Embed Size (px)
Citation preview

Cell Tissue Res (1990) 261:313-322
andT' Reseatrh �9 Springer-Verlag 1990
Immuno-electron microscopy of atrial natriuretic factor secretory pathways in atria and ventricles of control and cardiomyopathic hamsters with heart failure* M. Cantin**, G. Thibault, H. Haile-Meskel, M. Ballak, R. Garcia, G. Jasmin, and J. Genest
Laboratory of Pathobiology, Clinical Research Institute of Montreal, Department of Pathology, Universit6 de Montreal, Montreal, Qu6bec, Canada
Accepted April 12, 1990
Summary. The secretory pathways of atrial natriuretic factor have been investigated in atrial and ventricular cardiocytes of control and cardiomyopathic Syrian ham- sters in severe congestive heart failure with four anti- bodies: a monoclonal antibody (2H2) against rat syn- thetic atrial natriuretic factor (101-126), which is directed against region 101 103 of rat atrial natriuretic factor (99-126), and polyclonal, affinity-purified antibodies produced in rabbits against synthetic C-terminal atrial natriuretic factor (101-126), synthetic N-terminal atrial natriuretic factor (11-37) or the putative cleavage site of atrial natriuretic factor (98-99) : atrial natriuretic fac- tor (94~103). Application of the immunogold technique on thin frozen sections ( immunocryoultramicrotomy) re- vealed an identical picture with the four antibodies. In atria of both control and cardiomyopathic hamsters where atrial natriuretic factor secretion is regulated, the atrial natriuretic factor propeptide travels, uncleaved, from the Gotgi complex to immature and mature secre- tory granules. In ventricles of control hamsters, where secretion is constitutive, the atrial natriuretic factor pro- peptide travels from the Golgi complex to secretory vesi- cles. In the ventricles of hamsters with severe congestive heart failure, the Golgi complex is larger, secretory vesi- cles more abundant and a few secretory granules are present in ~ 20% of cardiocytes. Here again, the peptide travels uncleaved in all these pathways. These results reveal the pathways of secretion of atrial natriuretic fac- tor in atrial and ventricular cardiocytes and indicate that the propeptide is not cleaved intracellularly.
* Supported by a grant from the Medical Research Council of Canada to the Multidisciplinary Research Group on Hypertension, by the Canadian Heart Foundation and the Pfizer Company (Eng- land) ** Holder of the Jacques Genest Chair
Send offprint requests to : M. Cantin, Laboratory of Pathobiology, Clinical Research Institute of Montreal, 110 Pine Ave West, Mon- tr+al, Qu6bec, Canada H2W 1R7
Key words: Heart - Atrial natriuretic factor Cardio- myopathic animals - Golden hamster (Mesocricetus auratus)
The atria in mammals comprise an endocrine gland (Genest and Cantin 1988; de Bold 1985; Needleman and Greenwald 1986) producing in the rat atrial natriuretic factor (ANF), a 28-amino-acid peptide (99-126) (Schwartz et al. 1985; Thibault et al. 1985) which is diuretic, natriuretic, vasodilatory and inhibitory of al- dosterone, cortisol, arginine vasopressin and renin re- lease. In the rat (Katsube et al. 1986; Michener et al. 1986; Thibault et al. 1988) and the golden Syrian ham- ster (Thibault et al. 1989b), the N-terminal moiety (1-98) of the propeptide (1-126) of A N F is also present in the circulation. The atria in both rat (Cantin et al. 1987) and golden Syrian hamster (Ding et al. 1987) contain only the propeptide which is also present in rat atrial secretory granules (Thibault et al. 1987), as in the ven- tricular extracts of control rats (Cantin et al. 1987), con- trol hamsters (Ding et al. 1987) and hamsters with severe congestive heart failure (Ding et al. 1987). In cardiomyo- pathic hamsters with heart failure, the plasma levels of both C- and N-terminal A N F are markedly elevated and the ratio of IR-ANF between atria and ventricles, which is 132:1 in control hamster heart, becomes 4:1 in animals with severe congestive heart failure because of a decrease in the atrial levels and a concomitant in- crease in ventricular levels (Cantin et al. 1988). This is accompanied by a decrease of atrial ANF m R N A and marked increase in ventricular A N F m R N A (Thibault et al. 1989b).
In the present study, the immunogold technique on thin frozen sections (immunoultramicrotomy) and ap- propriate antibodies were used: (1) to delineate the secre- tory pathways of A N F in atria and ventricles of control and cardiomyopathic hamsters with severe heart failure; (2) to find out if cleavage of the ANF propeptide into

314
its plasma components (C- and N-terminal) occurs with- in these secretory pathways.
Materials and methods
Animals
Cardiomyopathic hamsters in severe heart failure were from the UM-X7.1 myopathic line established by cross-breeding diseased animals from the original B I t 14.6 strain of the Bit-Research Institute (Cambridge, Ma, USA) with unrelated healthy hamsters (Ding et al. 1987; Cantin et al. 1988; Jasmin and Eu 1979). Golden Syrian hamsters (Mesocricetus auratus) (Charles River, St-Con- stant, Quebec) of the same age (250-300 days) were used as con- trols. All animals were maintained on regular pellet chow (Purina, Richmond, Ind, USA) and tap water ad libitum in plastic cages at 22 ~ C with 60% humidity under a 6 a.m.-6 p.m. light regimen.
Assessment of the severity of congestive heart failure
The animals in severe heart failure (Ding et al. 1987; Cantin et al. 1988) showed generalized edema of the subcutaneous areas of the entire body (including the facial part) together with ascites, hydrothorax and hydropericardium. The majority of skeletal mus- cle groups, especially those of the hind legs and abdominal walls were markedly swollen. The mesentery of the intestine and the pancreas were highly edematous and the measurable exudates in the body cavities amounted to well over 2.5 ml. Diffuse necrotic and fibrotic lesions were present in the congested liver, and hemosi- derin-laden macrophages (heart-failure cells) were consistently seen in the lungs.
