixing 1.0 ml
of cell suspension with 0.5 ml of a 0.5 per cent water
solution of trypan blue, and the counting was done on a
haemocytometer. After the viable cell count was made, the
cell suspension was diluted to the desired cell number with
CMF-PBS.
Tumor cell suspensions were also prepared aseptically
by mincing the tissue over cracked ice with fine scissors.

22
using three changes of cold sterile CMF-PBS, totalling
approximately 10 ml. The tissue suspension was aspirated
into a 25-ml glass syringe and then inverted in ice for
10 to 20 min to allow larger tissue particles to settle out.
A Swinney hypodermic adapter was attached to the syringe
and the tissue suspension was forced through a wire mesh
support screen. Viable cell counts were determined. The
number of viable cells per cubic millimeter of suspension
was ascertained and multiplied by 1000 to obtain number
of viable cells per milliliter. Appropriate dilutions
were made with CMF-PBS to obtain the desirable number of
cells per milliliter for transplantation.
Preparation of Cell-Free Extract
Cell-free extract was obtained by placing minced
tumor tissue in a trypsinizing flask with a magnetic
stirring bar, and agitating at a moderate speed for 30
min. The suspension was centrifuged (Sorvall RC2B
Centrifuge) at 3000 X g for 10 min, and the supernatant
was recentrifuged at 8000 X g for 30 min. The supernatant
was recentrifuged (Beckman Model L Ultracentrifuge)
at 30,000 X g for one hr, using a Ti 50 rotor. One-ml

23
samples of the supernantants were injected subdermally
into DBA/IJ mice, which were observed periodically for
tumor progression. The supernatants were incubated for
one hour at 37 C with 100 mg/ml pancreatic deoxyribo-
nuclease (DNAase) or 100 mg/ml ribonuclease (RNAase).
The enzymes were removed by phenolic extraction, and the
solutions were injected subdermally into mice which
were observed for tumor progressions.
Preparation of Mitochondria
Mitochondria were isolated from tumor tissue by a
modification of the procedure of Schneider and Hogeboom
(80). The isolation medium (STEA) consisted of 0.25 M
sucrose, 1.0' mM tris-HCL, 1.0 mM ethylenediaminetetraacetate,
and 1.0 per cent bovine serum albumin.
The mice were sacrificed by cervical dislocation, and
the excised tumors were placed in a beaker of cold
STEA medium. The tissue was minced with sharp scissors,
and the STEA medium was decanted, and the minced tissue was
transferred to a tissue homogenizer. Approximately 10.ml
of STEA medium per gram of original tumor were added to
the vessel. The. tissue was homogenized in the cold.

24
throughout the procedure. The homogenate was transferred
to centrifuge tubes and centrifuged at 700 X g for 10 min
in a refrigerated centrifuge to remove cell debris. After
centrifugation, the supernatant was decanted through
cheesecloth into clean centrifuge tubes, and the cell
debris was discarded. The suspension was centrifuged
at 900 X g for 10 min. The supernatant was decanted
and discarded, and the mitochondrial pellet was resuspended
in approximately half the volume of STEA used in homog-
enization. The suspension was centrifuged two times at
9000 X g for 10 min to remove microsomal and other
cellular contamination. All operations in the procedure
were carried out in the cold (1-4 C). The final pellet
was resuspended in 0.5 ml STEA per gram of original tumor
tissue. The mice were inoculated subdermally with 1.0 ml
of the mitochondrial preparation. The control mice were
injected subdermally with 1.0 ml STEA medium. The
suspension was checked for purity by electron microscopy.
Preparation of Nuclei
Tumor tissue or liver from tumor-bearing mice
(tumor liver) was removed from DBA/lJ mice, weighed,
and immediately placed in a cold medium (SM) of 0.25

25
M sucrose and 3.3 mM mgCl2 . The tissue was minced
with sharp scissors, the SM medium was decanted, and
the minced tissue was transferred to a tissue homog-
enizer (Potter-Elvehjem with teflon pestle). Approximately
5 ml of SM medium were added per gram wet weight of
tissue. The tissue was homogenized and filtered through
two layers of gauze, and the filtrate was centrifuged at
1000 X g for 10 min. The crude nuclear sediment was
suspended in approximately 7.5 ml of 2.3 M sucrose and 3.3
mM MgCl2 per gram of original tissue and centrifuged at
40,000 X g for one hr. The supernatant was decanted, and
the nuclear sediment was suspended in 2 ml of SM medium
per gram of original tissue. The suspension was
centrifuged at 1000 X g for 5 min to remove contaminating
erythrocytes. The pellet of nuclei was suspended in SM
medium at a concentration of 35 X 107 nuclei per ml.
All procedures were performed in the cold (1-4C) under
aseptic conditions. The nuclear preparation was checked
for purity by electron microscopy. Mice were injected
subdermally, by means of a 22-gauge hypodermic needle,
with 0.5 ml of the nuclei suspension. The control mice
were injected by the same method with 0.5 ml of SM medium.

26
Some control animals were injected with. normal nuclei.
treated with the. nuclease as in the test 'samples.
Chamber Implants
Diffusion chambers were constructed by a modification
of Algire's method (5), by using Millipore lucite rings
and 25 mp Millipore filters (Millipore Filter Corporation).
The Millipore filters were glued to the lucite rings
with Millipore chamber adhesive. The chambers were
sterilized under ultraviolet light for 36 hr and were
turned over approximately every 12 hr. The chambers
were filled with a tumor cell suspension of 2 x 106 cells/
ml in CMF-PBS.' The cells were put into the chambers
through a hole in the lucite ring by means of a 25-gauge
needle fitted to a 2-ml syringe, and the opening was
sealed with a lucite rod and adhesive. The chamber was
inserted subdermally into the mouse through an incision
made with scissors on the dorsal side above the base of
the tail. After chamber transplantation, the incision
was sealed with lanolin. Control mice were implanted
with chambers containing CMF-PBS and CMF-PBS with normal
tissues. The implanted mice were sacrificed at two-day

27
intervals, starting the fourth day af ter implantation.
Liver, spleen, kidney, chamber tissues, and tissues in
contact with the chamber were removed and placed at once
in tissue fixative. The tissues were processed for
electron microscopy. The viable cells in the diffusion
chamber were counted.
Maintenance of Tumor Cells in Tissue Culture
Tumor was removed aseptically from a.ten-day-trans-
planted mouse, minced with a sharp, sterile razor blade,
rinsed two times in CMF-PBS, and placed in a solution
of 0.25 per cent trypsin in CMF-PBS in a trypsinization
flask with a magnetic stirrer. The mixture was allowed
to agitate until the tissue was dispersed into single
cells. The large particles were allowed to settle, and
the single cell suspension was collected and filtered
through cheesecloth to separate the fibrous tissue. The
cell suspension was. centrifuged at 600 X g for 3 min to
separate erythrocytes from the suspension. The cell
pellet was resuspended'and washed two times in 10 ml of
CMF-PBS. The cells were suspended in 10 ml of Eagle
minimum essential medium (Difco) at pH 6.3,and a sample

28
of cells was diluted in CMF-PBS to give a suspension which
was counted in a hemacytometer. Growth medium was prepared
with Eagle's minimum essential medium (Difco) or McCoy's
5A medium (Grande Island Biological. Company), supplemented
with 20 or 30 per cent fetal calf serum, and maintained
at pH 6.8. The cell suspension was diluted with the
complete growth medium to give 106 cells delivered in a
volume of 6 ml into 4-oz flat prescription bottles.
The bottles were sealed with silicon rubber stoppers to
maintain a closed atmosphere, and incubated at 36 C. All
procedures were performed under aseptic conditions, and
sterile reagents were used.
Parabiotic Chamber
The parabiotic system (Bellco Glass Company) utilized
contained two 150-ml chambers separated by a Millipore
filter and held together by a bolted collar clamp. Each
chamber had a suspended teflon-coated magnetic stirring
mechanism. A Millipore filter with a mean pore size of
25 mp was used to separate the chambers. Each side of
the chamber was filled with sterile water and autoclaved
at 121 C for 15 min at 15 lb. pressure, assuring complete

29sterilization without breaking the Millipore filter.
The water was aspirated from the chambers and replaced
with sterile 'growth medium, Eagle's minimum essential
medium or McCoy's 5A medium. Each medium was supplemented
with 20 per cent fetal calf serum and maintained at pH
6.8. Single-cell suspensions of tumor, normal liver, and
normal spleen tissues were prepared and counted. The
cells were added to the parabiotic chambers in a final
concentration of 106 cells/ml, with tumor cells in one
chamber and either normal liver or normal spleen cells
in the opposite chamber. The parabiotic chambers,
containing cell suspensions, were incubated with continuous
stirring at 36 C. Cell samples were taken from both
chambers, viable cell counts were made, and 1.0 ml of the
sample was transplanted subdermally into DBA/IJ mice.
Controls were injected subdermally with 1.0 ml of growth
medium.

CHAPTER III
RESULTS AN0 DISCUSSION
The transplantable murine lymphosarcoma used in
these studies demonstrated a rapid proliferation of neo-
plastic growth. A small segment of tumor tissue (2 to 3
mm in diameter) implanted into an isologous host did
not continue to develop, but degenerated and eventually
became necrotic, and finally was rejected by the host
(2, 20). Before the tissue reached total degeneration,
an apparent induced transformation of host tissue cells
surrounded the implant. The transformed cells developed
into new, rapidly multiplying tumor cells to form a
subcutaneous lymphosarcoma. The developing tumor is
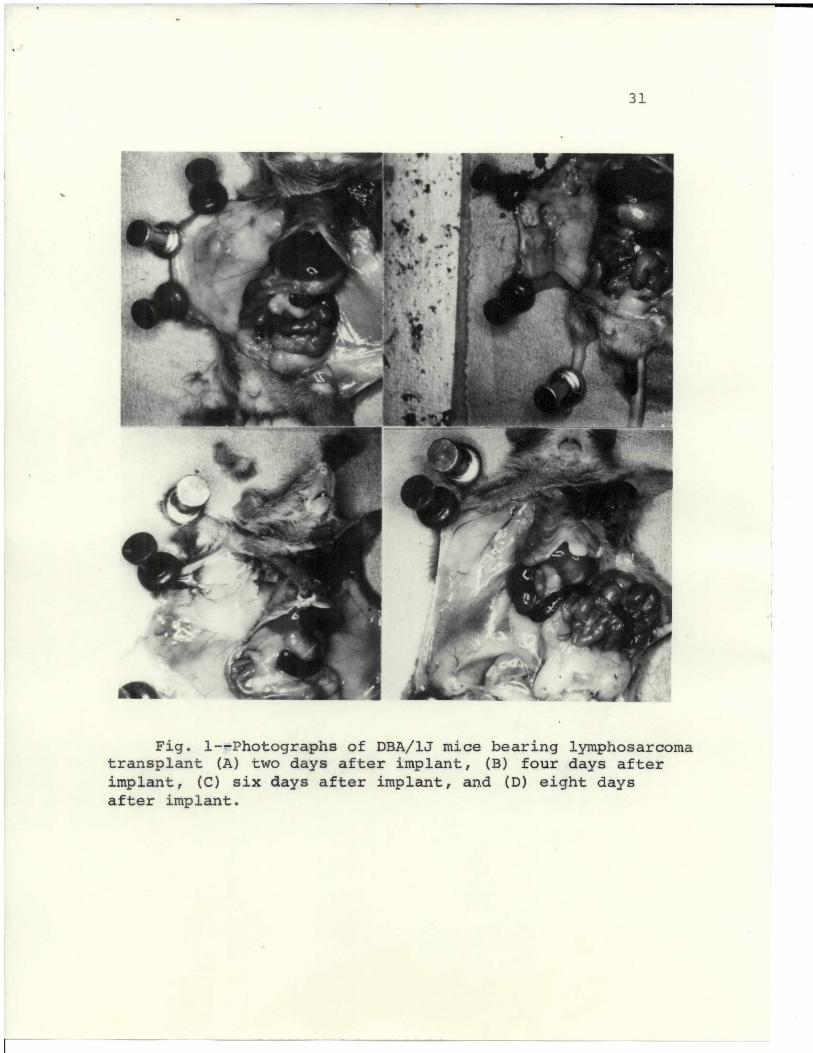
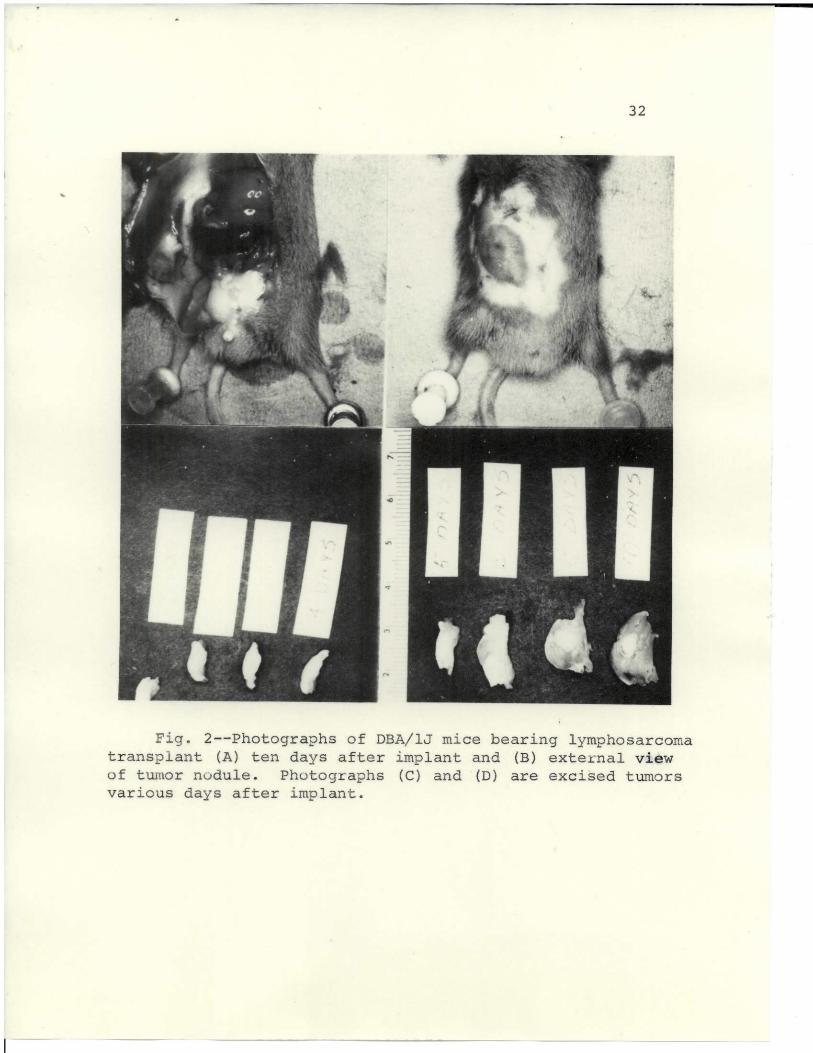
depicted in Figs. l and 2. Figs. la, lb, Ic, Id, and
Fig. 2a show typical examples of subcutaneous lymphosarcoma
tumor tissue after implantation of 2, 4, 6, 8, and 10
days, respectively. In Fig. 2b the animal has been shaved
to expose the subcutaneous tumor nodule 10 days after
transplantation. Excised tumors were placed in 10 per
30
31
4 411
' .i . '
I4
a
t 1
r
1
x
.
4 d;4
s
Y
_ A
i
1" q
9
V ^
1
s
14
Jk
t
Fig. 1--Photographs of DBA/lJ mice bearing lymphosarcomatransplant (A) two days after implant, (B) four days afterimplant, (C) six days after implant, and (D) eight daysafter implant.
'
C
32
I
~b j
k 4+ a'
Fig. 2--Photographs of DBA/1J mice bearing lymphosarcomatransplant (A) ten days after implant and (B) external viewof tumor nodule. Photographs (C) and (D) are excised tumorsvarious days after implant.

33
cent formalin, fixed, and divided into sections.. The
resulting tumor implant is seei in the tumor mass as a
white necrotic tissue in Figs. 2c and 2d. The implant
was very soft when removed from the tumor and was
separated from the surrounding cells by a capsular-
like membrane. The host tissues seemed to seal off or
reject the implant.
Anatomically, the mice showed an enlarged spleen,
a motley-colored and enlarged liver, and an unusually
light-colored kidney. The area surrounding the tumor
was highly vascularized, but the implanted tumor tissue
was devoid of capillaries. The lack of vascularization
into the original implanted tumor tissue probably
contributed to its necrosis. Folkman, et al. (32)
have isolated a tumor factor which is responsible for
angiogenesis. The tumor system used in this study was
not examined for the presence of that factor.
In Figs. 2c and 2d, the comparative size of the
excised tumor begins a rapid enlargement on approximately
the sixth day after implant. On the sixth and subsequent
days, the gross pathology of the internal organs became

34
obvious. At this point, evidence revealed .that the most
propitious time for successful transplantation of tissue
from the various organs of tumor-bearing mouse into an
isologous host in order to produce a tumor is on the sixth
day or later after implant. Many tumor systems with longer
durations for tumor development have been reported, but
the shorter duration of this system creates a superior
method for study. Observation on about the eighth day show-
ed the animals beginning to appear leukemic, characterized
by enlarged liver and spleen.
