Embed Size (px)
Citation preview

Immunocyto- and histochemical profiling ofnucleostemin expression: Marker of epidermalstem cells?
Lukas Lacina a,b, Karel Smetana Jr.a,c,*, Barbora Dvorankova a,c,Jirı Stork b, Zuzana Plzakova a,c, Hans-Joachim Gabius d
Journal of Dermatological Science (2006) 44, 73—80
www.intl.elsevierhealth.com/journals/jods
aCharles University, 1st Faculty of Medicine, Institute of Anatomy, Prague, Czech RepublicbCharles University, 1st Faculty of Medicine, Department of Dermatovenerology, Prague, Czech RepubliccCharles University, 2nd Faculty of Medicine, Center of Cell Therapy and Tissue Repair,Prague, Czech Republicd Ludwig-Maximilians-University, Faculty of Veterinary Medicine, Institute of Physiological Chemistry,Munich, Germany
Received 3 February 2006; received in revised form 11 August 2006; accepted 16 August 2006
KEYWORDSEpidermis;Hair follicle;Keratin;Nucleolus;Stem cells
Summary
Background and objective: Because the nucleolar protein nucleostemin is present inbone marrow and neuronal stem cells and malignancies originating thereof wemonitored its expression in frozen sections from normal human epidermis, basal cellcarcinomas, cultured keratinocytes and cells of the squamous carcinoma line FaDu. Inaddition, probing the value of this protein as a marker of epidermal stem cells was anaim of this study.Materials and methods: To further characterize cell features we added analysis ofexpression of keratins 10 or 19 as markers of terminal differentiation and Ki67 asmarker of proliferating cells as well as three adhesion/growth-regulatory galectins.Results: Immunohistochemical monitoring revealed expression of nucleostemin incells of both Ki67-positive and -negative nuclei regardless of the K10-expressionstatus. Cultured keratinocytes were positive, when they were prepared from hairfollicles and cultured in the presence of feeder cells. A small population of thesenucleostemin-positive cells also expressed galectin-1 but not galectins-3 and -9 intheir nucleoli. Part of these cells also expressed keratin 19. FaDu cells were stronglypositive, illustrating expression in malignant cells which require no feeder layer. Ofnote, the number of galectin-1-positive nucleoli was reduced in the course of culture.
* Corresponding author at: Charles University, 1st Faculty of Medicine, Institute of Anatomy, U Nemocnice 3, 128 00 Prague 2, CzechRepublic. Tel.: +420 2 24965873; fax: +420 2 24965770.
E-mail address: [email protected] (K. Smetana Jr.).
0923-1811/$30.00 # 2006 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.jdermsci.2006.08.008

1. Introduction
Tissue (adult) stem cells (SC) are involved in self-renewal of tissues and their regeneration afterinjury. Regarding therapeutic applications, theytherefore represent a potential source of cellmaterial for tissue engineering and regenerativemedicine, which when dysfunctional may also con-tribute to the origin of certain malignancies [1].Consequently, the study of the SC phenotype andcourses of maturation has attained a prominentstatus, especially the search for still elusive mar-kers. Interestingly, phenotypic fingerprints com-bining analysis of selected features for distinctpopulations of tissue SC including epidermal SCenabled a certain degree of characterization [2].It is within this quest for marker definition that wescreened the expression of nucleostemin (NuclS).This protein shuttles between nucleoplasm andnucleolus, binds GTP and interacts with p53[3,4]. It probably has regulatory functions, becauseit is not involved in the nucleolar synthesis of rRNAin the nucleolus [5], and its expression appears tocontrol proliferation and onset of apoptosis, lack ofexpression of nucleostemin being connected withlack of proliferation activity and overexpressionwith induction of apoptosis [3]. Expression of NuclSpresence was observed in bone marrow stromal,nervous and hematopoietic SC and malignanciesthereof [3,6]. Expression of nucleostemin in threetypes of SC with independent origin intimates thatthis protein may be an attribute of adult stem cellsas phenotypic marker. Of relevance in this respect,nucleostemin expression was reported for cancercells of epithelial origin such as cells of gastriccancer and HeLa cells [7,8]. At present, no dataabout its expression in normal epidermis andrelated stem cells are available, warranting to fillthis gap. Thus, we have addressed this issue in ourreport.
