Embed Size (px)
Citation preview

Cancer Therapeutics Insights
Impact of Tumor HER2/ERBB2 Expression Level on HER2-Targeted Liposomal Doxorubicin-Mediated Drug Delivery:Multiple Low-Affinity Interactions Lead to a Threshold Effect
Bart S. Hendriks, Stephan G. Klinz, Joseph G. Reynolds, Christopher W. Espelin, Daniel F. Gaddy, andThomas J. Wickham
AbstractNumerous targeted nanotherapeutics have been described for potential treatment of solid tumors.
Although attention has focused on antigen selection and molecular design of these systems, there has been
comparatively little study of how cellular heterogeneity influences interaction of targeted nanoparticles
with tumor cells. Antigens, such as HER2/ERBB2, are heterogeneously expressed across different
indications, across patients, and within individual tumors. Furthermore, antigen expression in nontarget
tissues necessitates optimization of the therapeutic window. Understanding the performance of a given
nanoparticle under different regimens of antigen expression has the ability to inform patient selection and
clinical development decisions. In this work, HER2-targeted liposomal doxorubicin was used as a model-
targeted nanoparticle to quantitatively investigate the effect of HER2 expression levels on delivery of
doxorubicin to the nucleus. We find quantitatively greater nuclear doxorubicin delivery with increasing
HER2 expression, exhibiting a threshold effect at approximately 2 � 105 HER2 receptors/cell. Kinetic
modeling indicated that the threshold effect arises from multiple low-affinity interactions between the
targeted liposome and HER2. These results support previous data showing little or no uptake into human
cardiomyocytes, which express levels of HER2 below the threshold. Finally, these results suggest that
HER2-targeted liposomal doxorubicin may effectively target tumors that fall below traditional definitions
of HER2-positive tumors, thereby expanding the potential population of patients that might benefit from
this agent. Mol Cancer Ther; 12(9); 1816–28. �2013 AACR.
IntroductionNumerous targeted nanoparticles against different
antigens have beendescribed in the literature for potentialtreatment of solid tumors (1–12). The majority of thosestudies focused on the selection and design of nanopar-ticles and/or targeting moieties. In contrast, the relation-ship between antigen expression level and the perfor-mance characteristics of targeted nanoparticles has notbeen investigated. Antigen expression levels vary greatlyacross patient tumors and often have heterogeneousexpression within tumors. Antigen expression level willaffect the ability of targeted nanoparticles to maximizetheir activity, with implications for indication and patientselection.
HER2/ERBB2, a member of the EGF receptor (EGFR)family of growth factor receptors, is a popular antigen formolecularly targeted therapies. It is highly overexpressedin about 20% of human breast cancers (13) and to a lesserdegree inmany other cancer types including gastric, lung,bladder, and endometrial (14). HER2 expression levelsvary across indications, across patients within an indica-tion, andwithin individual tumors themselves. Clinically,HER2 protein expression is assessed using HercepTest(Dako), or Pathway (Ventana Medical Systems), semi-quantitative immunohistochemical (IHC) assays thatscore samples as 0, 1, 2, or 3þ, with 3þ reflecting thehighest intensity and uniformity of staining. When exam-inedusingmore quantitativemethods, studies in cell lineshave mapped 0, 1þ, 2þ, and 3þ to approximate cellsurface receptors numbers of 2 � 104, 1 � 105, 5 � 105
andwell over 106 per cell (15). The fraction of cellswithin atumor considered to be HER2-positive can also vary fromnearly 0% to 100% depending on the patient sample andtumor type (16, 17).
The clinical development of HER2-targeted agentsrelies on the ability to understand the relationshipbetween HER2 expression level and activity of that par-ticular agent. Trastuzumab, a monoclonal antibody ther-apeutic targetingHER2, is approved inHER2 "3þ" tumors
Authors' Affiliation: Merrimack Pharmaceuticals, Cambridge, Massa-chusetts
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Corresponding Author: Bart Hendriks, Merrimack Pharmaceuticals, 1Kendall Square, Suite B7201, Cambridge, MA 02139. Phone: 617-441-7485; Fax: 617-491-1386; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-13-0180
�2013 American Association for Cancer Research.
MolecularCancer
Therapeutics
Mol Cancer Ther; 12(9) September 20131816
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

and a portion of "2þ" tumors that also show gene ampli-fication, as defined by IHC and FISH testing. There arealso recent efforts toward studying the performance oftrastuzumab in HER2-low breast cancer tumors that arenot gene amplified (18, 19). Gastric cancer, anotherapproved indication for trastuzumab, is known to haveparticularly heterogeneous expression of HER2 (16). Ingastric cancer, a distinct HER2-scoring criteria was creat-ed, which specifically lowered the fraction of cells thatneed to exhibit intense HER2 staining to be eligible forHER2-targeted therapies (20). Collectively, this under-scores the importance of understanding HER2 levels andheterogeneity in defining patient populations for HER2-directed therapies. A quantitative understanding of therelationship between antigen expression and activity ishighly desirable for optimizing the clinical developmentof molecularly targeted agents.One HER2-targeted agent currently in clinical devel-
opment is HER2-targeted PEGylated liposomal doxoru-bicin (HER2-tPLD; ref. 21). HER2-tPLD is designed todeliver doxorubicin directly toHER2-overexpressing can-cers and avoid uptake into nontarget tissues, such as theheart, which express low levels of HER2. HER2-tPLDcontains approximately 45 copies of mammalian-derivedF5-scFv (anti-HER2) per liposome (22). The F5-scFv wasselected for its ability to internalize while not inhibitingHER2 signaling (23). Doxorubicin, the active chemother-apeutic agent, is a backbone of cancer therapy in manyindications. It is known to work through multiplemechanisms, but the most direct is via interaction withtopoisomerase 2-a (topo2a). Doxorubicin intercalates inDNA and stabilizes the complex with topo2a, resulting indouble-stranded DNA breaks, preventing DNA replica-tion and ultimately leading to cell death (24).HER2-tPLD is believed to deliver its cytotoxic payload
via a two-step mechanism. First, it deposits in tumorsthrough the enhanced permeability and retention effect(EPR) similar to PLD (25–27). It has been shown for asimilar HER2-tPLD formulation that targeting does notaffect the EPR effect or the extent of total liposome depo-sition within a tumor (26). Second, targeting of HER2 re-sults in preferential accumulation of doxorubicin withinHER2-positive tumor cells and reduced nonspecific up-take by macrophages and other phagocytic cells (26). Pre-vious work has also shown that HER2-tPLD has little or nouptake into human cardiomyocytes, results in little or noevidence of cardiomyocyte cell death or dysfunction, andretains the low penetration into heart tissue of PLD (22)We and others have shown that targeting HER2-over-
expressing cellswithHER2-tPLD results in superior activ-ity relative to PLD in preclinical models expressing highlevels of HER2 (22, 28, 29). Correspondingly, the absenceof a targeting effect has been shown in HER2-negativemodels where activity was similar to PLD (29). However,the mechanism of action of HER2-tPLD has not beenquantitatively studied across the intermediate ranges ofHER2 expression. To understand the cellular character-istics of patient tumors that might benefit, and better
understand interactionswithnontarget tissues,we soughtto understand the quantitative relationship betweenHER2 expression level, cellular uptake, and nuclear deliv-ery of doxorubicin.
Materials and MethodsLiposome preparation
Liposomes were prepared and loaded with doxorubi-cin as previously described (26, 29). Briefly, the lipidcomponents were hydrogenated soy phosphatidyl-choline (HSPC), cholesterol, and PEG-DSPE (3:2:0.3,mol:mol:mol). The anti-ErbB2 (F5)-PEG-DSPE conju-gate was prepared and inserted into the liposome asdescribed previously (30, 31) to create HER2-tPLD. Lipo-somes were also prepared to contain a far red-fluorescentcarbocyanine tracer DiIC18(5)-DS (D12730—abbreviatedDiI5; Life Technologies), which intercalates into the lipidbilayer of the liposome during the extrusion process. TheDiI5-labeled liposomes, HER2-tPLD-DiI5 and PLD-DiI5,were prepared as above with DiI5 dye solubilized withthe lipid components at 0.3 mol% of total phospholipid.Free doxorubicin was removed using a Sephadex G-75size exclusion column eluted with HEPES-bufferedsaline (pH 6.5). The corresponding liposomes withoutdoxorubicin were prepared as well.
Cell cultureAdRr, AU565, Calu-3, HCC202, HCC1569, HCC1954,
HeLa, IGROV1, JIMT-1, MCF7,MDA-MB-361,MDA-MB-453, MKN-7, MKN-45, NCI-N87, OE19, SkBr3, SKOV3,U251, UACC812, ZR75-1, ZR75-30, and 4T1 cells wereobtained from American Type Culture Collection.OVCAR8 cells were obtained from the National CancerInstitute (NCI) cell repository. Cellswere cultured accord-ing to manufacturer’s recommendations. BT474-M3 cellswere a gift fromHermes Biosciences.MCF-7 clone 18 cellswere a gift from Dr. Christopher Benz (Buck Institute,Novato, CA). AdRr-Her2 and OVCAR8-Her2 are stableHER2-overexpressing cell lines derived from AdRr andOVCAR8 cells and were generated in-house. No furthercell line authentication was carried out.
Generation of HER2 clonesHER2-expressing clones of 4T1 and HeLa cells were
generated as follows: 4T1 andHeLa cells were transfectedwith a neomycin-selectable expression vector encodinghuman HER2 (Z2866; GeneCopeia) using transfectionreagent MegaTran 1.0 (OriGene) according to the manu-facturer’s instructions. Transfected cells were selected inmedia containing 400 to 500 mg/mL Geneticin/neomycin(Life Technologies) to eliminate nontransfected cells. Sur-viving cells were expanded under reduced Geneticinconcentrations and sorted via flow cytometry to obtainenriched cell populations with HER2 expression exceed-ing those observed in parental cell lines. The sort-enrichedcells were than subcloned by limited dilutions and colo-nies ranked by HER2 surface levels to obtain representa-tive populations that express different ranges of HER2.
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1817
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

Cell line HER2 quantificationCells were characterized for human HER2 expression
by quantitative fluorescence-activated cell sorting(qFACS) using an anti-HER2 antibody (#340552; BD Bios-ciences). Fluorescent intensity ofHER2 surface stainingbyflow cytometry was compared with staining with thesame antibody ofQuantumSimplyCellularmicrospheres(#815; Bangs Laboratories) to calculate the number ofHER2 surface receptors per cell according to the manu-facturer’s instructions.
Total cell doxorubicin quantification by HPLCEach cell line was plated at 25,000 cells per well and
incubated at 37�C for 24 hours. Cells were incubated with15 mg/mL of HER2-tPLD or PLD for 2 hours. The cellswere washed two times with PBS, solubilized in 1.0%acetic acid inmethanol at�80�C for 1 hour, centrifuged toremove debris and total doxorubicin quantified usinghigh-performance liquid chromatography (HPLC), asdescribed previously (22).
