Embed Size (px)
Citation preview

of April 2, 2017.This information is current as
MucosaCell Subset Function in the Intestinal on DendriticToxoplasma gondiiImpact of
Sara B. Cohen and Eric Y. Denkers
http://www.jimmunol.org/content/195/6/2754doi: 10.4049/jimmunol.1501137August 2015;
2015; 195:2754-2762; Prepublished online 17J Immunol
MaterialSupplementary
7.DCSupplementalhttp://www.jimmunol.org/content/suppl/2015/08/16/jimmunol.150113
Referenceshttp://www.jimmunol.org/content/195/6/2754.full#ref-list-1
, 29 of which you can access for free at: cites 59 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2015 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Impact of Toxoplasma gondii on Dendritic Cell SubsetFunction in the Intestinal Mucosa
Sara B. Cohen1 and Eric Y. Denkers
The function of mucosal dendritic cell (DC) subsets in immunity and inflammation is not well understood. In this study, we define
four DC subsets present within the lamina propria and mesenteric lymph node compartments based on expression of CD103 and
CD11b. Using IL-12p40 YFP (Yet40) reporter mice, we show that CD103+CD11b2 mucosal DCs are primary in vivo sources of IL-
12p40; we also identified CD1032CD11b2 mucosal DCs as a novel population producing this cytokine. Infection was preferentially
found in CD11b+ DCs that were negative for CD103. Lamina propria DCs containing parasites were negative for IL-12p40.
Instead, production of the cytokine was strictly a property of noninfected cells. We also show that vitamin A metabolism, as
measured by ALDH activity, was preferentially found in CD103+CD11b+ DC and was strongly downregulated in all mucosal DC
subsets during infection. Finally, overall apoptosis of lamina propria DC subsets was increased during infection. Combined, these
results highlight the ability of intestinal Toxoplasma infection to alter mucosal DC activity at both the whole population level and
at the level of individual subsets. The Journal of Immunology, 2015, 195: 2754–2762.
The intracellular protozoan Toxoplasma gondii infects∼20–50% of the U.S. population and is one of the mostcommon parasitic infections worldwide. After an acute
disseminating phase that originates from infection in the intestinalmucosa, long-term latency is established in tissues of the skeletalmuscle and CNS (1). The parasite is normally asymptomatic, but itcan reactivate, causing life-threatening disease in immunocompro-mised populations (including AIDS patients) (2). Congenital in-fection with Toxoplasma can cause catastrophic birth defects (3).Toxoplasma is currently the second most frequent cause of deathsfrom foodborne infections in the United States (4).Investigation of the host defense response following oral Toxo-
plasma inoculation has yielded many insights into function anddysfunction in the intestinal mucosal immune system (5, 6). Forexample, under low-dose infection conditions, the parasite triggersa protective Th1 response mediated by inflammatory monocytes inthe intestinal mucosa (7, 8). This response enables host survival,parasite dissemination, and establishment of latent infection. High-dose infection in the intestine triggers proinflammatory intestinalpathology from which mice do not recover. In this case, intestinaldamage is associated with a collapsing regulatory T cell (Treg) pop-ulation and epithelial lesions, concomitant with overgrowth andtranslocation of gram-negative species (9–11).The parasite is known to influence several lineages of innate
immunity, including dendritic cells (DC) (12). DC critically bridge
innate and adaptive immunity through their exquisite capacity todrive Ag-specific T cell activation and effector subset differentia-tion. Furthermore, DC are central players in determining toleranceversus immunity during inflammation and infection (13). For thecase of Toxoplasma, DC are obligatory for IL-12–dependent Th1induction and resultant host resistance to T. gondii (14–16).DC are understood to be a heterogeneous population with dis-
tinct surface markers, transcription factor requirements, and func-tions (17, 18). All DC originate from a common bone marrowprogenitor, but they subsequently differentiate into distinct subsets,includingmonocyte-derivedDC, conventionalDC, and plasmacytoidDC. Several key cytokines and transcription factors have been im-plicated in controlling DC developmental pathways, and recent genemapping studies have begun to elucidate the order in which thesefactors become expressed (19). For example, through the activity ofb-catenin, transcription factor IRF8 is involved in the generation ofsplenic CD8a+ and CD11b–CD103+ tissue DC, whereas IRF4 is im-portant for the differentiation of CD11b+CD103+ DC in the intesti-nal lamina propria (LP) (16, 20, 21). The transcription factor Zbtb46was recently identified as essential in generation of all classical DClineages (22, 23). Elegant studies are beginning to identify functionaldifferences among DC subsets. Splenic CD8+ DC appear specializedin cross-presentation and production of IL-12 during infections withmicrobes, including T. gondii (15, 24). CD103+ tissue DC possess theuniqueability to induce tolerogenicTregs (25,26).Nevertheless, thereis still much to be understood about howDC subsets differ in functionunder steady-state conditions, and, importantly, how this changes inresponse to infection and inflammation.In this study, we investigated the behavior of distinct subsets of
mucosal DC following oral inoculation with Toxoplasma. We ana-lyzed several distinct parameters including infection status, IL-12p40production, activation state, and retinoic acid production in intestinalDC subsets before and after infection. Our results indicate dynamicchanges brought about by Toxoplasma infection that are DC subsetspecific and that alter the DC compartment as a whole.
Materials and MethodsEthics statement
All experiments in this study were performed strictly according tothe recommendations of the Guide for the Care and Use of Laboratory
Department of Microbiology and Immunology, College of Veterinary Medicine,Cornell University, Ithaca, NY 14853
1Current address: Center for Infectious Disease Research, Seattle, WA.
Received for publication May 18, 2015. Accepted for publication July 20, 2015.
This work was supported by the National Institutes of Health–National Institute ofAllergy and Infectious Diseases Grant AI109061 (to E.Y.D.).
Address correspondence and reprint requests to Dr. Eric Y. Denkers, Cornell Uni-versity, College of Veterinary Medicine, Ithaca, NY 14853-6401. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: ALDH, aldehyde dehydrogenase; BGS, bovinegrowth serum; DC, dendritic cell; LP, lamina propria; MLN, mesenteric lymph node;PMN, polymorphonuclear leukocyte; Pru-RFP, Prugniaud-RFP OVA; SI, small intes-tine; STAg, soluble tachyzoite Ag; Treg, regulatory T cell.
Copyright� 2015 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/15/$25.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1501137
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

Animals of the National Institutes of Health. The protocols were approvedby the Institutional Animal Care and Use Committee at Cornell University(permit number 1995-0057). All efforts were made to minimize animalsuffering during the course of these studies.
Mice and infections
Female SwissWebster mice (6–8 wk old) were purchased from The JacksonLaboratory (Bar Harbor, ME), and female C57BL/6 were purchased fromTaconic Farms (Germantown, NY). C.129S4(B6)-Il12btm1Lky/J (IL-12p40-YFP) mice were obtained from The Jackson Laboratory and maintained asa breeding colony at Cornell University College of Veterinary Medicine.Infections were initiated in 8–12-wk-oldmice by oral inoculation of 30 cystsof the T. gondii type II ME49 strain or 40 cysts of the type II Prugniaud-RFP OVA (Pru-RFP) strain. Cysts were generated by an initial i.p. injectionof 100 tachyzoites into Swiss Webster mice to establish chronic infection.Cysts were subsequently harvested from chronically infected SwissWebstermice by homogenizing whole brain in PBS.
Preparation and purification of leukocytes
LP leukocytes were prepared as described previously (7). Small intestines wereexcised, flushed with PBS, opened longitudinally, and scraped with a bluntscalpel. The tissuewas thenminced into 1-cm pieces, resuspended in ice-coldHBSS supplemented with 5 mM EDTA and 10 mMHEPES, and shaken twiceat 37˚C for 15 min, removing the cloudy supernatant after each incubation. Thetissue was then resuspended in prewarmed DMEM (supplemented with 5%bovinegrowth serum [BGS] and250ml collagenase (50mg/ml); Sigma-Aldrich,St. Louis, MO) and rotated at 150 rpm for 45 min. The resulting suspensionwas passed through a 40-mm filter, washed, and maintained on ice until use.Mesenteric lymph nodes were minced in PBS, cultured with 0.1 mg/ml LiberaseTL (Roche Diagnostics, Indianapolis, IN) for 20 min at 37˚C, passed througha 40-mm filter, and washed to create a single-cell suspension. Splenocytesingle-cell suspensions were prepared by crushing spleens through a 40-mmfilter and lysing RBCs with ammonium-chloride-potassium buffer.
