Embed Size (px)
Citation preview

REGULAR ARTICLE
Impacts of litter and understory removal on soil propertiesin a subtropical Acacia mangium plantation in China
Yanmei Xiong & Hanping Xia & Zhi’an Li &Xi’an Cai & Shenglei Fu
Received: 25 July 2007 /Accepted: 20 December 2007 / Published online: 19 January 2008# Springer Science + Business Media B.V. 2007
Abstract In forest ecosystems, the effects of litter orunderstory on soil properties are far from being fullyunderstood. We conducted a study in a pure Acaciamangium Willd. plantation in southern China, byremoving litter or understory or both components andthen comparing these treatments with a control(undisturbed), to evaluate their respective effects onsoil physical, chemical and biological properties. Inaddition, a litter decomposition experiment wasconducted to understand the effects of understory onlitter decomposition. Our data showed that the
presence of understory favored litter decompositionto a large extent. In 1 year, 75.2 and 37.2% of litterwere decomposed in the control and understoryremoval treatment (UR), respectively. Litter had aprofound significance in retaining soil water andcontributing to soil fertility, including organic matter(OM), available phosphorus (P) and alkali-hydrolyzablenitrogen (N), but understory exerted less influence thanlitter on soil physical and chemical properties. Both litterand understory played an important role in soilbiological activity as indicated by microbial biomasscarbon (MBC), while there were no significant impactson soil exchangeable potassium (K) after either or bothwere removed. Contrary to our hypothesis, the effects ofunderstory or litter removal were not always negative. Asignificant soil pH increase with litter removal was apositive factor for acid soil in the studied site. Except forsoil moisture, significant effects, caused by removal oflitter or/and understory, on measured soil chemicalcharacteristics were only observed in the top 10 cm soillayer, but not in the 10–20 cm layer. Soil available P andexchangeable K contents were significantly higher inthe rainy season than in the dry season, however, for theother soil properties, not substantially affected byseason.
Keywords Litter decomposition . Soil fertility .
Soil microbial biomass . Soil pH . Soil water content
Plant Soil (2008) 304:179–188DOI 10.1007/s11104-007-9536-6
Responsible Editor: Barbara Wick.
Y. Xiong :H. Xia (*) : Z. Li :X. Cai : S. FuSouth China Botanical Garden,Chinese Academy of Sciences,Xingke Road #723, Tianhe District,Guangzhou 510650, Chinae-mail: [email protected]
Y. Xiong :H. Xia : Z. Li :X. Cai : S. FuHeshan National Field Research Station of ForestEcosystem,Heshan 529725, China
Y. Xiong :H. Xia : Z. Li :X. Cai : S. FuHeshan Hilly Land Interdisciplinary Experimental Station,Chinese Academy of Sciences,Heshan 529725, China

AbbreviationsUR understory removalLR litter removalUR + LR both understory and litter removalOM organic matterMBC microbial biomass carbon
Introduction
Forests are among the most productive ecosystems innature. In forests, components like shrubs and herbaceousplants of the understory and litter from plants areindispensable to ecosystem stability and ecologicalfunction. The roles that litter plays in ecosystems wererecognized as early as the 1850s and have beendocumented by a large number of studies worldwide(Sayer 2006). Litter contributes to forest ecosystemsmainly by nutrient and carbon turnover during decom-position, thus maintaining biogeochemical cycling in theecosystems. Litter cover acts as a protective layer formaintaining soil physical properties, like retention forsoil moisture (Ginter et al. 1979), buffering against soiltemperature and compaction change (MacKinney 1929),and soil conservation from erosion or leaching (Geddesand Dunkerley 1999; Mo et al. 2003). Litter alsoprovides habitats and substrates for soil fauna (Attignonet al. 2004) and microorganisms (Ruf et al. 2006).
The roles of understory in aboveground ecosystemshave been extensively studied. Due to the sensitivity ofunderstory species to site quality conditions, understoryhas been used as an indicator of landform character-istics, disturbance history, and altered environmentalconditions across the landscape and over time (Meilleuret al. 1992; Kamisako et al. 2007). Attempts have beenmade to understand the effect of understory onestablishment and growth of tree seedlings, whichaffect long term canopy structure and composition(Abe et al. 2001; Goto 2004). Understory functioningas a habitat for animals thus contributing to forestbiodiversity has also been reported (Camprodon andBrotons 2006). However, the impact of understory onsoil ecosystems has been studied less and focusedmainly on soil nitrogen (N) mineralization and Ncycling (Tripathi et al. 2005; Matsushima and Chang2007). Due to high turnover rates, understory vegeta-tion has been suggested to contribute substantially tonutrient cycling and soil fertility (Yarie 1980; ChapinIII 1983; Nilsson and Wardle 2005).
Due to the coexistence of litter and understory inforests, the previous studies have generally failed toseparate their respective contributions to soil ecosys-tems, and the evaluated effect of one componentpotentially included that of the other. Therefore, it isnecessary to develop new approaches to obtain aclearer understanding of the respective roles that thetwo components play in soil ecosystems.
