Embed Size (px)
Citation preview

41Cassava Genetic Improvement: Omics Approachesfor Facing Global ChallengesYoshimi Umemura, Rane Jagadish, Motoaki Seki, Yoshinori Utsumi,Jarunya Narangajavana, and Manabu Ishitani
Cassava is a staple food crop that ensures food security and is a source of incomegeneration for poor farmers in the tropics of many Asian and African countries thatare highly prone to harsh and unfavorable environments especially in the context ofclimate changes. Improving tolerance to abiotic stresses such as drought in this cropis a big challenge as it has substantial tolerance to drought compared to other crops.There has been little investment in employing advanced breeding technology forgenetic improvement of cassava, though we have now an increased access toemerging advanced tools in genomics, transcriptomics, phenomics, proteomics,and metabolomics for that can enhance our capacity to understand stress responsesand tolerance in plants.
In this chapter, we have reviewed challenges and opportunities for improvingproductivity of cassava by integrating omics with conventional breeding. The chapterhighlights efforts and vision of International Center for Tropical Agriculture (CIAT;http://www.ciat.cgiar.org/) toward a comprehensive evaluation of germplasm, estab-lishment of the transformation and phenotyping platform, and development ofgenomic tools for deriving the requisite information for developing effective strat-egies for molecular breeding for high-yield and high-value cassava under stressconditions. The chapter also deals with possible strategies to accelerate developmentof elite cassava cultivars by integration of the advanced technologies.
41.1Introduction
Cassava is an important tropical crop both for food security and for incomegeneration for poor farmers in many Asian and African countries, where itsproduction is more than 240 million ton a year [1]. This crop serves as the primaryfood source for more than 750 million people across the world. Cassava is welladapted to semiarid conditions and it needs adequate soil moisture mainly during
Improving Crop Resistance to Abiotic Stress, First Edition.Edited by Narendra Tuteja, Sarvajeet Singh Gill, Antonio F. Tiburcio, and Renu Tuteja� 2012 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2012 by Wiley-VCH Verlag GmbH & Co. KGaA.
j1049

planting. After establishment, it can withstand several months of drought.Generally, it is not irrigated, but in some areas responds markedly to irrigation [2].However, severe limitation of soil moisture can bring down the cassava yieldsignificantly [3–5]. So, the task of improving the productivity of this drought-tolerant crop under soil moisture stress environments is more challenging thanfor other crops.
In several investigations supported by the Generation Challenge Programme(GCP: http://www.generationcp.org/), in Brazil, Colombia, Tanzania and Ghana,contrasting cassava varieties have been screened and evaluated under droughtenvironments and physiological traits associated with drought tolerance have beenidentified [6–8]. Among these, there are traits related to developmental regulationthat permits the most drought-tolerant lines to optimally partition resources forstorage root initiation and maintenance during drought. However, despite substan-tial accumulation of basic knowledge of cellular and molecular level, there is nosignificant progress toward development of drought-tolerant varieties mainlybecause of limited success in dissecting the complex mechanisms underlyingtolerance to drought.
Recent remarkable innovations in omics-based research such as phenomics,genomics, transcriptomics, and application development can provide us new oppor-tunities to resolve the complexities associated with drought tolerance. Furthermore,these advances in science can also provide us crucial resources to promote researchboth in crop plants and inmodel plants [9–12]. All kinds of �omics,� arrays, and high-throughput technologies can now make it possible to carry out genetic and genomicanalysis for the investigation of gene function associatedwith phenotypic changes ona larger scale to accelerate crop breeding.
In this chapter, we have reviewed the recent efforts to improve cassavaespecially for drought tolerance and have presented perspectives for integratingthe knowledge that has been accumulated through the traditional research and thecurrent advanced technologies to improve cassava genotypes with an efficient andtargeted approach.
41.2What Makes the Cassava More Tolerant to Drought?
Cassava tolerates prolonged drought that often exceeds 5 months. This has beenattributed to partial closure of stomata, deep rooting systems, and small leaf canopy.These traits make cassava a desirable and adaptable crop, as a source for food andfeed, in the hot and dry tropics that are highly prone to predicted adverse globalclimate changes [13, 14].
Recent review by El-Sharkawy [8] reveals several attempts made at understandingthe mechanisms involved in the drought tolerance in cassava and its possibleapplication in breeding programs for enhancing its productivity [8]. Some of thetraits relevant to drought tolerance have been listed in Table 41.1. The challenge offurther improving cassava for the productivity of harvestable roots now largely lies in
1050j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

our efforts to phenotype known and unknown traits of drought tolerance. This canbe accomplished through optimized and novel phenotyping techniques that cangenerate highly reliable data on plant response and stress environments for com-plementing genomic information.
Table 41.1 Plant traits contributing to drought tolerance in cassava.
Trait Contribution to drought andproductivity
Conditions under which it isuseful
Photosynthesis High rate of photosynthesisunder favorable conditions andmaintenance even under pro-longed drought conditions andrecovery
General
Tight stomatal control Protection from both low soilmoisture and high evaporativedemand
Hot environment with no soilmoisture in deeper soil profile
Plant vigor High vegetative vigor enablesplants to recover from drought
Intermediate drought conditions
C3–C4 intermediatemechanisms
These characteristics, collective-ly, underpinned the high photo-synthetic rate (Pn) in normal air(Pn> 40mmolCO2m
�2 s�1) inhigh irradiances (>1800mmolm�2 s�1 of photosynthetic activeradiation), high leaf temperaturefrom 30 to 40 �C, and in highatmospheric humiditiesobserved in cassava grown infavorable environments [6, 15]
Environments where high evap-orative demand exist
Shoot morphologicaladjustments
During prolonged water deficits,cassava reduces its canopy byshedding older leaves and form-ing smaller new leaves leading toless light interception
Intermediate drought
Leaf area index Optimum leaf area for longerperiod during the drought andafter recovery
Normal and intermediatedrought conditions
Deep roots Cassava slowly extracts soilmoisture from deeper layers ofsoil
Drought environments withmoisture stored in deeper layersof soil
Predawn leaf waterpotential
Relative to unstressed plants Intermediate drought
41.2 What Makes the Cassava More Tolerant to Drought? j1051

