Embed Size (px)
Citation preview

Impact of single-nucleotide polymorphisms at the TP53-bindingand responsive promoter region of BCL2 gene in modulatingthe phenotypic variability of LGMD2C patients
Ikhlass Hadj Salem • Fatma Kamoun •
Nacim Louhichi • Moez Trigui • Chahnez Triki •
Faiza Fakhfakh
Received: 15 September 2011 / Accepted: 30 January 2012 / Published online: 25 February 2012
� Springer Science+Business Media B.V. 2012
Abstract Apoptosis of skeletal muscle fibers is a well-
known event occurring in patients suffering from muscular
dystrophies. In this study, we hypothesized that functional
polymorphisms in genes involved in the mitochondrial
apoptotic pathway might modulate the apoptotic capacity
underlying the muscle loss and contributing to intrafamilial
and interfamilial variable phenotypes in LGMD2C (Limb
Girdle Muscular Dystrophy type 2C) patients sharing the
same c.521delT mutation in SGCG gene. Detection of
apoptosis was confirmed on muscle biopsies taken from
LGMD2C patients using the TUNEL method. We geno-
typed then ten potentially functional SNPs in TP53, BCL-2
and BAX genes involved in the mitochondrial apoptotic
pathway. Potential genotype-dependent Bcl-2 and p53
protein expressed in skeletal muscle was investigated using
western blot and ELISA assays. The result showed that
muscle cells carrying the TP53-R72R and TP53-16 bp del/
del genotypes displayed an increased p53 level which
could be more effective in inducing apoptosis by activation
of the pro-apoptotic gene expression. In addition, the
BCL2-938 AA genotype was associated with increased
Bcl-2 protein expression in muscle from LGMD2C patients
compared to -938CC genotype, while there was no evi-
dence of significant difference in the BAX haplotype. Our
findings suggest that increased Bcl-2 protein expression
may counteract pro-apoptotic pathways and thus reduce the
muscle loss. To the best of our knowledge, this is a pioneer
study evaluating the role of apoptotic BCL-2 and TP53
genes in contributing to the phenotypic manifestation of
c.521delT mutation in LGMD2C patients. Larger studies
are needed to validate these findings.
Keywords Apoptosis � Bcl-2 � p53 � Single nucleotide
polymorphism � Phenotypic heterogeneity � LGMD2C
Introduction
Apoptosis is observed during normal development in most
tissues and it is a well-known event during skeletal muscle
myogenesis [1, 2]. In addition, apoptosis of skeletal muscle
fibers has been produced in rodents in various experimental
conditions whereas apoptosis of muscle fibers occurring in
humans suffering from different neuromuscular disorders
remains controversial [3, 4]. Some studies have reported
apoptosis of muscle fibers in patients suffering from spinal
muscular atrophy [5] as well as in those suffering from
muscular dystrophies [6]. However, other studies have
failed to demonstrate apoptosis of myonuclei in these dis-
orders and in muscle fibers of patients affected by
inflammatory myopathies [7, 8]. Since skeletal fibers are
mitochondria-rich, several signalling pathways of skeletal
muscle apoptosis are currently under intense investigation
with a particular focus on the role played by mitochondria
[9, 10].
I. Hadj Salem (&) � N. Louhichi � F. Fakhfakh (&)
Laboratoire de Genetique Moleculaire Humaine, Faculte de
Medecine de Sfax, Universite de Sfax, Avenue Magida Boulila,
3029 Sfax, Tunisia
e-mail: [email protected]
F. Fakhfakh
e-mail: [email protected]
F. Kamoun � C. Triki
Service de Neurologie Pediatrique, CHU Hedi Chaker de Sfax,
Sfax, Tunisia
M. Trigui
Service d’Orthopedie et Traumatologie, CHU Habib-Bourguiba
de Sfax, Sfax, Tunisia
123
Mol Biol Rep (2012) 39:7479–7486
DOI 10.1007/s11033-012-1581-4

Muscle atrophy is a genetically-controlled process
involving the activation of the apoptotic pathways and the
ubiquitin- proteasome systems [11]. Although the exact
mechanisms underlying the loss of muscle mass are far to
be unveiled, accumulating preclinical evidence suggests
that acceleration of myocytes loss via apoptosis might
represent a key mechanism driving the onset and progres-
sion of muscle loss and leading to phenotypic heteroge-
neity [11, 12].