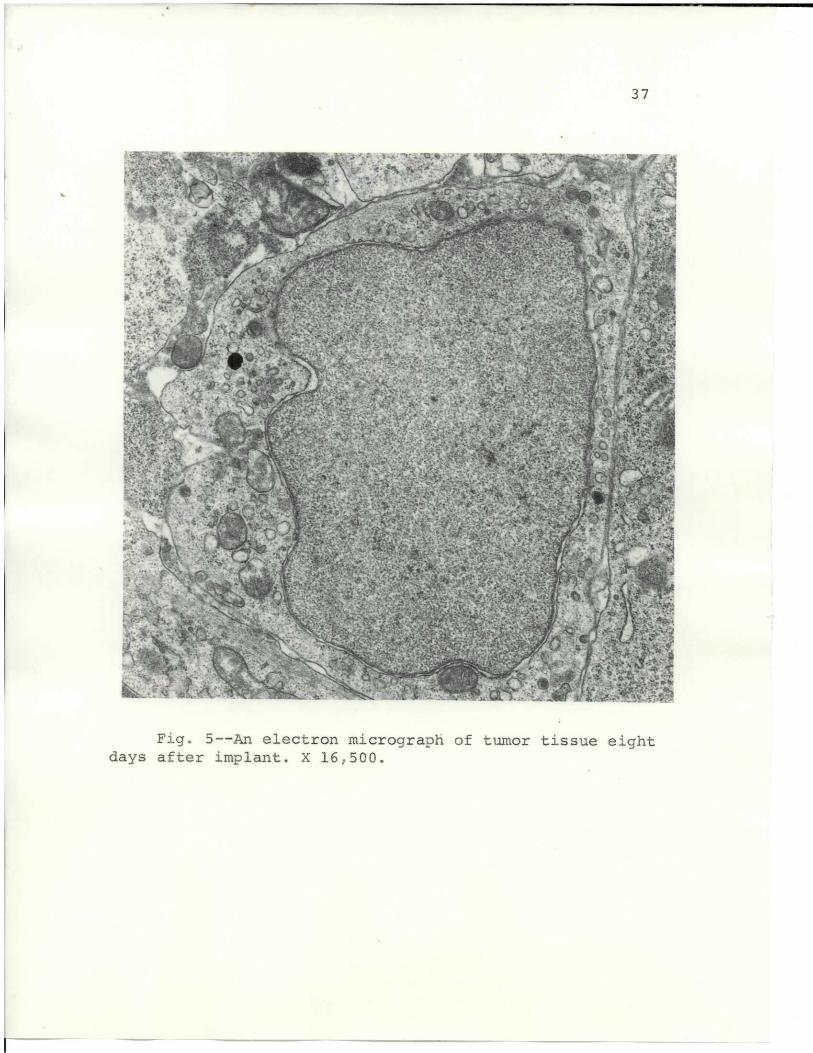
The fine-structural features of the murine lympho-
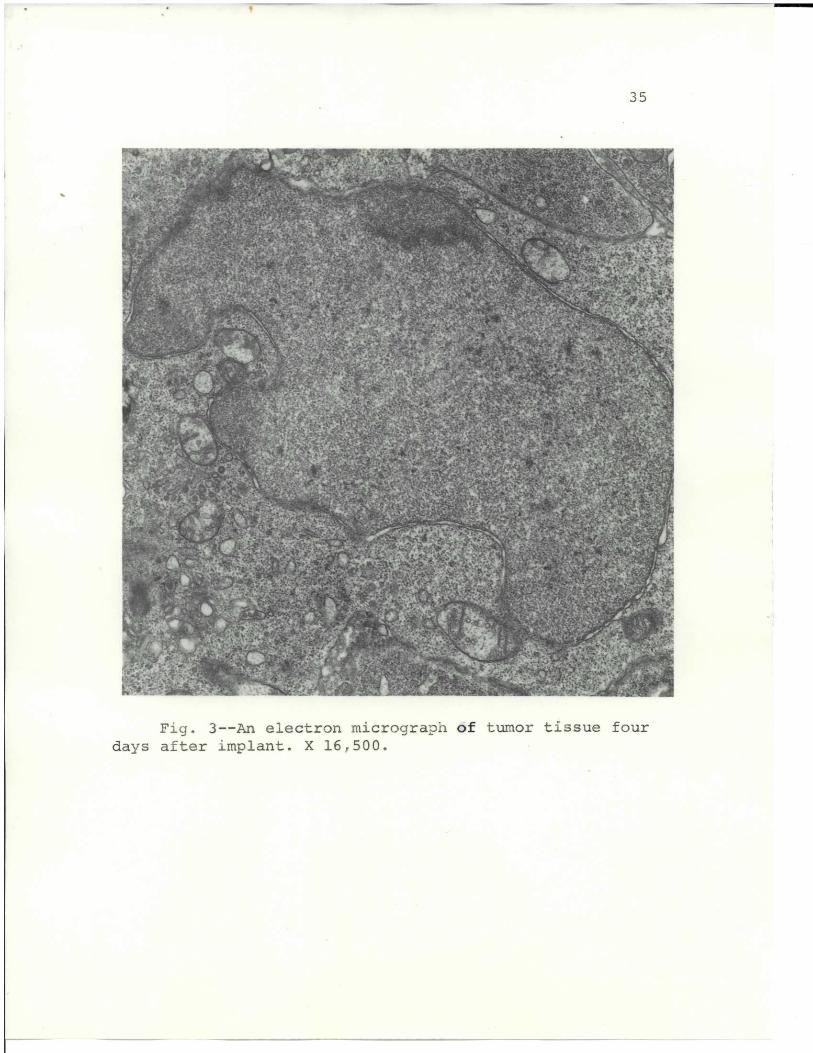
sarcoma are shown in Figs. 3, 4, 5, 6, 7, and 8. Fig. 3
is an electron micrograph of tumor tissue taken 4 days after
implant. The nucleus has a distinct nuclear envelope with
only slight separation. The nucleoplasm is homogeneous and
practically void of heterochromatin. The mitochondria
are almost normal and show limited pleomorphism or enlargement.
There is a marked lack of ergastoplasmic lamellae, but free
ribosomes are abundant. The tumor cells are in close contact
and do not have enlarged intercellular spaces.
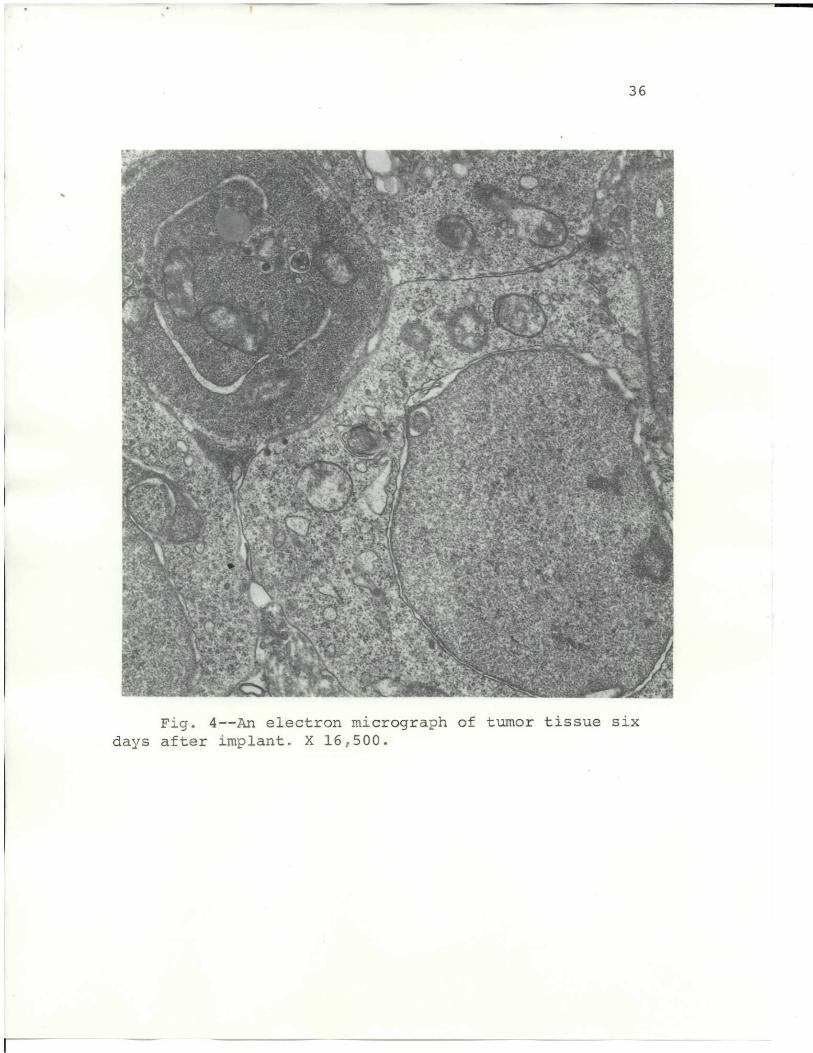
Fig. 4 depicts tumor tissue from a mouse 6 days after
transplant. The tissue differs from tissue in Fig. 3 in that
35
+ - " " " .r .""-+" T .. f. ,^- , " ri C " "*"- f. sue... .4 3 R pf
" . f ' ] , 'A .4 ,; s , ,.i / f' 4, ,5 ' w. " j""Tt;' " , '' " _,,, C1 i .1r,1" r i R sl 1 L y .' . .
. " . ' "a " " t i' " - rti!" :F t l t~' ! " _ . .nr ." f , . t "} / + . 1~! *"'.' "r'' G Te. = ~",y , + " r .
%.' " " - J Ar"+ '. ",
"+ Y. r'" w . i f a " "j",, ' f r ""t a/ s E;
a" i. ,}, yy" Ilk " J N r . ' .. ! .Y a r>a tvty. r i " 4 r , .r, " d l * t! yf
" " r " 4.. " a " ... / . 'S'a =. 4 yy .o
" v" 's r' a ... ""'t 4 ar.'1 " i n ' r
f l " " V ., . -, - sL . + ' R !'' r r "i S'" +.. Y; a a ti t + '" ; ti,. :. S . h ' r . i, JL: y,;
'+' "'" ._ *" " } " t" ., . fr'4-/ " ,'.' ".i _ . a.r Y .ri-+,R Y. " {. k a',- }a' ~ > }"' - ' t .- x' t
. " _' I a i,. , " , _,, i-" t"}} ,,, ,\/ 1.T T " iss "4 . '"a "
INS* tda "ff n' " ' " "'r+ v " Sv '; " " f It " "a-. -, " v s v,1 .' i Y '
1_ y {" " . r"'w7; 1"' t sZ " " "" tf^ : ": + _ it "T ti+.!.11 a'=M rs. 'Xa
+"t: ""' . 'S ' t .. i. tl+t" S - o' ^.. " ." " 's " ". 1 : .. lR f..t t .r '1, ." , t. "'C" t
': c '. j Y 1 f ". ft',r, +a j y "r14 .. S' .. 'rvc tir .'"aa "-', i : R C ' "' . ,".' "" "r ' 4'4-.6.' a".. ; J_ r v
f 1,4
oil,^P ".":' "aaT,=,1 t ' ."t' . .P' L" 'Sy .^r + _fCL f'.. " t.,ya c(? . Ij T )' . ^. .> ., i'.> '
' ',, , a"; : _ {3 '' . j _ " ff 1a ' o l v 2 1 " .y. ",c"Z." S .. -L r- - ]t F.
1-46 f "R ," 'r;!'lf y .'" " i- ' i Qrl +T : !\ rl" "a "r r ~j . "'' "'.f, vvR M. "
.yy ' r '"A 'ri ' '" ~' > r 'a w"f; a. f " t i~' + fr, x' ca. >
.. ". l .'Al71 f .' 4 1 ! ! " " it . t Y " iR + , v " "". Y i (c, sri>."4 3 y ,
[- t ". ,a 't r"a'a J * " :e -!" I ".,'isf' y t." ' 3 'l ' y:' ,,j 'rv .. IIQ. " , ti - " ar, " T(" F yf " !'v " X i
y, r f v R !' S" 1", l '" vf"" .. iJ . 'rjl t . f 1 ! ''r >, "s A? r_ , f f +.-',a t ', " ,
b ' " "Y ZaT"- . ' t" '7'tt " C Y '+S"it v . a "^r . ', , , , f ' .e I .. 1 .' i' . ". 1.:
.5 ~ r a 'a " ri 1, r. " '1 f."'i1.'. .a", 1'a j "' .a 1 'k '+" *- " ': "e 4,f:- l, r :t /,' " "
'e!f "" h. ,.1 '"1f' :dl= ' ,i A,,r. j ",a\'{"it'.a. Wes,>- .e a ( t Sr . '.' v '' "t .L ./,J. " NO" "+_t X11 ,.,,,..
_ il + , )..I: .". 'fi:a , ] :,'' ,\.. . ,"t . ;4,c 4L 1 + ; M.'.1{I' 1l .,' tx;«,. " .4.>.r.+ w ! .s r. a -. , ,, " .. pY{,y
" " d'4. w ~" -"a'trf } S laa.t > ' . s" ".".% i"j'h f,- l,..r+ s? t r r "a, "' M1 '1 . 7 1 c""+ _ 'Ia"
\' " "'+*>I '. ' F ' , i f _f f_'v ! r" " av' ,l" r." a " a tiF .:t V .' .. " i
r "". , - r t; w.._ r'y+i ta lya':i .1 hi r! N5 .f.a'tt" ""'T!! !'" - A . " " ". L .i " r ,",r ,,r" "t. ".
ir' ~ 'r 7 _ ie " w "'T" " ' + , 'f f. .t Y ! . V y f t ~ z ' " >t. . ' " ' ! - r"
y { " , h". "J4 lr . a -Ri" fi T". , ' " A!! ", '' -i' Jp, .,: ~ ifK..? ' , '.,iyF , , ^i" i 1 ,"u.' " : : " a +~' tatv ,
' '. i Kt <.i.'S1.'S^ "r. " - ",.':+. . V 1.,!l l . . , r ... " i S "" a'" 1"+ ' n 1w, .7 ; .. .
46 N% ",<. i ^" -- f .t"":. , ,,.1 F "1 f a f -'i / ti
"'ier 1K " j 'df" fA '" ' . " t i .'w. 1 +.i"7 ' v ' d "a - ti'a a l~
a"r "! ". \ i" 'O"'L vl ,1! .ar " 'Ir " "_ 4 y " < ", 4'. F a" 1^ ,4. , - ., V Pl. . F,," "S
TT ,, } maVy. { 'v '' / "Fr'. ",_ ij. '
.. "4..r../rt7 K'i ~ _ a. j '." }f k?} y'rf s''O? 7i , ] , . '. 'a"' .( G.MR aj'f" . ", +.. , .y1R '
.4 . f <]'"^e ra 1",Y"r I i M Zf " ".1. r . ".J" ?''l. "" "' .. t S:"y . ,"""
-, 1 a I" 1 . ' 1' fe -t+ b " a^ t s} " rf "."!" "r a. " e. t .,' . " t sw; +r i. s 1s 4l t _;Ice _b ' "'
Z"1" ,a, a'r!" A} ,+ r r". - +D. , 1a Y 1'yzy ." j[ y G; ? fi. ^ a =5 , v 11 ^ r t:l y, C.
'D "', - r - .. " , s C' " ''= i t y i + LP "' a ' " '' It r' " aa ; ' ,y . R '
.' f i.! ". ; > {l ay: .. A"i tfi sv "'Sr' Yy } = 1.",i i " ' , K+ . affl a ""]r
k TV} "a1 J .t- ,L. :'''" "' - .,e" 7. q'1.u1_ Y S:,ta =:.,"- "s , Rt 's f'-a'" f a" %;
.t : ;3i'/ ti " r,{' + ''" r'" ji "'fT ,. 1 a R -,' ' _ ''f > . " " ' C 'v- ' q" .r' , n ; . . '. t .a ,A ,1
a: ~ ._ " r,,. r . "." "'" 'v* " i ? , f f. '. ii, f "Tft ' .. a .s. " lI .. r 'SCr
_ 1R t a" r "' " # f "g, r 3' j . p t, = 'ttf< + v.'4(j" t ' ' l r'a jr,, f . " " ' ~ , p+ ff: .war' ". t r' _" "tfi " +1 " : " ",, ' a i!' al fJ " " J t ++ }f1 ". . v e ; a Y i l ; M '", a = '".-j .!'> . a + 7 ^ " " nS "C " '. a : Y" .,Y F" Z
" "A'. ti -". '""" .tie." T- 'ti" ti .. - ' >ty4" }''+R". ,3: t ' 'R A ' i y, - ' ,,yj .t ' 1 t,' " .v
" _ _ ry { V '"'v' j " KM f \" i!' v ,", e}{V=""r_ i2 1 ! r vt, - +.. f 1t? {.t "1 *ir"' 1''r \ ", j 7L t 9 . 1.> ""
, { '. : ' i" . ' 1' ,~tj:* " , . ,y/ .f ' ?, a 1 M '1i 'rrs", '1 iY 11..T' ' t.. .. ,:y J . f - y.
ti t T . _ <'r' l yr w ._. f .v~ ., i ; S*.4" Y, i. T + "" f " " ,d 1 s.i" 1:" ""+y : y r ! . T " a ' 'F . " e' e+ p " "t SS " Z
" / " A .i! " r ~- " ~ r : t'3'S t ', , " at, a ?' ~ ". # 1a 4" ]ts Al . r s rt a1 t i fta , ft" t " r, ''r , , e '' + , tt _ 1 f"L l. n .I,..v > a +! a "' #.i r" :/ " 7. , ti . ,.? , 'S " , i f rt ,_./ y 'R ,
- ^1i''"'> ' ' « r, 1 F ,l' : :'1.-.- 4,p" .1"F,"> " .L~4 , s .l-i , _? "7 H, "t"" t-t l ~ .a. s P'" F'TiT r , r y
; i r .y' . ~ z :E . s' wra _A "at .. 11{", ." !, a qi ! .. i
" ."{ "'!. O Is.{ _.. , i\ .A {{" "Tf ,t" .!"~+ 8+ 1- t.' 'J'am 4" i "' " l.fs +... a, "d, " -- t. Ti :F yt..