41.3Phenomics for Trait Discovery
Breeders� eye approach with large dependence on empirical selection for yield,disease, and pest resistance has significantly contributed to historical jump in foodproduction during the Green Revolution in the past. However, scope for continuingthis approach is gradually diminishing as evident from the recent yield plateau inmany of the staple foods such as rice, wheat, and maize. The rapidly developinggenomic knowledge and genotyping capacity can offer us opportunities to improvecrop varieties in a much more targeted way to grow them in harsh environments, formore productivity with fewer inputs. Genomics and transcriptomics in combinationwith bioinformatics can offer great opportunity to generate knowledge about thefunction of the genome, unraveling genomic pathways, but more importantly, toidentify useful diversity, genes, and alleles and transfer these rapidly into adaptedvarieties. However, these new approaches and tools need to be sufficiently comple-mented by screening protocols that can facilitate generation of high-quality data onplant phenotypes under given stress environment. This necessarily should includeunderstanding and management of stress environment and rapid and noninvasiveprocedures to measure response of the large number of genetically diverse genotypescreated through conventional breeding or genetic engineering approaches. Theseefforts with new terminology, �phenomics,� have now become the focal point ofmuchof the agricultural research aiming to get benefits from advances in genomics.
One of the major tasks in phenomics is the determination of plant responses tostress in well-defined environment that can clearly reveal the genetic variation. Thiswill lead to trait discovery that can be eventually associated with gene(s) throughquantitative trait locus (QTL) cloning or/and transgenic approach. In addition, thephenomics should focus on inherent gaps in understanding the magnitude ofdrought stress in terms of duration of soil moisture stress and level of soil moistureduring a specific growth period in crops such as cassava.
In a classical example of a field evaluation, it was shown that the reduction in rootyield in response to 120 days of drought imposed 60 days after planting could reducethe yield marginally compared to the control in which soil moisture level wasmaintained at field capacity. In this experiment, some of the genotypes had moregrain yield compared to control even after exposure to the drought [8]. Furthermore,compared to the control, water stress across all varieties caused a reduction in totalbiomass by 12%, no change in fresh root yields, a reduction in dried root yields by3.4%, a reduction in drymatter contents by 3.3%, and an increase in the harvest indexby 10%.Without any doubt, these data clearly demonstrate cassava�s ability to tolerateprolonged water stress when it is induced gradually at an early stage of growth andalso its ability to recover fromdrought.However, themagnitude of drought stress, theduration, and critical phase of growth in such experiments need a review as possiblythe soil moisture deficit was not sufficient enough to cause substantial impact onproductivity of cassava genotypes, to elucidate genetic variation in the response,which is fundamental to further genetic improvement. Alternatively, control plantsdid not have sufficient scope to express their yield potential. Hence, we suggest that
1052j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

the challenge of further improving the productivity of cassava under moisture stressshould consider a reoriented phenotyping protocol.
1) To dissect traits contributing to components of crop productivity under soilmoisture deficit [16].
2) Cassava root yield¼water uptake�water use efficiency� partitioning to rootsthat can be harvested.
3) To impose drought at critical stage of growth that determines productivity of crop.4) The time when number of productive roots determined.5) Root bulking.6) To reduce the influence of phenological variation in understanding genetic
variation in drought response, through staggered sowing.7) To differentiate genetic variation in recovery from drought stress.8) To understand the yield gap in the target environment that needs to be covered by
genetic improvement of traits associated with high root yield. The traits listed inTable 41.2 may be considered depending on the nature of drought stress.
41.4How Cassava Genomics Tool will help Develop Drought-Tolerant Variety
In 2007,CIATandRIKEN initiated an international collaborative research project andsince 2009, a 3-year cassava research project, between RIKEN (Japan, PI: Dr.MotoakiSeki), CIAT (Colombia, PI: Dr. Manabu Ishitani), andMahidol University (Thailand,PI: Dr. Jarunya Narangajavana), which is supported by the Japanese government, hasstarted to develop an integrated cassava genome analysis platform for the globalcassava research community and to apply it to molecular breeding for cassavaimprovement. In the collaborative work, the following cassava genomics tools arebeing developed: (1) large-scale collection of full-length cDNAs ([17], unpublisheddata), (2) cassava oligoarray analysis platform containing more than 30 000 genes,and (3) cassava database of international standard. For the collection of full-length cDNA clones, we have selected three genetically diverse varieties, namely,KU50 (Manihot esculenta Crantz), MEcu-72 (M. esculenta Crantz), and MPer 417-003(M. esculenta peruviana). KU50 is a commercial cultivar developed in Thailand,which is extensively grown in Southeast Asia. MEcu-72 is one of the naturallyoccurringwild landraces inEcuador reported to be resistant towhiteflies byCIAT [18].MPer 417-003 is a wild species from Brazil, which is probably involved in theevolution of cassava cultivar, M. esculenta Crantz. The MPer 417-003 has strongresistance to insects such as mealybugs, mites, and whiteflies [19]. One of theobjectives to choose these varieties is to understand common mechanisms ofdrought tolerance in cassava and use this knowledge for the improvement of othercropsmainly because cassava has some features that allow it to copewith stress betterthan other crops, for example, high stomatal sensitivity to environmental humid-ity [20] and quick recovery after stress [21]. Another objective is to identify novel traitsand superior genes using genetic diversity existing in cultivated and wild species ofcassava. So far, we have obtained 19 968, 29 952, and 19 968 end sequences of full-
41.4 How Cassava Genomics Tool will help Develop Drought-Tolerant Variety j1053

Table41.2
Cho
osingrigh
ttraitforrigh
tdrou
ghten
vironm
entwhilescreen
ingthegerm
plasm.A
sterisks
indicate
thelevelo
ftherelevancebetweentraitan
ddrou
ghttolerance(�¼low
,�� ¼
middlean
d��
� ¼high
)..
Major
trait
Associate
traits
Traitrelevanceto
drou
ghtat
diffe
rent
grow
thstages
Early
drou
ght
Mid
season
Late
drou
ght
Water
uptake
Deeproot
���
���
Tempo
ralroot
architecture
���
���
Coo
lcanop
y��
��
Water
use
efficien
cyPartial
closure
ofstom
ata
���
���
Highph
otosyn
thesis
���
���
Optim
umleaf
area
indexafterrecovery
���
���
�
Earlygrow
thvigo
r�
���
���
Betterpartitioningto
roots
Earlybu
lking
��
���
Stem
swithmorestoragecapacity
fornon
structural
carboh
ydrates
���
���
Efficien
ttran
slocationof
non
structuralcarboh
ydrates
toroots
����
�
1054j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