A wide clinical spectrum has been reported in patients
with LGMD2C (Limb Girdle Muscular Dystrophy type 2C)
ranging from Duchenne-like features with acute muscle
atrophy to adult-onset LGMD with minimal symptoms.
The age of onset, rate of progression and severity can vary
between and within affected families [13, 14]. This mul-
tiplicity of symptoms and signs cannot be explained on the
basis of the specific mutation alone. It depends on the
interactions with other genetic and/or epigenetic factors.
Therefore, we hypothesized that functional polymorphisms
in genes involved in the mitochondrial apoptotic pathway
might modulate the apoptotic capacity phenotype under-
lying the loss of muscle, thus contributing to intrafamilial
and interfamilial variable phenotypes caused by the iden-
tical mutation c.521delT.
Materials and methods
Study patients
The affected patients were all homozygous for the same
mutation c.521delT responsible for the LGMD2C. They
were clinically evaluated according to the Vignos scale
[15]. The main clinical features of the eight patients are
summarized in Table 1. Notable intrafamilial and interfa-
milial phenotypic variability existed among these patients
in the following parameters: age of onset, pace of pro-
gression and extent of limb weakness and disability
(Table 1).
Muscle tissue specimens
Biopsies were taken from the median head of the deltoid
muscle, for diagnosis purposes, and were used after the
patients’ informed consent. Four muscle biopsies from
healthy individuals and five from LGMD2C patients were
analyzed. A part of each biopsy specimen was frozen
immediately after excision in isopentane cooled by means
of liquid nitrogen and was stored at -80�C. Serial Sec-
tions 4 lm in thickness were cut in a cryostat microtome,
dried at room temperature, and directly stained or stored at
-80�C.
Detection of apoptosis
To detect apoptosis in muscle tissue we used the TUNEL
assay with an in situ Cell Death Detection kit (Fluorescein,
Roche) according to the manufacturer’s recommendations.
Positive controls were made by incubating some sections
with 0.5 mg/ml DNAse I (Promega) at room temperature
for 10 min before TUNEL staining. DNAse I treated sec-
tions incubated with fluorescein-labeled nucleotide mixture
without the addition of terminal deoxynucleotidyl trans-
ferase, were used as negative controls. The slides were
washed with phosphate buffer saline (PBS) and stained
with a 4,6-diamidino-2-phenylindole (DAPI: 100 ug/l)
solution for 20 min at room temperature. The slides were
analyzed using a fluorescence Microscope (Axioskop Z
plus Zeiss). All the experiments were conducted three to
four times with appropriate controls.
Genotyping
The genotyping methods used to distinguish the 10 selected
polymorphisms in 3 apoptosis-related genes TP53, BCL2
and BAX are presented in Table 2. Direct sequencing was
used for detecting four polymorphisms: c.459?1619 A[G
in BCL-2 and c.34?43 C[T; c.233?14 A[G and c.824
G[A in BAX genes. Genotyping methods for six SNPs
were previously described and detected by using either
PCR–RFLP (c.215 C[G and c.672?62 A[G in TP53
gene; -938 C[A and c.127 G[A in BCL-2 gene) or PCR-
PIRA (-248 G[A in BAX gene) or standard PCR (16-bp
ins/del in TP53 gene) [Table 2; [16, 17]]. The PCR
amplification was performed in a thermal cycler (Gene-
Amp PCR System 9700 (Applied Biosystems)) in a final
volume of 50 ll using 200 ng DNA, 8 pmol of each pri-
mer, 2 mM MgCl2, 500 lM dNTP, 1 9 PCR buffer, and
2 U Taq DNA polymerase. The conditions for the PCR
were as follows: initial denaturation at 95�C for 5 min,
followed by 10 cycles: 40 s at 95�C, 45 s at 66–60�C, and
45 s at 72�C, and then 35 cycles at 95�C for 40 s, 60�C for
45 s, 72�C for 45 s, and a final extension at 72�C for
10 min.
As regards the BCL2 (-938 C[A) polymorphism in the
promoter, the primers amplified a 262 bp DNA fragment.
Then, the PCR product was digested by BccI (New Eng-
land BioLabs) overnight at 37�C. The wild-type allele
(AA) produced two bands of 154 and 108 bp; wild-type/
variant allele (CA) produced three bands of 108, 154 and
262 bp and the variant allele (CC) produced a single
262 bp band (Table 2).