.a " . l . r , .,1 rxAi x+ ii ? ' C", r 'r ja'm' C x ( + " 71e 1"r j, ; 4p
" .;" ' .~ "a. "L J ',i }, "'. ', a l ir", .tl j+ Fes' M'l, " ,, .! " " " '} .1 1R "". R. -I t"' a .t7.1 . I' . [.
ra i " ", , ":' :is S a.f =: y'" t .ar y +'a Si O :t ,t?+ ""' S , \ r } . Wit' a Y= iX a ' ' . '3 4'. ' -'! ' " AA 1" " '' 'a..
yw, .{{," r ""L j "' L't*irty "' ~t ~ F . ,, . v ' .. i - d , F:i
' " * ,; , ;'ft j '" 'lti " wJ'' .> n r ' ; '"' E'' R_ 1P 't ' ;yS _ ' '" " : * ''
' .'9 a"" a ly ' " q v , tr ;. i't _ t. Y: / y ' , X . a I"" "R.s '- \ t L " . F 'a' R , f i a f-; "tj,7
f"1 " . ' "'R " ." '<.. 1"1,f(": "'!t "i "17. ' ' . ! f . .. t! r 'r ";. T " -rl4 + _ry ." T ,'t
W"s" " '! ' " ,,."". " -y J + t a- f:" t" " . 'i - " . F wj ,1r '' , _' ' S' " i.
e',. t ' " _ .'t . . "A " " " " ' a . aC i ' ^ A' .R r" ! t' 'Ja,,._f__ v,.y
A+-_ , " .r .}" .. fi r"r "ar , r: .j' " t rlt ,,i' : "' . {1l. rte' , " " <d ' , . .. .. j' f' . , 1r.Lt~"' >it- f"
a" 'X! ~s .. . t S ' "y 1 . .- . = s " v + _ S " " .: ' T, t 1" i " yf 1( 1't" s Y. 1 ,ie , -J6
0, 4M"a1 " ""r. ' " J "", "' " s ' Y . f r" .. j 4 "tS " i ""4.
'' t t ,t> " i . wLt ,, a i. "ti , tea. , a. w~'.. ,z ' .f ,.", to 7 y r . a""lV rf"7". rr (()) ? ,}"^i "'"w,. yyy s .a " 1 w ra a t\ ,
- '"..
s . F "
5 4 .+:\ ; ~ "' ' a '" ^, "a" ; " " i 'v++. S"t4"wS r ," t. i.--" . .N >_" v . '.
" T" .R 1T. ,' a x. 1'r S A f Y ",a r l,,r Via' +! -~ R A 1aa' ' " ,a>.
",$
" "" +"
iy KiF ,.. . " ''! L :i''"' a ; j t ,} , a1S a " i"! a . i '1 F..ic. ja1 " r".. y - f> -
Alt
' .s ' l + " : v ; i,* i ' :t. '"v'? " " ti :. t .' , ." Wit' lt , , C ; + 'i
1q ['.1, r] t A " +i (ri . , I-Q i f 1 ?": " f afT 1 2 .ti y R a.. . i, '>: Fs
f. '' . "y ,"r ~ { " i L .. " "> Il 4.. -+ '/1' r+ ' ' a . le" '! 1r t.M,. i "yi. ^' "fi ' A it'v ' " t_
"=r. : " .! . '" ".. _ _vf i "i+. " "fi l a .",".IS ' e." a L .a" ".f Rriy" .. t -j>. .5 ""11Ni'" i
r " " ,,,, - ' .sue - 1's,.." '"'; ;! 4 L , . ,. l r
" .v'. ' L'C +4 'f*"- 1 ' l";
7r7, g" "'.Art r t . '."'1.+/ -t: "i ?'i RS ~1' F cl -t r'"" '
,14r
.1
" ! : .r a, ! ea. i '-. ,;' ' W' !' .t. . 1 t'' , j+ " . a . +t _",i ryyrl h.': . tl !S 't' _ : ".
% I-S
41" lw :" 'f., . a " ""= w.C i a" j - +v ! 'f" .A u7 '1rt 'i- a 'i " 1} "4y- Y.. ,.,r a.
M" Y" ". '"." " + " ti ter " 'C . "a " .,,' 4" _ . * .. ".:' .. i .i " ", 7" . r }a~"ia
r . " s " ' 4 "" ti a " M " f!y .r! - J - M, #!K .:sfr _ 'oli r -3 *" 1+y. ""' , "''" f ! 7'r, * " 2 " y , 4 O, """ r "" ,. 'I"f.
.1'ar.f a.Z " C! i" f' '" R. v K . i " f A 'i , " '' ," .!_'"'s 1."j" " dt , f~' Y
.. ' ," r ". _ .ir " j P "" .i y a, . f rf . V" .Y s " "_ t . f4.
-. .rf!'"" a v , C" 1 x 1'I 'l \"^" + i. I+ w' + 'A,' "' ' K :s. I g , ~ 1 .i +.' v7y}1
^" , , dd " . " "ft _" " f " 1 A ' a'n'Z.i " ,'1 tV "! "w .F >"fA. " S." V!-' ., Si .t + " S." ?. '-": >
Fig. 3--An electron micrograph of tumor tissue four
days after :implant. X 16 /1500.
36
y r e .. fpZr '+s i %/'r7 ;;- ' ! >.: ,.'r
0 1
14 It. -. 4. ALV "jYr~ " '+% , ,, r . my . i ,. ". 'r t '' "'" a." "{u+ ~
r.i~
.. w . 4v i~ '' i , s A. su'l t, T 1" Z -
*4 rr.j!J~~ w ~r F " " t.106.'
f I . - + , ti S ? 5 " & " . " j " . - + rr" -t. "M y , rf
'a i j r~~, I A . '" .j r, "j/ y_ " -. . ." .ar . r "-
' M '+" rfr" '7 r r R" r * ". - .. l " r l"-: " t ": o " .fy. ' a Y " r ; ;
1A~-1' ..V% ['y '. a +t .! - C +Ei i ''7 "'..
4tqiY Y,.f t 'i +I 4^ a" .'" A ..."7 " r 3 ."' y ""
l" ,R i. . r? '"AV#," ; Y. _" x .91 '1 " ? ra ; "-. y: ,"If ld !
.l " k 1' ̂ *R " N" "r 1: t$ 7 _i " i li S * ""S. i . , +_ " S ' . ? , d4.rI T "Y aiT s ~
F""", ~ig. -- n le r n icrograph1", Xof- tumor rte ti"' 'f, ?-..issue s. ix rdays after implant. X 16,4" iy L "4}~i+"5OO .
37
94 rtom, ,..'" : _ ,. a . + ' * '
-o~ w ' : ~~ ~ igr;f * i i" - _ t.rl} .r " :.-." ',j;k''. 'i ' ; f I
"* "- . ~ ' " - 14 .-u eA- C14 3 w f>_ t" _ " V Y
! ' 1 ' f " r " .i ^ \ta_ . '- '':" f " _ r ., { a 1 t y
J , e1 ' 1 .- ,y + ," . ./r " , : ' v T tfLj ' + ....o.>11 < " }"'n1 ",t 1 - ' -' +
4 4L,.
,.a- '; W - ~r l f 'l Y."..f,.L J.. ->" "." V \ " kA" l i f""-- I
tEij'!1 Y ,ja". I.~"I " . *V a *' '"*':.* '- . '7 _ ' t " c. a=.\; t. . jt " t i ' ' ' "A . .t' " ' +". .." . .! "s '7
V "_ " .. -." t. -.l.. T -3 S fj: "rA " .!v -r 1i "!
414"- t Y ,- l ". a " + 1 ''{ "_--- .. *1 ' ' " -i " . " y . " .a t jr '/ " O $ S "
V , +' j eI / A , - " +Y a . -,3 _ " i N. '. " r .̂j " " "" #, fr. ! '1 = t! ',i1 Srai1i l V A k ' Ad. i aj >a~ ifL
Fi"g" . 5--An lectronmicroarph of tmor tisueeighdaysM. after6 implant X l6?.,"- " lr """~«".' ( . 5OOw# T' i
38
r ! '' ". 4 't a"A « - a" t.' a - y -. - f ': \ K..".. . .
r-U. ., 3
"ya ' " -.Y .t 1 "_J_ -{, h "i" t ' '"r2j2 , t "? .L <'- I" rv I A "^
'?'7 f^ y ., ar T" .-R , i Y .!'+ "_ ' . f . '+ 4" ya . " a !-- -.- .
A O.1
- -"+' '" 9 + '
ti ' a ' .t! A ,-3' " 5! " ̂ s> "i t .
-? " _ , . a " , t ,- '- - " I -, " " ' T - . : d ta '' " r 4 J1 Z
- , , r }- -~cT _v~r -'" .4
"r, r. ;t{f.c"- ' " t
4 ' .x r." ".n r' " ~ , ".'t ' -, , a r ."
k- *e
r" i A '- ' . 1. " y . -" . ", :' ,y.u . ' ' . f 7 A 4 ;~l<; . d_ &a "f y a A ,- - '
all -. ^ y - ,r ' ! r ' T_ r i V- r' " ' , " .,- '. ''' ,l~,"- "< ~ "!
" y: i .. -"~ r Z11. " " "S" . r_ r. f~ .. e-"S ' = '..: . .y rt
T, ; ,...77 _ " _.i.i. ' ' '_ M .
Fig. 6,<^Ar Selectron m, " or," aph of tu.. morIJtissue ten daysafter implant. X l6t5OO
39
f~ ,"I , \ . ' it"Ly I -- 'ry. ~; /.. 1 ~y t . f ' : 1LI
16do,
~<'b~~~ ~ ~~elA,'7N-it
i ; wf , ; J, A" ' .' .3 ; a'?.,p 7, "- ;t { "..' ,gam _ ^ t . ' . i $ '/," r .- Z Ar 7 ' -" . j
r~ . .!T rf ,1' * f1"' ' " " A-,1 I -"1.,' , " : _ -.4.r , F' 'a
" f 7 ' " '_ , .T " " " +" " "" _ " M .--f r -."
i-z. .f' -, :r "'s t s ' s - ' J . -j ?1 r
lI rti f.{ a "^. !.id j" i f "" ' ,'.L. .. 1," . , ~ J w , i '1
. L w " f i -~ '" q . 1" ' T ' " " . i ' * / _ Aj " r a r l "I S L
! , 7" S _ " ..1 ~%s ' iz ;; -,V
A
' 1 .* t _ ir t v i -
:S'o ". " ' -./ .Y~t "^'1Y of I " t>.'.ty , '" "' ." -'. , "" , y "" -' Z I", _*\.l )=~, . 7'. ". 't* J
' Cre "L 4 , ' s r ma 6 ' . . t .c! ' f ' ' k * ' r
13*t ! " ' .".r 4 q A '- f . . :i '' / "' t * " " v' " C. ,' VIPC** 0'6 -
"._ ' "l " ' t j" ; <f ilr , *tt ,, ew r> 1 rS
4 L , . ay.} ?" ~ , "1 1 I,3w " r. .. w ' r. R /
+r :i!"" ",zf > 'rS. '"~ y ' w _ ; M " "I f :C. '1+ ^t ;;t", 'f y.. '.~*-'-'yl~
Fig.7--n elctrn mirogaphof tmortisse telv
days after implant.a X 16,500.*
40
'. - '-
I ,ir /e J ' ; R S j G r J "r f "
~~~I, 4".w '1"' f. f ' " / " f dl ' r .1'Ir 'i '7" 3 'f i . F ..( ;, - "
t:" " l ' : d f.' ll{ ~. s 7 L , -.
- ~ --- I;~Apr
'NitA'~ _ " i, i \+
4 .'.i1' , , ,A , +r f'I /; f { "t ' ,- f,^r " J ,,
~ -19,
-A %"".. f:l~,y ! ! :'r - 17, . '; ."+ /' 9'.1'1" s, -7( /rJ'", '""" f i ' 'i"% 'f:- ,'" y"Ir. "v. rA."t'- f/.-"' . '- -' .,
,1 ~ ''1 " " ,(,'f v " Aye T , ~i 4 .. % i sy-i /"S* r "y
)" " P 4J T a' ~" ". if 4 , r ;, l ," 1 ̂ ".i ,/r f . , , z.r . r ri 1 r i , r . -.. , 'r l r t fw f 0 ,
9~~~ II 1.'- 9*
Fig. 8--An electron microraoho tecnta ecoiareaof tmor issu X 1,50

41
there is a separation of the nuclear envelope and a
slight increase in intercellular space. The cytoplasm
is devoid of ergastoplasmic lamellae, but abundant
polysomal formations are present.
Fig. 5 shows an increased separation between the
tumor cells. The cytoplasm shows an increase in vesicles
and a decrease in polysomal formations. The tissue was
taken from an 8-day tumor after original transplant.
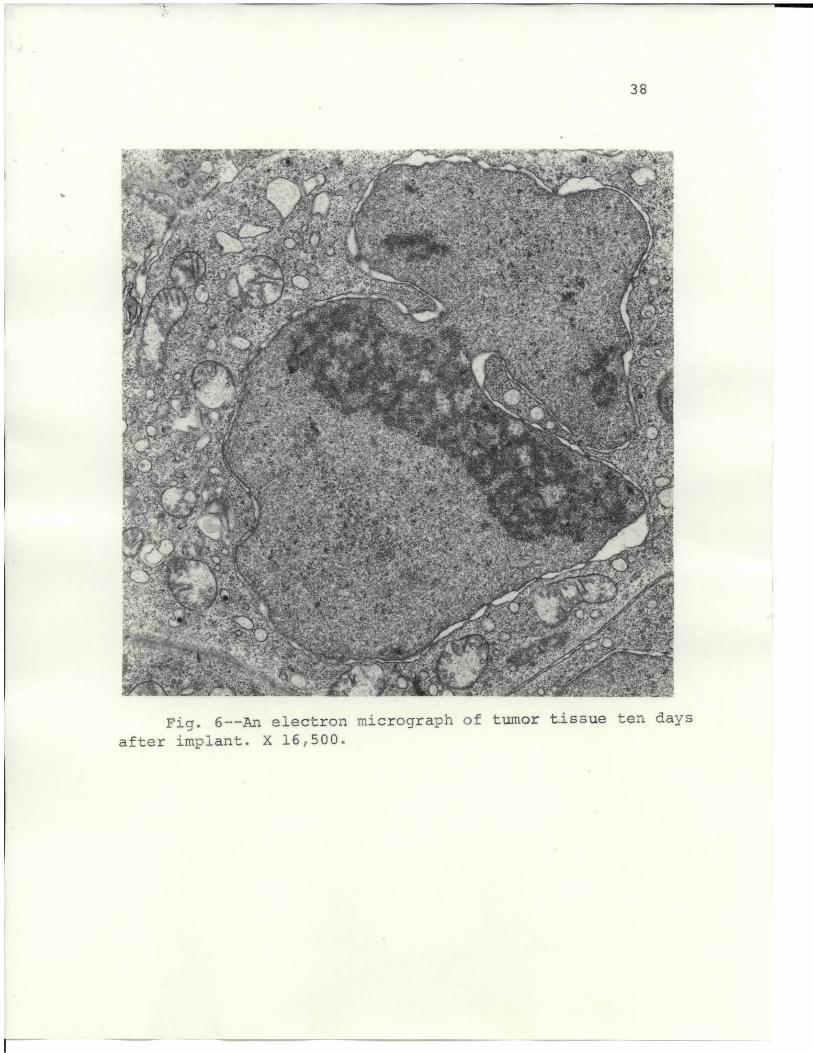
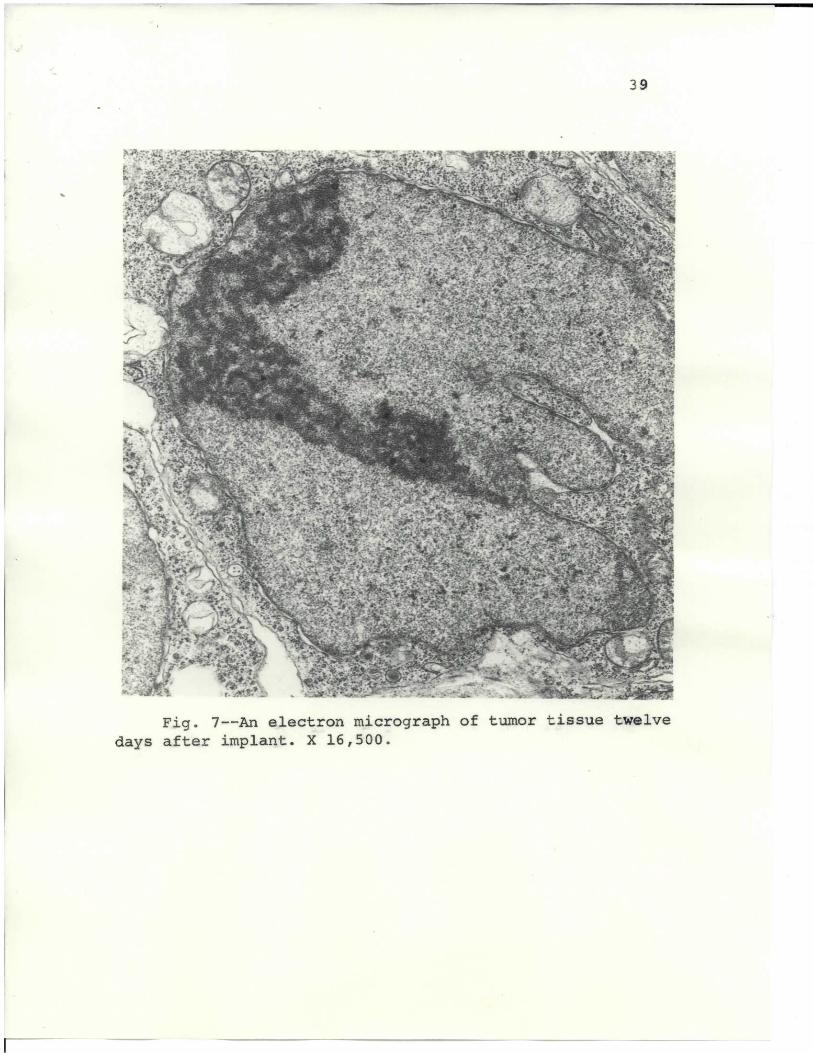
Tumor tissue taken from a 10-day mouse tumor implant
is shown in Fig. 6, and from a 12-day tumor in Fig. 7.
