Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 93, pp. 12611-12615, October 1996Physiology
A neurotensin antagonist, SR 48692, inhibits colonic responses toimmobilization stress in ratsIGNAZIO CASTAGLIUOLO*, SUsAN E. LEEMANt, ERZSEBET BARTOLAK-SUKIt, SIGFUS NIKULASSONt, BOSHENG QIU*,ROBERT E. CARRAWAY§, AND CHARALABOS POTHOULAKIS*¶*Division of Gastroenterology, Beth Israel Hospital, Harvard Medical School, Boston, MA 02215; Departments of tPharmacology and tPathology, BostonUniversity Medical Center Hospital, Boston University School of Medicine, Boston, MA 02118; and §Department of Physiology, University ofMassachusetts Medical Center, Worcester, MA 01655
Contributed by Susan E. Leeman, August 1, 1996
ABSTRACT We previously reported that short-term im-mobilization stress of rats causes increased colonic mucinrelease, goblet cell depletion, prostaglandin E2 secretion, andcolonic mast cell activation, as well as increased colonicmotility. The purpose of this study was to investigate whetherneurotensin (NT), a peptide expressed in both brain anddigestive tract, participates in these responses. Rats werepretreated with SR 48692 (1 mg/kg, i.p.), an NT antagonist,15 min before immobilization (30 min). The administration ofthe antagonist significantly inhibited stress-mediated secre-tion of colonic mucin, prostaglandin E2, and a product of ratmast cells, rat mast cell protease II (P < 0.05), but did notalter the increase in fecal pellet output caused by immobili-zation stress. Immobilization stress also resulted in a quan-tifiable decrease in the abundance ofNT receptormRNA in ratcolon compared with that in colonic tissues from nonimmo-bilized rats as measured by densitometric analysis of in situhybridization studies (P < 0.03). We conclude that the peptideNT is involved in colonic goblet cell release and mucosal mastcell activation after immobilization stress.
Neurotensin (NT), a 13-aa peptide originally isolated byCarraway and Leeman (1, 2), has a broad but discrete local-ization throughout the central nervous system and the diges-tive tract of animals and humans (2-4). NT has been identifiedin specific endocrine cells, named N-cells, in the ileal mucosaof dogs (5) and humans (6). The known intestinal effects ofNTinclude stimulation of growth of small and large bowel,pancreas, and stomach (7-9); stimulation of small intestinalsecretion (10); inhibition of small bowel and gastric motility(11); and stimulation of colonic motor activity (12). It iswell-established that restraint of animals stimulated motility ofthe large intestine, as measured by colonic transit and fecalpellet output (13, 14). We recently reported that restraint ofrats causes increased mucin release, as measured by [3H]glu-cosamine incorporation and goblet cell depletion, prostaglan-din E2 (PGE2) secretion, and mast cell activation in colonicexplants (15).A number of observations suggest that NT may participate
in stress-related responses. For example, central administra-tion of NT in freely moving rats stimulates release of adreno-corticotropin hormone and corticosterone levels (16-18),indicating that it may be involved in the regulation of hypo-thalamic-pituitary-adrenal axis activity. Ceccatelli et al. alsoshowed up-regulation of the NT precursor mRNA in theparaventricular nucleus of the hypothalamus after immobili-zation stress (19).Augeron et al. (20) reported that NT stimulates mucin
secretion from a human colonic goblet cell line by a receptor-mediated mechanism. Because our data showed that immo-
bilization stress also stimulated colonic goblet cell degranula-tion and mucin secretion from rat colon (15), we postulatedthat NT is involved in colonic mucin secretion as well in othercolonic responses after immobilization stress.An important advance in the field of NT research has been
the development of the potent, highly specific, nonpeptide NTreceptor antagonist SR 48692, which inhibits NT binding andfunctional effects ofNT (21, 22). We report here the effects ofSR 48692 on colonic responses after immobilization stress.
MATERIALS AND METHODS
Materials. The NT antagonist SR 48692 (provided by SanofiRecherche, Toulouse, France) was dissolved in dimethyl sul-foxide (Sigma) and diluted to the final concentration withisotonic saline immediately before i.p. injection to rats. Thefinal concentration of dimethyl sulfoxide in all experimentswas 10%. Protein concentrations were determined by thebicinchoninic acid protein assay reagent obtained from Pierce.