As one of the most important factors changing thestructure and function of ecosystems, human activityhas made severe disturbances to these forest compo-nents. Litterfall and understory vegetation wereintensively removed for fuel in the past and thisactivity even persists at present in some nations,particularly in developing countries (Katagiri et al.1997; Mo et al. 2003; Zhang et al. 2006). In addition,understory clearing is a major activity for forestmanagement in many regions, which aims not onlyto eliminate combustible loads to prevent fire, but alsoto control competition with trees and to favor saplingregeneration (Camprodon and Brotons 2006). Incontrast to the sustained disturbance throughouthuman history, only a small number of studies havebeen conducted to evaluate the consequences of litteror understory removal on forest ecosystems, and theresults obtained are far from sufficient to allow acomplete understanding (Mo et al. 2003; Zhang et al.2006; Matsushima and Chang 2007).
Therefore, we conducted this study in a pureAcacia mangium Willd. plantation in southern Chinaby removing litter or understory or both components,aiming to partition their respective effects on soilproperties. A litter decomposition experiment wasalso conducted to find out the potential effect ofunderstory on litter decomposition. We hypothesizedthat (1) litter and understory had different influenceson soil properties; (2) removal of either componentwould always cause negative effects on soil proper-ties; and (3) there might be a synergistic effect on soilfertility with the coexistence of litter and understory.
Materials and methods
Study site
This study was conducted at the Heshan Hilly LandInterdisciplinary Experimental Station, Chinese Acad-emy of Sciences in Guangdong province, P.R. China,
180 Plant Soil (2008) 304:179–188

located in 22°41′N and 112°54′E. The climate of theregion is subtropical monsoon with a mean annualprecipitation of 1,700 mm, falling mainly in the rainyseason from April to September, with the period fromOctober to March being the dry season. The meanannual temperature is 21.7°C with the mean maxi-mum monthly temperature of 29.2°C falling in Julyand the mean monthly minimum of 12.6°C falling inJanuary. The soil is acrisol (FAO 2006), and theselected soil chemical properties at the initiation ofthe experiment in 2002 are illustrated in Table 1.
The studied stand covered an area of about 3 hasloped at about 30°, in which A. mangium was theonly overstory species planted at a spacing of 3×4 min 1984. In the tropics and subtropics of China, A.mangium is one of the most commonly plantedlegume species, and A. mangium plantations areusually constructed as a means of restoring degradedhilly lands in southern China. The site had beennatural grassland before the A. mangium plantationwas set up, and then has been protected for scientificresearch since 1984. Measured litterfall from theoverstory of the plantation amounts to 10.4 t ha−1 a−1,much more than other plantations in the studied area(Zou et al. 2006). The canopy closure of A. mangiumwas about 60%, and the height and breast diameterwere 18.4 m and 22.1 cm on average, respectively.The understory species of the site were mainlyDicranopteris dichotoma, Miscanthus sinensis,Blechnum orientale, Ottochloa nodosa var. micrantha,Dianella ensifolia, Thysanolaena maxima, Litseacubeba, Euodia lepta, Ilex asprella, Embelia laeta,Mussaenda pubescens and Clerodendron fortunatum,without absolutely dominant species. The understorycoverage was about 75% at the beginning of the
experiment and most of the understory species wereshorter than 1 m in height.
Experimental design
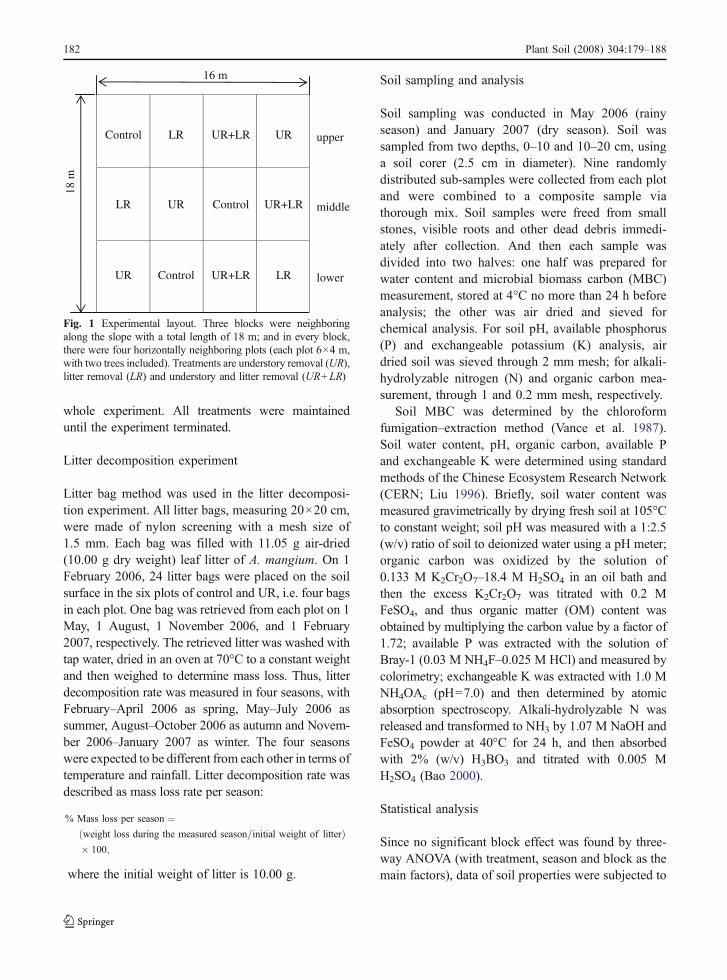
The experiment began from 2002 using a completerandomized block design. There were altogether threeneighboring blocks designed along the slope, present-ing the lower, medial and upper locations of the stand,respectively. Each block was equally divided into fourplots, and each plot had an area of 24 m2 (6×4 m).The control and three treatments were randomlyapplied to the four plots of each block, and thus therewere three replicates for control and each treatment(Fig. 1). The control was left undisturbed with intactunderstory and litter layer, and the three treatmentswere understory removal (UR), litter removal (LR),and both understory and litter removal (UR+LR). Theplot boundaries were enclosed by iron sheets of 25 cmhigh, leaving two drainage outlets at the two lowercorners of each plot for the purpose of rainwaterdrainage. Since the roots of understory species,especially of herbaceous plants, are mainly distributedat the top soil layer and their lateral growth is very littleat a depth of over 10 cm, the iron sheets were insertedonly 10 cm deep into soil and left 15 cm above ground.A net was set 3 m above ground for each LR and UR+LR plot and litter trapped on the net was removed onceevery 2 weeks. The mesh size was 2 cm and, therefore,the net could effectively prevent litter from dropping,but did not affect sunlight and rainfall. All understoryplants in each plot of UR and UR+LR were removedby uprooting at the beginning of the experiment, anduprooting was repeatedly done as long as newunderstory plants sprouted during the period of the