Total cell doxorubicin measurement by flowcytometry
Adherent cells were incubated with 5 mg/mL of HER2-tPLD for 3 hours followed by washing and incubation inculture media for 21 hours to allow internalization andunpacking of doxorubicin. Cells were then harvested andtotal uptake was assessed using flow cytometry by mea-suring the inherent fluorescence of doxorubicin. All flowcytometry data analysis was done with FlowJo v7.2.5(Tree Star, Inc.).
Quantification of nuclear doxorubicin by high-content analysis
Cells were fixed using 3.7% formaldehyde and washedwith PBS-T. Cells were stained with a 1:10,000 dilution ofHoechst 33342 and 1:1,000 dilution of Whole Cell Dye(Pierce) for 30 minutes at room temperature to allowvisualization of DNA and the whole cell, respectively.Plates were scanned using the Applied Precision Instru-ments ArrayWorx High Content Scanner with a 10�objective for Hoechst 33342/whole cell stain (460 nm),doxorubicin (595 nm), and APC/DiI5 (657 nm). Imageswere analyzed using the software ImageRail (32). Anintensity thresholdwas established for nuclear andwholecell signals. This threshold was applied to all images andused to segment individual cells.Data arepresented as themean pixel intensity for all cells in a given well for theindicated channel.
Liposome internalizationCell lines were grown and trypsinized using
versene:0.05% trypsin at a 1:1 ratio. Cells were pelletedand resuspended in10mL ice-coldmedium.Onemilliliterwas set aside as an untreated sample. DiI5-labeled HER2-tPLD (0.1 mg/mL) was loaded on cells on ice for 50 to 55minutes with mixing. Cells were pelleted and washedwith 1� 10 mL ice-cold PBS with 1% FBS (FACS buffer)
and resuspended in 900 mL ice-cold medium. A 100 mLsamplewas taken as the t¼ 0 time point. Eightmilliliter ofprewarmed RPMI–10%FBS culture media was added toeach tube and incubated at 37�C for up to 2 hours. At eachtime point, 1 mL of cell suspension was removed andadded to 9 mL ice-cold FACS buffer on ice.
Cells were pelleted and fixed with 100 mL cytofix solu-tion (#554655; Becton Dickinson) at 4�C. All samples werestainedwith an anti-PEG antibody (PEG-B-47 rabbit mAb2061-1; Epitomics) at 1:400 dilution for 30 minutes at 4�C.A goat-anti-rabbit Fab secondary detection reagent con-jugated to Alexa Fluor 488 (#4412; Cell Signaling Tech-nology) was used at 1:400 dilution for 30 minutes at 4�C.Cells were washed 3� with 200 mL of FACS bufferbetween incubations. Sampleswere resuspended in FACSbuffer and data were acquired via flow cytometry.
Internalization rates were estimated by assuming first-order internalization rates and fitting data with a singleexponential.
Kinetic computational modelThe model is based on mass-action kinetics, except
where indicated, and is composed of three components:a HER2 receptor trafficking model, a doxorubicin cellulartransport model, and amultivalent HER2-bindingmodel.All parameter values and their sources are listed in Sup-plementary Table S1.
HER2 trafficking. HER2 trafficking is modeled on thebasis of previous work (33). HER2 is synthesized at con-stant rate ksyn and inserted into the cell surfacemembrane.A fraction of free surface HER2 are internalized anddegraded at a net rate kdeg (the remaining fraction isinternalized and recycled; not explicitly represented).
Doxorubicin cellular transport. Doxorubicin cellulartransport is modeled on the basis of published work (34,35). Free doxorubicin partitions into the outer leaflet of theplasma membrane, characterized by association rate con-stant kf_ol, cell surface area (based on cell radius, assumingspherical cells), and a partition coefficient. Transport fromthe outer to inner leaflet of the membrane, and back, ismediated by flippases characterized by rate constantskflippase_in and kflippase_out, respectively. Dissociation fromthe membrane into the cytosol is similarly characterizedby rate constant kf_ol. In the cytosol, doxorubicin revers-ibly binds to DNA, with association rate kf_DNA anddissociation constant KD_DNA.
Multivalent HER2 binding. The binding of HER2-tPLD is modeled as a semirigid multivalent particle inter-acting with a surface. HER2-tPLD reacts with a singleHER2 receptor (the reversible interaction of a single scFvwith a single HER2), with association rate kf_LR anddissociation constant KD_LR. The singly bound HER2-tPLD may then cross-link additional receptors. Initialbinding enhances the diffusive rate of encounter withadditional HER2 by factor a. Each individual receptor–scFv interaction is characterized by dissociation constantKD_LR (28). Because of the large size of liposomes (100 nmin diameter), total binding is limited by physical space on
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1818
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

the cell surface. The total liposome projected area iscalculated and it is used to drive the association rate tozero as the surface becomes fully occupied: kf¼ kf,0� [1�(total liposome projected area)/(cell surface area)], wherekf,0 is the association rate in the absence of any stericlimitations (36). It is also assumed that liposomes cannotfreely access the portion of the cell surface that is adherentto the tissue culture plastic—roughly 50%.The (HER2-tPLD):HER2n (where nmay represent 1 or
more HER2 receptors) complex is subsequently inter-nalized via receptor-mediated endocytosis at rate ke.Because HER2-tPLD internalization rates (see Fig. 3)were similar to that of HER2 (33, 37, 38), it was assumedthat binding to and cross-linking of HER2 had no effecton internalization rate. Once inside the cell, doxorubicinis released from HER2-tPLD with first-order kinetics,characterized by rate krel_cell and stoichiometry of 20,000doxorubicin molecules per liposome. In the externalmedium, doxorubicin release occurs much more slowly,at rate krel_medium. Once released, the doxorubicinbehaves as described in the doxorubicin cellular trans-port model.Internalized HER2 that are bound by HER2-tPLD are
either degraded at rate kdeg_HER2_lipo or recycled back tothe surface at rate krecycle_HER2. Internalized liposomes areexocytosed from the cell at rate kdeg_lipo. Nonspecificuptake of liposomes is also represented in the model(33) as a convective transport process, as in fluid-phaseuptake, characterized by rate kfluidphase.Variation of scFv per liposome is simulated by creating
multiple models with different maximum numbers ofcross-linking events. The number of cross-linking eventscan be related to the interaction distance between a lipo-some and the cell surface, assuming uniform distributionof anti-HER2–binding moieties. This determines the frac-tion of the liposome surface that can engage in bindingevents and can be related to the total number of scFv perliposome.Implementation. The model was implemented in
MATLAB SimBiology R2011b (The Mathworks). Thecomplete model description is available in Supplementa-ry Materials both in a tabular descriptive. xls format aswell as an SBML model file.
ResultsBinding and uptakeTo define the levels of HER2 necessary for uptake of
HER2-tPLD into cells, HeLa and 4T1 (a murine cell line)were stably transfected to express human HER2.Although the parental 4T1 cell line expresses murineHER2, HER2-tPLD does not bind the murine receptor.Parental HeLa cells expressed approximately 2 � 105
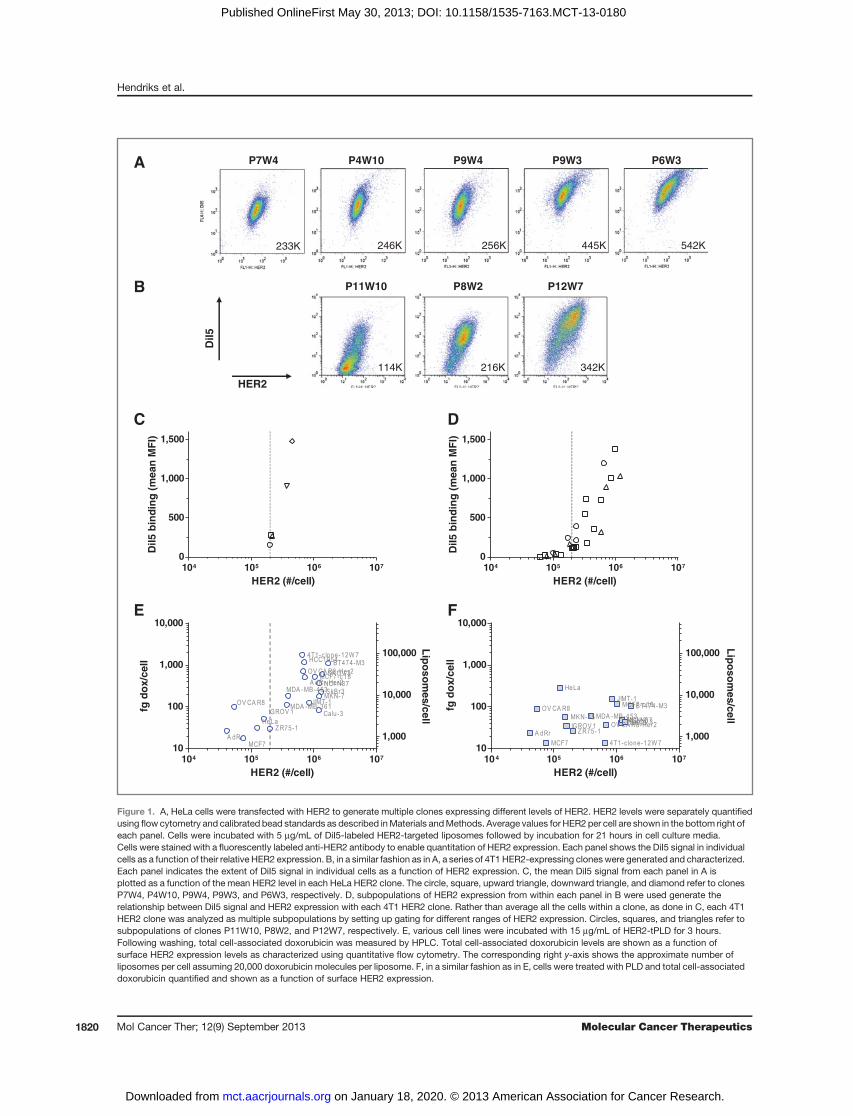
HER2per cell.HumanHER2-transfected cellswere sortedand subcloned to obtain cell populations that expressdifferent ranges of human HER2, shown in Fig. 1A and B.To quantify uptake into the different cell populations,
HER2-targeted empty liposomes were with the fluores-cent dye, DiI5. Empty liposomes were used as doxorubi-
cin has been reported to cause activation of the cytomeg-alovirus (CMV) promoter used in the HER2 construct (39,40). Each HeLa or 4T1-HER2 cell population was incu-bated with fluorescently labeled targeted liposomes for 3hours, washed, incubated in freshmedia for an additional21 hours, and then quantified via flow cytometry. Shownin Fig. 1C, five distinct HER2 clones derived from HeLacells were used to construct a curve of liposome uptakeas a function of mean surface HER2 expression level. Itwas observed that cell-associated DiI5 levels increasedwith increasing HER2 levels. In Fig. 1D, the HER2 clonesderived from 4T1 cells were used to construct a curve ofuptake as a function of surface HER2 expression. In thiscase, subpopulations of each of clone were used to definethe relationship betweenHER2 and uptake. In both cases,there was a sharp increase in targeted liposome uptake atroughly 2 � 105 HER2 per cell.