Regarding the BAX (-248 G[A) polymorphism in the
promoter, the forward primer (CATTAGAGCTGCG-
ATTGGACCG), in which the -2 C was introduced to
create a MspI restriction site, and the reverse primer
7480 Mol Biol Rep (2012) 39:7479–7486
123

(GCTCCCTCGGGAGGTTTGGT), amplified a 109 bp
DNA fragment (Table 2). Then, the PCR product was
digested by MspI (Fermentas) overnight at 37�C.
As for the TP53 (Arg72Pro) polymorphism in the pro-
moter, the primers amplified a 296 bp DNA fragment.
Then, the PCR product was digested by BstUI (New
England BioLabs) overnight at 60�C. The wild-type allele
(GG) produced two bands of 169 and 127 bp); wild-type/
variant allele (GC) produced three bands of 127, 169 and
296 bp and the variant allele (CC) produced a single
296 bp band (Table 2). Genotypes identified by PCR–
RFLP were confirmed with DNA sequencing using an ABI
PRISM� 3100 DNA sequencer.
Western-blot analysis of Bcl-2 expression in muscle
samples
We investigated the association between the CC, AC and
AC genotypes of polymorphism BCL-2 (-938 C[A) and
Bcl2 protein expression level in muscle. Nine muscle
biopsies were solubilized in treatment buffer in the pres-
ence of protease inhibitors. Protein concentration was
determined according to Bradford [18]. After denaturation
for 5 min at 100�C, sodium dodecyl sulphate (SDS)–
polyacrylamide gel electrophoresis was performed using of
equal amounts of muscular proteins, followed by transfer to
nitrocellulose filters. After blocking the reaction with 5%
milk powder and incubation with the primary antibodies
(anti-Bcl2 diluted titer of 1/1,000 and anti a-dystroglycan
diluted titer of 1/2,000), the nitrocellulose sheet was further
incubated with a mouse peroxidase-conjugated secondary
antibody (1/5,000) and developed using an enhanced
chemiluminescence system (Immun-Star HRP kit Bio-Rad)
according to the manufacturer’s instructions. Films were
scanned and densitometry was used to quantify the
immunoblot signals. To compare protein expression among
the different genotypes, the average intensity of the signal
was multiplied by the number of pixels in that area and
normalized to a-dystroglycan expression. The following
antibodies were used: mouse monoclonal anti-Bcl2 anti-
body (Invitrogen), mouse anti- a-dystroglycan antibody
(VIA4-1; Upstate Biotechnology), and mouse peroxidase-
conjugated secondary antibody (IgG H?L code 75031
Sanofi Diagnostics Pasteur).
Enzyme linked immunosorbent assay (ELISA)
Wells from microliter plates were coated with muscular
proteins for 2 h at 37�C and then overnight at 4�C. The
plates were washed with PBS-Tween and blocked with 1%
non-fat, powdered milk in PBS-Tween for 1 h at 37�C.
After three washings, monoclonal anti-Bcl-2, monoclonal
anti-p53 (Santa Cruz Biotechnology; diluted titer of
1:100)) and mouse anti- a-dystroglycan antibodies were
used for 2 h at 37�C. Dilution was the same as that used in
western blot assay. The plates were washed and peroxy-
dase-conjugated anti-mouse IgG (diluted titer of 1:5,000 in
the PBS-Tween) was added for 1 h at 37�C. Three wash-
ings were performed, and the chromogenic reagent tetra-
methylbenzidine (TMB) was used to reveal the binding of
the peroxidase-conjugated second antibody. The reaction
was stopped by adding 50 ll of 2 M H2SO4 to each well,
and the resulting absorbance was measured at 450 nm.
Statistical analysis
To test the genotype-dependent differences in protein
expression (Western-blot analysis), we performed pairwise
comparisons using Student t test. Differences were regar-
ded significant at P values less than 0.05.