The nucleus shows irregular shapes and an enlarged nuc-
leolus. The mitochondria are more pleomorphic than earlier
figures show, and have a heterogeneous matrix.
In the development of the tumor, the nucleus became
porgressively more irregular, featured deep invaginations
into the nucleoplasm, and formed nuclear blebs. In this
study, heterochromatin was not evident,as reported by
others (9, 69). On approximately the eighth day after
implant, the nucleolus became noticeably enlarged and
dark. According to reports, the nuclear membrane has not
been known to manifest a difference between normal and

42
malignant cells (9). But practically all tumor cells in
this study system demonstrated a separation of the nuclear
membrane that has not been reported before. This separation
.is not considered an artifact,because control tissue
prepared simultaneously did not exhibit nuclear membrane
separation.
The mitochondria became progressively pleomorphic
and distended. Their structural deficiencies were clearly
defined: heterogeneous matrix, irregular body, and cristae
shapes and swirls. Fewer mitochondria were present when
the tumor cell was less differentiated. And as the tumor
progressed, the mitochondria became increasingly degenerate.
Although there was a noticeable lack of ergastoplasmic
lamellae in the cytoplasm, free ribosomes and polysomal
formations were abundant. Swollen cisternae disrupted
into various-sized vesicles.
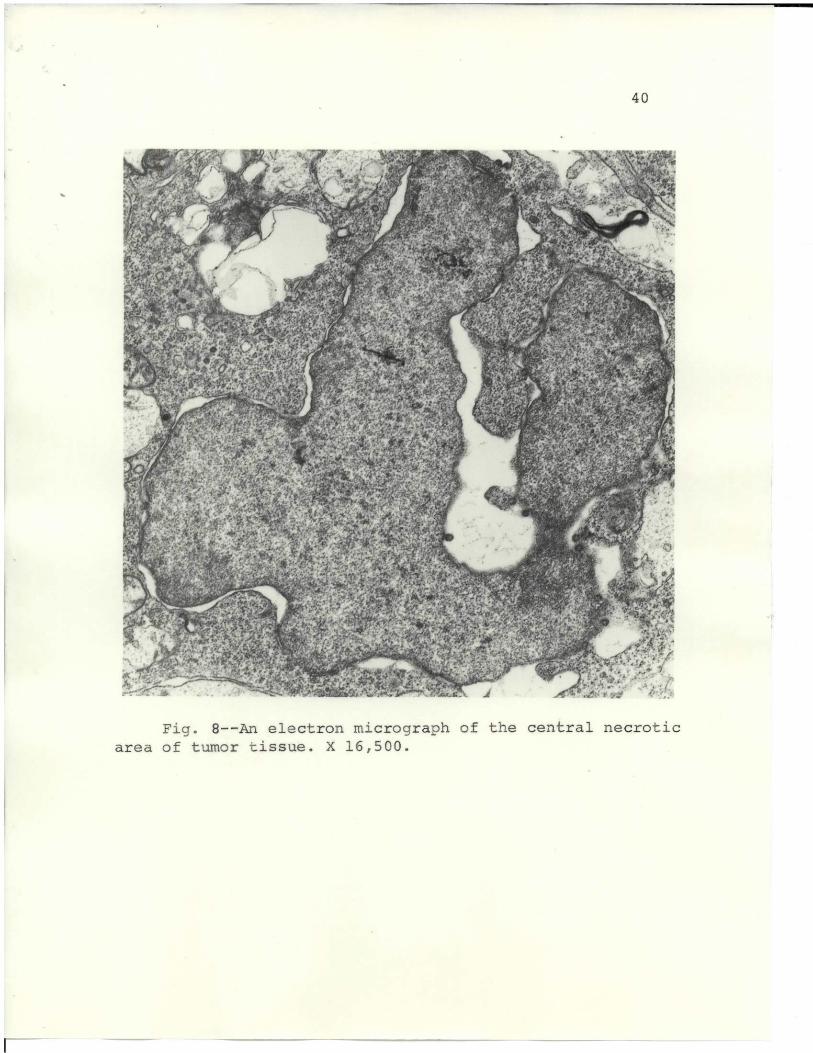
The tumor implant in mice became increasingly necrotic,
and autolytic processes took over. Fig. 8 shows tissue
taken from a necrotic area in the implant. The nuclear
membrane has become greatly separated and is characteristic
of the tissue. Nucleolar lesions were not observed, and
the nuclei were the last cellular organelle to undergo

43
degenerative changes. Cellular degeneration of the tumor
implant mass proceeded from the exterior to the interior.
Cytoplasmic alterations were evidenced in enlarged
cristae, fewer polysomal formations, and expanded vacular
spaces. The mitochondria were greatly swollen and showed
signs of degeneration.
The nuclear membrane separation seen in Figs. 4 and 6
may be an indication of necrosis in the tumor mass, which
is greatly expressed in Fig. 8. As the tumor mass enlarg-
ed, there was a decreased infiltration of capillaries
toward the interior. This would cause cell starvation,
leading to progressive deterioration and necrosis.
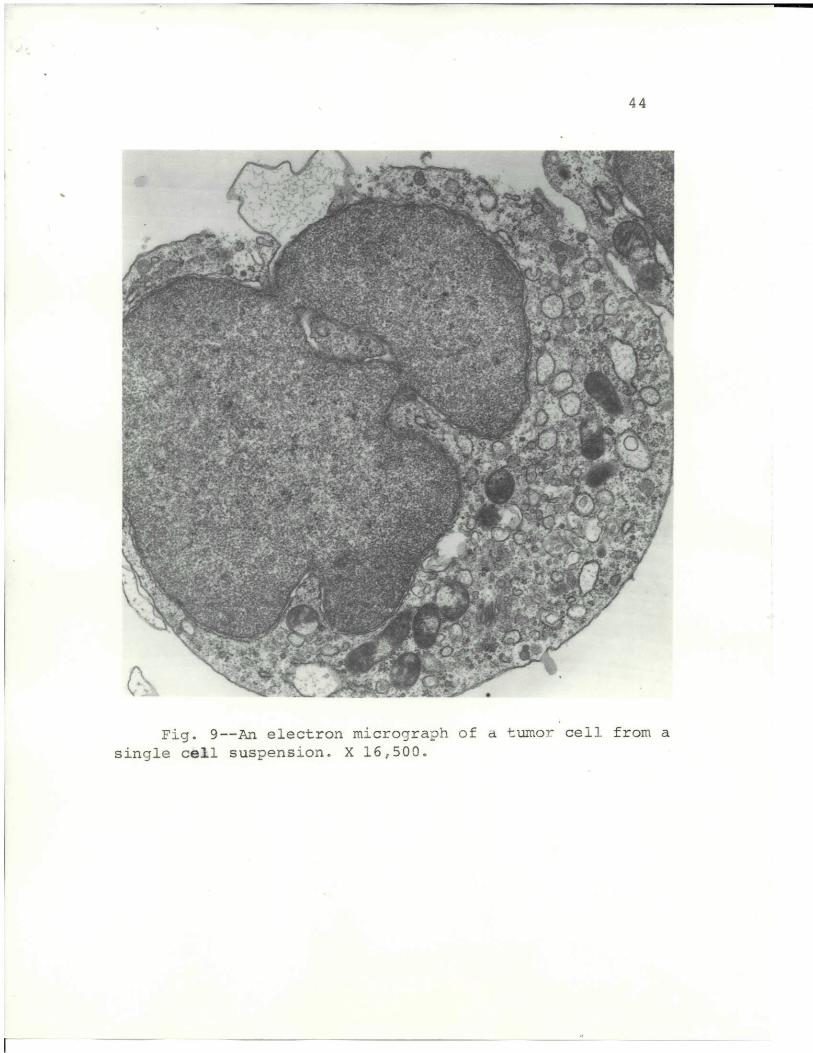
Fig. 9 is an example of an isolated tumor cell used
in the cell suspension experiments. The cells were
prepared by physical disruption of the tissue to produce
single cell suspensions.
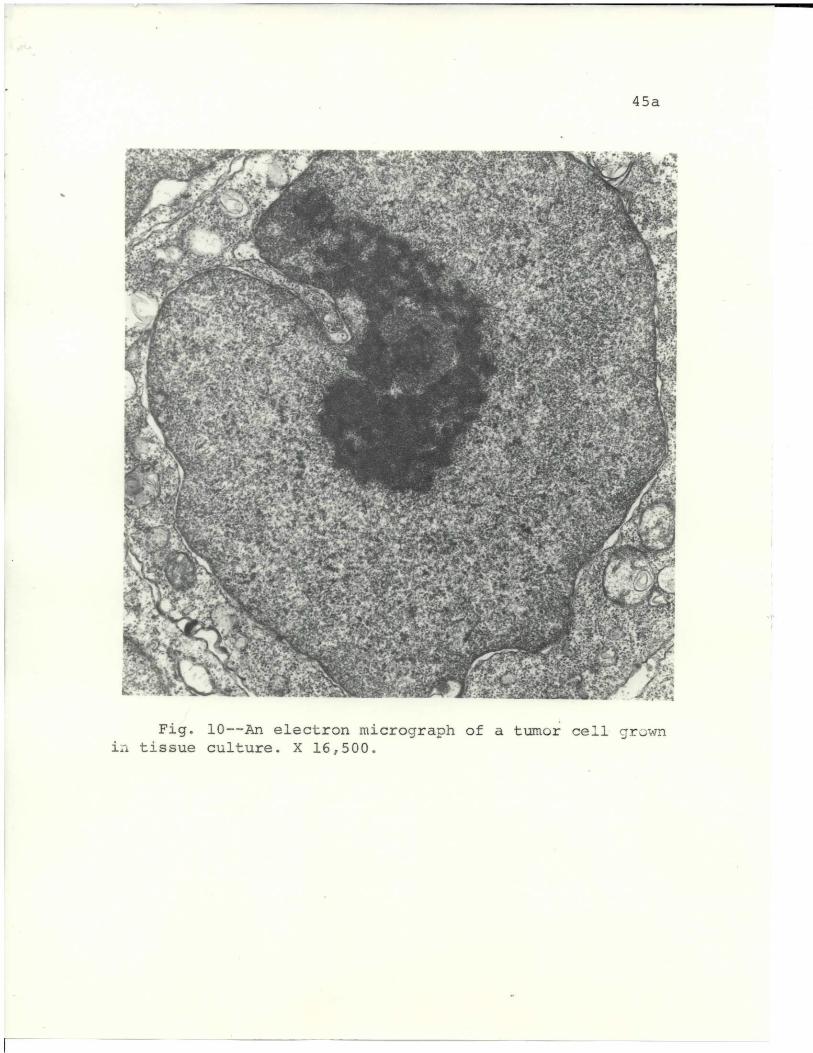
Fig. 10 is the representative appearance of a tumor
cell grown in culture. Cells from cell suspension
cultures, parabiotic chambers, and diffusion chambers
gave the same appearance. The most prominent feature of
the cells grown a vitro was the enlarged, darkened
nucleolus. When grown ia vitro, the mitochondria became
44
}" _ _ ' " i.i t.017. 1746&
~ ~ fT
M ./+ f.' r d 1 t, , 'st N^ wL
lip.$
' 1 "i'* 4 'I yy(: ' ~i" ' . ",{ ~
Ff k C t.,. Y Iitaa' ce . { - . y ; '+y '1+,",: ;::" r yew~ L-/V: . . , .. (. _i ' .. " y:
" r"f t {s i""' y a "t +"''i Sr if -e 'A , " r a r "' t ' wI -r : J ., r4 ' a Y Ia.- ",
Fig 9 An elec' ,'" "' tron microegraph'" of a tumor c cellfrom. asinge cel supenion.X 16500
-1
45a
f ' '
>t At' _ ""- -4 -
16VtA 4 I"P
' 7 ." 7. -I tI4
64r
At.~jr 'w %~3\' $@OP T
Fig. 10--An electron micrograph of a tumor cell gr wnin tissue culture. X 16,500.

45
increasingly degenerate with time. Numerous myelin-
like figures were found in many of the mitochondria.
In all investigations with the electron microscope,
virus particles or virus-like particles were never
positively identified in any of the specimens. The
observations were made on several different tissues,
subject to various treatments both in vivo and in vitro.
Table I gives the results for a lymphosarcoma
developing in DBA/lJ mice after transplantation of tumor
tissue. The tumor implant tissue was a histologically
diagnosed lymphosarcoma. The developing tumor was determined
by palpitation and visual observations. Four days after
the transplants were made, 34 per cent of the mice showed
tumor development. After 6 days, 82 per cent of the
animals had developed tumors, and by 10 days, 100 per
cent of the animals had tumors. The mean survival time
for the tumor-implanted mice was I1 days after transplant.
The exact cause of death has not been determined, but
several proposals may be considered: (A) toxic products
produced by the tumor, (B) necrotic debris from the tumor
causing renal failure, (C) metastasis of tumor cells to
other organs, (D) a leukemic condition, (E) size of tumor,

46
TABLE I
INCIDENCE OF LYMPHOSARCOMA TUMOR RESULTING FROM
TRANSPLANTS OF TUMOR IN DBA/IJ MICE
Days after Number of animals Per cent of animals
transplant of developing tumors developing tumors
tumor
2 0/50 0
4 17/50 34
6 41/50 82
8 49/50 98
10 50/50 100

47
and (F) viral or viral transformation of vital organs.
Tissues from various organs of the tumor-bearing
mice were implanted into isologous mice. The organs
selected were liver, spleen, and kidney.
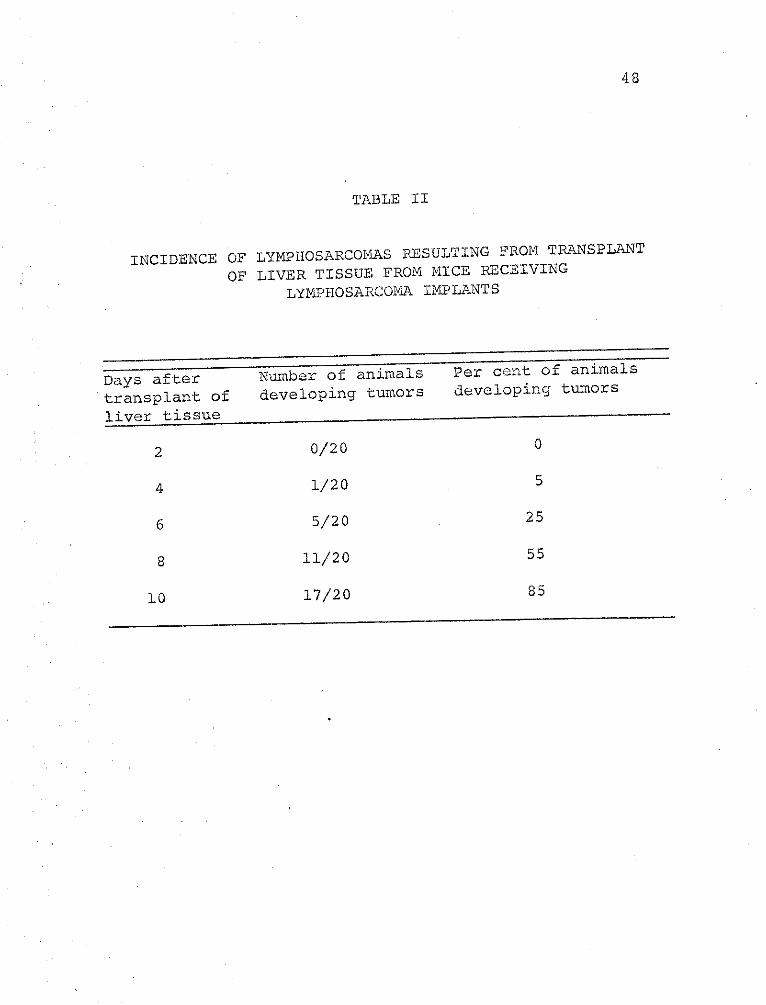
Table II shows that 6 days after implant, 25 per
cent of the recipients had developed a histologically
diagnosed lymphosarcoma. Ten days after implant, 85
per cent of the animals had developed a malignant tumor.