Peptides
Rat A N F (92-126) and (95-126) and human A N F (7%98) were purchased from Peninsula Laboratories Inc. (Belmont, Ca, USA). Rat A N F (94-103), (99-126) and human ANF (11-37) were syn- thetized by Bio-M6ga Inc. (Laval, Qu6bec, Canada), ANF (1-126) was purified by HPLC from rat atrial granules as reported earlier (Thibault et al. 1987) and ANF (1-98) was produced by enzymatic hydrolysis of the precursor (Seidah et al. 1986).
Antibody production and purification
Four different antibodies were used throughout. A monoclonal antibody (2H2) against rat synthetic ANF (101 126) which is di- rected against region 101-103 of rat ANF (99 126) (Milne et al. 1987) was used. Polyclonal antibodies were produced in rabbits either against the synthetic C-terminal A N F (101 126) or synthetic N-terminal A N F (11-37) or the putative cleavage site (residues 98 99); A N F (94-103). All these antibodies were purified by affini- ty chromatography and monospeciflcity determined by Western blot (Cantin et al. 1989; Thibault et al. 1989a). These results con- firmed several studies in which the cross-reactivity of each antibody was thoroughly assessed by other techniques for RIA or immuno- cytochemical purposes (Thibault et al. 1988; Chapeau et al. 1985; Gutkowska et al. 1986).
Radioimmunoassays
To test the cross-reactivity of the different peptides toward the antibodies, radioimmunoassays were set up. The ANF (101-126) antibodies (Ab 4~ 17) were tested using 125I-ANF (101-126) as al-
Table 1. Cross-reactivity of antibodies to A N F peptides (%)
ANF peptides Antibodies
#38 #30 #17 2H2 rat ANF human ANF rat ANF rat ANF (94-103) (11-37) (101-126) (101-126)
r ANF (1-126) 100 90 40 100 r ANF (1-98) <0.01 100 <0.01 <0.01 h ANF (1-30) - 100 - <0.01 h ANF (11-37) - 100 - <0.01 h ANF (79-98) 0.01 - - <0.01 r A N F (92-126) 80 < 0.01 100 r ANF (94-103) 20 - - - r ANF (95-126) 50 - - r A N F (99-126) <0.01 <0.01 100 100 h ANF (99-126) - <0.01 1 5 r A N F (103-126) - - 100 <0.01 r ANF (103-123) - 100 <0.01
ready reported (Gutkowska et al. 1986). The N-terminal antibodies (Ab 4t= 30) were tested using 125I-ANF (1 30) as tracer (Sundsjford et al. 1988). For the antibody against fragment 94-103 (Ab 4~38) labeled A N F (1 126) was used. Iodination of the peptides was performed by the lactoperoxidase method (Thibault et al. 1989a). The protocol of the RIAs is as follows: 100 gl of peptide were incubated with 100 gl of iodinated peptide (7000 cpm), 100 t-tl of diluted antibody and 100 gl of buffer (0.05 M NaPO4 pH 7.4, 0.13 M NaC1, 0.1% bovine serum albumin, 0.05% Triton X-100). After overnight incubation at 4 ~ C, the antibody-bound tracer was isolated by addition of 100 gl of normal rabbit serum (1:50), 100 gl of goat anti-rabbit IgG antiserum (1:50) and 1 ml of 12.5% poly- ethylene glycol 8000. The radioactivity present in the pellet was counted in a y-counter. The cross-reactivity of antibodies is given in Table 1.
Preparation for electron microscopy
Five control hamsters and five hamsters with severe congestive heart failure were perfused under pentobarbital anesthesia through the right and left ventricles first with 20 ml of Ringer-Locke fluid and then with 4% formaldehyde (prepared with paraformaldheyde) containing 0.1% glutaraldheyde and buffered with 0.1 M Na phos- phate (pH 7.2). Fragments of atria, of the interventricular septum and of the lateral portions of both right and left ventricles were kept in the same fixative for 2 h, washed in PBS three times for 15 min, embedded in 2.3 M sucrose and frozen in liquid nitrogen (Griffiths et al. 1983; Tokuyasu 1986).
