Embed Size (px)
Citation preview

ELSEVIER Advanced Drug Delivery Reviews 19 (1996) 445-467
advanced drugdeliiry reviews
Importance of receptor-mediated endocytosis in peptide delivery and targeting: kinetic aspects
Hitoshi Sate”‘” , Yuichi Sugiyamab, Akira Tsuji”, Isamu Horikoshi” A Department of Hospital Pharmacy, Toyama Medical and Pharmaceutical University, 2630 Sugitani. Toyama 9.30-01. Japan
hDepartment of Pharmaceutics, Faculty of Pharmaceutical Sciences, University of Tokyo, Tokyo, Japan ‘Department of Pharmaceutics, Faculty of Pharmaceutical Sciences. Kanazawa University. Kanazawa, Japan
Abstract
Receptor-mediated endocytosis (RME) is a general mechanism by which eukaryotic cells internalize peptide hormones, growth factors, cytokines, plasma glycoproteins, lysosomal enzymes, toxins and viruses concomitantly with their receptors on the cell surface. Receptor-mediated drug targeting is considered to show promise as a drug delivery system (DDS). The success of RME-utilizing drug targeting is dependent not only on the in vitro efficacy but on the in vivo behavior of the drug-polypeptide complex. In fact, there has not been much progress from the “natural” targeting of glycoproteins and LDL to hepatocytes or macrophage lineages, due to unfavorable effects on the pharmacokinetics and pharmacodynamics (PK/PD) of polypeptides. In this review, we considered the kinetic modelling of polypeptide RME in isolated or cultured cells in vitro, in perfused organs and in the whole body. Typical RME analyses are described to estimate the rate constants for polypeptide-receptor internalization, the degradation of internalized polypeptide and receptor externalization, based upon the “receptor recycling” model. The kinetic RME analysis enables not only the non-linear clearance and distribution in the liver to be predicted, but also the surface-bound and internalized polypeptide in the target cell, which is important for the intracellular efficacy of drug-polypeptide complex. In particular, the kinetics of down-regulation and subsequent recovery of surface receptors were precisely elucidated, because if the receptors are once down-regulated, the next dosage should be given only after the surface receptors are restored. The therapeutic efficacy/toxicity of a drug-carrier conjugate should be assessed from its microscopic pharmacology based on the RME mechanisms, in conjunction with macroscopic pharmacokinetic modelling.
Keywords: Peptide delivery and targeting; Receptor-mediated endocytosis (RME); Receptor recycling
Contents
1.
2. 3.
4. 5 . 6.
Introduction .......................................................................................................................................................................... 446
Factors influencing polypeptide pharmacokinetics/pharmacodynamics.. .............................................................................. 447
Kinetic analysis of RME in vitro and in perfused organs.. .................................................................................................... 449
3.1. Estimation of RME parameters from ligand movement.. .............................................................................................. 450
3.2. Estimation of RME parameters from receptor movement ............................................................................................ 452
Kinetic analysis of RME in vivo.. .......................................................................................................................................... 456
Implications of RME kinetics for clinical application of polypeptides .................................................................................. 459
Perspectives .......................................................................................................................................................................... 459
Acknowledgments ..................................................................................................................................................................... 460 References.. ............................................................................................................................................................................... 460
*Corresponding author. Fax: +81 764 345089; e-mail: [email protected]
0169-409X/96/$32.00 @ 1996 Elsevier Science B.V. All rights reserved
PII SO169-409X(96)00013-0

1. Introduction
Receptor-mediated endocytosis (RME) is a general mechanism by which animal cells inter- nalize a wide variety of selected extracellular materials concomitantly with their specific re- ceptors (for review, see Refs [l-S]). Such materi- als include peptide hormones, growth factors, cytokines, plasma glycoproteins, lysosomal en- zymes, toxins [9,10] and viruses [11,12]. First, a ligand binds to its specific receptor on the cell surface membrane: the ligand-bound receptors cluster in discrete regions called coated pits, which invaginate into the cell to form endo- cytotic vesicles or endosomes. The ligand and receptor then dissociate at an acidic pH (5-5.5) within the endosomes. The internalized receptor usually recycles back to the cell surface for further binding, while the internalized ligand is sorted and delivered for the most part to lyso- somes for degradation.
According to current understanding of RME, the receptor-mediated targeting of drugs has been considered promising as a drug delivery system (DDS) (for review, see Refs [13-201). In this DDS, drug or peptide is covalently or non- covalently coupled to a polypeptide carrier; the prepared drug-carrier complex should still bind to receptor, without significantly reducing the internalization capability. Moreover, gene fusion technology has produced cytokine-toxin com- plexes [21,22]. Fig. 1 indicates how a drug-poly- peptide complex may undergo cellular processing via the RME. The nature of receptor recycling enables the continuous delivery of drug-poly- peptide complexes into the target cells. Lysosom- al degradation of the drug-carrier complex (with an acid-sensitive linkage) may result in the re- lease of a drug into the cytosol, where it can exert its effect. Sometimes, endosomes merely cross polarized endothelial or epithelial cells in such a manner that the ligand remains intact (transcytosis). For example, transferrin remains bound to its receptor in the endosome because of its high affinity at acid pH, and the transferrin- receptor complex recycles back to the plasma membrane without degradation [23-261. where- upon the ligand dissociates from the receptor. Transcytosis of other polypeptides has also been
(outside)
WCeptClr binding
r* w+
.*
.* ‘r’r Cdl surface y y y y y y
membrane
(Inside) t recycling
internalization
Fig. I. A schematic diagram for receptor-mediated endo-
cytosis (RME) of drug-polypeptide complex. Intracellular
routings of polypeptide and receptor are described in the
text.
reported [27-301. Thus, drug-polypeptide com- plexes have been designed to cross the blood- brain barrier (BBB) by means of transcytosis. These include insulin [31] and transferrin [32,33], although the brain selectivity of these complexes in vivo remains unclear. The findings that ligand translocates from the endosome to the nucleus [34-371 has opened an avenue with which to develop a carrier for nuclear drug targeting. For example, albumin conjugated with the peptide RRRGL, corresponding to residues 31-35 of human angiogenin, has been specifically im- ported into the nucleus [38]. Endosome-disrup- tive peptides [39] may help drug-carrier com- plexes leave the endosome for the cytosol before sorting to lysosomes. This strategy is important for drugs sensitive to inactivation in lysosomes. In the RME-utilizing DDS, therefore. a poly- peptide ligand can be recognized as a transport carrier or a “vector” to efficiently deliver other- wise impermeable substances into, or across, cells. Gene transfer into endothelial cells and lymphocytes from patients ex vivo and grafting them to the same individual (i.e., gene therapy)

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 445-467 447
has allowed several kinds of human genetic diseases, especially those caused by single gene enzyme deficiencies, to be treated [40-421. Ac- cordingly, there is increasing usage of polypep- tides for DNAioligonucleotide delivery due to their capability to undergo RME in vitro. As an example, Fig. 2 illustrates an insulin-[N-acylurea albumin] conjugate bound electrostatically to NE0 gene, which was used for the DNA deliv- ery into HepGz cells via endocytosis with insulin receptors [43]. Table 1 summarizes applications of receptor-mediated drug delivery between 1990-1995 (prior to 1990, see [15] and [16]).
The success of the RME-utilizing drug target- ing is dependent not only on the in vitro efficacy of targeting but on the in vivo behavior of the drug-carrier complex. Therefore, it is important to kinetically analyze the RME processes of polypeptides in order to estimate both their targeting efficiency and pharmacokinetics in a comprehensive manner. In this review, therefore, we will focus upon the kinetic modeling analysis of polypeptide RME using isolated or cultured cells in vitro, perfused organs and the whole body in vivo, in order to provide kinetic insights
CELL MEMBRANE r 1 DNA
-
INSULIN RECEPTOR COMPLEX
, J lNsuLlN - [N-ACYLUREA
ALBUMIN] CONJUGATE
Fig. 2. Binding interactions of a conjugate consisting of a
polypeptide ligand such as insulin (designated as I) covalently
cross-linked to N-acylurea albumin. The N-acylurea albumin
(A-CDI). produced by carbodiimide modification of al-
bumin, bears numerous positive charges and is electrostatical-
ly bound with negatively charged DNA.
into receptor-mediated drug-polypeptide target- ing.
2. Factors influencing polypeptide phannacokinetics/pharmacodynamics
General considerations about the various fac- tors influencing peptide pharmacokinetics have been provided by McMartin and Bennett (44,451, and some of these factors have been incorpo- rated into physiologically based models [46-491 to predict the pharmacokinetics of peptides and proteins [50-521. If the target site (receptor-rich cells) resides within the vascular compartment where exogenously administered polypeptide can easily gain access, targeting efficiency may be relatively high. On the other hand, if the target site is located outside the vascular compartment, the polypeptide will encounter several limiting factors before reaching the target cells. These are enzymatic degradation either in blood or on the endothelial cell surface, non-specific elimination by the liver and kidney, binding with plasma proteins [53], especially a,-macroglobulin [54- 581, binding with heparin [59-611, blood flow to the target site, transcapillary permeability [62- 65] and interstitial diffusion [66,67]. Thus, the in vivo efficacy of polypeptides may be offset by some of these physiological barriers. Moreover, macromolecules with high receptor binding af- finity may not diffuse into the kernel of the target tissue due to efficient binding and uptake at the perivascular site (so-called “binding site barrier”), as demonstrated for antibody percola- tion into cancer tumors [68-701.
When a polypeptide is mainly cleared by the RME system, the RME should be considered as a predominant factor in its PK/PD. For example, the remarkable dose-dependence and analogue
( or antibody) blockade of peptide phar- macokinetics may result from saturable, recep- tor-mediated clearance [71-781. A clearance- mediating receptor (C-receptor) distinct from that (B-receptor) mediating biological actions has been reported for atria1 natriuretic factor (ANF) [79] and its pharmacokinetic role has been demonstrated [72,73,80]. Reversible recep- tor binding serves as a substantial depot of

448 H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 445%467
Table 1 Recent applications of receptor-mediated drug delivery (1990-1YYS)
Drug/gene Carrier Experimental method Ref No.
(i) Genes and antisense oligonucleotides
PEPCK-hFIX gene Galactosylated polylysine
CAT DNA Galactosylated histone
Luciferase gene Lactosylated polylysine
c-myb antisense Transferrin-polylysine
Luciferase gene
CAT DNA
CAT antisense
DTA”
Oligonucleotides
Neo gene
Factor IX gene
DTA”
Interleukin-6
Transferrin-polylysine
Asialoglycoprotein-polylysine
Asialoorosomucoid-polylysine
Asialoorosomucoid
Asialofetuin-polylysine
Insulin
Adenovirus
Virosome
Gene engineered diphteria toxin
Rat in vivo
HepGz cells
HepGz cells
Leukemia cells
Eukaryotic cells
Rat in vivo
NIH 3T3 cells
Hepatocytes
HepGl cells
HepGz cells
Hepatocytes
BHK-21 cells
Eukaryotic cells
11641
[I651 [166]-
[I671 [168,169]
11701 [I711 u721 [I731 I431 I1741 I1751 [I761
(ii) Antineoplastic agemr
Doxorubicin
Methotrexate
Eiliptinium-oleate
ILEh
Neocarzinostatin
(iii) Antiviral agents
ara-C
AZT
PMEA’
(iv) Peptides and proteins
Horseradish peroxidase
Superoxide dismutase
HIV-1 derived
synthetic peptide
(v) Miscellaneous agents
Chlorin e6
Naproxen
Maleylated BSA
Maleylated BSA
LDL
LDL
Transferrin
Glycosylated dextran
Mannosylated HSA
Mannosyl polymer
Insulin fragment
Mannosylated derivative
a,-Macroglobulin
Insulin
Lysozyme
Lymphoma cell line
Macrophages
916 melanoma
A549 cells
Leukemia K562 cells
[I771 [I781 [I791
[W [loll
Mouse in vivo
MT-4 cells
Macrophages
[1SIl
[I821
(1831
Mouse in vivo
Macrophages 1311 [I841
Mouse in vivo [I851
HepGL cells
Rat in vivo 1341
N61
” Diphtheria toxin fragment A.
h 2-morpholinomethyl-2’,3’,4’-trimethoxy acrylophenone hydrochloride.
’ 9-(2-phosphonylmethoxyethyl)adenine
polypeptide in its in vivo distribution, at the level of the whole body [81] and organs [82-861. Moreover, polypeptide pharmacokinetics may vary depending on alterations in receptor num- bers (up- and down-regulation) [87-891, endog- enous ligands [90] and soluble receptors [91-961. The importance of RME in the overall phar- macokinetics of polypeptide hormones has been emphasized from a kinetic viewpoint [97-991.
At the cellular level, the pharmacological
effect of a drug-polypeptide complex may be dependent on the targeting efficiency and cellular processing via the RME. Potential problems of receptor-mediated drug targeting have been de- scribed comprehensively in previous reviews [15,16,100]. Ultimately, post-endocytotic intracel- lular routing of a drug-polypeptide complex should affect RME-utilizing drug delivery. The use of transferrin as a carrier would be useful for helping cell-impermeable drugs traverse endo-