Experiments were also carried out to determine if theHER2 threshold for HER2-tPLD uptake would holdacross a panel of cell lines. Mean HER2 expression wasassessed for cell lines using qFACS. Each cell line wasincubatedwithHER2-tPLD, PLD, or free doxorubicin andtotal cellular doxorubicin quantified using HPLC. Figure1E shows the uptake of HER2-tPLD as a function of meanHER2 levels, whereas Fig. 1F shows the uptake of PLD.For select cell lines (4T1 12W7),more than100� increase intargeted liposome uptakewas observed. Increasing levelsof HER2 are associated with significant increases in totalcell association of HER2-tPLD. Conversely, the uptake ofPLDwas low and did not correlate with HER2 expression(Fig. 1F). These results suggest that HER2-tPLD is selec-tively taken up into cells that express HER2, especially incells which express more than approximately 2 � 105
receptors per cell. Below 2 � 105 receptors per cell, theuptake of HER2-tPLD is comparable with levels obtainedwith PLD.
InternalizationThe kinetics of association of HER2-tPLD with HER2-
expressing cells has been previously shown (22). We nextsought to quantify the internalization of HER2-tPLD.Cells were incubated at 4�C to halt endocytosis and thenincubated with DiI5-labeled HER2-tPLD to load thesurface with bound HER2-tPLD. Cells were washed andtransferred to 37�C to stimulate endocytosis. The amountof total cell-associated liposomes was measured by flowcytometry by quantifying changes in the DiI5 meanfluorescence intensity (MFI; Fig. 2A). Simultaneously,changes in surface-bound liposomeswere quantified overtime using an anti-PEG assay and are shown in Fig. 2B.The total cellular liposome content, measured by DiI5MFI remains constant, whereas the surface PEG can beshown to decrease over time, indicating internalization.Shown in Fig. 2C is the kinetics of HER2-tPLD internal-ization for multiple cell lines showing cell-specific inter-nalization rates. Internalization rates were estimatedusing early timedata (0–30minutes) to avoid confoundingeffects of recycling (Fig. 2D) and ranged from 0.008 to
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1819
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
DiI5
HER2
P4W10 P9W4 P9W3P7W4 P6W3
233K 246K 256K 445K 542K
P8W2 P12W7P11W10
114K 216K 342K
104 105 106 107
0
500
1,000
1,500
HER2 (#/cell)
DiI
5 b
ind
ing
(m
ea
n M
FI)
104 105 106 107
0
500
1,000
1,500
HER2 (#/cell)
DiI
5 b
ind
ing
(m
ea
n M
FI)
A
B
C D
E F
104 105 106 107
10
100
1,000
10,000
1,000
10,000
100,000
A dRr
OV CA R8
MCF7
HeLaIGROV 1
Z R75-1
MDA -MB-361
MDA -MB-453
4T1-c lone-12W 7
OV CA R8-Her2HCC1954
A dRr-Her2
JIMT-1
MCF7-c 18
Calu-3
MKN-7NCI-N87SkBr3
SKOV 3BT474-M3
HER2 (#/cell)
fg d
ox
/ce
ll
Lip
os
om
es
/ce
ll
104 105 106 107
10
100
1,000
10,000
1,000
10,000
100,000
A dRr
OV CA R8
MCF7
HeLa
MKN-45IGROV 1
Z R75-1
MDA -MB-453
4T1-c lone-12W 7
OV CA R8-Her2
JIMT-1MCF7-c 18
Calu-3MKN-7NCI-N87SKOV 3
BT474-M3
HER2 (#/cell)
fg d
ox
/ce
ll
Lip
os
om
es
/ce
ll
Figure 1. A, HeLa cells were transfected with HER2 to generate multiple clones expressing different levels of HER2. HER2 levels were separately quantifiedusing flow cytometry and calibrated bead standards as described inMaterials andMethods. Average values for HER2 per cell are shown in the bottom right ofeach panel. Cells were incubated with 5 mg/mL of DiI5-labeled HER2-targeted liposomes followed by incubation for 21 hours in cell culture media.Cells were stained with a fluorescently labeled anti-HER2 antibody to enable quantitation of HER2 expression. Each panel shows the DiI5 signal in individualcells as a function of their relative HER2 expression. B, in a similar fashion as in A, a series of 4T1 HER2-expressing cloneswere generated and characterized.Each panel indicates the extent of DiI5 signal in individual cells as a function of HER2 expression. C, the mean DiI5 signal from each panel in A isplotted as a function of the mean HER2 level in each HeLa HER2 clone. The circle, square, upward triangle, downward triangle, and diamond refer to clonesP7W4, P4W10, P9W4, P9W3, and P6W3, respectively. D, subpopulations of HER2 expression from within each panel in B were used generate therelationship between DiI5 signal and HER2 expression with each 4T1 HER2 clone. Rather than average all the cells within a clone, as done in C, each 4T1HER2 clone was analyzed as multiple subpopulations by setting up gating for different ranges of HER2 expression. Circles, squares, and triangles refer tosubpopulations of clones P11W10, P8W2, and P12W7, respectively. E, various cell lines were incubated with 15 mg/mL of HER2-tPLD for 3 hours.Following washing, total cell-associated doxorubicin was measured by HPLC. Total cell-associated doxorubicin levels are shown as a function ofsurface HER2 expression levels as characterized using quantitative flow cytometry. The corresponding right y-axis shows the approximate number ofliposomes per cell assuming 20,000 doxorubicin molecules per liposome. F, in a similar fashion as in E, cells were treated with PLD and total cell-associateddoxorubicin quantified and shown as a function of surface HER2 expression.
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1820
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
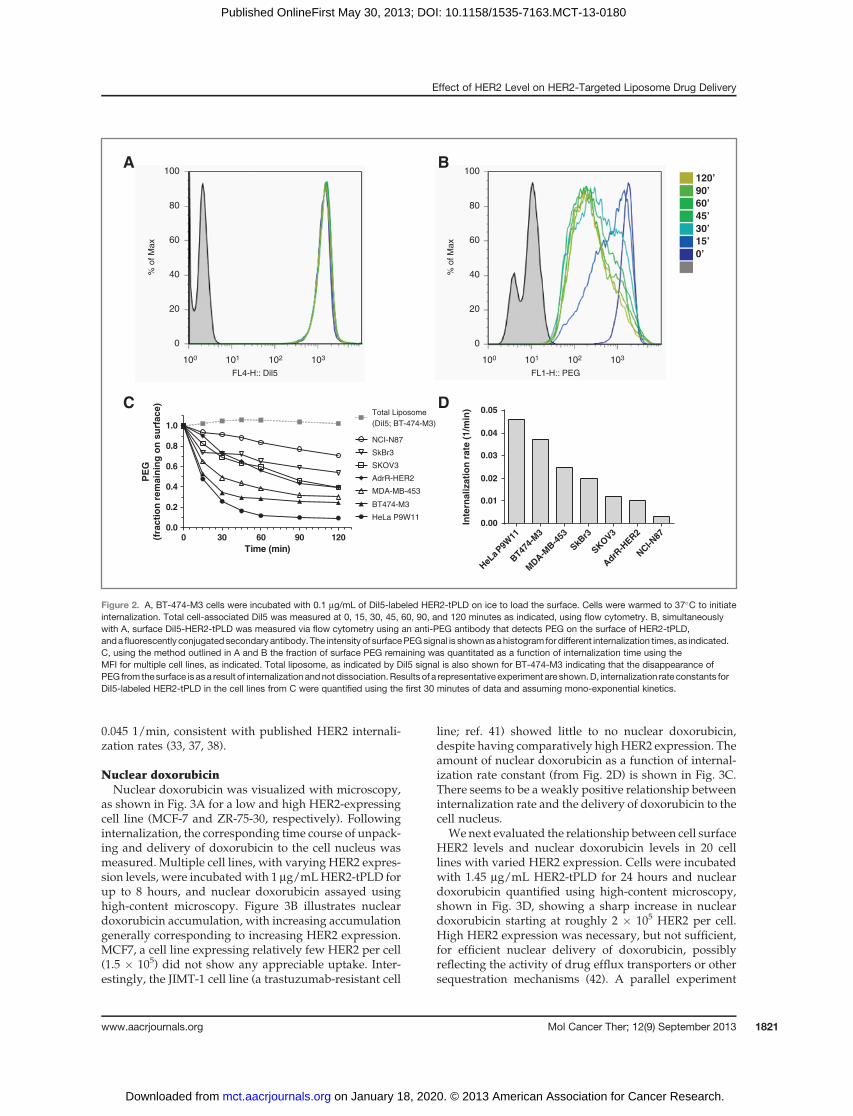
0.045 1/min, consistent with published HER2 internali-zation rates (33, 37, 38).