Table 1 Clinical and paraclinical data of LGMD2C patients sharing the c.521delT mutation
Patients Current age
[years]/sex
Age of onset
[years]
Serum CK
(UI)/age [years]aCalves
hypertrophy
Functional
gradebWheelchair bound
(age [years])
P1 17/M 4 11,900/8 – 10 10
P10 13/F 9 9,000/9 ? 2 –
P2 22/M 11 735/18 ? 10 14
P20 18/M 5 650/15 – 10 10
P3 13/F 5 8,110/7 – 10 9
P4 17/M 7 10,570/8 ? 10 12
P40 13/F 10 9,875/11 ? 3 –
P5 9/F 7 10,840/7 – 3 –
P6 8/F 6 10,485/7 – 3 –
M male, F female, CK Creatine Kinase, ? present, – absenta Normal value of CK: 25–350 IU/Lb Clinical testing are based on a scale of 10 [15]
Mol Biol Rep (2012) 39:7479–7486 7481
123

Results
Apoptosis assay
To investigate the mechanism of myofiber degeneration in
c-sarcoglycan deficiency, TUNEL staining was performed
on deltoid muscle biopsies from LGMD2C patients and
healthy individuals. No TUNEL positive myonuclei were
detected in healthy muscle (Fig. 1, HI). In contrast,
TUNEL-positive myonuclei were commonly found in
LGMD2C patients’ muscle (Fig. 1, P1 and P4) indicating
that apoptosis contributed to muscle degeneration in c-
Fig. 1 Apoptosis in LGMD2C deltoid muscular biopsies. Cross-
sections from healthy individual (HI) and LGMD2C patients (P1 and
P4) were labeled for apoptosis staining by TUNEL. Digitized images
were merged to superimpose the TUNEL labeled nuclei (greenpseudocolor field) with the DAPI staining to mark the cells and
discriminate myonuclei from outer nuclei. No TUNEL positive
nucleus was detected in the normal muscle whereas, in the LGMD2C
patients, positive nuclei can be observed in some fibers. Three
successive tissue sections for each sample were examined by
Fluorescence microscopy (magnification 940). Color figure online
Table 2 Genotyping assays for the selected polymorphisms
Gene Position and
base change
Genotyping method Primer PCR
product
Enzyme Gel band
pattern
TP53 p.R72P G[C
(rs1042522)
PCR–RFLP F: 50ATCTACAGTCCCCCTTGCCG30 296 bp BstUI G allele: 169
and 127 bp
R: 50GCAACTGACCGTGCAAGTCA30 C allele: 296 bp
16-bpins/del
(rs17878362)
PCR F: 50TGGGACTGACTTTCTGCTCTT30 Del: 180 bp
R: 50TCAAATCATCCATTGCTTGG30 Ins: 196 bp
c.672?62 G[A
(rs1625895)
PCR–RFLP F: 50TGGCCATCTACAAGCAGTCA30 404 bp MspI G allele:
336 bp,
68 bp
R: 50TTGCACATCTCATGGGGTTA30 A allele: 404 bp
BCL2 -938 C[A
(rs2279115)
PCR–RFLP F: 50TCCTGCCTTCATTTATCCAGCA30 262 bp Bcc I A allele:
154 bp,
108 bpR: 50CCAGGAGAGAGACAGGGGACA30
C allele: 262 bp
c.127 G[A
(rs1800477)
PCR–RFLP F: 50CCCGTTGCTTTTCCTCTGGGA30 178 bp Bgl I G allele: 157 bp,
21 bp
R: 50AGAAGATGCCCGCCGCGGGG30 A allele: 178 bp
c.458?1619 G[A
(rs4987853)
Direct
sequencing
F: 50TCTGTTGTCCCTTTGACCTTG30 196 bp
R: 50GGCCACGTAAAGCAACTCTC30
BAX -248 G[A
(rs4645878)
PIRA–PCR:
Mismatch,
sense primer
-2 G-to-C
F: 50CATTAGAGCTGCGATTGGACCG30 109 bp MspI G allele: 89 bp,
20 bp
R: 50GCTCCCTCGGGAGGTTTGGT30 A allele: 109 bp
c.34?43 C[T
(rs4645881)
Direct
sequencing
F: 50CATTAGAGCTGCGATTGG30 406 bp
R: 50CTCAGTGCTTGGAGATCG30
c.233?14 A[G
(rs1805419)
Direct
sequencing
F: 50CCGTCACTTTATCTGCTAGG30 521 bp
R: 50GGCCCAGACTCCTAGTTCTTAG30
c.824 G[A
(rs704243)
Direct
sequencing
F: 50AATGCCCGTTCATCTCAG30 371 bp
R: 50CCTCAAGACCACTCTTCC30
7482 Mol Biol Rep (2012) 39:7479–7486
123

sarcoglycan deficient muscular dystrophy. Apoptotic fea-
tures were seen outside the myofiber, presumably in
mononuclear cells of LGMD2C interstitial muscular tissue
and in some fibers, but were also seen in intact or partially
degenerated muscle fibers (Fig. 1). The presence of
TUNEL-positive myonuclei in muscle fibers suggests that
c-sarcoglycan deficiency leads to apoptosis as an event in
the dystrophic process.