length cDNAclones fromKU50,MEcu-72, andMPer 417-003, respectively and someof the sequences are not present on Phytozome Web site (www.phytozome.net/cassava), the cassava genome database. Using a set of unique genes from theexpressed sequence tag (EST) collections, an oligoarray analysis platform is beingdeveloped with more than 30 000 genes. These genomic and transcriptomic toolsenable us to dissect traits of importance for cassava improvement in a comprehensiveway. We believe that the platform could be applied for any cassava variety includingwild cassava species and will be useful to cassava research community globally.
In 2009, the first annotated draft cassava genome sequence became available at thePhytozomeWeb site (www.phytozome.net/cassava). The genome sequencing projecthas been initiated through theDOE-JGICommunity SequencingProgram (CSP) by a14-member consortium including DanForth Plant Center and CIAT. Furthermore,theUniversity ofArizonahas started a newproject funded by theBill &MelindaGatesFoundation in collaboration with DOE-JGI, 454 Life Sciences, and the University ofMaryland, Baltimore, to develop a genome variation database that will providebreeding tools such asmarkers. This variation databasewith comprehensive genomeinformation from different cassava genotypes will help make breeding schemesmore efficient throughmarker-assisted selection (MAS) (http://www.bio5.org/news/news_release/13-million-grant-ua-bill-melinda-gates-foundation-will-fund-next-phase-cassava-res). The full-length cassava cDNA sequence information derivedfrom different cassava varieties will significantly contribute to cassava genomesequence annotation and help know the source of genetic variation in differentcassava species. The high quality and correct annotation of the genome sequence canfurther accelerate identification of useful genes and promoters associated with traitscontributing to drought tolerance in cassava throughQTL, transcriptomic analysis (e.g., DNAmicroarrays), and transgenic approaches. In Arabidopsis and rice, molecularresponses to drought stress have been intensively analyzed and several key genesinvolved in the drought stress response and tolerance have been identified [22]. Asmost of the knowledge that has been obtained so far in Arabidopsis and rice can beapplied to other plants [23, 24], cassava homologues of the drought stress-relatedgenes in Arabidopsis and rice can allow us to look at genetic difference in terms ofgene and gene duplication betweenArabidopsis, rice, and cassava in terms of droughtstress response and its tolerance among the plant species.
41.5Gene Discovery for Drought Tolerance in Cassava
QTLmapping becomes a standard procedure in quantitative genetics [25] and recenttechnical progress in the area of molecular biology and genomics have allowed us toidentifyQTL gene. There are twomajor strategies for theQTL cloning: one is throughpositional cloning and another is through association mapping. Because of a longgrowth cycle and complex genetic nature of cassava, gene discovery through QTLcloning was not an easy task in the past and this will drastically change once all thegenomic sequences and genomics tools become available through current global
41.5 Gene Discovery for Drought Tolerance in Cassava j1055

efforts as described above. The tools will help develop fine geneticmapping, physicalmapping, or markers, which are strongly associated with trait of interest for cassavaimprovement [26–28]. In addition, plant comparative genomics and transcriptomicsapproach will largely benefit from research advances described above to discover keygenes associated with traits of interest [29–32]. Especially, sequence homology orsimilarity can serve as a strong evidence for detecting functional elements in genomicsequences. Here, we give an example to identify cassava genes, which may havemolecular and physiological functions similar to those of genes in other plants.
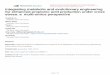
It was recently reported that transgenic cassava expressing IPT gene, whichencodes isopentenyl transferase from Agrobacterium tumefaciens, under the controlof the Arabidopsis promoter of the senescence-induced SAG12 gene, could trigger aseries of metabolic changes such as cytokinin metabolism that increased droughtresistance [33]. This study not only could prove the feasibility of delayed leafsenescence of woody cassava but also could reveal mechanisms underlying thecontrol of cytokinin homeostasis in cassava leaves. However, the origin of the IPTgene used in this study was A. tumefaciens, a bacterium. Therefore, we carried outhomologue search by BLAST program and phylogenetic analysis by NJ method toidentify orthologue genes of IPT in cassava, and nine sequences were identified fromcassava genome database as shown in Figure 41.1. Generally, IPTgenes are classifiedinto bacterial type or plant type according to their amino acid sequences [34].Interestingly, plant-type IPT genes in Arabidopsis are strongly involved in thecytokinin metabolism in contrast to bacterial-type IPT genes [34–36]. As shown inFigures 41.1 and 41.2, all the nine genes identified as cassava IPTgenes belong to theplant-type IPT group. Furthermore, some cassava EST sequences, which partiallyencode the plant-type IPTgenes, are now available.We are trying to isolate full-lengthcassava cDNAs encoding plant-type IPT genes for developing drought-tolerantcassava cultivars using transgenic approach.
It is recognized that traditional breeding ismost unlikely to provide all solutions forimproving the crop to meet the varying needs of small farmers and commercialproduction in the tropics [39, 40]. The transgenic technologies can accelerateintegration of desired traits into farmer preferred cultivars and landraces, as wellas elite breeding lines. Hence, this technique is one of the options to get maximumbenefit frommajor advances occurring in the genomics for improvement of cassava,which is a long-duration crop. Transgenic technologies allow beneficial traits to betransferred from one cassava cultivar to another and from wild relatives to cultivatedManihot, circumventing species boundaries and the problems of outcrossing andinbreeding depression, inherent to this vegetatively propagated crop.
The first series of cassava transgenic technologies were reported in 1996 [41–43].Since then, the transgenic technologies have been improved [44, 45] and utilized toproduce genetically transformed cassava expressing traits such as reduced cyano-genic content [46–48], enhanced starch production [49, 50], insect resistance [51],virus resistance [52–56], and herbicide resistance [57, 58]. Recently, it has beenreported that transgenic cassava expressing IPT gene from Agrobacterium increaseddrought resistance as mentioned in the previous section [33]. The transgenic plantswere useful both to investigate drought resistance in cassava and to increase storage
1056j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

