Embed Size (px)
Citation preview

Kidney International, Vol. 59 (2001), pp. 515–531
Immunofluorescent imaging of b1- and b2-adrenergic receptorsin rat kidney
VALERIE BOIVIN, ROLAND JAHNS, STEPAN GAMBARYAN, WINFRIED NESS, FRITZ BOEGE, andMARTIN J. LOHSE
Institute of Pharmacology and Toxicology; Department of Internal Medicine, Medizinische Poliklinik; Institute of ClinicalBiochemistry and Pathobiochemistry, Medizinische Klinik; and Institute of Anatomy, University of Wuerzburg,Wuerzburg, Germany
Immunofluorescent imaging of b1- and b2-adrenergic receptors participates in the regulation of several physiologicallyin rat kidney. important processes in the mammalian kidney. These
Background. b-Adrenergic receptors (b-ARs) are known include renal blood flow, glomerular filtration rate, NaClto participate in the regulation of glomerular filtration, NaCl reabsorption, acid-base balance, and most notably, thereabsorption, acid-base balance, and renin secretion; however,
secretion of renin from the renin-producing juxtaglomer-the precise histologic localization of b-AR at putative signalingular granular cells [reviewed in 1]. A number of attemptssites involved in these processes remains an open issue.
Methods. We used a set of subtype-specific rabbit antibodies have been made to elucidate further the specific role ofto visualize b1- and b2-AR in rat kidney by immunohistochemis- b-adrenergic receptors (b-ARs) in each of these proc-try and specified cells and segments of the nephron thought esses [2–4]. In this context, the localization and tissueto be regulated by catecholamines. In addition, the relative distribution of b-AR in the kidney have been main issuesproportion of b-AR subtypes in cortical and medullary portions
of investigation. Because antibodies suitable for immu-of rat kidney was determined by Western blotting and by com-nohistochemistry of b-AR in tissues have not been avail-peting [125I]-cyanopindolol binding with the b1- or b2-selective
antagonists bisoprolol or ICI 118,551, respectively. able so far [reviewed in 5], such studies had to be carriedResults. Immunoreactivity for b1-AR was found in mesan- out with radiolabeled [6, 7] or fluorescent receptor li-
gial cells, juxtaglomerular granular cells, the macula densa epi- gands [8] or, more recently, by in situ hybridization [9]thelium, proximal and distal tubular segments, and acid-secre- and/or reverse transcription-polymerase chain reactionting type A intercalated cells of the cortical and medullary
(RT-PCR) [10–12]. However, neither the resolution norcollecting ducts. Immunoreactivity for b2-AR was predomi-the specificity of these techniques have been sufficientnantly localized in the apical and subapical compartment ofto determine the localization and distribution of renalproximal and, to a lesser extent, distal tubular epithelia (sug-
gesting interactions with luminal fluid catecholamines). Both b-AR at the level of functional domains, specific cellsubtypes were dense in the membranes of smooth muscle cells types, or even subcellular structures. Recently, we pro-from renal arteries. Concordant data were obtained by radioli- duced a set of highly sensitive antibodies against b-AR,gand binding and immunoblotting of membranes prepared which are suitable for immunohistological studies andfrom cortical and medullary portions of the kidney.
distinguishing between receptor subtypes [13]. Here, weConclusion. Our data provide an immunohistochemical ba-used these tools to visualize b1- and b2-AR in rat kidneysis for the cellular targets of b-adrenergic regulation of renalat a high resolution. Our data provide a precise delineationfunction. Moreover, they could help to devise therapeutic strat-
egies directed at renal b-ARs. of the histomorphological distribution of b-AR withinthe kidney, specify some target cells upon which cate-cholamines can act via these receptors, and thus mightbe helpful in uncovering the specific roles of b1- andSince the early denervation experiments and pharma-b2-AR in the regulation of renal function. They mightcological studies on whole kidney and/or isolated kidneyalso contribute to a rational basis for influencing certaincells, it has been known that the b-adrenergic systemof the previously mentioned processes by pharmacologi-cal agents targeted at renal b-AR.
Key words: immunohistochemistry, receptor-antibodies, kidney func-tion, cell signaling, catecholamines.
METHODSReceived for publication May 5, 2000
Antibodiesand in revised form August 14, 2000Accepted for publication August 16, 2000 Production and characterization of subtype-specific
rabbit antibodies against the human b1- and b2-AR have 2001 by the International Society of Nephrology
515

Boivin et al: b-ARs in rat kidney516
previously been published [13]. Here, we used affinity- (2200 Ci/mmol [125I]-CYP; NEN, Zaventem, Belgium).purified antibodies directed against the aminoterminal Samples of 50 mg membrane protein were incubated (30(extracellular) domains of human b1- or b2-AR. The re- min, 308C) each with 400 pmol/L [125I]-CYP. Nonspecificceptors are highly conserved between rats [14, 15] and binding was determined in the presence of 10 mmol/Lhumans [16, 17] with a calculated amino acid sequence unlabeled l-propanolol. To determine the relative pro-identity of 93% for the b1-AR and 89% for the b2-AR, portion of either b1- or b2-AR, incubations with 100 pmol/Lrespectively. To control for specificity, the antibodies were [125I]-CYP were supplemented with various concentra-preincubated (12 hours, 48C) with baculovirus-infected tions of unlabeled receptor antagonists selective for b1-Sf9 insect cells expressing 1 to 2 3 106 intact recombinant (bisoprolol; Merck, Darmstadt, Germany) or b2-AR (ICIb1- or b2-AR per cell [13, 18, 19]. The unbound antibodies 118,551; RBI, Deisenhofen, Germany). The reactionsremaining in the supernatant after sedimentation (900 3 g, were stopped by adding an ice-cold buffer (0.1 mol/L48C, 10 min) of Sf9 cells expressing the corresponding Tris-HCl, pH 7.5) and rapid filtration (Whatman GF/Cb-AR subtype or not were used for these experiments. soaked in 0.3% polyethylenimine). Filter-bound radio-Immunolabeling of b1- and b2-AR was assigned to spe- activity was measured by gamma counting. Ligand bind-cific segments and cell types of the nephron by simultane- ing curves were fitted to the data by computer-aided non-ous or comparative immunostaining of various marker linear regression analysis.proteins. Calbindin (cytosolic vitamin D-dependent Ca21
binding protein) was detected with a mouse monoclonal Immunohistochemistryantibody D-28K (Sigma, Deisenhofen, Germany). Tamm- Large structures (that is, blood vessels and glomeruli)Horsfall protein was detected with a rabbit polyclonal were investigated in acetone-fixed kidney sections: Ratantibody (Biotrend, Koln, Germany). Renin was de- kidneys were rapidly frozen in isopentane at 2568C andtected by a rat antiserum, kindly provided by Professor cryostat sections of 3 to 4 mm were fixed with cold ace-A. Kurtz (Institute of Physiology, University of Regens- tone (4 min, 2208C). To investigate tubular structures,burg, Germany). Polyclonal rabbit antibodies against the the kidneys were fixed in situ by formaldehyde perfusion,endothelial nitric oxide synthetase were obtained from which provides a better preservation of the cell struc-Professor H. Schmidt (Institute of Pharmacology, Uni- tures. The kidneys of anesthetized rats were perfusedversity of Wurzburg, Germany). A mouse monoclonal through an aortic cannula (placed below the level ofantibody directed against synaptopodin was a gift from the kidneys), first with 4% formaldehyde in phosphate-Dr. P. Mundel (Albert Einstein College, New York, NY, buffered saline (PBS) for five minutes followed by 0.8USA). Specificity of a polyclonal rabbit antibody against mol/L sucrose in PBS for another five minutes. Subse-rat anion exchanger 1 (band 3 protein) has been pub-
quently, the kidneys were removed and treated as de-lished elsewhere [20].scribed previously in this article. Fixed kidney sectionswere rinsed three times with ice-cold PBS, blocked (30Western blots analysismin, 218C) with PBS containing 4% bovine serum albu-The kidneys of anesthetized male Sprague-Dawleymin (BSA), and 2% normal goat serum (Sigma), andrats (350 to 400 g) were removed and dissected intoincubated (12 h, 48C) with affinity-purified receptor anti-cortical, outer, and inner medullary portions. The differ-bodies diluted 1:200 in PBS containing 2% BSA and 2%ent portions were homogenized in ice-cold buffer [10goat serum. After thorough rinsing with PBS, tissue-mmol/L K2HPO4, 1 mmol/L ethylenediaminetetraaceticbound antibodies were detected with appropriate spe-acid (EDTA), 1 mmol/L EGTA] containing various pro-cies-specific secondary antibodies (Dianova) conjugatedtease inhibitors (5 mg/mL leupeptine, 1.5 mmol/L ben-to CY3 or CY2 (diluted 1:800 or 1:400, respectively).zamidine, 200 U/mL aprotinine, 2 mg/mL pepstatine A).After rinsing again with PBS (3 times 10 min, 218C), theSamples of 60 mg membrane protein of each of theseslides were mounted in mowiol (Sigma) and inspected atportions were analyzed by Western blot using affinity-200- to 630-fold magnification under an epifluorescencepurified rabbit antibodies against b1- or b2-AR dilutedmicroscope (Carl Zeiss, Oberkochen, Germany). Images1:1000 [13]. Immunoreactive bands were visualized bywere photographed with Kodak TMAX 400 black andhorseradish peroxidase-conjugated secondary antibodieswhite negative films. High-resolution images of cellular(diluted 1:5000; Dianova, Hamburg, Germany) and en-structures were obtained with a cooled CCD camerahanced chemiluminescence (ECL; Amersham, Little(Sensys, Photometrics, Munchen, Germany) equippedChelford, UK). Baculovirus-infected Sf9 insect cells ex-with an additional fourfold magnification lens, giving apressing recombinant b1- or b2-AR served as controlsfinal magnification of 2520-fold.[13, 18, 19].