Table 1 Means (standard error, n=3) of soil chemical properties (0–10 cm depth) at the study site at the initiation of the experimentin 2002
Treatment pH (water) Alkali-hydrolyzable N (mg kg−1) OM (g kg−1) Available P (mg kg−1) Exchangeable K (mg kg−1)
Control 4.1 (0.0) 152.1 (6.3) 37.9 (2.0) 3.6 (0.3) 50.7 (4.1)UR 4.1 (0.0) 140.1 (7.5) 35.6 (2.2) 3.4 (0.4) 47.1 (6.5)LR 4.1 (0.0) 141.8 (1.7) 37.1 (1.8) 3.1 (0.5) 43.9 (3.7)UR+LR 4.1 (0.1) 144.1 (1.4) 36.7 (1.2) 3.1 (0.5) 44.7 (8.6)Treatment effect ns ns ns ns nsBlock effect ns ns 0.043 ns ns
Treatment and block effects of the ANOVA (n=12) refer to P<0.05.
ns Not significant
Plant Soil (2008) 304:179–188 181
whole experiment. All treatments were maintaineduntil the experiment terminated.
Litter decomposition experiment
Litter bag method was used in the litter decomposi-tion experiment. All litter bags, measuring 20×20 cm,were made of nylon screening with a mesh size of1.5 mm. Each bag was filled with 11.05 g air-dried(10.00 g dry weight) leaf litter of A. mangium. On 1February 2006, 24 litter bags were placed on the soilsurface in the six plots of control and UR, i.e. four bagsin each plot. One bag was retrieved from each plot on 1May, 1 August, 1 November 2006, and 1 February2007, respectively. The retrieved litter was washed withtap water, dried in an oven at 70°C to a constant weightand then weighed to determine mass loss. Thus, litterdecomposition rate was measured in four seasons, withFebruary–April 2006 as spring, May–July 2006 assummer, August–October 2006 as autumn and Novem-ber 2006–January 2007 as winter. The four seasonswere expected to be different from each other in terms oftemperature and rainfall. Litter decomposition rate wasdescribed as mass loss rate per season:
% Mass loss per season ¼weight loss during the measured season=initial weight of litterð Þ� 100;
where the initial weight of litter is 10.00 g.
Soil sampling and analysis
Soil sampling was conducted in May 2006 (rainyseason) and January 2007 (dry season). Soil wassampled from two depths, 0–10 and 10–20 cm, usinga soil corer (2.5 cm in diameter). Nine randomlydistributed sub-samples were collected from each plotand were combined to a composite sample viathorough mix. Soil samples were freed from smallstones, visible roots and other dead debris immedi-ately after collection. And then each sample wasdivided into two halves: one half was prepared forwater content and microbial biomass carbon (MBC)measurement, stored at 4°C no more than 24 h beforeanalysis; the other was air dried and sieved forchemical analysis. For soil pH, available phosphorus(P) and exchangeable potassium (K) analysis, airdried soil was sieved through 2 mm mesh; for alkali-hydrolyzable nitrogen (N) and organic carbon mea-surement, through 1 and 0.2 mm mesh, respectively.
Soil MBC was determined by the chloroformfumigation–extraction method (Vance et al. 1987).Soil water content, pH, organic carbon, available Pand exchangeable K were determined using standardmethods of the Chinese Ecosystem Research Network(CERN; Liu 1996). Briefly, soil water content wasmeasured gravimetrically by drying fresh soil at 105°Cto constant weight; soil pH was measured with a 1:2.5(w/v) ratio of soil to deionized water using a pH meter;organic carbon was oxidized by the solution of0.133 M K2Cr2O7–18.4 M H2SO4 in an oil bath andthen the excess K2Cr2O7 was titrated with 0.2 MFeSO4, and thus organic matter (OM) content wasobtained by multiplying the carbon value by a factor of1.72; available P was extracted with the solution ofBray-1 (0.03 M NH4F–0.025 M HCl) and measured bycolorimetry; exchangeable K was extracted with 1.0 MNH4OAc (pH=7.0) and then determined by atomicabsorption spectroscopy. Alkali-hydrolyzable N wasreleased and transformed to NH3 by 1.07 M NaOH andFeSO4 powder at 40°C for 24 h, and then absorbedwith 2% (w/v) H3BO3 and titrated with 0.005 MH2SO4 (Bao 2000).
