Embed Size (px)
Citation preview

Journal of Agricultural Science and Technology B 8 (2018) 405-419 doi: 10.17265/2161-6264/2018.07.001
In Vitro Development and Germination of Immature
Embryos of Sweet Oranges Parson Brown and
Pineapple (Citrus sinensis)
Karim Mahmoudi1, 2, Najat Handaji1, Mohammed Ibriz2, Najat Arsalane1, Tarik Aderdour1, 2, Kawtar Label1, 2,
Hanae Ennaciri1, 2 and Hamid Benyahya1
1. Laboratory for the Improvement and Conservation of Plant Genetic Resources, National Institute of Agronomic Research (INRA),
Kenitra, BP 257, Morocco
2. Laboratory of Genetics and Biometry, Faculty of Sciences, University Ibn Tofail, Kenitra, Morocco
Abstract: Oranges are classified as the most traded fruit variety worldwide. The diversification of the varietal profile of this group is among the objectives of the Citrus breeding program. So, the creation of new triploid hybrids via immature embryos rescue, can lead to an increase in gene pool and produce seedless varieties. In addition, flow cytometry was successfully used for analyzing ploidy levels and genetic origin of triploid plants. The present study was aimed at optimizing immature embryos in vitro germination and plantlet development based on medium chemical composition of the two orange varieties, Pineapple and Parson Brown. When the fruit gets matured, they are harvested and the seeds extracted are classified according to their size. Only those which are small are cultured in a sterile Murashige and Tucker (MT) basal medium. The different concentrations of growth regulators are put to the test in order to gain the best medium for seeds development. M1 (MT + 1 mg/L gibberellic acid (GA3)), M2 (MT + 1 mg/L kenitin + 0.5 mg/L 6-benzylaminopurine (BAP) + 0.1 naphthalene acetic acid (NAA)), M3 (MT + 25 mg/L adenine sulfate (ADS)), M4 (MT + 0.5 mg/L kenitin + 0.5 mg/L BAP + 1 mg/L GA3). Percentage of germinated embryos (PGE) is the highest in M3 with 100% and 90% for both Pineapple and Parson Brown varieties, respectively. In addition, the germination interval average (GIA) is shorter in M1 for both varieties, which takes 6 d and 7 d, respectively. As regard to speed seedling growth (mm/week), both varieties undergo significant change in all four mediums. Similarly, the highest rate of acclimatization for the Pineapple and Parson Brown varieties is 80% and 90% in M1. In general, the smaller the embryos are, the more sensitive they are to culture medium composition. It is therefore essential to optimize the medium components in order to stimulate their in vitro development. Therefore, M1 (MT + 1 mg/L GA3) remains the best to promote short term good germination and produce a better rate of acclimatization. Key words: Citrus, orange, triploids, in vitro germination, immature embryos, flow cytometry, genetic.
1. Introduction
In Morocco, the Citrus sector is one of the main
fruit production and plays an important
socio-economic role, with orange varieties dominating
the production, estimated at 962,250 tons in 2016 [1].
Similarly, orange exports increased nearly to 120,500
tons, from which Maroc Late and Navel varieties are
the highest. Citrus varietal diversification based on
Corresponding author: Najat Handaji, Ph.D., research field:
biotechnology.
high quality criteria through extending production
period is considered as the central point around which
this breeding program is rotated. Quality is pursued
based both on the visual and organoleptic criteria
including color, skin texture, caliber, aspermia etc.
and juice content, sugar, acidity, aroma, which is
considered an issue of paramount importance in the
development of this sector. Moreover, the majority of
orange varieties (Sanguine, Navel and Valencia) are at
low seed rate, except Pineapple and Parson Brown
whose rates are high [2]. They are classified among
D DAVID PUBLISHING
In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
406
the most exploited varieties of juice industry [3].
Several methods are employed on these two varieties
in order to increase genetic variability, such as the
irradiation of seeds and axillary buds that produce
seedless clones for Pineapple [4, 5]. In vitro culture of
Pineapple buds (Citrus sinensis) results in the highest
bud induction according to Murashige and Skoog (MS)
medium which contains 3.0 mg/L
6-benzylaminopurine (BAP) [6-9]. Similarly, the
hybridization method which is employed on
Clementine and tetraploid variety ‘Pineapple’ results
in the formation of 17 triploid hybrids which are
obtained from undeveloped seeds [10, 11]. In addition,
the increase of genetic variability due to triploids has
become an interesting method for seedless cultivars
development [12, 13]. Soost and Cameron [14] stated
that triploid hybrids are vigorous, productive and
seedless. Thus, the gametic sterility which
characterizes triploid hybrids is due the fact that there
is an imbalance between chromosomes when the
embryonic development occurs in the ovum, as it was
claimed by Fatta et al. [15]. This meiotic disorder
removes seeds from fruits. In general, the triploidy
which is obtained through crossbreeding and
immature embryos rescue is considered the most
commonly used process [16]. In vitro germination of
immature mandarin embryos are obtained based on
Murashige and Tucker (MT) medium and
supplemented with gibberellic acid (GA3) (0.5 mg/L
GA3) [17] while orange varieties are obtained based
on MS medium which contains adenine sulfate (ADS)
(0.25 g /L) [18]. In general, seed germination is the
most frequently used procedure for propagation but
embryos rescue is way to produce seedless triploids
hybrids in Citrus [19, 20]. The present study was
aimed at optimizing in vitro germination of immature
embryos of Pineapple and Parson Brown varieties
based on different culture medium.
2. Materials and Methods
2.1 Plant Material Preparation
Two oranges varieties (Pineapple and Parson
Brown) are used to study in vitro germination
mediums for their immature embryos. The samples are
collected from the Citrus collection of the National
Institute of Agronomic Research “El Menzeh” during
the period from November to February.
2.2 Preparation and Sterilization of Seeds
At maturity, all seeds are extracted from the ripe
fruits and classified in two categories on the basis of
their morphology; either they are normal with fully
developed seeds, or abnormal with partially developed
seeds having flat shape or small one (Fig. 1).
Under the laminar flow cabinet, the flat and small
seeds were placed in tea bags and disinfected for 10
min in a solution of Lauryl mercyl followed by 70%
alcohol for 5 min, then surface sterilized with a
sodium hypochlorite solution 4% for 2 min. After that,
they are washed. The seeds are decorticated very
gently to avoid causing damage for the embryos. The
germination medium is composed of macro and micro
elements of MT medium (1969) [21]. Then they are
supplemented with 1 mg/L of GA3, 0.5 mg/L of
kinetin, 0.5 mg/L of BAP, and 0.1 mg/L of
naphthalene acetic acid (NAA) and 25 mg/L of ADS
(Table 1).
Table 1 Composition of the medium examined for immature orange embryos rescue
Code Basic medium Constituents Dose (mg/L)
M1 MT GA3 1
M2 MT Ken + BAP + NAA 0.5 + 0.5 + 0.1
M3 MT ADS 25
M4 MT Ken + BAP + NAA + GA3 0.5 + 0.5 + 0.1 + 2
MT: Murashige and Tucker; NAA: naphthalene acetic acid; BAP: 6-benzylaminopurine; ADS: adenine sulfate; Ken: kenitin; GA3: gibberellic acid.