root production in semiarid regions and leaf biomass as vegetable and animal feed.Furthermore, Saelim et al. [59] have established amarker-free cassava transformationprotocol using a cassava cultivar, KU50, which is extensively grown in Southeast Asia.This is the first demonstration of the efficacy of the system in promoting excision ofiptmarker gene in cassava species, with the consequent rapid production of marker-free transgenic plants [59]. The high efficiency of this system should facilitatepyramiding a number of transgenes by repeated transformation without having toundergo laborious, expensive, and time-consuming processes of sexual crossing andseed production. The generation of marker-free, thus environmentally safe, genet-ically modified cassava clones should also ease the public concerns regarding the useof transgenic cassava in both food and nonfood industries. Technologies fortransformation of plants are in high demand and are being increasingly employedby large seed and agrochemical sectors. Most of the academic research groups lack
Figure 41.1 Phylogenetic tree of IPT genes.The amino acid sequences of the IPT genes (9identified IPThomologous genes inM. esculenta(cassava), 19 IPT homologous genes in G. max(Gm) and 9 genes in A. thaliana (At) [34], 9genes in O. sativa (Os) [37], and 7 genes in Z.maiz (Zm) [38] were aligned by ClustalX2 and
the phylogenetic tree was constructed using theNJmethod. IPT gene in A. tumefaciens is a queryfor the BLAST search. Asterisks show the planttype putative IPT genes identified from cassavagenome database (phytozome). IPT genes areclassified into bacteria type or plant typeaccording to their amino acid sequences [34].
41.5 Gene Discovery for Drought Tolerance in Cassava j1057

infrastructure, logistics and human resources necessary to establish middle to high-throughput transformation pipeline for their own target crop. CIAT has beenworking in transgenic research on cassava and rice for more than a decade andhas scaled up its facilities to include confined fields for field evaluation, and it hasbeen uniquely placed to evaluate the gene technology with a generic permission fromNational Biosafety Committee of Colombia. The CIAT has now established theTransformation Platform to host all the in-house or collaborative research projectsthat aim to evaluate transgenic technology for important crops such as rice andcassava. The strategy is to accelerate evaluation of gene technology through trans-formation and phenotyping platforms by employing scaled up infrastructure,expertise, and knowledgewith the ultimate objective of complementing conventionalcrop improvement technology for cassava.
41.6Proteomics and Metabolomics Approach in Cassava
Several groups have shown the profiles of protein pattern of a storage root [60], fibrousand tuberous root [61], plant–pathogen interactions [62], and secondary somatic embryo-genesis [63] in cassava by using a technology of two-dimensional gel electrophoresis.Recently, Li et al. [64] provided a new insight into the proteomic patterns in the somaticembryos, plantlets, and tuberous roots [64]. These investigations have paved theway for acomprehensive, system-wide analysis of cassava. Large-scale proteome data sets are animportant resource for a better understanding of protein functions in terms of not onlygrowth and development but also response to environmental changes.
Metabolomics aims to understand metabolic systems based on comprehensiveand integrated approaches that take advantage of recent advances in instrumentationto characterizemetabolites.Metabolomic approaches can allow us to conduct parallelassessments of multiple metabolites and can facilitate quantitative analysis ofparticular metabolites with major advantages over chemical-level phenotyping anddiagnostic analysis. The plant metabolome represents an enormous chemicaldiversity due to the complex set of metabolites produced in each plant species[65, 66]. Various sources of information onmetabolomics have played a crucial role inmetabolome research and its synergistic integration with other omics approaches.Metabolomics information for cassava has not been launched yet, but for otherplants, there are Web sites on metabolome resources such as TAIR (http://www.arabidopsis.org/portals/metabolome/index.jsp), the KEGG PATHWAY Database(http://www.genome.jp/kegg/pathway.html) [67–69], the Plant Metabolic Network(PMN) (http://www.plantcyc.org/), PlantCyc including AraCyc and PoplarCyc(http://www.plantcyc.org:1555/PLANT/server.html) [70], KaPPA-View (http://kpv.kazusa.or.jp/kappa-view/) [71, 72], and PRIMe (http://prime.psc.riken.jp/) [73].However, the analysis of the ESTs for full-length cDNAs from abiotic and bioticstress-treated cassava tissues mentioned in the previous section has revealed thatsome of EST sequences encode novel plant genes and they do not belong to anypathways that are already known in plants (unpublished data). It indicates a
1058j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

possibility of the existence of some novel pathways related to stress tolerance incassava. Therefore, cassava metabolomics, which is a great analytical challenge, canserve as an immensely important tool to design molecular breeding strategies forimprovement of the productivity and value of cassava in the near future.
41.7Future Perspectives
Earlier investigationshaveoften indicated the relationshipof someof thephysiologicaltraits with root yield of cassava underwater deficit environment. The concepts evolvedso far need to be validated and transformed into crop improvement tools to ensureprecise phenotyping for trait discovery and gene identification. Phenomics platformwith tools and techniques that are emerging out of advances in digital imaging andimage analysis can significantly help in phenotyping long duration plants such ascassava with reduced cost. These noninvasive rapid methods can greatly complement
Genetic diversity in cassavaParental lines/breeding population
Omics tools• Phenomics
PhenotypingGenotypingOmics tools• Genomics
• Proteomics• MetabolomicsMarker development
• High-density mapTrait discovery• Novel trait
QTL
•Transcriptomics
• QTL
Trait evaluation and validation
Gene discoverySynteny
• Phenotypic marker
• Gene marker
Other crops• Candidate genes
Elite lines
Transgenics• GM cassava
Figure 41.2 Gene discovery for a particulartrait is the key to accelerating geneticimprovement of cassava; however, genotypingand phenotyping tools are the majorprerequisites that can determine the success.Recent advances in genomics can ensure high-density genetic maps, while precisionphenotyping is essential for identifying the keytraits associated with desired phenotype ofplants. This will also determine the precision ofassociation between the trait of interest and theQTL. Omics tools such as genomics and
transcriptomics can facilitate both high-qualitygenotyping and discovery of candidate genes byassessing synteny with other crops. Phenomics,proteomics, and metabolomics can improveefficiency of trait discovery. Thus, the genediscovery will be the product of integrationbetween efficient genotyping and phenotyping.It is necessary to further carry out traitevaluation and identify precise phenotypicmarker to carry forward the gene discovery intouseful genemarker and to supportmarker-aidedselection in the cassava breeding program.
41.7 Future Perspectives j1059

