Embed Size (px)
Citation preview

In Situ Hybridization Screen in Zebrafish for theSelection of Genes Encoding Secreted ProteinsPHILIP S. CROSIER,1 ANNE BARDSLEY,1 JULIA A. HORSFIELD,1 ANNA K. KRASSOWSKA,1
EDWARD R. LAVALLIE,2 LISA A. COLLINS-RACIE,2 JOHN H. POSTLETHWAIT,3 YI-LIN YAN,3
JOHN M. MCCOY,2 AND KATHRYN E. CROSIER1
1Division of Molecular Medicine, Faculty of Medical and Health Sciences, University of Auckland, Auckland, New Zealand2Genetics Institute/Wyeth-Ayerst Research, Cambridge, Massachusetts3Institute of Neuroscience, University of Oregon, Eugene, Oregon
ABSTRACT An in situ hybridization expres-sion screen using a signal sequence trap systemhas been conducted in zebrafish to isolate cDNAsthat encode secreted proteins. Random clones(secreted expressed sequence tags; sESTs) weresequenced from zebrafish embryonic (18–24 hrpostfertilization) and adult kidney libraries.From the two RNA sources, 627 random sESTcDNAs were identified as being homologous oridentical to known genes and 166 clones encodecurrently unidentified genes. The sESTs repre-sent a broad range of enzymes and other regula-tory molecules. Whole-mount in situ hybridiza-tion analysis was carried out by using antisenseprobes generated from 244 selected sESTs, and arange of expression patterns was obtained. Ge-netic mapping undertaken with sEST sequencesdemonstrated that assignment of map positionwas attainable by using 5* primers. The signalsequence trap system used in this work hasyielded a range of cDNAs that encode secretedproteins and, together with analysis of patternsof expression and genetic mapping, has the po-tential to facilitate analysis of signaling path-ways central to development and physiology.© 2001 Wiley-Liss, Inc.
Key words: expression screen; secreted proteins;signal sequence trap; zebrafish
INTRODUCTION
Genetic mutagenesis screens of Drosophila havebeen instrumental in identifying key regulators of em-bryonic development (Nusslein-Volhard and Wie-schaus, 1980; Eberl and Hilliker, 1988). Large scalemutagenesis screens have also been carried out in ze-brafish, providing hundreds of mutant lines for theanalysis of development in vertebrates (Driever andFishman, 1996; Haffter et al., 1996). A large number ofgenes identified in such screens have been shown tohave orthologs in higher organisms, including humans.Yet, whereas the “phenotype first” approach recognizesdevelopmentally indispensable genes, it can overlookgenes with subtle loss-of-function phenotypes (fre-quently due to partial gene redundancy), or strong
phenotypes in tissues or developmental stages not ex-amined by the screen. Moving from mutant stock tocloned gene is often difficult and time consuming, al-though genome projects are assisting significantly inthis area. To overcome these limitations, “gene first”approaches have been used in several organisms, suchas random expression screening (Lynch et al., 1995;Babin et al., 1997; Gawantka et al., 1998; Grammer etal., 2000; Thut et al., 2001), enhancer and promotertrapping (Wilson et al., 1989; Hope et al., 1998), differ-ential display (Zhang et al., 1998), and gene trapping inembryonic stem cells (Evans et al., 1997; Zambrowiczand Friedrich, 1998, for review). Some of these tech-nologies have been modified to select for moleculesexpressed in a specific subcellular location, includingnuclear, secreted, or transmembrane proteins (Skarneset al., 1995; Tate et al., 1998). Recently, gene trapapproaches have been extended to include a secretorytrap carried out in mice to analyze the in vivo functionsof genes encoding secreted and transmembrane pro-teins (Mitchell et al., 2001).
Many developmental processes are regulated by se-creted proteins and their membrane-bound receptors.For example, members of the hedgehog, wnt, nodal,and bone morphogenetic protein pathways are criticalin patterning and organogenesis. Because of the impor-tant developmental roles of secreted proteins, specificeffort has been made to identify genes encoding suchproteins. However, gene or promoter trapping strate-gies often use lacZ fusions and tend to identify predom-inantly nuclear or cytoplasmic proteins due to inacti-vation of b-gal activity upon passage through the cellmembrane (Fire et al., 1990). A common property ofsecreted and transmembrane proteins that can be usedfor their isolation is the presence of an amino-terminalsignal peptide; a charged, hydrophobic sequence con-ferring passage of the protein into the endoplasmic
Grant sponsor: Genetics Institute/Wyeth Research Contract,Health Research Council of New Zealand.
*Correspondence to: Philip S. Crosier, PhD, Division of MolecularMedicine, Faculty of Medical and Health Sciences, University ofAuckland, Private Bag 92019, Auckland, New Zealand.E-mail: [email protected]
Received 5 September 2001; Accepted 18 September 2001Published online 9 November 2001
DEVELOPMENTAL DYNAMICS 222:637–644 (2001)
© 2001 WILEY-LISS, INC.DOI 10.1002/dvdy.1218

reticulum (von Heijne, 1990). Although there is littlesequence relatedness among signal peptides, their uni-formity of function can be exploited in a gene isolationstrategy termed the signal sequence trap (SST). TheSST selects cDNAs encoding secreted proteins based ontheir ability to provide a functional signal peptide to atruncated secreted enzyme. In the construction of SSTlibraries, partial cDNAs encoding signal peptides arefused in-frame to a modified invertase gene lacking itsown leader peptide in a yeast expression vector. Secre-tion of invertase is required for yeast survival on raf-finose media, thereby selecting cDNA sequences capa-ble of encoding signal peptides and directing thesecretion of invertase protein. The effectiveness of thistechnique has been demonstrated for the identificationof cDNAs encoding secreted proteins from human pe-ripheral blood mononuclear cells (PBMC) (Jacobs et al.,1997), embryonic rat brain, and whole embryo tissue(Klein et al., 1996) and the isolation of a TGF-b familymember, derriere, required for posterior developmentin Xenopus (Sun et al., 1999).