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
407
Fig. 1 Seeds extracted from oranges varieties and classified according to their size. a: normal; b: flat; c: small.
2.3 Culture Conditions
Based on the all experiments carried out, the
medium which constitutes the basis is that of
Murashige and Tucker, 1969. Four compositions
come under examination.
All medium in this study are sterilized by an
autoclave of 120 °C under a pressure of 1 bar for 30
min. Then they are distributed into the testing tubes.
These tubes are incubated under 2,000 of lux light, a
16 h of photoperiod and a temperature of 27 °C during
the day and 19 °C at the night.
2.4 Studied Variables
Percentage of germinated embryos (PGE):
PGE (%) = NGS
TNS (1)
NGS: number of germinated seeds; TNS: total number
of seeds cultured.
Germination interval average (GIA):
GIA (d) = GD – CD (2)
CD: in vitro culture date; GD: germination date.
Speed seedling growth (SSG): The period of
development is generally identified from an initial
date that corresponds to the date of sowing and the
date of plantlets emergence. The development period
is the one which separates the two successive events.
The velocity is as follows:
SSG (mm/week) = D D
T (3)
D2: stem length (mm) of the second week; D1: stem
length (mm) of the first week; T: time (week).
Average number of plantlets leaves is counted after
seven weeks.
2.5 Acclimatization
The rooted in vitro plants go through a hardening
phase so that they become ready for the acclimation
stage; this phase involves the test tubes to be opened,
and the roots to be cleaned as the culture medium is
solidified by agar. Then the seeds are transplanted into
a
b c
In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
408
pots containing 50% of sand and 50% of peat in a
greenhouse; these plants are covered by plastics
during a period ranges from three weeks to a month so
that a high hygrometry is maintained around young
plantlets. Also, they are transplanted into bags to
promote their development.
2.6 Flow Cytometry Analyses
Ploidy levels of seedlings from the rescue of
immature embryos were evaluated by flow cytometry
using a Partec II cytometer. The leaf tissues of each
sample were finely cut in the presence of the triploid
control (Moroccan mandarin Hana) with a razor blade,
in a Petri dish containing 0.5 mL of
phosphate-buffered saline (PBS) buffer, dithiothreitol
1 mg/L and 0.1% Triton 100. The core suspension
was filtered using a 40 μm nylon filter. Half a
milliliter of a propidium iodide solution at 1 mg/mL
was added and the mixture was incubated at room
temperature for 5 min. This fluorochrome specifically
binds to the DNA and under ultraviolet (UV)
excitation at 365 nm; the fluorescence intensity
re-emitted by the nuclei is proportional to the amount
of visible DNA on the abscissa axis. Measurements of
several thousand nuclei are retranscribed as a
histogram.
2.7 Statistical Analysis of Data
Using statistical analysis system, several variance
analyses are carried out in order to draw a comparison
between the averages of the following variables:
germination rate (%), GIA (d), SSG (mm/week) and
the leaves average number in terms of culture medium
effects. For acclimation, the Chi-square test is used to
determine its rate.
3. Results
3.1 Study of the Effect of Culture Medium on the
Studied Parameters
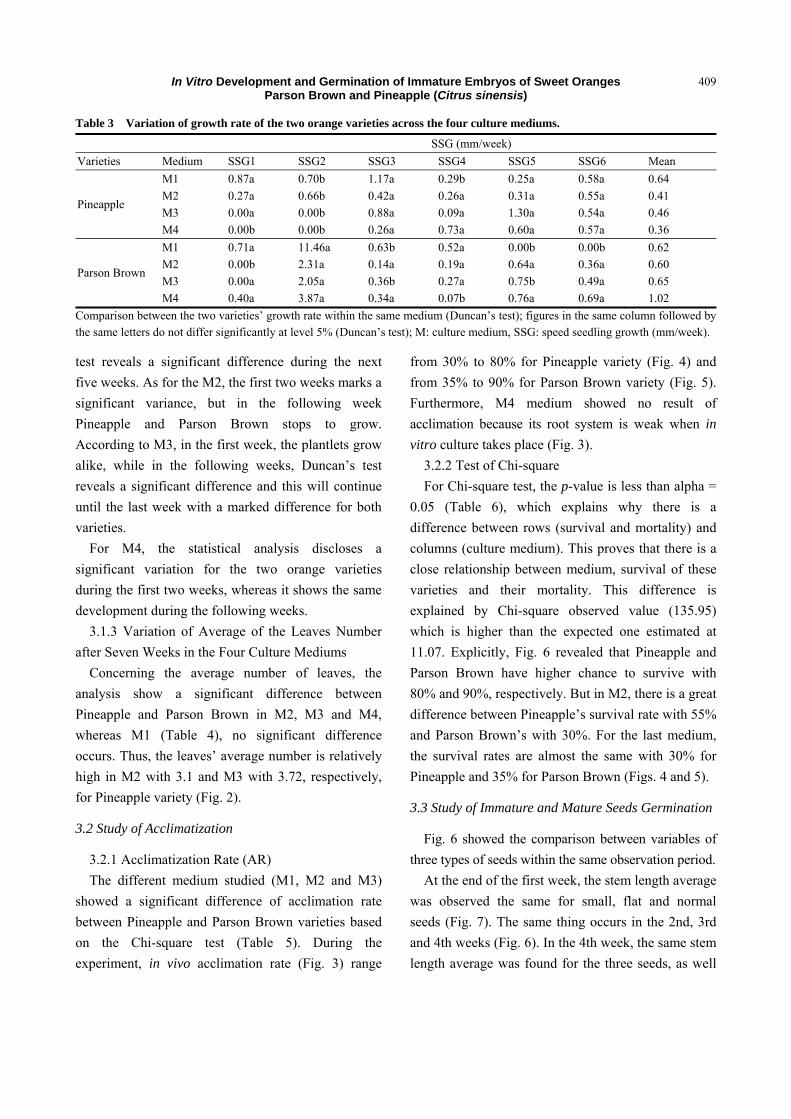
3.1.1 Percentage of Germination and GIA
The reason behind the germination of immature
embryos from the cultivars of the two oranges
varieties is that growth hormone exists in the four
culture mediums. PGE ranges from 65% to 100%
across the four culture mediums of both Pineapple and
Parson Brown varieties (Tables 2 and 3). The MT
medium which contains only ADS with
concentrations of 25 mg/L is the best performing
(100%).
For the GIA, there is no significant difference
between the two varieties “Pineapple and Parson
Brown” in the four cultures medium. The shortest and
most efficient one is the MT medium contains GA3 (1
mg/L), which takes 7 d.
3.1.2 Speed Seedling Growth (SSG)
SSG varies statistically based on the variety and the
medium (Table 3). Regarding M1, the growth rate is
the same for the two orange varieties during the first
week; however, statistical treatment through the Duncan’s
Table 2 Effect of different medium on the germination variables.