This study reveals expression of NuclS in theepidermis, in basal cell carcinoma (BCC) as wellas cultured epidermal and squamous cell carcinoma(SCC) cells. Expression of NuclS is correlated toexpression of markers of differentiation such askeratins (K) 10 and 19 and the marker of prolifera-tion Ki67. In addition we monitored expression of
three members of a family of adhesion/growth-regulatory endogenous lectins, i.e. the galectins[9—12]. Besides key roles at the cell surface, e.g.signaling G1 arrest via binding to glycans of thefibronectin receptor or growth inhibition by inter-action with ganglioside GM1 [13—16], the nuclearpresence of galectins-1 and -3 plays a role in splicingof pre-mRNA and is subject to regulation with thedifferentiation level [17,18]. These activities war-rant to study expression of relevant galectins fromeach of the three subfamilies, i.e. proto-type(galectin-1), chimera-type (galectin-3) and tan-dem-repeat-type (galectin-9), in cultured cells inrelation to NuclS presence.
2. Materials and methods
2.1. Sample collection and processing
Samples of normal human skin from five donors andBCC from seven donors were obtained with explicitconsent of the donors. The specimens were imme-diately cryoprotected by Tissue-Tek (Sakura, Zoe-terwoude, Netherlands) and frozen in liquidnitrogen for immunohistochemical analysis. Sec-tions at 7 mm thickness were obtained using aCryo-Cut-E microtome (Reichert Jung, Vienna, Aus-tria). These samples were used for in situ analysis.Other two samples of human skin obtained from theDepartment of Aesthetic Surgery of the third Facultyof Medicine of the Charles University with explicitconsent of donors were the source of cells for cellcultivation in vitro experiments.
2.2. Cell culture
Both types of keratinocytes, i.e. from hair folliclesand interfollicular epidermis, were prepared forstudy. Briefly, the tissue specimens were cut tosmall pieces and treated overnight with 0.3% solu-tion of trypsin (Sigma—Aldrich, Prague, CzechRepublic) at +4 8C. The epidermis was peeled offthereafter; a suspension of keratinocytes frominterfollicular epidermis was generated by carefulmechanical disintegration [19]. These cells werecultured in the absence or with mitomycin c
74 L. Lacina et al.
Conclusion: Nucleostemin positivity cannot be considered as marker for stem cells inskin sections. In cultured cells, nucleostemin is expressed in a distinct population ofthe epidermal cells from hair follicle kept in the presence of a feeder layer, intimatingan association of nucleostemin expression with this type of epithelio-mesenchymalinteraction which is not essential during propagation of malignant cells.# 2006 Japanese Society for Investigative Dermatology. Published by Elsevier IrelandLtd. All rights reserved.

Nucleostemin in epidermis 75
(Sigma—Aldrich, Prague, Czech Republic) treated3T3 feeder cells. The obtained pieces of dermiscontaining hair follicles [19] were rinsed and keptin a Petri dish in HMEM medium (Academy ofScience, Prague, Czech Republic) containing 10%of bovine serum (ZVOS, Hustopece, Czech Repub-lic). Within a few days, keratinocytes migratedfrom hair follicles and colonized the dish. Cellsof the second passage were processed. Keratino-cytes were grown on cover glass slips with or with-out the feeder layer. Feeder cells were seeded at adensity of 30,000 cells/cm2, keratinocytes at adensity of 20,000 cells/cm2. HMEM medium with10% bovine serum, 2% of fetal bovine serum (Bio-chrom, Berlin, Germany) supplemented with epi-dermal growth factor, hydrocortisone (Sigma—Aldrich, Prague, Czech Republic) and insulin wasused for culture at 37 8C and in an atmosphere with3.3% of CO2 for 3 and 5 days. Characteristics ofcultured keratinocytes were compared with thoseof cells of the FaDu line which originated from aSCC case. FaDu cells were seeded at a density of5000 cells/cm2 and cultured without feeder layer.E-MEM medium (Biochrom, Berlin, Germany) with10% of fetal bovine serum was used for culture at37 8C and 5% of CO2 for 2 or 7 days. Glass cover slipswith cells were rinsed with PBS, dried and storedfrozen at �20 8C.