Here, we present results from an SST selection ofcDNAs from zebrafish embryonic and adult kidneyRNA, combined with expression screening by whole-mount in situ hybridization. Our aim was to identifysecreted signaling molecules involved in embryonic de-velopment, especially hematopoiesis and neurogenesis,and to study these molecules in the genetically mallea-ble zebrafish system. In two separate screens, we haveisolated cytokines, numerous enzymes, and several un-known genes potentially encoding novel signaling pro-teins. The SST technology provides a straightforwardmethod for isolating genes encoding both new andknown secreted proteins, which, in combination withzebrafish embryology and genetics, should allow rapididentification of developmental regulatory moleculesand signaling pathways.
RESULTS AND DISCUSSION
We have carried out SST screens by using librariesconstructed from two different zebrafish RNA sources.For isolation of genes encoding secreted developmentalsignaling molecules (both hematopoietic and neurogen-ic), 18–24 hr postfertilization (hpf) embryonic RNAwas used. Adult kidney RNA was used as a source ofhematopoietic genes. Half of each RNA sample wasreverse-transcribed by using a random nonamerprimer, and the resulting cDNA was size-selected andcloned into the SST vector pSUC2T7M13ORI (see Fig.1 and Experimental Procedures section). Clones sur-viving the invertase selection ranged in size from ap-proximately 200 to 900 bp and should represent 59 endfragments of genes encoding proteins with amino ter-minal signal sequences. The remainder of the RNAsample was reverse-transcribed by using an oligo-dTprimer, and the resulting cDNA was cloned into a plas-mid library vector for isolation of full-length clones,after in situ hybridization screening of the SST posi-tives (secreted ESTs, or sESTs). From the embryonic
library, 286 sESTs were identified as potential or-thologs of known genes and 93 sESTs represent novelsequences, whereas from the adult kidney library therewere 341 potential orthologs and 73 novel sequences.
Expression analysis of the embryonic library sESTswas carried out by whole-mount in situ hybridization.Digoxigenin (DIG) -labeled riboprobes were tran-scribed from PCR products that were amplified fromeach sEST clone (see Fig. 1). Approximately half of theprobes detected gene expression in 18–24 hr embryosby this method, although in some cases the expressionwas generalized or primarily in the yolk. Approxi-mately 30% of all probes detected a clearly localizedexpression pattern, such as notochord staining or re-stricted expression in the brain. Failure to detect ahybridization signal may indicate that the sEST isexpressed transiently or below the level of detection bywhole-mount in situ hybridization in the developmen-tal stages tested.
We chose the embryonic time period of 18–24 hpf forour screen because it is during this time that hemato-poietic progenitor cells develop and are released intocirculation. This time is also a period of dramatic mor-phogenetic movements, including somite development,organogenesis, and neurogenesis. DIG-labeled sESTprobes were first tested by in situ hybridization on 18-and 24-hpf embryos, and those with restricted expres-
Fig. 1. The yeast signal sequence trap vector pSUC2T7M13ORI(Jacobs et al., 1997). Transcription of the invertase gene (SUC2) is drivenby the yeast alcohol dehydrogenase (ADH) promoter, but the gene lacksits initiating methionine codon and signal sequence. This defect rendersthe plasmid unable to rescue the growth of a SUC2- yeast strain on mediacontaining sucrose or raffinose as a sole carbon source. Insertion of acDNA fragment containing an initiator methionine and functional leaderpeptide in frame with the invertase gene restores invertase secretion,allowing cell survival and selection. PCR primer sites (INV-UNI andCOLPCR3) flanking the cDNA insert and including the T7 promoter wereused to amplify positively selected secreted expressed sequence tag(sEST) clone fragments. DIG-labeled antisense riboprobes were tran-scribed from the PCR products by using T7 RNA polymerase.
638 CROSIER ET AL.
sion patterns were examined at other embryonicstages.
Expression Patterns
Expression of the positive embryonic sEST cloneswere classed into 11 broad categories, including thedeveloping blood, nervous system, somites, tail bud,and yolk (see Table 1). Groups of genes with similar,complex expression patterns may indicate the presenceof common regulatory elements and may representgenes with shared function or those acting in a commonpathway. These gene sets, termed synexpressiongroups (Gawantka et al., 1998), may include both se-creted signaling molecules and their membrane boundreceptors.