Variables Medium Varieties
Mean Pineapple Parson Brown
Percentage of germination (%)
M1 89.00a 88.00b 88.50
M2 91.00a 87.00b 89.00
M3 100.00a 90.00b 95.00
M4 65.00b 85.00a 75.00
Germination interval average (GIA)
M1 6.48a 7.02a 6.75
M2 7.27a 9.18a 8.22
M3 24.09a 22.40a 23.24
M4 16.42a 13.15a 14.78
Comparison between the averages of the two varieties for the same medium (Duncan’s test); in the same column followed by the same letters do not differ significantly at the level 5% (Duncan’s test); M: culture medium.
In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
409
Table 3 Variation of growth rate of the two orange varieties across the four culture mediums.
SSG (mm/week)
Varieties Medium SSG1 SSG2 SSG3 SSG4 SSG5 SSG6 Mean
Pineapple
M1 0.87a 0.70b 1.17a 0.29b 0.25a 0.58a 0.64
M2 0.27a 0.66b 0.42a 0.26a 0.31a 0.55a 0.41
M3 0.00a 0.00b 0.88a 0.09a 1.30a 0.54a 0.46
M4 0.00b 0.00b 0.26a 0.73a 0.60a 0.57a 0.36
Parson Brown
M1 0.71a 11.46a 0.63b 0.52a 0.00b 0.00b 0.62
M2 0.00b 2.31a 0.14a 0.19a 0.64a 0.36a 0.60
M3 0.00a 2.05a 0.36b 0.27a 0.75b 0.49a 0.65
M4 0.40a 3.87a 0.34a 0.07b 0.76a 0.69a 1.02
Comparison between the two varieties’ growth rate within the same medium (Duncan’s test); figures in the same column followed by the same letters do not differ significantly at level 5% (Duncan’s test); M: culture medium, SSG: speed seedling growth (mm/week).
test reveals a significant difference during the next
five weeks. As for the M2, the first two weeks marks a
significant variance, but in the following week
Pineapple and Parson Brown stops to grow.
According to M3, in the first week, the plantlets grow
alike, while in the following weeks, Duncan’s test
reveals a significant difference and this will continue
until the last week with a marked difference for both
varieties.
For M4, the statistical analysis discloses a
significant variation for the two orange varieties
during the first two weeks, whereas it shows the same
development during the following weeks.
3.1.3 Variation of Average of the Leaves Number
after Seven Weeks in the Four Culture Mediums
Concerning the average number of leaves, the
analysis show a significant difference between
Pineapple and Parson Brown in M2, M3 and M4,
whereas M1 (Table 4), no significant difference
occurs. Thus, the leaves’ average number is relatively
high in M2 with 3.1 and M3 with 3.72, respectively,
for Pineapple variety (Fig. 2).
3.2 Study of Acclimatization
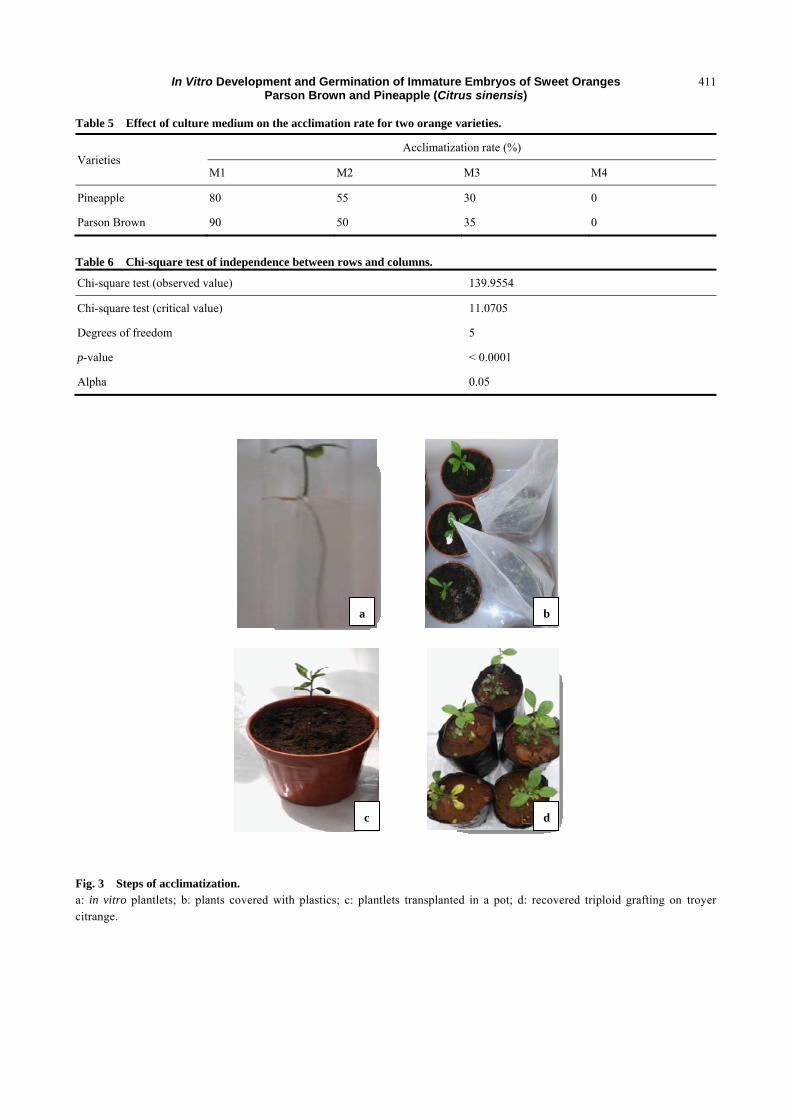
3.2.1 Acclimatization Rate (AR)
The different medium studied (M1, M2 and M3)
showed a significant difference of acclimation rate
between Pineapple and Parson Brown varieties based
on the Chi-square test (Table 5). During the
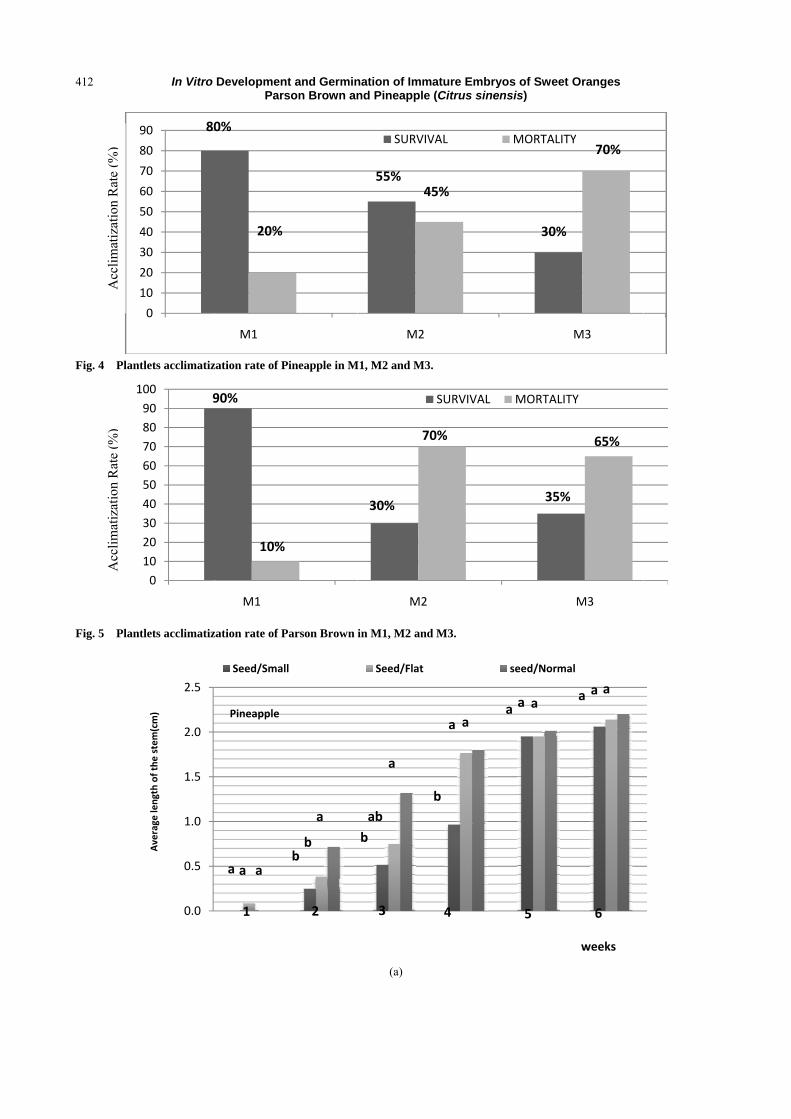
experiment, in vivo acclimation rate (Fig. 3) range
from 30% to 80% for Pineapple variety (Fig. 4) and
from 35% to 90% for Parson Brown variety (Fig. 5).