Immunocytochemical labeling
The fine sections ( ~ 90 nm) from ultramicrotomy were treated as described elsewhere (Thibautt et al. 1989a; Griffiths et al. 1983; Tokuyasu 1986). They were placed on Formvar- and carbon-coated grids, floated on a drop of 1% goat serum diluted in TRIS buffer (20 mM TRIS-HC1; 150 mM NaC1; 0.1% BSA) pH 8.2 for 30 min, on a drop of the purified antiserum (either C-terminal, N-terminal or cleavage site antibodies) diluted in BSA-TRIS buffer for 30 rain, rinsed in BSA-TRIS for 2 periods of 5 min each, incubated on a drop of goat anti-rabbit or anti-mouse immunoglobulin-G-gold (GARIG: 15 nm, diluted 1:15 in BSA-TRIS) (Janssen Life Science Products; SPI Supplies, West Chester, Pa, USA) for 30min; washed for 2 periods of 5 min each in BSA TRIS, washed twice for 5 min each in distilled H20, stained first for 10 min with uranyl oxalic acid (5 ml of 4% uranyl acetate; 5 ml of 0.3 M oxalic acid);

315
All pictures are electron micrographs of atrial and ventricular cardiocytes processed according to the immunocryoultramicrotomy technique using various primary antibodies
Fig. 1 Control hamster atrial cardiocyte with nucleus (N), Golgi complex (G), immature granules (P) and mature secretory granules (g) immunostained with cleavage site antibodies (94-103) (Ab 4#38). x 112350
T a b l e 2. Comparative diameter of secretory granules and vesicles in atria and ventricles of control and cardiomyopathic hamsters with severe heart failure (nm)
Atrial cardiocytes
Granules
Immature Mature (Trans Golgi)
Ventricular cardiocytes
Granules Vesicles
Immature Mature Immature Mature (Trans Golgi) (Trans Golgi)
Control hamsters 68.9 _+ 2.6 185.3 + 4.0** - 58.2 _+ 3.1 57.8 _+ 2.9
Cardiomyopathic hamsters 80.1 _+3.0* 176.7+_3.1" ** - 137.3_+3.1"** 56.9_+3.0 57.9_+3.1 with severe congestive heart failure
- = Not seen The diameter of secretory granules and vesicles was measured in 50 electron micrographs for each cell type at a final magnification of x 40 800. At least 100 secretory granules and vesicles of each type were measured; * p < 0.05 as compared to controls; ** p<0.05 as compared to immature atrial granules; *** p <0.05 as compared to immature and mature secretory vesicles of animals with severe heart failure. Statistical analysis of data was performed according to the unpaired (vertical comparisons) or paired (horizontal comparison) Student's t-test
a pH of 7 was obtained by adding a few drops of NH4OH (diluted 5x), rapidly washed in distilled water 3 times and post-stained on ice in a mixture of 0.5 ml of 4% uranyl acetate, 0.25 ml of 2% methyl cellulose and 3.25 ml of distilled H20. The grids were then removed from the methyl cellulose solution with a loop, excess fluid removed with a filter paper and they were allowed to dry at room temperature (Thibault et al. 1989a; Griffiths et al. 1983; Tokuyasu 1986). They were visualized in a JEOL 1200EX electron microscope. The monoclonal antibody was used at a concentration of 0.05 lag/ml; the antibodies against the C-terminal (101-126) por- tion of A N F were used at a concentration of 0.03 gg/ml; the anti- bodies against the N-termianl (11-37) portion of ANF were used at a concentration of 1.0 gg/ml; the antibodies against the cleavage site of ANF (94~103) were used at a concentration of 0.05 lag/ml.
The specificity of immunostaining was assessed with the follow- ing controls: (1) Incubation of thin sections with an antiserum previously adsorbed with the homologous antigen (10 lag synthetic ANF (99 126) or ANF (1-98) or ANF (94-103) and solid phase immunoadsorption (Chapeau et al. 1985); (2) incubation of thin sections with antiserum followed by 1 h incubation with goat anti- rabbit or anti-mouse immunoglobulin-G (diluted 1:50) and the Garig complexes; and (3) omission of the antiserum step and appli- cation of the Garig complexes alone. All of these controls gave negative results, i.e., insignificant random deposits of gold particles
on the section. Adsorption of the cleavage site antiserum with 10 lag of ANF (99-126) or human ANF (7948) as above did not produce any change in labeling of Golgi structures and secretory granules or vesicles with immunogold.
R e s u l t s
In al l cases, t he resul t s w e r e i den t i ca l w i t h t he f o u r an t i - b o d i e s a n d wil l be d e s c r i b e d a c c o r d i n g l y .
Control hamster heart
Atrium. All the s ec re to ry g r a n u l e s w e r e heav i l y labe led . G o l g i saccules a n d ves ic les w e r e w e a k l y l abe led (Fig . 1). I m m a t u r e g r a n u l e s l o c a t e d c lose to the t r ans face o f t he G o l g i c o m p l e x w e r e sma l l e r (Table 2) a n d m u c h less heav i l y l abe led t h a n m a t u r e s e c r e t o r y g r a n u l e s l o c a t e d fa r f r o m the nuc leus , b u t m o r e t h a n G o l g i saccules a n d vesicles . In m a n y cells, G o l g i c o m p l e x e s s u r r o u n d e d by
316
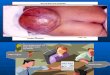
Fig. 2a-c. Ventricular cardiocytes from control hamsters. a Survey picture of ventricular cardiocyte immunostained with monoclonal (2H2) antibody with nucleus (N), labeled Golgi complex (G) and labeled secretory vesicles (v). Mitochondria (m), myofilaments with Z-bands (Z). x 26 300. b Lightly labeled secretory vesicles (v), labeled elements of the Golgi complex (G). m Mitochondrion. The fine-frozen section was labeled with cleavage- site antibodies (94~103) (Ab ~38). x 121900. e Labeled secretory vesicles (v) close to a nucleus (N). M filaments, m Mitochondrion. Polyclonal antibodies against N- terminal ANF (11 37) (Ab @30). x 46 400
secretory granules were found far f rom the nucleus, ordi- narily close to the sarcolemma. Here again, Golgi sac- cules and vesicles were labeled and there was an increas- ing gradient o f labeling f rom Golgi complex to small immature granules to larger mature secretory granules. The rough endoplasmic reticulum and all other cellular organelles were not labeled.
Ventricles. Sections made at various depths in the thick- ness of the mid-region of ventricular septum, and right and left ventricular free wall revealed an identical picture (Fig. 2 a-c). In practically every cell examined a few vesi- cles much smaller than atrial secretory granules and lo- cated either in between myofi laments or mitochondria in the center or at the periphery were labeled with gold particles. In all cases labeling was far less intense than
that seen over mature atrial secretory granules. In some cases, a few Golgi saccules or vesicles were also labeled at either nuclear pole or close to the sarcolemma. In several instances, these labeled Golgi structures were in close association with labeled vesicles. No structure re- sembling atrial secretory granules in size or intensity of labeling were seen.