H. Sato et al. I Advanced Drug Delivery Reviews 19 (19%) 445-467 449
thelial cells without degradation, but not for launching drugs within target cells. The complex- ity associated with RME, however, is that the kinetic properties of RME may vary depending on the ligands and cell types. For example, 25% of the internalized neocarzinostatin-transferrin conjugate was degraded in the lysosomes of leukemia cells, while transferrin alone was not degraded at all [loll. Moreover, fibroblasts de- grade complexes of internalized epidermal growth factor (EGF) and its receptor 11021, whereas pancreatic carcinoma cells recycle them back to the cell surface [103].
For any kinetic analysis of polypeptide, an adequate assay system has to be established with which to measure intact polypeptide. There are several means of doing so, including bioassays, trichloroacetic acid (TCA)-precipitation [104], gel chromatography, radioimmunoassays (RIA), radioreceptor assays (RRA), enzyme-linked im- munosorbent assays (ELISA) and high-perform- ance liquid chromatography (HPLC). A com- parison of these methods is not within the scope of this review. However, it must be pointed out that assay results can differ depending on the method used, due to differences in the resolution of metabolite separation [105,106] or in the recognizable locus of the polypeptide between the receptor and an antibody [107]. Moreover, when a radiolabeled peptide is used, the labeled material has to have sufficient receptor binding activity. Occasionally, labeled polypeptide loses its receptor binding activity due to a conforma- tional change, or iodination at a biologically important tyrosine residue. It is not surprising that receptor-mediated clearance and distribu- tion cannot be demonstrated using such a radio- labeled ligand in vivo. This problem has been experienced with some peptide hormones includ- ing insulin [108,109] and /3-endorphin [85]. Thus, experimental conditions (receptor-binding affini- ty, administered dose and the assay method) should also be included as variables influencing peptide pharmacokinetics. These factors should be considered especially when comparing pep- tide kinetic studies from different laboratories. Table 2 summarizes the factors which may in- fluence polypeptide PK/PD as mentioned above.
Table 2 Various factors which may influence the pharmacokinetics and pharmacodynamics of drug-polypeptide complex
(i) Physiological factors’ * Enzymatic degradation in the blood or on endothelial cells * Non-specific elimination by the liver and the kidney . Receptor-mediated clearance by the liver and the kidney - Binding to plasma proteins (such as cu,-macroglobulin) * Binding to heparin * Binding to soluble receptor . Blood flow to target site . Transcapillary permeability . Interstitial diffusion * Prevascular distribution (“binding-site barrier”)
(ii) RME activity of target cell * Competitive endogenous Iigand . Receptor number on cell surface . Rate of receptor-mediated endocytosis * Rate of receptor synthesis and degradation * Rate of receptor recycling * Up- and down-regulation of surface receptor
(iii) Intracellular fate in target cell Trafficking and translocation from endosome Disruption of drug-polypeptide linkage in lysosome Release of drug into cytosol Stability of drug in lysosome
iv) Experimental conditions Dose administered
. Route of administration * Molecular ratio of drug/carrier - Effect of labeling on receptor binding activity * Analytical methods
a Pathophysiological changes of these factors should be also considered.
These factors have to be considered to better incorporate PK/PD principles in the develop- ment of polypeptides and proteins and their drug delivery systems for therapeutic use.
3. Kinetic analysis of RME in vitro and in perfused organs
So far, a large number of kinetic studies have been performed with regard to the RME of polypeptides. Using epidermal growth factor (EGF) as an example, its in vitro RME kinetics

450 H. Suto et al. I Advanced Drug &livery Reviews 19 (1996) 44.f-467
were first elucidated by Wiley and Cunningham using human fibroblasts [llO,ll l] and later ex- tended to a state-of-the-art kinetic model (112,113]. Later, the hepatic handling of EGF via the RME mechanism was described kinetically using isolated and cultured hepatocytes [97,114- 1181 and perfused livers [83,89,97-99,114, 119,120]. As such, the RME processes of EGF in the liver have been extensively studied from various aspects. It is difficult to cover all of them here. Therefore, only typical RME analyses are described in this review.
Among the several processes of RME (Fig. l), binding of ligand to its receptor is the first step that triggers the internalization of ligand-recep- tor complex. Hence, the binding kinetic parame- ters of k,,, (association) and k,,rr (dissociation), as well as surface receptor number ([R,]), have to be measured in order to quantify the dynamic ligand-receptor interaction. However, intracellu- lar degradation of ligand and subsequent ac- cumulation of degradation products in the ex- tracellular medium may significantly hamper the accurate assay of receptor binding, based on unchanged ligand concentrations. Moreover, re- ceptor binding equilibrium should be altered by the internalization process. Even if the ligand concentration bound to receptors is appro- priately measured as opposed to that which was internalized, the apparent dissociation constant (K,,app) obtained by Scatchard analysis is not necessarily a valid measurement of ligand-re- ceptor affinity (i.e.. K, = k,,,,lk,,,) when RME is taking place. According to the formulation by Wiley and Cunningham [ill], K,,app can be expressed as follows:
K+pp = koff + kint ’ kt
k k Cl” Ill,
where k,,, and k, represent the endocytotic rate constants of occupied and unoccupied receptors, respectively. If the endocytosis of unoccupied receptors is neglected from the formulation pro- cess, then the following equation holds:
K,,app = k,” + km,
k 011
Therefore, it is a prerequisite to suppress RME
activity during the receptor binding assay. The temperature can be easily decreased to 4°C for this purpose. However, when receptor-binding kinetics are temperature-dependent, the use of phenylarsine oxide (PhAsO), NaN,, or KCN to inhibit RME may be an appropriate means of measuring true receptor-binding parameters in intact cells at 37°C (110,121,122].
The multiple indicator dilution (MID) tech- nique developed by Goresky et al. [123,124] has been employed to measure kinetic parameters representing the relatively rapid interactions be- tween polypeptides and the liver cell surface in the perfused liver [83,89,98,11.5,125]. In a MID experiment, a test polypeptide and an extracellu- lar reference compound ( 14C-labeled inulin) are administered as a bolus into the portal vein, and the outflow in the hepatic vein is sampled in about 0.5-l s aliquots. The concentration-time profiles (“dilution” curves) of both compounds in the outflow are simultaneously analyzed by a mathematical model [126]. Thus, some receptor- related parameters, including R,, k,, and k,,(,, can be determined. The key features of the MID method are, 1) the structural architecture of the liver as an in vivo system is maintained, 2) the effect of endogenous substances can be ignored, 3) multiple injectates can be consecutively tested, and 4) the concentration of drug, inhibitors, or plasma proteins, plasma flow-rate and tempera- ture can be controlled.
-3.1. Estimation of RME parameters ,from l&and movement
A mathematical model for the movement of extracellular and intracellular ligands, as well as of intracellular degradation products, is pre- sented in Fig. 3. This model is a topology of the RME scheme shown in Fig. 1 with respect to the movement of ligand, and is essentially the same as those previously developed for EGF [127] and insulin [128]. Since lysosomal delivery of poly- peptide is supposed to take place before peptide degradation (or digestion), the fraction (f) of internalized ligand that is sorted to lysosomes is parameterized. When the intracellular drug con- centration ( [Di]) is to be simulated, considera- tions should include the generation of the drug

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 445-467 4.51
ligand
cell surface degradation
products
kint A
extracellular extracellular space space
Fig. 3. A generic model for the internalization and degra- dation of drug-ligand (polypeptide) complex via the RME mechanism. 12,. L,, and Ldcg represent the amounts of surface-bound ligand, internalized ligand, and intracellular degradation products, respectively; k,“, represents the rate constant for internalization of drug-ligand-receptor com- plex; k_, is the externalization rate constant of intracellular receptor; k,,, and krV, define the degradation of L, and subsequent release of L,,, into the extracellular fluid, respec- tively: and f denotes the fraction of internalized ligand that is sorted to lysosomes; on the other hand, (1 -f) is assumed to be the fraction of internalized ligand that is not degraded but transcytosed (e.g., f=l for most polypeptides but f=O for transferrin).
from an intralysosomal drug-polypeptide com- plex (f.Li), with the rate constant, kgen, as well as the sequestration (degradation and extracellu- lar release) of the generated drug with the rate constant, kXeq. Thus, the equation d[D,]ldt= k,,;f[L,] -k,,,[D,] may hold for an intracellu- larly generated drug which is pharmacologically active. However, the feasibility of this equation remains to be confirmed due to the lack of corresponding experimental data. In fact, poly- peptide sorting in endosomes is a complex pro- cess and has been elaborately modelled [129,130].
Our task is to estimate the endocytotic rate constant (k,,,) and the intracellular degradation rate constant (kdcg) of the labeled polypeptide. The experimental procedure for this analysis is to incubate a labeled polypeptide (conjugated with drug) with isolated cells or perfused tissue at 37°C for a short period and then to measure the surface-bound and internalized ligands by an acid-wash [110,111,122,131]. The logic of the
acid-wash is that the radioactivity removed from the cells at low pH at O-PC represents “surface- bound” ligand, while the acid-resistant radioac- tivity associated with the cells represents “inter- nalized” ligand. In a perfused organ, the acid- wash should be performed after removing the extracellular polypeptide ligand that is not bound with surface receptors [119,128,132]. The inter- nalized ligand is further analyzed for the un- changed (L,) and degraded (Ldeg) ligands. For simplicity, let us assume that no transcytosis of internalized ligand (Li) with its receptor (i.e., f=O) takes place. Thus, the mass-balance equa- tions for the internalized ligand (L,) and its intracellular degradation products (Ldeg) can be written as:
g&l dt = kint[L,l - kdcg[Lil
Wdegl ~ = k&,1 - kelLegl dt
The summation of Eq. (3) and Eq. (4) and assuming the condition of kre,[Ldeg] <<k,,,[L,] (i.e., when the amount of intracellular degra- dation products is relatively low compared with surface-bound ligand) yields:
Wil + dL_J dt = kint[I. ] 5 (5)
The integration of Eq. (5) from time 0 to t yields:
[LB1 + [Ldegl= kint _/ Psldt 0
(6)
Thus, kint can be estimated from the initial slope of an “integration plot”, that is, a plot of inter- nalized radioactivity ([L,] + [Ldeg]) versus the integrated amount of surface-bound ligand
WJ j- 1 L, Id t can be calculated numerically by the trap%zoidal rule. As far as the acid-wash is used to measure the surface-bound and internal- ized ligands, the same equations can be applied to a perfused organ system. This is feasible when the intracellular degradation products cannot be neglected, but the release of degradation prod- ucts into the medium is not yet significantly increased [128,132]. Moreover, when intracellular degradation of the ligand is negligible during the