Furthermore, M4 medium showed no result of
acclimation because its root system is weak when in
vitro culture takes place (Fig. 3).
3.2.2 Test of Chi-square
For Chi-square test, the p-value is less than alpha =
0.05 (Table 6), which explains why there is a
difference between rows (survival and mortality) and
columns (culture medium). This proves that there is a
close relationship between medium, survival of these
varieties and their mortality. This difference is
explained by Chi-square observed value (135.95)
which is higher than the expected one estimated at
11.07. Explicitly, Fig. 6 revealed that Pineapple and
Parson Brown have higher chance to survive with
80% and 90%, respectively. But in M2, there is a great
difference between Pineapple’s survival rate with 55%
and Parson Brown’s with 30%. For the last medium,
the survival rates are almost the same with 30% for
Pineapple and 35% for Parson Brown (Figs. 4 and 5).
3.3 Study of Immature and Mature Seeds Germination
Fig. 6 showed the comparison between variables of
three types of seeds within the same observation period.
At the end of the first week, the stem length average
was observed the same for small, flat and normal
seeds (Fig. 7). The same thing occurs in the 2nd, 3rd
and 4th weeks (Fig. 6). In the 4th week, the same stem
length average was found for the three seeds, as well

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
410
as the same root length average (Fig. 6), which
explains that there is a growth harmony in stem and
root length.
The stem length average of the Parson Brown
variety shows a significant difference in the second
week and this continues in the sixth one. After that,
there is similarity between these seeds (Fig. 6).
Moreover, the root length average of Parson Brown
variety shows from the first week to the fifth one a
significant difference between small, flat and normal
seeds, whereas it reveals a similarity in the sixth week
(Fig. 6).
3.4 Result of Flow Cytometry Analysis
A significant number of polyploids recovered from
underdeveloped seeds of Citrus cultivars. Only the
Table 4 Effect of different basal medium on the leaves’ average number variation after seven weeks according to medium culture.
Varieties Average of leaves’ number
M1 M2 M3 M4
Pineapple 2.32bc 3.10ab 3.72a 2.66bc
Parson Brown 2.20bc 1.81c 2.80abc 1.80c
Comparison between averages of the two varieties of the same medium (Duncan’s test); the numbers in the same rows followed by the same letters do not differ significantly at level 5% (Duncan’s test); M: culture medium.
Fig. 2 In vitro plantlets development through embryo rescue after seven weeks of incubation on different basal medium. M: culture medium.
M2
M4M 3
M 1 M1
M3
In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
411
Table 5 Effect of culture medium on the acclimation rate for two orange varieties.
Varieties Acclimatization rate (%)
M1 M2 M3 M4
Pineapple 80 55 30 0
Parson Brown 90 50 35 0
Table 6 Chi-square test of independence between rows and columns.
Chi-square test (observed value) 139.9554
Chi-square test (critical value) 11.0705
Degrees of freedom 5
p-value < 0.0001
Alpha 0.05
Fig. 3 Steps of acclimatization. a: in vitro plantlets; b: plants covered with plastics; c: plantlets transplanted in a pot; d: recovered triploid grafting on troyer citrange.
a b
c d
412
Fig. 4 Plant
Fig. 5 Plant
1
2
3
4
5
6
7
8
9
4
1
Acc
lim
atiz
atio
n R
ate
(%)
Acc
lim
atiz
atio
n R
ate
(%)
In Vitro
tlets acclimatiz
tlets acclimatiz
0
10
20
30
40
50
60
70
80
90 80
0
10
20
30
40
50
60
70
80
90
009
0.0
0.5
1.0
1.5
2.0
2.5
Average
length of the stem(cm)
DevelopmenPars
zation rate of P
zation rate of P
M1
0%
20%
M1
90%
10%
Seed/Smal
1
a aa
Pineapple
nt and Germinson Brown an
Pineapple in M
Parson Brown
l
2
a
bb
nation of Immnd Pineapple
M1, M2 and M3
in M1, M2 and
(a)
M2
SURVIV
55%4
M2
7
30%
Seed/Flat
3
b
a
ab
mature Embrye (Citrus sinen
3.
d M3.