Flow cytometry
Single-cell suspensions were washed in PBS prior to resuspension inZombie Aqua viability dye (BioLegend, San Diego, CA) for 15 min at roomtemperature to exclude dead cells. Primary Abs (anti-CD11c eFluor610,anti-CD8a Pacific Blue or allophycocyanin-Cy7, anti-CD11b allophycocyanin-Cy7, anti-CD103 BV421, anti-MHCII Alexa700 or FITC, anti-CD64allophycocyanin, anti-CD45 PE-Cy7, Ly6C BV421, 1A8 PE-Cy7) re-suspended in ice-cold FACS buffer (1% BSA/0.01% NaN3 in PBS) wereadded directly to the cells for 30 min. Pru-RFP parasites were detected inthe PE channel. IL-12p40-YFP was detected in the FITC channel. For in-tracellular staining, cells were fixed using the Foxp3/transcription factorstaining kit fixative (eBioscience, San Diego, CA) and subsequently incu-bated with primary Abs resuspended in the Foxp3/transcription factor per-meabilization buffer (eBioscience). Abs used for intracellular cytokine stainingincluded anti-IRF8 PerCP and anti-IRF4 e450 (eBioscience). All sampleswere run on an LSRII flow cytometer (BD Biosciences, San Jose, CA), andthe data were analyzed using FlowJo software (FlowJo, Ashland, OR).
Ex vivo soluble tachyzoite lysate stimulation
LP leukocytes were harvested into a single-cell suspension from naiveYet40 reporter mice and stimulated ex vivowith 50mg/ml soluble tachyzoitelysate Ag (STAg). Cells were incubated at 37˚C for 4.5 h, and IL-12p40expression was measured by upregulation of YFP.
Retinoic acid metabolism measurement
To measure retinoic acid metabolism, an AldeFluor assay kit was usedaccording to the manufacturer’s instructions (StemCell Technologies,Vancouver, BC, Canada). Cells were resuspended in the provided assaybuffer, and 5 ml AldeFluor reagent was added with or without 5 ml of theDEAB inhibitor. The cells were then incubated for 45 min at 37˚C, washedwith assay buffer, and then incubated in primary Abs resuspended in assaybuffer for 20 min at 4˚C. Cells were washed once more in assay bufferand then analyzed with a LSRII flow cytometer (BD Biosciences).
Annexin labeling
For Annexin V labeling, cells were stained following the manufacturer’sinstructions for the Annexin FITC kit (eBioscience). Cells were washed in13 binding buffer, resuspended in 100ml binding buffer, and 5ml AnnexinV
FITC was then added. Cells were incubated for 10–15 min at room temper-ature and washed with binding buffer before analysis with an LSRII flowcytometer (BD Biosciences).
LP DC sorting
LP leukocytes were subjected to a Pan-DCEnrichment negative selection kit(EasySep)with 3mgbiotinylated anti-Epcam (eBioscience) added.Resultingenriched cells were resuspended in 50 ml Aqua Zombie viability dye (Bio-Legend) for 15 min at room temperature. Next, 500ml of a DCAbmixture inMACS buffer (5ml each: anti-CD45 PE-Cy7, anti-MHCII FITC, anti-CD11ce610, anti-CD64 allophycocyanin, anti-CD11b allophycocyanin-Cy7, anti-CD103 BV421) was added. Cells were incubated for 30 min at 4˚C, washedin MACS buffer, and passed through a filter-top FACS tube prior to sortingwith a FACSAria (BD) for LP-DC populations I–IV. Cells were sortedinto DMEM supplemented with 20% BGS, 25 mM HEPES, and 2 mMEDTA and cultured in complete DMEM (DMEM supplemented with 10%BGS, 1 mM sodium pyruvate, 0.1 mM nonessential amino acids, 0.05 mMb-mercaptoethanol, 300 U/ml penicillin, 300 mg/ml streptomycin, and30 mMHEPES) with or without STAg (50 mg/ml) stimulation for 24 h. IL-12p40 was measured from the supernatants using an in-house ELISA (27).
Statistical analyses
Statistical significance between groups was assessed using Student t test.Differences between three or more groups were determined using one-wayANOVAwith Neuman–Keuls posttest.
FIGURE 1. Small intestinal LP DC populations are stable during oral
Toxoplasma infection. (A) WT mice were orally infected with 30 ME49 Toxo-
plasma cysts, and live CD45+ cells from the SI-LP of naive and Toxoplasma-
infected mice were assessed for MHCII and CD11c expression. Total num-
bers of MHCII+CD11c+ cells were quantified (bar graph, n = 8 mice/group).
(B) CD64 expression was measured on MHCII+CD11c+ cells harvested from
naive and infected SI-LP to distinguish monocytic cells (CD64+) from bona-
fide DC (CD642). In the bar graphs, percentages of each group were con-
verted to absolute numbers (n = 8 mice/group). (C) After gating on bona fide
DC phenotype (Live+CD45+MHCII+CD11c+CD642) cells were assessed
for CD103 and CD11b expression to distinguish subpopulations of SI-LP
DC (designated populations I–IV). The bar graph shows average percent-
ages over multiple mice (n = 9 mice/group). (D) Total numbers of inflam-
matory monocytes (Ly6C+CD11b+) were quantified in the SI-LP of naive
and day 7–infected mice (naive, n = 3 mice; infected, n = 6 mice). Repre-
sentative data from individual mice and the means and SE for multiple mice
are shown.Data are representative of at least three experiments. ***p,0.001.
The Journal of Immunology 2755
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
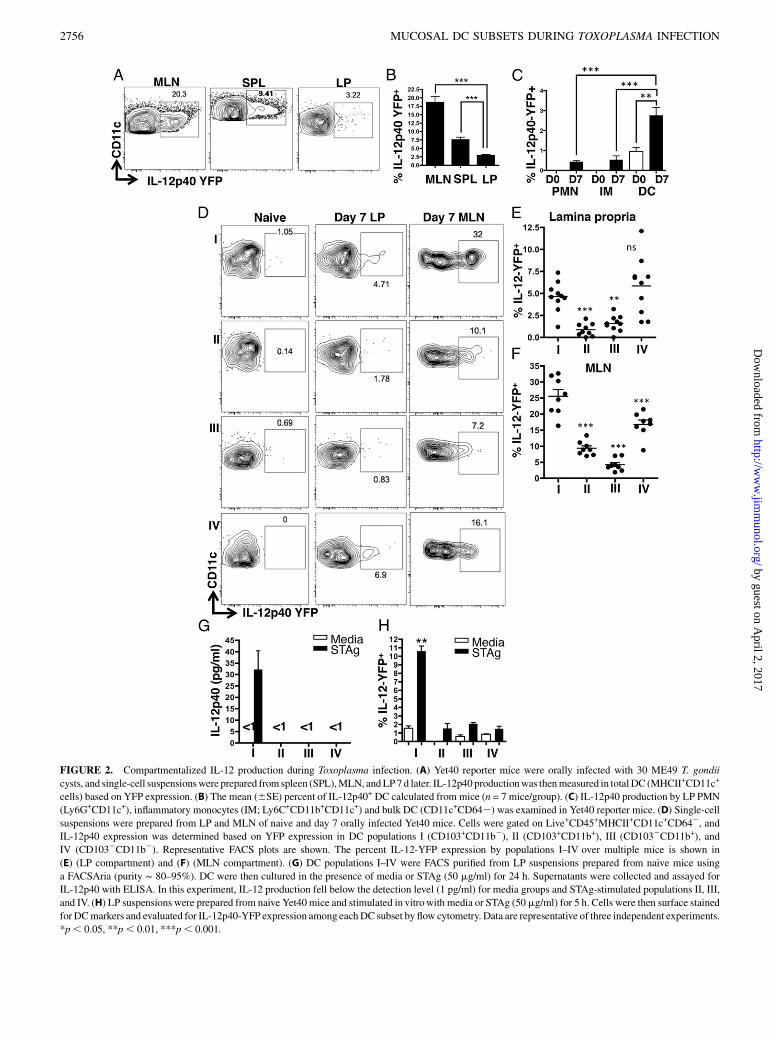
FIGURE 2. Compartmentalized IL-12 production during Toxoplasma infection. (A) Yet40 reporter mice were orally infected with 30 ME49 T. gondii
cysts, and single-cell suspensionswere prepared from spleen (SPL),MLN, andLP7 d later. IL-12p40 productionwas thenmeasured in totalDC (MHCII+CD11c+
cells) based on YFP expression. (B) The mean (6SE) percent of IL-12p40+ DC calculated from mice (n = 7 mice/group). (C) IL-12p40 production by LP PMN
(Ly6G+CD11c+), inflammatory monocytes (IM; Ly6C+CD11b+CD11c+) and bulk DC (CD11c+CD642) was examined in Yet40 reporter mice. (D) Single-cell
suspensions were prepared from LP and MLN of naive and day 7 orally infected Yet40 mice. Cells were gated on Live+CD45+MHCII+CD11c+CD642, and
IL-12p40 expression was determined based on YFP expression in DC populations I (CD103+CD11b2), II (CD103+CD11b+), III (CD1032CD11b+), and
IV (CD1032CD11b2). Representative FACS plots are shown. The percent IL-12-YFP expression by populations I–IV over multiple mice is shown in
(E) (LP compartment) and (F) (MLN compartment). (G) DC populations I–IV were FACS purified from LP suspensions prepared from naive mice using
a FACSAria (purity ∼ 80–95%). DC were then cultured in the presence of media or STAg (50 mg/ml) for 24 h. Supernatants were collected and assayed for
IL-12p40 with ELISA. In this experiment, IL-12 production fell below the detection level (1 pg/ml) for media groups and STAg-stimulated populations II, III,
and IV. (H) LP suspensions were prepared from naive Yet40mice and stimulated in vitrowith media or STAg (50mg/ml) for 5 h. Cells were then surface stained
for DCmarkers and evaluated for IL-12p40-YFP expression among eachDC subset by flow cytometry. Data are representative of three independent experiments.