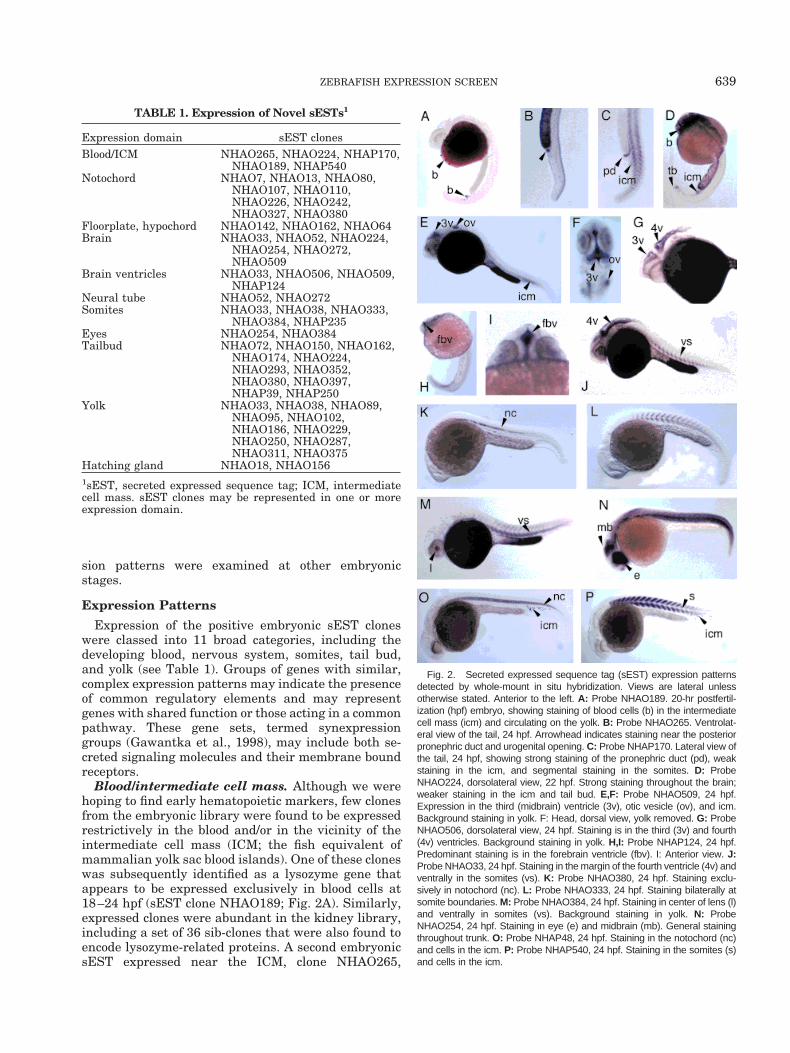
Blood/intermediate cell mass. Although we werehoping to find early hematopoietic markers, few clonesfrom the embryonic library were found to be expressedrestrictively in the blood and/or in the vicinity of theintermediate cell mass (ICM; the fish equivalent ofmammalian yolk sac blood islands). One of these cloneswas subsequently identified as a lysozyme gene thatappears to be expressed exclusively in blood cells at18–24 hpf (sEST clone NHAO189; Fig. 2A). Similarly,expressed clones were abundant in the kidney library,including a set of 36 sib-clones that were also found toencode lysozyme-related proteins. A second embryonicsEST expressed near the ICM, clone NHAO265,
TABLE 1. Expression of Novel sESTs1
Expression domain sEST clonesBlood/ICM NHAO265, NHAO224, NHAP170,
NHAO189, NHAP540Notochord NHAO7, NHAO13, NHAO80,
NHAO107, NHAO110,NHAO226, NHAO242,NHAO327, NHAO380
Floorplate, hypochord NHAO142, NHAO162, NHAO64Brain NHAO33, NHAO52, NHAO224,
NHAO254, NHAO272,NHAO509
Brain ventricles NHAO33, NHAO506, NHAO509,NHAP124
Neural tube NHAO52, NHAO272Somites NHAO33, NHAO38, NHAO333,
NHAO384, NHAP235Eyes NHAO254, NHAO384Tailbud NHAO72, NHAO150, NHAO162,
NHAO174, NHAO224,NHAO293, NHAO352,NHAO380, NHAO397,NHAP39, NHAP250
Yolk NHAO33, NHAO38, NHAO89,NHAO95, NHAO102,NHAO186, NHAO229,NHAO250, NHAO287,NHAO311, NHAO375
Hatching gland NHAO18, NHAO1561sEST, secreted expressed sequence tag; ICM, intermediatecell mass. sEST clones may be represented in one or moreexpression domain.
Fig. 2. Secreted expressed sequence tag (sEST) expression patternsdetected by whole-mount in situ hybridization. Views are lateral unlessotherwise stated. Anterior to the left. A: Probe NHAO189. 20-hr postfertil-ization (hpf) embryo, showing staining of blood cells (b) in the intermediatecell mass (icm) and circulating on the yolk. B: Probe NHAO265. Ventrolat-eral view of the tail, 24 hpf. Arrowhead indicates staining near the posteriorpronephric duct and urogenital opening. C: Probe NHAP170. Lateral view ofthe tail, 24 hpf, showing strong staining of the pronephric duct (pd), weakstaining in the icm, and segmental staining in the somites. D: ProbeNHAO224, dorsolateral view, 22 hpf. Strong staining throughout the brain;weaker staining in the icm and tail bud. E,F: Probe NHAO509, 24 hpf.Expression in the third (midbrain) ventricle (3v), otic vesicle (ov), and icm.Background staining in yolk. F: Head, dorsal view, yolk removed. G: ProbeNHAO506, dorsolateral view, 24 hpf. Staining is in the third (3v) and fourth(4v) ventricles. Background staining in yolk. H,I: Probe NHAP124, 24 hpf.Predominant staining is in the forebrain ventricle (fbv). I: Anterior view. J:Probe NHAO33, 24 hpf. Staining in the margin of the fourth ventricle (4v) andventrally in the somites (vs). K: Probe NHAO380, 24 hpf. Staining exclu-sively in notochord (nc). L: Probe NHAO333, 24 hpf. Staining bilaterally atsomite boundaries. M: Probe NHAO384, 24 hpf. Staining in center of lens (l)and ventrally in somites (vs). Background staining in yolk. N: ProbeNHAO254, 24 hpf. Staining in eye (e) and midbrain (mb). General stainingthroughout trunk. O: Probe NHAP48, 24 hpf. Staining in the notochord (nc)and cells in the icm. P: Probe NHAP540, 24 hpf. Staining in the somites (s)and cells in the icm.
639ZEBRAFISH EXPRESSION SCREEN

stained a small group of cells just posterior to the yolkextension at 24 hpf (Fig. 2B), possibly the posterior endof the pronephric duct. An sEST from the kidney li-brary, clone NHAP170, appeared to be expressed in theposterior pronephric duct and ICM, as well as in thesomites (Fig. 2C). Several other sESTs were expressedweakly in the ICM in addition to other sites. For ex-ample, sEST NHAO224 was expressed throughout thebrain, and weakly in the ICM and at the tip of the tail(Fig. 2D), whereas probe NHAO509 stained strongly inthe third (midbrain) ventricle and the otic vesicleand moderately in the ICM (Fig. 2E). Additionally,two clones were expressed in individual cells in theICM region as well as either in the notochord(NHAP48; Fig. 2O) or segmentally in the somites(NHAP540; Fig. 2P).