research efforts of both the genomic analysis platform and the transformationplatform to enhance our understanding about molecular mechanisms of droughttolerance in cassava. However, these technologies and knowledge that can save timeand money in developing elite cultivars have not been effectively integrated intocassava breeding programs. This can be partially because of heterozygosity and longgrowth cycle, which are the major constraints in cassava breeding programs. Fur-thermore, as shown in Figure 41.2, the key lies in integration of genomics, tran-scriptomics, metabolomics, proteomics, and other large-scale �omics� data withsystems biology approaches, and it can open new avenues for engineering cassavato enhance yields, improve nutritional value, and overcome the problems of posthar-vest physiological deterioration as well as biotic and abiotic stresses.
Through integrated omics approach, gene markers can be identified rapidly andat low cost in the near future. Furthermore, MAS that focuses on key traitscontributing to drought tolerance can become more robust for accelerating geneticimprovement of cassava. Since drought tolerance is a highly complex trait, pyr-amiding multiple sources of genes for drought tolerance into a set of newprogenitors can lead to a much desirable impact. CIAT will increasingly adopttechnologies evolving from recent developments in single-nucleotide polymorph-isms (SNPs) to accelerate the application of MAS strategies in cassava breedingprograms. Marker genotyping based on SNP resources are being developed byusing next-generation sequencers (NGS) such as 454 DNA sequencer and Illuminagenome analyzer. The computational discovery of EST-based SNPs and/or EST-SNPmarkers is now in progress for numerous species [74–82]. EST data sets for full-length cDNA, which we have obtained (see Section 41.5), will serve as an importantresource for discovery of SNPs, especially for locating expressed genes onto agenetic map. This will lead topositional cloning of genes of interest that can beuseful as efficient markers for accelerating improvement of cassava for toleranceagainst abiotic and biotic stresses.
Acknowledgments
We thank Dr. Sankaran Murugan for discovering genes related to droughttolerance. This work was supported in part by Strategic Funds for the Promotionof Science and Technology of theMinistry of Education, Culture, Sports, Science andTechnology (MEXT), Japan.
References
1 FAO. FAOSTAT Statistical Database,Agriculture Data.
2 Vries, C.A., Ferweds, J.D., and Flash, M.(1976) Choice of crops in relation to actualand potential production in tropics. Neth.J. Agric. Sci., 15, 241–246.
3 de Oliveira, S.L., Macedo, M.M.C., andPorto, M.C.M. (1982) Effect of waterdeficit on cassava root yield. Pesqui.Agropecu. Bras., 17, 121–124.
4 Connor, D.J. and Cock, J.H. (1981)Response of cassava to water shortage. II.
1060j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

Canopy dynamics. Field Crops Res., 4,285–296.
5 Porto, M.C.M. (1983) Physiologicalmechanisms of drought tolerance incassava (Manihot esculenta Crantz). PhDdissertation. The University of Arizona,Tucson.
6 El-Sharkawy, M.A. (1993) Drought-tolerant cassava for Africa, Asia, andLatin America: breeding projects work tostabilize productivity without increasingpressures on limited natural resources.BioScience, 43, 441–451.
7 Lenis, J.I., Calle, F., Jaramillo, G.,Perez, J.C., Ceballo, S.H., and Cock, J.H.(2006) Leaf retention and cassavaproductivity. Field Crops Res., 95, 126–134.
8 El-Sharkawy, M.A. (2007) Physiologicalcharacteristics of cassava tolerance toprolonged drought in the tropics:implications for breeding cultivarsadapted to seasonally dry and semiaridenvironments. Braz. J. Plant Physiol., 19,257–286.
9 Fukushima, A., Kusano, M., Redestig, H.,Arita, M., and Saito, K. (2009) Integratedomics approaches in plant systemsbiology. Curr. Opin. Chem. Biol., 13 (5–6),532–538.
10 Keurentjes, J.J.B., Koornneef, M., andVreugdenhil, D. (2008) Quantitativegenetics in the age of omics. Curr. Opin.Plant Biol., 11 (2), 123–128.
11 Mochida, K., Furuta, T., Ebana, K.,Shinozaki, K., and Kikuchi, J. (2009)Correlation exploration of metabolic andgenomic diversity in rice. BMC Genomics,10, 568.
12 Urano, K., Kurihara, Y., Seki, M., andShinozaki, K. (2010) �Omics� analyses ofregulatory networks in plant abiotic stressresponses. Curr. Opin. Plant Biol., 13 (2),132–138.
13 El-Sharkawy, M.A. (2005) How cancalibrated research-based models beimproved for use as a tool inidentifying genes controlling croptolerance to environmental stresses in theera of genomics – from anexperimentalist�s perspective.Photosytnthetica, 43, 161–176.
14 IPCC – Intergovernmental Panel onClimate Change (2006) IPCC Special
Report on theRegional Impacts ofClimatechange – An Assessment of Vulnerability.UNEP, WMO (http://www.grida.no/climate/ipcc/regional/502.htm).
15 EL-Sharkawy, M.A., De Tafur, S.M., andCadavid, L.F. (1992) Potentialphotosynthesis of cassava as affectedby growth conditions. Crop Sci., 32 (6),1336–1342.
16 Passioura, J.B. (1977) Grain yield, harvestindex, and water use of wheat. J. Aust. Inst.Agri. Sci., 43, 117–120.
17 Sakurai, T., Plata, G., Rodr�ıguez-Zapata,F., Seki, M., Salcedo, A., Toyoda, A.,Ishiwata, A., Tohme, J., Sakaki, Y.,Shinozaki, K., and Ishitani, M. (2007)Sequencing analysis of 20,000 full-lengthcDNA clones from cassava reveals lineagespecific expansions in gene familiesrelated to stress response. BMC PlantBiol., 7, 66.
18 Bellotti, A. and Arias, B. (2001) Host plantresistance to whiteflies with emphasis oncassava as a case study. Crop Prot., 20,813–823.
19 Burbano, M.M., Carabali, M.A.,Montoya, L.J., and Belloti, A. (2007)Resistance of Manihot species toMononychellus tanajoa (Acariformes),Aleurotrachelus socialis, and Phenacoccusherreni (Hemiptera). Rev. Colomb.Entomol., 33, 110–115.
20 Oguntunde, P.G. (2005) Whole-plantwater use and canopy conductance ofcassava under limited available soil waterand varying evaporative demand. PlantSoil, 278, 371–383.
21 El-Sharkawy, M.A. and Cock, J.H. (1987)Response of cassava to water stress. PlantSoil, 100, 345–360.
22 Umezawa, T., Fujita, M., Fujita, Y.,Yamaguchi-Shinozaki, K., andShinozaki, K. (2006) Engineering droughttolerance in plants: discovering andtailoring genes to unlock the future. Curr.Opin. Biotechnol., 17, 113–122.
23 Xiong, L., Schumaker, K.S., and Zhu, J.-K.(2002)Cell signaling during cold, drought,and salt stress. Plant Cell, 14 (Suppl.),S165–S183
24 Yamaguchi-Shinozaki, K. and Shinozaki,K. (2006) Transcriptional regulatorynetworks in cellular responses and
References j1061