StatisticsRadioligand bindingRadioligand-binding experiments were analyzed byAll binding experiments were performed using the
nonselective b-AR antagonist [125I]-iodocyanopindolol computer-aided nonlinear regression analysis (LIGAND-

Boivin et al: b-ARs in rat kidney 517
Table 1. Binding of b-adrenergic receptor ligands to membranes prepared from rat kidney
Ligand binding sites (fmol/mg) %Receptor Ki
b
Ligand subtypea nmol/L Cortex Outer medulla Inner medulla
[125I]-CYP b1 1 b2 ND 45.562.2 (53%) 33.361.2 (39%) 7.561.8 (8%)Bisoprolol b1 7.3 30.961.4 (56%) 24.261.1 (44%) ND
b2 2100 14.660.7 (62%) 9.160.5 (38%) NDb1:b2 — 68%:32% 73%:27% ND
ICI 118,551 b1 199 29.161.1 (55%) 23.361.1 (45%) NDb2 0.3 16.460.8 (63%) 9.960.7 (37%) ND
b1:b2 — 64%:36% 70%:30% ND
The concentration of b-AR in membranes prepared from the indicated portions of rat kidney was determined by [125I]-CYP binding. The relative proportions ofb1 and b2-AR were determined by radioligand binding competition experiments using unlabeled b1- (bisoprolol) or b2-selective antagonists (ICI 118,551). The resultingbiphasic displacement curves allowed to determine a high- and low-affinity component for each b-AR subtype [25, 26]. The data given are the mean 6 SD of threeexperiments; ND, not determined.
a Deduced from the proportion of high- and low-affinity bindingb Derived from nonlinear regression analysis of [125I]-CYP-displacement curves
program) [21]. All data are given as mean 6 SD. Where with a high- and a low-affinity component, which is inperfect agreement with previous radioligand bindingquantitative statistics could not be applied (immunoflu-
orescence and immunoblotting), the examples given are studies on rat tissues [25, 26]. The relative proportionsof b1- and b2-AR derived from these curves differedrepresentative of at least three independent experiments
done on different days and with kidneys obtained from slightly between cortex (b1:b2 < 65:35%) and medulla(b1:b2 < 70:30%; Table 1), with both subtypes beingdifferent animals. Unless otherwise stated, fluorescent
images are representative of the respective area of a more frequent in cortical (b1 5 56% and b2 5 62%)than in medullary membranes (b1 5 44% and b2 5 38%;section inspected in at least ten separate fields of view.
The schemes given throughout this article depict simpli- Table 1). It should be noted that within a same kidneyzone, similar proportions of b1- and b2-AR were obtainedfied nephrons not drawn to scale (modified from Kriz and
Kaissling [22]) and do not represent the exact anatomy of with bisoprolol and with ICI 118,551.From the subtype-specific receptor antibodies thatthe rat nephron (that is, short looped vs. long looped).
were raised against selected extramembraneous domainsof the human b1- or b2-AR [13], those directed against the
RESULTSamino-terminal domains exhibited the highest degree of
Characterization of b1- and b2-adrenergic receptors in specificity and subtype selectivity in cell systems express-rat kidney by radioligand binding and immunoblotting ing recombinant human b-AR. Since these domains are
highly conserved between humans [16, 17] and ratsTo define our experimental system, we determined theamount and subtype distribution of b-ARs in rat kidney [14, 15], we expected that these antibodies would be
suitable for immunohistological studies on rat tissues.by conventional radioligand binding. For this purpose,we separated cortical, outer, and inner medullary por- However, before embarking on such studies, this as-
sumption was tested by probing Western blots of mem-tions of the kidney by macroscopic dissection, isolatedthe cell membranes from these portions, and measured branes prepared from cortical and medullary portions
of rat kidney. Antibodies against the b1-AR (anti–b1-AR)the overall concentration of membrane-bound b-AR byusing the nonselective receptor antagonist [125I]-cyano- labeled a single protein band with an apparent size of
68 kD (Fig. 1, lanes 1 through 3), which is in good agree-pindolol. In agreement with previous reports [7, 23], weobserved the highest density of b-AR in cortical mem- ment with the molecular mass deduced from the amino
acid sequence of the rat b1-AR [14], when taking intobranes (53%) and somewhat lower receptor densitiesin membranes prepared from the outer medulla (39%; account some variations in electrophoretic mobility
caused by glycosylation [27]. The apparent decrease inTable 1). Because only a rather faint amount of b-ARwas found in the inner medulla (8%), in this portion, signal intensity from the cortex to the inner medulla is
in good correlation with the receptor densities in thea further subtyping of the receptors was not meaning-ful. In the cortex and outer medulla, the proportion of corresponding kidney zones determined by radioligand
binding (Table 1) and reflects the different expressionb1- and b2-AR subtypes was determined by competing[125I]-cyanopindolol binding with unlabeled subtype- of this receptor subtype in the corresponding kidney
zones. Antibodies against the b2-AR (anti–b2-AR) la-selective receptor antagonists, that is, using bisoprololfor the b1-AR [24] and ICI 118,551 for the b2-AR sub- beled a protein band of slightly higher mobility (Fig. 1,
lanes 6 through 8), which is also plausible in the lighttype, respectively. In each case, there were biphasic dis-placement curves suggestive of a two-sited binding model of the (slightly shorter) amino acid sequence of the rat

Boivin et al: b-ARs in rat kidney518
munoreactivity of smooth muscle in a consecutive sec-tion was largely abolished by preadsorption with recom-binant b1-AR expressed in Sf9 insect cells (Fig. 2b). Athigher magnification, the antibody label was seen to bemainly confined to small dot-like structures located atthe cell surface (Fig. 2d, arrows), which corresponds tothe b-AR staining pattern previously observed in cul-tured human epidermoid carcinoma A 431 cells [13].Vascular smooth muscle cells were also stained by
Fig. 1. Immunoblots of b1- and b2-adrenergic receptors (b1- and b2-AR)anti–b2-AR (albeit somewhat less intense; Fig. 2c), andin rat kidney. Membranes prepared from cortical (lanes 1 and 6), outer
medullary (lanes 2 and 7), and inner medullary (lanes 3 and 8) portions this staining could likewise be abolished by preadsorp-of rat kidney were subjected to Western blotting and probed with tion with recombinant b2-AR (data not shown).antibodies against b1-AR (lanes 1 through 5) or b2-AR (lanes 6 through
Glomeruli. By immunohistochemistry, several loca-10). Bound antibodies were detected by horseradish peroxidase-conju-gated secondary antibodies and enhanced chemiluminescence. Mem- tions within and adjacent to the glomerulus could bebranes prepared from Sf9 cells expressing recombinant b1- (49 kD; lane identified, where b1-AR appeared to be clustered: a very4 and 10) or b2-AR (47 kD; lane 5 and 9) served as specificity controls
strong immunostaining by anti–b1-AR was observed in(note that receptor glycosylation is known to be different and heteroge-nous in Sf9 insect cells) [13, 27]. Apparent molecular sizes (kD) are the juxtaglomerular portion of the afferent arteriole (Fig.denoted on the right margin. 3a). These immunofluorescent signals could be clearly
assigned to the renin-producing juxtaglomerular granu-lar cells themselves by staining of serial sections withrenin-antibodies (Fig. 3, compare circled areas in a andb2-AR [15] when taking protein glycosylation into ac-b). In addition, b1-AR appeared to be densely expressedcount. In this context, it should be noted that b-AR arein the walls of arterioles entering the glomeruli, whichknown to be less glycosylated when expressed in Sf9could be assigned to the afferent arterioles by their inter-insect cells [13, 18], giving protein bands with apparentnal elastic membrane (visible by green autofluores-sizes of 51 and 47 kD for the recombinant human b1-cence). Anti–b1-AR also highlighted banded structuresand b2-AR, respectively (Fig. 1, lanes 4 and 9). Thesein the glomerular tuft, which seemed to be interspaceddata indicated that these antibodies did recognize rat b1-between the capillaries (Fig. 4a, c, e, g, i). These signalsand b2-AR, and thus they appeared to be suitable forseemed to be specific for b1-AR in as much as they couldan immunohistological study on rat kidney.be abolished by preadsorption with recombinant b1-AR
Immunolocalization of b1- and b2-adrenergic receptors (Fig. 4, compare c and d). Such structures were not la-in rat kidney beled by anti–b2-AR (Fig. 4, compare a and b), which
reacted with only certain tubular structures adjacent toProcedural strategy. Initial experiments were carriedthe glomerulus, as will be discussed later. Moreover,out on cryostat sections (3 to 4 mm) of rat kidney, whichthe banded pattern of b1-AR in the glomerular tuft waswere subsequently fixed with acetone. Staining of suchclearly different from that obtained with antibodiessections with antibodies against b-AR led to a bright andagainst endothelial nitric oxide synthetase (eNOS) as adistinct labeling of vascular and glomerular structures,specific marker of capillaries (Fig. 4, compare e with f)which could readily be interpreted at the cellular level.and could not be assigned to podocytes by counter-Certain tubular structures within the cortical and medul-staining with synaptopodin antibodies (Fig. 4, comparelary nephron segments were also labeled by the antibod-g with h). Considering that within the glomerular tuft,ies, but were not sufficiently conserved for an interpreta-only three cell types are found (endothelial cells, podo-tion at the cellular level. Immunohistochemistry of thesecytes, and mesangial cells) [22] and that the signal ob-structures had to be carried out on sections of kidneystained with anti–b1-AR could not be associated withfixed in situ by formaldehyde perfusion. However, inendothelial cells or with podocytes, most likely it is asso-such perfusion-fixed sections, the staining of vascularciated with mesangial cells. This argument is further sup-and glomerular structures was generally much less in-ported by high resolution images of the glomerular capil-tense, most likely because these tissue compartments arelary tuft (Fig. 4i, k), suggesting that b1-AR are situatedmore strongly fixed by vascular perfusion than extravas-in the membranes of mesangial cells that are clusteredcular tissues.in the intercapillary space and extend processes joiningBlood vessels. The cell types most brilliantly stainedthe capillary walls. Most likely, the latter structures cor-by antibodies against b1- and b2-AR in rat kidney sectionsrespond to mesangial cell processes extending towardwere the smooth muscle cells in the walls of large bloodthe peripherally located capillaries (Fig. 4, compare ivessels. Figure 2a shows a representative oblique section
through an arcuate artery stained with anti–b1-AR. Im- and k).
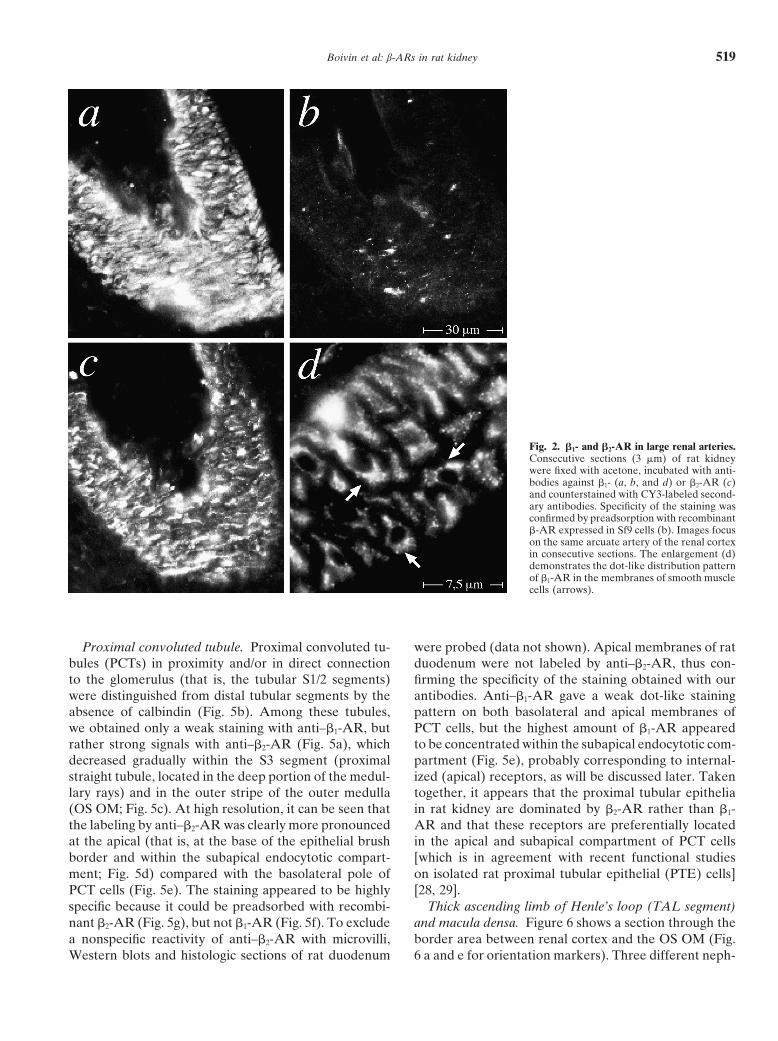
Boivin et al: b-ARs in rat kidney 519
Fig. 2. b1- and b2-AR in large renal arteries.Consecutive sections (3 mm) of rat kidneywere fixed with acetone, incubated with anti-bodies against b1- (a, b, and d) or b2-AR (c)and counterstained with CY3-labeled second-ary antibodies. Specificity of the staining wasconfirmed by preadsorption with recombinantb-AR expressed in Sf9 cells (b). Images focuson the same arcuate artery of the renal cortexin consecutive sections. The enlargement (d)demonstrates the dot-like distribution patternof b1-AR in the membranes of smooth musclecells (arrows).