3. Immuno- and galectinhistochemistry
Tissue sections and cultured cells grown on coverslipswere fixed with 2% paraformaldehyde in PBS (pH 7.2)for 5 min.NuclS expressionwasdetectedusing a goatpolyclonal antibody (Neuromics, Bloomington, MN,USA) at a concentration of 3 mg/ml of incubationmedium. K10, K19 and the proliferation markerKi67 were detected with respective mouse monoclo-nal antibodies (DAKO, Brno, Czech Republic). Galec-tins-1, -3 and -9 were detected with non-cross-reactive rabbit polyclonal antibody fractions [20—22]. Donkey anti-goat antibody labeled with thefluorochrom TRITC (Jackson Laboratories, WestGrove, PA, USA) and porcine anti-rabbit or anti-mouse antibodies labeled with FITC (AlSeVa, Prague,Czech Republic) were used as second-step reagents.Specificity controls includedomissionof thefirst-stepreagent from the protocol, assaying of other anti-bodies not reactive in this tissue to excludebinding toFc receptors and saturation of non-specific protein-binding sites to exclude antigen-independent bind-ing, as described previously together with details ofthe staining protocol [23]. Cell nuclei were counter-stained with DAPI (Sigma—Aldrich, Prague, Czech
Republic), and the specimens were finally mountedto Vectashield (Vector Laboratories, Burlingame, CA,USA). Analysis of staining profiles was performedusing a Nikon Eclipse 90i fluorescence microscopeequipped with filterblocks specific for the spectro-scopic properties of DAPI, FITC and TRITC. A cooledhigh-resolution CCD camera Vosskhuler and a com-puter-assisted image analysis system LUCIA 5.1, bothfrom Laboratory Imaging (Prague, Czech Republic),were employed for analysis and filing of data, whichwere then subjected to quantitative analysis [24].The data for statistical analysis were acquired bymeasuring of fluorescence intensity of 300 cells persection. Three sections for each sample were ana-lyzed in all frozen samples of normal human skin orbasal cell carcinomas.