Cardiomyopathic hamster heart: severe congestive heart failure
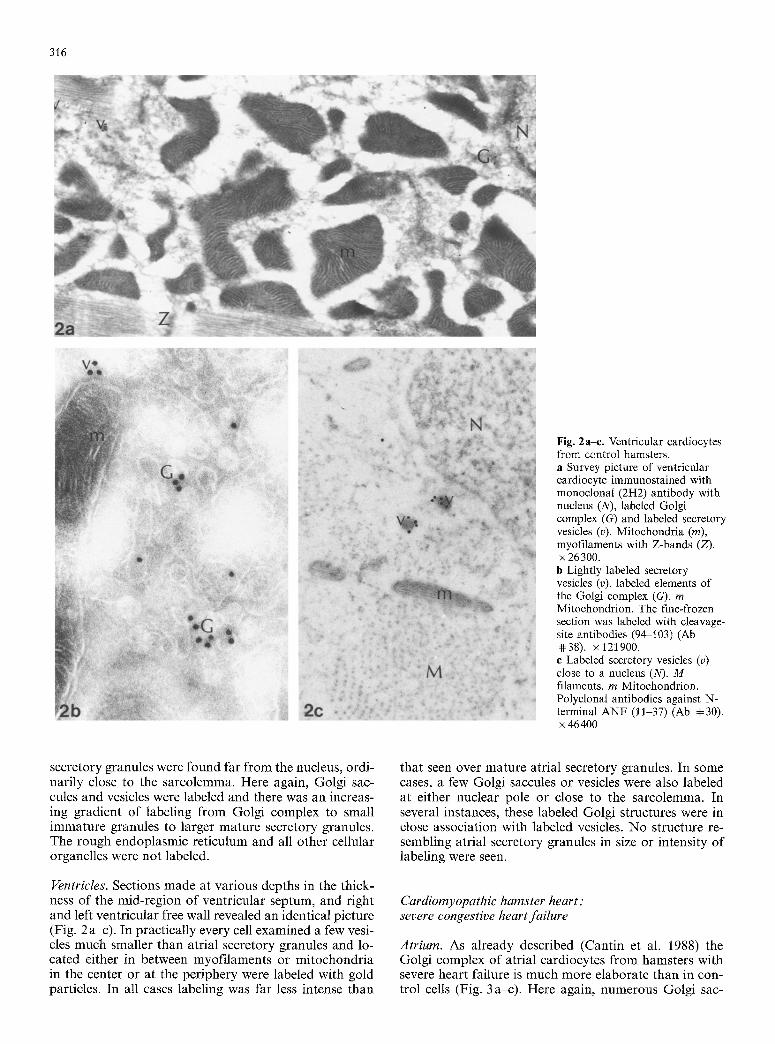
Atrium. As already described (Cantin et al. 1988) the Golgi complex of atrial cardiocytes f rom hamsters with severe heart failure is much more elaborate than in con- trol cells (Fig. 3 a-c). Here again, numerous Golgi sac-
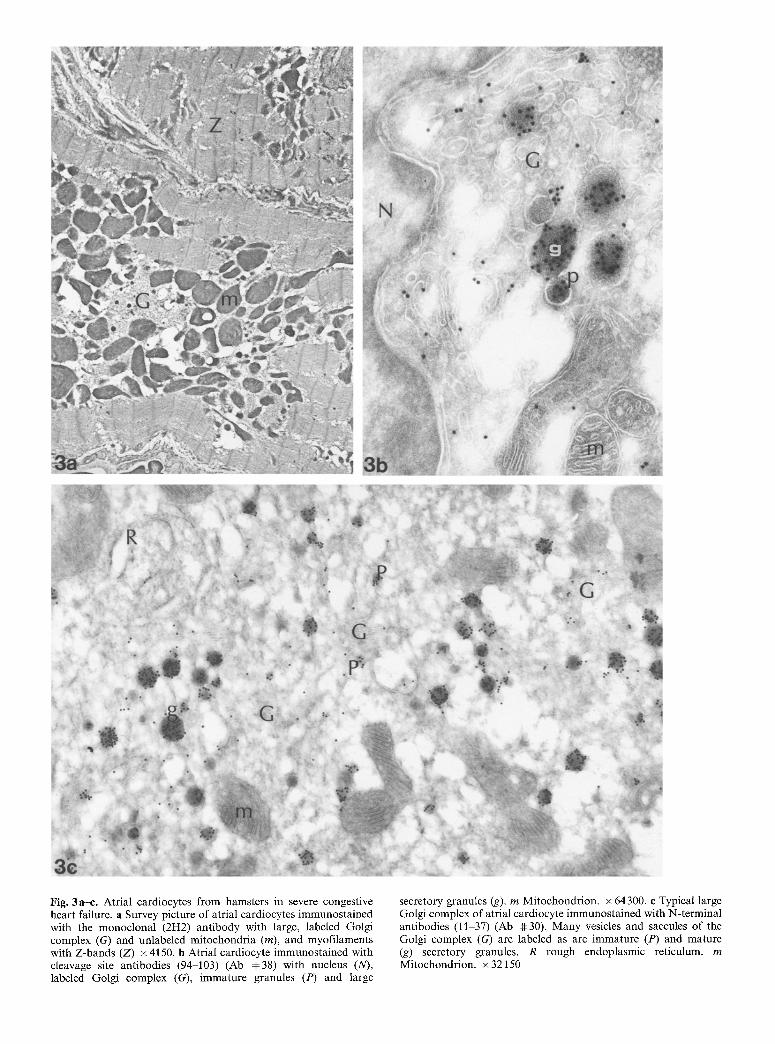
Fig. 3a-e. Atrial cardiocytes from hamsters in severe congestive heart failure, a Survey picture of atrial cardiocytes immunostained with the monoclonal (2H2) antibody with large, labeled Golgi complex (G) and unlabeled mitochondria (m), and myofilaments with Z-bands (Z) x4150, b Atrial cardiocyte immunostained with cleavage site antibodies (94-103) (Ab #38) with nucleus (N), labeled Golgi complex (G), immature granules (P) and large
secretory granules (g). m Mitochondrion. x 64300. e Typical large Golgi complex of atrial cardiocyte immunostained with N-terminal antibodies (11-37) (Ab ~ 30). Many vesicles and saccules of the Golgi complex (G) are labeled as are immature (P) and mature (g) secretory granules. R rough endoplasmic reticulum, m Mitochondrion. • 32150
318
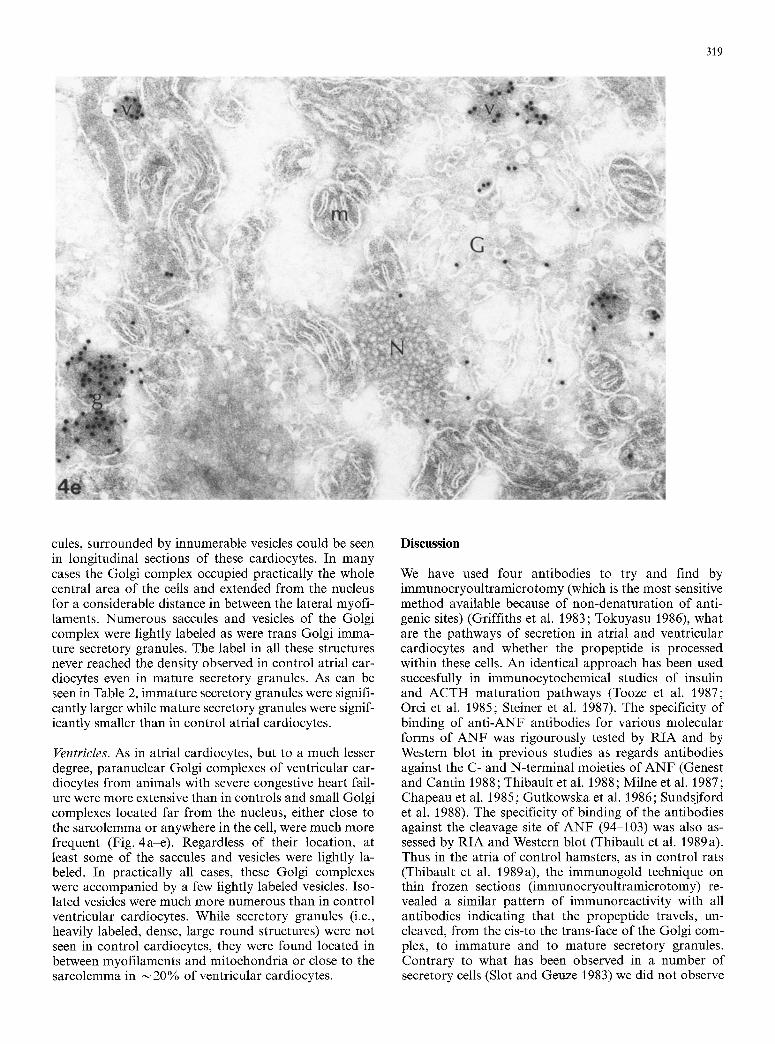
Fig. 4a-e. Ventricular cardiocytes from hamsters in severe congestive heart failure. a Paranuclear (N) labeled Golgi complex (G). Use of potyclonal antibodies against the C-terminal (101-126) (Ab 4t:17). x 110100. b Small labeled vesicles (v) near myofilaments (M). Use of polyclonal antibodies against the N-terminal (11-37) (Ab 4t: 30). x 115700.
e Labeled Golgi complex far from a nucleus with lightly labeled vesicles (v) and large labeled secretory granules (g). Use of polyclonal cleavage site (94-103) antibodies (Ab 4~38). x 94050. d High-power view of secretory vesicles (v). Some unlabeled structures possibly correspond to lysosomes (L). Use of monoclonal antibody (2H2). x84500. e Labeled Golgi complex (G) close to a nucleus with pores (N), with numerous saccules and vesicles, some of which are labeled. Two secretory granules (g) are more heavily labeled and larger than secretory vesicles (v). m Mitochondrion. Use of cleavage-site antibodies (94-103) (Ab 4#38). x68100