Proximal convoluted tubule. Proximal convoluted tu- were probed (data not shown). Apical membranes of ratduodenum were not labeled by anti–b2-AR, thus con-bules (PCTs) in proximity and/or in direct connection
to the glomerulus (that is, the tubular S1/2 segments) firming the specificity of the staining obtained with ourantibodies. Anti–b1-AR gave a weak dot-like stainingwere distinguished from distal tubular segments by the
absence of calbindin (Fig. 5b). Among these tubules, pattern on both basolateral and apical membranes ofPCT cells, but the highest amount of b1-AR appearedwe obtained only a weak staining with anti–b1-AR, but
rather strong signals with anti–b2-AR (Fig. 5a), which to be concentrated within the subapical endocytotic com-partment (Fig. 5e), probably corresponding to internal-decreased gradually within the S3 segment (proximal
straight tubule, located in the deep portion of the medul- ized (apical) receptors, as will be discussed later. Takentogether, it appears that the proximal tubular epithelialary rays) and in the outer stripe of the outer medulla
(OS OM; Fig. 5c). At high resolution, it can be seen that in rat kidney are dominated by b2-AR rather than b1-AR and that these receptors are preferentially locatedthe labeling by anti–b2-AR was clearly more pronounced
at the apical (that is, at the base of the epithelial brush in the apical and subapical compartment of PCT cells[which is in agreement with recent functional studiesborder and within the subapical endocytotic compart-
ment; Fig. 5d) compared with the basolateral pole of on isolated rat proximal tubular epithelial (PTE) cells][28, 29].PCT cells (Fig. 5e). The staining appeared to be highly
specific because it could be preadsorbed with recombi- Thick ascending limb of Henle’s loop (TAL segment)and macula densa. Figure 6 shows a section through thenant b2-AR (Fig. 5g), but not b1-AR (Fig. 5f). To exclude
a nonspecific reactivity of anti–b2-AR with microvilli, border area between renal cortex and the OS OM (Fig.6 a and e for orientation markers). Three different neph-Western blots and histologic sections of rat duodenum
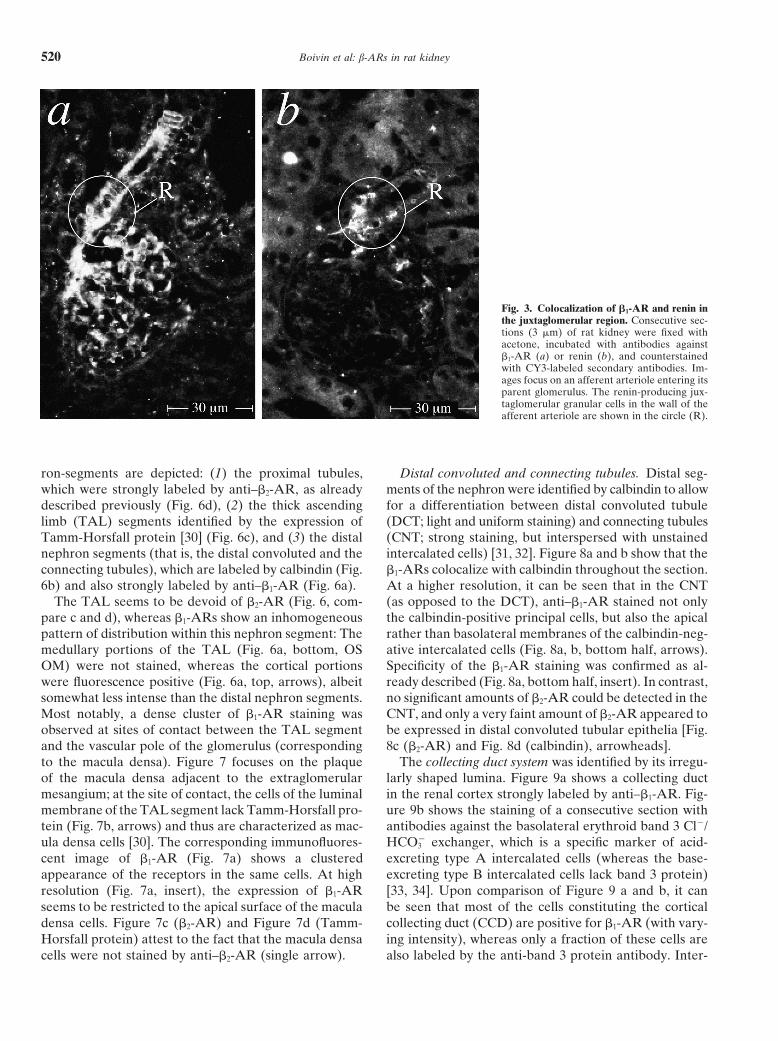
Boivin et al: b-ARs in rat kidney520
Fig. 3. Colocalization of b1-AR and renin inthe juxtaglomerular region. Consecutive sec-tions (3 mm) of rat kidney were fixed withacetone, incubated with antibodies againstb1-AR (a) or renin (b), and counterstainedwith CY3-labeled secondary antibodies. Im-ages focus on an afferent arteriole entering itsparent glomerulus. The renin-producing jux-taglomerular granular cells in the wall of theafferent arteriole are shown in the circle (R).
ron-segments are depicted: (1) the proximal tubules, Distal convoluted and connecting tubules. Distal seg-ments of the nephron were identified by calbindin to allowwhich were strongly labeled by anti–b2-AR, as already
described previously (Fig. 6d), (2) the thick ascending for a differentiation between distal convoluted tubule(DCT; light and uniform staining) and connecting tubuleslimb (TAL) segments identified by the expression of
Tamm-Horsfall protein [30] (Fig. 6c), and (3) the distal (CNT; strong staining, but interspersed with unstainedintercalated cells) [31, 32]. Figure 8a and b show that thenephron segments (that is, the distal convoluted and the
connecting tubules), which are labeled by calbindin (Fig. b1-ARs colocalize with calbindin throughout the section.At a higher resolution, it can be seen that in the CNT6b) and also strongly labeled by anti–b1-AR (Fig. 6a).
The TAL seems to be devoid of b2-AR (Fig. 6, com- (as opposed to the DCT), anti–b1-AR stained not onlythe calbindin-positive principal cells, but also the apicalpare c and d), whereas b1-ARs show an inhomogeneous
pattern of distribution within this nephron segment: The rather than basolateral membranes of the calbindin-neg-ative intercalated cells (Fig. 8a, b, bottom half, arrows).medullary portions of the TAL (Fig. 6a, bottom, OS
OM) were not stained, whereas the cortical portions Specificity of the b1-AR staining was confirmed as al-ready described (Fig. 8a, bottom half, insert). In contrast,were fluorescence positive (Fig. 6a, top, arrows), albeit
somewhat less intense than the distal nephron segments. no significant amounts of b2-AR could be detected in theCNT, and only a very faint amount of b2-AR appeared toMost notably, a dense cluster of b1-AR staining was
observed at sites of contact between the TAL segment be expressed in distal convoluted tubular epithelia [Fig.8c (b2-AR) and Fig. 8d (calbindin), arrowheads].and the vascular pole of the glomerulus (corresponding
to the macula densa). Figure 7 focuses on the plaque The collecting duct system was identified by its irregu-larly shaped lumina. Figure 9a shows a collecting ductof the macula densa adjacent to the extraglomerular
mesangium; at the site of contact, the cells of the luminal in the renal cortex strongly labeled by anti–b1-AR. Fig-ure 9b shows the staining of a consecutive section withmembrane of the TAL segment lack Tamm-Horsfall pro-
tein (Fig. 7b, arrows) and thus are characterized as mac- antibodies against the basolateral erythroid band 3 Cl2/HCO2
3 exchanger, which is a specific marker of acid-ula densa cells [30]. The corresponding immunofluores-cent image of b1-AR (Fig. 7a) shows a clustered excreting type A intercalated cells (whereas the base-
excreting type B intercalated cells lack band 3 protein)appearance of the receptors in the same cells. At highresolution (Fig. 7a, insert), the expression of b1-AR [33, 34]. Upon comparison of Figure 9 a and b, it can
be seen that most of the cells constituting the corticalseems to be restricted to the apical surface of the maculadensa cells. Figure 7c (b2-AR) and Figure 7d (Tamm- collecting duct (CCD) are positive for b1-AR (with vary-
ing intensity), whereas only a fraction of these cells areHorsfall protein) attest to the fact that the macula densacells were not stained by anti–b2-AR (single arrow). also labeled by the anti-band 3 protein antibody. Inter-
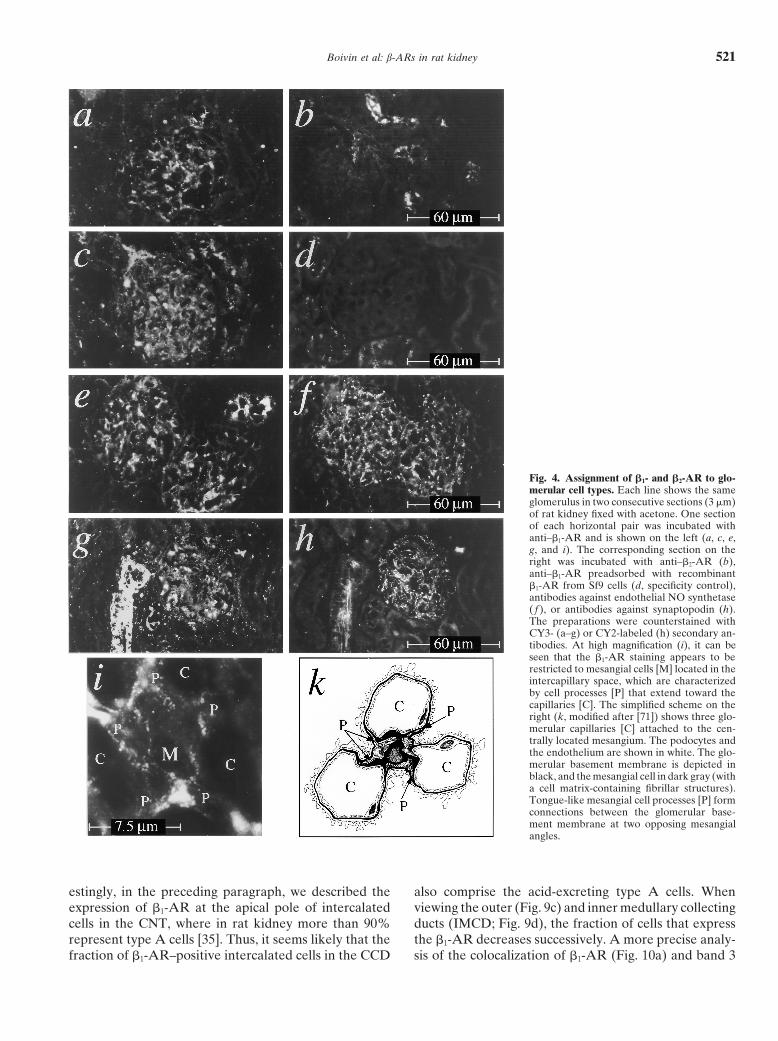
Boivin et al: b-ARs in rat kidney 521
Fig. 4. Assignment of b1- and b2-AR to glo-merular cell types. Each line shows the sameglomerulus in two consecutive sections (3 mm)of rat kidney fixed with acetone. One sectionof each horizontal pair was incubated withanti–b1-AR and is shown on the left (a, c, e,g, and i). The corresponding section on theright was incubated with anti–b2-AR (b),anti–b1-AR preadsorbed with recombinantb1-AR from Sf9 cells (d, specificity control),antibodies against endothelial NO synthetase( f ), or antibodies against synaptopodin (h).The preparations were counterstained withCY3- (a–g) or CY2-labeled (h) secondary an-tibodies. At high magnification (i), it can beseen that the b1-AR staining appears to berestricted to mesangial cells [M] located in theintercapillary space, which are characterizedby cell processes [P] that extend toward thecapillaries [C]. The simplified scheme on theright (k, modified after [71]) shows three glo-merular capillaries [C] attached to the cen-trally located mesangium. The podocytes andthe endothelium are shown in white. The glo-merular basement membrane is depicted inblack, and the mesangial cell in dark gray (witha cell matrix-containing fibrillar structures).Tongue-like mesangial cell processes [P] formconnections between the glomerular base-ment membrane at two opposing mesangialangles.