4. Results
4.1. Normal skin and basal cell carcinoma
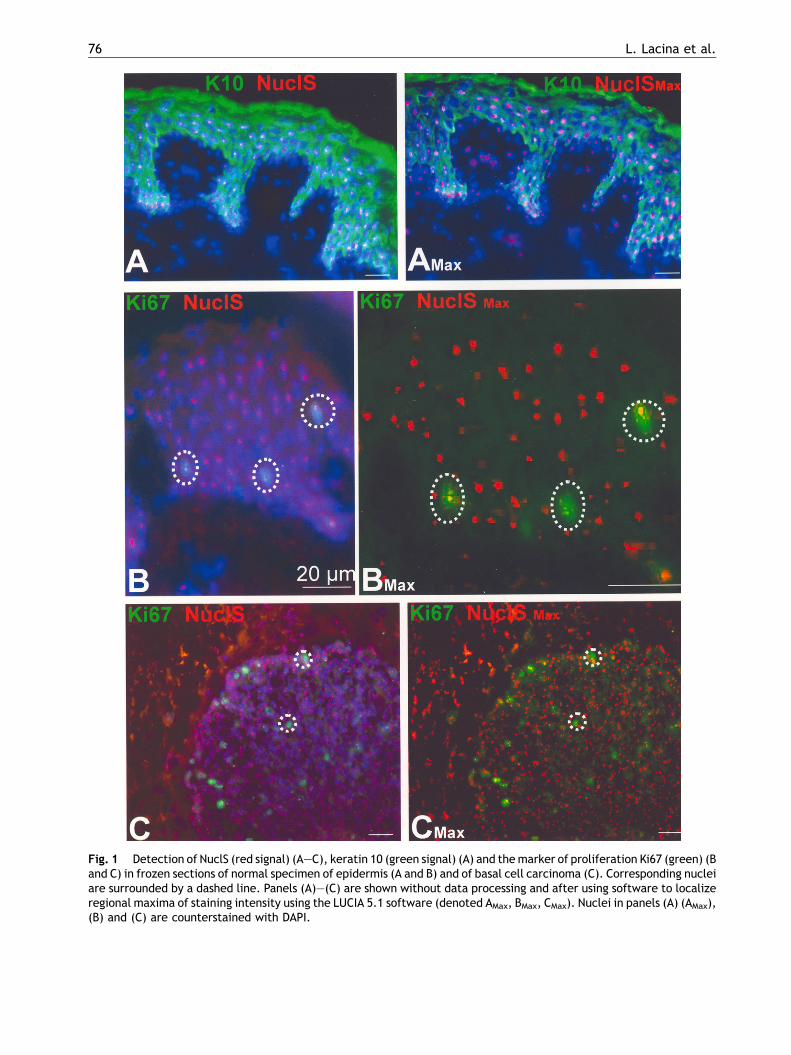
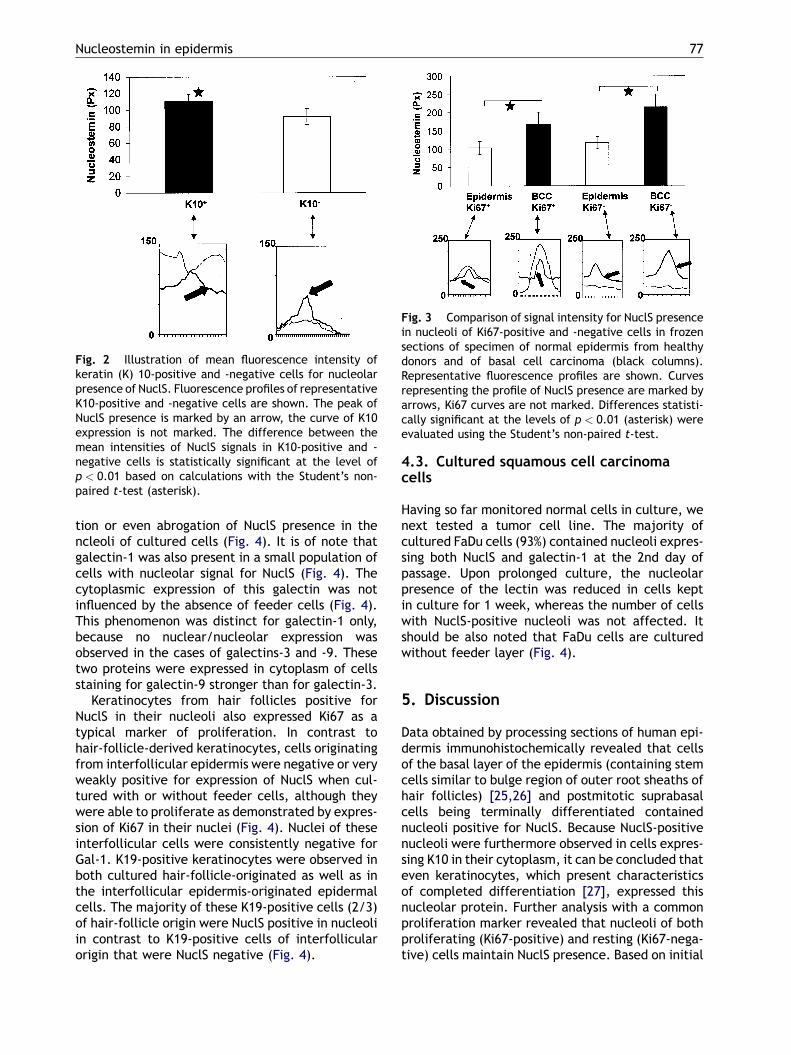
Nuclei of cells of the basal layer assumed to consti-tute the proliferatively active compartment of theepidermis and nuclei of suprabasal cells of the spi-nous layer, where the terminally differentiated cellsreside, contained nucleoli positive for NuclS (Fig. 1)in both interfollicular epidermis and hair follicles. Tofurther characterize the functional status of cellswith nucleostemin-positive nucleoli, we examinedthe expression of K10 as a marker of terminal differ-entiation. NuclS-positive nuclei were found in bothK10-positive and -negative cells (Fig. 1). In quanti-tative terms, the signal of NuclS expression wassignificantly lower in K10-negative than in K10-posi-tive cells (Fig. 2). The interfollicular regions contain-ing keratinocytes with a positive signal fornucleostemin are known to be negative for K19, aswas verified by studying this type of intermediatefilaments in parallel sections.Whenwecompared theexpression of NuclS and Ki67, used as a marker ofproliferating cells, both cells with and also withoutKi67 expression exhibited a signal for NuclS in theirnucleoli (Fig. 1). Comparing signal intensity for NuclSin normal epithelium and cells of BCC the expressionlevel of NuclS was significantly higher in tumor cellsthan in normal epithelium (Figs. 1 and 3). Havinghereby monitored NuclS presence in frozen sections,we will next look at this feature in cells in vitro.