Statistical analysis
Since no significant block effect was found by three-way ANOVA (with treatment, season and block as themain factors), data of soil properties were subjected to
middle
upper
18 m
16 m
Control LR UR+LR UR
LR UR Control UR+LR
UR Control UR+LR LR lower
Fig. 1 Experimental layout. Three blocks were neighboringalong the slope with a total length of 18 m; and in every block,there were four horizontally neighboring plots (each plot 6×4 m,with two trees included). Treatments are understory removal (UR),litter removal (LR) and understory and litter removal (UR+LR)
182 Plant Soil (2008) 304:179–188

two-way ANOVA (treatment and season being themain factors) to assess their respective effects andalso the interaction between them. Levene’s test wasused to test for homogeneity of variance, and data ofsoil moisture in the 10–20 cm soil layer were logtransformed due to heteroscedasticity. Differencesamong treatments and control were compared by theLSD test. Differences of soil properties between the top10 cm and deeper 10–20 cm soil layers and differenceof mass loss rate per season between control and URwere determined by paired sample t test. For statisticalsignificance, P=0.05 was chosen. All analyses wereconducted using the SPSS 13.0 software.
Results
Effects of understory on litter decomposition rate
The decomposition rates of litter were significantlydifferent between control and UR; furthermore theyproduced distinct changes along with seasons (Fig. 2).In spring 12.6 and 15.7% of litter were decomposedin control and UR, respectively, but their decomposi-tion rates showed completely different changes insummer and autumn. Litter in control was decom-posed at an increasing rate during summer andautumn, resulting in 18.9% mass loss in summer,and peaking at 32.6% in autumn; on the contrary, thedecomposition rate in UR decreased during the twoseasons, to 12.5% in summer and to a minimum of2.2% in autumn. In winter, the decomposition rates ofboth treatments became similar, with control falling to
11.1% and UR rising to 6.8%. Cumulative annualdecomposition rates of litter were 75.2 and 37.2% incontrol and UR, respectively. Paired sample t testshowed that control was significantly higher than URwith regard to litter decomposition rate (P<0.05).
Effects of removal on soil physical and chemicalproperties
In the top 10 cm layer, soil pH, moisture content,alkali-hydrolyzable N, OM and available P weresignificantly different among treatments (P<0.05),but not significant with regard to soil exchangeable K(Table 2; Fig. 3). Litter removal significantly influ-enced soil pH (P<0.05), moisture content (P<0.01),alkali-hydrolyzable N (P<0.05), OM (P<0.05) andavailable P (P<0.01), with higher pH values andlower values of the other properties in LR and UR+LRthan that in control and UR (Fig. 3a–e). However,understory removal had no significant effect on thesesoil properties since there was no significant differencebetween control and UR or between LR and UR+LR.
In the 10–20 cm soil layer, there was a significanteffect of different treatments on soil moisture content,with UR moister than control and the other twotreatments (P<0.05; Table 2). But neither understorynor litter removal exerted a significant influence onthe other soil properties in this layer.
Season significantly influenced soil available P inthe top 10 cm layer and exchangeable K in both soillayers, with higher available P and exchangeable Kvalues in the rainy season than in the dry season (P<0.001; Table 2; Fig. 3e and f). The effects of seasonon soil available P were more marked in control andUR than in LR and UR+LR, as indicated by thesignificant interaction between treatment and season(P<0.01; Table 2; Fig. 3e). Neither season norinteraction between treatment and season produced asignificant effect on soil pH, moisture content, alkali-hydrolyzable N and OM in both soil layers (Table 2).
Soil alkali-hydrolyzable N, OM, available P andexchangeable K were much higher in the top 10 cmlayer than in the 10–20 cm layer regardless of season(P<0.001; Table 3). In contrast, soil pH was lower(P<0.001) in the top 10 cm layer compared to thedeeper layer. Soils of control and UR, with littercovering, were moister in the top 10 cm layer, however,for LR and UR+LR, without litter, wetter in the deeperlayer (Table 3).