Brain and neural tube. sESTs expressed in thebrain can be divided into several categories, includingclones that were expressed throughout the brain(NHAO224; Fig. 2D), clones with restricted expressionin the ventricles (Fig. 2E–I), and clones with strongestexpression in the midbrain and hindbrain, extendingcaudally along the dorsal neural tube (see Fig. 4E–G).Among the sESTs showing expression in the neuroep-ithelium of the ventricles, regional differences in ex-pression also existed. For example, clone NHAO509was expressed primarily in the third ventricle and theotic vesicles (Fig. 2E,F). sEST NHAO506 was ex-pressed in both the third and the fourth ventricles (Fig.2G), whereas the kidney library clone NHAP124 wasmost strongly expressed in the forebrain ventricle (Fig.2H,I). Embryonic clone NHAO33 is expressed at themargin of the fourth (hindbrain) ventricle, as well as inthe ventral region of the somites (Fig. 2J).
Notochord. Several sESTs were expressed in thenotochord (Table 1; Figs. 2K,O, 3K). The notochord, astructural feature of the phylum Chordata, is a sourceof signals that pattern neural, somitic, and dermomyo-tomal tissue by specifying ventral neural tube fatesand inhibiting dorsal fates (Goulding et al., 1993). Sig-naling molecules that are expressed in the notochordinclude Sonic hedgehog (shh), noggin, and the bonemorphogenetic proteins BMP-2 and BMP-7 (Krauss etal., 1993; Dale et al., 1997; McMahon et al., 1998).Some of our notochord-expressed clones may encodenovel notochord signaling molecules, whereas othersmay represent presently unknown extracellular matrixor adhesion molecules specifically secreted by the no-tochord.
Somites. Intercellular signaling underlies segmen-tation of the vertebrate body, including subdivision ofparaxial mesoderm into somites. Several sESTs wereexpressed in or between the somites and may haveroles in differentiation of somitic tissue and/or providesignals for neural crest migration. Some of these wererestricted to the ventral region of the somites, part ofwhich gives rise to sclerotome (vertebral cartilage)(NHAO33, Fig. 2J; NHAO384, Fig. 2M). NHAO384 wasalso expressed in the center of the lens (see below),
whereas NHAO33 was expressed in the brain. Oneembryonic sEST showed restricted expression at thesomite boundaries, possibly in the intersegmentalblood vessels (NHAO333, Fig. 2L). sEST cloneNHAP170, from the kidney library, appeared to beexpressed segmentally in the somites, as well as in thepronephric duct and ICM (Fig. 2C), whereas NHAP540was also expressed in the somites and individual cellslocated in the posterior ICM (Fig. 2P).
Eyes. None of the sESTs appeared to stain exclu-sively in the eye; however, several clones were found tobe expressed in the lens in addition to other sites. Forexample, clone NHAO254 (Fig. 2N) was strongly ex-pressed in the eye, midbrain, and throughout thesomites. Clone NHAO384 was expressed in the centerof the lens (Fig. 2M) as well as in the ventral portion ofthe somites.
Fig. 3. Differential secreted expressed sequence tag (sEST) expres-sion in the tail bud. A–C: Probe NHAO72. Staining is strong in theposterior tail bud (tb), beginning around 12 hr postfertilization (hpf), andcompletely absent from anterior regions. A: 12 hpf, posterior view. Thehead (h) is pointing down, under the yolk. B: 12 hpf, lateral view. Anterioris to the left. C: 24 hpf, tail lateral view. Staining remains in the distal tipof the tail, outside of the notochord. D–F: Probe NHAO150. Stainingsurrounds the tail bud at 12 hpf (D: 12 hpf, posterior view; and E, 12 hpf,lateral view) and is absent from the extreme posterior tip of the tail at 24hpf (arrowhead in F). G: Probe NHAO397, 24 hpf. Expression throughoutthe posterior tail and along the caudal fin ectoderm (cfe). H–J: ProbeNHAP39. H: 18 hpf. Strong staining around the posterior notochord (nc),diminishing anteriorly. Expression in tail at 22 hpf (I) and 24 hpf (J).Caudal fin ectoderm (cfe) also stains. K: Probe NHAP18. Tail of 24-hpfembryo with staining in the posterior notochord.
640 CROSIER ET AL.

Posterior tail. Another large set of sESTs was ex-pressed in the region of the distal tail bud (see Fig. 3).These clones may represent a synexpression group ofsignaling molecules required for development in verte-brates. Fate mapping of the zebrafish tail bud hasshown that this tissue contributes to the formation oftail and trunk structures as far anterior as somite11–12, and that it is not a homogenous population ofcells (Kanki and Ho, 1997). Restricted expression ofgenes such as no tail (ntl) (Schulte-Merker et al., 1994),snail (Thisse et al., 1993), and eve1 (Joly et al., 1993) inthe posterior domain of the tail bud is consistent withthe mesodermal fates of cells originating at this site. InXenopus, the tip of the tail is a derivative of the blas-topore lip (the embryonic organizer), and retains potentorganizer activity, including the ability to induce for-mation of notochord (Gont et al., 1993). Genes markingtip of tail in Xenopus include Xnot2, a homeobox generelated to Xnot. Xnot is expressed in the dorsal blas-topore lip and notochord and is regulated by fibroblastgrowth factor and activin (von Dassow et al., 1993). Inzebrafish, the floating head mutation abrogates thezebrafish homolog of Xnot, and results in abnormalnotochord development (Talbot et al., 1995). Brachyuryis expressed in both notochord and tail bud in mouseand zebrafish (Herrmann, 1991; Schulte-Merker et al.,1992). Some of the sESTs were restricted to the poste-rior notochord (e.g., NHAP18, Fig. 3K), whereas otherswere expressed outside of the notochord at the tip ofthe tail (e.g., NHAO72; Fig. 3A–C and NHAP39; Fig.3H–J). Varied expression of sESTs in this region isconsistent with the idea that the tail bud does notconstitute a blastema, or uniform population of cells(Griffith et al., 1992). Rather, cell fate restrictions haveoccurred and patterning events are likely to be pro-ceeding due to communication between cells expressingdifferent signaling molecules. Analysis of expression ofthe sESTs and their corresponding genes in zebrafishmutants defective in posterior development should as-sist in unraveling these signaling pathways.