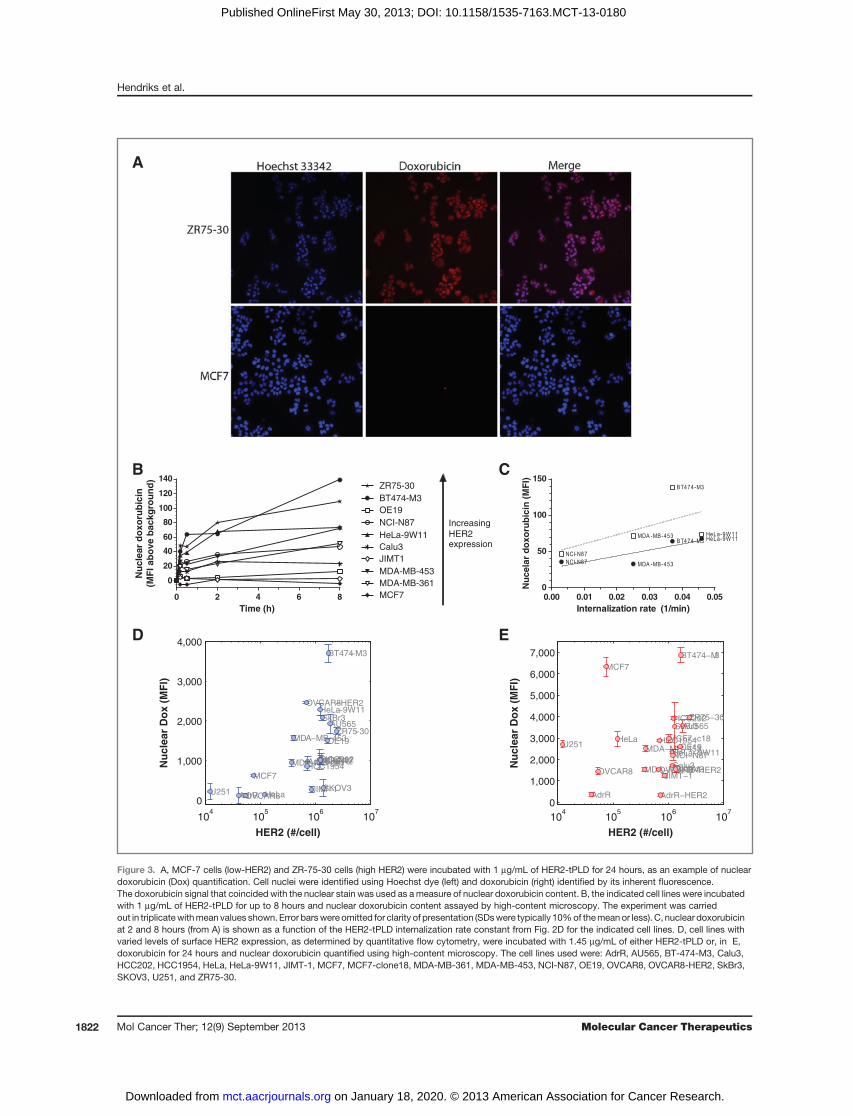
Nuclear doxorubicinNuclear doxorubicin was visualized with microscopy,
as shown in Fig. 3A for a low and high HER2-expressingcell line (MCF-7 and ZR-75-30, respectively). Followinginternalization, the corresponding time course of unpack-ing and delivery of doxorubicin to the cell nucleus wasmeasured. Multiple cell lines, with varying HER2 expres-sion levels, were incubatedwith 1 mg/mLHER2-tPLD forup to 8 hours, and nuclear doxorubicin assayed usinghigh-content microscopy. Figure 3B illustrates nucleardoxorubicin accumulation, with increasing accumulationgenerally corresponding to increasing HER2 expression.MCF7, a cell line expressing relatively few HER2 per cell(1.5 � 105) did not show any appreciable uptake. Inter-estingly, the JIMT-1 cell line (a trastuzumab-resistant cell
line; ref. 41) showed little to no nuclear doxorubicin,despite having comparatively highHER2 expression. Theamount of nuclear doxorubicin as a function of internal-ization rate constant (from Fig. 2D) is shown in Fig. 3C.There seems to be a weakly positive relationship betweeninternalization rate and the delivery of doxorubicin to thecell nucleus.
We next evaluated the relationship between cell surfaceHER2 levels and nuclear doxorubicin levels in 20 celllines with varied HER2 expression. Cells were incubatedwith 1.45 mg/mL HER2-tPLD for 24 hours and nucleardoxorubicin quantified using high-content microscopy,shown in Fig. 3D, showing a sharp increase in nucleardoxorubicin starting at roughly 2 � 105 HER2 per cell.High HER2 expression was necessary, but not sufficient,for efficient nuclear delivery of doxorubicin, possiblyreflecting the activity of drug efflux transporters or othersequestration mechanisms (42). A parallel experiment
120’
90’
60’
45’
30’
15’
0’
HeL
a P9W
11
BT47
4-M
3
MDA-M
B-4
53
SkBr3
SKOV3
AdrR
-HER
2
NCI-N
87
0.00
0.01
0.02
0.03
0.04
0.05
Inte
rnali
zati
on
rate
(1/m
in)
100
80
60
40
20
0
100 101
FL4-H:: Dil5
% o
f M
ax
% o
f M
ax
FL1-H:: PEG
102 103 100 101 102 103
100
80
60
40
20
0
0 30 60 90 1200.0
0.2
0.4
0.6
0.8
1.0
HeLa P9W11
BT474-M3
AdrR-HER2
NCI-N87
SKOV3
MDA-MB-453
SkBr3
Total Liposome
(DiI5; BT-474-M3)
Time (min)
PE
G
(fra
cti
on
rem
ain
ing
on
su
rfa
ce
)A B
C D
Figure 2. A, BT-474-M3 cells were incubated with 0.1 mg/mL of DiI5-labeled HER2-tPLD on ice to load the surface. Cells were warmed to 37�C to initiateinternalization. Total cell-associated DiI5 was measured at 0, 15, 30, 45, 60, 90, and 120 minutes as indicated, using flow cytometry. B, simultaneouslywith A, surface DiI5-HER2-tPLD was measured via flow cytometry using an anti-PEG antibody that detects PEG on the surface of HER2-tPLD,andafluorescently conjugatedsecondary antibody. The intensity of surfacePEGsignal is shownasahistogram for different internalization times, as indicated.C, using the method outlined in A and B the fraction of surface PEG remaining was quantitated as a function of internalization time using theMFI for multiple cell lines, as indicated. Total liposome, as indicated by DiI5 signal is also shown for BT-474-M3 indicating that the disappearance ofPEG from thesurface is as a result of internalizationandnot dissociation.Results of a representative experiment are shown.D, internalization rate constants forDiI5-labeled HER2-tPLD in the cell lines from C were quantified using the first 30 minutes of data and assuming mono-exponential kinetics.
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1821
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
104
105
106
107
0
1,000
2,000
3,000
4,000
5,000
6,000
7,000
AdrR AdrR−HER2
AU565
BT474−M3
Calu3
HCC202
HCC1954HeLa
HeLa−9W11
JIMT−1
MCF7
MCF7−c18
MDA−MB−361
MDA−MB−453NCI−N87
OE19
OVCAR8 OVCAR8−HER2
SkBr3
SKOV3
U251
ZR75−30
HER2 (#/cell)
Nu
cle
ar
Do
x (
MF
I)
104
105
106
107
0
1,000
2,000
3,000
4,000
AdrR
AdrR−HER2
AU565
BT474−M3
Calu3HCC202HCC1954
HeLa
HeLa−9W11
JIMT−1
MCF7
MDA−MB−361
MDA−MB−453
NCI−N87
OE19
OVCAR8
OVCAR8−HER2
SkBr3
SKOV3U251
ZR75−30
HER2 (#/cell)
Nu
cle
ar
Do
x (
MF
I)
D E
B C
A
0 2 4 6 8
0
20
40
60
80
100
120
140
BT474-M3
HeLa-9W11
MDA-MB-453
MCF7
NCI-N87
OE19
MDA-MB-361
JIMT1
Calu3
ZR75-30
Time (h)
Nu
cle
ar
do
xo
rub
icin
(MF
I ab
ove b
ackg
rou
nd
)
IncreasingHER2expression
Internalization rate (1/min)
Nu
ce
lar
do
xo
rub
icin
(M
FI)
0.00 0.01 0.02 0.03 0.04 0.050
50
100
150
BT474-M3 HeLa-9W 11
MDA -MB-453NCI-N87
BT474-M3
HeLa-9W 11MDA -MB-453
NCI-N87
Figure 3. A, MCF-7 cells (low-HER2) and ZR-75-30 cells (high HER2) were incubated with 1 mg/mL of HER2-tPLD for 24 hours, as an example of nucleardoxorubicin (Dox) quantification. Cell nuclei were identified using Hoechst dye (left) and doxorubicin (right) identified by its inherent fluorescence.The doxorubicin signal that coincided with the nuclear stain was used as ameasure of nuclear doxorubicin content. B, the indicated cell lines were incubatedwith 1 mg/mL of HER2-tPLD for up to 8 hours and nuclear doxorubicin content assayed by high-content microscopy. The experiment was carriedout in triplicatewithmean values shown. Error barswere omitted for clarity of presentation (SDswere typically 10%of themeanor less). C, nuclear doxorubicinat 2 and 8 hours (from A) is shown as a function of the HER2-tPLD internalization rate constant from Fig. 2D for the indicated cell lines. D, cell lines withvaried levels of surface HER2 expression, as determined by quantitative flow cytometry, were incubated with 1.45 mg/mL of either HER2-tPLD or, in E,doxorubicin for 24 hours and nuclear doxorubicin quantified using high-content microscopy. The cell lines used were: AdrR, AU565, BT-474-M3, Calu3,HCC202, HCC1954, HeLa, HeLa-9W11, JIMT-1, MCF7, MCF7-clone18, MDA-MB-361, MDA-MB-453, NCI-N87, OE19, OVCAR8, OVCAR8-HER2, SkBr3,SKOV3, U251, and ZR75-30.
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1822
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

using free doxorubicin showed no relationship betweenHER2 expression and nuclear doxorubicin uptake, shownin Fig. 3E.