319
cules, surrounded by innumerable vesicles could be seen in longitudinal sections of these cardiocytes. In many cases the Golgi complex occupied practically the whole central area of the cells and extended from the nucleus for a considerable distance in between the lateral myofi- laments. Numerous saccules and vesicles of the Golgi complex were lightly labeled as were trans Golgi imma- ture secretory granules. The label in all these structures never reached the density observed in control atrial car- diocytes even in mature secretory granules. As can be seen in Table 2, immature secretory granules were signifi- cantly larger while mature secretory granules were signif- icantly smaller than in control atrial cardiocytes.
Ventricles. As in atrial cardiocytes, but to a much lesser degree, paranuclear Golgi complexes of ventricular car- diocytes from animals with severe congestive heart fail- ure were more extensive than in controls and small Golgi complexes located far from the nucleus, either close to the sarcolemma or anywhere in the cell, were much more frequent (Fig. 4a-e). Regardless of their location, at least some of the saccules and vesicles were lightly la- beled. In practically all cases, these Golgi complexes were accompanied by a few lightly labeled vesicles. Iso- lated vesicles were much more numerous than in control ventricular cardiocytes. While secretory granules (i.e., heavily labeled, dense, large round structures) were not seen in control cardiocytes, they were found located in between myofilaments and mitochondria or close to the sarcolemma in ~ 20% of ventricular cardiocytes.