452 H. Saio et al. I Advanced Drug Delivery Reviews 1Y (1996) 44-T-467
experiment, Eq. (6) is approximated to [L,], so that:
This equation has been readily applied to a relatively stable polypeptide, EGF [ 114,119]. Moreover, if we assume that [L,] is almost constant, Eq. (7) can be solved as:
[L,I kl
= k,,, . t
This equation corresponds to the simple plot of the “In/Sur ratio” as a function of time. which was first derived by Wiley and Cunningham for EGF [llO,lll] and that has been applied to other polypeptides [122,133-1361.
Assuming in Eq. (4) that [L,,,] is small enough for the condition of k,,,[L,,,]<k,,,[L,] and subsequent integration from time 0 to t yields:
lLdegl = kdeg 1 CLildt 0
(9)
Thus, kdrg can be estimated from the initial slope of a plot of the intracellular degradation products versus the integrated amount of internalized ligand, as demonstrated for EGF [128,137].
These methods of estimating k,,, and kdeg are feasible only for a short time interval (e.g., 5-20 min), where a significant amount of degradation products may not be generated and released from the cell. Therefore, the calculated values may serve as “initial” estimates for the overall fitting of the data to Eq. (3) and Eq. (4). Moreover, these methods cannot be used to monitor the intracellular routing of a ligand for a relatively long interval. Alternatively, “pulse- chase” can be applied to investigate the intracel- lular itinerary of ligand more precisely [26,138- 1421. Briefly, labeled polypeptide is incubated with cells at 0°C for 60 min, and the extracellular polypeptide is removed by washing. Endocytosis of the surface-bound ligand is initiated by warm- ing the cells to 37°C. At various times after the start of incubation, the cells are harvested and the surface-bound (L,) and internalized (L,)
ligands are separated by an acid-wash at 4°C. Thus, the pulse-chase method allows the “single- wave” internalization of surface receptors to be observed, whereas the conventional steady-state method [ 1 LO,11 l] allows the continuous internali- zation (or multiple rounds) of receptors.
3.2. Estimation of RME parameters from receptor movement
Generally, internalized receptors behave dif- ferently from their ligand. A mathematical model for the surface and intracellular receptors is presented in Fig. 4. This model is a topology of the RME scheme shown in Fig. 1 with respect to the movement of the receptor. Our aim is to estimate the endocytotic rate constants (kinr and k,) and the externalization rate constant of the internalized receptor (k,,,). The experimental procedure for this analysis is to incubate isolated cells or perfused tissue with a large excess of unlabeled polypeptide to cause rapid internaliza- tion of the surface receptors, then to measure the ligand binding activity on the cell surface at 4°C. For simplicity, both the degradation rate con- stant of internalized receptor (k,,,,,) and the
unoccupied occupied
1 II\
k ext
+, } 1 $ kinl
intracellular space
Fig. 4. A generic model for internalization and externaliza-
tion of receptor movement via the RME mechanism. R, and
R, define the number of surface and internalized receptors,
respectively; v represents the fraction of receptor occupancy
on the cell surface; k,,, and k, represent the endocytotic rate
constants of occupied and unoccupied receptors on the cell
surface, respectively; v,~” denotes the de novo synthesis rate
of receptor; and k,,,,, is the degradation rate constant of
internalized receptor.

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 445-467 453
new receptor synthesis rate (v,,,) are assumed to be zero, so that the total number of mobile receptors ( [RT]) is constant (= [R,] + [Ri]), for a short time. The mass-balance equations for the surface receptor number ([R,]) can then be written as:
dR1 dt = kext[‘il - {kt(l - v) + kint . ~>[kl
(10)
where Y represents the fraction of receptor occupancy (= [L,]/[R,]). When a high concen- tration of ligand is exposed to cells, v falls to unity and Eq. (10) reduces to:
4R51 dt = kextlIR~l - kint[Rsl (11)
The integration of Eq. (11) from time 0 to t and rearrangement yields:
ln ( Rlll - Psleq = (k + k
RI - RL, ) tnt ext
)t
(12)
where subscripts 0 and eq represent the initial condition (t =0) and the equilibrium condition where [R,] reaches the minimum, respectively. According to Eq. (11) [R,]” and [R,],, are given by:
No = k, :;Iex, ’ rRTl (13)
[R~leq = k,,, + k,,, ’ LRTI
The rearrangement of Eq. (12) yields:
In [R,]:[R,;1J, - (Y = (kin, + k,,,t)
= k down-regulation *t
where (Y and kdown_regu,at,on are defined
RI,, k + Lt a=[R,lo= kint + kcxt
(14)
(15)
as:
(16)
(17)
The kdown-rcgulatmn value can be determined from the slope of the plot of ln{(l-(Y)/([R,]/ [R,],,-a)} versus the time curve when the sur-
face receptors are being “down-regulated”. Di- viding Eq. (14) by [RT] gives:
kxt [RJe, _ RT km, + Lt
(18)
Thus, the RME parameters can be estimated as follows:
down-regulation
=al/3.k down-regulatmn
k,,, = k down-rrgulatm - kext
k,=a.k down-regulation - L,
where p is defined as:
(19)
(20)
(21)
LRTI
p = ~W,, (22)
Standaert and Pollet [143] derived some equa- tions equivalent to Eqs. (10-22) and determined the values of CX, p and kdown_regu,at,on as 0.55, 1.11 and 0.22 h ’ , respectively, according to the rapid “down-regulation” of surface receptors in vitro (Fig. 5). The values of k,,,, kint and k, were estimated as 0.11, 0.11 and 0.011 h- ’ , respective- ly, using Eqs. (19-21) for the movement of insulin receptors in myocytes (Fig. 5) [143].
The next objective is to examine whether the down-regulation of surface receptors is revers- ible. The extracellular ligand should be removed and the cells washed by ligand-free medium after the maximum down-regulation of the surface receptors. The surface receptor number may then increase with time depending on the rate of externalization of internalized ligand. However, it should be confirmed by using cyclohexamide that the increase in the receptors is attributed not to newly synthesized receptor but receptor reap- pearance. Now the extracellular ligand is re- moved and there should be no surface-bound ligand, so the receptor occupancy (v) can be set at zero and Eq. (10) reduces to:
(23)

44 H. Sate et al. I Advanced Drug Delivery Reviews 19 (1996) 44.7-467
I. I. I. 10 20 30 40
TIME (hours)
Fig. 5. Kinetics of insulin-induced down-regulation and suh-
sequent recovery of cell surface insulin receptors in the
myocytes. The cells were incubated at 37°C for the indicated
times in the presence or absence (control) of 20 nM insulin.
After washing extensively. surface “rf-labeled insulin binding
was determined. Following maximum down-regulation of
surface receptors (0.45 at 20 h), the cells were washed. and
insulin-free conditioned media was added. The inset shows
these data plotted as first-order kinetic processes as described
in the text. Reproduced from Ref [143] with permission.
The integration of Eq. (23) from time 0 to t and rearrangement yields:
,n (24)
where subscript ss represents the steady-state condition where [R,] is maximum. According to Eq. (23) [R,],, is given by:
k, [Rslss = k, ;;,,, ’ lRTl (25)
receptor recovery reflects not only k,,, but also k,. However, krecovery can be approximated as k,,,, since k, is usually smaller than k,,,. The value of krCCOVCrV can be determined from the slope of the plot’of ln{(r- l)l(y- [R,]/[R,],,)} versus time. This parameter, as well as k down-rcgulallon’ is very important to optimize the interval of drug-polypeptide administration, be- cause if the receptors are once down-regulated, the next dosage should be given only after the surface receptors are restored. Standaert and Pollet [143], determined krecovery as 0.12 h-’ from the kinetics of surface receptor recovery (Fig. 5). This value is entirely consistent with the sum of k, and k,,, estimated from the kinetics of re- ceptor down-regulation (Fig. 5) indicating the feasibility of the “receptor-recycling” model (Fig.
4). Rearrangement of Eq. (1.5) gives:
z = (1 - LY). exp{ - (k,,, + k,,,)t} + a (29) \ 0
Essentially the same approach was employed to estimate k,,, (0.22 min-‘) and k,,, (0.11 mini’) [128] to accommodate the ligand-induced “rapid down-regulation” of surface receptors in vivo (Fig. 6) [144]. Insulin was injected into rats at a high dose and the time course of ligand- binding activity on the cell surface was measured. In a perfused organ system. the receptor re- covery can be chased with the single-pass ex- traction measurement [83] after the surface re- ceptor is down-regulated. Analysis of the re- covery phase of the curve in Fig. 6 with Eq. (24)
Rearrangement of Eq. (24) yields:
In Y-l
y - [R,] / [R,],, = (k’ + kCx’)r
= krec<,oerr . t
where y and k I
rL‘o\‘CT\I are defined as: 8 16 24 40 80 120 180
k.. 1 =k,+k,,, (27) Time of sacrifice after insulin injection (min) r“OL’LI”
VU,
’ = RI,, (28)
The apparent slope (krCCOVelV) of the surface
Fig. 6. Liver insulin binding activity at various times following
insulin injection in rats. The binding assay was performed
both on intact (filled circles) and Triton X-lOO-solubilized
(untilled circles) liver particles. Reproduced from Ref [I441
with permission.

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996j 445-467 45s
is not adequate, because the receptor recovery process is inevitably affected by high insulin concentrations in plasma (delayed recovery of surface receptors due to continuous internaliza- tion). For the same reason, the discrepancy between the k,,, of the insulin receptor (0.035 min-‘) estimated by Frank et al. [145] and that (0.11 mini’) we estimated as above might be attributed to the high plasma concentration (45 nM) of insulin in their study.
Moreover, a dynamic RME model of poly- peptide in an intact, perfused organ (Fig. 7) can be developed by combining a recirculatory sys- tem with generic RME models (Fig. 3 and Fig. 4) under some of the assumptions described above (i.e., f=O; kdeg, R=O; vSyn=O). The non- linear clearance and surface binding of insulin in the perfused livers were analyzed [128] based on this model. The mass-balance differential equa- tions for Fig. 7 are as follows:
d[R,l dt = ‘ext[R,l- ‘in,&,1 - ‘,([R,l- &,I)
(30)
Reservoir (V,)
Fig. 7. The “receptor-recycling” model of hepatic insulin
clearance and distribution. Solid and broken arrows indicate
the movement of insulin and its receptor, respectively. The
liver is subdivided into three compartments. i.e., extracellular
fluid. cell surface and intracellular (vesicular) space, which
are assumed to be well-mixed. C, and C, denote the poly-
peptide concentrations in the perfusate and extracellular
fluid, respectively. The other nomenclature is presented in
the legends to Fig. 3 and Fig. 4. The total number of mobile
receptors R., (= R, + R,) and receptor affinity K, (=k,,,, lk,,,)
are assumed to be constant.
WI 2 = k,nt[L,l + k,([R,l - Psi) - kcxt[Ril dt
(31)
v W,l - = Q,W,l - [C,l> p dt (32)
v d[C,l - = Q,W,l - [Ccl> e dt
- k,“~cJRI - [U + k&s1 (33)
$$ = k,,[C,IRI - [Ll) - (k,,, + k,Ll
(34)
d[Lil dt = k,nt[Lsl - kdeg[Lil
4Lde,l ~ = kcieg[Lil - krel[Ldrgl dt
(35)
(36)
The nomenclature is presented in the legends to Fig. 3 and Fig. 4. These equations were simul- taneously fitted to the observed time courses of
C,, L L, and Ldel! by a nonlinear least-squares regression analysis, to optimize R,, k,,, and k_. For this fitting procedure, the initial estimates of
k,“, and k&g were used as obtained by the “ratio plot” analysis (i.e., Eq. (6) and (9)). The initial estimation of R, is described in Ref [128]. The other parameters (VP, V,, Q,, k,,, kc,,.,, k, and k,,,) were experimentally measured, cited, or calculated from published references. The theo- retical curves for C,, L,, L, and Ldeg were in good agreement with the observed data (Fig. 8) [128]. The optimized values of R,, k,,, and kdcg were 8.66 pmol per g liver, 0.62 min -’ and 0.91 min -‘, respectively. Thus, the k,,, of insulin in different types of cell is in the order of hepatocytesbkidney (0.022 mini ‘) [132]> adipocytes (0.0027 min-‘) [146] >myocytes (0.0018 mini’) [143], in consistent with the ability of these tissues to degrade insulin. The subsequent computer-simulation (Fig. 9) clearly showed that the recovery half-life of the surface receptor is not constant, but varies with the initial dose added to the perfusate, reflecting the insulin disappearance from the circulation. The dynamic “receptor-recycling” model (Fig. 7) is

4% H. Sate et al. I Advanced Drug Delivery Reviews 19 (19%) 44-f-467
Time (min) Time (min)
Fig. 8. The disappearance of “‘I-labeled insulin by cyclically
perfused mouse livers at low (0.018 nM) and high (2 nM)
initial concentrations (in panel A). In panel B, the timc-
courses of surface-bound (unfilled circles) and internalized
(filled circles). as well as of intracellular degradation products
(tilled triangles), are presented. Solid lines show computer-
generated simulation curves using the model shown in Fig. 6.
Reproduced from Ref 1128) with permission.
useful when predicting the changes in surface receptor number and surface/intracellular ligand concentrations in the target tissue, which should be important for the pharmacodynamics (onset, offset and duration of effect) of a drug-poly- peptide complex in general. An important advan-
0.01 I 0 40 80 120
Time (min)
tage of physiologically based models relative to the traditional compartmental types is the possi- bility of predicting the changes in drug levels and physiological factors in practically inaccessible or specific tissue sites, thus making it possible to predict data by changing the selected parameters involved in the model.
4. Kinetic analysis of RME in vivo
Among several means of evaluating the in vivo effect of tissue uptake via the RME [97-99,147], in vivo tissue uptake [84] is a rapid and simple procedure applicable to various polypeptides and tissues. The experimental protocol is as follows. Animals receive an intravenous (i.v.) injection of labeled polypeptide with several doses of un- labeled polypeptide, and arterial blood is col- lected at designated times (4-5 points over 20- 150 s). The amount of labeled ligand taken up by
40 80
Time (min)
120
Fig. 9. Computer simulation of the time-dependent changes in the perfusate concentration of insulin (A) and surface receptor
number (B) at varying initial concentrations of insulin in the reservoir. using the “receptor-recycling” model of hepatic insulin
clearance and distribution.