4.2. Cultured epidermal cells
Cultured cells prepared from hair follicles werepositive for NuclS in nucleoli when these cells werecultured with 3T3 feeder cells (Fig. 4). Absence offeeder cells from the culture led to a strong reduc-
76 L. Lacina et al.
Fig. 1 Detection of NuclS (red signal) (A—C), keratin 10 (green signal) (A) and themarker of proliferation Ki67 (green) (Band C) in frozen sections of normal specimen of epidermis (A and B) and of basal cell carcinoma (C). Corresponding nucleiare surrounded by a dashed line. Panels (A)—(C) are shown without data processing and after using software to localizeregional maxima of staining intensity using the LUCIA 5.1 software (denoted AMax, BMax, CMax). Nuclei in panels (A) (AMax),(B) and (C) are counterstained with DAPI.
Nucleostemin in epidermis 77
Fig. 2 Illustration of mean fluorescence intensity ofkeratin (K) 10-positive and -negative cells for nucleolarpresence of NuclS. Fluorescence profiles of representativeK10-positive and -negative cells are shown. The peak ofNuclS presence is marked by an arrow, the curve of K10expression is not marked. The difference between themean intensities of NuclS signals in K10-positive and -negative cells is statistically significant at the level ofp < 0.01 based on calculations with the Student’s non-paired t-test (asterisk).
Fig. 3 Comparison of signal intensity for NuclS presencein nucleoli of Ki67-positive and -negative cells in frozensections of specimen of normal epidermis from healthydonors and of basal cell carcinoma (black columns).Representative fluorescence profiles are shown. Curvesrepresenting the profile of NuclS presence are marked byarrows, Ki67 curves are not marked. Differences statisti-cally significant at the levels of p < 0.01 (asterisk) wereevaluated using the Student’s non-paired t-test.
tion or even abrogation of NuclS presence in thencleoli of cultured cells (Fig. 4). It is of note thatgalectin-1 was also present in a small population ofcells with nucleolar signal for NuclS (Fig. 4). Thecytoplasmic expression of this galectin was notinfluenced by the absence of feeder cells (Fig. 4).This phenomenon was distinct for galectin-1 only,because no nuclear/nucleolar expression wasobserved in the cases of galectins-3 and -9. Thesetwo proteins were expressed in cytoplasm of cellsstaining for galectin-9 stronger than for galectin-3.
Keratinocytes from hair follicles positive forNuclS in their nucleoli also expressed Ki67 as atypical marker of proliferation. In contrast tohair-follicle-derived keratinocytes, cells originatingfrom interfollicular epidermis were negative or veryweakly positive for expression of NuclS when cul-tured with or without feeder cells, although theywere able to proliferate as demonstrated by expres-sion of Ki67 in their nuclei (Fig. 4). Nuclei of theseinterfollicular cells were consistently negative forGal-1. K19-positive keratinocytes were observed inboth cultured hair-follicle-originated as well as inthe interfollicular epidermis-originated epidermalcells. The majority of these K19-positive cells (2/3)of hair-follicle origin were NuclS positive in nucleoliin contrast to K19-positive cells of interfollicularorigin that were NuclS negative (Fig. 4).
4.3. Cultured squamous cell carcinomacells
Having so far monitored normal cells in culture, wenext tested a tumor cell line. The majority ofcultured FaDu cells (93%) contained nucleoli expres-sing both NuclS and galectin-1 at the 2nd day ofpassage. Upon prolonged culture, the nucleolarpresence of the lectin was reduced in cells keptin culture for 1 week, whereas the number of cellswith NuclS-positive nucleoli was not affected. Itshould be also noted that FaDu cells are culturedwithout feeder layer (Fig. 4).
5. Discussion
Data obtained by processing sections of human epi-dermis immunohistochemically revealed that cellsof the basal layer of the epidermis (containing stemcells similar to bulge region of outer root sheaths ofhair follicles) [25,26] and postmitotic suprabasalcells being terminally differentiated containednucleoli positive for NuclS. Because NuclS-positivenucleoli were furthermore observed in cells expres-sing K10 in their cytoplasm, it can be concluded thateven keratinocytes, which present characteristicsof completed differentiation [27], expressed thisnucleolar protein. Further analysis with a commonproliferation marker revealed that nucleoli of bothproliferating (Ki67-positive) and resting (Ki67-nega-tive) cells maintain NuclS presence. Based on initial