tolerance to dehydration and cold stresses.Annu. Rev. Plant Biol., 57, 781–803.
25 Lynch, M. and Walsh, B. (eds) (1998)Genetics and Analysis of Quantitative Traits,Sinuaer Associates.
26 Salvi, S. and Tuberosa, R. (2005) To cloneor not to clone plant QTLs: present andfuture challenges. Trends Plant Sci., 10,297–304.
27 Martienssen, R.A., Rabinowicz, P.D.,O�Shaughnessy, A., and McCombie, W.R.(2004) Sequencing the maize genome.Curr. Opin. Plant Biol., 7, 102–107.
28 Van der Hoeven, R., Ronning, C.,Giovannoni, J., Martin, G., and Tanksley,S. (2002) Deductions about the number,organization, and evolution of genes in thetomato genome based on analysis of alarge expressed sequence tag collectionand selective genomic sequencing. PlantCell, 14, 1441–1456.
29 Mochida, K. and Shinozaki, K. (2010)Genomics and bioinformatics resourcesfor crop improvement. Plant Cell Physiol.,51, 497–523.
30 Sato, S. and Tabata, S. (2006) Lotusjaponicus as a platform for legumeresearch. Curr. Opin. Plant Biol., 9,128–132.
31 Itoh, T., Tanaka, T., Barrero, R.A.,Yamasaki, C., Fujii, Y., Hilton, P.B.,Antonio, B.A., Aono, H., Apweiler, R.,Bruskiewich, R., Bureau, T., Burr, F.,Costa de Oliveira, A., Fuks, G., Habara, T.,Haberer, G., Han, B., Harada, E., Hiraki,A.T., Hirochika, H., Hoen, D., Hokari, H.,Hosokawa, S., Hsing, Y.-I., Ikawa, H.,Ikeo, K., Imanishi, T., Ito, Y., Jaiswal, P.,Kanno, M., Kawahara, Y., Kawamura, T.,Kawashima, H., Khurana, J.P., Kikuchi,S., Komatsu, S., Koyanagi, K.O., Kubooka,H., Lieberherr, D., Lin, Y.-C., Lonsdale, D.,Matsumoto, T., Matsuya, A., McCombie,W.R., Messing, J., Miyao, A., Mulder, N.,Nagamura, Y., Nam, J., Namiki, N.,Numa, H., Nurimoto, S., O�Donovan, C.,Ohyanagi, H., Okido, T., Oota, S.,Osato, N., Palmer, L.E., Quetier, F.,Raghuvanshi, S., Saichi, N., Sakai, H.,Sakai, Y., Sakata, K., Sakurai, T., Sato, F.,Sato, Y., Schoof, H., Seki, M., Shibata, M.,Shimizu, Y., Shinozaki, K., Shinso, Y.,Singh,N.K., Smith-White, B., Takeda, J.-I.,
Tanino, M., Tatusova, T., Thongjuea, S.,Todokoro, F., Tsugane, M., Tyagi, A.K.,Vanavichit, A., Wang, A., Wing, R.A.,Yamaguchi, K., Yamamoto, M.,Yamamoto, N., Yu, Y., Zhang, H., Zhao,Q., Higo, K., Burr, B., Gojobori, T., andSasaki, T. (2007) Curated genomeannotation of Oryza sativa ssp. japonicaand comparative genome analysis withArabidopsis thaliana. Genome Res., 17,175–183.
32 Neale, D.B. and Ingvarsson, P.K. (2008)Population, quantitative and comparativegenomics of adaptation in foresttrees. Curr. Opin. Plant Biol., 11,149–155.
33 Zhang, P., Wang, W.-Q., Zhang, G.-L.,Kaminek, M., Dobrev, P., Xu, J., andGruissem, W. (2010) Senescence-inducible expression of isopentenyltransferase extends leaf life, increasesdrought stress resistance and alterscytokinin metabolism in cassava. J. Integr.Plant Biol., 52, 653–669.
34 Takei, K., Sakakibara,H., andSugiyama,T.(2001) Identification of genes encodingadenylate isopentenyltransferase, acytokinin biosynthesis enzyme, inArabidopsis thaliana. J. Biol. Chem., 276,26405–26410.
35 Sun, J., Niu, Q.-W., Tarkowski, P., Zheng,B., Tarkowska, D., Sandberg, G., Chua,N.-H., and Zuo, J. (2003) The ArabidopsisAtIPT8/PGA22 gene encodes anisopentenyl transferase that is involved inde novo cytokinin biosynthesis. PlantPhysiol., 131, 167–176.
36 Miyawaki, K., Tarkowski, P.,Matsumoto-Kitano, M., Kato, T., Sato, S.,Tarkowska, D., Tabata, S., Sandberg, G.,and Kakimoto, T. (2006) Roles ofArabidopsis ATP/ADPisopentenyltransferases and tRNAisopentenyltransferases in cytokininbiosynthesis. Proc. Natl. Acad. Sci. USA,103, 16598–16603.
37 Sakamoto, T., Sakakibara, H., Kojima, M.,Yamamoto, Y., Nagasaki, H., Inukai, Y.,Sato, Y., and Matsuoka, M. (2006)Ectopic expression of KNOTTED1-likehomeobox protein induces expression ofcytokinin biosynthesis genes in rice. PlantPhysiol., 142, 54–62.
1062j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