Three of the animals, after 10 days, did not have a tumor,
but one of these mice had a nodule that developed and
regressed. Tumor development, in these investigations,
is defined as a terminal malignancy. Normal liver,
transplanted into mice, did not produce a tumor, nor
did any of the control animals develop tumors spontaneously.
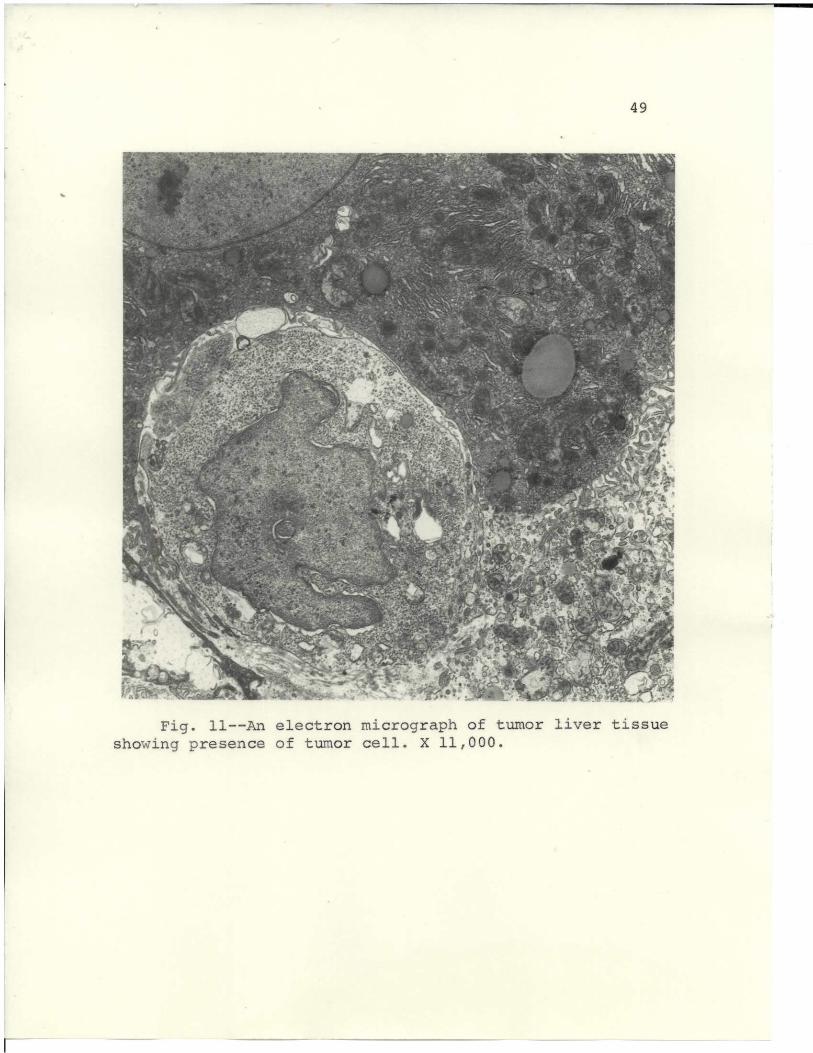
Liver tissues from tumor-bearing mice were examined
by electron microscopy. Four to five days after subdermal
tumor implants in mice, tumor cells appeared in the liver,
as seen in Fig. 1I. Several more days of tumor progression
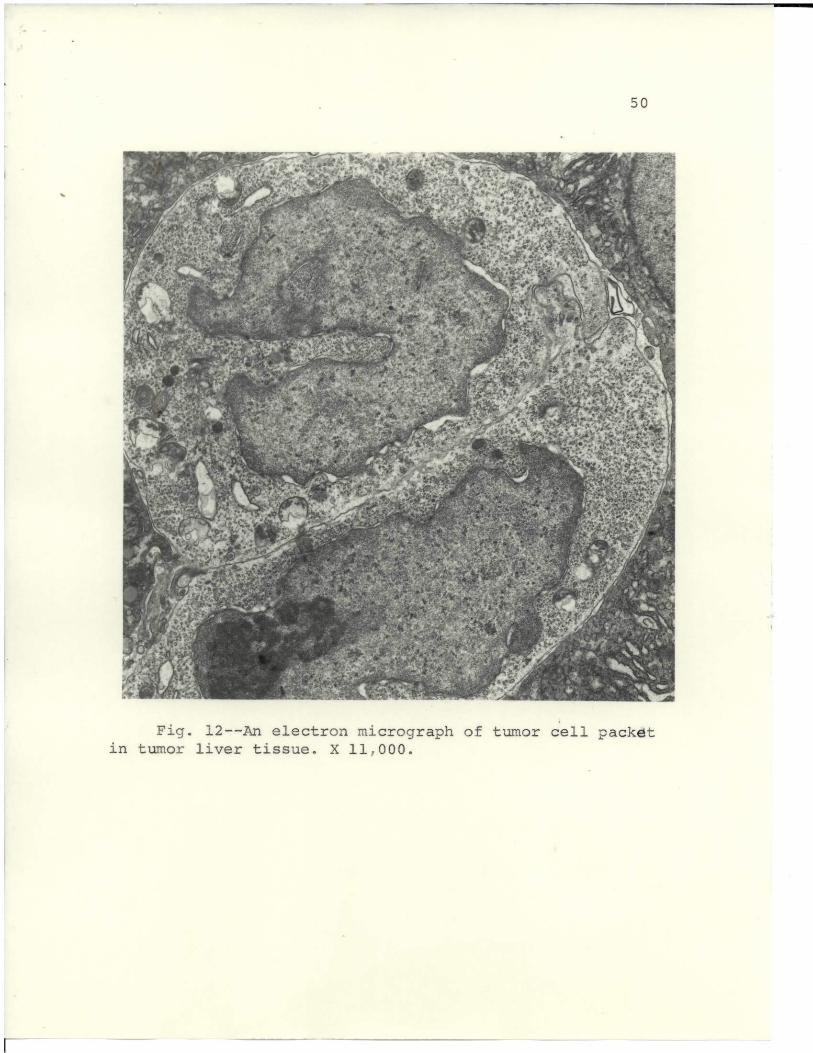
revealed foci of several tumor cells, as shown in Fig. 12.
The foci were not demonstratable by light microscopy when
the tissues were stained with hematoxylin and eosin.
43
TABLE II
INCIDENCE OF LYMPHOSARCOMAS RESULTING FROM TRANSPLANT
OF LIVER TISSUE FROM MICE RECEIVING
LYM.PHOSARCOA IMPLANTS
Das after Number of animals Per cent of animals
transplant of developing tumors developing tumors
liver tissue
2 0/20 0
4 1/20 5
6 5/20 25
8 11/20 55
8517/2010
49
'oil
~. & " 'j fI" fi..A , j ',a-"s:' Y"
. -17~
* A^' .. a.if', a 1 j 'f . 'a .
~ ' . . S i"' r z K . ,,F kj - .w.- . 9 i- e : ' .:--lt .t " l Ns tf"N ..f " .Io _ . " " "~fi:
140
\ .. f, i " - P *: , 4 's lS .x }, -' S"t.4 J r "!'ti-' a r " 1rC~i -. " ~a" _a 'L _
M K;c t .' k. h ft ' yt a=FC a
'4 , t 1 ~l'+r , i-tds _ ra j . f «I-}ypf,
Fig.a 11--An electron S " mi %/2 " !" ta- acogs.rp f uorlve isushowing" presen R, ce oJ ,.af f tumor- cell. ';,. X 11,000
-M
50
r+ 1 3, r i s a ;
G , '', , ' * A 'y,' " " F s "r SL. r'K'"" IWO 4 . "" iy R:"'; -'.t ,:
i4o ' *I' a" ti "e J s, 'it' ' ^'r f'i '' .. : ' f." j', i . "4.
'J" ' ". Y Y
i a- - _ .. : "r,. '. iia f ;
t j ; .. .re + r: t- r "M ht.t t, 3' 'i
q/ I xSP""'1 ."y' " pppS j y , r> ".'" - " irf ; p c"''All -4 -01
2 KKK!!l o i f . !a 1^l f a.a+ sl k+y t+'k4! '*r " :*: 3
".! ' ., ',. f . , " } 'J-fir .S ,' :" r {',} ' - " a "" ", ' ' 4"" f " " f
i 4r Gi " .: + 4.:. t. 7 At 'S. t ". } R .. .,a ,/ ityt' 4 ",
lR w "..M l.rf I: 1 " 1,Iti y - Ji'., .' ". '/ 7.. ".' -11f
f / ' ". y
, «d . , Y rs: , r' '. '+4 :; 1 : c ~' db .. ic G i~- "" r" . d.r . r , yyt it r , ; , f
"' ~. r ' " r< ' . "
*,' Y !:"' " Y' y ; . ,6 r i / . : f.1+ll,++ 'i . ' t ;1 y .a
fr '" 4 91' T Ia r , t ran t / r '.V Ys s t' 4 / It I 11 4 , ' a' r i! fL'tC..
r , y y b Y + +14. t " ./ x. a ;' "' f e e }Pr tiv w - AV ," ; .t ay/ :y s
g y i .' -".g, l ,j ,:/Ji''"f ~ct '"' i P "r . ""V t'- S"'- i ' J% T "IO,,jr {'- i. ' v /ste" M~ r ,f f ,.; "1' .j; " , _" ' ' _ t-'"J' 4 ''+ R1" / at ' r {}.i {. .0 i. " 4; . y r r y > ' ."," .. , !M . ." - 1} '; ' t o r ". t f ' i , . 9 > :.[
p ' i "a r T '.," ' r." . : . r .,, 3: ' .- ,i.s A.?j 4w, ,.," ^ r ,e. 4 3'" f-r ; ' t'>=as'p.
,j I ,,. " ;r' , r<' i. "" VIN "h !;' I f..+}{.d r. .' y " "E a t,..i 41 r,. W' *' '-'tart Ar
jf*i !. ! r IF Gr' Y t a 1 A".}
" fr fk ' . , - 4e ' y. :R !s, ) v < '' " .. F Mt: r 'i(/ Yj+ r" 7
f fa ' 'E1C, ", ) l} 4 fir! 3 L! /,j". f{'eF"P' %a3f 'i'- } b \ I n"
'Ilk V III,
116
: ,u . ' = ". "y :"Y 1~ YST .' ,t, .;1 3 '. 1 Yr . f ,r yr t "Par , ;_
v r C 1 I f.r _ ,"o .R ' T Y ' a 'r .. "l' , . ' ".v a" . k" a - N!R( A" ^"i+ _ T; ,d .? '"3 ,"." / _r I
r 1 ea k a' ,., " - , t . *' "'4t ". F :; 'Y.:N. ,f."rY i !"i ' - : . - " 4 " y". . r =, ri 1 _
A . t Sr .. } r' s r G 'a .* ." .. -"'; . - + 4 a3i Ir S - i, " al ,
r' " i" a' s a « '~t :"T b +' ' f , k." ,Y' , ' .Y a , s to '"+ V' ' i y y
', t Y' /" :. ' .4 x i r+x 's. r -,yam "" . t4- :. ,t" w.,,r r ;''h ti, r 'b "Y T t
.' "f t .'> " ' ' ,. i" .. s' '' 4 rs , . . ,. i - af; it ,s ue :"P cJ! y.
y R .. i , a+ , X i , a3 " F .r1 " f , 1L + i 1,, a 7" f
s3 ' y. a (i/,w /y1 /( .'r i i t. i ti4 a q a" ' ",.e." ' f ~ , -! - tt IrYi , 's (" " f, :9.'
1 .;/ '"e"-1 + _ : !' *r .w + (!, " " 9 ja^. 1 +C I" i. W . 4,.j - y , ! . awr+ .
f r y P 3 Y' r?'=' +v'} i8" a ' 'A * " - y^ i!. a" .4 'f ._' 3 ,,,."74a'L'",t!"r' ' " !' y . a( .Y r j'' .A - 3 "R '' .ef T'. - " 1i{ ~ " !f - k { t /' 1 , 3 - .' !(nL y A. ,.i - t - ' '1 S f Sl.- ; ' Li !'"i, .. ' > Yy :r"aF - e.r -i. '
J d ': .i r, f.t; a : "' "'S3 S .}.. f. MS".-'? Y ""F' t" - i' f"' . .yr R f3 y ir + 'Z- .V; 1 ; K T 1S
41 Sh y f r ; 4 " .a , y:; .i Sy "- f 3 vt i 4f ! stpA Qs 1 >, j+ ' r i 1 rl 'f c( AJ. K' : o
«
"- S .'r"t a. t ;+ i . ' '"ti fe, "f "r t'. ey. . t " r ,p. . na " , '" }, I +l Ar ','tii.."
"' ' ~l i'_ '
.i " r".
" v / ," , ' " ,y!'
i:II 'r, 1"e ' '
,7' "4
'
R
-, , I
v' y
E": i .t
," t,. , { ! 1 siR - Tl ". r t, ; r " j r " : y r " "tr'. .' : Iy 'M -a +'- r"-rjk ' j( " 1 jam r'
1 r' id' v: " i1., r t/t /'; t er M.. ,. ' "g " "s 'R r "1 t' ''a _ '"t T"
-! ! "y - ' " i i r'y, ' ' , ,+' Y'' ~"" r .# v 'r, . r " . "G: F-R :t ,r" ~i .~' f e t , . , +Ib ; ",r NC' " -i, y
i '.rw " ""S f a .r 5 . "1 Y r t "( 'N1 +t r t , "," - -S" y r, " i ti, .{f k " "= " " t +k .
" i >lr. Qom- Ar'r y+/ " i.. ' ,a"_ i 9 yy ,.yam. a ? r -, y 7' i.' t fi " .r.., , M' a'
_ _ f ."' MR «, a ? 4" " - . csr N Z'T f'4 -," a. 1 ^ ,,. .1r. r+F " ' j ' j"t 7.%'- tt t I' , jr
f . r 3. . ! . - -3 r, aM ' - ,1. ,.r' it d"" . -" ;a. t R! J a.4 " " al
L a.. r "1l'4 . Z'G. I. ' L ".. .. q ty'! +tX ,p a'R 1 ,r r
i f [4 vr. y4Y ,,. ,;,f ,ti i y.f(i ' "'", " I":, 3'F rte. 3 f? .'s-a ' # t} i +r
A . a! R I a . r + ',."1} ':' 3 '3 ". f' 3 " . .n - _'r >" .' "r o ' 1t
' i "vr %i: ' ". .fp'.* e f "if , +"ar "."- r.. y kRdb..'7: _', i "' >! 'R"1I:"
It'llyti M y M.-} n+ltrt L r r " ffY . y?4;' F i+t t A^yf.75" s, t4 l e i 4'M
a i'L" y'" . r , '',N' '.t i 4' Il 4R ,cat r , ' 71 ' a T.K + " r
;, .' , ,i' f. " 1 x, *+ q !1 . j,.p yy .r SFr; }N .^ .q' rr ti'k ,",.a' ! a' :,
,, t f , aa' MMM {+y L"-: ' ' .f , /.i- , " I, f.1 . 5" " ; y R. ; " '" v /f' s , ; ' 9r . +71",.+# -
+. + Tj Ft a " J w 4 r 4. A 1,a"J- " qy _ p }"
"+ ~f J r > 'r. "n " " . 4. - r "M R ' : a6''- Pf ' . *', ., .. , r Ti ti "1. " 1" 1 1.4 ' e I
y!r , 1 .i" . r " .a . .. .y47ni 1. !...,t rF7 ? . F Si "yf yC. y' f .t.
:.rte J v - t' a =T ryr w , " < + Y-'LYi . + - Jlf'. r'"'r i ""^'' 4" " 7i'-" j ri'' , s,,; i - q
~'. " ,". ! '/!T. i i r. M ;s .
i y1 " A OA f N i j"... -. 'Z, yf ," , 4 t. N 414%1." i'~ !w1 YcP '. k "!aE ,, 3, . ^ f. . " r . r th 'r
Eb y +' R A. t y 1 ., !t wit , f: /:. {_ yT ,r . 2 7 11 ti. '. "i i/ til"." a SI t i
fq+f i'', ,R t}'(- !/ ' LL 4 i^_ .r _ t' .",r '" .i:.+. ^,.., r .. ? .: a+w ' . .'+KM1 1 x:.+ " t , 'c r-s Y. ,T } y "" t , .pY" "Jf
eJ " ^ T+ i..I 7"i,. '6' I -"" ""+''!1"y + it: i, .t,,a ," f"4' ;7 f",S i '! "" Q " : -fl' , r a t + r. i
10
S ;.f r , I " y+Rrai r ,lq' .s , '' v..Clthy'S iii',: l 3;? ir T'" .+i .i;iL.' .5, ,F .y
-. t_" ' c "+ V"' Y1 {v yF'' 4.y : +f+1 k '' .3, . , ,.. , .. j p,
,: ! " r , .. R f '',F, .} ' f .4. ,J v R: '~""+ + .a "'s"i. 1 a'drT, _ 1'.f; , u+ " I S I.9 i' I"- G: YN{ ("': 's,.,,t ,gf i / ",[ a
"!" {,r
"
*
, :art it " i - . !"T a"r4 ,; / wY " %NY' y "F .T+/ pf.a a .,. ti , "
" "I ! e . V " a:: "-N0, ;:rte, .%" .ryr, '" *"y ry .. , 7 " yet , '" ' '!t-f 411" ate r ,
q x .!{ T 4, "NNi i, e , :"'I < :4+. -i.y ' . ra: qf, { ,' , a.. !v 4 r:
r a, w 5 r . "R'V 3 r- " t 4 rf . :. , f ': ! jy4 Y-'b ur ,I + J % .' _' . "i A .' " , ,.