estingly, in the preceding paragraph, we described the also comprise the acid-excreting type A cells. Whenviewing the outer (Fig. 9c) and inner medullary collectingexpression of b1-AR at the apical pole of intercalated
cells in the CNT, where in rat kidney more than 90% ducts (IMCD; Fig. 9d), the fraction of cells that expressthe b1-AR decreases successively. A more precise analy-represent type A cells [35]. Thus, it seems likely that the
fraction of b1-AR–positive intercalated cells in the CCD sis of the colocalization of b1-AR (Fig. 10a) and band 3
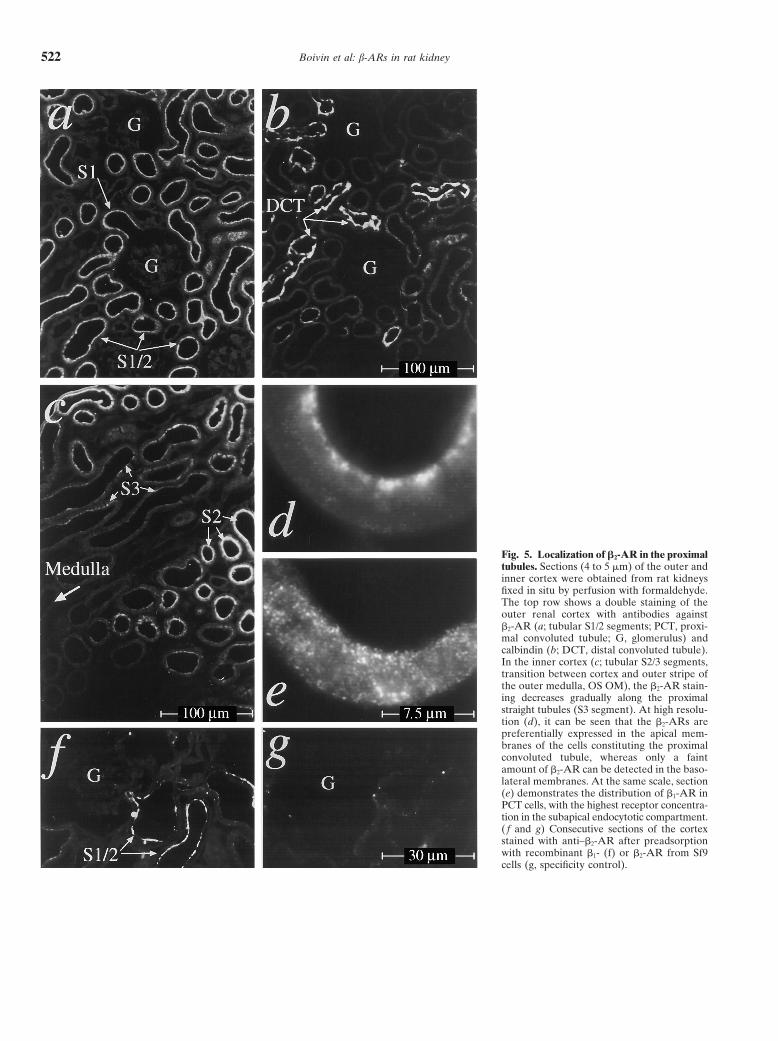
Boivin et al: b-ARs in rat kidney522
Fig. 5. Localization of b2-AR in the proximaltubules. Sections (4 to 5 mm) of the outer andinner cortex were obtained from rat kidneysfixed in situ by perfusion with formaldehyde.The top row shows a double staining of theouter renal cortex with antibodies againstb2-AR (a; tubular S1/2 segments; PCT, proxi-mal convoluted tubule; G, glomerulus) andcalbindin (b; DCT, distal convoluted tubule).In the inner cortex (c; tubular S2/3 segments,transition between cortex and outer stripe ofthe outer medulla, OS OM), the b2-AR stain-ing decreases gradually along the proximalstraight tubules (S3 segment). At high resolu-tion (d), it can be seen that the b2-ARs arepreferentially expressed in the apical mem-branes of the cells constituting the proximalconvoluted tubule, whereas only a faintamount of b2-AR can be detected in the baso-lateral membranes. At the same scale, section(e) demonstrates the distribution of b1-AR inPCT cells, with the highest receptor concentra-tion in the subapical endocytotic compartment.( f and g) Consecutive sections of the cortexstained with anti–b2-AR after preadsorptionwith recombinant b1- (f) or b2-AR from Sf9cells (g, specificity control).
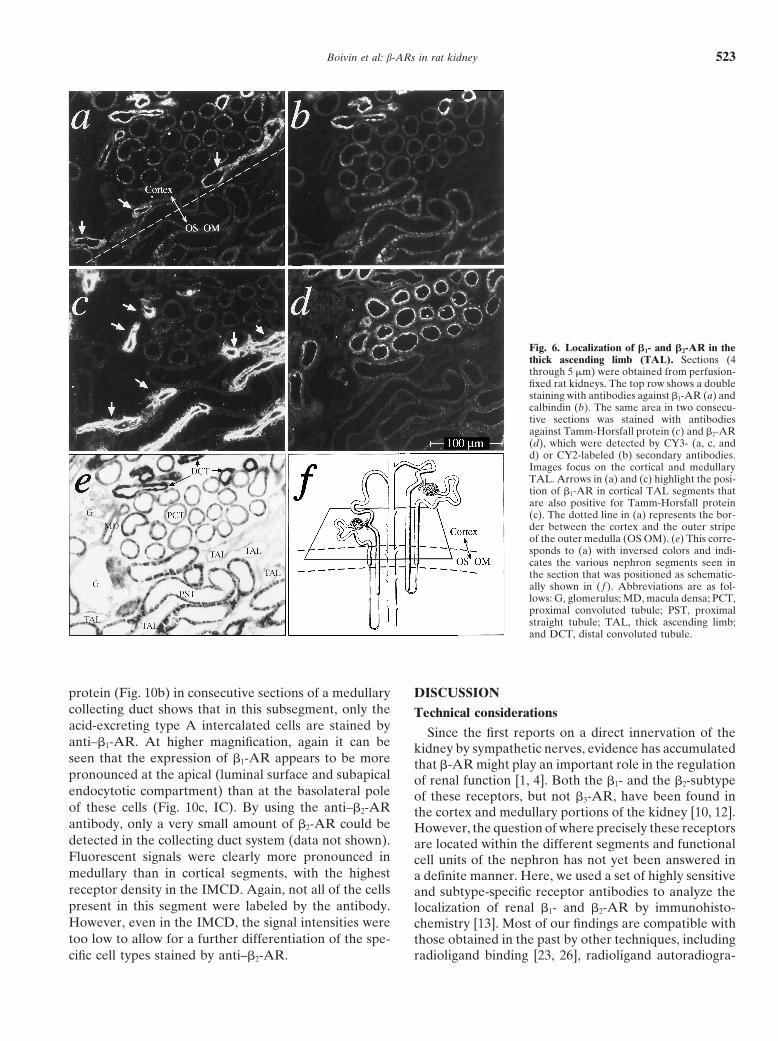
Boivin et al: b-ARs in rat kidney 523
Fig. 6. Localization of b1- and b2-AR in thethick ascending limb (TAL). Sections (4through 5 mm) were obtained from perfusion-fixed rat kidneys. The top row shows a doublestaining with antibodies against b1-AR (a) andcalbindin (b). The same area in two consecu-tive sections was stained with antibodiesagainst Tamm-Horsfall protein (c) and b2-AR(d), which were detected by CY3- (a, c, andd) or CY2-labeled (b) secondary antibodies.Images focus on the cortical and medullaryTAL. Arrows in (a) and (c) highlight the posi-tion of b1-AR in cortical TAL segments thatare also positive for Tamm-Horsfall protein(c). The dotted line in (a) represents the bor-der between the cortex and the outer stripeof the outer medulla (OS OM). (e) This corre-sponds to (a) with inversed colors and indi-cates the various nephron segments seen inthe section that was positioned as schematic-ally shown in ( f ). Abbreviations are as fol-lows: G, glomerulus; MD, macula densa; PCT,proximal convoluted tubule; PST, proximalstraight tubule; TAL, thick ascending limb;and DCT, distal convoluted tubule.
protein (Fig. 10b) in consecutive sections of a medullary DISCUSSIONcollecting duct shows that in this subsegment, only the Technical considerationsacid-excreting type A intercalated cells are stained by Since the first reports on a direct innervation of theanti–b1-AR. At higher magnification, again it can be kidney by sympathetic nerves, evidence has accumulatedseen that the expression of b1-AR appears to be more that b-AR might play an important role in the regulationpronounced at the apical (luminal surface and subapical of renal function [1, 4]. Both the b1- and the b2-subtypeendocytotic compartment) than at the basolateral pole of these receptors, but not b3-AR, have been found inof these cells (Fig. 10c, IC). By using the anti–b2-AR the cortex and medullary portions of the kidney [10, 12].antibody, only a very small amount of b2-AR could be However, the question of where precisely these receptorsdetected in the collecting duct system (data not shown). are located within the different segments and functionalFluorescent signals were clearly more pronounced in cell units of the nephron has not yet been answered inmedullary than in cortical segments, with the highest a definite manner. Here, we used a set of highly sensitivereceptor density in the IMCD. Again, not all of the cells and subtype-specific receptor antibodies to analyze thepresent in this segment were labeled by the antibody. localization of renal b1- and b2-AR by immunohisto-However, even in the IMCD, the signal intensities were chemistry [13]. Most of our findings are compatible withtoo low to allow for a further differentiation of the spe- those obtained in the past by other techniques, including
radioligand binding [23, 26], radioligand autoradiogra-cific cell types stained by anti–b2-AR.