38 Brugi�ere, N., Humbert, S., Rizzo, N.,Bohn, J., and Habben, J.E. (2008) Amember of the maize isopentenyltransferase gene family, Zea maysisopentenyl transferase 2 (ZmIPT2),encodes a cytokinin biosyntheticenzyme expressed during kerneldevelopment. Cytokininbiosynthesis inmaize. PlantMol. Biol., 67,215–229.
39 Jennings, D.L. and Iglesias, C. (2002)Breeding for crop improvement, inCassava:Biology, Production andUtilization(eds R.J. Hillocks and J.M. Thresh), CABI,Oxon, pp. 149–166.
40 Kawano, K. (2003) Thirty years of cassavabreeding for productivity: biological andsocial factors for success. Crop Sci., 43,1325–1335.
41 Li, H.Q., Sautter, C., Potrykus, I., andPuonti-Kaerlas, J. (1996) Genetictransformation of cassava (Manihotesculenta Crantz). Nat. Biotechnol., 14,736–740.
42 Taylor, N.J., Edwards, M., Kiernan, R.J.,Davey, C.D., Blakesley, D., andHenshaw, G.G. (1996) Development offriable embryogenic callus andembryogenic suspension culturesystems in cassava (Manihot esculentaCrantz). Nat. Biotechnol., 14,726–730.
43 Vasil, I.K. (1996) Milestones in cropbiotechnology: transgenic cassava andAgrobacterium-mediated transformationof maize. Nat. Biotechnol., 14, 702–703.
44 de Vetten, N., Wolters, A.-M., Raemakers,K., van der Meer, I., ter Stege, R., Heeres,E., Heeres, P., and Visser, R. (2003) Atransformation method for obtainingmarker-free plants of a cross-pollinatingand vegetatively propagated crop. Nat.Biotechnol., 21, 439–442.
45 Bull, S.E., Owiti, J.A., Niklaus, M.,Beeching, J.R., Gruissem, W., andVanderschuren, H. (2009) Agrobacterium-mediated transformation of friableembryogenic calli and regeneration oftransgenic cassava. Nat. Protoc., 4,1845–1854.
46 Andersen, M.D., Busk, P.K., Svendsen, I.,and Møller, B.L. (2000) Cytochromes P-450 from cassava (Manihot esculenta
Crantz) catalyzing the first steps in thebiosynthesis of the cyanogenic glucosideslinamarin and lotaustralin. Cloning,functional expression in Pichia pastoris,and substrate specificity of the isolated.J. Biol. Chem., 275, 1966–1975.
47 Siritunga, D. and Sayre, R. (2007)Transgenic approaches for cyanogenreduction in cassava. J. AOAC Int., 90,1450–1455.
48 Siritunga, D. and Sayre, R. (2004)Engineering cyanogen synthesis andturnover in cassava (Manihot esculenta).Plant Mol. Biol., 56, 661–669.
49 Raemakers, C.J.J.M., Schreuder, M.M.,Pereira, I., Suurs, L., Vincken, J.P.,Jacobsen, E., and Visser, R.G.F. (2003)Production of amylose-free cassava plantsby genetic modification, in Cassava: AnAncient Crop for Modern Times Proceedingsof the 5th International Meeting of theCassava Biotechnology Network, St LouisMO, 2001 (eds C.M. Fauquet and N.J.Nigel).
50 Ihemere, U., Arias-Garzon, D.,Lawrence, S., and Sayre, R. (2006) Geneticmodification of cassava for enhancedstarch production. Plant Biotechnol. J., 4,453–465.
51 Ladino, J.J., Echeverry, M., Mancilla, L.I.,Lopez, D., Chavarriaga, P., Tohme, J., andRoca,W. (2002) Genetic transformation ofcassava: confirmation of transgenesis inclone 60444 and analysis of CRY1Abprotein in transgenic lines. Preliminarydata on transformation of farmer-preferred cultivars SM1219-9 andCM3306-4. Annual Report, CIAT, Cali,Colombia.
52 Pita, J.S., Fondong, V.N., Sangar�e, A.,Otim-Nape, G.W., Ogwal, S., and Fauquet,C.M. (2001) Recombination,pseudorecombination and synergism ofgeminiviruses are determinant keys to theepidemic of severe cassavamosaic diseasein Uganda. J. Gen. Virol., 82,655–665.
53 Chellappan, P., Masona, M.V.,Vanitharani, R., Taylor, N.J., andFauquet, C.M. (2004) Broad spectrumresistance to ssDNA viruses associatedwith transgene-induced gene silencing incassava. Plant Mol. Biol., 56, 601–611.
References j1063