Season
Spring Summer Autumn Winter
% m
ass
loss
rat
e pe
r se
ason
0
10
20
30
40
50
Control UR
Fig. 2 Percent mass loss rate per season during one year’s litterdecomposition process in control and understory removal (UR).Error bars are standard errors (n=3)
Plant Soil (2008) 304:179–188 183

Table 2 P values of two-way ANOVA analysis for treatment effect, season effect and the interaction between treatment and season(n=24), with respect to soil physical and chemical properties in 0–10 and 10–20 cm soil layer, respectively
Source pH Moisture (%) Alkali-hydrolyzable N(mg kg−1)
OM(g kg−1)
Available P(mg kg−1)
Exchangeable K(mg kg−1)
0–10 cm
10–20 cm
0–10 cm
10–20 cm
0–10 cm
10–20 cm
0–10 cm
10–20 cm
0–10 cm
10–20 cm
0–10 cm
10–20 cm
Treatment <0.05 ns <0.01 <0.05 <0.05 ns <0.05 ns <0.01 ns ns nsSeason ns ns ns ns ns ns ns ns <0.001 ns <0.001 <0.01Treatment ×season
ns ns ns ns ns ns ns ns <0.01 ns ns ns
ns Not significant
pH
3.6
3.8
4.0
4.2
4.4
4.6 Rainy seasonDry season
a
aab
bc c
Control UR LR UR+LRControl UR LR UR+LR
Exc
hang
eabl
e K
(m
g K
kg-
1 )
0
10
20
30
40
50
60
70 f
Ava
ilabl
e P
(m
g P
kg-
1 )
0
1
2
3
4
5
a
a
bb
e
Moi
stur
e co
nten
t (%
)0
5
10
15
20
25
30 Rainy seasonDry season
a
bb
ab
Alk
ali-h
ydro
lyza
ble
N (
mg
N k
g-1 )
60
80
100
120
140
160
180
200 c
a
a
b b
Org
anic
mat
ter
(g O
M k
g-1 )
0
10
20
30
40
50d
a
b b
a
Fig. 3 Soil physical andchemical properties, soilpH (a), moisture content(b), alkali-hydrolyzable N(c), organic matter(d), available P (e), andexchangeable K (f), in thetop 10 cm layer during therainy and dry season. Errorbars are standard errors(n=3). Treatments are un-derstory removal (UR), litterremoval (LR) and understo-ry and litter removal (UR+LR). Treatments superscrip-ted with different letters aresignificantly different atP=0.05 level by LSD test
184 Plant Soil (2008) 304:179–188

Effects of removal on soil microbial biomass
Soil MBC was higher in the top 10 cm soil layer thanin the deeper 10–20 cm layer (P<0.05), with theexception of UR+LR which had the reverse pattern inboth rainy and dry seasons (Fig. 4). ANOVA in thetop 10 cm layer failed because of missing data in tworeplicates of UR+LR in both seasons. In the 10–20 cm layer there was no significant difference amongtreatments or between the two seasons. Soil MBC wasgreatest in control for both soil layers.
Discussion
Function of understory on litter decomposition
Understory was of great importance to litter decom-position in our study. It is well accepted that physicalenvironmental conditions such as moisture andtemperature strongly affect litter decomposition(Meentemeyer 1978; de Santo et al. 1993). In thisarea, the rainy period (summer and autumn) coincideswell with high temperature. With an overstory coverof about 60% in the studied plantation, understoryremoval caused the direct exposure of the litter layer
to sunshine and high temperature, and thus water inthe litter layer might be lost quickly, which made URadverse to decomposers during this period. The decayrate of litter seems more closely related to the watercontent of litter layer than to that of soil (de Santoet al. 1993). Therefore, although soil water content inUR was not distinctly different from that in control(Fig. 3b), it was probably the relatively low watercontent in the litter layer induced by understoryremoval that caused litter in UR plots to decomposeat a declining rate in summer and autumn (Fig. 2). Incontrast, the rate of litter mass loss in controlincreased continuously during the period (Fig. 2). Itcould be attributed to the shading effect of understory,which protected the litter layer from water loss,creating a favorable microclimate for decomposersthat accelerated litter decomposition in the rainyperiod.
Effects of litter removal on soil physical and chemicalproperties
Compared with the contribution of litter to soilfertility and nutrient cycling, its function in retainingsoil water content has been paid less attention (Ginteret al. 1979). In the present study, litter functioned as
Table 3 Percent differences between means of top (0–10 cm) and deeper (10–20 cm) layers with respect to soil properties in rainy anddry season, respectively, with analysis of variance (P values) by top–deeper paired sample t test (n=12)
Treatment pH (%) Moisture (%) Alkali-hydrolyzable N (%) OM (%) Available P (%) Exchangeable K (%)
Rainy Dry Rainy Dry Rainy Dry Rainy Dry Rainy Dry Rainy Dry
Control −6.7 −7.8 7.9 5.5 36.6 45.6 41.5 42.7 65.1 37.0 36.5 16.0UR −4.9 −6.9 8.8 4.1 50.0 45.8 43.8 30.7 56.6 27.3 28.3 17.2LR −5.8 −4.9 −10.0 −5.5 33.2 30.8 30.9 14.7 26.0 27.3 34.0 26.9UR+LR −6.6 −4.9 −13.0 −15.1 31.9 38.3 29.3 30.8 31.1 32.4 26.7 20.2P values <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.001 <0.001 <0.001 <0.001
% difference=100×(meantop−meandeeper)/mean top; positive figures indicate reduction of values from the top layer to deeper layerand negative figures represent the reverse pattern.