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 44.5-467 451
each tissue is determined at 3 min postdose. The apparent tissue uptake rate is described by:
dL1, - = ~Lptake~qJ, dt (37)
where [C,],, [L,], and CLuprakt: represent the plasma concentration, the amount of polypeptide associated with tissue at time t and the tissue uptake clearance, respectively. Integration of Eq. (37) from time 0 to t yields:
Ll, = CLptake . AUC”-, + MO (38)
where AUC,_, and [L,], represent the area under the plasma concentration (AUC)-time profile from time 0 to t and the amount of polypeptide in tissue at time 0, respectively.
Thus, CLuptakc can be estimated from the initial slope of an “integration plot” of [L,], versus
AUC,,_,. In each organ, the integration plot gives
0
-2
-4 t
Liver (22 min) \
%
\
+,
Jejunum (2.4 hr)
2 4 6 2 4 6
Ileum (2.2 hr) Spleen 14.5 hrl
0 2 4 6 0 2 4 6 0 2 4 6 0
Stomach (3.5 hr)
0 2 4 6
Or
a straight line passing through the y-axis inter- cept, [L,],. Therefore, dividing the [L,],, value by the initial plasma concentration of the polypep- tide ([C,],) yields the initial distribution volume of the tissue ([K,],,), which represents the vol- ume to which polypeptide can be distributed in the tissue of interest within a short time. The tissue uptake of EGF [84] and hepatocyte growth factor (HGF) [148] have been analyzed by this procedure. From the CLuptak, values in various tissues, it was found that the hepatic and renal uptake clearances were approximately 50% and 15% of the early-phase clearance of EGF, re- spectively [88]. A useful application of this tech- nique is to analyze the down-regulation and subsequent recovery of surface receptors (Fig. 10) [Ml. To do so, an i.v. injection of a high dose of polypeptide is followed either by a binding assay of the tissue surface receptors or by a second injection of tracer polypeptide to measure
I Kidney (4.6 hrl I Duodenum (2.0 hr)
The (h)
Fig. 10. Sigma-minus plot for the determination of recovery half-lives of tissue uptake clearance of [‘ZSI]-labeled epidermal growth
factor (EGF). In normal rat, tissue uptake clearance [CL,,,,,_(O)] was determined after intravenous injection of [“‘I]-labeled
EGF. At time t after the administration of excess (300 pglkg) unlabeled EGF, tissue uptake clearance [CL,,,,,,(t)] of
[“SI]-labeled EGF was determined. Adopted from Ref [88] with permission.

Drliverv Rrview.c 1Y (lYY6) 44.5-467
CLupltik, during the recovery of surface receptors. The “sigma-minus” plot of ln[CL,,,,,,(O) - CL uptake(t)] (difference of CLuprakc between time 0 and t) versus time will give a straight descend- ing line, and the ratio of 0.693/t,,, represents
krccWcry which is the sum of k,,, and k,, as mentioned above. It is assumed that the plasma concentration of exogenously administered poly- peptide becomes low enough to neglect the continuous endocytosis of surface receptors at time t. As shown in Fig. 10, the rate constant for the surface receptor recovery in the liver is much higher than those in other tissues, suggesting that the kinetic properties of EGF receptors differ among tissues,
If we further consider the capillary permeabili- ty (PS) and intrinsic uptake clearance (CL,,,) of a ligand under a pseudo steady-state condition, CL,,,,,,, can be also expressed as follows [149]:
PS . CL,“, CLupNX = ps + CL
I I, I
This equation assumes that the blood flow is not rate-limiting the ligand uptake into the tissue. If PS is relatively large compared with CL,,,,. CL uptnkc is approximately equal to CL,,,,, and
CL,,,, reflects the endocytotic uptake of the ligand as:
CLJC,l = k,,,,F-,I (40)
Since [L,] is the concentration of occupied sur- face receptors. Y [R,] (v=[C,], Eq. (40) can be formulated as:
CLI”JC,J = k”, . [WqJ
K d’ app + [C P ]
Substituting K,,app using Eq. (2:
~(&app + [C,l b
(41)
1 (a simpler form of K,,app) and solving for CL,,, yields:
k ,n, kJR,l cLm’ = k,, + k ,,,, + k,,,[C,,l (42)
If [C,] is low (i.e.. at a tracer concentration) enough for the condition k,,,, + k,,, >> k ,,,, [C,] to hold, then Eq. (42) simplifies to:
k . km l&l cL1nl = i:,,, + k,,,, (43)
Eq. (43) was already utilized in a previous paper [87] without an exact formulation. On the other hand, if [C,] is high enough for the condition k,,f, +k ,,,, <<k,,,[C,,] to hold, then Eq. (42) re- duces to:
cL _ k,Rl I”[ --Kg- (44)
This equation indicates that the uptake rate, CL,,,,[C,], is almost constant at high ligand concentrations. According to Eq. (18), [R,] at high ligand concentrations under a steady-state condition ([R,],,) is expressed as:
L, L&l IR& = k,,, + k,,,
Therefore, Eq. (44) can be formulated as:
cL~~~~[c,~l = km, . LPGI
k,,, + k,,,
(45)
(46)
Since k,,, is usually smaller than k,,,, the above equation can be approximated to:
CL,,[C,l = k,,[%l (47)
This equation suggests that ligand uptake via RME is limited for the most part by receptor externalization when the ligand concentration is very high and the surface receptor is significantly down-regulated. Therefore, receptor recycling (or reutilization) may be important for the poly- peptide clearance at its higher concentrations. but not at lower concentrations, as suggested in a previous paper which compared “receptor-recy- cling” and “non-receptor recycling” models for insulin clearance in the perfused liver [128]. Eqs. (42-44) and Eqs. (46,47) are useful when pre- dicting explicitly the in vivo clearance of poly- peptides in an organ using the in vitro parame- ters of RME, as follows:
Q . fs . CL,,,, cLr,an = Q + fB . CL,,, (48)
where Q is the blood flow rate and fB is the fraction of a polypeptide unbound in blood.

H. Sate et al. I Advanced Drug Delivery Reviews 19 (1996) 44.5-467 459
5. Implications of RME kinetics for clinical application of polypeptides
From the RME kinetics of insulin in the perfused liver [128], the mean time of receptor recycling (T,,,) was calculated to be 11 min (= I lki”t + l lk,,,), suggesting that the recycling of insulin receptors will harmonize with plasma insulin concentrations, if insulin is administered in pulses at about ll-min intervals. In fact, reports [150-1.541 have shown that the pulsatile administration of insulin with 11-13 min inter- vals reduces hypoglycemia (or hyperinsulinemia) compared with continuous administration. More- over, the pulsatile administration of insulin may reduce insulin resistance by reversing the down- regulation of insulin receptors. The general no- tion that harmonization of drug administration with receptor recycling could provide more effi- cient and safer therapy, will be useful for the clinical application of other polypeptides and proteins.
In practice, the clinical application of poly- peptides will be targeted to patients for a par- ticular disease. Therefore, the PKlPD analysis of polypeptides in diseased model animals is im- portant [87,155]. For example, a significant de- crease and subsequent recovery of CLuptak, in the liver was observed after Ccl, treatment, whereas no change was seen in kidney, spleen, or lung [155].
6. Perspectives
In fact, there has not been much progress from the “natural” targeting of glycoproteins and LDL to hepatocytes or macrophage lineages, for the most part due to the effects of unfavorable factors on the PK/PD of polypeptides (see Table 2). Drug delivery using neoglycoproteins [156], such as mannosylated and lactosylated albumin, and nonprotein macromolecules such as galac- tosylated polylysine, has also been limited to the same types of cells (Table 1). Nevertheless, the ability to manipulate even these two cell types would have a major impact in the therapy of a large number of disease states related to these cells (such as hepatocellular carcinoma). To fur-
ther accomplish extrahepatic drug targeting via RME, we need to identify or engineer other types of polypeptide carriers that are little taken up by the liver and macrophages, but efficiently distributed into selected target sites such as tumors. Polypeptides such as cytokines and growth factors have been identified and cloned, and their structural and functional characteristics have been classified and stored in databases [157,158]. Some of the available polypeptides, including soluble receptors for cytokines [91-961, might be good carriers for receptor-mediated drug delivery.
Although there are some physiological barriers for polypeptides as listed in Table 2, it may be possible to overcome these factors by miniaturiz- ing polypeptides to increase their tissue/capillary diffusibility, and by modification of polypeptides to protect against non-receptor-mediated seques- tration mechanisms. For instance, the systemic administration of HGF as a heparin-bound form was proposed (Fig. 11) [13]. The heparin-HGF complex was two-three times more stable than HGF itself, with the hepatocyte proliferative activity retained. The same strategy could be applied to other heparin binding polypeptides, such as tumor necrosis factor (TNF) [59] and fibroblast growth factor (FGF) [60,61]. In gener- al, the polypeptides classified as “paracrines” and “autocrines” are not appropriate for systemic administration unless they are stabilized some- how.
Intracellular trafficking of receptors is directly influenced by the pH sensitivity of the receptor/ ligand dissociation [159,160]. The gene-en- gineered LDL receptor without a property of ligand release at acid pH was no longer recycled back, but was degraded intracellularly [160]. Moreover, molecular mechanisms of lysosomal sorting have been clarified [161-1631. Therefore, neo polypeptides that can regulate their own destination in the cell may be engineered in future. Perhaps the ideal complex for receptor- mediated drug delivery is a multi-functional protein, which consists of a homing device (“navigator”), receptor targeting device (“recog- nize?), intracellular targeting device (“door- opener”) and a drug.
Finally, the therapeutic efficacy/toxicity of a