Description of the Restraint Stress Model. Adult maleWistar rats (Charles River Breeding Laboratories) werehoused under controlled conditions on a 12-h light/12-h darkcycle and provided with food and water at libitum. Rats werehandled daily for 7 days before experiments to minimize thestress of handling. All experiments were performed between10 and 11 a.m. to minimize any influence of the circadianrhythm. Rats were injected i.p. with vehicle (saline containing10% dimethyl sulfoxide) or vehicle-containing 1 mg/kg SR48692. Stress effects were examined using the "wrap-restraint"model described by Williams et al. (13). Animals were re-strained by gentle wrapping of their upper and lower limbs withmasking tape, and then returned to their cages. Control ratswalked freely. After 30 min, the animals were killed with abolus of sodium pentobarbital (120 mg/kg, i.p.), the colon wasexcised and cut longitudinally, and sections (approximately 2 x2 mm) were cut and cultured for measurements of mucin,PGE2, and rat mast cell protease II (RMCPII) as describedbelow. Colonic motility was estimated by counting the numberof fecal pellets expelled during the immobilization period (14,23). (This study was approved by the Boston UniversityMedical Center and Beth Israel Hospital Institutional AnimalCare and Use Committee.)Measurement of Colonic Mucin Glycoprotein Release. Co-
lonic mucosal-explants were cultured in 35-mm tissues culturedishes (Fisher) in 1.5 ml of Trowell's medium (GIBCO/BRL)containing 10 ,Ci/ml [3H]glucosamine (New England Nu-clear; 1 Ci = 37 GBq) and 1% penicillin/streptomycin. Ex-plants (three sections per dish) were incubated at 37°C in a
Abbreviations: NT, neurotensin; RMCPII, rat mast cell protease II;PGE2, prostaglandin E2; NTR, NT receptor; CRF, corticotropin-releasing factor.ITo whom reprint requests should be addressed at: Beth IsraelHospital, Division of Gastroenterology, Dana 501, 330 BrooklineAvenue, Boston, MA 02215.
12611
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.

12612 Physiology: Castagliuolo et al.
5%/95% C02/02 atmosphere. After 18 h, the medium wasaspirated and trichloroacetic acid or phosphotungstic acid(Sigma) was added to a final concentration of 10% and 1%,respectively. Glycoproteins were allowed to precipitate over-night at 4°C. The mixture was then centrifuged (1600 x g for30 min at 4°C), and the pellet was resuspended in coldtrichloroacetic acid or phosphotungstic acid and centrifugedagain. This washing step was repeated three times to ensurecomplete removal of unincorporated [3H]glucosamine. Thefinal pellet was dissolved in 1 ml of 0.3 M NaOH. Samples wereneutralized with 0.1 ml of 0.6 M acetic acid, and radioactivitywas measured by scintillation counting. In some samples, toconfirm that [3H]glucosamine was incorporated into mucin,the trichloroacetic acid or phosphotungstic acid precipitatewas examined by density gradient ultracentrifugation as de-scribed (24, 25).Mucin secretion was also estimated in histological sections
by counting the number of goblet cells depleted of mucin.Colonic samples obtained at the end of the 30-min immobili-zation stress were fixed in formalin, paraffin-embedded, andstained with hematoxylin and eosin, and Alcian blue. For eachsample, the number of goblet cells containing mucus wasestimated in an area that included 10 parallel colonic crypts.Because previous results from our laboratory indicated thatimmobilization stress reduces the number of surface, but notcrypt, goblet cells (15), we restricted our measurements to thesurface goblet cells only. Sections were examined in a blindedfashion by a gastrointestinal pathologist (S.N.), and resultswere expressed as the number of goblet cells containing mucinper 100 colonic surface epithelial cells.Measurement of PGE2 and RMCPII. Mucosal explants
(three sections per dish) were incubated (at 37°C for 30 min)in 1 ml of modified Kreb's buffer (26). After 30 min, themedium was replaced with fresh medium, and explants wereincubated for an additional 2 h at 37°C. RMCPII and PGE2were measured in 100-,ul aliquots of the supernatants. RMC-PII was measured by a sandwich ELISA kit (MORENDUNAnimal Health LTD, Edinburgh) as we described previously(27, 28), and results were expressed as nanograms of RMCPIIper milligram of tissue wet weight. PGE2 was measured by animmunoenzymatic assay (PerSeptive Diagnostics, Cambridge,MA), and results were expressed as picograms of PGE2 permilligram of tissue wet weight.