Rainy season
Control UR LR UR+LR Control UR LR UR+LR
Mic
robi
al b
iom
ass
C (
mg
C k
g-1)
0
100
200
300
400
Dry season
Mic
robi
al b
iom
ass
C (
mg
C k
g-1)
0
100
200
300
400
0–10 cm10–20 cm
0–10 cm10–20 cm
Fig. 4 Soil microbialbiomass carbon (MBC) intwo soil layers for the rainyand dry season. Error barsare standard errors (n=3).Treatments are: understoryremoval (UR), litter removal(LR) and understory andlitter removal (UR+LR)
Plant Soil (2008) 304:179–188 185

the major determinant for soil water content, and litterremoval significantly decreased soil moisture(Fig. 3b). It is consistent with Eckstein and Donath(2005), which showed the positive relation betweensoil water content and litter amount. In fact, mulchingor covering the soil surface with a layer of plantresidues has been found an effective method ofconserving soil water because it reduces surfacerunoff and evaporation, and increases infiltration ofwater into the soil (Ghawi and Battikhi 1986;Adekalu et al. 2007).
The effect of plant litter or residues on soil pH hasbeen extensively studied, but some results have beenconflicting. Acid neutralization effects were founddue to the production of ammonium during plantmaterial decomposition (Hoyt and Turner 1975), ordue to an excess of cations over inorganic anions inplant materials (Noble et al. 1996; Noble andRandall1999). On the contrary, plant materials havealso been suggested as one cause of soil acidificationby nitrification and the subsequent loss of NO�
3 or byan increased cation exchange capacity and thecorresponding increase in exchangeable acidity(Bolan et al. 1991; Tang and Yu 1999). As a result,the direction and magnitude of pH change dependlargely on the balance between nitrification andammonification. It was found by Li et al. (2001) thatin an A. mangium plantation nitrification dominatedover ammonification which might be due to thedecomposition of A. mangium litter. Therefore, thesignificant elevation of soil pH due to litter removal(Fig. 3a) might be the result of mitigated nitrificationby litter removal.
It has been well documented that nutrient return bylitter decomposition is a very important process forsoil fertility and nutrient cycling in ecosystems(Didham 1998; Wardle et al. 2003). In the presentstudy, the significant decline of soil alkali-hydrolyzableN, OM and available P by litter removal has furtherproved the contribution of litter to soil fertility andnutrient cycling (Fig. 3c–e). To our surprise, litterremoval exerted no significant influence on soilexchangeable K (Fig. 3f). It has been reported thatacidification usually leads to depletion of base cations(e.g. K+, Mg2+, Ca2+) because of the exchange betweenH+ and them (Bailey et al. 2005). Considering that soilin the studied site was strong acidic (Table 1), thesignificant soil pH elevation by litter removal (Fig. 3a)might mitigate this depletion to some extent. As a
result, despite the reduced K return caused by litterremoval, the mitigated K depletion might compensatefor this loss.
The significant decreases of soil available P andexchangeable K from the rainy season to dry seasonmight be due to different mechanisms (P<0.001;Table 2; Fig. 3e and f). Rewetting after drying seasonhas been reported to induce a large pulse of Pmineralization (Lodge et al. 1994). In the tropicalmonsoon forests of India, the flux of P mineralizedfrom microbial biomass induced by rewetting in thefirst 4 weeks of the rainy season was greater thanthe total flux of P mineralized from litterfall over theentire rainy season (Singh et al. 1989). Since our firstsampling was in May 2006, the onset of the rainyseason, the pulse of soil available P in the rainyseason was reasonable. However, the strong Kleaching during the peak rainy season might beresponsible for the lower values of soil exchangeableK in the dry season, since K is susceptible toleaching. During the dry season, with reduced leach-ing and plant uptake, mechanisms like capillary risecan lead to K accumulation, which contributes to theseasonal cycle of soil K (Zwikel et al. 2007).
Effects of understory removal on soil physicaland chemical properties
Since the presence of plants and their influence onsoil conditions may depend on original soil propertiesat both microsite and landscape scales (Rhoades1997), the evaluation of original soil spatial variabil-ity is necessary to determine understory effects on soil(Wick et al. 2000). In the present study, the measuredsoil properties at the initiation of experiment were notsignificantly different among treatments and, there-fore, the original soil properties of all studied plotscould be considered similar (Table 1). The effect ofunderstory on measured soil properties was thusseparated from the effect of variation of original soilproperties.
Understory could contribute to soil fertility byaccelerating the litter decomposition rate as shown inthe litter decomposition experiment (Fig. 2). Howev-er, understory removal had no significant effect onmost of the physical and chemical properties studied(Fig. 3), which is inconsistent with the conclusion insome studies that understory plays an important rolein nutrient and carbon cycling (Yarie 1980; Nilsson
186 Plant Soil (2008) 304:179–188

and Wardle 2005). The substantial uptake of soilnutrients and water by understory could be responsiblefor our result. Nutrient uptake by undergrowth issubstantial and even induces soil fertility depletionand inhibits tree growth in some cases (Nambiar andSands 1993).
Effects of litter and understory on soil biologicalproperties
As the primary decomposer, microorganisms play afundamental role in carbon and nutrient cycling andthe formation of soil structure and, therefore, a highmicroorganism biomass is necessary for the mainte-nance of soil quality (Berg et al. 2001). Based on theimportance for utilization by soil microbes, there arethree main carbon sources in forest ecosystems: plantroot exudates, litter, and soil organic matter (Ruf et al.2006). As a result, either understory or litter is ofparamount importance to maintain a large amountof soil microbe biomass. Due to the greater effect ofunderstory roots and litter on top soil layer than ondeeper layer, the significant decrease of MBC fromthe 0–10 cm layer to 10–20 cm layer was found inUR, LR and control, which were covered with litter,understory and both of them, respectively (Fig. 4). Asfor UR+LR, since the more favorable conditions forsoil microbes in the top soil layer were removed bythe removal of both understory and litter, soil microbepopulation was limited by other adverse conditions,and soil MBC might be lower in the top layeraccordingly (Fig. 4).