*p, 0.05, **p, 0.01, ***p, 0.001.
2756 MUCOSAL DC SUBSETS DURING TOXOPLASMA INFECTION
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

ResultsIntestinal monocytes rapidly expand while DC numbers remainstable during oral Toxoplasma gondii infection
To understand the dynamics of intestinal monocytes/DC followingoral inoculation with T. gondii, levels of CD11c+MHCII+ cells werequantified from the small intestinal LP of naive and mice infectedon day 7. Naive mice possessed a relatively small population ofCD11c+MHCII+ cells, but that cell population significantly ex-panded in the intestines of infected mice (Fig. 1A). Myeloid cellsin the intestine are composed of CD64+ monocytes–macrophagesand CD642 DC. The CD11c+MHCII+ cells from naive mice wereprimarilyCD642, suggesting identitywithDC (Fig. 1B).However,the vast majority (∼80–90%) were CD64+ in infected mice, sug-gesting a monocyte–macrophage origin (Fig. 1B). Indeed, the totalnumber of CD642 bona fide intestinal DC was unchanged over thecourse of infection, whereas there was a clear expansion of CD64+
monocytes–macrophages in the LP (Fig. 1B, bar graphs).To assess whether DC were altered by infection at the level of
individual subsets, DC (defined as MHCII+CD11c+CD642) fromnaive and infected small intestinal (SI)-LP were classified into fourgroups (designated I–IV hereafter) based on expression of CD103and CD11b (28) (Fig. 1D). Similar to the unchanged total numbersof DC, numbers of individual DC subsets also remained stable dur-ing infection (Fig. 1C). These data suggest that T. gondii at 7 d post-infection does not influence levels of DC at the global or individualsubset level, whereas monocytes are rapidly expanded by infection.Inflammatory monocytes, which coexpress Ly6C and CD11b,
contribute to host defense in the intestinal mucosa following Toxo-plasma infection (7, 8). Indeed, we found a pronounced increase inLy6C+CD11b+ SI-LP cells triggered by T. gondii (Fig. 1D). Nev-ertheless, we note that total numbers of Ly6C+CD11b+ inflamma-tory monocytes expanded in response to infection (∼1.5 3 105)was significantly less than the increase in total numbers of MHCII+
CD11c+CD64+ monocytes–macrophages (∼9 3 105), suggestingpresence of other monocyte–macrophage lineage cell types.
IL-12p40 is produced primarily by CD103+CD11b2 andCD1032CD11b2 DC subsets
IL-12p40 production is a hallmark of T. gondii infection, and splenicCD8a+ DC are an essential source during i.p. infection (15, 16, 29).To address the functional effects of T. gondii infection on IL-12induction by specific intestinal DC subsets, IL-12p40-YFP re-porter mice were orally infected, and YFP expression was exam-ined in DC from the SI-LP, mesenteric lymph nodes (MLN), andspleen. The Day 7 IL-12p40 response by DC from the MLN andspleen was relatively robust, with 15–20% of MLN DC and 8–10%of spleen DC expressing the cytokine (Fig. 2A, 2B). Despite theposition of SI-LP DC at the frontline of infection, their IL-12p40expression was strikingly lower (∼3%) than that observed in the
MLNor spleen, suggesting decreased capacity byLP-residentDC toproduce IL-12p40 during infection.We then examined the LP compartment, focusing on IL-12p40
production by polymorphonuclear leukocytes (PMN), inflamma-tory monocytes, and total DC. Each of these subsets is known toproduce IL-12 during T. gondii infection (7, 30). Nevertheless, theDC population was significantly enhanced in IL-12p40 productionrelative to PMN and inflammatory monocytes (Fig. 2C).We determined IL-12p40 expression in DC subsets I (CD103+
CD11b2), II (CD103+CD11b+), III (CD103-CD11b+), and IV(CD1032CD11b2). As expected, in noninfected mice, IL-12p40expression was minimal (1% or less) in each SI-LP DC subset(Fig. 2D). In day 7 SI-LP, we observed IL-12p40 induction thatwas restricted to populations I (CD103+CD11b2) and IV (CD1032
CD11b2) DC (Fig. 2D, 2E). As expected, and based on Fig. 2Aand 2B, IL-12p40 responsiveness was greater overall in MLN DCsubsets than in SI-LP DC. As in the SI-LP, DC populations I and IVproduced the greatest amount of IL-12p40 (Fig. 2D, 2F).The phenotype of DC in the MLN can be resident (CD11chigh
MHCint) or migratory (CD11cintMHCIIhigh) (31). Using this dis-crimination, we assessed IL-12p40 expression among MLN DCpopulations I–IV (Supplemental Fig. 1). In cells from naive IL-12p40reporter mice, migratory DC populations I (CD103+CD11b2) andIV (CD1032CD11b2) possessed a phenotype that was positivefor IL-12p40 production, possibly indicating steady-state levelsof stimulation of DC arriving from the gut. In contrast, the residentMLN DC in noninfected mice did not express IL-12p40. In in-fected mice, resident and migratory DC subsets I and IV ex-pressed IL-12p40 (Supplemental Fig. 1).We next assessed whether similar patterns of IL-12p40 produc-
tion were observed by SI-LP DC subsets in vitro. The four popula-tions of SI-LP DC were sort-purified from naive mice and stimulatedex vivo with STAg, a sonicated extract derived from T. gondii tachy-zoites. As expected, population I (CD103+CD11b2) DC secretedIL-12p40 after STAg stimulation as determined with ELISA. Sur-prisingly, and in contrast to the analysis of in situ IL-12p40 produc-tion (Fig. 2D, 2E), no detectable IL-12p40 was found in populationIV nor in any of the other DC populations (Fig. 2G). To determinewhether this result could be replicated using Yet40 IL-12 reporterDC in vitro, LP cells from naive Yet40 mice were stimulated withSTAg, and levels of YFP were measured using flow cytometry. Con-sistent with the sort experiment, IL-12p40-YFP was restricted to pop-ulation I in vitro, and population IV failed to express cytokine (Fig.2H). Therefore, whereas CD103+CD11b2 DC are the primary pro-ducers of IL-12p40 in response to T. gondii, the CD1032CD11b2 DCsubset is also capable of upregulating this cytokine during in vivo in-fection, although this response is not apparent during invitro culture.We also examined splenic IL-12p40 production in infected IL-
12p40-YFP mice (Fig. 3). Both CD8a+ and CD11b+ splenic DC in-
FIGURE 3. CD8a+ DC are the primary IL-12
producers in the spleen during Toxoplasma infec-
tion. Yet40 reporter mice were orally infected with
30 ME49 T. gondii cysts, and splenocytes were har-
vested 7 d later. (A) Splenic DC (MHCII+CD11c+)
were assessed for expression of CD8a+ and CD11b+
markers. (B) CD8a+ and CD11b+ DC from naive
and Day 7-infected Yet40 reporter mice were eval-
uated for IL-12p40-YFP expression using flow
cytometry. Shown are representative FACS plots
for each subset and the means (6SE) of percent
IL-12p40+ DC in naive and infected mice (naive,
n = 2 mice; infected, n = 4). Data are representative
of three experiments. **p, 0.01, ****p, 0.001.
The Journal of Immunology 2757
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
creased IL-12p40 expression after infection. Nevertheless, theCD8a+ DC subset was the major IL-12p40–positive population,which is in agreement with previous studies by others (15, 16,21, 32). Splenic CD8a+ DC and mucosal population I DC are de-velopmentally related insofar as they depend on transcription fac-tors IRF8 and BATF3 for differentiation, and mice lacking thesemolecules are defective in IL-12 production during Toxoplasmainfection (15, 33). Therefore, our observation that populationI (CD103+CD11b2) DC are major sources of IL-12 in the intestinalmucosa is in agreement with these previous studies.