78 L. Lacina et al.
Fig. 4 Detection of NuclS (red signal) (A—F), galectin-1 (green signal) (A, B, F), Ki67 (green signal) (C) and keratin 19(green signal) (D and E) in cells originated in hair follicles (HF) cultured in the presence of feeder cells (A and D) or withoutfeeder cells (B), in interfollicular cells (IF) (C) and cells of the squamous cell carcinoma line FaDu (F) cultured for 3, 5 and7 days, respectively. Detection of galectin-1 in cells from panel (A) was refined by data processing for visualization of

Nucleostemin in epidermis 79
observations of Tsai and McKay [3], malignanciesoriginating from neural and hematopoietic stemcells also contain nucleoli positive for NuclS. Ourdata extend the expression profile to cells of basalcell carcinomas, in situ and in vitro.
The results of our study part on cultured cellsdemonstrated that nucleoli positive for NuclS werefound in cells prepared from hair follicles when theywere co-cultured with feeder cells. Exclusion of fee-dercells substantially reducedoreven impairedNuclSexpression in nucleoli. Of potential relevance in thisrespect is to note that epithelio-mesenchymal inter-actions are important for the development of theepidermis including its functional integrity extendingto stem cell function [28,29]. Looking at the well-studied case of keratin expression, palm/plant spe-cific keratin type 9 is expressed in cultured keratino-cytes only when they are cultured in the presence offeeder-cell fibroblasts originating from this body site[29]. Expression of nucleostemin in hair-follicle ker-atinocytes cultured in presence of feeder cellsextends the panel of proteins regulatable by contactto/vicinity ofmesenchymal cells in ontogeny andalsopostnatally. In our study, cells prepared from theinterfollicular epidermis showed a significantlyreduced expression level or even absence of NuclS.No direct relationship between signals for K19 repre-senting a marker of epidermal stem cells and NuclSwas found discernable. Previously, we had observeddifferences in other phenotypic markers and func-tional aspects of behavior of the cells prepared fromhair follicle and interfollicular epidermis, particu-larly the expression of distinct keratins and galectinsincluding galectin-reactive sites [19,30,31]. Namely,galectin-1 and epitopes reactivewith the lectinwereselectively expressed in nuclei/nucleoli of cells pre-pared from hair follicles phenotypically related toepidermal SC [19,30,31]. In addition to cell surfaceglycans this lectin is known to have affinity, too, todistinct peptide motifs, making it capable to beengaged in protein—protein recognition in the cell[17,32—34]. Extending the previously reportedresults, nucleolar expression was observed for galec-tin-1 in a small population of hair-follicle-derivedcells lightmicroscopically colocalizing with NuclS.When comparing the obtained results on normal ker-atinocytes with those of the FaDu cells growing with-out feeder layer, a strong nucleolar signal for NuclSwasobserved. This propertywas kept constant duringculture,whereaspositivity forgalectin-1wasstronglyreduced.
Nucleostemin expression is an indicator of con-siderable proliferative capacity of cells (including
regional maxima (AMax). Fluorecsence profiles of FaDu cell nuc(I1—I6). Nuclei in panels (A), (B) and (D) are counterstained
malignant cells) [8,35]. This feature, i.e. almostnon-limited proliferative capacity, represents afundamental property of stem cells. Because weobserved NuclS positivity in cells prepared fromhair-follicle-containing cells with stem-cell pheno-type [19], the pool of NuclS-positive cells mayprobably include epidermal stem cells. On theother hand, NuclS expression is not directly con-nected to proliferation, because interfollicularproliferating (Ki67-positive) keratinocytes areNuclS-negative. The observation that malignantFaDu cells express this protein should be viewedunder the aspect that these cells harbor phenotypicmarkers of epidermal SC [18]. To relate strongexpression of NuclS in post-proliferative K10-posi-tive and K19-negative suprabasal cells with func-tionality it may be relevant to note thatoverexpression of this nucleolar protein has proa-poptotic activity [3]. Because NuclS is widelyexpressed in cells of normal and malignant renaltissue [36], it can be assumed that its functionalspectrum includes more tasks than simply regula-tion of proliferation in stem cells. In conclusion,despite positivity of epidermal stem cells in vitro,this protein cannot be considered as reliable mar-ker of epidermal stem cells, because it is expressedby suprabasal, terminally differentiated keratino-cytes harboring K10 in situ.