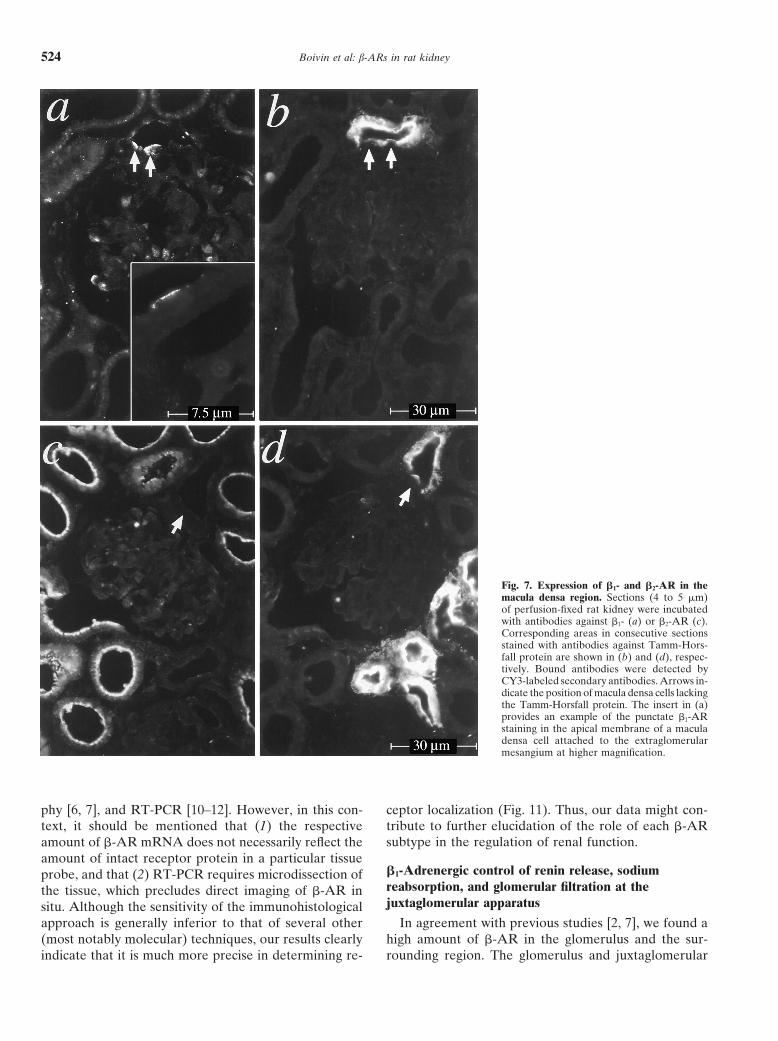
Boivin et al: b-ARs in rat kidney524
Fig. 7. Expression of b1- and b2-AR in themacula densa region. Sections (4 to 5 mm)of perfusion-fixed rat kidney were incubatedwith antibodies against b1- (a) or b2-AR (c).Corresponding areas in consecutive sectionsstained with antibodies against Tamm-Hors-fall protein are shown in (b) and (d), respec-tively. Bound antibodies were detected byCY3-labeled secondary antibodies. Arrows in-dicate the position of macula densa cells lackingthe Tamm-Horsfall protein. The insert in (a)provides an example of the punctate b1-ARstaining in the apical membrane of a maculadensa cell attached to the extraglomerularmesangium at higher magnification.
phy [6, 7], and RT-PCR [10–12]. However, in this con- ceptor localization (Fig. 11). Thus, our data might con-tribute to further elucidation of the role of each b-ARtext, it should be mentioned that (1) the respectivesubtype in the regulation of renal function.amount of b-AR mRNA does not necessarily reflect the
amount of intact receptor protein in a particular tissueb1-Adrenergic control of renin release, sodiumprobe, and that (2) RT-PCR requires microdissection ofreabsorption, and glomerular filtration at thethe tissue, which precludes direct imaging of b-AR injuxtaglomerular apparatussitu. Although the sensitivity of the immunohistological
approach is generally inferior to that of several other In agreement with previous studies [2, 7], we found ahigh amount of b-AR in the glomerulus and the sur-(most notably molecular) techniques, our results clearly
indicate that it is much more precise in determining re- rounding region. The glomerulus and juxtaglomerular
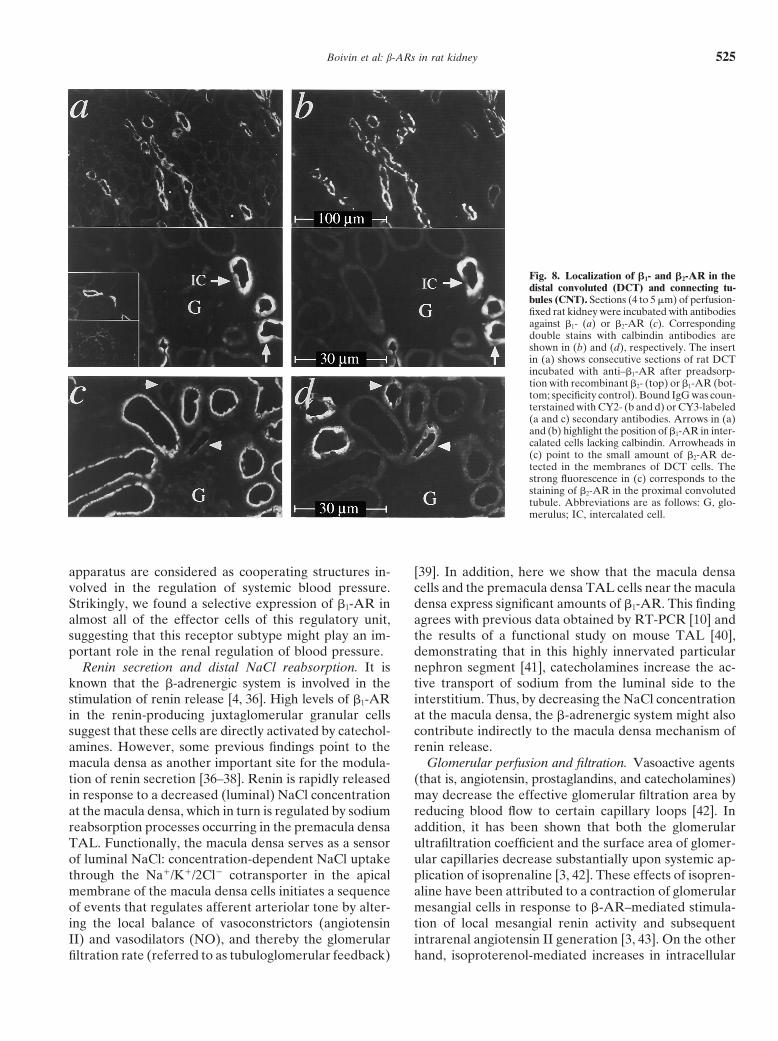
Boivin et al: b-ARs in rat kidney 525
Fig. 8. Localization of b1- and b2-AR in thedistal convoluted (DCT) and connecting tu-bules (CNT). Sections (4 to 5 mm) of perfusion-fixed rat kidney were incubated with antibodiesagainst b1- (a) or b2-AR (c). Correspondingdouble stains with calbindin antibodies areshown in (b) and (d), respectively. The insertin (a) shows consecutive sections of rat DCTincubated with anti–b1-AR after preadsorp-tion with recombinant b2- (top) or b1-AR (bot-tom; specificity control). Bound IgG was coun-terstained with CY2- (b and d) or CY3-labeled(a and c) secondary antibodies. Arrows in (a)and (b) highlight the position of b1-AR in inter-calated cells lacking calbindin. Arrowheads in(c) point to the small amount of b2-AR de-tected in the membranes of DCT cells. Thestrong fluorescence in (c) corresponds to thestaining of b2-AR in the proximal convolutedtubule. Abbreviations are as follows: G, glo-merulus; IC, intercalated cell.
apparatus are considered as cooperating structures in- [39]. In addition, here we show that the macula densacells and the premacula densa TAL cells near the maculavolved in the regulation of systemic blood pressure.
Strikingly, we found a selective expression of b1-AR in densa express significant amounts of b1-AR. This findingagrees with previous data obtained by RT-PCR [10] andalmost all of the effector cells of this regulatory unit,
suggesting that this receptor subtype might play an im- the results of a functional study on mouse TAL [40],demonstrating that in this highly innervated particularportant role in the renal regulation of blood pressure.