DC IL-12p40 production correlates with expression of IRF8and inversely correlates with IRF4
Previous studies identified roles for IRF4 and IRF8 in respectivedevelopment of CD103+CD11b+ (population II) and CD103+
CD11b2 (population I) DC (20, 34, 35). It is also known that IRF8in splenic DC controls IL-12 production during T. gondii infection(16, 21). Therefore, it was of interest to determine intracellular ex-pression of IRF8 and IRF4 in DC populations I–IV. Overall, infec-tion had little or no effect on IRF8 (Supplemental Fig. 2A, 2B) orIRF4 (Supplemental Fig. 2C, 2D) levels amongDC subpopulations.However, IRF8 expression was greatest in population I (CD103+
CD11b2) DC (Supplemental Fig. 2B), whereas this DC subsetexpressed the lowest level of IRF4 (Supplemental Fig. 2D). Pop-ulation II (CD103+CD11b+) expressed the highest levels of IRF4with a low amount IRF8. Interestingly, IRF8 and IL-12p40 expres-sion were not strictly correlated, inasmuch as population IV wasa major source of IL-12p40 in SI-LP DC during in vivo infection(Fig. 2E), yet these DC expressed only modest levels of IRF8compared with population I DC (Supplemental Fig. 2A, 2B). We
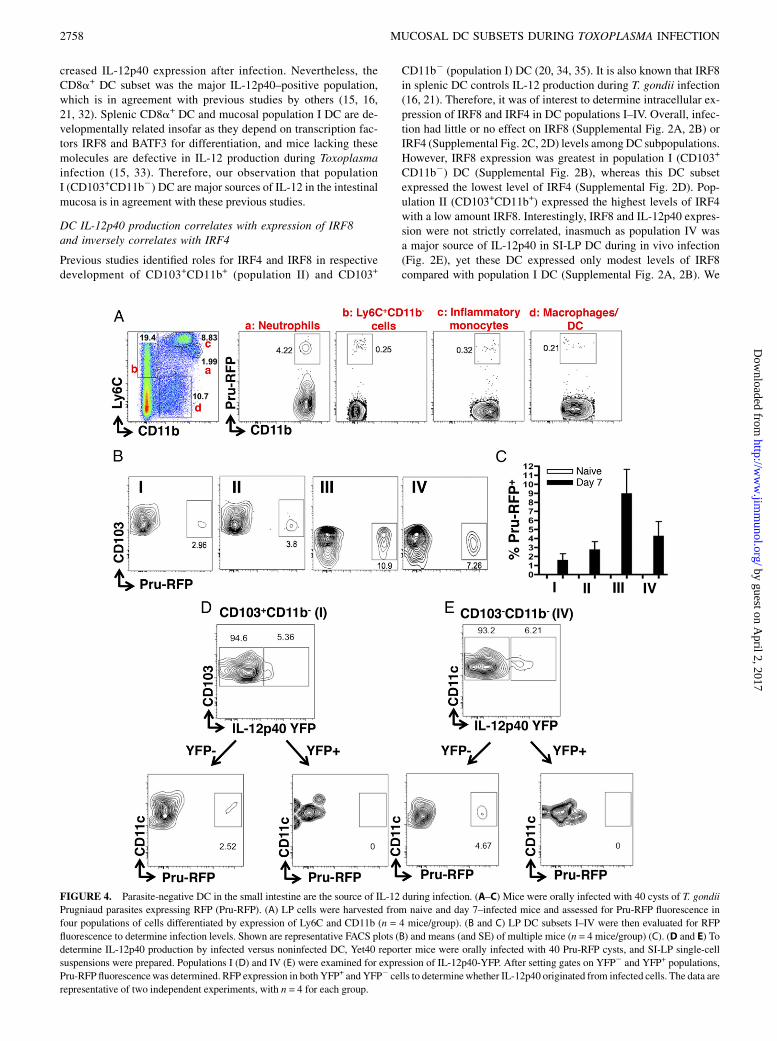
FIGURE 4. Parasite-negative DC in the small intestine are the source of IL-12 during infection. (A–C) Mice were orally infected with 40 cysts of T. gondii
Prugniaud parasites expressing RFP (Pru-RFP). (A) LP cells were harvested from naive and day 7–infected mice and assessed for Pru-RFP fluorescence in
four populations of cells differentiated by expression of Ly6C and CD11b (n = 4 mice/group). (B and C) LP DC subsets I–IV were then evaluated for RFP
fluorescence to determine infection levels. Shown are representative FACS plots (B) and means (and SE) of multiple mice (n = 4 mice/group) (C). (D and E) To
determine IL-12p40 production by infected versus noninfected DC, Yet40 reporter mice were orally infected with 40 Pru-RFP cysts, and SI-LP single-cell
suspensions were prepared. Populations I (D) and IV (E) were examined for expression of IL-12p40-YFP. After setting gates on YFP2 and YFP+ populations,
Pru-RFP fluorescencewas determined. RFP expression in both YFP+ and YFP2 cells to determinewhether IL-12p40 originated from infected cells. The data are
representative of two independent experiments, with n = 4 for each group.
2758 MUCOSAL DC SUBSETS DURING TOXOPLASMA INFECTION
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
also observed relatively heterogeneous IRF4 staining in naivepopulation IV DC, possibly suggesting further complexity in thisDC subpopulation.
IL-12p40 production by SI-LP DC is confined to noninfectedcells
Production of IL-12 is a critical step in establishing immunity toToxoplasma, but it is unclear whether parasite-positive or parasite-negative DC produce IL-12 (36–39). To gain insight into this issue,we inoculated mice with fluorescent Pru-RFP parasites and ex-amined cells in the intestinal LP 7 d later. We first asked whichcells harbored parasites overall based on Ly6C versus CD11b ex-pression. Interestingly, although neutrophils (CD11b+Ly6Clo; pop-ulation a) composed aminor proportionofLP leukocytes (2%), therewas a clear preferential infection of this cell type in that ∼4% har-bored parasites (Fig. 4A).CD11b2Ly6C+ cells (population b), likelyactivated T cells (40), and CD11b+Ly6C+ inflammatory monocytes(population c) both exhibited low levels of infection (0.2–0.3%).The macrophage/DC subset (CD11b+Ly6C2; population d) alsocontained a subpopulation of infected cells (0.2–0.3%).Because of the importance of DC-derived IL-12 during Toxo-
plasma infection, and given the ability of T. gondii to producemolecules triggering this cytokine, we examined infection statusas a potential correlate of IL-12p40 production in each DC subset.As shown in Fig. 4B and 4C, we could readily identify sub-populations of infected DC among each of the four DC subsets.Interestingly, there was a preference for infection in population IIIDC (CD1032CD11b+), with a less striking tropism for infection inpopulation IV DC (CD1032CD11b2). Notably, the most heavilyinfected DC subset (population III) was not a strong producer ofIL-12p40 during infection (Fig. 2E).Population I and IVDC are themajor sources of IL-12p40 among
DC subsets (Fig. 2E); therefore, we specifically examined the cor-relation between infection status and cytokine production in thesesubsets using Pru-RFP infection of IL-12p40-YFP reporter mice. InFig. 4D, we identified ∼5% IL-12p40–expressing population I DC.After gating on YFP2 and YFP+ cells, we found that all Pru-RFP–positive cells were contained within the IL-12p40–negative pop-ulation (Fig. 4D). Similarly, we identified 6% IL-12p40–positivepopulation IV DC (Fig. 4E). After gating on YFP+ and YFP2 cells,there were no detectable parasites within the IL-12–expressingpopulation.We next examined the correlation between infection and IL-12
production in splenic DC. As shown in Fig. 5A, ∼18% of CD11c+
DC expressed IL-12p40. Interestingly, when we gated on YFP2 (noIL-12) and YFP+ (IL-12 expressing) DC, tachyzoite infection wasfound with .10-fold greater proportion in the YFP+ DC subset(Fig. 5A, 5B). This could also be demonstrated using the reverseapproach (Fig. 5C, 5D). Thus, ∼0.3% of splenic DC were parasitePru-RFP-positive; however, when we compared IL-12 expressionin RFP2 and RFP+ DC, ∼14% of the noninfected DC popula-tion expressed IL-12p40, whereas .50% of parasite-positive DCexpressed IL-12p40 (Fig. 5C, 5D). Thus, in the spleen most of theinfected DC also produce IL-12p40, although there are clearlynoninfected DC that produce IL-12p40.