Floorplate and hypochord. Two sESTs were ex-pressed in midline structures adjacent the notochord,termed the floorplate and hypochord. The floorplateconsists of epithelial cells located at the ventral midlineof the neural tube, above the notochord. Its differenti-ation depends on signals from the notochord. The floor-plate is implicated in the control of neural cell patternand axonal pathfinding and expresses adhesion mole-cules such as F-spondin (Klar et al., 1992). The hypo-chord is a transient rod-like structure situated imme-diately under the notochord and is linked with thesomites by means of extracellular matrix fibrils(Lofberg and Collazo, 1997). The hypochord may beinvolved in positioning the dorsal aorta, which formsunder it. The secreted protein collagen 2 a1 (col2a1) isexpressed in both hypochord and floorplate (Yan et al.,1995).
Clones NHAO64 and NHAO162 (Fig. 4A–D) wereexpressed in the hypochord and floorplate, as well as
the posterior notochord. NHAO64 and its full-lengthcDNA show sequence similarity to follistatin-relatedprotein (FRP), originally identified as a gene up-regu-lated by TGF-b in osteoblastic cell lines (Shibanuma etal., 1993). In Xenopus the FRP ortholog is localized inthe Spemann organizer, notochord, floor plate, hypo-chord, and somites (Okabayashi et al., 1999). Expres-sion of the avian ortholog (termed follistatin-like pro-tein or Flik) has been shown to require signalsemanating from the neural tube (Amthor et al., 1996).
Orthologs of Known Genes
In addition to the novel genes identified in the SSTscreens, we have isolated several probable zebrafishorthologs of genes identified in other organisms (Table2). From sequence analysis of positive clones from theembryonic SST library, we have identified a member ofthe midkine/pleiotrophin family of heparin bindinggrowth factors (Muramatsu, 1993) which is expressedpredominantly in dorsal aspects of the midbrain andhindbrain of 18–28 hpf embryos clone NHAO72 (seeFig. 4E). A related but not identical clone (NHAO52)shows a similar expression pattern, as does the novel
Fig. 4. Comparison of unknown secreted expressed sequence tags(sESTs) with known genes. A,B: In situ hybridization by using a full-length cDNA corresponding to sEST NHAO64, which shows strong sim-ilarity to follistatin-related protein (FRP). Expression is seen in the pos-terior notochord (nc) and along the hypochord (hc) and floorplate (fp).The caudal fin ectoderm (cfe) also exhibits weak staining. Unknown sESTNHAO162 C,D: A similar pattern. (A,B,D: 24 hr postfertilization [hpf]; C:20 hpf). E–G: Clones with expression in dorsal brain regions and alongthe dorsal neural tube at 24 hpf. E: In situ hybridization using a full-lengthcDNA corresponding to sEST NHAO12, a zebrafish midkine. Expressionis in the olfactory placode (op), telencephalon (t), cerebellum (c), andhindbrain (hb; arrowheads indicate segmental staining of the rhom-bomeres). Staining continues caudally along the dorsal neural tube,diminishing in the trunk. F: Midkine-related sEST NHAO52 shows asimilar expression pattern, as does unknown sEST NHAO272 (G).
641ZEBRAFISH EXPRESSION SCREEN

sEST NHAO272. Orthologous genes encoding knownsignaling molecules isolated from the embryonic SSTlibrary include Wnt5, Wnt11, and follistatin-relatedprotein (FRP; see Fig. 4A). Zebrafish wnt5 and wnt11have been independently isolated by others and shownto harbor mutations in the pipetail and silberblick phe-notypes, respectively (Rauch et al., 1997; Heisenberg etal., 2000). The adult kidney library has yielded a po-tential chemokine with similarity to human SDF-1(sEST clone NHAP454), a pre-B cell stimulatory mole-cule and T-lymphocyte chemoattractant (Tashiro et al.,1993; Nagasawa et al., 1996). Other cDNAs encodingknown secreted proteins isolated from the SST librar-ies include apolipoprotein A1 and D, cysteine protein-ase, and cathepsin L.
Genetic Mapping
Mapping of the sESTs will facilitate identification ofcandidate genes affected in zebrafish mutants, whosemap positions are either known or are in the process ofbeing determined. In addition, it is now apparent thatsyntenic groups (groups of genes on a single chromo-some) have been conserved throughout vertebrate evo-lution (Postlethwait et al., 1998). It is possible thatmapped genes in zebrafish may be identified as candi-date disease genes based on their map location corre-sponding to a known human disease locus. A selectionof sEST clones are being mapped by using either 59 or39 (if a corresponding full-length cDNA is available)primers. Assignment of map position was possiblewhen only 59 sequence was available.