Acknowledgments
The authors are grateful to Dr. J. Boucek for gift ofcells of the commercial FaDu line and to Eva Vancovafor excellent technical assistance. This study wassupportedby theGrantAgencyof theCzechRepublic,project no. 304/04/0171, the Ministry of Education,Youth and Sport of the Czech Republic, projects no.MSM0021620806 and no. 1M0021620803 and an ECMarie Curie Research Training Network grant (con-tract no. MRTN-CT-2005-019561).
References
[1] Sell S. Stem cell origin of cancer and differentiation therapy.Crit Rev Oncol Hematol 2004;51:1—28.
[2] Tumbar T, Guasch G, Greco V. Defining the epithelial stemcell niche in skin. Science 2004;303:359—63.
[3] Tsai RYL, McKay RDG. A nucleolar mechanism controlling cellproliferation in stem cells and cancer cells. Gene Develop2002;16:2991—3003.
[4] Tsai RYL, McKay RDG. A multistep GTP-driven mechanismcontrolling the dynamic cycling of nucleostemin. J Cell Biol2005;168:179—84.
leoli (D) were measured and expressed as intensity curveswith DAPI.

80 L. Lacina et al.
[5] Politz JC, Polena I, Trask I, Bazett-Jones DP, Pedwerson T. Anonribosomal landscape in the nucleolus revealedby the stemcell protein nucleostemin. Mol Biol Cell 2005;16:3401—10.
[6] KafienahW, Mistry S, Williams C, Hollander AP. Nucleosteminis amarker of proliferating stromal stem cells in adult humanbone marrow. Stem Cells 2006;24:1113—20.
[7] Sijin L, Ziwei C, Yajun L, Meiyu D, Hongwei Z, Guofa H, et al.The effect of knocking-down nucleostemin gene expressionon the in vitro proliferation and in vivo tumorigenesis ofHeLa cells. J Exp Clin Cancer Res 2004;23:529—38.
[8] Liu S-J, Cai Z-W, Liu Y-J, Dong M-Y, Sun L-Q, Hu G-F, et al.Role of nucleostemin in growth regulation of gastric cancer,liver cancer and other malignancies. World J Gastroenterol2004;10:1246—9.
[9] Gabius H-J. Animal lectins. Eur J Biochem 1997;243:543—76.[10] Kaltner H, Stierstorfer B. Animal lectins as cell adhesion
molecules. Acta Anat 1998;161:162—79.[11] Gabius H-J, Siebert H-C, Andre S, Jimenez-Barbero J, Rudi-
ger H. Chemical biology of the sugar code. Chem Bio Chem2004;5:740—64.
[12] Lahm H, Andre S, Hoeflich A, Kaltner H, Siebert H-C, SordatB, et al. Tumor galectinology: insights into the complexnetwork of a family of endogenous lectins. Glycoconj J2004;20:227—38.
[13] Kopitz J, von Reitzenstein C, Andre S, Kaltner H, Uhl J,Ehemann V, et al. Negative regulation of neuroblastoma cellgrowth by carbohydrate-dependent surface binding ofgalectin-1 and functional divergence from galectin-3. J BiolChem 2001;276:35917—23.
[14] Andre S, Kaltner H, Lensch M, Russwurm R, Siebert H-C,Fallsehr HC, et al. Determination of structural and func-tional overlap/divergence of five proto-type galectins byanalysis of the growth-regulatory interaction with ganglio-side GM1 in silico and in vitro on human neuroblastoma cells.Int J Cancer 2005;114:46—57.
[15] Fischer C, Sanchez-Ruderisch H, Welzel M, Wiedenmann B,Sakai T, Andre S, et al. Galectin-1 interacts with the a5b1
fibronectin receptor to restrict carcinoma cell growth viainduction of p21 and p27. J Biol Chem 2005;280:37266—77.
[16] Villalobo A, Nogales-Gonzales A, Gabius H-J. A guide tosignaling pathways connecting protein—glycan interactionwith the emerging versatile effector functionality of mam-malian lectins. Trends Glycosci Glycotechnol 2006;18:1—37.
[17] Wang JL, Gray RM, Haudek KC, Patterson RJ. Nucleocyto-plasmic lectins. Biochim Biophys Acta 2004;1673:75—93.
[18] Smetana Jr K, Dvorankova B, Chovanec M, Boucek J, Klıma J,Motlık J, et al. Nuclear presence of adhesion-/growth-reg-ulatory galectins in normal/malignant cells of squamousepithelial origin. Histochem Cell Biol 2006;125:171—82.