VAL
45%
SURVIVAL
0%
4
aa
b
yos of Sweet nsis)
MORTALITY
30%
MORTALIT
35%
seed/Norma
5
aaa
Oranges
M3
Y70%
M3
TY
65%
weeks
l
6
a a a
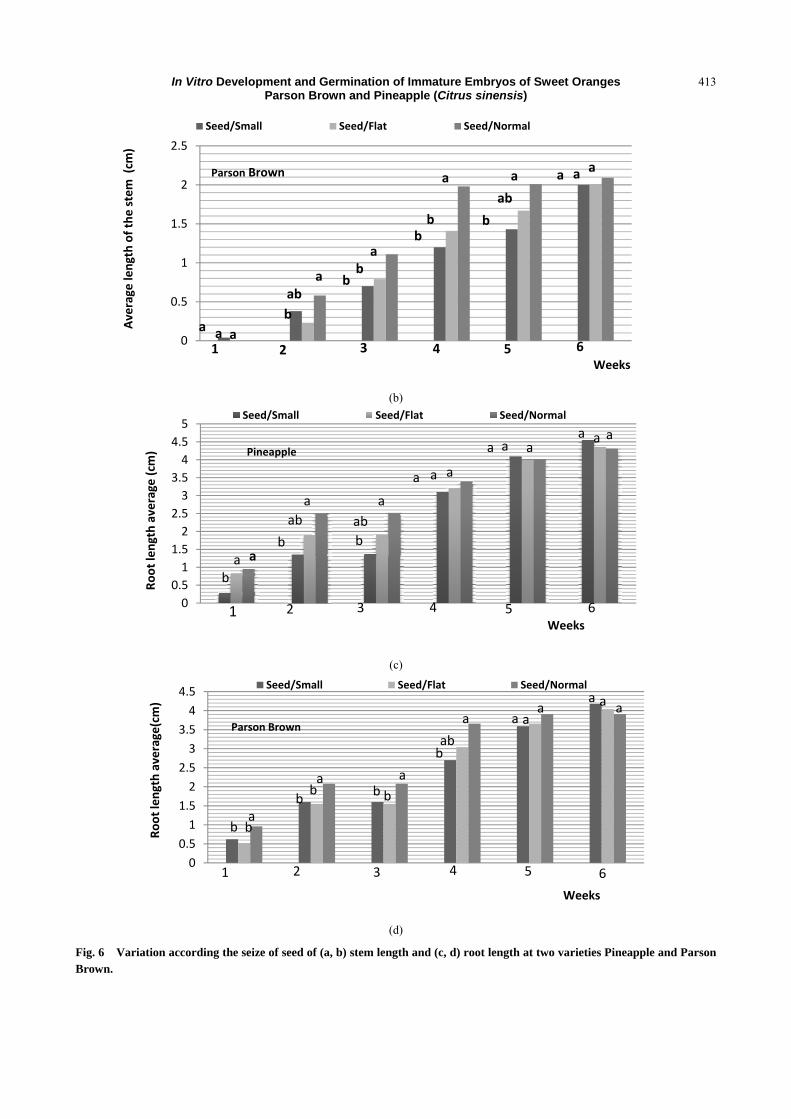
Fig. 6 VariaBrown.
Average
length of the stem (cm
)
In Vitro
ation according
0
0.5
1
1.5
2
2.5
Se
a
1
P
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Root length average
(cm
)
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
Root length average(cm)
DevelopmenPars
g the seize of s
eed/Small
b
aa1 2
Parson Brown
Seed/Sm
1
ab
a
b
Pineapp
Seed
1
bab
Parson Brow
nt and Germinson Brown an
eed of (a, b) st
Seed
bba
b
ab
2
mall
2
a
b
a
ab
ple
d/Small
2
a
bb
wn
nation of Immnd Pineapple
(b)
(c)
(d)
tem length and
d/Flat
bb
ab
3
Seed/Flat
3
a
a
ab
b
Seed/F
3
a
bb
mature Embrye (Citrus sinen
d (c, d) root len
Seed/N
a
b
a
b
4
S
4
aa
a
Flat
4
b
a
ab
yos of Sweet nsis)
ngth at two var
ormal
aa
ab
5
Wee
Seed/Normal
5
aa
W
Seed/Norm
5
aa
a
Oranges
rieties Pineapp
Weeks
a a
6
eks
6
aa a
Weeks
mal
6
a a a
413
ple and Parson
3
n

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
414
medium M1 which is the best medium containing
GA3 in terms of the embryo rescue and recovering of
plant triploids, Citrus cultivars were examined for
seed size underdeveloped. Maximum triploids plants
66.67% were found in small seed of variety Parson
Brown, followed by flat seed 40.00%. Furthermore,
25.00% of triploids plants were observed in flat seed
of variety Pineapple whereas the minimum value
(0.0%) was observed in small seed (Table 7).
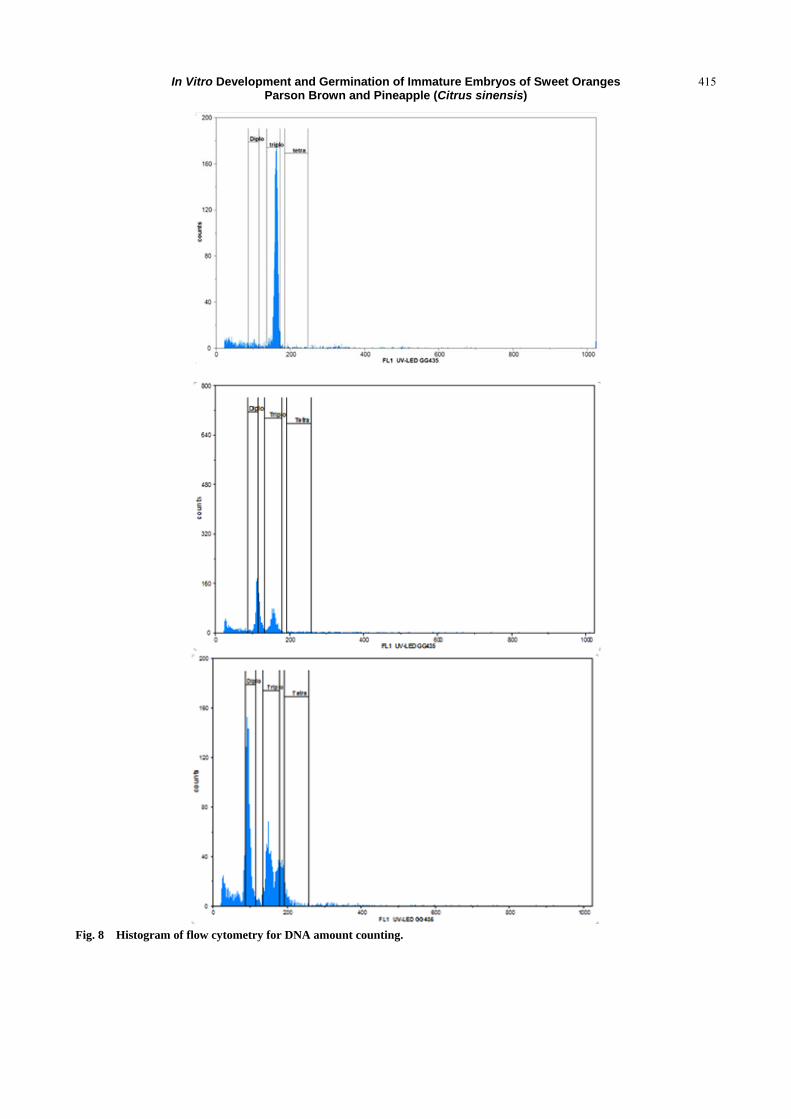
The ploidy level profiles of two Parson Brown and
Pineapple varieties from the immature embryo rescue
with that of the control are presented in the form of
histograms (Fig. 8), the fluorescence intensity emitted
appearing on the abscissa axis and the number of
nuclei on the ordinate.
Triploids seedling of two orange varieties Pineapple
and Parson Brown selected by cytometry analysis
were budded on Troyer citrange and planted in
National Institute of Agronomic Research (INRA)
domain (Fig. 9).
Fig. 7 Seedling regeneration after six weeks of culture. a: small; b: flat; c: normal.
Table 7 Number of plants analyzed and ploidy level of plants recovered in oranges ‘Parson Brown’ and ‘Pineapple’ in medium M1.