Renin secretion and distal NaCl reabsorption. It is nephron segment [41], catecholamines increase the ac-tive transport of sodium from the luminal side to theknown that the b-adrenergic system is involved in the
stimulation of renin release [4, 36]. High levels of b1-AR interstitium. Thus, by decreasing the NaCl concentrationat the macula densa, the b-adrenergic system might alsoin the renin-producing juxtaglomerular granular cells
suggest that these cells are directly activated by catechol- contribute indirectly to the macula densa mechanism ofrenin release.amines. However, some previous findings point to the
macula densa as another important site for the modula- Glomerular perfusion and filtration. Vasoactive agents(that is, angiotensin, prostaglandins, and catecholamines)tion of renin secretion [36–38]. Renin is rapidly released
in response to a decreased (luminal) NaCl concentration may decrease the effective glomerular filtration area byreducing blood flow to certain capillary loops [42]. Inat the macula densa, which in turn is regulated by sodium
reabsorption processes occurring in the premacula densa addition, it has been shown that both the glomerularultrafiltration coefficient and the surface area of glomer-TAL. Functionally, the macula densa serves as a sensor
of luminal NaCl: concentration-dependent NaCl uptake ular capillaries decrease substantially upon systemic ap-plication of isoprenaline [3, 42]. These effects of isopren-through the Na1/K1/2Cl2 cotransporter in the apical
membrane of the macula densa cells initiates a sequence aline have been attributed to a contraction of glomerularmesangial cells in response to b-AR–mediated stimula-of events that regulates afferent arteriolar tone by alter-
ing the local balance of vasoconstrictors (angiotensin tion of local mesangial renin activity and subsequentintrarenal angiotensin II generation [3, 43]. On the otherII) and vasodilators (NO), and thereby the glomerular
filtration rate (referred to as tubuloglomerular feedback) hand, isoproterenol-mediated increases in intracellular
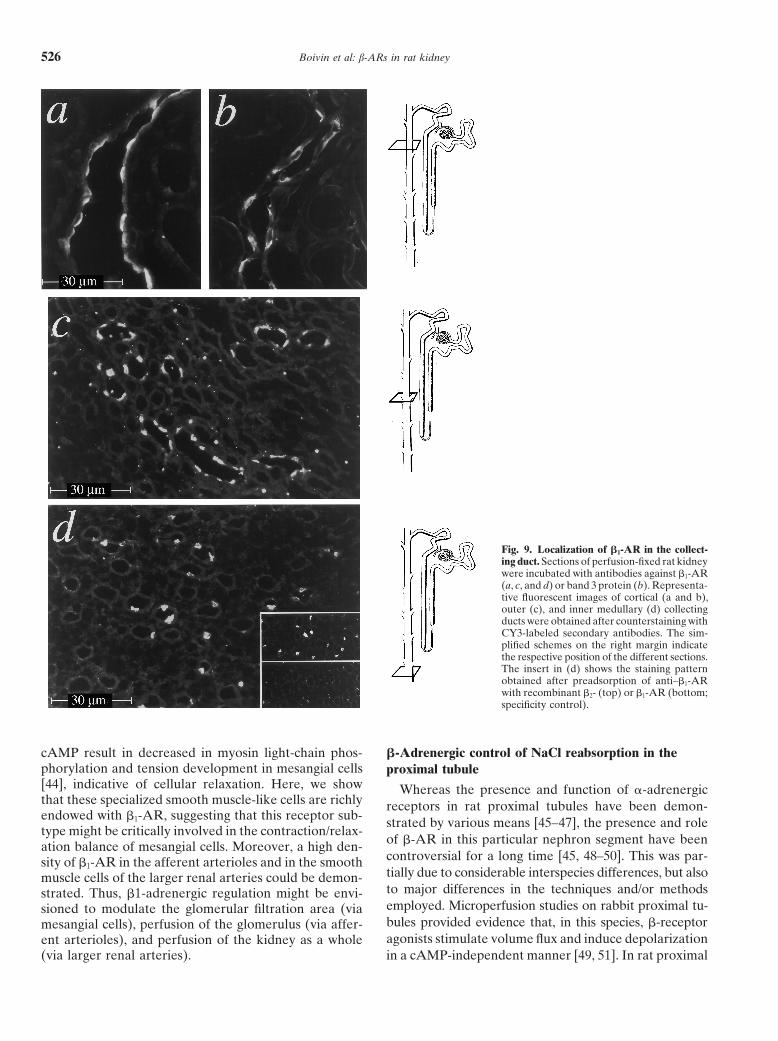
Boivin et al: b-ARs in rat kidney526
Fig. 9. Localization of b1-AR in the collect-ing duct. Sections of perfusion-fixed rat kidneywere incubated with antibodies against b1-AR(a, c, and d) or band 3 protein (b). Representa-tive fluorescent images of cortical (a and b),outer (c), and inner medullary (d) collectingducts were obtained after counterstaining withCY3-labeled secondary antibodies. The sim-plified schemes on the right margin indicatethe respective position of the different sections.The insert in (d) shows the staining patternobtained after preadsorption of anti–b1-ARwith recombinant b2- (top) or b1-AR (bottom;specificity control).
cAMP result in decreased in myosin light-chain phos- b-Adrenergic control of NaCl reabsorption in thephorylation and tension development in mesangial cells proximal tubule[44], indicative of cellular relaxation. Here, we show Whereas the presence and function of a-adrenergicthat these specialized smooth muscle-like cells are richly receptors in rat proximal tubules have been demon-endowed with b1-AR, suggesting that this receptor sub- strated by various means [45–47], the presence and roletype might be critically involved in the contraction/relax-
of b-AR in this particular nephron segment have beenation balance of mesangial cells. Moreover, a high den-controversial for a long time [45, 48–50]. This was par-sity of b1-AR in the afferent arterioles and in the smoothtially due to considerable interspecies differences, but alsomuscle cells of the larger renal arteries could be demon-to major differences in the techniques and/or methodsstrated. Thus, b1-adrenergic regulation might be envi-employed. Microperfusion studies on rabbit proximal tu-sioned to modulate the glomerular filtration area (viabules provided evidence that, in this species, b-receptormesangial cells), perfusion of the glomerulus (via affer-agonists stimulate volume flux and induce depolarizationent arterioles), and perfusion of the kidney as a whole
(via larger renal arteries). in a cAMP-independent manner [49, 51]. In rat proximal
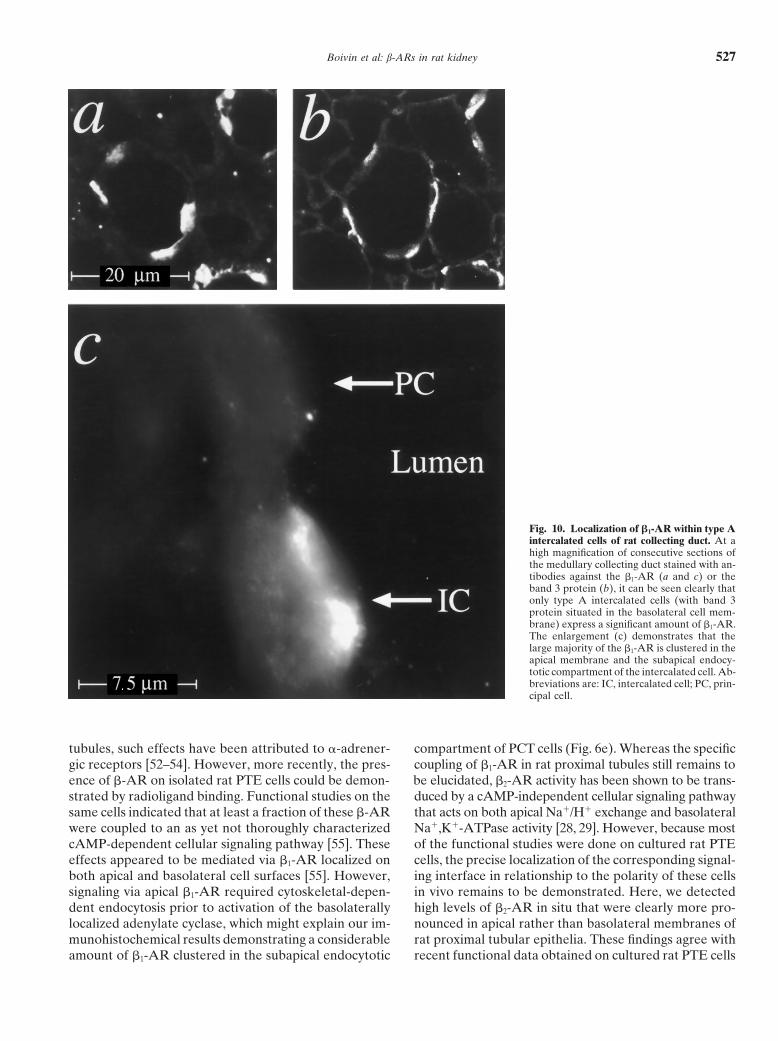
Boivin et al: b-ARs in rat kidney 527
Fig. 10. Localization of b1-AR within type Aintercalated cells of rat collecting duct. At ahigh magnification of consecutive sections ofthe medullary collecting duct stained with an-tibodies against the b1-AR (a and c) or theband 3 protein (b), it can be seen clearly thatonly type A intercalated cells (with band 3protein situated in the basolateral cell mem-brane) express a significant amount of b1-AR.The enlargement (c) demonstrates that thelarge majority of the b1-AR is clustered in theapical membrane and the subapical endocy-totic compartment of the intercalated cell. Ab-breviations are: IC, intercalated cell; PC, prin-cipal cell.
tubules, such effects have been attributed to a-adrener- compartment of PCT cells (Fig. 6e). Whereas the specificcoupling of b1-AR in rat proximal tubules still remains togic receptors [52–54]. However, more recently, the pres-
ence of b-AR on isolated rat PTE cells could be demon- be elucidated, b2-AR activity has been shown to be trans-duced by a cAMP-independent cellular signaling pathwaystrated by radioligand binding. Functional studies on the
same cells indicated that at least a fraction of these b-AR that acts on both apical Na1/H1 exchange and basolateralNa1,K1-ATPase activity [28, 29]. However, because mostwere coupled to an as yet not thoroughly characterized
cAMP-dependent cellular signaling pathway [55]. These of the functional studies were done on cultured rat PTEcells, the precise localization of the corresponding signal-effects appeared to be mediated via b1-AR localized on
both apical and basolateral cell surfaces [55]. However, ing interface in relationship to the polarity of these cellsin vivo remains to be demonstrated. Here, we detectedsignaling via apical b1-AR required cytoskeletal-depen-
dent endocytosis prior to activation of the basolaterally high levels of b2-AR in situ that were clearly more pro-nounced in apical rather than basolateral membranes oflocalized adenylate cyclase, which might explain our im-
munohistochemical results demonstrating a considerable rat proximal tubular epithelia. These findings agree withrecent functional data obtained on cultured rat PTE cellsamount of b1-AR clustered in the subapical endocytotic

Boivin et al: b-ARs in rat kidney528
Fig. 11. Simplified scheme indicating the ma-jor locations of b1- and b2-AR in rat nephron,as determined by immunohistochemistry. Sig-nificant amounts of b1-AR were found in renin-producing juxtaglomerular granular cells (inthe wall of the afferent arteriole), mesangialcells, proximal tubular cells, cortical TAL seg-ments (including the macula densa cells), allportions of the distal tubular nephron segments,and the intercalated cells of the collecting ductsystem (depicted as dark dots). b2-ARs werefound in the membranes of proximal tubularepithelia and, to a lesser extent, in the distalconvoluted tubules and the medullary collect-ing ducts.