In Situ Hybridization for NT Receptor mRNA. cDNA tem-plate. Sections were hybridized with a 383-base RNA probecomplimentary to the distal 3' untranslated portion of NTreceptor (NTR) mRNA. The NTR mRNA was synthesizedfrom a cDNA that was provided by S. Nakanishi (KyotoUniversity, Kyoto, Japan) and subsequently subcloned by us.11To generate antisense and sense cRNA probes, the cDNA waslinearized with XbaI and XhoI, respectively. Experiments withRNase pretreatment followed by hybridization gave no hybrid-ization signal with either antisense or sense probes.Probe synthesis. The 35S-CTP-labeled RNA probes (Amer-
sham) were synthesized using a MAXIscript T3/T7 kit (Am-bion, Austin, TX). Labeled cRNA was purified with QuickSpin columns (Boehringer Mannheim) according to the man-ufacturer's instructions.
Hybridization. Control rats (n = 8) and rats after 30 min ofimmobilization stress (n = 7) were killed by i.p. injection ofsodium pentobarbital. Colonic tissues were removed, dis-sected, and then frozen in isopeptane on dry ice. Longitudinalsections (20-gm thick) were cut in a cryostat, thaw-mountedinto aminoalkylsilane-coated slides, and kept at 70°C. In situhybridization cytochemistry was carried out according to themethod of Miller et al. (29), which was modified by E.B.-S. and
M. Alexander (unpublished). Briefly, sections were prehybrid-ized for 3 h and hybridized for 18 h at 56°C using a probeconcentration of 1 pmol/ml. The hybridization buffer con-tained 47% formamide/9.4% dextran sulfate/0.28 M NaCl/1.25 x Denhardt's solution/9.4 mM Tris, pH 8/0.94 mMEDTA, pH 8/10mM DTT/tRNA to give a final concentrationof 0.5 mg/ml. The posthybridization steps included a 30-minRNase wash followed by washes with decreasing concentra-tions of standard saline citrate (SSC; from 2x to 0.1 X) at roomtemperature and a 60-min 0.1 x SSC wash at 66°C. Sectionswere dehydrated in alcohol series. Sections used for semiquan-titation were exposed to Hyperfilm ,Bmax (Amersham) for 10days and counterstained with thionin.
Semiquantitation of 35S-CTP-cRNA hybridization. Imageswere obtained by apposing slide-mounted plane sections and14C-labeled standards to Hyperfilm f3max for 10 days. Com-puter-aided microdensitometry was performed with aMCID-MI system (Imaging Research, St. Catherine's, ON,Canada) interfaced with a Nikon CCD video camera equippedwith a X2 Nikon objective. After the system had been cali-brated, a standard curve that was used for relative comparisonsof mRNA was created to convert optical density to linearizedintensity. Intensity values were obtained from the entire tissuesections and were then corrected for background intensity bysubtracting the intensity of areas showing no labeling. Dupli-cate readings were obtained, and mean values were calculated.
Statistical Analyses. Statistical analyses were performedusing SIGMASTAT (version 1.00, Jandel, San Rafael, CA).ANOVA was used for intergroup comparisons.
RESULTSInhibition by the NT Antagonist, SR 48692, on Stress-
Induced Colonic Mucin Release. Mucin release was assessedby measuring [3H]glucosamine-labeled glycoproteins in super-natants of colonic explants and by histologic examination. Asexpected (15), colonic explants from rats restrained for 30 minreleased significantly more [3H]glucosamine-labeled glycopro-teins in vitro than controls (Fig. 1, P < 0.01). Histologicexamination of colonic tissues also showed a significant re-duction in the number of superficial goblet cells containingmucus per 100 colonic surface epithelial cells in stressed rats
30000
4)
a_ CL
6 E
ISO cnC~c
.COm
O(3oC
Az
25000
20000
15000
10(000
5000
0 .1
+L~~1~~~
Stress
I.
_ISR 48692+ Stress
FIG. 1. Effect of the NT antagonist SR 48692 on immobilizationstress-induced colonic mucin glycoprotein release. Rats were injectedi.p. with saline (control) or saline containing 1 mg/kg SR 4869215 minbefore immobilization stress. Rats were then immobilized, whereascontrol rats moved freely in their cages. After 30 min, rats were killed,colon was removed, and colonic explants (n = 3 per dish) werecultured in medium containing [3H]glucosamine and incubated at37°C for 18 h. Mucin release was measured by incorporation of[3H]glucosamine into trichloroacetic acid or phosphotungstic acidprecipitates of culture supernatants. Six to twelve rats were tested foreach condition. Results are expressed as mean ± SEM per group. +,P < 0.05; + +, P < 0.01 versus control; *, P < 0.05 versus stress.