Cultivars Seed size Percentage of polyploids
Haploids (N) Diploids (2N) Triploid (3N) Tetraploids (4N)
Parson Brown Small - 33.33 66.67 -
Flat - 60.00 40.00 -
Pineapple Small - 100.00 0.00 -
Flat - 75.00 25.00 -
a b c
In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
415
Fig. 8 Histogram of flow cytometry for DNA amount counting.

416
Fig. 9 Recov
4. Discussi
In fact, th
significantly
obtained fro
varieties. Th
by Ziani [22
mandarin va
who found a
when using
additive in p
seed germin
MS supplem
addition, sev
on immatur
containing 2
and germina
Phrase longu
of Carica p
[28].
GIA is s
which conta
to those foun
study of av
showed the
that GA3 p
believe that
thus reduces
In Vitro
vered triploid
ions
he favored MT
y stimulate im
om undevelo
hese results a
2] for argan p
arieties. In c
a low germina
ADS. It is no
plant tissue cu
nation was op
mented with
veral germin
e orange em
20 mg/L AD
ation result w
ue ADS also
papaya, Mus
significant in
ains GA3. For
nd by Vishwa
vocado carrie
same results.
promotes dorm
t GA3 reduc
s the time of
DevelopmenPars
grafting on tro
T medium wi
mmature emb
oped seed of
are similar to
lant and Jask
ontrary to M
ation percent
ormally used
ulture [24, 25
ptimum 66.2%
h 25 mg/L
nation studies
mbryos based
S and 500 m
was significan
used for in v
sa sp. [27] a
n the favore
r 7 d, these r
akarma [29].
ed out by Ku
. Some schola
mancy, whil
es the dorm
f preservation
nt and Germinson Brown an
oyer citrange.
ith ADS seem
ryos germina
f the two ora
o those obta
kani et al. [16
Mahdy et al.
age for Zea m
also as a gro
5]. In other st
% in the med
ADS [26].
s are carried
on MS med
mg/L malt ext
nt with 75% [
vitro propaga
and Uraria p
ed MT med
results are sim
The germina
ucera et al.
ars [31-33] c
le others [34
mancy period
n before plant
nation of Immnd Pineapple
ms to
ation
ange
ained
] for
[23]
mays
owth
udy,
dium
. In
d out
dium
tract
[16].
ation
picta
dium
milar
ation
[30]
laim
4-36]
and
ting.
An
GA
foun
mg/
germ
sho
resu
T
rate
thes
al.
whi
con
lead
the
acc
in M
whi
[41
[42
can
Ham
be
pro
imp
frui
pres
the
mature Embrye (Citrus sinen
experiment
A3 reduce the
nd the same
/L GA3 to
mination of
wed that add
ult in good ge
The medium
e (%) for Pine
se results are
[38] and Zdru
ich indicates
ntribute to th
ding to better
average nu
ording to Du
M2 and M3
ich are simila
] for pistach
], the results
n produce a
mad and Tah
the most su
liferation, a
portant role o
its [44, 45]. S
sence of ADS
other hand, t
yos of Sweet nsis)
on Kiwi fru
average tim
results, whic
o the medi
citrus embr
ding 1 mg/L G
ermination of
M1 reveals
eapple and P
e similar to th
ujkovskaja-ri
s that addin
he germinati
r survival of
umber of lea
uncan’s test, t
for Pineapp
ar to those ob
hio varieties.
showed that
large numbe
a [43] states
uccessful cyto
and many
of BAP in fl
Some researc
S encourages
tissue culture
Oranges
uit seeds whi
e of germina
ch indicate t
ium contrib
ryos. Some
GA3 accordin
f citrus varieti
a significant
Parson Brown
hose found b
ichter [39]. A
ng GA3 to
ion of apple
seedling [40
aves, the res
that there is g
ple variety w
tained by Ab
According t
t the auxin/cy
er of leaves.
that BAP is c
okinin horm
scientists s
lower inducti
chers also sho
s shoot induc
e trials using
ile using the
ation [37, 38]
hat adding 2
bute to the
reports also
ng to Ref. [39
ies.
t acclimation
n varieties, as
by Gmitter et
An experience
the medium
e seeds and
0]. Regarding
sults showed
good number
without GA3,
bousalim et al
to Sim et al.
ytokinin ratio
In addition,
considered to
mone for leaf
showed the
ion of Citrus
owed that the
ction [46], on
MS medium
e
]
2
e
o
9]
n
s
t
e
m
d
g
d
r
,
l.
o
,
o
f
e
s
e
n
m

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
417
with supplements of various hormonal concentrations
including BAP, kinetin and ADS are useful in shoot
proliferation [47]. The use of additives such as ADS
as seen in the study is a key factor in the production of
shoots from Jatropha curcas, a concept that supports
reports by Sarika and Meenakshi [48]. Munde and
Gajbhiye [49] studied the effect of regulators GA3 on
seedling growth of mango stones and producing more
number of leaves.
Furthermore, the combination of three hormones
namely BAP, kinetin, NAA leads to a better leaf
production for of mandarin variety [50]. Also, the
medium which contains GA3 is the one which
stimulate root growth [41], besides, Singh and Kaur
[51] showed that the use of auxins is so important for
root system for citrus species such as limonia, sinensis
and jambhiri citrus.
The study of immature and mature seeds
germination revealed a significant variation between
seed type (small, flat and normal), so their
development was different according to the time. In
fact, embryo rescue depends on the stages of
embryonic development and the composition of the
nutritional medium [52]. A study carried out by
Caraimi et al. [53] showed that the embryos sized (<
0.3 mm) had 76% of germination based on MT
medium which contains 150 mm of sucrose and 14.4
μM of GA3. While embryos whose size is 0.3-0.8 mm
had 95% of germination based on the same medium
supplemented with 150 mm of sucrose and 2.9 μM of
GA3.
MT basal medium containing 1 mg/L GA3 recovered
85 triploid plants Citrus [54]. Triploids were obtained
from underdeveloped seeds ranging 1/3 to 1/6 [55, 56].
Similarly spontaneous triploids were obtained among
progenies of Lisbon and Eureka lemon, Ruby sweet
orange and Imperial grapefruit [57].
5. Conclusions
Triploid plants are rare in nature because of their
inviable seeds and resulting lack of progeny, so it is
challenging to detect naturally occurring triploid
plants. However, due to their faster growth and
seedlessness, they will be useful for improving
biomass, fruit and flower traits, and other qualities of
economically important food, The use of in vitro
embryo rescue and ploidy level assessment by flow
cytometry gave highly efficient recovery of Citrus
triploid hybrids from small and flat seeds. This study
of in vitro germination of immature embryos for two
orange varieties Pineapple and Parson Brown showed
that the favored MT medium containing GA3 is
efficient in terms of the GIA great advantage for in
vitro germination of citrus varieties. For MT medium
which contains ADS, a large average number of
leaves are produced for both varieties, while the M4
could not produce any adequate roots in Pineapple and
Parson Brown acclimatization with a value of 0%.