demonstrating that (1) selective stimulation of b2-AR b1-Adrenergic receptors in the distal segmentsof the nephronwith metaproterenol increases both apical Na1/H1 ex-
change and basolateral Na1,K1-ATPase activity, and (2) In agreement with previous data obtained by radioli-that these effects are most probably mediated through gand binding and autoradiography [6, 7, 12], we demon-apical rather than basolateral b2-AR [29]. strate here that b1-ARs are predominant in all distal
The S1 (and S2) segments of the PCT are known to segments of the rat nephron. Again, the expression ofbe less densely innervated [56]. Therefore, it has been this subtype was more pronounced at the apical surface
of the cells forming the DCT. In the CNT, a similar levelsuggested that catecholamines gain access to their recep-of b1-AR was found in the membranes of both cell typestors on the tubular epithelia mainly via the cellular inter-that constitute this segment, that is, the principal cellsstitium and/or the circulation [1, 52], suggesting a pre-and the intercalated cells, whereas in the initial portionsdominance of basolaterally localized b-AR. However,of the CCD the b1-AR seemed to be expressed preferen-our study shows that an important fraction of b1-AR andtially in the intercalated cells. We also identified somethe large majority of b2-AR are located in the apical cellb2-AR in the membranes of DCT cells, but neither inmembrane. One possible explanation for this peculiarthe CNT nor in the initial portions of the CCD wasorientation would be that the receptors are meant toa significant amount of this receptor subtype detected.interact with catecholamines gaining access to the tubu-Previous studies on cell lines derived from the DCTlar fluid. This assumption is supported further by phar-suggest that the b-AR of these cells are functionallymacological studies on rat kidney, which have clearlycoupled to the adenylate cyclase and stimulate sodiumshown that apical tubular surfaces are exposed to cate-reabsorption [12]. Moreover, direct innervation of the
cholamines (and their methylated derivatives) eitherdistal tubule has been demonstrated [1, 58], suggesting
through glomerular filtration or through direct secretion that the b-ARs in these segments are stimulated by cate-of these substances by organic cation transporters from cholamines released from synapses. However, in addi-the peritubular capillaries into the tubular luminal fluid tion to the predicted basolateral localization, our study[52, 53, 57]. In addition, from functional studies on iso- shows that the majority of these receptors appear to belated rat PTE cells, it was concluded that cellular signal- expressed in the apical membranes of DCT cells, whiching through apical b-AR is strictly mediated by the apical again leads to speculations about a functional relevanceb-AR themselves rather than by paracellular ligand leak of tubular fluid catecholamines.or transcellular ligand transport [55]. Taken together, it
b-Adrenergic receptors in the collecting duct systemseems conceivable that both apical b1- and b2-AR areinvolved in the regulation of the NaCl and fluid balance High amounts of b1-AR were found in the collecting
duct system, which is composed of the distal portionsin the proximal tubular nephron segments.

Boivin et al: b-ARs in rat kidney 529
of the CCD, and the outer medullary collecting duct line activates both a cAMP-dependent (increase in baso-(OMCD) and IMCD, respectively. In the medullary col- lateral Cl2/HCO2
3 exchange) and a cAMP-independentlecting duct, we also detected some b2-AR, but failed to cellular signaling pathway (increase in apical H1-ATPasedetect this receptor subtype in the CCD. This observa- activity), which resulted in increased luminal H1 secre-tion seems to disagree with a previous study demonstra- tion. Our study revealed that in rats the b1-AR subtypeting equal amounts of b1- and b2-AR–specific mRNA was clearly restricted to the acid-excreting type A inter-in microdissected tissue samples of rat CCD [11]. The calated cells, suggesting that b-adrenergic stimulation ofdiscrepancy may be due to contaminations of the isolated acid secretion in the collecting duct is mediated throughCCD fragments used for RT-PCR with small vessels this receptor subtype. Besides the b1-AR subtype, a faintand/or capillaries rich in b2-AR. Discrepancies between amount of b2-AR in the medullary portions of the collect-mRNA and protein levels might also arise from differen- ing duct was detected, with an apparent higher expres-tial translation and/or protein turnover of b1- and b2-AR sion of this receptor subtype in the IMCD. Correspondingin the collecting duct. Finally, it should be noted that data were obtained by Yasuda et al, who demonstratedsimilar PCR studies carried out on CCD of hamster [50] expression of both functional b1-AR and, to a lesserand rat kidney [9] failed to detect significant levels of extent, b2-AR in the initial and terminal portions of ratb2-AR specific mRNA.
IMCD [67].Until recently, the only well-established catecholamine
actions in the collecting duct were derived from func- Pharmacological considerationstional studies on the rabbit. In these studies, b-adrenergic
One striking feature in this study is that an importantagonists were shown to decrease the transepithelialfraction of the b-AR expressed in the tubular cells ofpotential difference [59], to inhibit K1 secretion [60],the nephron and in the intercalated cells of the collectingand to stimulate Cl2/HCO2
3 exchange and Cl2/Cl2 self-duct system is located apically (in addition to the pre-exchange [61] in a cAMP-dependent manner. In addition,dicted basolateral location) and thus is likely to be ex-it was suggested that isoprenaline increases HCO2
3 excre-posed to the tubular luminal fluid. Implications of thistion into the tubular fluid through stimulation of b1-ARpeculiar orientation for the delivery of the endogenous[62]. All of these processes are thought to occur in typeligands have already been discussed. However, oneB intercalated cells, which are more abundant in rabbitshould also consider implications for the systemic admin-than in rat CCD [35, 63]. In the rat, little is knownistration of synthetic b-AR agonists and antagonists.about the b-adrenergic control of this particular nephron
segment. Here, we show a high expression of the b1-AR Such considerations apply particularly to b1-selective an-subtype in rat CCD. The receptors are predominantly tagonists of the first and second generations, such aslocalized on intercalated (rather than principal) CCD metoprolol and bisoprolol, which are standard agents incells, including both the band 3-positive type A and very the treatment of hypertension, coronary heart disease,probably also the band 3-negative type B intercalated and heart failure [68, 69]. Although a majority of thesecells. Thus, in conjunction with functional data derived water-soluble drugs are metabolized to inactive metabo-from the rabbit, our immunohistochemical results indi- lites, up to 50% of an oral dose of bisoprolol (and aboutcate that in rat CCD, the b-adrenergic system might be 3% in the case of metoprolol) is eliminated via the kid-involved in the regulation of both the acid-excreting type neys as an unchanged substance [24]. It is likely thatA and the base-excreting type B intercalated cells. such agents will exert pharmacological effects during
their passage through the nephron. For instance, theyb1-Adrenergic control of acid excretion in thecould block b1-AR at the macula densa, in distal tubularmedullary collecting ductepithelia, or in type A intercalated cells of the collecting
The OMCD has been shown to be an important siteduct and thus inhibit renin secretion, NaCl reabsorption,for urine acidification, whereas the IMCD is known toand also urine acidification. Such pharmacological effectsrepresent the site of final adjustment of the urine compo-have, to our knowledge, not been taken into consider-sition. The numbers of intercalated cells in these segmentsation when using these compounds in clinical practiceare less than in the CCD, but they are all acid-excretingbecause the expression of b-AR at the luminal surfacestype A cells (identified by H1-ATPase in the apical, andof the nephron has not been known. In light of a recentband 3 protein in the basolateral membrane) [33]. Inreport on the effects of metoprolol on renal NaCl han-the rabbit, there is increasing evidence that b-ARs aredling in an experimental rat model of chronic heart fail-critically involved in regulating OMCD function [64, 65]ure [70] and the data presented here, it is tempting toand particularly in regulating the secretion of H1 into thespeculate how the pharmacological potential of b-recep-tubular fluid. Recently, those studies were extended bytor antagonists in tubular luminal fluid could be exploitedManger, Pappas and Koeppen [66], who demonstrated
that in cultured rabbit type A intercalated cells isoprena- further in therapy.

Boivin et al: b-ARs in rat kidney530
by automated DNA sequence analysis: Further evidence for aACKNOWLEDGMENTSmultigene family. Proc Natl Acad Sci USA 84:8296–8300, 1987
16. Frielle T, Collins S, Daniel KW, et al: Cloning of the cDNAThe work was supported by the Deutsche Forschungsgemeinschaft(DFG Ja 706/2-1; Bonn, Germany). Valerie Boivin was recipient of a for the human b1-adrenergic receptor. Proc Natl Acad Sci USA
84:7920–7924, 1987grant from the Alexander-von-Humboldt Foundation (Bonn, Germany).A portion of this study was presented at the European Conference 17. Kobilka BK, Dixon RA, Frielle T, et al: cDNA for the human
b2-adrenergic receptor: A protein with multiple membrane-span-on Management of Coronary Heart Disease, Nice, France, April, 2000.ning domains and encoded by a gene whose chromosomal locationis shared with that of the receptor for platelet-derived growthReprint requests to Valerie Boivin, Ph.D., Institute of Pharmacology
and Toxicology, University of Wuerzburg, Versbacher Str. 9, D-97078 factor. Proc Natl Acad Sci USA 84:46–50, 198718. Reilander H, Boege F, Vasudevan S, et al: Purification and func-Wuerzburg, Germany.