Discussion
We have used four antibodies to try and find by immunocryoultramicrotomy (which is the most sensitive method available because of non-denaturation of anti- genic sites) (Griffiths et al. 1983; Tokuyasu 1986), what are the pathways of secretion in atrial and ventricular cardiocytes and whether the propeptide is processed within these cells. An identical approach has been used succesfully in immunocytochemical studies of insulin and ACTH maturat ion pathways (Tooze et al. 1987; Orci et al. 1985; Steiner et al. 1987). The specificity of binding of ant i-ANF antibodies for various molecular forms of ANF was rigourously tested by RIA and by Western blot in previous studies as regards antibodies against the C- and N-terminal moieties of ANF (Genest and Cantin 1988; Thibault et al. 1988; Milne et al. 1987; Chapeau et al. 1985; Gutkowska et al. 1986; Sundsjford et al. 1988). The specificity of binding of the antibodies against the cleavage site of ANF (94-103) was also as- sessed by RIA and Western blot (Thibault et al. 1989a). Thus in the atria of control hamsters, as in control rats (Thibault et al. 1989a), the immunogold technique on thin frozen sections (immunocryoultramicrotomy) re- vealed a similar pattern of immunoreactivity with all antibodies indicating that the propeptide travels, un- cleaved, from the cis-to the trans-face of the Golgi com- plex, to immature and to mature secretory granules. Contrary to what has been observed in a number of secretory cells (Slot and Geuze 1983) we did not observe

320
any increase in labeling from the cis-to the trans-face of the Golgi complex. The results, however, indicate that the number of gold particles was increased over mature secretory granules as compared to immature ones locat- ed close to the trans Golgi face. This is in agreement with previous studies. In the exocrine pancreas, condens- ing vacuoles, budding from the Golgi complex, aggre- gate to form mature secretory granules (Farquhar and Palade 1981). In prolactin cells of the anterior pituitary, small immature secretory granules, budding from the trans-face of the Golgi complex, aggregate and fuse to form mature secretory granules (Farquhar and Palade 1981). The same is true for the processing of proopiome- lanocortin in intermediate cells of the rat pituitary gland (Kalina et al. 1988) and in the rabbit parotid gland (Cope and Williams 1981 ; Williams and Cope 1981). The same phenomenon has also been observed in mouse juxtaglo- merular cells (Lacasse et al. 1985). These results thus reveal that the ANF propeptide is not cleaved either in the Golgi complex or in secretory granules whether immature or mature. The same conclusion emerges from the study of atrial cardiocytes in severe congestive heart failure even though the Golgi complex is much larger and more eleborate. In this situation, the immature se- cretory granules are significantly larger and the mature ones, as already described with another technique (Can- tin et al. 1988), are slightly but significantly smaller than in control atrial cardiocytes. These differences in size may be explained by a difference in the degree and speed of fusion of immature granules to form mature ones. The smaller size and lesser labeling of mature granules may be due to the very rapid transit and fast release in a situation characterized by lesser atrial levels of IR- ANF and ANF mRNA (Thibault et al. 1989b; Cantin et al. 1988) and markedly increased plasma levels (Can- tin et al. 1988).
In control hamster ventricular cardiocytes, as in the atria, ANF travels, uncleaved, from the Golgi complex to small trans Golgi secretory vesicles and to small vesi- cles which are found anywhere in the cells. The identical size of immature (trans Golgi) and mature secretory vesi- cles suggest absence of fusion. In ventricular cardiocytes from animals with severe heart failure, the same pattern is observed. In addition, typical secretory granules which are larger and more heavily labeled are also present in
20% of cardiocytes as already described with another technique (Cantin et al. 1988). In all cases, the propep- tide travels, uncleaved, from the Golgi complex to imma- ture and then to mature vesicles and granules. This is at variance with other cell types where constitutively secreted proteins like albumin from hepatocytes (Ed- wards et al. 1976; Ikehara et al. 1976) or a-mating factor from yeast (Julius et al. 1984) are properly cleaved. The same is true for viral membrane glycoproteins which are constitutively secreted and yet undergo proper pro- teolytic processing (Burgess and Kelly 1987). In any event, the pattern of ANF constitutive secretion in con- trol ventricular cardiocytes is identical to that found in the constitutive secretory pathway of albumin in hepato- cytes (Brands et al. 1983). The presence of a few secreto- ry granules in the ventricular cardiocytes of animals with
severe congestive heart failure suggest that in the case of hypersecretion, the cells return, at least partially, to an embryonal and neonatal type of secretory pathways (Hassall et al. 1988). In atria, generally, it may be de- duced by inference to the maturation of bioactive pep- tide hormones that ANF is initially translated as a large 150 to 152 amino acid preprohormone from which a signal sequence is probably co-translationally cleaved from the N-terminal to yield the 126 to 128 amino acid propeptide. The subsequent maturation steps are un- known except for the fact that intact prohormone has been isolated from atrial tissues (Glembotski et al. 1985; Trippodo et al. 1984; Miyata et al. 1985) and from atrial secretory granules (Thibault et al. I987) and is secreted from cultured rat atrial cardiocytes (Bloch et al. 1985; Glembotski and Gibson 1985). In the Langendorff prep- aration, however, the C-terminal is released from the perfused rat heart (Thibault et al. 1986). When, however, the propeptide is added to the tissue preparation, both the presence (Michener et al. 1986) and absence (Gibson et al. 1987) of proteolytic cleavage products have been reported in the perfusate. Analysis of the molecular forms of ANF in rat plasma has revealed unequivocally two major fragments: the C-terminal (99-126) (Schwartz et al. 1985; Thibault et al. 1985) and N-terminal (1-98) (Thibault et al. 1988) and a minor constituent (103-126) (Schwartz et al. 1985). In the hamster, both the C-termi- nal and the N-terminal portions of the ANF propeptide are also found in the circulation (Thibault et al. 1989 b). Previous studies have also revealed that ventricular car- diocytes from rat (Cantin et al. 1987) and hamster (Ding et al. 1987) contain the ANF propeptide and, in culture, rat ventricular cardiocytes release the propeptide in a constitutive fashion. The isolated ventricles from ham- sters in severe congestive heart failure release as much as 74% of the ANF found in the perfusate from the whole heart in the Langendorff preparation (Thibault et al. 1989b).