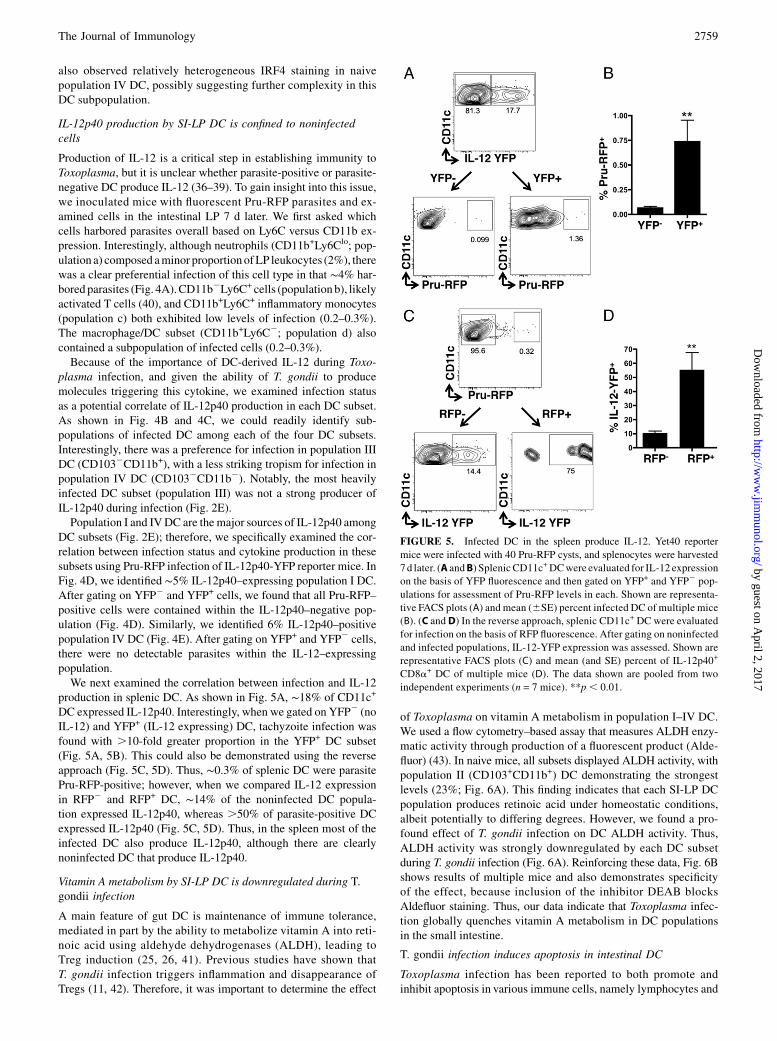
Vitamin A metabolism by SI-LP DC is downregulated during T.gondii infection
A main feature of gut DC is maintenance of immune tolerance,mediated in part by the ability to metabolize vitamin A into reti-noic acid using aldehyde dehydrogenases (ALDH), leading toTreg induction (25, 26, 41). Previous studies have shown thatT. gondii infection triggers inflammation and disappearance ofTregs (11, 42). Therefore, it was important to determine the effect
of Toxoplasma on vitamin A metabolism in population I–IV DC.We used a flow cytometry–based assay that measures ALDH enzy-matic activity through production of a fluorescent product (Alde-fluor) (43). In naive mice, all subsets displayed ALDH activity, withpopulation II (CD103+CD11b+) DC demonstrating the strongestlevels (23%; Fig. 6A). This finding indicates that each SI-LP DCpopulation produces retinoic acid under homeostatic conditions,albeit potentially to differing degrees. However, we found a pro-found effect of T. gondii infection on DC ALDH activity. Thus,ALDH activity was strongly downregulated by each DC subsetduring T. gondii infection (Fig. 6A). Reinforcing these data, Fig. 6Bshows results of multiple mice and also demonstrates specificityof the effect, because inclusion of the inhibitor DEAB blocksAldefluor staining. Thus, our data indicate that Toxoplasma infec-tion globally quenches vitamin A metabolism in DC populationsin the small intestine.
T. gondii infection induces apoptosis in intestinal DC
Toxoplasma infection has been reported to both promote andinhibit apoptosis in various immune cells, namely lymphocytes and
FIGURE 5. Infected DC in the spleen produce IL-12. Yet40 reporter
mice were infected with 40 Pru-RFP cysts, and splenocytes were harvested
7d later. (A andB) SplenicCD11c+DCwere evaluated for IL-12 expression
on the basis of YFP fluorescence and then gated on YFP+ and YFP2 pop-
ulations for assessment of Pru-RFP levels in each. Shown are representa-
tive FACS plots (A) and mean (6SE) percent infected DC of multiple mice
(B). (C and D) In the reverse approach, splenic CD11c+ DC were evaluated
for infection on the basis of RFP fluorescence. After gating on noninfected
and infected populations, IL-12-YFP expression was assessed. Shown are
representative FACS plots (C) and mean (and SE) percent of IL-12p40+
CD8a+ DC of multiple mice (D). The data shown are pooled from two
independent experiments (n = 7 mice). **p , 0.01.
The Journal of Immunology 2759
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
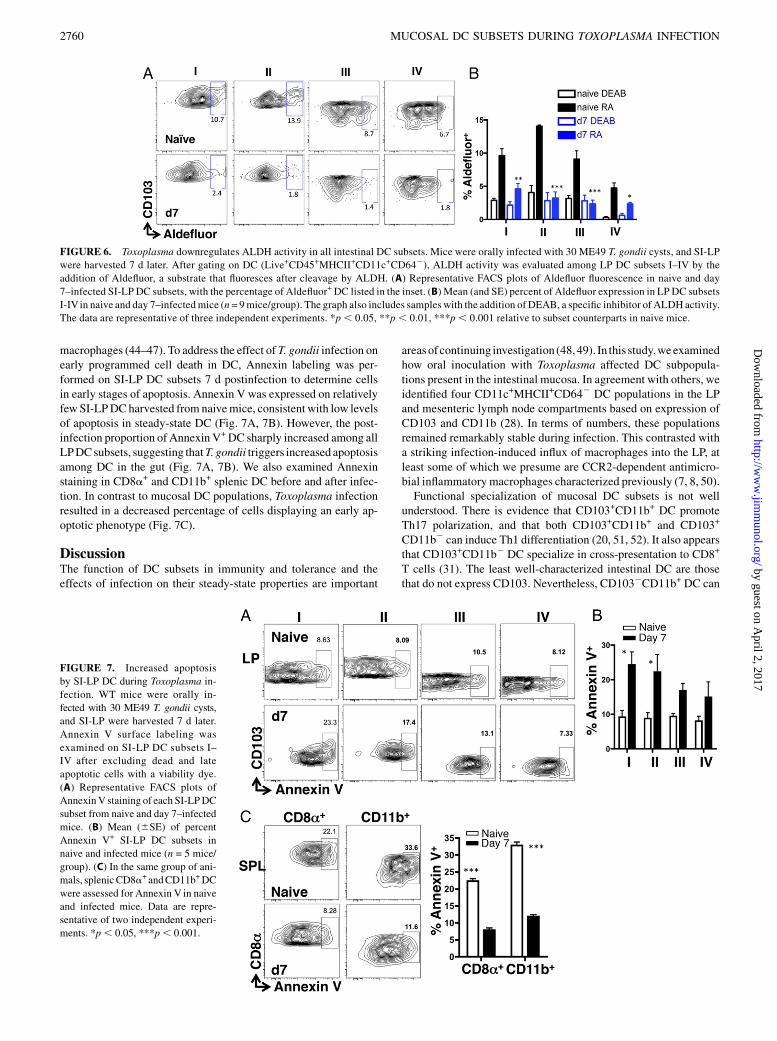
macrophages (44–47). To address the effect of T. gondii infection onearly programmed cell death in DC, Annexin labeling was per-formed on SI-LP DC subsets 7 d postinfection to determine cellsin early stages of apoptosis. Annexin V was expressed on relativelyfewSI-LPDCharvested from naivemice, consistent with low levelsof apoptosis in steady-state DC (Fig. 7A, 7B). However, the post-infection proportion of Annexin V+DC sharply increased among allLPDCsubsets, suggesting thatT. gondii triggers increased apoptosisamong DC in the gut (Fig. 7A, 7B). We also examined Annexinstaining in CD8a+ and CD11b+ splenic DC before and after infec-tion. In contrast to mucosal DC populations, Toxoplasma infectionresulted in a decreased percentage of cells displaying an early ap-optotic phenotype (Fig. 7C).
DiscussionThe function of DC subsets in immunity and tolerance and theeffects of infection on their steady-state properties are important
areasof continuing investigation (48, 49). In this study,weexaminedhow oral inoculation with Toxoplasma affected DC subpopula-tions present in the intestinal mucosa. In agreement with others, weidentified four CD11c+MHCII+CD642 DC populations in the LPand mesenteric lymph node compartments based on expression ofCD103 and CD11b (28). In terms of numbers, these populationsremained remarkably stable during infection. This contrasted witha striking infection-induced influx of macrophages into the LP, atleast some of which we presume are CCR2-dependent antimicro-bial inflammatory macrophages characterized previously (7, 8, 50).Functional specialization of mucosal DC subsets is not well
understood. There is evidence that CD103+CD11b+ DC promoteTh17 polarization, and that both CD103+CD11b+ and CD103+
CD11b2 can induce Th1 differentiation (20, 51, 52). It also appearsthat CD103+CD11b2 DC specialize in cross-presentation to CD8+
T cells (31). The least well-characterized intestinal DC are thosethat do not express CD103. Nevertheless, CD1032CD11b+ DC can
FIGURE 6. Toxoplasma downregulates ALDH activity in all intestinal DC subsets. Mice were orally infected with 30 ME49 T. gondii cysts, and SI-LP
were harvested 7 d later. After gating on DC (Live+CD45+MHCII+CD11c+CD642), ALDH activity was evaluated among LP DC subsets I–IV by the
addition of Aldefluor, a substrate that fluoresces after cleavage by ALDH. (A) Representative FACS plots of Aldefluor fluorescence in naive and day
7–infected SI-LP DC subsets, with the percentage of Aldefluor+ DC listed in the inset. (B) Mean (and SE) percent of Aldefluor expression in LP DC subsets
I-IVin naive and day 7–infectedmice (n = 9mice/group). The graph also includes samples with the addition of DEAB, a specific inhibitor of ALDH activity.