Conclusions
As was expected, an effect of understory on litter wasfound. Presence of the understory favored litterdecomposition to a large extent, thereby increasingthe litter contribution to soil fertility and nutrientcycling. And the enhanced litter decomposition, inturn, implied a contribution of the understory to soilfertility. This interaction of litter and understoryshould be considered a synergistic effect in compar-ison with their sole presence.
As for their respective roles in the soil ecosystem,litter had the most profound significance in retainingsoil moisture and nutrients, since litter removalcaused a significant decrease of soil moisture content
and fertility, particularly to soil OM, alkali-hydrolyzableN and available P. However, A. mangium litter is likelyto enhance soil acidification, because the soil in thestudied site was acid with a pH range of 4 to 4.2(Table 1). One implication of this result is that a mixedplantation of A. mangium and other species capable ofacid neutralization is a potential solution for thisproblem. Apart from the litter-reliant contribution ofunderstory to soil fertility as described above, theunderstory exerted less influence on soil physical andchemical properties studied. However, both litter andunderstory were important in sustaining soil micro-organisms as the most important carbon sources forsoil microbes.
Contrary to our hypothesis, effects caused by litteror understory removal were not necessarily negative. Inview of the soil acidification common in the studiedarea, the significant soil pH increase with litter removalis doubtlessly positive. And the lightened soil acidifi-cation is expected to mitigate depletion of soil basecations, thereby favoring their retention in soil.
Acknowledgements The project was funded by the NaturalScience Foundation of China (30630015) and the KnowledgeInnovation Program of the Chinese Academy of Sciences(KZCX2-YW-413). We thank Prof. Murray B. McBride andDr. Samran Sombatpanit for correction and comments on themanuscript. Special thanks to two anonymous reviewers forvaluable comments that significantly improved the manuscript.We are grateful to Messrs Yongbiao Lin, Bi Zou, and XingquanRao for their technical helps.
References
Abe M, Miguchi H, Nakashizuka T (2001) An interactive effectof simultaneous death of dwarf bamboo, canopy gap, andpredatory rodents on beech regeneration. Oecologia127:281–286
Adekalu KO, Olorunfemi IA, Osunbitan JA (2007) Grass mulch-ing effect on infiltration, surface runoff and soil loss of threeagricultural soils in Nigeria. Bioresour Technol 98:912–917
Attignon SE, Weibel D, Lachat T, Sinsin B, Nagel P, PevelingR (2004) Leaf litter breakdown in natural and plantationforests of the Lama forest reserve in Benin. Appl Soil Ecol27:109–124
Bailey SW, Horsley SB, Long RP (2005) Thirty years of changein forest soils of the Allegheny Plateau, Pennsylvania. SoilSci Soc Am J 69:681–690
Bao S (2000) Agricultural and chemical analysis of soil. ChinaAgricultural, Beijing, pp 56–58 (in Chinese)
Berg M, de Ruiter P, Didden W, Janssen M, Schouten T,Verhoef H (2001) Community food web, decompositionand nitrogen mineralisation in a stratified Scots pine forestsoil. Oikos 94:130–142
Plant Soil (2008) 304:179–188 187

Bolan NS, Hedley MJ, White RE (1991) Processes of soilacidification during nitrogen cycling with emphasis onlegume based pastures. Plant Soil 134:53–63
Camprodon J, Brotons L (2006) Effects of undergrowthclearing on the bird communities of the NorthwesternMediterranean Coppice Holm oak forests. For EcolManage 221:72–82
Chapin FS III (1983) Nitrogen and phosphorus nutrition andnutrient cycling by evergreen and deciduous understoryshrubs in an Alaskan black spruce forest. Can J For Res13:773–781
de Santo AV, Berg B, Rutigliano FA, Alfani A, Fioretto A(1993) Factors regulating early-stage decomposition ofneedle litters in five different coniferous forests. Soil BiolBiochem 25:1423–1433
Didham RK (1998) Altered leaf-litter decomposition rates intropical forest fragments. Oecologia 116:397–406
Eckstein RL, Donath TW (2005) Interactions between litter andwater availability affect seedling emergence in fourfamilial pairs of floodplain species. J Ecol 93:807–816
FAO (2006) World reference base for soil resources 2006.World Soil Resources Report 103. FAO, Rome
Geddes N, Dunkerley D (1999) The influence of organic litteron the erosive effects of raindrops and of gravity dropsreleased from desert shrubs. Catena 36:303–313
Ghawi I, Battikhi A (1986) Water melon production undermulch and trickle irrigation in the Jordan valley. J AgronCrop Sci 157:145–155
Ginter DL,Mcleod KW, Sherrod C (1979) Water stress in longleafpine induced by litter removal. For Ecol Manage 2:13–20