Genotyping
Since apoptotic phenotype was confirmed with TUNEL
assay and in order to explain the phenotypic heterogeneity
observed in the LGMD2C patients sharing the same
mutation c.521delT, we performed a genotype-phenotype
analysis using polymorphisms in apoptotic genes. Poly-
morphisms were included in this study if they might the-
oretically result in amino acid changes (nonsynonymous
SNP, nsSNP), or they are located at regulating regions such
as promoters, or reportedly associated with known pheno-
typic effects. We selected 10 potentially functional poly-
morphisms in 3 genes that were, TP53, BCL-2 and BAX
genes involved in the mitochondrial pathway. Figure 2
shows the haplotypes construction performed for all vari-
ants. With regard to SNPs in TP53 gene, we note two
different haplotypes: one haplotype includes variant het-
erozygotes for TP53 c.215C/G (p.R72P), TP53 intron 3
16-bp ins/del and TP53 c.672 ? 62A/G (TP53 intron 6
A/G), whereas the other haplotype includes variant in
homozygous state: c.215G (p.72R), 16 bp del/del and
c.672 ? 62G (Fig. 2).
The study of genotyping of the polymorphisms sites in
Bcl-2 family members’ genes showed similar genotypes for
-248 G[A; c.34?43 C[T; c.233?14 A[G; and c.824
G[A in Bax and c.127 G[A and c.458?1619 G[A in Bcl-
2 genes whereas the most studied polymorphism site -938
C[A in Bcl-2 gene showed different genotypes: CC, AA
and CA (Fig. 2).
Effects of the BCL2-938 C[A polymorphism on Bcl-2
protein expression in muscular biopsies
We investigated potential genotype-dependent Bcl-2 pro-
tein expression in skeletal muscle from LGMD2C patients
and healthy individuals. As shown in Fig. 3a, expression of
Bcl-2 in patients with -938 AA genotype was increased
significantly compared with that in muscle from LGMD2C
patients with CC genotypes. Protein expression associated
with the AC genotype was somehow more variable,
showing low expression compared to that seen in CC
genotypes, intermediate expression in another sample and
higher expression in one sample resembling the AA
genotype. To correct the variations in total protein, blots
were also probed with an anti-a-dystroglycan antibody.
After densitometry of Bcl-2 and a-dystroglycan specific
bands, the ratios of Bcl2/a-dystroglycan were computed
Fig. 2 Pedigree of the Tunisian
families harboring the
c.521delT mutation and
showing the haplotypes of
regulatory SNPs in apoptotic
genes TP53, BCL-2 and BAX.
m: mutation c.521delT in SGCGgene. We note two different
haplotypes for TP53 SNPs
(simple and double framed in
red) and three different
genotypes CC, AA and CA for
BCL2-938C/A polymorphism
(simple and double framed in
blue). Non-boxed genotypes for
the remaining SNPs are non-
informative for all patients.
Color figure online
Mol Biol Rep (2012) 39:7479–7486 7483
123

and associated with BCL2 genotypes (Fig. 3b). The ratios
Bcl2/a-dystroglycan were almost threefold higher when
comparing AA (0.980 ± 0.07) with CC genotypes
(0.31 ± 0.07; P = 0.0009; Student’s t test) whereas
intermediate ratios were found associated with the AC
genotype (0.54 ± 0.11). Thus, an increased activity of the
inhibitory P2 promoter appears to result in an accordingly
reduced bcl-2 protein expression associated with the CC
genotype.