The data are representative of three independent experiments. *p, 0.05, **p , 0.01, ***p , 0.001 relative to subset counterparts in naive mice.
FIGURE 7. Increased apoptosis
by SI-LP DC during Toxoplasma in-
fection. WT mice were orally in-
fected with 30 ME49 T. gondii cysts,
and SI-LP were harvested 7 d later.
Annexin V surface labeling was
examined on SI-LP DC subsets I–
IV after excluding dead and late
apoptotic cells with a viability dye.
(A) Representative FACS plots of
AnnexinV staining of each SI-LPDC
subset from naive and day 7–infected
mice. (B) Mean (6SE) of percent
Annexin V+ SI-LP DC subsets in
naive and infected mice (n = 5 mice/
group). (C) In the same group of ani-
mals, splenicCD8a+ andCD11b+DC
were assessed for Annexin V in naive
and infected mice. Data are repre-
sentative of two independent experi-
ments. *p, 0.05, ***p, 0.001.
2760 MUCOSAL DC SUBSETS DURING TOXOPLASMA INFECTION
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

trigger both IFN-g and IL-17–producing effector T cells. In addi-tion, there is evidence that CD1032CD11b2 DC originate fromgut-associated lymphoid tissue, whereas CD1032CD11b+ may beresident within the intestinal LP (53).DC are critical for IL-12 production and induction of protec-
tive immunity to Toxoplasma (14). In the spleen, CD8a+ DC areamajor source of this cytokine during infection (15). Among theDCsubsets in the intestinal mucosa, two DC subsets (population I,CD103+CD11b2; population IV, CD1032CD11b2) were the majorsources of infection-induced IL-12p40. Population I DC are be-lieved to be developmentally related to splenic CD8aDC, insofar asthey share a requirement for transcription factors Id2, IRF8, andBatf3 for their differentiation (54–56). Furthermore, IRF8 itself isimplicated in parasite driven DC IL-12 production (16, 21). Thus,our finding that mucosal CD103+CD11b2DC produce IL-12p40 inresponse to Toxoplasma is in line with these previous studies.There is currently limited understanding of the function of
CD1032CD11b2 (population IV) DC. Accordingly, it is of interestthat these cells were a source of IL-12p40 during intestinal Toxo-plasma infection. It is possible that this DC subset is somehow re-lated to the population I DC. However, the distinct pattern of IRF8/IRF4 expression in population I versus population IV DC suggestsotherwise. Furthermore, when DC subsets from noninfected micewere subjected to in vitro STAg stimulation, population IV DCfailed to respond with IL-12p40 production in marked contrast topopulation I DC.We propose that population IVDC are triggered toproduce IL-12 during in vivo infection because of the intestinal in-flammatory milieu triggered by the parasite, rather than a directresponse to parasite molecules.Among intestinal DC responding to T. gondii, we determined
that IL-12 production was a response of noninfected rather thaninfected cells. This determination supports a bystander model ofT. gondii–induced IL-12 inwhich cells respond to soluble parasitefactors, parasite debris, or host factors, as suggested by other re-cent studies (57). In contrast, both infected and noninfected DCproduced IL-12p40 in the spleen. In fact, most infected splenicDC were found to be positive for IL-12p40 (Fig. 5). The under-lying basis for these differing responses is not clear, but it is in-teresting to note that GRA15 and GRA24, two Toxoplasma proteinsrecently implicated in IL-12 induction, are directly injected into thecytoplasm of infected host cells (38, 58). In contrast, T. gondii pro-filin, an IL-12-inducing TLR11/12 ligand, is believed to functionwithin endosomal compartments resulting from phagocytosis ofparasite material (16). Accordingly, we hypothesize that IL-12 pro-duction in infected DC could indicate a GRA15/GRA24 dominatedpathway, whereas IL-12 production in parasite negative DC mightreflect a response driven by profilin, or, especially for the case ofpopulation IVDC, a response triggered by host-derived factors.Whythese responsesmight differ could be due to several factors includingdifferences in phagocytic activity or unique patterns of TLR ex-pression among DC subsets. These possibilities are currently underexamination.Retinoic acid has a key role in Foxp3+ Treg generation in the
intestinal mucosa, and CD103+ DC are believed to be an importantsource of this vitamin A metabolite (25, 26). We found that insteady state, the fourDC subsets all produced retinoic acid, althoughlevels differed for each. Nevertheless, we found that the highestlevels of retinoic acid were expressed by the CD11b+CD103+ pop-ulation. Strikingly, there was a profound decrease in retinoic acidexpression following T. gondii infection in each DC subset. Thisfinding is consistentwith the observed collapse in intestinal Treg andemergence of Th1 activity in the residual population that is associ-ated with intestinal infection with Toxoplasma (11). Loss of retinoicacid producing activity in mucosal DCmay underlie the emergence
of fulminant proinflammatory pathology that is triggered by highinfectious doses of T. gondii (6, 59).We found an overall increase in DC subsets in the LP compart-
ment undergoing apoptosis, as defined by Annexin V staining. Thismay be a consequence of the overwhelming proinflammatory cy-tokine response elicited by Toxoplasma in the small intestine andbeyond (60). For example, it has been shown that parasite-triggeredapoptosis in the intestinal mucosa is blocked by treatment with anti–IFN-gmAb (46). It is also possible that apoptosis reflects cytotoxicactivity of CD8+ T lymphocytes and NK cells. In this regard,perforin-triggered egress of parasites from infected cells has beenproposed as a method used by T. gondii to promote rapid dissemi-nation during early infection (61, 62). Conversely, the decrease inDC apoptosis in the spleen after infection could be related to theantiapoptotic effects of the parasite in infected cells (47). Theseissues await further investigation.Infection with Toxoplasma in the intestinal mucosa can activate
innate immune cells and induce highly efficient protective Th1immunity with little overt inflammatory damage (5). At the otherextreme, once an infectious dose threshold is crossed, the parasitetriggers dysbiosis and translocation of luminal bacteria that culmi-nates in fulminant proinflammatory cytokine pathology (6). Un-derstanding how these opposite extreme endpoints are controlledby DC subsets in the intestinal mucosa is an area of ongoing in-vestigation with potential for great translational impact in the clinic.
DisclosuresThe authors have no financial conflicts of interest.
References1. Dubey, J. P. 2013. The history and life-cycle of Toxoplasma gondii. In Toxo-
plasma gondii The model apicomplexan: Perspective and methods, 2nd Ed.,L. M. Weiss, and K. Kim, eds. Academic Press, San Diego, p. 1–17.
2. McLeod, R. M., C. van Tubbergen, J. G. Montoya, and E. Petersen. 2013. Hu-man Toxoplasma infection. In Toxoplasma gondii The Model Apicomplexan:Perspectives and Methods, 2nd Ed., L. M. Weiss, and K. Kim, eds. AcademicPress, San Diego, p. 100–159.
3. Remington, J. S., R. McLeod, P. Thuliez, and G. Desmonts. 2001. Toxo-plasmosis. In Infectious Diseases of the fetus and newborn infant, 3rd Ed.,J. S. Remington, and J. O. Klein, eds. W. B. Saunders Co., Philadelphia,p. 205–346.
4. Scallan, E., R. M. Hoekstra, F. J. Angulo, R. V. Tauxe, M. A. Widdowson,S. L. Roy, J. L. Jones, and P. M. Griffin. 2011. Foodborne illness acquired in theUnited States–major pathogens. Emerg. Infect. Dis. 17: 7–15.
5. Cohen, S. B., and E. Y. Denkers. 2014. Border maneuvers: deployment ofmucosal immune defenses against Toxoplasma gondii. Mucosal Immunol. 7:744–752.
6. Egan, C. E., S. B. Cohen, and E. Y. Denkers. 2012. Insights into inflammatorybowel disease using Toxoplasma gondii as an infectious trigger. Immunol. CellBiol. 90: 668–675.
7. Cohen, S. B., K. J. Maurer, C. E. Egan, S. Oghumu, A. R. Satoskar, andE. Y. Denkers. 2013. CXCR3-dependent CD4⁺ T cells are required to activateinflammatory monocytes for defense against intestinal infection. PLoS Pathog. 9:e1003706.
8. Dunay, I. R., R. A. Damatta, B. Fux, R. Presti, S. Greco, M. Colonna, andL. D. Sibley. 2008. Gr1(+) inflammatory monocytes are required for mucosalresistance to the pathogen Toxoplasma gondii. Immunity 29: 306–317.