HER2-tPLD cellular uptake modelWe constructed a kinetic computational model to solid-
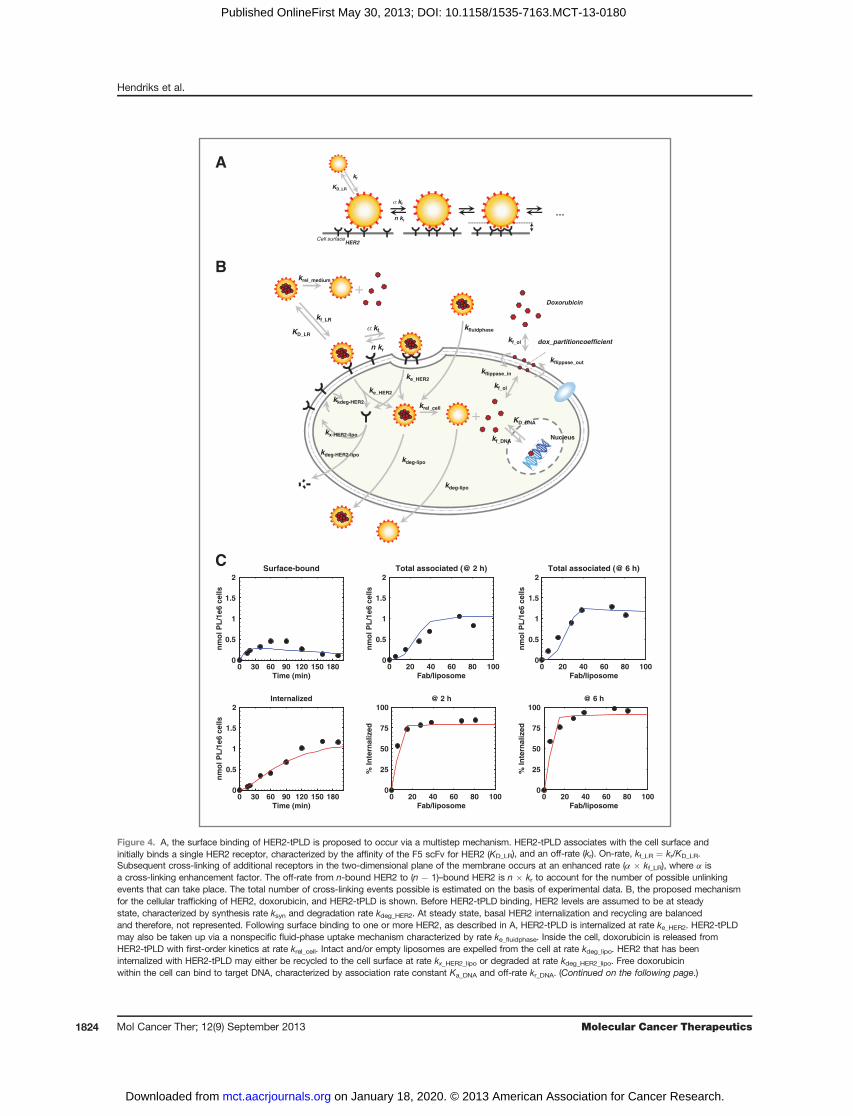
ify our understanding of the various kinetic steps fromHER2-tPLD binding (including its dependence on HER2expression level), internalization, doxorubicin release,and nuclear binding, described inMaterials andMethodsand shown in Fig. 4A and B.Model training. We incorporated model parameters
available from the literature, however, several HER2-tPLD and cell-specific parameters required estimation.A publication on a highly related form of HER2-tPLD(differing only in the antibody used) from Kirpotin andcolleagues (43) contains a rich dataset on the effect ofvarying antibody density and time courses of surface-bound versus internalized liposomes. Using these data,andaccounting for thedifferent affinity of the 4D5 scFv [22nmol/L (44) versus 160 nmol/L for F5 (28)], we were ableto generate estimates for the liposome-cell association rate(kf_LR), the maximum number of cross-linking events (n),the cross-linking enhancement factor (a), the recyclingand degradation rate of internalized HER2 followingHER2-tPLD binding (kx_HER2_lipo, kdeg_HER2_lipo), the lipo-some/lipid expulsion rate (kdeg_lipo), the cell-specificHER2 internalization rate (ke), and the rate of nonspecificuptake (ke_fluidphase). Shown in Fig. 4C is the simultaneousfit of the HER2-tPLD cellular traffickingmodel to the datafrom Kirpotin and colleagues (43). Parameter values arelisted in Supplementary Table S1. Among the parametersis the estimation that the maximum number of cross-linking (F5-HER2 binding) events is four per liposome–cell interaction. This indicates that four F5-scFv, or rough-ly 10% of liposome surface, interacts with the cell in casesinwhichHER2 is not limiting. The value is also consistentwith experimental observations, that it is not possible tostrip HER2-tPLD off of high HER2-expressing cells withlow pH solutions, as one might do with immunoglobulinG (IgG) molecules (data not shown).Effect of HER2 expression level on nuclear doxorubicin
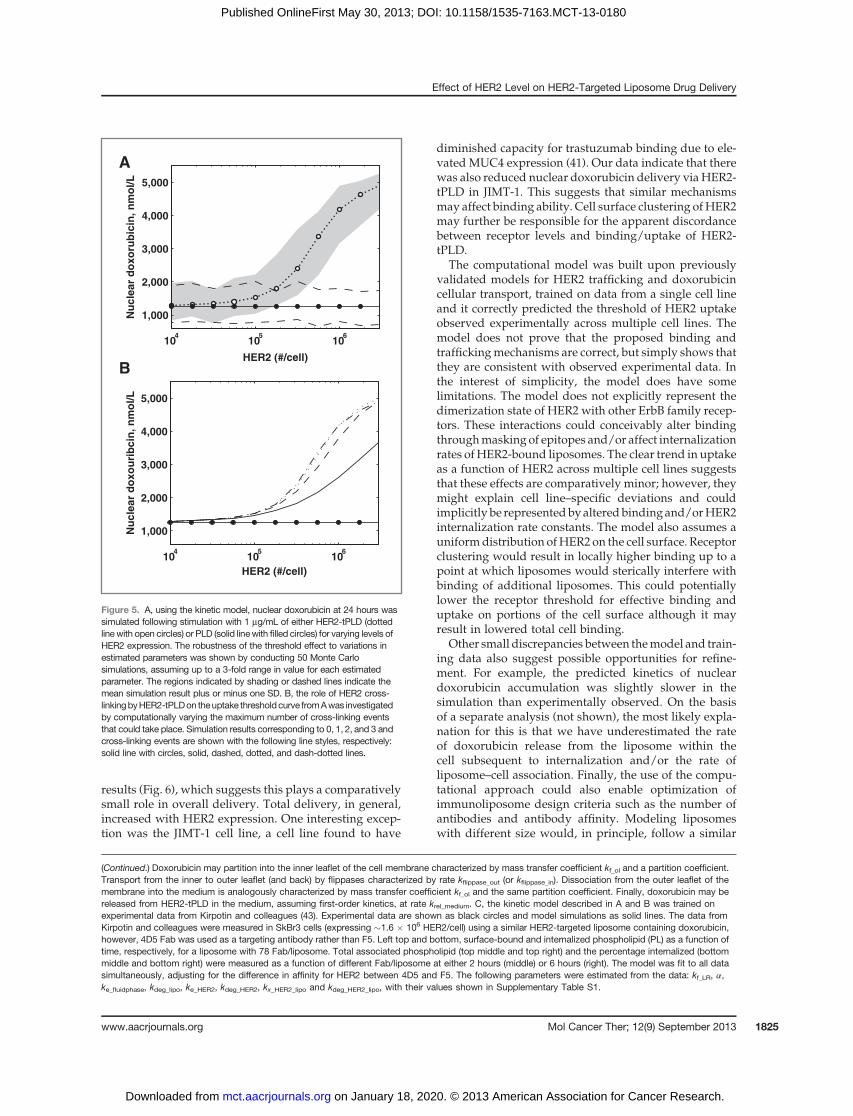
delivery. Weused themodel to simulate the relationshipbetween nuclear doxorubicin and HER2 expression levelfollowing incubation with 0.1 mg/mL of HER2-tPLD,shown in Fig. 5A. The model predicts a threshold ofuptake of approximately 2 � 105 HER2 per cell, highlyconsistent with the experimental data from Figs. 1 and 3.Also, indicated are confidence intervals derived fromMonteCarlo simulations assumingup to 3-fold variabilityin the values of the estimated parameters, illustrating therobustness of this result.At lowHER2expression, bindingis inefficient and the small amount of nuclear doxorubicincontent arises from two components: nonspecific lipo-some uptake and external doxorubicin release. In Fig.5B, we simulate the effect of different numbers of cross-linking events on the delivery of doxorubicin to thenucleus. This indicates that multiple receptor interactions
enhance binding and delivery, although there are dimin-ishing returns after two cross-linking events. Finally, at 2� 105 HER2 per cell, there would, on average, only be twoHER2 under the projected area of each liposome. This isconsistent with the beginning of an avidity effect andsubsequent threshold-like behavior for binding anduptake.
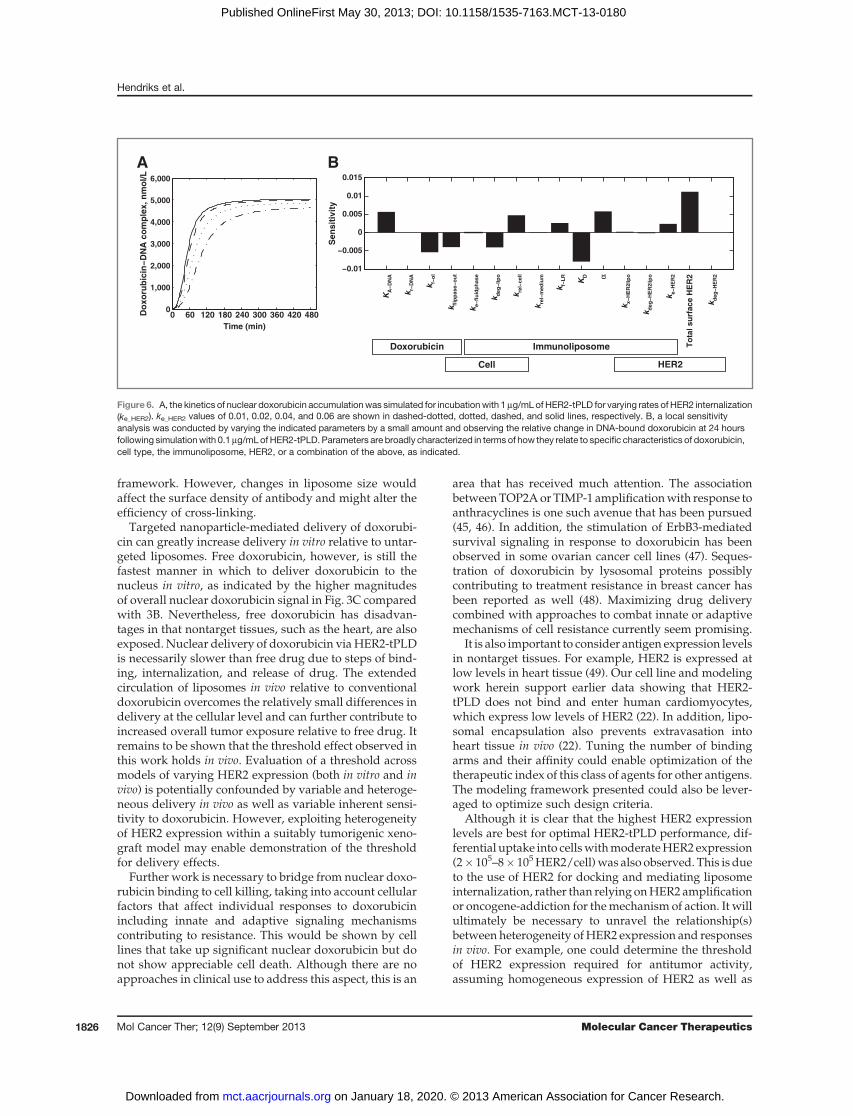
Effect of HER2 internalization rate on nuclear doxoru-bicin delivery. Because of the interest in antigen selec-tion for the development of targeted nanoparticles, wealso explored the importance of internalization rate.Shown in Fig. 6A, we simulated the kinetics of nucleardoxorubicin delivery for a range of internalization rates,based on Fig. 2D. Here, we observed that, within thephysiologic range of internalization rates for HER2, thereis a weakly positive relationship between internalizationrate and nuclear doxorubicin delivery. This result wasconfirmed by experimental observations that internaliza-tion rates were modestly correlated with increased nucle-ar doxorubicin (see Fig. 3C). A sensitivity analysis, shownin Fig. 6B, further indicates that the rate of internalizationplays a comparatively small role relative to other kineticsteps. HER2 expression level is themost important factor,followed by two liposome-specific parameters: the scFvaffinity (KD) and the cross-linking enhancement factor (a).Internalization is a necessary step for effective delivery,but it is not predicted to be kinetically limiting.