E-mail: [email protected] tional characterization of the human b2-adrenergic receptor pro-duced in baculovirus-infected insect cells. FEBS Lett 282:441–444,1991
19. Jahns R, Boivin V, Siegmund C, et al: Autoantibodies activatingAPPENDIXhuman b1-adrenergic receptors are associated with reduced cardiacfunction in chronic heart failure. Circulation 99:649–654, 1999Abbreviations used in this article are: b-AR, b-adrenergic re-
ceptors; BSA, bovine serum albumin; CCD, cortical collecting duct; 20. Drenckhahn D, Schluter K, Allen DP, et al: Colocalizationof band 3 with ankyrin and spectrin at the basal membrane ofCNT, connecting tubule; DCT, distal convoluted tubule; ECL, en-
hanced chemiluminescence; eNOS, endothelial nitric oxide synthase; intercalated cells in the rat kidney. Science 230:1287–1289, 198521. Munson PJ, Rodbard D: LIGAND: A versatile computerized[125I]-CYP, [125I]-iodocyanopindolol; IMCD, inner medullary collecting
duct; OMCD, outer medullary collecting duct; OSOM, outer stripe of approach to characterization of ligand binding systems. Anal Bio-chem 107:220–239, 1980the outer medulla; PBS, phosphate-buffered saline; PCT, proximal
convoluted tubule; PTE, proximal tubular epithelial cells; RT-PCR, 22. Kriz W, Kaissling B: Structural organisation of the mammaliankidney, in The Kidney: Physiology and Pathophysiology, editedreverse transcription-polymerase chain reaction; TAL, thick ascending
limb of Henle’s loop. by Seldin DW, Giebisch G, New York, Raven Press, 1992, pp707–777
23. Snavely MD, Motulsky HJ, Moustafa E, et al: b-AdrenergicREFERENCES receptor subtypes in the rat renal cortex. Circ Res 51:504–513, 1982
24. Leopold G, Kutz K: Bisoprolol: Pharmacokinetic profile. Rev1. DiBona GF, Kopp UC: Neural control of renal function. PhysiolContemp Pharmacother 8:35–43, 1997Rev 77:75–197, 1997
25. Brodde O-E: The pharmacology of bisoprolol. Rev Contemp Phar-2. Engel G, Maurer R, Perrot K, et al: b-Adrenoceptor subtypesmacother 8:21–23, 1997in sections of rat and guinea-pig kidney. Naunyn Schmiedebergs
26. Michel MC, Wang XL, Schlicker E, et al: Increased b2-adreno-Arch Pharmacol 328:354–357, 1985ceptor density in heart, kidney, and lung of spontaneously hyper-3. Pelayo J, Tucker B, Blantz R: Effects of b-adrenergic stimulationtensive rats. J Auton Pharmacol 7:41–51, 1987with isoproterenol on glomerular hemodynamics. Am J Physiol
27. Boege F, Jurss R, Cooney D, et al: Functional and structural257:F866–F873, 1989characterization of the two b1-adrenoceptor forms in turkey eryth-4. Torretti J: Sympathetic control of renin release. Ann Rev Pharma-rocytes with molecular masses of 50 and 40 kilodaltons. Biochemis-col Toxicol 22:167–192, 1982try 26:2418–2425, 19875. Malbon CC, Moxham CP, Brandwein HJ: Antibodies to b-adren-
28. Singh H, Linas S: b2-Adrenergic function in cultured rat proximalergic receptors, in The b-Adrenergic Receptors, edited by Perkinstubule epithelial cells. Am J Physiol 271(1 Pt 2):F71–F77, 1996JP, Baltimore, Humana Press, 1991, pp 181–261
29. Singh H, Linas S: Role of protein kinase C in b2-adrenoceptor6. Healy D, Munzel P, Insel P: Localization of b1- and b2-adrenergicfunction in cultured rat proximal tubule epithelial cells. Am Jreceptors in rat kidney by autoradiography. Circ Res 57:278–284,Physiol 273:F193–F199, 19971985
30. Hoyer JR, Sisson SP, Vernier RL: Tamm-Horsfall glycoprotein:7. Summers R, Stephenson J, Kuhar M: Localization of b-adrenocep-Ultrastructural immunoperoxidase localization in the rat kidney.tor subtypes in rat kidney by light microscopic autoradiography.Lab Invest 41:168–173, 1979J Pharmacol Exp Ther 232:561–569, 1985
31. Taylor AN, McIntosh JE, Bourdeau JE: Immunocytochemical8. Amenta F, Cavallotti C, De Rossi M, et al: b-Adrenoceptors inlocalization of vitamin D-dependent calcium-binding protein inthe rat kidney. Immunohistochemical study. Naunyn Schmiede-renal tubules of rabbit, rat, and chick. Kidney Int 21:765–773, 1982bergs Arch Pharmacol 324:94–98, 1983
32. Kaissling B, Kriz W: Morphology of the loop of Henle, distal9. Meister B, Dagerlind A, Nicholas A, et al: Patterns of messengertubule, and collecting duct, in Handbook of Physiology: RenalRNA expression for adrenergic receptor subtypes in the rat kidney.Physiology, edited by Windhager EE, New York, Oxford Univer-J Pharmacol Exp Ther 268:1605–1611, 1994sity Press, 1992, pp 109–16710. Elalouf J, Buhler J, Tessiot C, et al: Predominant expression
33. Clapp WL, Madsen KM, Verlander JW, et al: Intercalated cellsof b1-adrenergic receptor in the thick ascending limb of rat kidney:of the rat inner medullary collecting duct. Kidney Int 31:1080–1087,Absolute mRNA quantitation by reverse transcription and poly-1987merase chain reaction. J Clin Invest 91:264–272, 1993
34. Alper SL, Natale J, Gluck S, et al: Subtypes of intercalated cells11. Mandon B, Siga E, Champigneulle A, et al: Molecular analysisin the rat kidney collecting duct defined by antibodies againstof b-adrenergic receptor subtypes in rat collecting duct: Effectserythroid band 3 and renal vacuolar H1-ATPase. Proc Natl Acadon cell cAMP and Ca21 levels. Am J Physiol 268:F1070–F1080,Sci USA 86:5429–5433, 19891995
35. Verlander JW, Madsen KM, Tisher CC: Effect of acute respira-12. Gesek F, White K: Molecular and functional identification oftory acidosis on two populations of intercalated cells in rat corticalb-adrenergic receptors in distal convoluted tubule cells. Am J Phys-collecting duct. Am J Physiol 253:F1142–F1456, 1987iol 272:F712–F720, 1997
36. Osborn JL, Thames MD, DiBona GF: Role of macula densa in13. Jahns R, Siegmund C, Jahns V, et al: Probing human b1- andrenal nerve modulation of renin secretion. Am J Physiol 242:R367–b2-adrenoceptors with domain-specific fusion protein antibodies.R371, 1982Eur J Pharmacol 316:111–121, 1996
37. Skott O, Briggs JP: Direct demonstration of macula densa-medi-14. Machida CA, Bunzow JR, Searles RP, et al: Molecular cloningated renin secretion. Science 237:1618–1620, 1987and expression of the rat b1-adrenergic receptor gene. J Biol Chem
38. Schnermann J, Briggs JP: The macula densa is worth its salt.265:12960–12965, 1990J Clin Invest 104:1007–1009, 199915. Gocayne J, Robinson DA, Fitz GM, et al: Primary structure of rat
cardiac b-adrenergic and muscarinic cholinergic receptors obtained 39. Wright FS, Schnermann J: Interference with feedback control of

Boivin et al: b-ARs in rat kidney 531
glomerular filtration rate by furosemide, triflocin and cyanide. mal tubule epithelial cells in culture. Am J Physiol 268:F553–F560,1995J Clin Invest 53:1695–1708, 1974
56. Barajas L, Powers K: Innervation of the renal proximal convo-40. Bailly C, Imbert-Teboul M, Roinel N, et al: Isoproterenol in-luted tubule of the rat. Am J Anat 186:378–388, 1989creases Ca, Mg, and NaCl reabsorption in mouse thick ascending
57. Baines AD, Craan A, Chan W, et al: Tubular secretion andlimb. Am J Physiol 258:F1224–F1231, 1990metabolism of dopamine, norepinephrine, methoxytyramine and41. Barajas L, Powers K: Innervation of the thick ascending limb ofnormetanephrine by the rat kidney. J Phamacol Exp Ther 208:144–Henle. Am J Physiol 255:F340–F348, 1988147, 197942. Schor N, Ichikawa I, Brenner BM: Mechanisms of action of
58. Barajas L, Powers K, Wang P: Innervation of the renal corticalvarious hormones and vasoactive substances on glomerular ultra-tubules: A quantitative study. Am J Physiol 247:F50–F60, 1984filtration in the rat. Kidney Int 20:442–451, 1981 59. Iino Y, Troy JL, Brenner BM: Effects of catecholamines on elec-43. Dzau VJ, Kreisberg J: Cultured glomerular mesangial cells con- trolyte transport in cortical collecting tubules. J Membr Biol 61:67–
tain renin: Influence of calcium and isoproterenol. J Cardiovasc 73, 1981Pharmacol 8(Suppl 10):S6–S10, 1986 60. Kimmel PK, Goldfard S: Effects of isoproterenol on potassium
44. Kreisberg JI, Venkatachalam MA, Radnik RA, et al: Role of secretion by the cortical collecting tubule. Am J Physiol 246:F804–myosin light-chain phosphorylation and microtubules in stress fiber F810, 1984morphology in cultured mesangial cells. Am J Physiol 249:F227– 61. Tago K, Schuster VL, Stokes JB: Regulation of chloride selfF235, 1985 exchange by cAMP in cortical collecting tubule. Am J Physiol
251:F40–F48, 198645. Sundaresan P, Fortin T, Kelvie S: a- and b-adrenergic receptors62. Hayashi M, Yamaji Y, Iyori M, et al: Effect of isoproterenolin proximal tubules of rat kidney. Am J Physiol 253:F848–F856,
on intracellular pH of the intercalated cells in the rabbit cortical1987collecting ducts. J Clin Invest 87:1153–1157, 199146. Clark JD, Cragoe EJ, Limbird LE: a2-adrenergic receptors regu-
63. Schuster VL, Bonsib SM, Jennings ML: Two types of collectinglate Na1-H1 exchange via cAMP-dependent mechanism. Am Jduct mitochondria-rich (intercalated) cells: Lectin and band 3 cyto-Physiol 259:F977–F985, 1990chemistry. Am J Physiol 251:C347–C355, 198647. Feng F, Pettinger WA, Abel PW, et al: Regional distribution of
64. Burnatowska-Hledin MA, Spielman WS: Immunodissection ofa1-adrenoceptor subtypes in rat kidney. J Pharmacol Exp Ther mitochondria-rich cells from rabbit outer medullary collecting tu-258:263–268, 1994 bule. Am J Physiol 254:F907–F911, 1988
48. Jacobs W, Chan Y: Evidence for the presence of functional 65. Hays S, Kokko JP, Jacobson R: Hormonal regulation of protonb-adrenoceptor along the proximal tubule of the rat kidney. Bio- secretion in rabbit medullary collecting duct. J Clin Invest 78:1279–chem Biophys Res Commun 141:334–339, 1986 1286, 1986
49. Kudo K, Kondo Y, Abe K, et al: Evidence for presence of func- 66. Manger T, Pappas C, Koeppen B: b-Adrenergic regulation of H1
tional b-adrenoceptor in rabbit S2 proximal straight tubules. Am secretion by cultured outer medullary collecting duct cells. Am JJ Physiol 261:F393–F399, 1991 Physiol 263:F1011–F1019, 1992
67. Yasuda G, Sun L, Lee H, et al: The b1- and b2-adrenoceptor50. Taniguchi S, Watanabe T, Nakao A, et al: Distribution ofsubtypes in cultured rat inner medullary collecting duct cells. Amb2-adrenergic receptor mRNA expression along the hamster neph-J Physiol 271:F762–F769, 1996ron segments. FEBS Lett 318:65–70, 1993
68. MERIT-HF Study Group: Effect of metroprolol CR/XL in chronic51. Bello-Reuss E: Effect of catecholamines on fluid reabsorptionheart failure: Metroprolol CR/XL randomised intervention trialby the isolated proximal convoluted tubule. Am J Physiol 238:in congestive heart failure (MERIT-HF). Lancet 353:2001–2007,F347–F352, 1980199952. Chan YL: The role of norepinephrine in the regulation of fluid
69. CIBIS-II Investigators and Committees: The cardiac insufficiencyabsorption in the rat proximal tubule. J Pharmacol Exp Ther bisoprolol study II (CIBIS-II): A randomised trial. Lancet 353:9–13,215:65–70, 1980 1999
53. Weinman EJ, Sansom SC, Knight TF, et al: a and b-adrenergic 70. DiBona GF, Sawin LL: Effect of metoprolol administration onagonists stimulate water absorption in the rat proximal tubule. renal sodium handling in experimental congestive heart failure.J Membr Biol 69:107–111, 1982 Circulation 100:82–86, 1999
54. Gesek FA, Schoolwerth AC: Hormonal interaction with the prox- 71. Kriz W, Elger M, Lemley K, et al: The structure of the glomerularimal Na1-H1 exchanger. Am J Physiol 258:F514–F521, 1990 mesangium: A biomechanical interpretation. Kidney Int 38(Suppl
30):S2–S9, 199055. Hanson A, Linas S: b-Adrenergic receptor function in rat proxi-





