9. Craven, M., C. E. Egan, S. E. Dowd, S. P. McDonough, B. Dogan, E. Y. Denkers,D. Bowman, E. J. Scherl, and K. W. Simpson. 2012. Inflammation drives dys-biosis and bacterial invasion in murine models of ileal Crohn’s disease. PLoSOne 7: e41594.
10. Heimesaat, M. M., S. Bereswill, A. Fischer, D. Fuchs, D. Struck, J. Niebergall,H. K. Jahn, I. R. Dunay, A. Moter, D. M. Gescher, et al. 2006. Gram-negativebacteria aggravate murine small intestinal Th1-type immunopathology followingoral infection with Toxoplasma gondii. J. Immunol. 177: 8785–8795.
11. Oldenhove, G., N. Bouladoux, E. A. Wohlfert, J. A. Hall, D. Chou, L. DosSantos, S. O’Brien, R. Blank, E. Lamb, S. Natarajan, et al. 2009. Decrease ofFoxp3+ Treg cell number and acquisition of effector cell phenotype during lethalinfection. Immunity 31: 772–786.
12. Denkers, E. Y., B. A. Butcher, L. Del Rio, and S. Bennouna. 2004. Neutrophils,dendritic cells and Toxoplasma. Int. J. Parasitol. 34: 411–421.
13. Manicassamy, S., and B. Pulendran. 2011. Dendritic cell control of tolerogenicresponses. Immunol. Rev. 241: 206–227.
14. Liu, C. H., Y. T. Fan, A. Dias, L. Esper, R. A. Corn, A. Bafica, F. S. Machado,and J. Aliberti. 2006. Cutting edge: dendritic cells are essential for in vivo IL-12
The Journal of Immunology 2761
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

production and development of resistance against Toxoplasma gondii infection inmice. J. Immunol. 177: 31–35.
15. Mashayekhi, M., M. M. Sandau, I. R. Dunay, E. M. Frickel, A. Khan,R. S. Goldszmid, A. Sher, H. L. Ploegh, T. L. Murphy, L. D. Sibley, andK. M. Murphy. 2011. CD8a(+) dendritic cells are the critical source ofinterleukin-12 that controls acute infection by Toxoplasma gondii tachyzoites.Immunity 35: 249–259.
16. Raetz, M., A. Kibardin, C. R. Sturge, R. Pifer, H. Li, E. Burstein, K. Ozato,S. Larin, and F. Yarovinsky. 2013. Cooperation of TLR12 and TLR11 in theIRF8-dependent IL-12 response to Toxoplasma gondii profilin. J. Immunol. 191:4818–4827.
17. Belz, G. T., and S. L. Nutt. 2012. Transcriptional programming of the dendriticcell network. Nat. Rev. Immunol. 12: 101–113.
18. Satpathy, A. T., X. Wu, J. C. Albring, and K. M. Murphy. 2012. Re(de)fining thedendritic cell lineage. Nat. Immunol. 13: 1145–1154.
19. Miller, J. C., B. D. Brown, T. Shay, E. L. Gautier, V. Jojic, A. Cohain, G. Pandey,M. Leboeuf, K. G. Elpek, J. Helft, et al; Immunological Genome Consortium.2012. Deciphering the transcriptional network of the dendritic cell lineage. Nat.Immunol. 13: 888–899.
20. Persson, E. K., H. Uronen-Hansson, M. Semmrich, A. Rivollier, K. Hagerbrand,J. Marsal, S. Gudjonsson, U. Hakansson, B. Reizis, K. Kotarsky, and W. W. Agace.2013. IRF4 transcription-factor-dependent CD103(+)CD11b(+) dendritic cellsdrive mucosal T helper 17 cell differentiation. Immunity 38: 958–969.
21. Cohen, S. B., N. L. Smith, C. McDougal, M. Pepper, S. Shah, G. S. Yap,H. Acha-Orbea, A. Jiang, B. E. Clausen, B. D. Rudd, and E. Y. Denkers. 2015.Beta-catenin signaling drives differentiation and proinflammatory function ofIRF8-dependent dendritic cells. J. Immunol. 194: 210–222.
22. Meredith, M. M., K. Liu, G. Darrasse-Jeze, A. O. Kamphorst, H. A. Schreiber,P. Guermonprez, J. Idoyaga, C. Cheong, K. H. Yao, R. E. Niec, andM. C. Nussenzweig. 2012. Expression of the zinc finger transcription factor zDC(Zbtb46, Btbd4) defines the classical dendritic cell lineage. J. Exp. Med. 209:1153–1165.
23. Satpathy, A. T., W. Kc, J. C. Albring, B. T. Edelson, N. M. Kretzer,D. Bhattacharya, T. L. Murphy, and K. M. Murphy. 2012. Zbtb46 expressiondistinguishes classical dendritic cells and their committed progenitors from otherimmune lineages. J. Exp. Med. 209: 1135–1152.
24. Shortman, K., and W. R. Heath. 2010. The CD8+ dendritic cell subset. Immunol.Rev. 234: 18–31.
25. Coombes, J. L., K. R. Siddiqui, C. V. Arancibia-Carcamo, J. Hall, C. M. Sun,Y. Belkaid, and F. Powrie. 2007. A functionally specialized population of mu-cosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and ret-inoic acid-dependent mechanism. J. Exp. Med. 204: 1757–1764.
26. Sun, C. M., J. A. Hall, R. B. Blank, N. Bouladoux, M. Oukka, J. R. Mora, andY. Belkaid. 2007. Small intestine lamina propria dendritic cells promote de novogeneration of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 204: 1775–1785.
27. Butcher, B. A., L. Kim, P. F. Johnson, and E. Y. Denkers. 2001. Toxoplasmagondii tachyzoites inhibit proinflammatory cytokine induction in infected mac-rophages by preventing nuclear translocation of the transcription factor NF-kappa B. J. Immunol. 167: 2193–2201.
28. Persson, E. K., C. L. Scott, A. M. Mowat, and W. W. Agace. 2013. Dendritic cellsubsets in the intestinal lamina propria: ontogeny and function. Eur. J. Immunol.43: 3098–3107.
29. Yarovinsky, F., D. Zhang, J. F. Andersen, G. L. Bannenberg, C. N. Serhan,M. S. Hayden, S. Hieny, F. S. Sutterwala, R. A. Flavell, S. Ghosh, and A. Sher.2005. TLR11 activation of dendritic cells by a protozoan profilin-like protein.Science 308: 1626–1629.
30. Bliss, S. K., Y. Zhang, and E. Y. Denkers. 1999. Murine neutrophil stimulationby Toxoplasma gondii antigen drives high level production of IFN-g-indepen-dent IL-12. J. Immunol. 163: 2081–2088.
31. Cerovic, V., S. A. Houston, J. Westlund, L. Utriainen, E. S. Davison, C. L. Scott,C. C. Bain, T. Joeris, W. W. Agace, R. A. Kroczek, et al. 2015. Lymph-borneCD8a+ dendritic cells are uniquely able to cross-prime CD8+ T cells with an-tigen acquired from intestinal epithelial cells. Mucosal Immunol. 8: 38–48.
32. Reis e Sousa, C., S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest,R. N. Germain, and A. Sher. 1997. In vivo microbial stimulation induces rapidCD40 ligand-independent production of interleukin 12 by dendritic cells andtheir redistribution to T cell areas. J. Exp. Med. 186: 1819–1829.
33. Aliberti, J., O. Schulz, D. J. Pennington, H. Tsujimura, C. Reis e Sousa,K. Ozato, and A. Sher. 2003. Essential role for ICSBP in the in vivo developmentof murine CD8alpha + dendritic cells. Blood 101: 305–310.
34. Scott, C. L., C. C. Bain, P. B. Wright, D. Sichien, K. Kotarsky, E. K. Persson,K. Luda, M. Guilliams, B. N. Lambrecht, W. W. Agace, et al. 2015. CCR2(+)CD103(-) intestinal dendritic cells develop from DC-committed precursors andinduce interleukin-17 production by T cells. Mucosal Immunol. 8: 327–339.
35. Vander Lugt, B., A. A. Khan, J. A. Hackney, S. Agrawal, J. Lesch, M. Zhou,W. P. Lee, S. Park, M. Xu, J. DeVoss, et al. 2014. Transcriptional programmingof dendritic cells for enhanced MHC class II antigen presentation. Nat. Immunol.15: 161–167.
36. McKee, A. S., F. Dzierszinski, M. Boes, D. S. Roos, and E. J. Pearce. 2004.Functional inactivation of immature dendritic cells by the intracellular parasiteToxoplasma gondii. J. Immunol. 173: 2632–2640.
37. Pifer, R., A. Benson, C. R. Sturge, and F. Yarovinsky. 2011. UNC93B1 is es-sential for TLR11 activation and IL-12-dependent host resistance to Toxoplasmagondii. J. Biol. Chem. 286: 3307–3314.