DiscussionIn this work, we examined the quantitative relationship
between HER2 expression levels and the performance ofHER2-tPLD. We show that HER2-tPLD shows increasedcellular uptake with increasing HER2 expression andcorresponding increases in delivery of doxorubicin to thenucleus.Most notably,we found a threshold effect, takingplace at approximately 2 � 105 HER2 per cell, at whichthere was enhanced binding and uptake of HER2-tPLDrelative to PLD. This threshold held both within a singlecell background and across multiple cell lines. This sug-gests that a critical density of HER2 receptors is necessaryon the cell surface for efficient binding of HER2-tPLD andcross-linking of receptors. The presence of a thresholdeffect is further shown using a computational model thatcaptures the key steps in HER2-tPLD–mediated deliveryof doxorubicin. Using the model, we are able to show thatthe threshold effect arises from multiple low-affinityinteractions leading to high aviditiy, estimate the extentof cross-linking of HER2, and characterize the role ofinternalization on nuclear doxorubicin delivery.
Experimentally, we found variability across cell linesat all levels. Internalization of HER2-tPLD varied consid-erably, but was within ranges previously observed. Dif-ferences in internalization rate may reflect differentextents of heterodimerization with EGFR; increased het-erodimerization with EGFR might result in slightlyincreased internalization of HER2-tPLD (33). There wasaweak correlation between internalization rate andnucle-ar doxorubicin accumulation confirmed via simulation
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1823
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
0 20 40 60 80 1000
0.5
1
1.5
2
Fab/liposome
nm
ol
PL
/1e
6 c
ell
s
Total associated (@ 2 h)
0 20 40 60 80 1000
0.5
1
1.5
2
Fab/liposome
nm
ol
PL
/1e
6 c
ell
s
Total associated (@ 6 h)
0 20 40 60 80 1000
25
50
75
100
Fab/liposome
% I
nte
rna
lize
d
@ 2 h
0 20 40 60 80 1000
25
50
75
100
Fab/liposome
% I
nte
rna
lize
d
@ 6 h
0 30 60 90 120 150 1800
0.5
1
1.5
2
Time (min)
nm
ol
PL
/1e
6 c
ell
s
Surface-bound
0 30 60 90 120 150 1800
0.5
1
1.5
2
Time (min)
nm
ol
PL
/1e
6 c
ell
s
Internalized
Cell surfaceHER2
…
kf
kf
n kr
KD_LR
A
B
C
Nucleuskf_DNA
KD_DNA
Doxorubicin
kf_ol dox_partitioncoefficient
kf_ol
kflippase_in
kflippase_out
kf_LR
kf
n kr
KD_LR
ke_HER2
ke_HER2
kx-HER2-lipo
krel_cell
kdeg-lipo
kdeg-HER2-lipo
+
+krel_medium
kkdeg-HER2
kfluidphase
kdeg-lipo
Figure 4. A, the surface binding of HER2-tPLD is proposed to occur via a multistep mechanism. HER2-tPLD associates with the cell surface andinitially binds a single HER2 receptor, characterized by the affinity of the F5 scFv for HER2 (KD_LR), and an off-rate (kr). On-rate, kf_LR ¼ kr/KD_LR.Subsequent cross-linking of additional receptors in the two-dimensional plane of the membrane occurs at an enhanced rate (a � kf_LR), where a isa cross-linking enhancement factor. The off-rate from n-bound HER2 to (n � 1)–bound HER2 is n � kr to account for the number of possible unlinkingevents that can take place. The total number of cross-linking events possible is estimated on the basis of experimental data. B, the proposed mechanismfor the cellular trafficking of HER2, doxorubicin, and HER2-tPLD is shown. Before HER2-tPLD binding, HER2 levels are assumed to be at steadystate, characterized by synthesis rate ksyn and degradation rate kdeg_HER2. At steady state, basal HER2 internalization and recycling are balancedand therefore, not represented. Following surface binding to one or more HER2, as described in A, HER2-tPLD is internalized at rate ke_HER2. HER2-tPLDmay also be taken up via a nonspecific fluid-phase uptake mechanism characterized by rate ke_fluidphase. Inside the cell, doxorubicin is released fromHER2-tPLD with first-order kinetics at rate krel_cell. Intact and/or empty liposomes are expelled from the cell at rate kdeg_lipo. HER2 that has beeninternalized with HER2-tPLD may either be recycled to the cell surface at rate kx_HER2_lipo or degraded at rate kdeg_HER2_lipo. Free doxorubicinwithin the cell can bind to target DNA, characterized by association rate constant Ka_DNA and off-rate kr_DNA. (Continued on the following page.)
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1824
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
results (Fig. 6), which suggests this plays a comparativelysmall role in overall delivery. Total delivery, in general,increased with HER2 expression. One interesting excep-tion was the JIMT-1 cell line, a cell line found to have
diminished capacity for trastuzumab binding due to ele-vated MUC4 expression (41). Our data indicate that therewas also reduced nuclear doxorubicin delivery viaHER2-tPLD in JIMT-1. This suggests that similar mechanismsmay affect binding ability. Cell surface clustering ofHER2may further be responsible for the apparent discordancebetween receptor levels and binding/uptake of HER2-tPLD.
The computational model was built upon previouslyvalidated models for HER2 trafficking and doxorubicincellular transport, trained on data from a single cell lineand it correctly predicted the threshold of HER2 uptakeobserved experimentally across multiple cell lines. Themodel does not prove that the proposed binding andtraffickingmechanisms are correct, but simply shows thatthey are consistent with observed experimental data. Inthe interest of simplicity, the model does have somelimitations. The model does not explicitly represent thedimerization state of HER2 with other ErbB family recep-tors. These interactions could conceivably alter bindingthroughmasking of epitopes and/or affect internalizationrates ofHER2-bound liposomes. The clear trend in uptakeas a function of HER2 across multiple cell lines suggeststhat these effects are comparatively minor; however, theymight explain cell line–specific deviations and couldimplicitly be representedbyalteredbindingand/orHER2internalization rate constants. The model also assumes auniformdistribution ofHER2 on the cell surface. Receptorclustering would result in locally higher binding up to apoint at which liposomes would sterically interfere withbinding of additional liposomes. This could potentiallylower the receptor threshold for effective binding anduptake on portions of the cell surface although it mayresult in lowered total cell binding.
Other small discrepancies between themodel and train-ing data also suggest possible opportunities for refine-ment. For example, the predicted kinetics of nucleardoxorubicin accumulation was slightly slower in thesimulation than experimentally observed. On the basisof a separate analysis (not shown), the most likely expla-nation for this is that we have underestimated the rateof doxorubicin release from the liposome within thecell subsequent to internalization and/or the rate ofliposome–cell association. Finally, the use of the compu-tational approach could also enable optimization ofimmunoliposome design criteria such as the number ofantibodies and antibody affinity. Modeling liposomeswith different size would, in principle, follow a similar
HER2 (#/cell)
Nu
cle
ar
do
xo
rub
icin
, n
mo
l/L
104
105
106
1,000
2,000
3,000
4,000
5,000
Nu
cle
ar
do
xo
uri
bcin
, n
mo
l/L
A
B
104
105
106
1,000
2,000
3,000
4,000
5,000
HER2 (#/cell)
Figure 5. A, using the kinetic model, nuclear doxorubicin at 24 hours wassimulated following stimulation with 1 mg/mL of either HER2-tPLD (dottedline with open circles) or PLD (solid linewith filled circles) for varying levels ofHER2 expression. The robustness of the threshold effect to variations inestimated parameters was shown by conducting 50 Monte Carlosimulations, assuming up to a 3-fold range in value for each estimatedparameter. The regions indicated by shading or dashed lines indicate themean simulation result plus or minus one SD. B, the role of HER2 cross-linkingbyHER2-tPLDon theuptake thresholdcurve fromAwas investigatedby computationally varying the maximum number of cross-linking eventsthat could take place. Simulation results corresponding to 0, 1, 2, and 3 andcross-linking events are shown with the following line styles, respectively:solid line with circles, solid, dashed, dotted, and dash-dotted lines.
(Continued.) Doxorubicin may partition into the inner leaflet of the cell membrane characterized by mass transfer coefficient kf_ol and a partition coefficient.Transport from the inner to outer leaflet (and back) by flippases characterized by rate kflippase_out (or kflippase_in). Dissociation from the outer leaflet of themembrane into the medium is analogously characterized by mass transfer coefficient kf_ol and the same partition coefficient. Finally, doxorubicin may bereleased from HER2-tPLD in the medium, assuming first-order kinetics, at rate krel_medium. C, the kinetic model described in A and B was trained onexperimental data from Kirpotin and colleagues (43). Experimental data are shown as black circles and model simulations as solid lines. The data fromKirpotin and colleagues were measured in SkBr3 cells (expressing �1.6 � 106 HER2/cell) using a similar HER2-targeted liposome containing doxorubicin,however, 4D5 Fab was used as a targeting antibody rather than F5. Left top and bottom, surface-bound and internalized phospholipid (PL) as a function oftime, respectively, for a liposome with 78 Fab/liposome. Total associated phospholipid (top middle and top right) and the percentage internalized (bottommiddle and bottom right) were measured as a function of different Fab/liposome at either 2 hours (middle) or 6 hours (right). The model was fit to all datasimultaneously, adjusting for the difference in affinity for HER2 between 4D5 and F5. The following parameters were estimated from the data: kf_LR, a,ke_fluidphase, kdeg_lipo, ke_HER2, kdeg_HER2, kx_HER2_lipo and kdeg_HER2_lipo, with their values shown in Supplementary Table S1.
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1825
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180
framework. However, changes in liposome size wouldaffect the surface density of antibody and might alter theefficiency of cross-linking.
Targeted nanoparticle-mediated delivery of doxorubi-cin can greatly increase delivery in vitro relative to untar-geted liposomes. Free doxorubicin, however, is still thefastest manner in which to deliver doxorubicin to thenucleus in vitro, as indicated by the higher magnitudesof overall nuclear doxorubicin signal in Fig. 3C comparedwith 3B. Nevertheless, free doxorubicin has disadvan-tages in that nontarget tissues, such as the heart, are alsoexposed. Nuclear delivery of doxorubicin viaHER2-tPLDis necessarily slower than free drug due to steps of bind-ing, internalization, and release of drug. The extendedcirculation of liposomes in vivo relative to conventionaldoxorubicin overcomes the relatively small differences indelivery at the cellular level and can further contribute toincreased overall tumor exposure relative to free drug. Itremains to be shown that the threshold effect observed inthis work holds in vivo. Evaluation of a threshold acrossmodels of varying HER2 expression (both in vitro and invivo) is potentially confounded by variable and heteroge-neous delivery in vivo as well as variable inherent sensi-tivity to doxorubicin. However, exploiting heterogeneityof HER2 expression within a suitably tumorigenic xeno-graft model may enable demonstration of the thresholdfor delivery effects.