CONCLUSIONS
Efforts are in place toward whole genome sequencingof zebrafish. Once genes are identified, the next chal-lenge is to ascertain their function. It has become ap-parent that important clues can be gained from knowl-edge of gene expression pattern, as many genes withdevelopmental regulatory function are expressed in alocalized manner. Zebrafish is particularly well suitedto this type of analysis. In situ hybridization screening
can identify differentially expressed developmentalregulators regardless of functional redundancy, whichmay cause them to be missed in screens for lethalmutant phenotypes. It is also a potential method ofidentifying unknown genes affected in zebrafish mu-tants. Genes expressed in multiple tissues may corre-spond to composite phenotypes of mutants and may becandidates for the affected gene or genes acting in thesame pathway. In addition, groups of genes withshared, complex expression patterns, termed synex-pression groups, may represent genes with sharedfunction, as is evident with the components of Notch-Delta signaling in Xenopus (reviewed in Niehrs, 1997).This finding raises the possibility that, by identifyingsynexpression groups, entire molecular pathways maybe identified, regardless of the functional redundancyof individual components. In this study, we have com-bined a yeast signal sequence trap selection for se-creted factors with zebrafish in situ hybridizationscreening to focus on potentially new developmentalsignaling molecules. Our screen has identified severalnovel genes and several possible synexpression groups.This type of screening approach clearly complementssystematic mutagenesis in the effort to understandgene function on a large scale.
EXPERIMENTAL PROCEDURESLibrary Construction
The signal sequence trap (SST) vector pSUC2T7M13ORIwas designed to identify sequences in cDNA clones thatmediate transport of proteins into the secretion path-way (Jacobs et al., 1997). The vector carries an inver-tase gene (SUC2), lacking both its initiating methio-nine codon and its signal peptide. Transcription ofinvertase is mediated by the yeast alcohol dehydroge-nase promoter, but the invertase translation and secre-tion defect renders the plasmid unable to rescue thegrowth of a SUC2- yeast strain on media containingsucrose or raffinose as the sole carbon source. Insertionof a cDNA fragment containing an initiator methionineand functional leader peptide in frame with the inver-
TABLE 2. sESTs Homologous to Known Genes1
sEST clones Homology Expression at 24 hpf Percentage identityNHAO64 Follistatin-related protein Hypochord, floorplate, tailbud notochord, caudal
fin ectoderm98(AI588548)2
NHAO12/429 Midkine (human) Dorsal brain and neural tube 98(AW280954)NHAP454 SDF-1 chemokine Brain, thymic primordia, pronephric ducts,
lateral line primordia98(BG799709)
NHAO63 wnt11 Posterior notochord 99(AF067429)NHAO173 wnt5 n.d. 98(U51268)NHAO101 Apolipoprotein A-1 n.d. 98(BG799224)NHAO122 Cathepsin-L n.d. 99(AI476900)NHAO22 Cysteine proteinase n.d. 98(BF158340)NHAO161 grp78 Hypochord, floorplate, tailbud notochord, caudal
fin ectoderm98(AW421396)
NHAO189 Lysozyme Circulating macrophages 98(AF402599)1sEST, secreted expressed sequence tag; hpf, hours postfertilization; n.d., not done.2Accession number of sequence with highest percentage identity to zebrafish sEST clone.
642 CROSIER ET AL.

tase gene restores invertase secretion, allowing cellsurvival and selection.
One microgram of poly A1 RNA was isolated fromadult zebrafish kidney and 18- to 24-hpf embryos andseparately reverse transcribed to cDNA by using Su-perScript II reverse transcriptase (Gibco/Life Technol-ogies). The oligonucleotide primer 59-CGATTGAAT-TCTAGACCTGCCTCGAGNNNNNNNNN-39 (N denotesG, A, T, or C) consisted of nine randomized bases at the39 end joined to an XhoI restriction site to allow fordirectional cloning. The resulting cDNA was ligated toEcoRI adapters, digested with XhoI, and size selectedto isolate cDNAs between 300 and 900 bp. The cDNAwas then ligated to pSUC2T7M13ORI vector predi-gested with EcoRI and XhoI to create a library of ran-dom-primed cDNA fragments cloned 59 to the trun-cated invertase gene. The cDNA libraries constructedfrom zebrafish embryonic and kidney RNAs inpSUC2T7M13ORI were transformed into yeast strainYT455 (Kaiser et al., 1987), which was converted toTRP- and then to URA- (Jacobs et al., 1997), and plas-mids were isolated from colonies that survived theinvertase selection. Positive clones were sequencedrandomly and analyzed for similarity to known genes.
Probe Synthesis and In Situ Hybridization
Selected expressed SST clones (sESTs) were PCRamplified by using primers flanking the insert andincluding the T7 promoter, allowing for transcription ofDIG-labeled antisense RNA probes. (sEST clones gen-erated as part of this investigation are available toacademic researchers from Genetics Institute/Wyeth-Ayerst Research.) Initial in situ hybridizations wereperformed according to Oxtoby and Jowett (1993) onbatches of embryos staged 18–24 hpf to test the probesand determine expression patterns. Clones were se-lected for further analysis based on specific expressionin blood cell precursors or the nervous system. Full-length cDNAs have been isolated for selected sESTclones for further characterization by in situ hybridiza-tion, morpholino knock-down studies, and analysis ofexpression in mutants.
Mapping
sEST clones were mapped to the zebrafish genomemap by single-strand conformation polymorphismanalysis (SSCP) or restriction fragment length poly-morphism (RFLP) mapping. Genomic DNA from C32and SJD parental strains were amplified by usingprimers specific to 39UTR sequences of full-lengthclones, or 59 sequences of the sESTs. Identified SSCPsor RFLPs were scored on either the MOP cross haploidmapping panel or the HS doubled haploid diploid panelfor genotyping (Postlethwait et al., 2000; Woods et al.,2000).
ACKNOWLEDGMENTS
We acknowledge the strong support of Dr. SteveClark during this work. Graca Cabreira-Hansen, Tim-
othy Fountaine, and Ana Ramachandran provided ex-cellent technical assistance with this project. We thankKristina Maconaghie for help with the preparation ofthis manuscript. sEST clones generated as part of thisinvestigation are available to academic researchersfrom Genetics Institute/Wyeth-Ayerst Research.
REFERENCES
Amthor H, Connolly D, Patel K, Brand-Saberi B, Wilkinson DG,Cooke J, Christ B. 1996. The expression and regulation of follistatinand a follistatin-like gene during avian somite compartmentaliza-tion and myogenesis. Dev Biol 178:343–362.