The isolated ventricles from both control and ham- ster with severe heart failure release a variable portion of ANF as the propeptide while the rest is properly cleaved as shown by HPLC analysis (Thibault et al. 1989b). These results thus suggest that while ANF from atria is properly cleaved, that from ventricles is only partially cleaved and some portion is released as the propeptide.
Taken together these results indicate that in both atria while the secretion is by definition regulated (Bur- gess and Kelly 1987) and in ventricles where the secretion is mostly constitutive, the ANF propeptide is not cleaved intracellularly. This is true even in cases of intense stimu- lation of secretory activity as in severe congestive heart failure. Experiments are now in progress to find out if cleavage occurs during exocytosis.
References
Bloch KD, Scott JA, Zisfein JB, Fallon JT, Seidman CE, Matsueda GR, Margolis MN, Homcy CJ, Graham RM, Seidman JG (1985) Biosynthesis and secretion of proatrial natriuretic factor by cultured rat cardiocytes. Science 230:1168 1171

321
Bold de AJ (1985) Atrial natriuretic factor: a hormone produced by the heart. Science 230 : 767 770
Brands RJ, Slot W, Geuze HJ (1983) Albumin localization in rat liver parenchymal cells. Eur J Cell Biol 32:99-107
Burgess TL, Kelly RB (1987) Constitutive and regulated secretion of proteins. Ann Rev Cell Biol 3 : 243-293
Cantin M, Ding J, Thibault G, Gutkowska J, Salmi L, Garcia R, Genest J (1987) Immunoreactive atrial natriuretic factor is present in both atria and ventricles. Mol Cell Endocrinol 52:105 113
Cantin M, Thibault G, Ding J, Gutkowska J, Garcia R, Jasmin G, Hamet P, Genest J (1988) ANF in experimental congestive heart failure. Am J Pathol 130:552-568
Cantin M, Thibault G, Haile-Meskel H, Ding J, Milne RW, Ballak M, Charbonneau C, Nemer M, Drouin J, Garcia R, Genest J (1989) Atrial natriuretic factor in the impulse-conduction sys- tem of rat cardiac ventricles. Cell Tissue Res 256:309-325
Chapeau C, Gutkowska J, Schiller PW, Milne RW, Thibault G, Garcia R, Genest J, Cantin M (1985) Localization of immuno- reactive synthetic atrial natriuretic factor (ANF) in the heart of various animal species. J Histochem Cytochem 33 : 541-550
Cope HC, Williams MA (1981) Secretion granule formation in the rabbit parotid gland after isoprenaline-induced secretion. Stereological reconstructions of granule populations. Anat Rec 199:377 387
Ding J, Thibault G, Gutkowska J, Garcia R, Karabatsos T, Jasmin G, Genest J, Cantin M (1987) Cardiac and plasma atrial na- triuretic factor (ANF) in experimental congestive heart failure. Endocrinology 121 : 248-257
Edwards K, Fleischer B, Dryburg H, Fleischer S, Schreider G (1976) The distribution of albumin precursor protein and albu- min in liver. Biochem Biophys Res Commun 72:310-318
Farquhar MG, Palade GE (1981) The Golgi apparatus (complex) (1954-1981). From artifact to center stage. J Cell Biol 19:77S- 103S
Genest J, Cantin M (1988) The atrial natriuretic factor: its physiol- ogy and biochemistry. Rev Physiol Biochem Pharmacol 110:1- 145
Gibson TR, Shields PP, Glembotski CC (1987) The conversion of atrial natriuretic peptide (ANP) (1-126) to ANP (99-126) by rat serum: contribution to ANP cleavage in isolated perfused rat hearts. Endocrinology 120:764-772
Glembotski CC, Gibson TR (1985) Molecular forms of immunore- active atrial natriuretic peptides released from cultured rat atrial myocytes. Biochem Biophys Res Commun 132:1008 1017
Glembotski CC, Wildey CM, Gibson TR (1985) Molecular forms of immunoreactive atrial natriuretic peptide in the rat hypotha- lamus and atrium. Biochem Biophys Res Commun 129:671- 678
Griffiths G, Simons K, Warren G, Tokuyasu KT (1983) Immuno- electron microscopy using thin, frozen sections: application to studies of the intracellular transport of Semliki Forest virus spike glycoproteins. Methods Enzymol 96:466M85
Gutkowska J, Horky K, Schiffrin EL, Thibault G, Garcia R, De L6an A, Hamet P, Temblay J, Anand-Srivastava MB, Janus- zewicz P, Genest J, Cantin M (1986) Atrial natriuretic factor: radioimmunoassay and effects on adrenal and pituitary glands. Fed Proc 45:2101-2105
Hassall CJS, Wharton J, Gulbenkian S, Anderson JV, Frater J, Bailey D J, Merichi A, Bloom SR, Polak JM, Burnstock G (1988) Ventricular and atrial myocytes of newborn rats synthe- size and secrete atrial natriuretic peptide in culture: light and electron microscopical localization and chromatographic exam- ination of stored and secreted molecular forms. Cell Tissue Res 251:161 169
Ikehara Y, Oda K, Kato K (1976) Conversion of proalbumin into serum albumin within secretory vesicles of rat liver. Biochem Biophys Res Commun 72:319-326
Jasmin G, Eu HY (1979) Cardiomyopathy in hamster dystrophy. Ann N Y Acad Sci 317:46-58