Goto Y (2004) Early post-fire vegetation regeneration in Larixkaempferi artificial forests with an undergrowth of Sasasenanensis. Ecol Res 19:311–321
Hoyt PB, Turner RC (1975) Effects of organic materials addedto very acid soils on pH, aluminum, exchangeable NH4,and crop yields. Soil Sci 119:227–237
Kamisako M, Sannoh K, Kamitani T (2007) Does understoryvegetation reflect the history of fluvial disturbance in ariparian forest? Ecol Res 22:67–74
Katagiri S, Li C, Nagayama Y, Iwatsubo G (1997) Influences ofhuman impact on nutrient cycling in deteriorated pineforest in southern China. In: Iwatsubo G, Li C (eds)Ecological and hydrological study on a forested watershedin southern China. Kyoto University, Kyoto, pp 165–178
Li Z, Peng S, Rae DJ, Zhou G (2001) Litter decomposition andnitrogenmineralization of soils in subtropical plantation forestsof southern China, with special attention to comparisonsbetween legumes and non-legumes. Plant Soil 229:105–116
Liu G (1996) Analysis of soil physical and chemical propertiesand description of soil profiles. China Standard, Beijing,pp 121–265 (in Chinese)
Lodge DJ, McDowell WH, McSwiney CP (1994) The impor-tance of nutrient pulses in tropical forests. Tree 9:384–387
MacKinney AL (1929) Effects of forest litter on soil temperatureand soil freezing in autumn and winter. Ecology 10:312–321
Matsushima M, Chang SX (2007) Effects of understoryremoval, N fertilization, and litter layer removal on soilN cycling in a 13-year-old white spruce plantation infestedwith Canada bluejoint grass. Plant Soil 292:243–258
Meentemeyer V (1978) Macroclimate and lignin control oflitter decomposition rates. Ecology 59:465–472
Meilleur A, Bouchard A, Bergeron Y (1992) The use ofunderstory species as indicators of landform ecosystemtype in heavily disturbed forest: an evaluation in the Haut-Saint-Laurent, Quebec. Vegetation 102:13–32
Mo J, Brown S, Peng S, Kong G (2003) Nitrogen availability indisturbed, rehabilitated and mature forests of tropicalChina. For Ecol Manage 175:573–583
Nambiar EKS, Sands R (1993) Competition for water andnutrients in forests. Can J For Res 23:1955–1968
Nilsson MC, Wardle DA (2005) Understory vegetation as aforest ecosystem driver: evidence from the northernSwedish boreal forest. Front Ecol Environ 3:421–428
Noble AD, Randall PJ (1999) Alkalinity effects of different treelitters incubated in an acid soil of N.S.W., Australia.Agrofor Syst 46:147–160
Noble AD, Zenneck I, Randall PJ (1996) Leaf litter ashalkalinity and neutralisation of soil acidity. Plant Soil179:293–302
Rhoades CC (1997) Single-tree influences on soil properties inagroforestry: lessons from natural forest and savannaecosystems. Agrofor Syst 35:71–94
Ruf A, Kuzyakov Y, Lopatovskaya O (2006) Carbon fluxes insoil food webs of increasing complexity revealed by 14Clabelling and 13C natural abundance. Soil Biol Biochem38:2390–2400
Sayer EJ (2006) Using experimental manipulation to assess theroles of leaf litter in the functioning of forest ecosystems.Biol Rev 81:1–31
Singh JS, Raghubanshi AS, Singh RS, Srivastava SC (1989)Microbial biomass acts as a source of plant nutrients in drytropical forest and savanna. Nature 338:499–500
Tang C, Yu Q (1999) Impact of chemical composition oflegume residues and initial soil pH on pH change of a soilafter residue incorporation. Plant Soil 215:29–38
Tripathi SK, Sumida A, Shibata H, Uemura S, Ono K, Hara T(2005) Growth and substrate quality of fine root and soilnitrogen availability in a young Betula ermanii forest ofnorthern Japan: effects of the removal of understory dwarfbamboo (Sasa kurilensis). For Ecol Manage 212:278–290
Vance ED, Brookes AC, Jenkinson DS (1987) An extractionmethod for measuring soil microbial biomass C. Soil BiolBiochem 19:703–707
Wardle DA, Nilsson MC, Zackrisson O, Gallet C (2003)Determinants of litter mixing effects in a Swedish borealforest. Soil Biol Biochem 35:827–835
Wick B, Tiessen H, Menezes RSC (2000) Land quality changesfollowing the conversion of the natural vegetation intosilvo-pastoral systems in semi-arid NE Brazil. Plant Soil222:59–70
Yarie J (1980) The role of understory vegetation in the nutrientcycle of forested ecosystems in the mountain hemlockbiogeoclimatic zone. Ecology 61:1498–1514
Zhang G, Jiang H, Niu G, Liu X, Peng S (2006) Simulating thedynamics of carbon and nitrogen in litter-removed pineforest. Ecol Modell 195:363–376
Zou B, Li Z, Ding Y, Tan W (2006) Litterfall of commonplantations in south subtropical China. Acta Ecol Sin26:715–721 (in Chinese)
Zwikel S, Lavee H, Sarah P (2007) Temporal evolution of saltsin Mediterranean soils transect under different climaticconditions. Catena 70:282–295
188 Plant Soil (2008) 304:179–188





