References
[1] Adam, H. P. 2016. “Citrus Production by County.” United States Agriculture Department.
[2] Mahmoudi, K., Handaji, N., Arsalane, N., Ibriz, M., Aderdour, T., Label, K., Ait El Aouad, B., and Benyahya, H. 2017. “Preliminary Selection of the Orange Cultivars Using as Female Parents in Diploids Crosses and in Triploidy Program.” International Journal of Botany Studies 2 (6): 232-40.
[3] Handaji, N., Benyahia, H., Arsalane, N., Ben Azouz, A., and Gaboun, F. 2013. “A Pomological and Organoleptic Evaluation of 34 Orange Variants (Citrus sinensis (L.) Osbeck) from Apomictic Seedlings Tested in the Gharb Region.” Al Awamia 127: 45-70.
[4] Hearn, C. J. 1984. “Development of Seedless Orange and
Grapefruit Cultivars through Seed Irradiation.” J. Am.
Soc. Hort. Sci. 109: 270-3.
[5] Hearn, C. J. 1985. “Citrus Scion Improvement Program.”
Fruits Var. J. 39: 34-7.
[6] Duran-vila, N., and Navarro, L. 1989. “Morphogenesis and Tissue Culture of Three Citrus Species.” Plant Cell, Tissue and Organ Culture 16: 123-33.
[7] Canto, A. M., Souza, F. V., Costa, M. A., Souza, A. S., Ledo, C. A., and Cabral, J. R. 2004. “In Vitro Conservation of Abacaxitrated Germplasm with Paclobutrazol.” Pesquisa Agropecuária Brasileira 39 (7): 717-20.
[8] Marin, M. L., and Duran-vila, N. 1991. “Conservation of Citrus Germplasm in Vitro.” Amer. Soc. Hort. Sci. 116

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
418
(4): 740-6. [9] Kurt, S., and Ulger, S. 2014. “Production of Common
Sour Orange: Carrizo Citrange Hybrids Using Embryo Rescue.” International Journal of Fruit Science 14: 42-8.
[10] Aleza, P., Juárez, J., Ollitrault, P., Navarro, L., and Bot,
A. 2010. “Polyembryony in Non-apomictic Citrus
Genotypes.” Annals of Botany 106 (4): 533-45.
[11] Ollitrault, P., Dambier, D., Luro, F., and Froelicher, Y.
2008. “Ploidy Manipulation for Breeding Seedless
Triploid Citrus.” Plant Breeding Reviews 20: 323-54.
[12] Kamiri, M., Stift, M., Srairi, I., Costantino, G., El
Moussadik, A., Hmyene, A., Bakry, F., Ollitrault, P., and
Froelicher, Y. 2011. “Evidence for Non-disomic
Inheritance in a Citrus Interspecific Tetraploid Somatic
Hybrid between C. reticulate and C. limon Using SSR
Markers and Cytogenetic Analysis.” Plant Cell Reports
30 (8): 1415-25.
[13] Esen, A., and Soost, R. K. 1971. “Unexpected Triploids in Citrus: Their Origin, Identification and Possible Use.” Journal of Heredity 62: 329-33.
[14] Soost, R. K., and Cameron, J. W. 1975. Advances in Fruit Breeding. United States of America: Purdue University Press.
[15] Fatta, D., El Bosco., S. M., Atranga, G., and Geraci, G. 1992. “Micro and Macro Sporogenesis of Two Triploid Hybrids of Citrus.” Proc. Int. Soc. Citriculture 2 (1): 2-124.
[16] Jaskani, M. J., Khan, I. A., and Khan, M. M. 2005. “Fruit Set, Seed Development and Embryo Germination in Interploid Crosses of Citrus.” Sci. Hort. 107 (1): 51-7.
[17] Tan, M., Song, J., and Deng, X. 2007. “Production of Two Mandarin × Trifoliate Orange Hybrid Populations via Embryo Rescue with Verification by SSR Analysis.” Euphytica 157 (1-2): 155-60.
[18] Mehmood, A., Jaskani, M. J., Sajjad, Y., Ahmad, I., and Hussain, Z. 2011. “In Vitro Germination of Hybrid Embryos from Interploidal Crosses of Citrus.” Pakistan Journal of Botany 43 (4): 2045-9.
[19] Handaji, N., Benyahia, H., Arsalane, N., and Benaouda, H. 2018. “AYA and HANA: New Seedless Mandarin’s Triploid Hybrids Selected in Morocco.” Biotechnology Journal International 21 (1): 1-9.
[20] Handaji, N., Benyahia, H., Arsalane, N., and Benaouda, H. 2017. “Selection of New Promising Seedless Mandarins Triploid Hybrids from Crosses between Monoembryonic Diploid Female and Diploid Male Parents.” Research in Biotechnology 8: 37-44.
[21] Murashige, T., and Tucker, D. P. H. 1969. “Growth Factor Requirements of Tissue Culture.” In Proceedings of the First International Citrus Symposium, Vol. 3. University of California, Riverside, 1155-61.
[22] Ziani, S. 2014. Multiplication of Argan (Arganiaspinosa
L. Skeels) by in Vitro Sowing, Microbouturing, Micrografting, Organogenesis and/or Somatic Embryogenesis. Algeria. University of Chlef Hassiba Benbouali.
[23] Mahdy, A. M., Elkhatib, E. A., and Fathi, N. O. 2007. “Drinking Water Treatment Residuals as Amendment to Alkaline Soils, Effects on Growth of Corn and Phosphorus Extractability.” International Journal of Environmental Science and Technology 4 (4): 489-96.
[24] Deora, N. S., and Shekhawat, N. S. 1995. “Micropropagation of Capparis decidua (Forsk.) Edgew.—A Tree of Arid Horticulture.” Plant Cell Reports 15 (3-4): 278-81.
[25] Das, S., Jha, T. B., and Jha, S. 1996. “Organogenesis and Regeneration from Pigmented Callus in Camellia sinensis (L.) O. Kuntze cv. Nandadevi, an Elite Darjeeling Tea Clone.” Plant Science 121 (2): 207-12.
[26] Park, S. Y., Murthy, H. N., and Paek, K. Y. 2000. “In Vitro Seed Germination of Calanthe sieboldii, an Endangered Orchid Species.” Journal of Plant Biology 43 (3): 158-61.
[27] Schmildt, O., Schmildt, E. R., and Jose, A. T. 2007. “Adeninan Sulfate in Vitro Multiplication of Papaya “Tainung 01”.” Scientia Agraria 8 (2): 141-7.
[28] Anand, A., Srinivasa, R. C., Latha, R., Josekutty, P. C., and Balakrishna, P. 1998. “Micropropagation of Uraria picta, a Medicinal Plant, through Axillary Bud Culture and Callus Regeneration.” In Vitro Cellular and Developmental Biology Plant 34 (2): 136-40.
[29] Vishwakarma, D. 2013. “Effect of Growing Media and GA3 on Seed Germination, Growth and Survival of Acid Lime (Citrus aurantifolia Swingle) var. Kagzi.” MSc. thesis, Jabalpur College of Agriculture Jabalpur (M.P.), 28-9.