Julius D, Schekman R, Thornor R (1984) Glycosylation and pro-
cessing of preproalpha-factor through the yeast secretory path- way. Cell 36:309-318
Kalina M, Elmalek M, Hammel I (1988) Intragranular processing of proopiomelanocortin in the intermediate cells of rat pituitary glands. A quantitative immunocytochemical approach. Histo- chemistry 89:193 198
Katsube N, Schwartz P, Needleman P (1986) Atriopeptin turnover: quantitative relationships between '~ in vivo" changes in plasma levels and atrial content. J Pharmacol Exp Ther 239:474479
Lacasse J, Ballak M, Mercure C, Gutkowska J, Chapeau C, Foote S, M+nard J, Corvot P, Cantin M, Genest J (1985) Immunocy- tochemical localization of renin in juxtaglomerular cells. J Histochem Cytochem 33:323-332
Michener ML, Gierse JK, Seetharam R, Rok KF, Olins PO, Mai MS, Needleman P (1986) Proteolytic processing of atriopeptin prohormone. Mol Pharmacol 30: 552-557
Milne RW, Gutkowska J, Thibault G, Schiller PW, Charbonneau C, Genest J, Cantin M (1987) A murine monoclonal antibody against rat atrial natriuretic factor (ANF) which cross-reacts with mouse ANF. Mol Immunol 24:127-132
Miyata A, Kangawa K, Toshimori T, Katoh T, Matsuo H (1985) Molecular forms of atrial natriuretic polypeptide in mammalian tissues and plasma. Biochem Biophys Res Commun 129:248- 255
Needleman P, Greenwald JE (1986) Atriopeptin: a cardiac hor- mone intimately involved in fluid, electrolyte and blood pres- sure homeostasis. N Engl J Med 314:828-834
Orci L, Ravazzola M, Amherdt M, Madson O, Vassali JD, Perrelat A (1985) Direct identification of prohormone conversion site in insulin secreting cells. Cell 42:671-681
Schwartz D, Geller D, Manning PT, Siegel WR, Fok KF, Smith CF, Needleman P (1985) Ser-Leu-Arg-Arg-Atriopeptin III the major circulating form of atrial peptide. Science 229 : 397-400
Seidah NG, Cromlish JA, Hamelin J, Thibault G, Chr6tien M (1986) Homologous IRCM-serine protease @ 1 from pituitary, heart atrium and ventricles: a common prohormone maturation enzyme? Biosci Rep 6:835-844
Slot JW, Geuze NJ (1983) Immunoelectron microscopic explora- tion of the Golgi complex. J Histochem Cytochem 31 : 1049- 1086
Steiner DF, Michael J, Houghten R, Mathieu M, Gardner PR, Ravazzola M, Orci L (1987) Use of a synthetic peptide antigen to generate antisera reactive with a proteolytic processing site in native human proinsulin: demonstration of cleavage within clathrin-coated (pro) secretory vesicles. Proc Natl Acad Sci USA 84:6184-6188
Sundsjford JA, Thibault G, Larochelle P, Cantin M (1988) Purifi- cation and plasma determination of N-terminal fragment of pro-atrial natriuretic factor in human. J Clin Endocrinol Metab 66:605 610
Thibault G, Lazure C, Schiffrin EL, Gutkowska J, Chartier L, Garcia R, Seidah NG~ Chr6tien M, Genest J, Cantin M (1985) Identification of a biologically active circulating form of rat atrial natriuretic factor. Biochem Biophys Res Commun 130:981-986
Thibault G, Garcia R, Gutkowska J, Lazure C, Seidah NG, Chr6- tien M, Genest J, Cantin M (1986) Identification of the released form of atrial natriuretic factor by the perfused rat heart. Proc Soc Exp Biol Med 182;: 137-141
Thibault G, Garcia R, Gutkowska J, Bilodeau J, Lazure C, Seidah NG, Chr6tien M, Genest J, Cantin M (1987) The propeptide Asn 1-Tyr 126 is the storage form of rat atrial natriuretic factor. Biochem J 241:265-272
Thibault G, Murthy KK, Gutkowska J, Seidah NG, Lazure C, Chr6tien M, Cantin M (1988) NH2-terminal fragment of rat pro-atrial natriuretic factor in the circulation: identification, radioimmunoassay and half-life. Peptides 9:47 53
Thibault G, Haile-Meskel H, Wrobel-Konrad E, Ballak M, Garcia R, Genest J, Cantin M (1989a) Processing of the ANF propep- tide by atrial cardiocytes as revealed by immunocryoultramicro- tomy. Endocrinology 124: 3109-3116

322
Thibault G, Nemer M, Drouin J, Lavigne JP, Ding J, Haile-Meskel H, Charbonneau C, Garcia R, Genest J, Jasmin G, So!e M, Cantin M (1989 b) Ventricles are a major site of ANF synthesis and release in cardiomyopathic hamsters with heart failure. Circ Res 65:71-82
Tokuyasu KT (1986) Application of cryoultramicrotomy to im- munocytochemistry. J Microsc 143:139-149
Tooze J, Hollinshead M, Frank R, Burke B (1987) An antibody specific for an endoproteolytic cleavage site provides evidence
that pro-opiomelanocortin is packaged into secretory granules in ART20 cells before its cleavage. J Cell Biol 105:155-162
Trippodo NC, Ghai RD, MacPhee AA, Cole FE (1984) Atrial natriuretic factor: atrial conversion of high to low molecular weight forms. Biochem Biophys Res Commun 119:282-288
Williams MA, Cope HC (1981) Membrane dynamics in the par- otid acinar cells during regranulation: a stereological study following isoprenaline-induced secretion. Anat Rec 199:389 401