Babin PJ, Thisse C, Durliat M, Andre M, Akimenko MA, Thisse B.1997. Both apolipoprotein E and A-I genes are present in a non-mammalian vertebrate and are highly expressed during embryonicdevelopment. Proc Natl Acad Scie U S A 94:8622–8627.
Dale JK, Vesque C, Lints TJ, Sampath TK, Furley A, Dodd J, PlaczekM. 1997. Cooperation of BMP7 and SHH in the induction of fore-brain ventral midline cells by prechordal mesoderm. Cell 90:257–269.
Driever W, Fishman MC. 1996. The zebrafish: heritable disorders intransparent embryos. J Clin Invest 98:S41–S46.
Eberl DF, Hilliker AJ. 1988. Characterization of X-linked recessivelethal mutations affecting embryonic morphogenesis in Drosophilamelanogaster. Genetics 118:109–120.
Evans MJ, Carlton MB, Russ AP. 1997. Gene trapping and functionalgenomics. Trends Genet 13:370–374.
Fire A, Harrison SW, Dixon D. 1990. A modular set of lacZ fusionvectors for studying gene expression in Caenorhabditis elegans.Gene 93:189–198.
Gawantka V, Pollet N, Delius H, Vingron M, Pfister R, Nitsch R,Blumenstock C, Niehrs C. 1998. Gene expression screening in xe-nopus identifies molecular pathways, predicts gene function andprovides a global view of embryonic patterning. Mech Dev 77:95–141.
Gont LK, Steinbeisser H, Blumberg B, de Robertis EM. 1993. Tailformation as a continuation of gastrulation: the multiple cell pop-ulations of the Xenopus tail bud derive from the late blastopore lip.Development 119:991–1004.
Goulding MD, Lumsden A, Gruss P. 1993. Signals from the notochordand floor plate regulate the region-specific expression of two Paxgenes in the developing spinal cord. Development 117:1001–1016.
Grammer TC, Liu KJ, Mariani FV, Harland RM. 2000. Use of large-scale expression cloning screens in the Xenopus laevis tadpole toidentify gene function. Dev Biol 228:197–210.
Griffith CM, Wiley MJ, Sanders EJ. 1992. The vertebrate tail bud:three germ layers from one tissue. Anat Embryol (Berl) 185:101–113.
Haffter P, Granato M, Brand M, Mullins MC, Hammerschmidt M,Kane DA, Odenthal J, van Eeden FJ, Jiang YJ, Heisenberg CP,Kelsh RN, Furutani-Seiki M, Vogelsang E, Beuchle D, Schach U,Fabian C, Nusslein-Volhard C. 1996. The identification of geneswith unique and essential functions in the development of thezebrafish, Danio rerio. Development 123:1–36.
Heisenberg C-P, Tada M, Rauch G-J, Saude L, Concha ML, Geisler R,Stemple DL, Smith JC, Wilson SW. 2000. Silberblick/Wnt11 medi-ates convergent extension movements during zebrafish gastrula-tion. Nature 405:76–81.
Herrmann BG. 1991. Expression pattern of the Brachyury gene inwhole-mount TWis/TWis mutant embryos. Development 113:913–917.
Hope IA, Arnold JM, McCarroll D, Jun G, Krupa AP, Herbert R. 1998.Promoter trapping identifies real genes in C. elegans. Mol GenGenet 260:300–308.
Jacobs KA, Collins-Racie LA, Colbert M, Duckett M, Golden-Fleet M,Kelleher K, Kriz R, Lavallie ER, Merberg D, Spaulding V, Stover J,Williamson MJ, McCoy JM. 1997. A genetic selection for isolatingcDNAs encoding secreted proteins. Gene 198:289–296.
Joly JS, Joly C, Schulte-Merker S, Boulekbache H, Condamine H.1993. The ventral and posterior expression of the zebrafish ho-
643ZEBRAFISH EXPRESSION SCREEN

meobox gene eve1 is perturbed in dorsalized and mutant embryos.Development 119:1261–1275.
Kaiser CA, Preuss D, Grisafi P, Botstein D. 1987. Many randomsequences functionally replace the secretion signal sequence ofyeast invertase. Science 235:312–317.
Kanki JP, Ho RK. 1997. The development of the posterior body inzebrafish. Development 124:881–893.
Klar A, Baldassare M, Jessell TM. 1992. F-spondin: a gene expressedat high levels in the floor plate encodes a secreted protein thatpromotes neural cell adhesion and neurite extension. Cell 69:95–110.
Klein RD, Gu Q, Goddard A, Rosenthal A. 1996. Selection for genesencoding secreted proteins and receptors. Proc Natl Acad Sci U S A93:7108–7113.
Krauss S, Concordet JP, Ingham PW. 1993. A functionally conservedhomolog of the Drosophila segment polarity gene hh is expressed intissues with polarizing activity in zebrafish embryos. Cell 75:1431–1444.
Lofberg J, Collazo A. 1997. Hypochord, an enigmatic embryonic struc-ture: study of the axolotl embryo. J Morphol 232:57–66.
Lynch AS, Briggs D, Hope IA. 1995. Developmental expression pat-tern screen for genes predicted in the C. elegans genome sequencingproject. Nat Genet 11:309–313.
McMahon JA, Takada S, Zimmerman LB, Fan CM, Harland RM,McMahon AP. 1998. Noggin-mediated antagonism of BMP signal-ing is required for growth and patterning of the neural tube andsomite. Genes Dev 12:1438–1452.
Mitchell KJ, Pinson KI, Kelly OG, Brennan J, Zupicich J, Scherz P,Leighton PA, Goodrich LV, Lu X, Avery BJ, Tate P, Dill K, Pang-ilinan E, Wakenight P, Tessier-Lavigne M, Skarnes WC. 2001.Functional analysis of secreted and transmembrane proteins criti-cal to mouse development. Nat Genet 28:241–249.