From the result of ELISA assay, and as found in western
blot analysis, we found that CC genotype correlated with
low Bcl-2 expression, while AA genotype correlated with
the highest expression which confirms the western blot
result (Fig. 3c). We noted also the expression level of p53
in each genotype. Figure 3c shows the rate of p53
expression in each genotype. P53 expression was signifi-
cantly higher in TP53 R72R, 16 bp-del/del haplotype car-
riers (Fig. 3c). The expression of Bcl-2 and p53 was found
to be slightly lower in normal muscle.
Discussion
The Tunisian families described here bear the LGMD2C
after clinical, genetic and molecular investigation.
Although the patients shared the same mutation c.521delT
in SGCG gene and displayed similar mild limb girdle
muscular dystrophy phenotypic features, heterogeneity was
expressed by variability in the age of onset, in the age of
wheelchair-bound and in the course of the disease.
Understanding the intrafamilial clinical variability in
patients carrying the same mutation remains a great
challenge.
In this study, we performed genotype–phenotype anal-
ysis to examine the role of potentially functional variants in
TP53, BCL-2 and BAX apoptotic genes in modulating the
phenotype expression. An intriguing finding in the present
study is the significant difference in TP53 and BCL-2
haplotypes found between LGMD2C patients. TUNEL
assay was performed on histologically affected muscle
biopsies of five LGMD2C patients. The results demonstrate
the presence of myofibers presenting TUNEL-positive
nuclei. An increase in apoptosis and detection of TUNEL-
positive nuclei were found in biopsies from gsg-/-mice
suggesting that programmed cell death contributes to
myofiber degradation [19].
The bcl-2 family of proto-oncogenes encodes specific
proteins which are a well-known group of death-modulat-
ing factors regulated by the p53 transcriptional factor and
involved in the mitochondrial apoptotic pathway. Bcl-2
inhibits cell death in a variety of mammalian cell types,
whereas its homologous protein, Bax, promotes cell death
via apoptosis [20].
A total number of 14 polymorphisms have been identi-
fied in the TP53 gene but the most intensively studied ones
being R72P (c.215 C[G), p53-16 bp del/ins (intron 3), and
c.672?62 A[G (intron 6). Here we show for the first time
that biopsy muscle from LGMD2C patients carrying TP53
R72 and 16-bp deletion variants may display an increased
p53 level which is more effective to induce apoptosis by
inducing pro-apoptotic gene expression. In fact, the Pro72
and Arg72 variants were reported to differ in their
Fig. 3 Correlation between Bcl2-938 C/A, R72P, 16 bp ins/del TP53
genotypes and protein expression. a Genotype-dependent expression
of Bcl-2 protein in muscle from LGMD2C patients. Western-blot
analyses of muscle from patients with genotype BCL2-938 AA
showed increased expression of Bcl-2 protein (28 kDa) compared to
those from patients with BCL2-938 AC and BCL2-938 CC genotypes.
Alpha-dystroglycan (156 kDa) was used as a standard to allow for
normalization to differential protein loading. b Quantitative analysis
of protein expression shown in panel A. Densitometry was performed
using Scion Image statistical analysis. Horizontal bars represent the
mean value of the three values of each genotype. c The p53 and Bcl-2
expression levels were measured using the ELISA technique for the
different genotypes found in our genotype–phenotype analysis. HIHealthy individuals. The absorbance was read at 450 nm
7484 Mol Biol Rep (2012) 39:7479–7486
123

functional activity: the Arg72 variant suppressed effec-
tively cellular transformation, and was more efficient than
the Pro72 variant at inducing apoptosis [21, 22]. The
expression of p53 pro-apoptotic target genes and the
mitochondrial apoptotic response in p53-null cell lines
stably transfected with constructs that encode p53-R72 is
higher than in cell lines expressing the p53-P72 variant,
supporting the current view that p53-R72 is a more potent
inducer of apoptosis than p53-P72 [21]. Thus, the muscle
cells carrying the arginine allele may have greater sus-
ceptibility to apoptosis (Fig. 4). In addition, the 16 bp
deletion allele was described to be associated with high
level of TP53 transcript in lymphoblastoid cell lines,
whereas the 16 bp insertion allele might influence alter-
native splicing of p53 protein, leading to unstable tran-
scripts or proteins with altered activities and reduction of
interaction with the pro-apoptotic members [23].