[30] Kucera, B., Cohn, M. A., and Leubner-Metzger, G. 2005. “Plant Hormone Interactions during Seed Dormancy Release and Germination.” Seed Science Research 15 (4): 281-307.
[31] Rappaport, L., Lippert, L. F., and Timm, H. 1957. “Sprouting, Plant Growth and Tuber Production as Affected by Chemical Treatment of White Patata Seed Pieces.” American Potato Journal 34 (9): 254-60.
[32] Tsukamoto, Y., Asahira, T., and Namiki, T. 1961. “Studies on the Dormancy of the Potato Tuber: The Effect of Gibberellic on Breaking Dormancy of Potatoes Lifted at Different Times.” Mem. Res. Inst. Fd. Sci. Kyoto Univ. 23: 23-7.
[33] Timm, H., Rappaport, L., Bishop, J. C., and Hoyle, B. J. 1962. “Sprouting, Plant Growth, and Tuber Production as Affected by Chemical Treatment of White Potato Seed Species. IV. Responses of Dormant and Sprouted Seed Potatoes to Gibberellic Acid.” Amer. Potato J. 39:

In Vitro Development and Germination of Immature Embryos of Sweet Oranges Parson Brown and Pineapple (Citrus sinensis)
419
107-15. [34] Doorenbos, J. 1958. “Effect of Gibberellic Acid on
Sprouting of Potatoes.” Neth. J. Agric. Sci. 6: 267-70. [35] Marinus, J., and Bodlaender, K. B. A. 1978. “Growth and
Yield of Seed Potatoes after Application of Gibberellic Acid to the Tubers before Planting.” Neth. J. Agric. Sci. 26: 354-65.
[36] De Fernandez, O. S., Borgo, R., and Tizio, R. 1990. “Rupture of Dormancy of Potato Tubers (Solanum tuberosum L.): Effects of Injuries, Gibberellic Acid, Kinetin and Combined Treatments of These Two Growth Regulators.” C.R. Soc. Biol. 184: 259-68.
[37] Ynoue, C. K., Ono, E. O., and Marchi, L. D. O. S. 1999. “The Effect of Gibberellic Acid on Kiwi (Actinidia chinensis Pl.) Seed Germination.” Scientia Agricola 56 (1): 9-12.
[38] Gmitter, F. G., Ling, X. B., and Deng, X. X. 1990. “Induction of Triploid Citrus Plants from Endosperm Cali in Vitro.” Theor. Appl. Genet. 80: 785-90.
[39] Zdrujkovskaja-richter, A. I. 1981. Embryo Cultures and Development of New Forms of Plants. Moscow: University Moscow. (Abstr.)
[40] Wani, R. A., Malik, T. H., Malik, A. R., Baba, J. A., and Dar, N. A. 2014. “Studies on Apple Seed Germination and Survival of Seedlings as Affected by Gibberellic Acid under Cold Arid Conditions.” International Journal of Scientific and Technology Research 3 (3): 210-6.
[41] Abousalim, A., El mahboul, B., and Walali, L. D. 1992. “In Vitro Germination of Seeds and Growth of Pistachio Seedlings (Pistacia vera L.).” Res. Impr. Prod. Agr. Arid Environment 4: 17-23.
[42] Sim, G. E., Goh, C. J., and Loh, C. S. 1988. “Multiple Bud Formation from Shoot and Root Explants in the Presence of 6-benzylaminopurine.” Annual Review of Plant Physiology 12 (2): 108-11.
[43] Hamad, A. M., and Taha, R. M. 2008. “Effect of Benzylaminopurine (BAP) on in Vitro Proliferation and Growth of Pineapple (Ananas comosus L. Merr.) cv. Smooth Cayenne.” Journal of Applied Sciences 8 (22): 4180-5.
[44] Saini, H. K., and Gill, M. S. 2010. “Direct Shoot Organogenesis and Plant Regeneration in Rough Lemon (Citrus jambhiri Lush.).” Ind. J. Biotech. 9 (419): 423.
[45] Rattanpal, H. S., Kaur, G., and Gupta, M. 2011. “In Vitro Plant Regeneration in Rough Lemon (Citrus jambhiri
Lush.) by Direct Organogenesis.” Afr. J. Biotech. 10 (63): 13724-8.
[46] Kumar, N., Anand, K. G. V., and Reddy, M. P. 2010. “Shoot Regeneration from Cotyledonary Leaf Explants of Jatropha curcas: A Biodiesel Plant.” Acta Physiologiae Plantarum 32 (5): 917-24.
[47] Khan, M. K., Misra, P., Sharma, T., Shukla, P. K., and Ramteke, P. W. 2014. “Effect of Adenine Sulphate on in Vitro Mass Propagation of Stevia rebaudiana Bertoni.” Journal of Medicinal Plant Research 8 (13): 543-9.
[48] Sarika, S., and Meenakshi, B. 2008. “In Vitro Propagation of Physic Nut (Jatropha curcas L.): Influence of Additives.” International Journal of Intergrative Biology 3 (1): 73-9.
[49] Munde, G. R., and Gajbhiye, R. P. 2010. “Effect of Plant Growth Regulators on Seedling Growth of Mango Stones.” Green Farming 1 (3): 288-9.
[50] Singh, S., Ray, B. K., Bhattacharyya, S., and Deka, P. C. 1994. “In Vitro Propagation of Citrus reticulata Blanco and Citrus limon Burm. f.” Hort Sci. 29 (3): 214-6.
[51] Singh, B., and Kaur, A. 2011. “Comparison of Agar and Gum Karaya as Gelling Agent for in Vitro Regeneration of Rough Lemon (Citrus jambhiri Lush.) Plantlets from Nodal Explants.” J. Crop Sci. Biotech. 14 (4): 297-303.
[52] Rangan, T. S., Murashige, T., and Bitters, W. P. 1969. “In Vitro Studies of Zygotic and Nucellar Embryogenesis in Citrus.” In Proceeding of the First International Citrus Symposium, edited by Chapman, H. D. University of California, Riverside, 225-9.
[53] Carimi, F., Pasquale, F., and Puglia, A. M. 1998. “In Vitro Rescue of Zygotic Embryos of Sour Orange (Citrus aurantium L.) and Their Detection Based on RFLP Analysis.” Plant Breeding 117 (3): 261-6.
[54] Vasil, I. K., and Thorpe, T. A., eds. 1994. Plant Cell and Tissue Culture. Kluwer. Acad. Publ., Dordrecht, The Netherlands, 483.
[55] Esen, A., and Soost, R. K. 1973. “Seed Development in Citrus with Special Reference to 2X × 4X Crosses.” American Journal of Botany 60 (5): 448-62.
[56] Esen, A., and Soost, R. K. 1973. “Precocious Development and Germination of Spontaneous Triploid Seed in Citrus.” Journal of Heredity 64 (3): 147-54.
[57] Esen, A., and Soost, R. K. 1971. “Unexpected Triploids in Citrus: Their Origin, Identification and Possible Use.” Journal of Heredity 62 (6): 329-33.





