[19] Purkrabkova T, Smetana Jr K, Dvorankova B, Holıkova Z,Dvorankova B, Bock C, et al. New aspects of galectin func-tionality in nuclei of cultured bone marrow stromal andepidermal cells: biotinylated galectins as tool to detectspecific binding sites. Biol Cell 2003;95:535—45.
[20] Nagy N, Legendre H, Engels O, Andre S, Kaltner H, Wasano K,et al. Refined prognostic evaluation in colon carcinoma usingimmunohistochemical galectin fingerprinting. Cancer 2003;97:1849—58.
[21] Kayser K, Trott J, BohmG, Huber M, Kaltner H, Andre S, et al.Localized fibrous tumors (LFTs) of the pleura: clinical data,asbestos burden, and syntactic structure analysis applied tonewly defined angiogenic/growth-regulatory effectors.Pathol Res Pract 2005;201:791—801.
[22] Lensch M, Lohr M, Russwurm R, Vidal M, Kaltner H, Andre S,et al. Unique sequence and expression profiles of rat galec-tin-5 and -9 as a result of species-specific gene divergence.Int J Biochem Cell Biol 2006;38:1741—58.
[23] Fronkova V, Holıkova Z, Liu F-T, Homolka J, Rijken DC, AndreS, et al. Simultaneous detection of endogenous lectins andtheir binding capability at the single cell-level-a technicalnote. Folia Biol 1999;45:157—62.
[24] Dubovy P, Svızenska I, Klusakova I. Computer-assisted quan-titative analysis of immunofluorescence staining of theextracellular matrix in rat dorsal and ventral spinal roots.Acta Histochem 2002;104:371—4.
[25] Watt FM. The stem cell compartment in human interfolli-cular epidermis. J Dermatol Sci 2002;28:173—80.
[26] Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors:stem cells and their niche. Cell 2004;116:769—78.
[27] Kanitakis J. Immunohistochemistry of normal skin. Eur JDermatol 1998;8:539—47.
[28] Tureckova J, Sahlberg C, Aberg T, Ruch JV, Thesleff I,Peterkova R. Comparison of expression of the msx-1, msx-2, BMP-2 and BMP-4 genes in the mouse upper diastemal andmolar tooth primordia. Int J Develop Biol 1995;39:459—68.
[29] Yamaguchi Y, Itami S, Tarutani M, Hosokawa K, Miura H,Yoshikawa K. Regulation of keratin 9 in nonpalmoplantarkeratinocytes by palmoplantar fibroblasts through epithe-lial-mesenchymal interactions. J Invest Dermatol 1999;112:483—8.
[30] Chovanec M, Smetana Jr K, Dvorankova B, Plzakova Z, AndreS, Gabius H-J. Decrease of nuclear reactivity to growth-regulatory galectin-1 in senescent human keratinocytes anddetection of non-uniform staining profile alterations uponprolonged culture for galectins-1 and -3. Anat Histol Embryol2004;33:348—54.
[31] Klıma J, Smetana Jr K, Motlık J, Plzakova Z, Liu F-T, Stork J,et al. Comparative phenotypic characterization of kerati-nocytes originating from hair follicles. J Mol Histol 2005;36:89—96.
[32] Gabius H-J. Glycohistochemistry: the why and how of detec-tion and localization of endogenous lectins. Anat HistolEmbryol 2001;30:3—31.
[33] Rotblat B, Niv H, Andre S, Kaltner H, Gabius H-J, Kloog Y.Galectin-1 (L11A) predicted from a computed galectin-1farnesyl-binding pocket selectively inhibits Ras-GTP. CancerRes 2004;64:3112—8.
[34] Gabius H-J. Cell surface glycans: the why and how of theirfunctionality as biochemical signals in lectin-mediatedinformation transfer. Crit Rev Immunol 2006;26:43—79.
[35] Yang H-X, Jin G-L, Meng L, Zhang J-Z, Liu W-B, Shou C-C.Screening and identification of proteins interacting withnucleostemin. World J Gastroenterol 2005;11:4812—4.
[36] Fan Y, Liu Z, Zhao S, Lou F, Nilsson S, Ekman P, et al.Nucleostemin mRNA is expressed in both normal and malig-nant renal tissues. Br J Cancer 2006;94:1658—62.