Anti-apoptotic BCL-2 gene plays an important role in
the regulation of apoptosis. Previous studies had examined
the significance of apoptosis in muscular dystrophies
pathogenesis by inhibiting apoptosis through either inacti-
vation of the pro-apoptosis protein Bax or overexpression
of the anti-apoptosis protein Bcl-2 [20]. In this study, we
found that muscle from patients homozygous for the
-938A allele displayed an increased Bcl-2 protein
expression while -938CC genotype correlated with the
lowest expression. As indicated in previous literature, this
polymorphism was located in the second promoter (P2) of
the BCL-2 gene, which worked as a negative modulator on
the first promoter (P1) and subsequently, BCL-2 gene
expression [24].This may suggest that the significantly
higher activity of the C allele may decrease more effi-
ciently the activation of the predominant promoter, P1. We
speculate then that C allele may result in a down-regulation
of bcl-2 mRNA transcript levels, a decrease in cellular
levels of bcl-2 protein, unregulated programmed cell death
which enhances loss of the muscle mass leading to a more
severe phenotype (Fig. 4).
A role for Bcl-2 in muscle pathology has been suggested
indicating that Bcl-2 is expressed in response to injury in
order to counter-act pro-apoptotic pathways and thus to
reduce the muscle loss [25]. Indeed, an overexpression of
Bcl-2 in myoblasts from mdx-mice was shown to prevent a
rise in intracellular Ca2?, and thus inhibiting apoptosis
[26]. It has been suggested that Bcl-2 may also act as an
antioxidant, thus protecting cells from free radicals [25].
Since free radical toxicity has been hypothesized as being
related to muscle fiber damage in several muscle disorders,
we cannot rule out the possibility that the expression of
these apoptosis-related proteins may reflect some protec-
tive mechanism of the injured muscle fiber against free-
radical cytotoxicity. Interestingly, this has been supported
in interventional animal studies [25].
Our observation indicates the possibility of apoptotic
BCL-2 and TP53 genes contributing to the phenotypic
manifestation of homozygous c.521delT mutation in our
patients, which, of course, requires confirmation from
independent studies to support this investigation. Although
the number of patients in each family is small and this
phenotypic expression could occur by chance, perhaps the
intrafamilial heterogeneity observed in our patients, toge-
ther with the interfamilial heterogeneity, could help us to
better understand which modifying factors play a role in
the different manifestations of the disease.
Acknowledgments We are grateful to the family members for their
invaluable cooperation. We are grateful to Pr. Raja Mokdad Gargouri
from Centre of Biotechnology of Sfax, for providing us the p53
antibody. We also acknowledge Mr. Jamil JAOUA, founder and
former head of the English Unit at the Sfax Faculty of Science,
Tunisia, for proofreading this paper.
References
1. Webb J (1977) Cell death in developing skeletal muscle: histo-
chemistry and ultrastructure. J Pathol 123:175–180
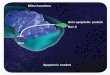
Fig. 4 Hypothesized mechanism for interaction between the Bcl2-
938 C/A and R72P, 16 bp ins/del TP53 polymorphisms and severity
of phenotype. LGMD2C patients who possess the proposed trans-
criptionally inactive Bcl-2-938C allele and R72R, 16-bp del/del
haplotype in TP53 gene may have reduced local BCL-2 levels,
resulting in suppressed Bcl-2-mediated inhibition of apoptosis,
enhanced programmed cell death by inducing expression of pro-
apoptotic genes, loss of muscle mass and subsequent more severe
phenotype
Mol Biol Rep (2012) 39:7479–7486 7485
123

2. Fidzianska A, Goebel H (1990) Human ontogenesis: cell death in
fetal muscle. Acta Neuropathol 81:572–577
3. Carraro U (1995) Apoptotic death of dystrophic muscle fibers
after exercise: a new hypothesis on the early events of muscle
damage. Basic Appl Myol 5:371–374
4. Podhorska-Olokov M, Sandri M, Bruson A et al (1995) Apoptotic