460 H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 44-f-467
l HGF
0 ep Heperin
$J HCF recepror
y tleperin-Ike substance
l P L$Aa eP
eP l P ,BQv v
/
00 a. I
out I
I VV
In HGF receptor Heparin-like substance
receptor-mediated endocytosle
\/ 1
StitWfation of DNA Synthesis non-specific uptake
Fig. 11. Schematic diagram of a drug delivery system of HGF based on the mechanism of its hepatic handling. HGF molecules
carried in the blood flow can bind to both HGF receptor and a heparin-like substance [ 1871, followed by internalization via each
binding site. On the other hand, the binding of the heparin-HGF complex to a heparin-like substance is reduced by pre-masking
of the heparin-binding domain of HGF with heparin. Adopted from Ref [13] with permission.
drug-carrier conjugate should be assessed from its microscopic pharmacology based on the RME mechanisms, in conjunction with macroscopic pharmacokinetic modelling.
Acknowledgments
This work was supported by a grant from the Research Foundation for Pharmaceutical Sci- ences and Uehara Memorial Foundation, awarded to H.S.
References
[II
PI
PI
141
PI
Stahl. P. and Schwartz, A.L. (1986) Receptor-mediated
endocytosis. J. Clin. Invest. 77, 657-662.
Wileman, T.. Harding, C. and Stahl, P. (1985) Receptor-
mediated endocytosis. Biochem. J. 232, 1-14.
Goldstein, J.L.. Brown, M.S., Anderson, R.G., Russell,
D.W. and Schneider. W.J. (198.5) Receptor-mediated
endocytosis: concepts emerging from the LDL receptor
system. Ann. Rev. Cell Biol. 1. l-39.
Carpentier, J.L. (1984) Receptor-mediated endocytosis
of polypeptide hormones: mechanism and significance.
Mol. Immunol. 21, 1157-llS9.
Pastan, I.H. and Willingham, M.C. (1981) Journey to the
center of the cell: role of the receptosome. Science 214.
504-509.
PI
[71
PI
I91
1101
IllI
[I21
Gorden, P., Carpentier, J.L.. Freychet, P.O. and Orci. L. (1980) Internalization of polypeptide hormones: mecha-
nism, intracellular localization and significance. Dia-
betologia 18, 263-274. Brown, M.S. and Goldstein, J.L. (1979) Receptor-me-
diated endocytosis: insights from the lipoprotein recep-
tor system. Proc. Natl. Acad. Sci. U.S.A. 76, 3330-3337.
Goldstein, J.L., Anderson, R.G. and Brown, M.S. (1979)
Coated pits, coated vesicles, and receptor-mediated
endocytosis. Nature 279, 679-685.
Olsnes. S., Kozlov, J.V., van Deurs, B. and Sandvig, K.
(1991) Bacterial protein toxins acting on intracellular targets. Sem. Cell Biol. 2. 7-14.
FitzGerald. D. and Pastan, I. (1991) Redirecting Pseudo-
monas exotoxin. Sem. Cell Biol. 2, 31-37.
Carrasco, L. (1994) Entry of animal viruses and macro- molecules into cells. FEBS Lett. 350, 151-154.
Pauza, C.D. (1991) The endocytic pathway for human
immunodeticiency virus infection. Adv. Exp. Med. Biol.
300, 11 l-138.
[I31 Sugiyama, Y. and Kato. Y. (1994) Pharmacokinetic
aspects of peptide delivery and targeting: importance of
receptor-mediated endocytosis. Drug Develop. Ind.
Pharm. 20, 591-614.
1141
[ISI
Pardridge, W.M. (1992) Recent developments in peptide
drug delivery to the brain. Pharmacol. Toxicol. 71, 3-10.
Basu, S.K. (1990) Receptor-mediated endocytosis of
macromolecular conjugates in selective drug delivery.
Biochem. Pharmacol. 40. 1941-1946.
[I’51 Meijer, D.K. and van der Sluijs, P. (1989) Covalent and
noncovalent protein binding of drugs: implications for
hepatic clearance, storage, and cell-specific drug deliv- ery. Pharm. Res. 6, 105-118.
1171 Fiume, L.. Busi, C., Mattioli, A. and Spinosa. G. (1988)

1181
u91
PO1
1211
WI
1231
P41
W
(271
P91
(301
1311
H. Sato et al. / Advanced Drug Delivery Review 19 (1996) 445-467
Targeting of antiviral drugs bound to protein carriers. Crit. Rev. Ther. Drug Carrier Sys. 4, 265-284. Dragsten, P.R., Mitchell, D.B., Covert, G. and Baker, T. (1987) Drug delivery using vesicles targeted to the hepatic asialoglycoprotein receptor. Biochim. Biophys. Acta 926, 270-279. Fallon, R.J. and Schwartz, A.L. (1985) Receptor-me- diated endocytosis and targeted drug delivery. Hepato- logy 5. 899-901. Fiume, L., Busi, C. and Mattioli, A. (1983) Targeting of antiviral drugs by coupling with protein carriers. FEBS Lett. 153, 6-10.
(1987) Chimeric peptides as a vehicle for peptide phar- maceutical delivery through the blood-brain barrier. Biochem. Biophys. Res. Commun. 146, 307-313.
[34] Akhlynina, T.V., Rosenkranz, A.A., Jans, D.A. and Sobolev, A.S. (1995) Insulin-mediated intracellular targeting enhances the photodynamic activity of chlorin e6. Cancer Res. 55, 1014-1019.
[351 Moroianu, J. and Riordan, J.F. (1994) Nuclear transloca- tion of angiogenic proteins in endothelial cells: an essential step in angiogenesis. Biochemistry 33, 12535- 12539.
[361
Pastan, 1. and FitzGerald, D. (1991) Recombinant toxins for cancer treatment. Science 254, 1173-I 177. Pastan, 1. and FitzGerald, D. (1989) Pseudomonas exotoxin: chimeric toxins. J. Biol. Chem. 264, 1X57- 15160.
[37J
Ciechanover. A., Schwartz, A.L., Dautry-Varsat, A. and Lodish, H.F. (1983) Kinetics of internalization and recycling of transferrin and the transferrin receptor in a human hepatoma cell line. Effect of lysosomotropic agents. J. Biol. Chem. 258, 9681-9689. Klausner, R.D., Ashwell, G., van Renswoude, J., Har- ford, J.B. and Bridges, K.R. (1983) Binding of apo- transferrin to K562 cells: explanation of the transferrin cycle. Proc. Natl. Acad. Sci. U.S.A. 80, 2263-2266. Dautry-Varsat, A., Ciechanover, A. and Lodish, H.F. (1983) pH and the recycling of transferrin during re- ceptor-mediated endocytosis. Proc. Natl. Acad. Sci. U.S.A. 80. 2258-2262.
[381
1391
Hopkins, C.R. and Trowbridge, I.S. (1983) lnternaliza- tion and processing of transferrin and the transferrin receptor in human carcinoma A431 cells. J. Cell Biol. 97, 508-521.
1401
[411
1421
[431
Laduron, P.M. (1994) From receptor internalization to nuclear translocation. New targets for long-term phar- macology. Biochem. Pharmacol. 47, 3- 13. Rosenkranz, A.A., Yachmenev, S.V., Jans, D.A.. Serebryakova, N.V., Murav’ev, V.I., Peters, R. and Sobolev, A.S. (1992) Receptor-mediated endocytosis and nuclear transport of a transfecting DNA construct. Exp. Cell Res. 199, 323-329. Moroianu, J. and Riordan, J.F. ( 1994) Identification of the nucleolar targeting signal of human angiogenin. Biochem. Biophys. Res. Commun. 203, 1765-1772. Plank, C., Oberhauser, B., Mechtler, K., Koch, C. and Wagner. E. (1994) The influence of endosome-disruptive peptides on gene transfer using synthetic virus-like gene transfer systems. J. Biol. Chem. 269, 12918-12924. Friedmann, T. (1994) Gene therapy for neurological disorders. Trends Genet. 10, 210-214. Roemer, K. and Friedmann, T. (1992) Concepts and strategies for human gene therapy. Eur. J. Biochem. 208, 211-225.
Dan, N. and Cutler. D.F. (1994) Transcytosis and processing of intrinsic factor-cobalamin in Caco-2 cells. J. Biol. Chem. 269, 18849-18855. Ghinea, N., Mai, T.V., Groyer-Picard, M.T. and Milgrom, E. (1994) How protein hormones reach their target cells. Receptor-mediated transcytosis of hCG through endo- thelial cells. J. Cell Biol. 125, 87-97. Magnusson, S., Faerevik, 1. and Berg, T. (1992) Charac- terization of retroendocytosis in rat liver parenchymal cells and sinusoidal endothelial cells. Biochem. J. 287, 241-246.
Friedmann, T. (1989) Progress toward human gene therapy. Science 244, 1275-128 I Huckett, B., Ariatti, M. and Hawtrey, A.O. (1990) Evidence for targeted gene transfer by receptor-me- diated endocytosis. Stable expression following insulin- directed entry of NE0 into HepG2 cells. Biochem. Pharmacol. 40, 253-263.
]44
(45
McMartin, C. (1989) Molecular sieving, receptor pro- cessing and peptidolysis as major determinants of pep- tide pharmacokinetics in vivo. Biochem. Sot. Trans. 17, 931-934. Bennett, H.P. and McMartin, C. (1978) Peptide hor- mones and their analogues: distribution, clearance from the circulation. and inactivation in vivo. Pharmacol. Rev. 30, 247-292.
Maratos-Flier, E., Kao, C.Y., Verdin, E.M. and King, G.L. (1987) Receptor-mediated vectorial transcytosis of epidermal growth factor by Madin-Darby canine kidney cells. J. Cell Biol. 105, 1595-1601. Fukuta. M., Okada, H.. Iinuma. S., Yanai, S. and Toguchi, H. (1994) insulin fragments as a carrier for peptide delivery across the blood-brain barrier. Pharm. Res. II. 1681-1688.
]461
]471
Gerlowski, L.E. and Jain, R.K. (1983) Physiologically based pharmacokinetic modeling: principles and applica- tions. J. Pharm. Sci. 72, 1103-1127. Lin, J.H.. Sugiyama, Y., Awazu, S. and Hanano. M. (1982) Physiological pharmacokinetics of ethoxyben- zamide based on biochemical data obtained in vitro as well as on physiological data. J. Pharm. Sci. IO, 649-661.
[48] Tsuji, A.. Yoshikawa, T.. Nishide, K., Minami, H., Kimura, M., Nakashima, E., Terasaki, T.. Miyamoto, E., Nightingale, C.H. and Yamana, T. (1983) Physiologically based pharmacokinetic model for p-lactam antibiotics I: Tissue distribution and elimination in rats. J. Pharm. Sci. 72, 123991252.
[32] Bickcl. U., Yoshikawa, T., Landaw, E.M., Faull, K.F. and Pardridge, W.M. (1993) Pharmacologic effects in vivo in brain by vector-mediated peptide drug delivery. Proc. Natl. Acad. Sci. U.S.A. 90, 2618-2622.
[33] Pardridge, W.M.. Kumagai, A.K. and Eisenberg, J.B.
461