. ... :Jf l+C. .+Y;s ri',A''=; iY k" " r'' -'" '.,i . tZ. a ' s {i1a, r -' ' ., ' { 4, , " - t, 'r r"' r
, /- . "a ., ,.,. 4P. .. , ^J :Y' 4 ''ir > 'R 's .*' rej.. '7' .a T Y(ar'. DI, -
;f *" > ?rte * 'P ' , _ , ut -f ' _ ' "' ' ,sr 5~ " L a f" 'J %'';j* "' " .
- }, Ct f. s s : .dr "- """ " y.," yttiL M i iy'. 1 i 'E.4 . , //'r r. n -
ILL
'..S
$'
,_ ;y>r: aZ '4' Tt ' H i .:' "" .r fa., / 9 411'' r ' "I.. " a -' ; 'P';? - ;' '.4b
3 t
f t" Y i
. d _.. ..:}' '- - *f-. -o .'^,wM r R " r .. TIL f L + (r
ry~ g r f' r % ~' r g*T'hjfI& JO&:..a
"4 '/ r vim t A y . tF" .+t '.WK , ,i .a , .\ ,(,;" .' i " " t " j I ' y
- i' + l \ _.} l,7 _ "1r ,r a 'L"i i" .Y ny t , " ' 1 " TA" ?
Fig. 12--An electron micrograph of tumor cell packet
in tumor liver tissue. X 117000*
-M

51
Liver tissues, used in the results 'of Table II were
taken from animals with a 0-day tumor. A tumor of this
duration had many tumor cell foci, which evidently
developed into lymphosarcoma when implanted into isologous
mice.
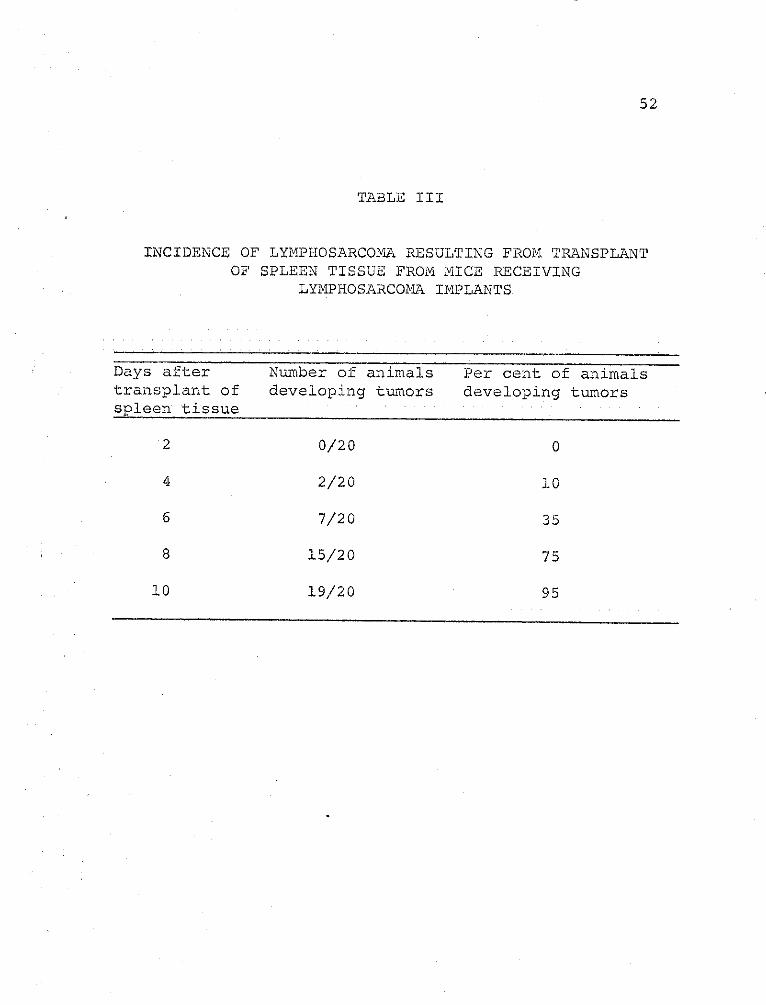
Table III gives the incidence of tumor development
with spleen tissues taken from a tumor-bearing animal.
A high percentage of animals developed tumors by the
eighth day, 75 per cent as compared to 55 per cent found
with liver tumor tissue (Table II). The spleen tissues
gave a consistently high incidence of tumor, almost as
high as that of the tumor tissues. The 10-day tumor
spleen tissues, when examined microscopically, were
pathologically diagnosed as having myeloid metaplasia.
Normal spleen tissue did not produce a lymphosarcoma in
any recipient animal.
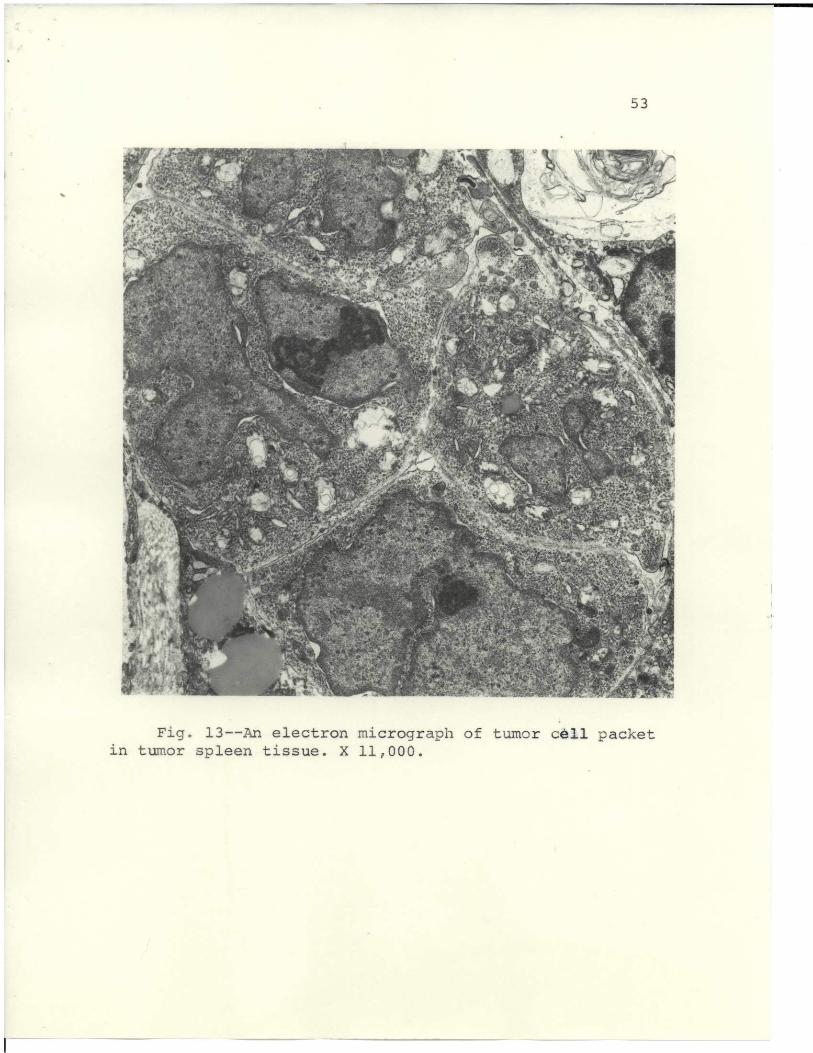
When the spleen tissues were examined ultrastructurally,
specimens showed that the tumor cell foci were present
and resembled those in liver (Fig. 12). In Fig. 13, the
foci are more numerous in the spleen, and this may have
contributed to its high incidence of tumor production.
52
TABLE III
INCIDENCE OF LYMPHOSARCOMA RESULTING FROM TRANSPLANTOF SPLEEN TISSUE FROM MICE RECEIVING
LYMPHOSARCOMA IMPLANTS
Days after Number of animals Per cent of animalstransplant of developing tumors developing tumorsspleen tissue
2 0/20 0
4 2/20 10
6 7/20 35
8 15/20 75
10 19/20 95
53
I,' ~%
">" ' t .~ ft+p'^:b li' JA ir .";' s4. t i
"rNH ' !" A 1? . K}^ } y'1 ; , ."i.}', " 1T S[ I i"i
li " .Y 3, " t . S - r. <'r + 'hr A 4 r ." iI" v " " "
e t1 f "t r E t~ 'r ; ,:}# A N~$
, ., - .. . 'te. . ;,'AA <,~ i. '*
! : i « t Y G~,p~ F ,y~ .i oE"" "- , + % ; "
.~ " " M " '-fv " ,' ' 'S l :1'*vA ,. .) i"r''s" :1"" ,^ ~t ' ; L .y _*" " '1' ,A V it r" a y Gt :. ' " A. , .. Ia
-4 4 A {r4'~ t yt '. :w _' ',, r -, ' _ ? Fn y L _
! i " t - "" , .h s r", t -'": ' i ~ . 7 i" a t _ Aic_ -e~. " rq,. r ,t " k y rt "' 14Jr
-"_ . ~f J r E" r ,-y~~ .~V W y . vr' } low-A5 .. ' -yf t er <,3~.i ^ t;"-
f X; ,'." jd , "r A l y in r '" '4 , , ,. r "e 0 0. .', Y " k ,1.i "a . _ .". f : Ay ' ) :. a_ ~ J ,tt4 Y', i" ':E4R. . > ^ : t' ,. L - .!f _t' ylt}t } "37 "r r _ I i-
r o. " 't ttFa }} .w" j 'iA( A" : 1 .as .i'n L ,.,": ' ,,jt~j=- ."!'" o ." ., .J-' ^',.A
kk ,t r 7
' fa, a {:k.. : Q , . .it ." s ." YT~y X h : . r'
7s~"F P r;,' f" .: " ,Fi. 13--nyelctro micrgrapof'umor l acke
in tumor spleen tissue. X 11,000. .7.

54
Cells resembling the Stermberg-Reed cell, Fig. 14, were
frequently found in tumor spleen tissues. Large, multi-
nucleated cells were found in the spleen 8 to 10 days after
tumor implant.
Table IV gives production of tumors from tumor kidney
implants. The kidney tissues appeared histologically
normal when viewed by light and electron microscopy, but
passage of the tissue produced a tumor in 70 per cent of
the animals 12 days after implant. Two of the mice had
regressive tumors. Kidney tissue from a normal mouse did
not induce a tumor.
Tumor cell suspensions were prepared enzymatically
(trypsin) from an excised tumor, diluted to the desired
number of cells per ml, and injected subdermally into mice.
Table V shows that 103 cells/ml induced tumors in only
10 per cent of the animals. At higher concentrations of
cells, 108 cells/ml, 90 per cent of the mice developed
tumors 16 days after transplant.
Tumor cell suspensions prepared physically gave
consistently higher activity for tumor initiation than
did enzymatically prepared tumor cell suspensions. The
trypsin may have inactivated some of the transforming
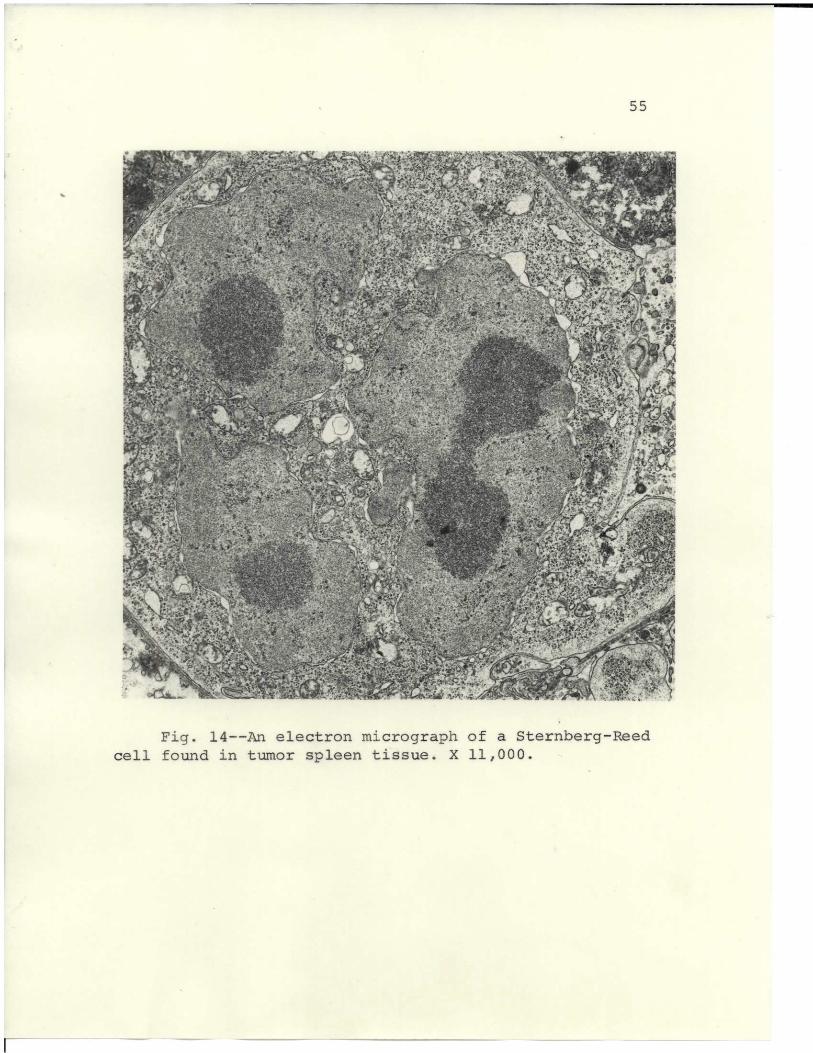
55
g1 Tr''tf45Stt ~~ V ~ A',f , '" t 4 > ijj yI 4 ' tN 4Ca+. ' ft"1 1'',\_ "f, ,i.7[ ^ .r
arv " r; j .? .. .{ y I y 2 A 1't AI 9.
.. : y . ,+ p .' :, i "'} + i ,s: ' E-'~ "' i.' r , s f. , '.. .' a "f .
^f Y "" tY ' '- .r.4z 4 -'Y jrn't t' Ja , '"V : i ''r , , ": w"- :ty " ." f 5 .: ,I .r:v" '4 W - ." y _ v " " - a' t : ' ' ' ' ;. _ "J 'tt " r Z ' x .-f u t * i t'j.:4 7yr 1, '. f it 'toI-A4 : 'f.lf; , t : " 4.tv}( ygg
y ~ ~ ~ ~ ~ ~ ~ ~ ~ ' t . . .. 3 " . "> ."" 1a.Aw J iJ "" ' c "1t / ')
t tJi' jil. 1 a:jw Y.. ;f ^..;.:-y -'1f iai e ''4 ,t,.< " [*
'7 ' r "tt {G y+' ;> h> ~ 4rA i 4t 'M . 1 "j, h' ' t" 'S l. '1 ' ' ~ L!' i -- t \ f~ 4t '<
$ *'i } T' ' " ""4!- !''.t ' *-=1 3"r'. _ t 4+""! ." & y "- , .
',( tSp""" w. yI .a """ v '"" 'D S t} . m y gm.r . : ^ Pj 4 . " Z . . $ ;
ti : i '{ ;r f., V f, ' v i 't - " : f' - s t " . "T" . ."a'- & !:1- i s.-c ." t r.:.y. } - s" c -7't . {.j" r };..r 41S "'_4%i v , f" > 6
7 " _ r a
." '!). : ary ;, ,"r"J r -6 i <- :Y ' i Y,: I" s - * #7 ! ' " Yi =.J _" 4 c t % L ( " i t' 'i . .t- 4 A K. r ' i , s , s f' - ' : : . r rj ' ; i " ' ; : _, '-'. " . .li . 4 , " "' f s' i-: y ' r'-S
X .':' ' 1 r" _"a" ' v. i s , ', '" ~ i "Lp "rk _ -::
'r ' ..;v . .. 4 1'J.); r Yt1'' ." .". yw " ^ "c- +_ , Y%. j, .' S '" rti ~ '"', r r . . , y .'' -,,. '"s " Yc " s' " . .i ',='s' . ,4t y-F } .. / " " , ; l'w " L!"" y4iriL. .. ~r ir 4Ar'" 4t'. s'~ i" ' 0 . ir r~a r .. ~~'; : I':, fhy. , .