Muramatsu T. 1993. Midkine (MK), the product of a retinoic acidresponsive gene, and pleiotrophin constitute a new protein familyregulating growth and differentiation. International J Dev Biol37:183–188.
Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S,Kitamura Y, Yoshida N, Kikutani H, Kishimoto T. 1996. Defects ofB-cell lymphopoiesis and bone-marrow myelopoiesis in mice lackingthe CXC chemokine PBSF/SDF-1. Nature 382:635–638.
Niehrs C. 1997. Perspectives in genome technology; gene-expressionscreens in vertebrate embryos: more than meets the eye. GenesFunction 1:229–231.
Nusslein-Volhard C, Wieschaus E. 1980. Mutations affecting segmentnumber and polarity in Drosophila. Nature 287:795–801.
Okabayashi K, Shoji H, Onuma Y, Nakamura T, Nose K, Sugino H,Asashima M. 1999. cDNA cloning and distribution of the Xenopusfollistatin-related protein. Biochem Biophys Res Commun 254:42–48.
Oxtoby E, Jowett T. 1993. Cloning of the zebrafish krox-20 gene(krx-20) and its expression during hindbrain development. NucleicAcids Res 21:1087–1095.
Postlethwait JH, Yan Y-L, Gates MA, Horne S, al. e. 1998. Vertebrategenome evolution and the zebrafish gene map. Nat Genet 18:345–349.
Postlethwait JH, Woods IG, Ngo-Hazelett P, Yan Y-L, Kelly PD, ChuF, Huang H, Hill-Force A, Talbot WS. 2000. Zebrafish comparativegenomics and the origins of vertebrate chromosomes. Genome Res10:1890–1902.
Rauch G-J, Hammerschmidt M, Blader P, Schauerte HE, Strahle U,Ingham PW, McMahon AP, Haffter P. 1997. Wnt5 is required fortail formation in the zebrafish embryo. Cold Spring Harbor SympQuant Biol 62:227–234.
Schulte-Merker S, Ho RK, Herrmann BG, Nusslein-Volhard C. 1992.The protein product of the zebrafish homologue of the mouse T geneis expressed in nuclei of the germ ring and the notochord of theearly embryo. Development 116:1021–1032.
Schulte-Merker S, van Eeden FJ, Halpern ME, Kimmel CB, Nusslein-Volhard C. 1994. no tail (ntl) is the zebrafish homologue of themouse T (Brachyury) gene. Development 120:1009–1015.
Shibanuma M, Mashimo J, Mita A, Kuroki T, Nose K. 1993. Cloningfrom a mouse osteoblastic cell line of a set of transforming-growth-factor-beta 1-regulated genes, one of which seems to encode a fol-listatin-related polypeptide. Eur J Biochem 217:13–19.
Skarnes WC, Moss JE, Hurtley SM, Beddington RS. 1995. Capturinggenes encoding membrane and secreted proteins important formouse development. Proc Natl Acad Sci U S A 92:6592–6596.
Sun BI, Bush SM, Collins-Racie LA, La Vallie ER, DiBlasio-SmithEA, Wolfman NM, McCoy JM, Sive HL. 1999. derriere: a TGF-betafamily member required for posterior development in Xenopus.Development 126:1467–1482.
Talbot WS, Trevarrow B, Halpern ME, Melby AE, Farr G, Postleth-wait JH, Jowett T, Kimmel CB, Kimelman D. 1995. A homeoboxgene essential for zebrafish notochord development. Nature 378:150–157.
Tashiro K, Tada H, Heilker R, Shirozu M, Nakano T, Honjo T. 1993.Signal sequence trap: a cloning strategy for secreted proteins andtype I membrane proteins. Science 261:600–603.
Tate P, Lee M, Tweedie S, Skarnes WC, Bickmore WA. 1998. Captur-ing novel mouse genes encoding chromosomal and other nuclearproteins. J Cell Sci 111:2575–2585.
Thisse C, Thisse B, Schilling TF, Postlethwait JH. 1993. Structure ofthe zebrafish snail1 gene and its expression in wild-type, spadetailand no tail mutant embryos. Development 119:1203–1215.
Thut CJ, Rountree RB, Hwa M, Kingsley DM. 2001. A large-scale insitu screen provides molecular evidence for the induction of eyeanterior segment structure by the developing lens. Dev Biol 231:63–76.
von Dassow G, Schmidt JE, Kimelman D. 1993. Induction of theXenopus organizer: expression and regulation of Xnot, a novel FGFand activin-regulated homeo box gene. Genes Dev 7:355–366.
von Heijne G. 1990. The signal peptide. J Membr Biol 115:195–201.Wilson C, Pearson RK, Bellen HJ, O’Kane CJ, Grossniklaus U, Ge-
hring WJ. 1989. P-element-mediated enhancer detection: an effi-cient method for isolating and characterizing developmentally reg-ulated genes in Drosophila. Genes Dev 3:1301–1313.
Woods IG, Kelly PD, Chue F, Ngo-Hazelett N, Yan Y-L, Huang H,Postlethwait JH, Talbot WS. 2000. A comparative map of the ze-brafish genome. Genome Res 10:1903–1914.
Yan YL, Hatta K, Riggleman B, Postlethwait JH. 1995. Expression ofa type II collagen gene in the zebrafish embryonic axis. Dev Dyn203:363–376.
Zambrowicz BP, Friedrich GA. 1998. Comprehensive mammalian ge-netics: history and future prospects of gene trapping in the mouse.Int J Dev Biol 42:1025–1036.
Zhang JS, Duncan EL, Chang AC, Reddel RR. 1998. Differentialdisplay of mRNA. Mol Biotechnol 10:155–165.
644 CROSIER ET AL.





