462 H. Sate et nl. I Advanced Drug Delivery Reviews 19 (IW5) 445-467
[49] Bischoff, K.B. and Dedrick, R.L. (1968) Thiopental
pharmacokinetics. J. Pharm. Sci. 57, 1346-1351.
[SO] Baxter. L.T.. Zhu. H., Mackensen, D.G. and Jain. R.K.
(1994) Physiologically based pharmacokinetic model for
specific and nonspecific monoclonal antibodies and frag-
ments in normal tissues and human tumor xenografts in
nude mice. Cancer Res. 54. 1517-1528.
[Sl] Sato, H., Sugiyama, Y.. Sawada, Y.. Iga, T. and Hanano,
M. (1987) Physiologically based pharmacokinetics of
radioiodinated human P-endorphin in rats. An applica-
tion of the capillary membrane-limited model. Drug
Metab. Dispos. 15, 540-550.
[S2] Covell. D.G.. Barbet. J.. Holton, G.D., Black. C.D.V.,
Parker, R.J. and Weinstein. J.N. (1986) Phar-
macokinetics of monoclonal immunoglobulin Gl.
F(ab’)2, and Fab’ in mice. Cancer Res. 46. 3969-397X.
IS31 Sato, H., Sugiyama, Y.. Sawada, Y.. Iga. ‘I‘. and Hanano.
M. (1985) Binding of radioiodinated human p-endor-
phin to serum proteins from rats and humans, detcr-
mined by several methods. Life Sci. 37. 1309-131X.
[S4] Soker, S.. Svahn, C.M. and Neufeld. G. (1993) Vascular
endothelial growth factor is inactivated by binding to
Luz-macroglobulin and the binding is inhibited by
heparin. J. Biol. Chem. 268. 76X5-7691.
[SS] Wollenberg. G.K.. LaMarre. J.. Rosendal, S.. Cionias.
S.L. and Hayes. M. (1991) Identification of cr-macro
globulin as a carrier of human tumor necrosis factor-
alpha: preferential binding to cu,-macroglobulin-plasma
or cuz-macroglobulin-methylamine. Am. J. Pathol. 13X.
265.
(561 Borth. W. and Luger. T.A. (1989) Identilication of CY,-
macroglobulin as a cytokine binding plasma protein.
Binding of interleukin-18 to “F” cu,-macroglobulin. J.
Biol. Chem. 264. 5818.
[S7] Dennis, P.. Saksela, O., Harpcl. P. and Rifkin. D.B.
(1989) a:-Macroglobulin is a binding protein for basic
tibroblast growth factor. J. Biol. Chcm. 264. 7210-7216.
[5X] Huang. J.S.. Decker, S.S. and Deuel. T.F. (19X4) Specific
covalent binding of platelet-derived growth factor to
human plasma cu?-macroglobulin. Proc. Natl. Acad. Sci.
IJ.S.A. 81, 342.
159) Lantz, M., Thysell. H., Nilsson. E. and Glsson. I. (1991)
On the binding of lumor necrosis factor (TNF) to
heparin and the release in vivo of the TNF-binding
protein I by heparin. J. Clin. Invest. 88, 2026-2031.
[60] Copeland. R.A., Ji. H., Halfpenny. A.J.. Williams. R.W..
Thompson, KC.. Herber. W.K., Thomas K.A.. Brunei-,
M.W., Ryan, J.A.. Marquis-Omer. D. et al. (1901) The
structure of human acidic fibroblast growth factor and its
interaction with heparin. Arch. Biochem. Biophys. 2X9.
53-61.
(611 Baird, A., Schubert, D.. Ling. N. and Guillemin. R.
(1988) Receptor- and heparin-binding domains of basic
fibroblast growth factor. Proc. Natl. Acad. Sci. U.S.A.
8.5, 2324-232X.
[62] Ader, M. and Bergman, R.N. (1994) Importance of
transcapillary insulin transport to dynamics of insulin
action after intravenous glucose. Am. J. Physiol. 266.
E I7-E2S.
1631 Sung, C.. Youle, R.J. and Dedrick, R.L. (1990) Phar- macokinetic analysis of immunotoxin uptake in solid
tumors: role of plasma kinetics, capillary permeability,
and binding. Cancer Res. SO, 7382-7392.
1641 Yang, Y.J., Hope. I.D., Ader, M. and Bergman, R.N. ( 1989) Insulin transport across capillaries is rate limiting
for insulin action in dogs. J. Clin. Invest. 84. 1620-1628.
[6S] Rasio, E. (1982) The capillary barrier to circulating insulin. Diabetes Care 5. 15XXlhl.
[66] Jain, R.K. (1989) Delivery of novel therapeutic agents in
tumors: physiological barriers and strategies. J. Natl.
Cancer Inst. 81, 570-576.
1671 Jain, R.K. (1987) Transport of molecules in the tumor
interstitium: a review. Cancer Res. 47, 3039-3051.
[6X] Juweid, M., Neumann, R., Paik, C., Perez-Bacete. M.J..
Sam, J., van Osdol, W. and Weinstein. J.N. (1992)
Micropharmacology of monoclonal antibodies in solid
tumors: direct experimental evidence for a binding site
barrier. Cancer Res. 52, 5144-5153.
[69] Weinstein. J.N. and van Osdol. W. (1992) Early intcrven-
tion in cancer using monoclonal antibodies and other
biological ligands: micropharmacology and the “binding
site barrier”. Cancer Res. 52, 2747s-2751s.
1701 Fujimori. K., Fisher. D.R. and Weinstein, J.N. (1991)
Integrated microscopic-macroscopic pharmacology of
monoclonal antibody radioconjugates: the radiation dose
distribution. Cancer Res. 51, 4821-4827.
(711 Kuwabara. T.. Kato, Y., Kobayashi, S., Suzuki, H. and
Sugiyama, Y. (1994) Nonlinear pharmacokinetics of a
recombinant human granulocyte colony-stimulating fac-
tor derivative (nartograstim): species differences among
rats. monkeys and humans. J. Pharmacol. Exp. Ther.
271. 1535-1543.
1721 Abassi. Z.A.. Tate, J., Hunsberger, S.. Klein, H., Trach-
twsky. D. and Keiser. H.R. (1992) Pharmacokinetics of
ANF and urodilatin during cANF receptor blockade and
neutral endopeptidase inhibition. Am. J. Physiol. 263.
EX70-EX76.
173) Barclay. P.L.. Bennett, J.A., Greengrass, P.M.. Griffin,
A.. Samuels. G.M. and Shepperson, N.B. (1992) The
pharmacokinetics of “‘I-atria1 natriuretic factor in
anaesthetized rats. Effects of neutral cndopeptidase
inhibition with candoxatrilat and of ANF-C receptor
blockade. Biochem. Pharmacol. 44, 1013-1022.
1741 Chiu, P.J.. T&off. G.. Romano, M.T.. Foster, C.J. and
Sybertz. E.J. (lY91) Influence of C-ANF receptor and
neutral endopeptidase on pharmacokinetics of ANF in
rats. Am. J. Physiol. 260, R208-R216.
17.51 Kiyokawa, H.. Kono, N., Hamaguchi, T., Kawachi, M.. Tajima, K., Yamada, Y., Hara, N., Mineo. I., Kuwajima.
M. and Tarui. S. (1989) Autoantibodies to the insulin
receptor impair clearance of plasma endogenous insulin.
Diab. Res. Clin. Prac. 7. S41-S44.
1761 Stevenson. R.W., Cherrington, A.D. and Steiner, K.E.
(1985) The relationship between plasma concentration
and disappearance rate of immunoreactive insulin in the
conscious dog. Horm. Metab. Res. 17, 551-553.
(771 Eaton. R.P.. Friedman, N., Allen, R.C. and Schade, D.S.
( 1984) Insulin removal in man: in vivo evidence for a

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 44-Y-467 46.3
receptor-mediated process. .I. Clin. Endocrinol. Metab. 69, 555-559.
[78] Weitzman. R.E. and Fisher. D.A. (1978) Arginine vas- opressin metabolism in dogs. I. Evidence for a receptor- mediated mechanism. Am. J. Physiol. 235, E591-E597.
[79] Nussenzveig, D.R., Lewicki, J.A. and Maack. T. (1990) Cellular mechanisms of the clearance function of type C receptors of atria1 natriuretic factor. .I. Biol. Chem. 265, 20952-20958.
[80] Okolicany, J., McEnroe, G.A., Koh, G.Y., Lewicki, J.A. and Maack, T. (1992) Clearance receptor and neutral endopeptidase-mediated metabolism of atria1 natriuretic factor. Am. J. Physiol. 263, F546-F553.
[St] Zeleznik, A.J. and Roth, J. (1978) Demonstration of the insulin receptor in vivo in rabbits and its possible role as a reservoir for the plasma hormone. J. Clin. Invest. 61. 136331374.
[82] Sato, H., Terasaki, T. and Tsuji, A. (1990) Specific binding and clearance of [‘Hldynorphin (1-13) in the perfused rat lung: an application of the multiple-in- dicator dilution method. J. Pharm. Pharmacol. 42, 879- 882.
[83] Sato, H., Sugiyama, Y., Sawada, Y., Iga, T., Sakamoto, S.. Fuwa, T. and Hanano, M. (1988) Dynamic determi- nation of kinetic parameters for the interaction between polypeptide hormones and cell-surface receptors in the perfused rat liver by the multiple-indicator dilution method. Proc. Natl. Acad. Sci. U.S.A. 85, 8355-8359.
[84] Kim, D.C., Sugiyama, Y., Satoh, H., Fuwa, T., Iga, T. and Hanano, M. (1988) Kinetic analysis of in vivo receptor- dependent binding of human epidermal growth factor by rat tissues. J. Pharm. Sci. 77, 200-207.
[85] Sato. H., Sugiyama, Y., Sawada, Y., Iga, T. and Hanano, M. (1988) In vivo evidence for the specific binding of human P-endorphin to the lung and liver of the rat. Biochem. Pharmacol. 37, 2273-2278.
[86] Whitcomb, D.C., O’Dorisio, T.M., Cataland, S., Shet- zline, M.A. and Nishikawara, M.T. (1985) Identification of tissue insulin receptors: use of a unique in vivo radioreceptor assay. Am. J. Physiol. 249, E561-E567.
[87] Sato. H.. Terasaki, T., Okumura, K. and Tsuji, A. (1991) Effect of receptor up-regulation on insulin phar- macokinetics in streptozotocin-treated diabetic rats. Pharm. Res. 8, 563-569.
1881 Yanai, S., Sugiyama. Y., Iga, T., Fuwa. T. and Hanano, M. (1990) Kinetic analysis of the downregulation of epidermal growth factor receptors in rats in vivo. Am. J. Physiol. 258, C593-C598.
[89] Sato, H., Sugiyama, Y., Kim, D.C., Yanai, S., Kurita, M., Fuwa, T., Iga, T. and Hanano, M. (1989) Decrease in the number of receptors for epidermal growth factor in the liver of D-galactosamine-intoxicated rats. Biochem. Pharmacol. 38, 2663-2671.
[90] Wong. M.W. and Jamieson, J.C. (1979) Evidence for reduced uptake of asialo-al-acid glycoprotein during the acute phase response to inflammation. Life Sci. 25. 827- 834.
[91] Stasi, R., Zinzani, L., Galieni, P., Lauta, V.M., Damasio, E., Dispensa, E., Dammacco, F.,Venditti, A., Del Poeta,
G., Cantonetti, M. et al. (1995) Clinical implications of cytokine and soluble receptor measurements in patients with newly-diagnosed aggressive non-Hodgkin’s lym- phoma. Eur. J. Haematol. 54, 9-17.
[92] Klein, B. and Brailly, H. (1995) Cytokine-binding pro- teins: stimulating antagonists. Immunol. Today 16, 216- 220.
[93] Debets, R. and Savelkoul, H.F. (1994) Cytokine antago- nists and their potential therapeutic use. Immunol. Today 15, 455-458.
[94] Mohler, K.M., Torrance, D.S., Smith, C.A., Goodwin, R.G.. Stremler, K.E., Fung, VP., Madani, H. and Widmer, M.B. (1993) Soluble tumor necrosis factor (TNF) receptors are effective therapeutic agents in lethal endotoxemia and function simultaneously as both TNF carriers and TNF antagonists. J. Immunol. 151, 154881561.
[95] Beguin, Y. (1992) The soluble transferrin receptor: biological aspects and clinical usefulness as quantitative measure of erythropoiesis. Haematologica 77, l-10.
[96] Campbell, P.G. and Novak, J.F. (1991) Insulin-like growth factor binding protein (IGFBP) inhibits IGF action on human osteosarcoma cells. J. Cell. Physiol. 149, 293-300.
[97] Sugiyama, Y. and Kato, Y. (1995) In vitro models of hepatic clearance: Methods to determine the kinetic parameters for receptor-mediated hepatic uptake. In: M.D. Taylor and G.L. Amidon (Eds.), Peptide Based Drug Design: Controlling Transport and Metabolism, ACS Professional Reference Books, Washington, DC. pp. 525-551.
[98] Sugiyama, Y.. Kim, D.-C., Satoh. H., Yanai, S., Sato. H., Iga, T. and Hanano, M. (1990) Receptor-mediated disposition of polypeptides: Kinetic analysis of the transport of epidermal growth factor as a model peptide using in vitro isolated perfused organs and in vivo system. J. Control. Rel. 13, 157-174.
[99] Sugiyama. Y. and Hanano, M. (1989) Receptor-mediated transport of peptide hormones and its importance in the overall hormone disposition in the body. Pharm. Res. 6, 192-202.
[lOO] Fallon. R.J. and Schwartz, A.L. (1989) Receptor-me- diated delivery of drugs to hepatocytes. Adv. Drug Delivery Rev. 4, 49-63.
[loll Kohgo, Y., Kondo, H., Kato, J., Sasaki, K., Tsushima, N., Nishisato, T., Hirayama, M., Fujikawa, K., Shintani. N., Mogi, Y. et al. (1990) Kinetics of internalization and cytotoxicity of transferrin-neocarzinostatin conjugate in human leukemia cell line, K562. Jpn. J. Cancer Res. 81, 91-99.
[102] Stoscheck, C.M. and Carpenter, G. (1984) Down regu- lation of epidermal growth factor receptors: direct demonstration of receptor degradation in human ti- broblasts. J. Cell Biol. 98, 1048-1053.
[103] Korc, M. and Finman, J.E. (1989) Attenuated process- ing of epidermal growth factor in the face of marked degradation of transforming growth factor-a. J. Biol. Chem. 264, 14990-14999.
[104] Chen, M., Berman, J.R., MaCarroll, M.C. and Chen,