,'~~~~~~ ~ ~ ~ "?t I Vrrf . "'.i " { "{
i 7?~St ,)h'j1s A1j Ati
_.r24 T h Vr"t< 9 t t " - . -t 1 t " -r 7 * .4g t k * K '"" " ' ar t t'' ':Y s + 4%t. T ' : " " )* .''ftk . t ' $ ' f 1' C '. ."~ N4 : ! ' .'g ' .-'y& -- "i . y r f '
( : -. C "1 . "" '. " FI . 1r y +s "/. r4t'. L'."t. ! ' fr:":x.}' "_: e. . : iA'c jh vt1"r /i"b; 't't's -r'. a r . - S "~v7 .:t
Fig :"14 An eliectron micrograph o...."{,, :; ft-" f a S teinberg-Reedcell found in tumort ":. yfs! { "spleen t" f ."ri ssueyI X 11,000,, .. '' .': ' J 6" .n

56
TABLE IV
INCIDENCE OF LYMPHOSARCOMA RESULTING FROM TRANSPLANTOF KIDNEY TISSUE FROM MICE RECEIVING
LYMPHOSARCOMA IMPLANTS
Days after Number of animals Per cent of animalstransplant of developing tumors developing tumorskidney tissue
2 0/20 0
4 0/20 0
6 3/20 15
8 7/20 35
10 13/20 65
12 14/20 70
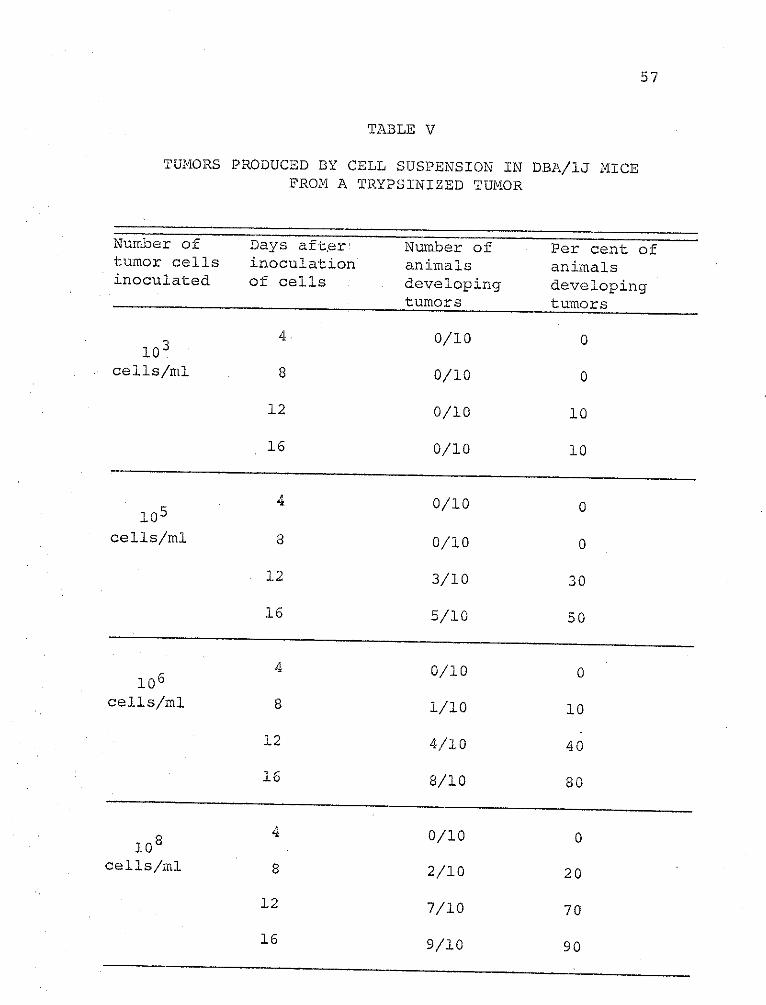
57
TABLE V
TUMORS PRODUCED BY CELL SUSPENSION INFROM A TRYPSINIZED TUMOR
DBA/IJ MICE
Number of Days after Number of Per cent oftumor cells inoculation animals animalsinoculated of cells developing developing
tumors tumors
0/10 0
cells/ml 8 0/10 0
12 0/10 10
16 0/10 10
4 0/10 0105
cells/ml 8 0/10 0
L2 3/10 30
16 5/10 50
104 0/10 0
cells/ml 8 1/10 10
12 4/10 40
16 8/10 80
108cells/ml
4id
8
12
16
0/10
2/10
7/10
9/10
0
20
70
90

58
activity. The cell suspension of 105 cells/ml gave a
per cent tumor development comparable to 108 cells/ml
trypsinized cell suspensions. When both cell suspensions
were examined ultrastructurally, there were no observable
differences revealed in their fine structure.
Cell-free preparations of tumor tissue were used to
inoculate mice subdermally for induction of tumor. The
suspension was centrifuged at 3000 X g for 10 min, and
then 1.0 ml of the supernatant was injected subdermally
into the animals. Tumor appeared in 48 per cent of the
mice. The superatant, when centrifuged at 8,000 X g for
30 min and inoculated into mice, initiated tumors in 23
per cent of the animals. A third centrifugation of 30,000
X g and inoculation produced no tumors. When the solutions
were treated with RNAase, tumor production was inhibited.
But DNAase did not inhibit production. The nuclease
treatments indicated that the transforming "factor" is
RNA, or a ribonucleoprotein.
Isolated mitochondria suspensions from tumor tissues
were injected subdermally into mice. After 6 months,
no tumors were observed in the recipients. Investigations

59
of mitochondria-transferring virus and virosomes have
been reported (48, 58).
Tumor nuclei, when transplanted into mice, produced
tumors in 90 per cent of the animals 6 days after implant.
Nuclei from livers of tumor-bearing mice were transplanted
into normal recipients, and 85 per cent had tumors 8 days
after implant.
The nuclei from the tumor disrupted, and the
membrane-Cree sample was incubated with RNase or DNase.
The suspension treat d with RNase did not induce tumor
formation, but the suspension incubated with DNase produced
tumors in 83 per cent of the mice. On the other hand,
liver nuclei gave an opposite effect. DNase treatment
inhibited tumor production, whereas RNase did not. Tumors
appeared in 56 per cent of the RNase-treated suspensions.
Tumor cell extracts, treated with heating to 60-70 C
for 10-30 min, then treated with iodine or formalin
solutions, and injected into mice ,induced tumor in 55
per cent of the animals tested. If the treated solutions
were further treated with RNase, the induction of tumor
was inhibited. DNase did not inhibit.

60
Table I shows tpat after 16 days, 5 per cent of
+t.4 mice which ha received diffusion chambers had tumors,
and that the percentage increased to 80 per cent after 22
days. The tumor-cell suspension originally added to the
chamber was 2 x lo6 cells/mi. During the first 10 days,
the viable count in the chamber decreased to 104 cells/mI,
but by the fifteenth day increased to 106 cells/ml. The
viable cells were found in close contact with the Millipore
filters adjacent to the host's tissues.
The diffusion-chamber methods were developed to
permit grafts or cellsto survive and grow when separated
from host tissues by means of porous but cell-impenetrable
filters (32). In some of the investigations reported earlier,
the average fi'-er pore size covered a wide range (3, 61),
but none were in the 25 my range. Millipore filters used
in this study would not allow cell-to-cell contact, a fact
confirmed by electron microscopy. The average pore size
of Millipore filter (25 mpa) used in these experiments
was less than the smallest known mouse tumor viruses
(60-i5O mp).
Liver and spleen tissues from tumor mice were examined
by electron microscopy. The tumor was induced by diffusion
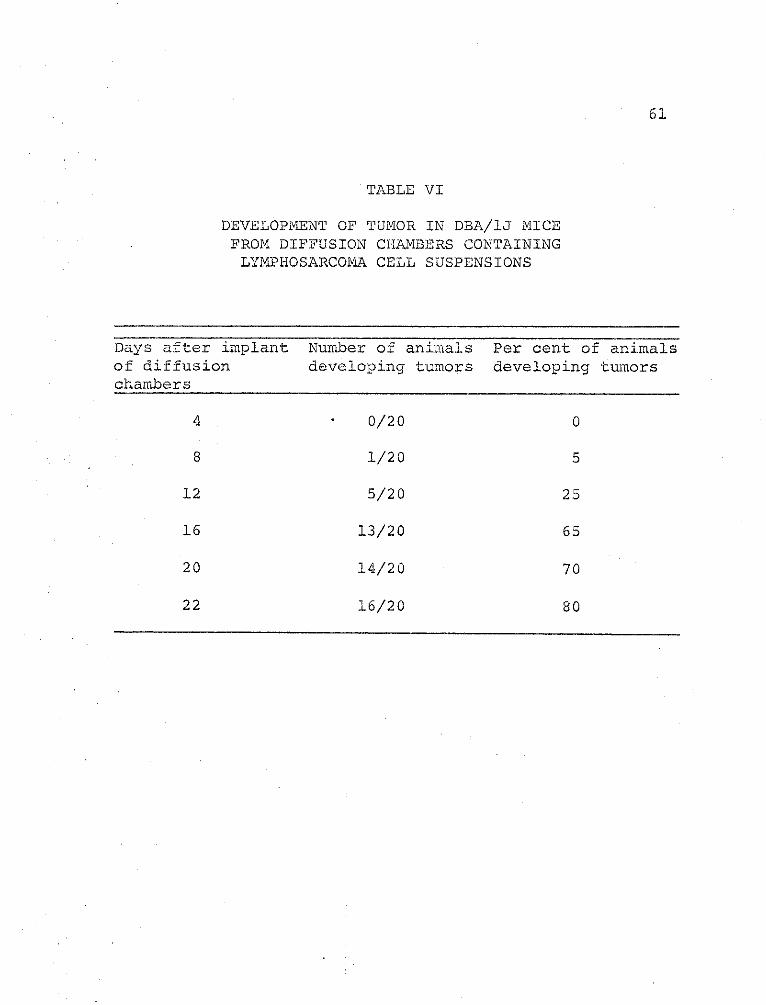
61
TABLE VI
DEVELOPMENT OF TUMOR IN DBA/IJ MICE
FROM DIFFUSION CHAMBERS CONTAINING
LYMPHOSARCOMlA CELL SUSPENSIONS
Days after implant Number of animalsof diffusion developing tumorschambers
4
8
12
16
20
22
0/20
1/20
5/20
13/20
14/20
16/20
Per cent of animalsdeveloping tumors
0
5
25
65
70
80
........

62
chamber implants. Tumor cells like those found in
Figs. ll, 12, and 13 were observed in the tissues. The
chambers were impenetrable to the tumor cells; therefore
the transforming "factor" diffused from the chamber to
infect and transform cells. Where the transformation
occurred is not known. Perhaps transformation was performed
at a site adjacent to the chamber, and then the transformed
cells metastasized to the soil tissue. It is even possible
that the "factor" entered the circulation and transformed
cells at the tissue site.
Tumor cells were cultivated in suspended-cell
cultures through several passages. The primary cell
inoculum was 106 cells/ml, which increased to 2.6 x 106
cells/ml during the first 8 hr growth. Cells not transferred
to fresh medium after 10 hr primary growth died within
18-24 hr. Cells transferred to new media after 10 hr
growth survived to 60 hr before decreasing in viability.
Cells carried through to the fourth transfer failed to
survive for more than a few hours. The pH of the medium
shifted rapidly toward the acid within the first few hours,
which indicated a high metabolic rate. Later transfers did
not give pronounced pH shifts.

63
Viable cell counts. from the parabiotic. chamber
indicated an increase from 100 cells/ml to 3 x 106 cells/ml
within 12 hr after initial inoculum of cells. During the
first 12 hr, normal liver cells increased to 1.2 x 106
cells/ml, and normal spleen increased to 1.4 x 106 cells/ml.
After 24 hr, the viable cell count decreased in the liver
cell suspension from 1.2 x 106 to 105 cells/mI, and the
spleen cell culture remained nearly the same at 1.2 x
106 cells/ml.
Cell samples of transformed liver or spleen cells
(106 cells/ml) were inoculated subdermally into mice.
The animals receiving 12-hr transformed liver cells or
spleen cells developed tumors in 65 per cent of the
recipients 10 days after transplant. Cell samples taken
after 24 hr produced tumors in 45 per cent of the animals.
The Millipore filter separating the parabiotic
chambers had an average pore size of 25 np. The transforming
"factor" was smaller than known tumor virions.
The experiments in this investigation have shown the
transforming "factor" to be a RNA or a ribonucleoprotein
of molecular size. RNA from purified avian and mouse

64
oncogenic;viruses have approximately the same molecular
weight. There seems to be very little difference between
avian and marine RNA in regard to base composition, and
there is no difference between those avian viruses;
this confirms their close relationship (75, 76).
Investigations have revealed that heat and dimethyl
sulfoxide (DMSO), two agents able to break hydrogen bonds
between bases, can break the 65-70S RNA of Rous sarcoma
virus (RSV) into pieces of similar size, 2.5 x 106 daltons;
this suggested that the large RNA molecule consists of
four (or possibly five) subunits held together by hydrogen
bonds (1.3, 26, 65, 30).
Montagneir, et al. (65) found that the target size of
the transforming capacity of RSV, in X-ray inactivation
experiments, was about the same as that found in tobacco
mosaic virus (TMV) infectivity (53). Moreover, the
oncogenic capacity of RSV may be coded by a RNA subunit
the size of TMV RNA, which is approximately 2.5 x 106
daltons.
Duesberg (26) reported 5-6 subunits by electrophoretic
mobility for influenza virus RNA, and each subunit appeared
to have a corresponding double-stranded replicative form.

6.5
Hence, if RNA subunits, obtained with DMS or by some
other means, replicate independently, and if each actually
carries distinct information, one may expect to find
recombination between RNA oncogenic viruses.
Viral PNAs have an interesting feature in their
capacity to hybridize with cellular DNA. Temin (90, 91)
reported that RSV-RNA hybridized with DNA from RSV infected
cells. He concluded that viral RNA could be synthesizedon a viral RNA-primed DNA template. Later investigations
showed that RNA annealed equally well with DNA from
transformed and normal cells (41). Harel, et al. (41)
showed the hybridized RNA, consisting of a single piece
about 7 x 105 daltons, with a high content of adenine.
This finding demonstrated the existence of a segment of
viral RNA, rich in adenine, that could anneal with homol-
ogous thymine-rich region (s) of cell DNA existing prior
to infection.
Shibley, et al. (84) reported that leukemia could be
induced in mice by nonneutralizeable suspensions of viral
nucleoids obtained by ether extraction from plasma or
viremic mice, and also from multiple in vitro passages.

66
smaller than mammary tumor virus (MTV) can carry the
viral activity (64, 66, 68). These last data suggest,
even to a critical mind, that subviral components may
be actually infectious.
The results with a murine lymphosarcoma presented in
this investigation indicate that the transforming "factor"
from the tuior is an RNA. However, tumor liver or tumor
spleen indicated that DNA is the transforming agent.
One explanation may be that the genome of the virus is
incorporated into the tumor tissue as a provirus. The
provirus then produces a viral genome of RNA capable of
transforming other cells. The transformed cells have
RNA-dependent DNA polymerase to produce DNA complement
of the infectious RNA. But the transformed cells lack the
ability to incorporate the DNA into their genetic machinery;
thus an infectious DNA remains in the cells. For instance,
when the transformed tissue was used for transplants, the
surrounding tissue was transformed. The surrounding
transformed tissue then incorporated the infectious DNA
into its genome as a provirus capable of producing infectious
RNA as found in the tumor tissue.

67
Variations were neither found in nor isolated from
any of th tissues. of the tumor-befring mice used for
study. Perhaps, under certain conditions, the virus
has lost its ability to form a protein coat; consequently,
it would not be demonstrable as a virion. Therefore the
transforming and tumor-inducing agent was not considered
to be a vi-rion, but an infectious PIA or a segment of
RNA oncogenic viral genome.

CHAPTER IV
SUMMARY
A search was made to find an agent that maintains
and induces a ly mphosarcoma in DBA/IJ mice and transforms
cells in vivo and ia vitro. The ultrastructures of the
transplantable tumors and the transformed cells were
examined for cellular alterations and the presence of
visions. Virions were not demonstrable, and the trans-
forming "factor" was determined to be an infectious RNA
or a nucleoprotein. The agent was able to pass through
Millipore filters (25 m) employed on diffusion chambers
and parabiotic chambers to transform cells in vivo and
in vitro. When transplanted into normal mice, the
transformed cell induced a lymphosarcoma.
This evidence indicated that it is not necessary to
have a complete virion to transform a mammalian cell.
Thus the "factor" in cell transformation may be an
incomplete vision which has lost its ability to develop
a protein coat.
68

LITERATURE CITED
1. Abercrombie, M., and Ambrose, E.J. 1962. The surfaceproperties of cancer cells: A review. CancerRes. 22:525-548.
2. Abrams, J.A. 1969. The degeneration of implanted tumortissue in mice. Unpublished Master's Thesis,Department of Biology, North Texas State University,Denton, Texas.
3. Achong, B.G., and Epstein , M.A. 1966. Fine structure ofthe Burkitt tumor. J. Nat. Cancer Inst. 36:887-894.
4. Alexander, H.E. , Koch, G., Mountain, I.M. , Sprunt, K.,and Damme, 0. von. 1958. Infectivity of ribonucleicacid of polio virus on HeLa cell monolayers.Virology 5:172-187.
5. Algire, G.H. , Weaver, J.M. , and Prehn, R.T. 1954.Growth of cells in vivo in diffusion chambers.I. Survival of homografts in immunized mice.J. Nat. Cancer Inst. 15:493-507.
6. Ashenazi, A., and Melnick, J.L. 1962. Induced latentinfection of monkeys with vacuolating SV-40papova virus. Virus in kidneys and urine.Proc. Soc. Exp. Biol. N.Y. 111:367-372.