myonuclei appear in adult skeletal muscles of normal and mdxmice after mild exercise. Basic Appl Myol 5:87–90
5. Tews DS, Goebel H (1997) Apoptosis-related proteins in skeletal
muscle fibers of spinal muscular atrophy. J Neuropathol Exp
Neurol 56:150–156
6. Tews DS, Goebel H (1997) DNA-fragmentation and expression
of apoptosis-related proteins in muscular dystrophies. Neuropa-
thol Appl Neurobiol 23:331–338
7. Schneider C, Gold R, Dalakas M et al (1996) MHC class
I-mediated cytotoxicity does not induce apoptosis in muscle
fibers nor in inflammatory T cells: studies in patients with pol-
ymyositis, dermatomyositis and inclusion body myositis. J Neu-
ropathol Exp Neurol 55:1205–1209
8. Olive M, Mtnez-Matos JA, Montero J et al (1997) Apoptosis is
not the mechanism of cell death of muscle fibers in human
muscular dystrophies and inflammatory myopathies. Muscle
Nerve 20:1328–1330
9. Pellegrino MA, Desaphy JF, Brocca L et al (2011) Redox
homeostasis, oxidative stress and disuse muscle atrophy. J Phys-
iol 589:2147–2160
10. Dam AD, Mitchell AS, Rush JWE, Quadrilatero J (2011) Ele-
vated skeletal muscle apoptotic signaling following glutathione
depletion. Apoptosis. doi:10.1007/s10495-011-0654-5
11. Marzetti E, Leeuwenburgh C (2006) Skeletal muscle apoptosis,
sarcopenia and frailty at old age. Exp Gerontol 41:234–238
12. Marzetti E, Privitera G, Simili V et al (2010) Multiple pathways
to the same end: mechanisms of myonuclear apoptosis in sarco-
penia of aging. Sci World J 19:340–349
13. Angelini C, Fanin M, Freda MP et al (1999) The clinical spec-
trum of sarcoglycanopathies. Neurology 52:176–179
14. Sandona D, Betto R (2009) Sarcoglycanopathies: molecular
pathogenesis and therapeutic prospects. Expert Rev Mol Med
11:e28. doi:10.1017/S1462399409001203
15. Vignos PJ, Spencer GE, Archibald KC (1963) Management of
progressive muscular dystrophy of childhood. J Am Med Assoc
194:89–96
16. Zhibin H, Chunying L, Kexin C et al (2008) Single nucleotide
polymorphisms in selected apoptotic genes and BPDE-induced
apoptotic capacity in apparently normal primary lymphocytes: a
genotype–phenotype correlation analysis. J Cancer Epidemiol.
doi:10.1155/2008/147905
17. Chen K, Hu Z, Wang LE et al (2007) Single-nucleotide poly-
morphisms at the TP53-binding or responsive promoter regions
of BAX and BCL2 genes and risk of squamous cell carcinoma of
the head and neck. Carcinogenesis 28:2008–2012
18. Rajini S, Barkha P, Hamadeh J (2009) Comparison of total pro-
tein concentration in skeletal muscle as measured by the Bradford
and Lowry Assays. J Biochem 145:791–797
19. Hack A, Chantal T, Fang J et al (1998) c-Sarcoglycan deficiency
leads to muscle membrane defects and apoptosis independent of
dystrophin. J Cell Biol 142:1279–1287
20. Miller JB, Girgenrath M (2006) The role of apoptosis in neuro-
muscular diseases and prospects for anti-apoptosis therapy.
Trends in Mol Med 12:279–286
21. Dumont P, Leu J, Della Pietra A et al (2003) The codon 72
polymorphic variants of p53 have markedly different apoptotic
potential. Nat Genet 33:357–365
22. Thomas M, Kalita A, Labrecque A et al (1999) Two polymorphic
variants of wild-type p53 differ biochemically and biologically.
Mol Cell Biol 19:1092–1100
23. Gemignani F, Moreno V, Landi S et al (2004) A TP53 poly-
morphism is associated with increased risk of colorectal cancer
and with reduced levels of TP53 mRNA. Oncogene
23:1954–1956
24. Young RL, Korsmeyer SJ (1993) A negative regulatory element
in the bcl-2 50-untranslated region inhibits expression from an
upstream promoter. Mol Cell Biol 13:3686–3697
25. Basset O, Boittin FX, Cognard C et al (2006) Bcl-2 overex-
pression prevents calcium overload and subsequent apoptosis in
dystrophic myotubes. Biochem J 395:267–276
26. Danielssona O, Nilssonb C, Lindvallc B et al (2009) Expression
of apoptosis related proteins in normal and diseased muscle: a
possible role for Bcl-2 in protection of striated muscle. NMD
19:412–417
7486 Mol Biol Rep (2012) 39:7479–7486
123