464
[W
IW
[IO71
[lOU
[IO91
[1101
I1111
[114
[113]
]ll41
[1151
[1161
[I171
[1181
H. Suto et al. I Advanced Drug Delivery Reviews I9 (1996) 445-467
I.W. (1981) Method for rapidly estimating specific
radioactivity and radiochemical purity of radioiodi-
nated peptide hormones used in radioimmunoassays.
Clin. Chem. 27, 632-633.
factor by rat hepatocytes: receptor pathway. J. Cell
Biol. 102, 24-36.
Sato, H.. Tsuji, A., Hirai, K. and Kang, Y.S. (1990)
Application of HPLC in disposition study of A,,-“CI-
labeled insulin in mice. Diabetes 39. 563-569.
Stentz. F.B.. Harris, H.L. and Kitabchi. A.E. (1983)
Early detection of degraded A,,-“il-insulin in human
Iibroblasts by the use of high performance liquid
chromatography. Diabetes 32. 474-477.
Besser, G.M., Orth, D.N., Nicholson. W.E., Byyny.
R.L., Abe, K. and Woodham, J.P. (1971) Dissociation
of the disappearance of bioactive and radioim-
munoreactive ACTH from plasma in man. J. Clin.
Endocrinol. Metab. 32. 595-603.
[ 1191 Sato, H., Sugiyama, Y., Sawada, Y., Iga, T., Fuwa, T.
and Hanano. M. (1990) Internalization of EGF in
perfused rat liver is independent of the degree of
receptor occupancy. Am. J. Physiol. 258. G682-G689.
[120] Dunn. W.A. and Hubbard, A.L. (1984) Receptor-me-
diated endocytosis of epidermal growth factor by
hepatocytes in the perfused rat liver: ligand and re-
ceptor dynamics. J. Cell Biol. 98, 2148-2159.
I1211
Cockram, C.S.. Jones, R.H., Boroujerdi, M.A. and
Sonksen, P.H. (1984) Evidence for separate handling in
vivo of different regions of the insulin molecule using
A14- and Bl-labeled insulin tracers. Diabetes 33, 721-
727.
Wardzala. L.J., Simpson, I.A.. Rechler, M.M. and
Cushman, S.W. (1984) Potential mechanism of the
stimulatory action of insulin on insulin-like growth
factor II binding to the isolated rat adipose cell.
Apparent redistribution of receptors cycling between a
large intracellular pool and the plasma membrane. J.
Biol. Chem. 259, 8378-8383.
Berman, M., McGuire, E.A.. Roth, J. and Zeleznik.
A.J. (1980) Kinetic modeling of insulin binding to
receptors and degradation in vivo in the rabbit. Dia-
betes 29, 50-59.
Draznin, B., Trowbridge, M. and Ferguson, L. (1984)
Quantitative studies of the rate of insulin internaliza-
tion in isolated rat hepatocytes. Biochem. J. 218, 307-
312.
11231 Goresky, C.A. (1983) Kinetic interpretation of hepatic
multiple-indicator dilution studies. Am. J. Physiol. 245,
Gl-G12.
Wiley, H.S. and Cunningham, D.D. (1982) The endo-
cytotic rate constant. A cellular parameter for quan-
titating receptor-mediated endocytosis. J. Biol. Chem.
257, 4222-4229.
Wiley, H.S. and Cunningham, D.D. (1981) A steady
state model for analyzing the cellular binding, internali-
zation and degradation of polypeptide ligands. Cell 25.
433-440.
11241 Goresky, C.A., Bach, G.C. and Nadeau, B.E. (1973)
On the uptake of materials by the intact liver. The
transport and net removal of galactose. J. Clin. Invest.
52, 991-1009.
[I251 Whiteside, C.I., Lumsden, C.J. and Silverman, M.
(1988) In vivo characterization of insulin uptake by dog
renal cortical epithelium. Am. J. Physiol. 255, E357-
E365.
Gex-Fabry, M. and DeLisi, C. (1986) Regulation of
interacting populations during endocytosis: models of
growth factor-tumor promoter dynamics. Am. J. Phy-
siol. 250. R1123-R1132.
11261 Zeleznik, A.J. and Roth, J. (1978) Demonstration of
the insulin receptor in vivo in rabbits and its possible
role as a reservoir for the plasma hormone. J. Clin.
Invest. 61, 1363-1374.
Gex-Fabry, M. and DeLisi, C. (1984) Receptor-me-
diated endocytosis: a model and its implications for
experimental analysis. Am. J. Physiol. 247, R768-R779.
Kato, Y., Sato, H.. Ichikawa, M.. Suzuki. H., Sawada. Y.,
Hanano, M., Fuwa, T. and Sugiyama. Y. (1992) Exist-
ence of two pathways for the endocytosis of epidermal
growth factor by rat liver: phenylarsine oxide-sensitive
and -insensitive pathways. Proc. Natl. Acad. Sci. U.S.A.
89, 8507-851 I. Yanai, S., Sugiyama, Y., Iga. T., Fuwa. 7‘. and Hanano.
M. (1991) Comparison of the methods for determining
cell-surface and intracellular receptors for epidermal
growth factor in the rat liver. Pharm. Res. 8, 557-562.
Jackie, S., Runquist, E.A., Miranda-Brady, S. and
Havel, R.J. (1991) Trafficking of the epidermal growth
factor receptor and transferrin in three hepatocytic
endosomal fractions. J. Biol. Chem. 266, 1396-1402.
Rush, G.F. and Alberts, D. (1987) The hepatic binding
and uptake kinetics of epidermal growth factor: studies
with isolated rat hepatocytes. Life Sci. 40, 679-685.
Dunn, W.A., Connolly, T.P. and Hubbard. A.L. (1986)
Receptor-mediated endocytosis of epidermal growth
11271 Yanai, S., Sugiyama, Y., Kim. D.C., Iga, T., Fuwa, T.
and Hanano, M. (1991) Kinetic analysis of receptor-
mediated endocytosis of epidermal growth factor by
isolated rat hepatocytes. Am. J. Physiol. 260, C457-
C467.
[1281
11291
11301
[l31]
]l321
Sato, H., Terasaki, T., Mizuguchi, H., Okumura, K. and
Tsuji, A. (1991) Receptor-recycling model of clearance
and distribution of insulin in the perfused mouse liver.
Diabetologia 34, 613-621.
Linderman, J.J. and Lauffenburger, D.A. (1988) Analy-
sis of intracellular receptor/ligand sorting in endo-
somes. J. Theor. Biol. 132, 203-245.
Linderman, J.J. and Lauffenburger, D.A. (1986) Analy-
sis of intracellular receptor/ligand sorting. Calculation
of mean surface and bulk diffusion times within a
sphere. Biophys. J. 50, 295-305.
Haigler, H.T., Maxfield, F.R.. Willingham, M.C. and
Pastan, I. (1980) Dansylcadaverine inhibits internaliza-
tion of 1251-epidermal growth factor in BALB 3T3
cells. J. Biol. Chem. 255, 1239-1241.
Sato, H., Yoshioka, K., Terasaki, T. and Tsuji, A.
(1991) Receptor-mediated endocytosis of A,,-“iI-in-

H. Sato et al. I Advanced Drug Delivery Reviews 19 (1996) 445-467 465
sulin by the nonfiltering perfused rat kidney. Biochim.
Biophys. Acta 1073, 442-450. (1331 Cozza, E.N., Vila, M.C. and Gomez-Sanchez, C.E.
(1990) ET-I receptors in C-6 cells: homologous down-
regulation and modulation by protein kinase C. Mol.
Cell. Endocrinol. 70, 155-164.
Bagheri, A., Harris, S. and Reed, B.C. (1989) An
analysis of the relationship between the cellular dis-
tribution and the rate of turnover for the separate
classes of unoccupied, noncovalently occupied, and
covalently occupied insulin receptor. J. Biol. Chem.
264, 12673-12679.
[134] Eskild, W.. Kindberg, G.M., Smedsrod, B., Blomhoff, [147] Lai, W.H., Cameron, P.H., Wada, I., Doherty, J.J., Kay,
R.. Norum, K.R. and Berg, T. (1989) Intracellular D.G., Posner, B.1. and Bergeron, J.J. (1989) Ligand-
transport of formaldehyde-treated serum albumin in mediated internalization, recycling, and downregulation
liver endothelial cells after uptake via scavenger re- of the epidermal growth factor receptor in vivo. J. Cell
ceptors. Biochem. J. 258, 511-520. Biol. 109, 2741-2749.
(1351 Zoon, K.C., Zur Nedden, D., Hu, R. and Arnheiter, H.
(1986) Analysis of the steady state binding, internaliza-
tion, and degradation of human interferon-a,. J. Biol.
Chem. 261, 4993-4996.
[136] Sharma, R.J. and Grant, D.A. (1986) A differential
effect between the acute and chronic administration of
ethanol on the endocytotic rate constant, ke, for the
internalisation of asialoglycoproteins by hepatocytes.
Biochim. Biophys. Acta 862, 199-204.
(1371 Kato, Y. and Sugiyama, Y. (1993) Binding, internaliza-
tion, degradation, and mitogenic effect of epidermal
growth factor in cultured rat hepatocytes. S.T.P.
Pharma Sci. 3, 75-82.
[148] Liu, K.X., Kato, Y., Narukawa, M., Kim, D.C., Hanano.
M., Higuchi, O., Nakamura, T. and Sugiyama, Y. (1992)
Importance of the liver in plasma clearance of hepat-
ocyte growth factors in rats. Am. J. Physiol. 263, G642-
G649.
[149] Sato, H., Sugiyama, Y., Miyauchi, S., Sawada. Y.. Iga, T.
and Hanano, M. (1986) A simulation study on the
effect of a uniform diffusional barrier across hepat-
ocytes on drug metabolism by evenly or unevenly
distributed uni-enzyme in the liver. J. Pharm. Sci. 75.
3-8.
11381 Decker, S.J. (1990) Epidermal growth factor and trans-
forming growth factor-a induce differential processing
of the epidermal growth factor receptor. Biochem.
Biophys. Res. Commun. 166, 615-621.
[139] Gladhaug, I.P. and Christoffersen, T. (1988) Rapid
constitutive internalization and externalization of epi-
dermal growth factor receptors in isolated rat hepat-
ocytes. Monensin inhibits receptor externalization and
reduces the capacity for continued endocytosis of
epidermal growth factor. J. Biol. Chem. 263. 12199-
12203.
(1501 Logan-Darrough, M. (1995) Pulsatile iv. insulin
therapy for severely out of control diabetes. J. Intrav.
Nurs. 18, 124-128.
[151] Schmitz, O., Pedersen, S.B., Mengel, A.. Porksen, N.,
Bak, J., Moller, N., Richelsen, B., Alberti, K.G.. Butler,
P.C. and Orskov, H. (1994) Augmented effect of short-
term pulsatile versus continuous insulin delivery on
lipid metabolism but similar effect on whole-body
glucose metabolism in obese subjects. Metab. Chn.
Exp. 43, 842-846.
[140] Presky, D.H. and Schonbrunn, A. (1986) Receptor-
bound somatostatin and epidermal growth factor are
processed differently in GH4Cl rat pituitary cells. J.
Cell Biol. 102, 878-888.
[152] Paolisso, G., Scheen, A.J., Giugliano, D., Sgambato, S.,
Albert, A.,Varricchio, M., D’Onofrio, F. and Lefebvre,
P.J. (1991) Pulsatile insulin delivery has greater meta-
bolic effects than continuous hormone administration
in man: importance of pulse frequency. J. Clin. Endo-
crinol. Metab. 72, 607-615.
[141] Clarke, B.L. and Weigel, P.H. (1985) Recycling of the
asialoglycoprotein receptor in isolated rat hepatocytes.
ATP depletion blocks receptor recycling but not a
single round of endocytosis. J. Biol. Chem. 260, 128-
133.
[I531 Bratusch-Marrain, P.R., Komjati, M. and Waldhausl,
W.K. (1986) Efficacy of pulsatile versus continuous
insulin administration on hepatic glucose production
and glucose utilization in type I diabetic humans.
Diabetes 35, 922-926.
[142] Klausner, R.D., Van Renswoude, J., Ashwell, G.,
Kempf, C., Schechter, A.N., Dean, A. and Bridges,
K.R. (1983) Receptor-mediated endocytosis of trans-
ferrin in K562 cells. J. Biol. Chem. 258, 4715-4724.
[143] Standaert, M.L. and Pallet, R.J. (1984) Equilibrium
model for insulin-induced receptor down-regulation.
Regulation of insulin receptors in differentiated BC3H-
1 myocytes. J. Biol. Chem. 259, 2346-2354.
[ 1441 Desbuquois, B., Lopez, S. and Burlet, H. (1982) Lig-
and-induced translocation of insulin receptors in intact
rat liver. J. Biol. Chem. 257, 10852-10860.
[145] Frank, H.J., Donohoe, M.T. and Morris, W.L. (1985)
Effect of glyburide on in vivo recycling of the hepatic
insulin receptor. Am. J. Med. 79, 53-58.
[146] Reed, D.K.. Newton, C., Fraga, M.. Glastad, K.,
[154] Matthews, D.R., Naylor, B.A., Jones, R.G., Ward, G.M.
and Turner, R.C. (1983) Pulsatile insulin has greater
hypoglycemic effect than continuous delivery. Diabetes
32, 617-621.
[155] Liu, K.X., Kato, Y., Yamazaki, M., Higuchi. O., Naka-
mura, T. and Sugiyama, Y. (1993) Decrease in the
hepatic clearance of hepatocyte growth factor in carbon
tetrachloride-intoxicated rats. Hepatology 17, 651-660.
[156] Palomino, E. (1994) “Carobohydrate handles” as natu-
ral resources in drug delivery. Adv. Drug Deliv. Rev.
13, 311-323.
[157] Casciari, J., Sato, H., Durum. S.K., Fiege, J. and
Weinstein, J.N. (1996) Tabular lexicon of cytokine
structure and function. In: B.A. Chabner and D. Longo
(Eds.), Cancer Chemotherapy: Principles and Practice.
Lippincott-Raven Publ., Philadelphia, pp. 785-808.