Further work is necessary to bridge from nuclear doxo-rubicin binding to cell killing, taking into account cellularfactors that affect individual responses to doxorubicinincluding innate and adaptive signaling mechanismscontributing to resistance. This would be shown by celllines that take up significant nuclear doxorubicin but donot show appreciable cell death. Although there are noapproaches in clinical use to address this aspect, this is an
area that has received much attention. The associationbetweenTOP2AorTIMP-1 amplificationwith response toanthracyclines is one such avenue that has been pursued(45, 46). In addition, the stimulation of ErbB3-mediatedsurvival signaling in response to doxorubicin has beenobserved in some ovarian cancer cell lines (47). Seques-tration of doxorubicin by lysosomal proteins possiblycontributing to treatment resistance in breast cancer hasbeen reported as well (48). Maximizing drug deliverycombined with approaches to combat innate or adaptivemechanisms of cell resistance currently seem promising.
It is also important to consider antigen expression levelsin nontarget tissues. For example, HER2 is expressed atlow levels in heart tissue (49). Our cell line and modelingwork herein support earlier data showing that HER2-tPLD does not bind and enter human cardiomyocytes,which express low levels of HER2 (22). In addition, lipo-somal encapsulation also prevents extravasation intoheart tissue in vivo (22). Tuning the number of bindingarms and their affinity could enable optimization of thetherapeutic index of this class of agents for other antigens.The modeling framework presented could also be lever-aged to optimize such design criteria.
Although it is clear that the highest HER2 expressionlevels are best for optimal HER2-tPLD performance, dif-ferential uptake into cellswithmoderateHER2 expression(2� 105–8� 105HER2/cell)was also observed. This is dueto the use of HER2 for docking and mediating liposomeinternalization, rather than relying onHER2 amplificationor oncogene-addiction for themechanism of action. It willultimately be necessary to unravel the relationship(s)between heterogeneity ofHER2 expression and responsesin vivo. For example, one could determine the thresholdof HER2 expression required for antitumor activity,assuming homogeneous expression of HER2 as well as
−0.01
−0.005
0
0.005
0.01
0.015
Sen
sit
ivit
y
KA
−D
NA
k r−D
NA
k f−o
l
k flip
pase−
ou
t
k e−
flu
idp
hase
k deg
−lip
o
k rel−
cell
k rel−
med
ium
k f−L
R
KD α
k x−
HE
R2lip
o
k deg
−H
ER
2lip
o
To
tal su
rface H
ER
2
k e−
HE
R2
k deg
−H
ER
2
Doxorubicin
Cell
Immunoliposome
HER2
Do
xo
rub
icin
−D
NA
co
mp
lex, n
mo
l/L
A B
0 60 120 180 240 300 360 420 4800
1,000
2,000
3,000
4,000
5,000
6,000
Time (min)
Figure 6. A, the kinetics of nuclear doxorubicin accumulationwas simulated for incubationwith 1mg/mLof HER2-tPLD for varying rates of HER2 internalization(ke_HER2). ke_HER2 values of 0.01, 0.02, 0.04, and 0.06 are shown in dashed-dotted, dotted, dashed, and solid lines, respectively. B, a local sensitivityanalysis was conducted by varying the indicated parameters by a small amount and observing the relative change in DNA-bound doxorubicin at 24 hoursfollowing simulationwith 0.1mg/mLof HER2-tPLD. Parameters are broadly characterized in terms of how they relate to specific characteristics of doxorubicin,cell type, the immunoliposome, HER2, or a combination of the above, as indicated.
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1826
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

determine the percentage of cells expressing high HER2levels that are necessary to observe responses to HER2-tPLD. Nonetheless, this work suggests an opportunity totarget patient tumors with moderate levels of HER2expression where few treatment options exist. This couldgreatly increase the potential patient population beyondthat of traditionally defined "HER2-positive" disease.In these cases, it may be especially important to under-stand the ability to target small metastases and/or iden-tify tumors that show high degrees of overall liposomedeposition.
Disclosure of Potential Conflicts of InterestT.J. Wickham has ownership interest (including patents) in Merrimack
Pharmaceuticals. No potential conflicts of interest were disclosed by theother authors.
Authors' ContributionsConception and design: B.S. Hendriks, S.G. Klinz, J.G. Reynolds, C.W.Espelin, T.J. WickhamDevelopment of methodology: S.G. Klinz, J.G. Reynolds, D.F. Gaddy, T.J.Wickham
Acquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): S.G. Klinz, J.G. Reynolds, D.F. GaddyAnalysis and interpretation of data (e.g., statistical analysis, biostatis-tics, computational analysis): B.S. Hendriks, S.G. Klinz, J.G. Reynolds,C.W. Espelin, D.F. Gaddy, T.J. WickhamWriting, review, and/or revision of the manuscript: B.S. Hendriks,S.G. Klinz, J.G. Reynolds, D.F. Gaddy, T.J. WickhamAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): S.G. Klinz, J.G. ReynoldsStudy supervision: J.G. Reynolds
AcknowledgmentsThe authors thank Isabelle Eckelhofer for technical assistance and
Dmitri Kirpotin, Daryl Drummond, John Park, Chris Benz, Ulrik Nielsen,Elena Geretti, Shannon Leonard, Helen Lee, and Nancy Dumont forinsightful discussions.
Grant SupportThis work was funded by Merrimack Pharmaceuticals.The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received March 8, 2013; revised April 22, 2013; accepted May 24, 2013;published OnlineFirst May 30, 2013.
References1. Paszko E, Senge MO. Immunoliposomes. Curr Med Chem 2012;19:
5239–77.2. Mamot C, Ritschard R, Wicki A, Stehle G, Dieterle T, Bubendorf L,
et al. Tolerability, safety, pharmacokinetics, and efficacy of doxo-rubicin-loaded anti-EGFR immunoliposomes in advanced solidtumours: a phase 1 dose-escalation study. Lancet Oncol 2012;13:1234–41.
3. Mamot C, Drummond DC, Greiser U, Hong K, Kirpotin DB, MarksJD, et al. Epidermal growth factor receptor (EGFR)-targeted immu-noliposomes mediate specific and efficient drug delivery to EGFR-and EGFRvIII-overexpressing tumor cells. Cancer Res 2003;63:3154–61.
4. Lee RJ, Low PS. Delivery of liposomes into cultured KB cells via folatereceptor-mediated endocytosis. J Biol Chem 1994;269:3198–204.
5. Zhai G, Wu J, Yu B, Guo C, Yang X, Lee RJ. A transferrin receptor-targeted liposomal formulation for docetaxel. J Nanosci Nanotechnol2010;10:5129–36.
6. Dunne M, Zheng J, Rosenblat J, Jaffray DA, Allen C. APN/CD13-targeting as a strategy to alter the tumor accumulation of liposomes.J Control Release 2011;154:298–305.
7. Pastorino F, Di Paolo D, Piccardi F, Nico B, Ribatti D, Daga A, et al.Enhanced antitumor efficacy of clinical-grade vasculature-targetedliposomal doxorubicin. Clin Cancer Res 2008;14:7320–9.
8. Brignole C, Marimpietri D, Di Paolo D, Perri P, Morandi F, Pastorino F,et al. Therapeutic targeting of TLR9 inhibits cell growth and inducesapoptosis in neuroblastoma. Cancer Res 2010;70:9816–26.
9. Wang L, Su W, Liu Z, Zhou M, Chen S, Chen Y, et al. CD44 antibody-targeted liposomal nanoparticles for molecular imaging and therapy ofhepatocellular carcinoma. Biomaterials 2012;33:5107–14.
10. Mitchell MJ, Chen CS, Ponmudi V, Hughes AD, King MR. E-selectinliposomal and nanotube-targeted delivery of doxorubicin to circulatingtumor cells. J Control Release 2012;160:609–17.
11. Shroff K, Kokkoli E. PEGylated liposomal doxorubicin targeted toalpha5beta1-expressing MDA-MB-231 breast cancer cells. Langmuir2012;28:4729–36.
12. PetersenAL, Binderup T, Jolck RI, Rasmussen P,Henriksen JR, PfeiferAK, et al. Positron emission tomography evaluation of somatostatinreceptor targeted 64Cu-TATE-liposomes in a human neuroendocrinecarcinoma mouse model. J Control Release 2012;160:254–63.
13. Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A, McGuire WL.Human breast cancer: correlation of relapse and survival with ampli-fication of the HER-2/neu oncogene. Science 1987;235:177–82.
14. Scholl S, BeuzebocP,Pouillart P. TargetingHER2 inother tumor types.Ann Oncol 2001;12(Suppl 1):S81–7.
15. Ross JS, Fletcher JA, Linette GP, Stec J, Clark E, Ayers M, et al. TheHer-2/neu gene and protein in breast cancer 2003: biomarker andtarget of therapy. Oncologist 2003;8:307–25.
16. Koltz BR, Hicks DG, Whitney-Miller CL. HER2 testing in gastric andesophageal adenocarcinoma: new diagnostic challenges arising fromnew therapeutic options. Biotech Histochem 2012;87:40–5.
17. HercepTest Interpretation Manual – Breast. (Dako, Carpinteria, CA)Literature Order No. 29036. Available from: http://www.dako.com/us/29036_herceptest_interpretation_manual-breast_ihc_us.pdf.
18. Rossi V, Sarotto I, Maggiorotto F, Berchialla P, Kubatzki F, Tomasi N,et al. Moderate immunohistochemical expression of HER-2 (2þ) with-out HER-2 gene amplification is a negative prognostic factor in earlybreast cancer. Oncologist 2012;17:1418–25.
19. NSABP Protocol B-47. A Randomized Phase III Trial of AdjuvantTherapy Comparing Chemotherapy Alone (Six Cycles of DocetaxelPlus Cyclophosphamide or Four Cycles of Doxorubicin Plus Cyclo-phosphamide Followed by Weekly Paclitaxel) to Chemotherapy PlusTrastuzumab in Women with Node-Positive or High-Risk Node-Neg-ative HER2-Low Invasive Breast Cancer. Available from: http://www.nsabp.pitt.edu/B-47.asp.