54 Zhang, P., Vanderschuren,H., F€utterer, J.,and Gruissem, W. (2005) Resistance tocassava mosaic disease in transgeniccassava expressing antisense RNAstargeting virus replication genes. PlantBiotechnol. J., 3, 385–397.
55 Vanderschuren, H., Akbergenov, R.,Pooggin, M.M., Hohn, T., Gruissem, W.,and Zhang, P. (2007) Transgenic cassavaresistance to African cassava mosaic virusis enhanced by viral DNA-A bidirectionalpromoter-derived siRNAs. Plant Mol.Biol., 64, 549–557.
56 Vanderschuren, H., Alder, A., Zhang, P.,and Gruissem,W. (2009) Dose-dependentRNAi-mediated geminivirus resistance inthe tropical root crop cassava. Plant Mol.Biol., 70, 265–272.
57 Sarria, R., Torres, E., Angel, F.,Chavarriaga, P., and Roca, W.M. (2000)Transgenic plants of cassava (Manihotesculenta) with resistance toBasta obtainedby Agrobacterium-mediatedtransformation. Plant Cell Rep., 19,339–344.
58 Echeverry, M., Mancilla, L.I., Cortes, D.F.,Chavarriaga, P., and Tohme, J. (2002)Preliminary evaluation of the expressionof the transgene bar in cassava plantsmaintained under asexual propagation for10 years. Annual Report, CIAT, Cali,Colombia.
59 Saelim, L., Phansiri, S.,Suksangpanomrung, M., Netrphan, S.,andNarangajavana, J. (2009) Evaluation ofa morphological marker selection andexcision system to generate marker-freetransgenic cassava plants. Plant Cell Rep.,28, 445–455.
60 Cabral, G.B. and Carvalho, L.J.C.B. (2001)Analysis of proteins associated withstorage root formation in cassava usingtwo-dimensional gel electrophoresis. Rev.Bras. Fisiol. Veg., 13, 41–48.
61 Sheffield, J., Taylor, N., Fauquet, C., andChen, S. (2006) The cassava (Manihotesculenta Crantz) root proteome: proteinidentification and differential expression.Proteomics, 6, 1588–1598.
62 Mehta, A., Magalhaes, B.S., Souza, D.S.L.,Vasconcelos, E.A.R., Silva, L.P.,Grossi-de-Sa, M.F., Franco, O.L.,da Costa, P.H.A., and Rocha, T.L. (2008)
Rooteomics: the challenge of discoveringplant defense-related proteins in roots.Curr. Protein Pept. Sci., 9, 108–116.
63 Baba, A., Nogueira, F., Pinheiro, C.,Brasil, J., Jereissati, E., Juca, T., Soares, A.,Santos, M., Domont, G., and Campos, F.(2008) Proteome analysis of secondarysomatic embryogenesis in cassava(Manihot esculenta). Plant Sci., 175,717–723.
64 Li, K., Zhu, W., Zeng, K., Zhang, Z., Ye, J.,Ou, W., Rehman, S., Heuer, B., andChen, S. (2010) Proteomecharacterizationof cassava (Manihot esculenta Crantz)somatic embryos, plantlets and tuberousroots. Proteome Sci., 8, 10.
65 Bino, R.J., Hall, R.D., Fiehn, O., Kopka, J.,Saito, K., Draper, J., Nikolau, B.J.,Mendes,P., Roessner-Tunali, U., Beale, M.H.,Trethewey, R.N., Lange, B.M., Wurtele,E.S., and Sumner, L.W. (2004) Potential ofmetabolomics as a functional genomicstool. Trends Plant Sci., 9, 418–425.
66 Roepenack-Lahaye, E., von Degenkolb, T.,Zerjeski, M., Franz, M., Roth, U.,Wessjohann, L., Schmidt, J., Scheel, D.,and Clemens, S. (2004) Profiling ofArabidopsis secondary metabolites bycapillary liquid chromatography coupledto electrospray ionization quadrupoletime-of-flight mass spectrometry. PlantPhysiol., 134, 548–559.
67 Aoki-Kinoshita, K.F. and Kanehisa, M.(2007) Gene annotation and pathwaymapping in KEGG. Methods Mol. Biol.,396, 71–91.
68 Kanehisa, M. (2002) The KEGG database.Novartis Found. Symp., 247, 91–101,discussion 101–103, 119–128, 244–252.
69 Kanehisa, M., Araki, M., Goto, S.,Hattori, M., Hirakawa, M., Itoh, M.,Katayama, T., Kawashima, S., Okuda, S.,Tokimatsu, T., and Yamanishi, Y. (2008)KEGG for linking genomes to life and theenvironment. Nucleic Acids Res., 36,D480–D484
70 Mueller, L.A., Zhang, P., and Rhee, S.Y.(2003) AraCyc: a biochemical pathwaydatabase for Arabidopsis. Plant Physiol.,132, 453–460.
71 Tokimatsu, T., Sakurai, N., Suzuki, H.,Ohta, H., Nishitani, K., Koyama, T.,Umezawa, T., Misawa, N., Saito, K., and
1064j 41 Cassava Genetic Improvement: Omics Approaches for Facing Global Challenges

Shibata, D. (2005) KaPPA-view: a web-based analysis tool for integration oftranscript and metabolite data on plantmetabolic pathway maps. Plant Physiol.,138, 1289–1300.
72 Sakurai, N., Ara, T., Ogata, Y., Sano, R.,Ohno, T., Sugiyama, K., Hiruta, A.,Yamazaki, K., Yano, K., Aoki, K.,Aharoni, A., Hamada, K., Yokoyama, K.,Kawamura, S., Otsuka, H., Tokimatsu, T.,Kanehisa, M., Suzuki, H., Saito, K., andShibata, D. (2010) KaPPA-View4: ametabolic pathway database forrepresentation and analysis of correlationnetworks of gene co-expression andmetabolite co-accumulation and omicsdata. Nucleic Acids Res., 39, D677–D684
73 Akiyama, K., Chikayama, E., Yuasa, H.,Shimada, Y., Tohge, T., Shinozaki, K.,Hirai, M.Y., Sakurai, T., Kikuchi, J., andSaito, K. (2008) PRIMe: a Web site thatassembles tools for metabolomics andtranscriptomics. In Silico Biol., 8, 339–345.
74 Ching, A. and Rafalski, A. (2002) Rapidgenetic mapping of ESTs using SNPpyrosequencing and indel analysis. Cell.Mol. Biol. Lett., 7, 803–810.
75 Duran, C., Appleby, N., Clark, T., Wood,D., Imelfort, M., Batley, J., and Edwards,D. (2009) AutoSNPdb: an annotated singlenucleotide polymorphism database forcrop plants. Nucleic Acids Res., 37,D951–D953
76 Kota, R., Varshney, R.K., Prasad, M.,Zhang,H., Stein,N., andGraner, A. (2008)EST-derived single nucleotidepolymorphism markers for assemblinggenetic and physical maps of the barley
genome. Funct. Integr. Genomics, 8,223–233.
77 Kota, R., Varshney, R.K., Thiel, T.,Dehmer, K.J., and Graner, A. (2001)Generation and comparison of EST-derived SSRs and SNPs in barley(Hordeum vulgare L.). Hereditas, 135,145–151.
78 Lopez, C., Pi�egu, B., Cooke, R.,Delseny, M., Tohme, J., and Verdier, V.(2005) Using cDNA and genomicsequences as tools to develop SNPstrategies in cassava (Manihot esculentaCrantz). TAG Theor. Appl. Genet., 110,425–431.
79 Rafalski, A. (2002) Applications of singlenucleotide polymorphisms in cropgenetics. Curr. Opin. Plant Biol.,5, 94–100.
80 Schmid, K.J., Sorensen, T.R., Stracke, R.,Torjek, O., Altmann, T., Mitchell-Olds, T.,and Weisshaar, B. (2003) Large-scaleidentification and analysis of genome-wide single-nucleotide polymorphismsfor mapping in Arabidopsis thaliana.Genome Res., 13, 1250–1257.
81 Tang, J., Vosman, B., Voorrips, R.E.,Linden, C.G., and van der Leunissen,J.A.M. (2006) QualitySNP: a pipeline fordetecting single nucleotidepolymorphisms and insertions/deletionsin EST data from diploid and polyploidspecies. BMC Bioinformatics, 7 438.
82 Useche, F.J., Gao, G., Harafey, M., andRafalski, A. (2001) High-throughputidentification, database storage andanalysis of SNPs in EST sequences.Genome Inform., 12, 194–203.
References j1065