38. Rosowski, E. E., D. Lu, L. Julien, L. Rodda, R. A. Gaiser, K. D. Jensen, andJ. P. Saeij. 2011. Strain-specific activation of the NF-kappaB pathway by
GRA15, a novel Toxoplasma gondii dense granule protein. J. Exp. Med. 208:195–212.
39. Dupont, C. D., D. A. Christian, E. M. Selleck, M. Pepper, M. Leney-Greene,G. Harms Pritchard, A. A. Koshy, S. Wagage, M. A. Reuter, L. D. Sibley, et al.2014. Parasite fate and involvement of infected cells in the induction of CD4+and CD8+ T cell responses to Toxoplasma gondii. PLoS Pathog. 10: e1004047.
40. Gumley, T. P., I. F. McKenzie, and M. S. Sandrin. 1995. Tissue expression,structure and function of the murine Ly-6 family of molecules. Immunol. CellBiol. 73: 277–296.
41. Manicassamy, S., and B. Pulendran. 2009. Retinoic acid-dependent regulationof immune responses by dendritic cells and macrophages. Semin. Immunol. 21:22–27.
42. Benson, A., S. Murray, P. Divakar, N. Burnaevskiy, R. Pifer, J. Forman, andF. Yarovinsky. 2012. Microbial infection-induced expansion of effector T cellsovercomes the suppressive effects of regulatory T cells via an IL-2 deprivationmechanism. J. Immunol. 188: 800–810.
43. Yokota, A., H. Takeuchi, N. Maeda, Y. Ohoka, C. Kato, S. Y. Song, andM. Iwata. 2009. GM-CSF and IL-4 synergistically trigger dendritic cells to ac-quire retinoic acid-producing capacity. Int. Immunol. 21: 361–377.
44. Carmen, J. C., and A. P. Sinai. 2007. Suicide prevention: disruption of apoptoticpathways by protozoan parasites. Mol. Microbiol. 64: 904–916.
45. Gavrilescu, L. C., and E. Y. Denkers. 2001. IFN-g overproduction and high levelapoptosis are associated with high but not low virulence Toxoplasma gondiiinfection. J. Immunol. 167: 902–909.
46. Liesenfeld, O., J. C. Kosek, and Y. Suzuki. 1997. Gamma interferon induces Fas-dependent apoptosis of Peyer’s patch T cells in mice following peroral infectionwith Toxoplasma gondii. Infect. Immun. 65: 4682–4689.
47. L€uder, C. G. K., and U. Gross. 2005. Apoptosis and its modulation during in-fection with Toxoplasma gondii: molecular mechanisms and role in pathogen-esis. Curr. Top. Microbiol. Immunol. 289: 219–237.
48. Bekiaris, V., E. K. Persson, and W. W. Agace. 2014. Intestinal dendritic cells inthe regulation of mucosal immunity. Immunol. Rev. 260: 86–101.
49. Mildner, A., and S. Jung. 2014. Development and function of dendritic cellsubsets. Immunity 40: 642–656.
50. Schulthess, J., B. Meresse, E. Ramiro-Puig, N. Montcuquet, S. Darche,B. Begue, F. Ruemmele, C. Combadiere, J. P. Di Santo, D. Buzoni-Gatel, andN. Cerf-Bensussan. 2012. Interleukin-15-dependent NKp46 innate lymphoidcells control intestinal inflammation by recruiting inflammatory monocytes.Immunity 37: 108–121.
51. Denning, T. L., B. A. Norris, O. Medina-Contreras, S. Manicassamy, D. Geem,R. Madan, C. L. Karp, and B. Pulendran. 2011. Functional specializations ofintestinal dendritic cell and macrophage subsets that control Th17 and regulatoryT cell responses are dependent on the T cell/APC ratio, source of mouse strain,and regional localization. J. Immunol. 187: 733–747.
52. Fujimoto, K., T. Karuppuchamy, N. Takemura, M. Shimohigoshi, T. Machida,Y. Haseda, T. Aoshi, K. J. Ishii, S. Akira, and S. Uematsu. 2011. A new subsetof CD103+CD8alpha+ dendritic cells in the small intestine expresses TLR3,TLR7, and TLR9 and induces Th1 response and CTL activity. J. Immunol.186: 6287–6295.
53. Cerovic, V., S. A. Houston, C. L. Scott, A. Aumeunier, U. Yrlid, A. M. Mowat,and S. W. Milling. 2013. Intestinal CD103(-) dendritic cells migrate in lymphand prime effector T cells. Mucosal Immunol. 6: 104–113.
54. Edelson, B. T., W. Kc, R. Juang, M. Kohyama, L. A. Benoit, P. A. Klekotka,C. Moon, J. C. Albring, W. Ise, D. G. Michael, et al. 2010. Peripheral CD103+dendritic cells form a unified subset developmentally related to CD8alpha+conventional dendritic cells. J. Exp. Med. 207: 823–836.
55. Jackson, J. T., Y. Hu, R. Liu, F. Masson, A. D’Amico, S. Carotta, A. Xin,M. J. Camilleri, A. M. Mount, A. Kallies, et al. 2011. Id2 expression delineatesdifferential checkpoints in the genetic program of CD8a+ and CD103+ dendriticcell lineages. EMBO J. 30: 2690–2704.
56. Murphy, T. L., R. Tussiwand, and K. M. Murphy. 2013. Specificity throughcooperation: BATF-IRF interactions control immune-regulatory networks. Nat.Rev. Immunol. 13: 499–509.
57. Christian, D. A., A. A. Koshy, M. A. Reuter, M. R. Betts, J. C. Boothroyd, andC. A. Hunter. 2014. The use of transgenic parasites and host reporters to dissectthe events that promote IL-12 production during toxoplasmosis. Infect. Immun.82: 4056–4067.
58. Braun, L., M. P. Brenier-Pinchart, M. Yogavel, A. Curt-Varesano, R. L. Curt-Bertini, T. Hussain, S. Kieffer-Jaquinod, Y. Coute, H. Pelloux, I. Tardieux, et al.2013. A Toxoplasma dense granule protein, GRA24, modulates the early im-mune response to infection by promoting a direct and sustained host p38 MAPKactivation. J. Exp. Med. 210: 2071–2086.
59. Liesenfeld, O., J. Kosek, J. S. Remington, and Y. Suzuki. 1996. Association ofCD4+ T cell-dependent, interferon-g-mediated necrosis of the small intestinewith genetic susceptibility of mice to peroral infection with Toxoplasma gondii.J. Exp. Med. 184: 597–607.
60. Cohen, S. B., and E. Y. Denkers. 2015. The gut mucosal immune response toToxoplasma gondii. Parasite Immunol. 37: 108–117.
61. Persson, C. M., H. Lambert, P. P. Vutova, I. Dellacasa-Lindberg, J. Nederby,H. Yagita, H. G. Ljunggren, A. Grandien, A. Barragan, and B. J. Chambers.2009. Transmission of Toxoplasma gondii from infected dendritic cells to naturalkiller cells. Infect. Immun. 77: 970–976.
62. Persson, E. K., A. M. Agnarson, H. Lambert, N. Hitziger, H. Yagita,B. J. Chambers, A. Barragan, and A. Grandien. 2007. Death receptor ligation orexposure to perforin trigger rapid egress of the intracellular parasite Toxoplasmagondii. J. Immunol. 179: 8357–8365.
2762 MUCOSAL DC SUBSETS DURING TOXOPLASMA INFECTION
by guest on April 2, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
![Toxoplasma gondii - PCRmax · Toxoplasma gondii is a species of parasitic protozoa in the genus Toxoplasma.[1] The definitive host of T. gondii is the cat, but the parasite can be](https://img.pdfslide.us/doc/110x75/5cc21bb288c993ed078d60e8/toxoplasma-gondii-toxoplasma-gondii-is-a-species-of-parasitic-protozoa-in.jpg)












![Research Article Toxoplasma gondii Pave the Road for Dementia?downloads.hindawi.com/journals/jpr/2020/8859857.pdf · including Toxoplasma gondii (T. gondii) [6], Herpes simplex virus-1](https://img.pdfslide.us/doc/110x75/5f9376ed91220772b35c9b7d/research-article-toxoplasma-gondii-pave-the-road-for-dementia-including-toxoplasma.jpg)
![Primerdesign Ltd TM Toxoplasma gondii - Home : genesig · Toxoplasma gondii is a species of parasitic protozoa in the genus Toxoplasma.[1] The definitivehostofT.gondiiisthecat,buttheparasitecanbecarriedbythevastmajorityof](https://img.pdfslide.us/doc/110x75/5cc21bb288c993ed078d60da/primerdesign-ltd-tm-toxoplasma-gondii-home-toxoplasma-gondii-is-a-species.jpg)