466 H. Safe et rd. I Advunced Drug Delivrrv Reviews 10 (lYY6) 445-467
[15X] Casciari, J.. Sate. H.. Durum. SK.. Ficge. J. and
Weinstein. J.N. (1996) Reference databases of cytokine
structure and function. In: B.A. Chabner (Ed.).
Chemotherapy and Biological Response Modifiers An-
nual. Elsevier. pp. in press.
[ 1591 French. A.R.. Tadaki, D.K., Niyogi, S.K. and Lauffen-
burger. D.A. (1995) Intracellular trafficking of epider-
ma1 growth factor family ligands is directly influenced
by the pH sensitivity of the receptorlligand interaction.
J. Biol. Chem. 270. 4334-4340.
[I601 Davis. C.G.. Goldstein, J.L., Sudhof, T.C., Anderson.
R.G.. Russell. D.W. and Brown MS. (1987) Acid-
dependent ligand dissociation and recycling of LDL
receptor mediated by growth factor homology region.
Nature 326. 760-765.
[ 1611 D’Souza-Schorey, C.. Li, G.. Colombo. M.1. and Stahl,
I1621
[I631
11641
I1651
[1661
[I671
11681
[I691
(1701
P.D. (1995) A regulatory role for ARF6 in receptor-
mediated endocytosis. Science 267. 1175-l 178.
French, A.R.. Sudlow. G.P., Wiley, S. and Lauffen-
burger, D.A. (1994) Postendocytic trafficking of epider-
ma1 growth factor-receptor complexes is mediated
through saturable and specific endosomal interactions.
J. Biol. Chem. 269. 15749-15755.
Dintzis. SM., Velculescu. V.E. and Pfeffers, S.R. (1994)
Receptor extracellular domains may contain trafficking
information. Studies of the 300.kDa mannosc h-phos-
phate receptor. J. Biol. Chem. 269. 12159-12166.
Perales, J.C., Ferkol. T., Beegen. H., Ratnoff. O.D. and
Hanson, R.W. (1994) Gene transfer in vivo: sustained
expression and regulation of genes introduced into the
liver by receptor-targeted uptake. Proc. Natl. Acad. Sci.
U.S.A. 91. 4086-4090.
Chen, J., Stickles, R.J. and Daichendt, K.A. (1994)
Galactosylated histone-mediated gene transfer and
expression. Hum. Gene Ther. 5, 429-435.
Midoux. P., Mendes, C.. Legrand, A.. Raimond, J..
Mayer, R.. Monsigny, M. and Roche, A.C. ( 1993)
Specific gene transfer mediated by lactosylated poly-t
lysine into hepatoma cells. Nucl. Acid Res. 21. 871-
878.
Citro, G., Perrotti. D., Cucco, C.. D’Agnano, I.. Sacchi.
A.. Zupi. Cr. and Calabretta, B. (1992) Inhibition of
leukemia cell proliferation by receptor-mediated up-
take of c-myb antisense oligodeoxynucleotides. Proc.
Nat]. Acad. Sci. U.S.A. 89, 7031-7035.
Wagner. E., Zenke. M.. Cotten. M., Beug. H. and
Birnstiel. M.L. (1990) Transferrin-polycation conju-
gates as carriers for DNA uptake into cells. Proc. Natl.
Acad. Sci. 1J.S.A. 87, 3410-3414.
Zenke. M., Steinlein, P., Wagner, E.. Cotten, M.. Beug.
H. and Birnstiel, M.L. (1990) Receptor-mediated endo-
cytosis of transferrin-polycation conjugates: an effi-
cient way to introduce DNA into hematopoietic cells.
Proc. Nat]. Acad. Sci. U.S.A. 87, 3655-3659.
Chowdhury. N.R., Wu, C.H., Wu, G.Y., Yerneni, P.C.,
Bommineni, V.R. and Chowdhury. J.R. (1993) Fate of
DNA targeted to the liver by asialoglycoprotem re-
ceptor-mediated endocytosis in vivo. Prolonged per-
11711
11721
[173)
11741
11751
I1771
[I781
11791
[1801
I1811
[W
(I831
sistence in cytoplasmic vesicles after partial hepatec-
tomy. J. Biol. Chem. 268, 11265-11271.
Bunnell. B.A., Askari, F.K. and Wilson. J.M. ( 1992)
Targeted delivery of antisense oligonucleotides by molecular conjugates. Somat. Cell. Mol. Genet. IX, SSO-569.
Chang, T.M., Hossain, A. and Chang, C.H. (1994)
Effect of rate of intracellular transport and diacytosis
on cytotoxicity of hybrid toxins. Study with hybrids
using hepatic asialoglycoprotein receptor-mediated
endocytosis. Biochim. Biophys. Acta 1224, 77-88.
Reinis, M.. Damkova. M. and Korec, E. (1993) Re-
ceptor-mediated transport of oligodeoxynucleotides
into hepatic cells. J. Virol. Methods 42, 99-105.
Cristiano, R.J.. Smith, L.C., Kay, M.A.. Brinkley. B.R.
and Woo, S.L. (1993) Hepatic gene therapy: efficient
gene delivery and expression in primary hepatocytes
utilizing a conjugated adenovirus-DNA complex. Proc.
Nat]. Acad. Sci. U.S.A. YO, 11548-11552.
Bron, R.. Ortiz. A. and Wilschut, J. (1994) Cellular
cytoplasmic delivery of a polypeptide toxin by reconsti-
tuted influenza virus envelopes (virosomes). Biochemis-
try 33, Yl lo-91 17.
Jean, L.F. and Murphy. J.R. (1991) Diphtheria toxin
receptor-binding domain substitution with interleukin
6: genetic construction and interleukin 6 receptor-spe-
cific action of a diphtheria toxin-related interleukin 6
fusion protein. Prot. Eng. 4. 989-994.
Basu. S.. Mukhopadhyay, B., Basu, S.K. and
Mukhopadhyay. A. (1994) Enhanced intracellular de-
livery of doxorubicin by scavenger receptor-mediated
endocytosis for preferential killing of histiocytic lym-
phoma cells in culture. FEBS Lett. 342, 249-2.54.
Mukhopadhyay, A. and Basu, S.K. (1990) Enhanced
intracellular delivery of methotrexate by a receptor-
mediated process. Biotechnol. App. Biochem. 12. 529%
536.
Samadi-Baboli, M., Favre, G., Canal. P. and Soula, G.
(1993) Low density lipoprotein for cytotoxic drug
targeting: improved activity of elliptinium derivative
against 816 melanoma in mice. Br. J. Cancer 68. 319-
326.
Lestavel-Delattre. 8, Martin-Nizard, F., Clavey. V,
Testard. P., Favre. G., Doualin, G., Houssaini. H.S.,
Bard, J.M.. Duriez, P., Delbart, C., et al. (1992) Low-
density lipoprotein for delivery of an acrylophenone
antineoplastic molecule into malignant cells. Cancer
Res. 52, 3629-3635.
Nishikawa. M., Kamijo, A.. Fujita, T., Takakura, Y.,
Sezaki, H. and Hashida, M. (1993) Synthesis and
pharmacokinetics of a new liver-specific carrier, glyco-
sylated carboxymethyl-dextran, and its application to
drug targeting. Pharm. Res. 10, 1253-1261.
Molema, G., Jansen, R.W., Pauwels, R., de Clercq. E.
and Meijer, D.K. (lY90) Targeting of antiviral drugs to
T4-lymphocytes. Anti-HIV activity of neoglycoprotein-
AZTMP conjugates in vitro. Biochem. Pharmacol. 40.
260332610.
Midoux. P., Negre, E., Roche, A.C., Mayer, R., Mon-

H. Sam et al. I Advanced Drug Delivery Reviews 19 (1996) 44.G467 467
signy, M., Balzarini, J., De Clercq, E., Mayer, E.,
Ghaffar. A. and Gangemi, J.D. (1990) Drug targeting:
anti-HSV-1 activity of mannosylated polymer-bound
9-(2-phosphonylmethoxyethyl)adenine. Biochem. Bio-
phys. Res. Commun. 167, 1044-1049.
[184] Takakura. Y., Masuda, S., Tokuda, H.. Nishikawa, M.
and Hashida, M. (1994) Targeted delivery of superox-
ide dismutase to macrophages via mannose reccptor-
mediated mechanism. Biochem. Pharmacol. 47. %3-
858.
[lSS] Mitsuda, S.. Nakagawa. T., Osada, T., Shimamoto, T.,
Nakazato, H. and Ikai, A. (1993) A receptor mediated
delivery of an HIV-l derived peptide vaccine. Biochem.
Biophys. Res. Commun. 194, 11551160.
[186] Franssen. E.J.. Moolenaar, F., de Zeeuw. D. and
Meijer, D.K. (1993) Low molecular weight proteins as
carriers for renal drug targeting: naproxen coupled to
lysozyme via the spacer I -lactic acid. Pharm. Res. 10.
963-969.
[187] Zarnegar. R.. DeFrances. M.C.. Oliver. 1,. and Mich-
alopoulos, G. (1990) Identification and partial charac-
terization of receptor binding sites for HGF on rat
hepatocytes. Biochem. Biophys. Res. Commun. 173.
1179%85.





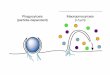









![Intracellular Trafficking Network of Protein Nanocapsules: Endocytosis… · 2016-09-13 · endocytosis, recycling endocytosis and exocytosis pathways [22]. Rab5 and Rab7 have been](https://img.pdfslide.us/doc/110x75/5f34351cd6125f288673d8b5/intracellular-trafficking-network-of-protein-nanocapsules-endocytosis-2016-09-13.jpg)