7. Basilico, C., and DiMayorca, G. 1965. Radiation targetsize of the lytic and the transforming ability ofpolyoma virus. Proc. Nat. Acad. Sci,. 54:125-127.
8. Benjamin, T.L. 1965. Relative target sizes for theinactivation of the transforming and reproductiveabilities of polyoma virus. Proc. Nat. Acad. Sci.54:121-124.
69

70
9. .Bernard, W., and Granboulan, N. 1963. The finestructure of the cancer nucleus. Exptl. CellRes. Supply. 9:19-31.
10. Bielka, H., and Graffi A. 1959. Untersuchungeniber die leukamogene wirkung von nucleinsaurenaus virusindizierten leukamiegewebe. Acta.Biol. Med. Germ. 3:515-521.
11. Bittner, J.J. 1935. The breeding behavior and tumorincidence of an inbred albino strain of mice.Am. J. Cancer 25:113-121.
12. Black, P.H., and Rowe, W.P. 1964. Viral studies ofSV40 tumorigenesis in hamsters. J. Nat. CancerInst. 32:253-265.
13. Blair, C.D., and Duesberg, P.H. 1968. Structure ofRauscher mouse leukemia virus RNA. Nature 220:396-399.
14. Borrel, A. 1907. Le problem du sander. Inst.Pasteur Bull. 5:497-545.
15. Brown, F., and Stewart, D.L. 1959. Studies withinfective ribonucleic acid from tissues and cellcultures infected with the virus of foot andmouth disease. Virology 7:408-413.
'16. Colter, I.S., Bird, H.H., and Brown, R.A. 1957.Infectivity of ribonucleic acid from Ehrlichascites tumor cells infected with Mengo encephalitis.Nature 179:859-861.
17. Dalton, J.A. 1964. An electron microscopical studyof a series of chemically induced hepatomas. In:P. Emmelot and 0. Muhlbock, eds: Cellular ControlMechanisms and Cancer. Elsevir Pub. Co., NewYork. p.211.
18. , and Haguenau, F. 1968. The nucleus.Academic Press, New York. 244 p.

71
19. Defendi, V., and Gasic, G. 1963.' Surface mucopoly-saccharides of polyoma virus transformed cells.J. Cellular Comp. Physiol. 62:23-28.
20. Delarus, J., Mignol, J., and Caulet, T. 1971.Vascularization and the appearance of malignanttumors: An experimental study. Cancer (Paris)Bull. 56(4):381-396.
21. Diderholm, H., Berg, R., and Wesslen, T. 1966.Transformation of rat and guinea-pig cells invitro by SV40, and the transplantability of thetransformed cells. Int. J. Cancer 1:139-148.
22. DiMayorca, G.A., Eddy, B.E., Stewart, S.E., Hester,W. S. , Freind, C., and Bendich, A. 1959.Isolation of infectious deoxyribonucleic acidfrom S.E. polyoma-infected tissue culture. Proc.Nat. Acad. Sci. 12:1805-1812.
23. Dipple, A., Lawyley, P.D., and Brooks, P. 1968.Theory of tumour initiation of chemical carcinogens:Dependence of activity on sturcture of ultimatecarcinogen. Europ. J. Cancer 4:493-506.
24. Dmochoski, L. 1963. The search for human tumourviruses. Texas Rep. Biol. Med. 21:113-126.
25. , Grey, C.E., and Gross, L. 1959. Studiesof submicroscopic structure of parotid glandtumors of mice. Acta Un. Int. Cancer 15:791-798.
26. Duesberg, P. H. 1968. Physical properties of Roussarcoma virus RNA. Proc. Nat. Acad. Sci.60 : 1511-1518.
27. Eddy, B.E. 1964. Simian virus 40 (SV40): An Oncogenicvirus. Prog. Exp. Tum. Res. 4:1-26..
28. , Borman, G.S., Grubbs, G.E., and Young, R.D.1962. Identification of the oncogenic substance inrhensus monkey kidney cell cultures as simianvirus 40. Virology 17:65-75.

29. Ellermann, V. , and Bang, 0. 1908. Experimentelleleukame bet Hubnern. Zbl. Bakt..46:595.
30. Erikson, R.UL. 1969. Studies on the RNA from avianmyeloblastosis virus. Virology 37:124-131.
31. Fernandes, MV., and Moorhead, P. S. 1965. Tran formationof African green monkey kidney cultures infectedwith simian vacuolating virus (SV40). Tex. Rep.Biol. Med. 23:242-258.
32. Folkman, Judah, Merler, Ezio, Abernathy, Charles,and Williams, Gretchen. 1971. Isolation of atumor factor responsible for angiogeneis. J.Exp. Med. 133 (2) :275-288.
33. Fraenke1 -Conrat, H. 1956. The role of nucleic acidin the reconstitution of active tobacco mosaicvirus. J. am. Chem. Soc. 78:882-893.
34. Franklin, R.M., Wecker, E., and Henry, C. 1959.Some properties of an infectious ribonucleicacid from mouse encephalomyelitis virus. Virology7:220-229.
35. Gierer, A., and Schramm, G. 1956. Die infektiositgtder nucleinsaure aus tabakmosaik-virus -Zeitschr,Naturforsch. 1 16:138-143.
36. Girardi, A.J., Hilleman, M.R. and Zwickey, R.E. 1964.Test in hamster for oncogenic quality of ordinaryviruses including adenovirus type 7. Proc. Soc.Exp. Biol. N.Y. 115:1141-1150.
37. , Sweet, B.H., Slotnick, V.B., andHilleman, M.R. 1962. Development of tumors inhamsters inoculated in the neonatal period withvacuolating virus SV40. Proc. Soc. Exp. Biol.N.Y. 109:649-660.
38. Grace, u. T. 1962. Virus and neoplasia. Surg. Clin.N. Amer. 4 2 : 2 73-285.
72

73
39. Graffi, A., and Fri t z, D. 1960.' Essaid du transmissionpar acide nucliques extraits. de la souche BB'-T2du virus de la tumour protidienne polyome.Revue Franc. d'etud. Clin. Biol. 5:388-396.
40. Hanafusa, H., and Hanafusa, T. 1966. Determiningfactor in the capacity of Rous sarcoma virus toinduce tumors in mammals. Proc. Nat. Acad. Sci.55:532-538.
41. Harel, L., Harel, J., and Huppert, J. 1967. Partialhomology between RNA from Rauscher mouse leukemiavirus and cellular DNA. Biochem. Biophy. Res.Commun. 28:44-49.
42. Hays, E.F., Simmons, N.S., and Beck, W.S. 1957.Induction of mouse leukemia with purifiednucleic acid preparation. Nature 180:1419-1423.
43. Hilleman, H.R. 1964. Prospects for the role of virusin human malignancy. Health Labour. Science 1:70-73.
44. Hruban, Z. , Kirsten, W.H., and Sleser, A. 1966. Finestructure of spontaneous hepatic tumors of maleC3H/fGs mice. Lab. Invest. 15:576-581.
45. Huebner, R.J., Casey, N.J., Chanock, R.M., and Schell,K. 1965. Tumors induced in hamsters by a strainadenovirus type 3: Sharing of tumor antigens and"neoantigens" with those produced by adenovirustype 7 tumors. Proc. Nat. Acad. Sci. 54:381-388.
46. , and Todaro, G. 1969. Oncogenes of RNAtumor viruses as determinants of cancer. Proc.Nat. Acad. Sci. 64:1087-1094.
47. , Rowe, W.P., and Lane, W.T. 1962.Oncogenic effects in hamsters of human adenovirustype 12 and 18. Proc. Nat. Acad. Sci. 48:2051-2058.

74
48. Kara, J. , Mach, ,0., and Cerna, H. 1971. Replication
of rous sarcoma virus and the biosynthesis of the
oncogenic subviral ribonucleoprotein particles
("virosomes") in the mitochondria isolated from
rous sarcoma ti ssue. Biochem. Biophys. Res.
Commun. 44(1):162-170.
49. Lacour, F., and Hafel, J. 1962. Lesions neoplastiques(erythroblastose aviare). C.R. Acad. Sci. 254:
4231-4238.
50. Lacour, F., Lacour, J., Harel, J., and Huppert, J.
1960. Transplantable malignant tumours in mice
induced by preparations containing ribonucleic
acid extracted from human and mouse tumours. J.
Nat. Cancer Inst. 24:301-309.
51. Larson, V.M., Girardi A.J., Hilleman, M.R., andZwickey, R.E. 1965. Studies of oncogenicity of
adenovirus type 7 viruses in hamsters. Proc.
Soc. Exp. Biol. N.Y. 118:15-24.
52. Latarjet, J. 1964. Possible mechanism of radiation
induced malignancy at the cellular level, p.
322-328. In: Cellular Control Mechanisms andCancer. Elsevir Pub. Co., Amsterdam.
53. , and Chamaillard, L. 1962. Inactivation
du virus de Friend par les rayonsX et les ultra-
violets. Cancer Bull. 49:382-389.
54. , Reveyrotte, N., and Moustacchi, E. 1958.
Production de cancers multiples chez des socires
ayant recu de l'acid nucleique extrait de tissues
leucemiques isologues ou homologues. C.R. Acad.
Sci. 246:853-867.
55. Leduc, E.H., and Willson, J.W. 1964. Effect of
essential fatty acid deficiency on ultrastructural
and growth of transplantable mouse hepatoma BRL.J. Natl. Cancer Inst. 33:721-727.

75
56. Luft, J.H. 1961. Improvements in epoxy resin embeddingmethods. J. Biophys. Biochem. Cytol. 9:409-414.
57. Koch, G., Loenig, S., and Alexander, H.E. 1960.
Quantitative studies on the infectivity ribonucleicacid from partially purified and highly purifiedpoliovirus preparations. Virology 10:329-343.
58. Mach, 0., and Kara, J. 1971. Presence of rous sarcoma
virus inside the mitochondria isolated by zonal
and differential centrifugation from rous sarcomacells . Folta Biol. (Praha) 17(2):65-72.
59. Maldonado, J.F., Brown, A.L., Bayrd, E.D., and Pease,G.L. 1966. Ultrastructure of the myeloma cell.Cancer 19:1613-1617.
60. McBride, W.D., and Wiener, A. 1964. In vitro transfor-mation of hamster kidney cells by human adenovirustype 12. Proc. Soc. Biol. N.Y. 115:870-874.
61. McLeod, D.L., and Ham, A.W. 1963. Search for oncogenicproperties in.various viruses found in man: Positiveresults with adenovirus type 12 and 18. Canad. Med.Assoc. J. 89:799-805.
62. Merwin, R.M., and Redmon, L.W. 1963. Induction ofplasma cell tumors and sarcomas in mice diffusionchambers placed in the peritoneal cavity. J. Natl.Cancer Inst. 31:997-1017.
63. Millonig, G. 1961. A modified procedure for leadstaining of thin sections. J. Biophys. Biochem.Ctyol..11:736-739.
64. Miroff, G., and Magdoff-Fiarchild, B.S. 1965. Studyof B particles in stomach milk of nursing mice.J. Natl. Cancer Inst. 34:777-793.
65. Montagnier, L., Golde, A., and Vigier, P. 1969. Apossible subunit structure of Rous sarcoma virus.J. Gen. Virol. 4:449-452.

76
66. Moore, D.R. , Pollard, E.C., and Haagensen, C.D. 1962.
Further correlation of physical and biological
properties of mouse mammary tumor agent. Fed.
Proc. 21:942-946.
67. Murad, T.M., and Scarpetli, D.G. 1967. The ultra-
structure of medullary and scirrhous mammaryduct carcinoma. Am. J. Pat hol. 50:335-344.
68. Nandi, S. 1967. Host-virus interactions in the mouse
mammary tumor system, p.295-314. In: Carcino-
genesis: A broad critique. Williams and Wilkins,Baltimore.
69. Novikoff, A.B., and Biempica 1966. Cytochemical and
electron microscopic examination of Morris 5123
and Reuber H-35 hepatomas after several yearstransplantation. Gann Monograph 1:65.
70. Oberling, C., and Bernhard, W. 1961. The morphologyof the cancer cell, p.405. In: J. Barchet and A.E.
Mirsky, eds., The Cell, Vol. V., Academic Press.
71. , and Guerin, M. 1954. The role of viruses
in the production of cancer. Adv. Cancer Res.
2:253-272.
72. Pease, P. 1970. Microorganisms associated with malignancy.Ann. N.Y. Acad. Sci. 174:782-785.
73. Ponten, J. 1971. Spontaneous and virus induced trans-formation in cell culture. Virol. Monogr., Springer-Verlag, New York. p.1-253.
74. Rapp, F., and Melnick, J.L. 1966. Papovavirus SV40,
Adenovirus and their hybrids: Transformation,complementation, and transcapsidation. Progr.Med. Virol. 8:349-399.
75. Robinson, W.S., and Duesberg, P. 1967. Tumor virus
RNA; in subviral carcinogenesis monografe 1stInt. Symp. on Tumor viruses, p.3-18.

77
76. Robinson, W.S., Robinson, H.L., and Dipesberg, P.1967. Tumor virus RNA's. Proc. Natl. Acad. Sci.
58:825-834.
77. Rous, P. 1911. A sarcoma of the fowl transmissibleby an agent separable from the tumor cells. J.Exp.Med. 13:397-411.
78. Sabitini, D.D., Bensch, K., and Barnett, R.J. 1963.The preservation of cellular ultrastructure andenzymatic activity by adenhyde fixation. J. CellBiol. 17:19-58.
79. Sach, L., and Medina, D. 1960. Polyoma virus mutantwith a reduction in tumour formation. Nature187:107-109.
80. Schneider, W.C., and Hogeboom, G.H. 1951. Cytochemicalstudies of mammalian tissue: The isolationof cell components by differential centrifugation:a review. J. Biol. Chem. 183:123-128.
81. Seljelid, R., and Ericsson, J.L.E. 1965. Electronmicroscopic observations on specializations ofthe cell surface in renal clear cell carcinoma.Lab. Invest. 14:435-439.
82. Scholes, V.E. 1969. Skin nucleic acid phosphorusmetabolism of DBA/1J mice during implanted tumordevelopment and methylcholanthrene carcinogenesis.Cancer Res. 29:1416-1419.
83. Shelton, E., and Rice, M.E. 1958. Studies on mouselymphomas. II. Behavior of three lymphomas indiffusion chambers in relation to their invasivecapacity in the host. J. Natl. Cancer Inst. 21:137-149.
84. Shibley, G.P., Durr, t.E., Schidlovsky, G., Wright,B.S., and Schmitter, R. 1967. Leukemogenic activityof ether-extracted Rauscher leukemia virus.Science 156:1610-1612.

78
85. Shope, R.E. 1932. A filterable virus causing tumor
like condition in rabbits and its relationship to
virus myxomatosum. J. Exp. Med. 56:803-822.
86. Southam, C. 1963. The role of viruses in neoplasia
with emphasis"on human leukemia. J. Pediatrics
63:138-149.
87. Svododa, J. Chyle, P., Simkovic, K., and Hilgert, J.
1963. Demonstration of the absence of infectious
Rous sarcoma virus in rat tumor XC, whose structural-
ly intact cells produce Rous sarcoma when trans-
ferred to chick. Folia Biol. 9:77-83.
88. __. 1964. Fine structure of hepatomas induced
in rats with P-dimethylaminoazobenzene. J. Natl.Cancer inst. 33:315-319.
89. Tandler, B., and Shipkey, F.H. 1966. Ultrastructureof Warthin's tumor I. Mitochondria. J. Ultrastruct.Res. 11:292-298.
90. Temin, H.M. 1964. Homology between RNA from Rous
sarcoma virus and DNA from Rous sarcoma virus
infected cells. Proc. Natl. Acad. Sci. 52:323-329.
91. . 1964. Nature of the provirus of Rous sarcoma.
Natl. Cancer Inst. Monograph 17:557-570.
92, . 1965. Carcinogenesis by avian sarcoma
virus. I. Cell multiplication and differentiation.J. Natl. Cancer Inst. 35:679-693.
93. .1970. Malignant transformation of cells by
viruses. Prospect. Biol. Med. 14(l):ll-26.
94. . 1971. The protovirus hypothesis: Spec-
ulations on the significance of RNA-directed DNA
synthesis for normal development for carcinogenesis.
(B) J. Nat. Cancer Inst. 46 (2) :III-VII.
95. . 1971. Mechanism of cell transforination by
RNA tumor viruses. Ann. Rev. of Microbiol. 25:609-648.

79
96. Trentin, J.J., Yabe, Y., and Taylor, G. 1962. Thequest for human cancer viruses. Science 137:
835-841.
97. Vogt, M., and Dulbecco, R. 1963. Steps in the neo-pl.astic transformation of hamster embryo cellsby polyoma virus. Proc. Natl. Acad. Sci. 49:171-177.
98. , and Rubin, H. 1962. The cytology of Roussarcoma virus infection. Cold Spring Harbor Symp.27:395-401.
99. Wecker, E. 1959. The extraction of infectious virusnucleic acid with hot phenol. Virology 7:241-249.
100. Zilber, L.A. 1945. Viruses and tumours. Kiln. Med.1:79-87.
101. , and Abelev, G.1. 1968. Theories ofCarcinogenesis, p.3-8. In: The virology andimmunology of cancer. Pergamon Press Inc., NewYork.





