20. Kunz PL, Mojtahed A, Fisher GA, Ford JM, Chang DT, Balise RR, et al.HER2 expression in gastric and gastroesophageal junction adenocar-cinoma in a US population: clinicopathologic analysis with proposedapproach to HER2 assessment. Appl ImmunohistochemMol Morphol2012;20:13–24.
21. Munster P, Miller K, Krop IE, Dhindsa N, Niyikiza C, Nielsen U, et al.A phase 1 study of MM-302, a HER2-targeted liposomal doxoru-bicin, in patients with advanced HER2-positive (HER2þ) breastcancer. In: Proceedings of the Thirty-Fifth Annual CTRC-AACR SanAntonio Breast Cancer Symposium; 2012 Dec 4–8; San Antonio, TX.Philadelphia (PA): AACR. Cancer Res 2012;72(24 Suppl):Abstract nrP5-18-09.
22. Reynolds JG, Geretti E, Hendriks BS, Lee H, Leonard SC, Klinz SG,et al. HER2-targeted liposomal doxorubicin displays enhanced anti-tumorigenic effects without associated cardiotoxicity. Toxicol ApplPharmacol 2012;262:1–10.
23. Neve RM, Nielsen UB, Kirpotin DB, Poul MA, Marks JD, Benz CC.Biological effects of anti-ErbB2 single chain antibodies selected forinternalizing function. Biochem Biophys Res Commun 2001;280:274–9.
Effect of HER2 Level on HER2-Targeted Liposome Drug Delivery
www.aacrjournals.org Mol Cancer Ther; 12(9) September 2013 1827
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

24. Pommier Y, Leo E, Zhang H, Marchand C. DNA topoisomerases andtheir poisoning by anticancer and antibacterial drugs. ChemBiol 2010;17:421–33.
25. Peer D, Karp JM, Hong S, Farokhzad OC, Margalit R, Langer R.Nanocarriers as an emerging platform for cancer therapy. Nat Nano-technol 2007;2:751–60.
26. KirpotinDB,DrummondDC,ShaoY,ShalabyMR,HongK,NielsenUB,et al. Antibody targeting of long-circulating lipidic nanoparticles doesnot increase tumor localization but does increase internalization inanimal models. Cancer Res 2006;66:6732–40.
27. Drummond DC, Meyer O, Hong K, Kirpotin DB, Papahadjopoulos D.Optimizing liposomes for delivery of chemotherapeutic agents to solidtumors. Pharmacol Rev 1999;51:691–743.
28. Nielsen UB, Kirpotin DB, Pickering EM, Hong K, Park JW, RefaatShalabyM, et al. Therapeutic efficacy of anti-ErbB2 immunoliposomestargeted by a phage antibody selected for cellular endocytosis. Bio-chim Biophys Acta 2002;1591:109–18.
29. Park JW, Hong K, Kirpotin DB, Colbern G, Shalaby R, Baselga J, et al.Anti-HER2 immunoliposomes: enhanced efficacy attributable to tar-geted delivery. Clin Cancer Res 2002;8:1172–81.
30. Nellis DF, Giardina SL, Janini GM, Shenoy SR, Marks JD, Tsai R, et al.Preclinical manufacture of anti-HER2 liposome-inserting, scFv-PEG-lipid conjugate. 2. Conjugate micelle identity, purity, stability, andpotency analysis. Biotechnol Prog 2005;21:221–32.
31. Nellis DF, Ekstrom DL, Kirpotin DB, Zhu J, Andersson R, Broadt TL,et al. Preclinical manufacture of an anti-HER2 scFv-PEG-DSPE, lipo-some-inserting conjugate. 1. Gram-scale production and purification.Biotechnol Prog 2005;21:205–20.
32. Millard BL, Niepel M, Menden MP, Muhlich JL, Sorger PK. Adaptiveinformatics for multifactorial and high-content biological data. NatMethods 2011;8:487–93.
33. Hendriks BS, Opresko LK,Wiley HS, Lauffenburger D. Coregulation ofepidermal growth factor receptor/human epidermal growth factorreceptor 2 (HER2) levels and locations: quantitative analysis of HER2overexpression effects. Cancer Res 2003;63:1130–7.
34. Eytan GD, Kuchel PW. Mechanism of action of P-glycoprotein inrelation to passive membrane permeation. Int Rev Cytol 1999;190:175–250.
35. Hendriks BS, Reynolds JG, Klinz SG, Geretti E, Lee H, Leonard SC,et al. Multi-scale kinetic modeling of liposomal doxorubicin deliveryquantifies the role of tumor and drug-specific parameters in localdelivery to tumors. Clin Pharmacol: Pharmacometrics & Sys Pharma-col 2012;1:e15.
36. Wickham TJ, Granados RR, Wood HA, Hammer DA, Shuler ML.General analysis of receptor-mediated viral attachment to cell sur-faces. Biophys J 1990;58:1501–16.
37. Austin CD, De Maziere AM, Pisacane PI, van Dijk SM, Eigenbrot C,Sliwkowski MX, et al. Endocytosis and sorting of ErbB2 and the site of
action of cancer therapeutics trastuzumabandgeldanamycin.Mol BiolCell 2004;15:5268–82.
38. Alcantara CS, Yang CH, Steiner TS, Barrett LJ, Lima AA, ChappellCL, et al. Interleukin-8, tumor necrosis factor-alpha, and lactoferrinin immunocompetent hosts with experimental and Brazilian childrenwith acquired cryptosporidiosis. Am J Trop Med Hyg 2003;68:325–8.
39. Kinoshita A, Kobayashi D, Hibino Y, Isago T, Uchino K, Yagi K, et al.Regulation of CMV promoter-driven exogenous gene expression withdoxorubicin in genetically modified cells. J Pharm Pharmacol 2008;60:1659–65.
40. Kim KI, Kang JH, Chung JK, Lee YJ, Jeong JM, Lee DS, et al.Doxorubicin enhances the expression of transgene under control ofthe CMV promoter in anaplastic thyroid carcinoma cells. J Nucl Med2007;48:1553–61.
41. Nagy P, Friedlander E, Tanner M, Kapanen AI, Carraway KL, Isola J,et al. Decreased accessibility and lack of activation of ErbB2 in JIMT-1,a herceptin-resistant, MUC4-expressing breast cancer cell line.Cancer Res 2005;65:473–82.
42. Li Y, Zhang Q, Tian R, Wang Q, Zhao JJ, Iglehart JD, et al.Lysosomal transmembrane protein LAPTM4B promotes autophagyand tolerance to metabolic stress in cancer cells. Cancer Res 2011;71:7481–9.
43. Kirpotin D, Park JW, Hong K, Zalipsky S, Li WL, Carter P, et al.Sterically stabilized anti-HER2 immunoliposomes: design and tar-geting to human breast cancer cells in vitro. Biochemistry 1997;36:66–75.
44. Park JW, Hong K, Carter P, Asgari H, Guo LY, Keller GA, et al.Development of anti-p185HER2 immunoliposomes for cancer thera-py. Proc Natl Acad Sci U S A 1995;92:1327–31.
45. Hertel PB, TuD, Ejlertsen B, JensenMB,Balslev E, JiangS, et al. TIMP-1 in combination with HER2 and TOP2A for prediction of benefit fromadjuvant anthracyclines in high-risk breast cancer patients. BreastCancer Res Treat 2012;132:225–34.
46. Ejlertsen B, Jensen MB, Nielsen KV, Balslev E, Rasmussen BB, Will-emoe GL, et al. HER2, TOP2A, and TIMP-1 and responsiveness toadjuvant anthracycline-containing chemotherapy in high-risk breastcancer patients. J Clin Oncol 2010;28:984–90.
47. Bezler M, Hengstler JG, Ullrich A. Inhibition of doxorubicin-inducedHER3–PI3K–AKT signalling enhances apoptosis of ovarian cancercells. Mol Oncol 2012;6:516–29.
48. Li Y, Zou L, Li Q, Haibe-Kains B, TianR, Desmedt C, et al. Amplificationof LAPTM4BandYWHAZcontributes to chemotherapy resistance andrecurrence of breast cancer. Nat Med 2010;16:214–8.
49. OnsumM, Geretti E, Paragas V, Kudla A,Moulis S, Luus L, et al. Singlecell quantitative HER2 measurement in patient samples identifiesheterogeneity and distinct sub-groups within traditionally-definedHER2-positive patients. Am J Pathol. In press, 2013.
Hendriks et al.
Mol Cancer Ther; 12(9) September 2013 Molecular Cancer Therapeutics1828
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180

Correction
Correction: Impact of Tumor HER2/ERBB2Expression Level on HER2-Targeted LiposomalDoxorubicin-Mediated Drug Delivery: MultipleLow-Affinity Interactions Lead to a ThresholdEffect
In this article (Mol Cancer Ther 2013;12:1816–28), which appeared in theSeptember 2013 issue of Molecular Cancer Therapeutics (1), the authors regretthat the article title was incorrect.
The authors forgot tomention the company code ofHER2-targeted liposomaldoxorubicin (MM-302) for reference. The companycode is important for otherinvestigators to link this workwith other ongoing clinical trials that are usingMM-302. The publisher recommended that correcting the title was the mostappropriate means for making this link. The correct title is given below.
"Impact of Tumor HER2/ERBB2 Expression Level on HER2-TargetedLiposomal Doxorubicin (MM-302)-Mediated Drug Delivery: MultipleLow-Affinity Interactions Lead to a Threshold Effect".
Reference1. Hendriks BS, Klinz SG, Reynolds JG, Espelin CW, Gaddy DF, Wickham TJ. Impact of tumor
HER2/ERBB2expression level onHER2-targeted liposomal doxorubicin-mediateddrugdelivery:multiple low-affinity interactions lead to a threshold effect. Mol Cancer Ther 2013;12:1816–28.
Published online November 12, 2013.doi: 10.1158/1535-7163.MCT-13-0885�2013 American Association for Cancer Research.
MolecularCancer
Therapeutics
Mol Cancer Ther; 12(12) December 20132962

2013;12:1816-1828. Published OnlineFirst May 30, 2013.Mol Cancer Ther Bart S. Hendriks, Stephan G. Klinz, Joseph G. Reynolds, et al. Low-Affinity Interactions Lead to a Threshold EffectLiposomal Doxorubicin-Mediated Drug Delivery: Multiple Impact of Tumor HER2/ERBB2 Expression Level on HER2-Targeted
Updated version
10.1158/1535-7163.MCT-13-0180doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2013/05/30/1535-7163.MCT-13-0180.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/12/9/1816.full#ref-list-1
This article cites 45 articles, 18 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/12/9/1816.full#related-urls
This article has been cited by 4 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/12/9/1816To request permission to re-use all or part of this article, use this link
on January 18, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 30, 2013; DOI: 10.1158/1535-7163.MCT-13-0180





