I1Bartolak-Suki, E., Alexander, M. J. & Leeman, S. E., 15th AnnualMeeting of the Society for Neuroscience, Nov. 11-16, 1995, SanDiego, p. 42.7 (abstr.).
Proc. Natl. Acad. Sci. USA 93 (1996)
Control

Proc. Natl. Acad. Sci. USA 93 (1996) 12613
-
.1."I --vr,-!rA V'* i.,..4", -7
, -W.,;, 4"
.7-.- !, 4ro, .1 1
FIG. 2. Inhibition of stress-induced goblet cell depletion of mucin by the NT antagonist SR 48692. Rats were injected i.p. with either saline orsaline containing the NT antagonist SR 48692 15 min before immobilization stress. Rats were then immobilized, whereas control rats moved freelyin their cages. After 30 min, rats were killed and full thickness samples of colon were fixed in formalin and stained with hematoxyline and eosin,and Alcian blue. (a) Colon from nonimmobilized rats showing normal mucosal architecture with many goblet cells containing mucin in the cryptsas well as in the superficial epithelium. (b) Colon obtained from a rat exposed to 30 min of immobilization stress showing disappearance ofmucin-containing goblet cells in the superficial mucosal layer while goblet cells are unaffected. (c) Colon from a rat pretreated with SR 48692 15min before immobilization showing an increased number of mucin-containing superficial goblet cells. (X85.)
as compared with controls (Fig. 2 and Table 1), consistent withour previous observation (15). Pretreatment of rats with theNT antagonist SR 48692 15 min before immobilization stresssignificantly inhibited mucin release from cultured colonicexplants (Fig. 1, P < 0.05). In nonstressed animals, pretreat-ment with SR 48692 (1 mg/kg, i.p., n = 4) had no significanteffect on basal levels of colonic mucin (12,060 ± 180 dpm/mgprotein) as compared with levels of vehicle-injected controlanimals (11,750 ± 520 dpm/mg protein). Further confirmationof inhibition of stress-mediated mucin secretion by the NTantagonist was observed histologically. Treatment with SR48692 inhibited the reduction in the number of surface gobletcells containing mucin (Fig. 2 and Table 1).
Inhibition by the NT Antagonist on Stress-Mediated PGE2and RMCPII Release. Thirty minutes of immobilization stressresulted in a 2.3-fold and 7.1-fold increase in PGE2 (Fig. 3) andRMCPII (Fig. 4), respectively. Pretreatment with the NTantagonist significantly inhibited stress-mediated PGE2 (Fig.3, P < 0.05) and RMCPII release (Fig. 4, P < 0.05). Innonstressed animals, pretreatment with SR 48692 (1 mg/kg,i.p., n = 4) had no significant effect on basal levels of colonicPGE2 (10.2 ± 1.0 pg/mg of tissue wet weight) or RMCPII
Table 1. Effect of the NT antagonist SR 48692 on immobilizationstress-induced goblet cell depletion
No. of superficial goblet cellscontaining mucus per 100
colonocytes ± SEMNo stress (n = 7) 7.9 ± 1.0Stress (n = 10) 2.6 ± 0.5*Stress + SR 48692 (n = 7) 4.7 ± 0.5tt
Rats were injected i.p. with either saline or saline containing SR48692 15 min before immobilization stress. Rats were then immobi-lized, whereas control rats moved freely in their cages. After 30 min,rats were killed and full thickness samples of the colon were stainedwith hematoxyline and eosin, and Alcian blue. For each section, thenumber of superficial goblet cells containing mucus per 100 colono-cytes was counted by a single "blinded" histopathologist. Results areexpressed as means ± SEM per group.*, P < 0.01 versus no stress; tP < 0.05 versus stress; tP < 0.05.
(0.9 ± 0.08 ng/mg of tissue wet weight) as compared withlevels of vehicle-injected control animals (9.6 ± 1.5 pg/mg and0.74 ± 0.1 ng/mg of tissue wet weight for PGE2 and RMCPII,respectively).Lack of Effect of the NT Antagonist on Stress-Mediated
Effects in Colonic Transit. Immobilization stress for 30 minresulted in a 9-fold increase in fecal pellet output as comparedwith that for control rats (n = 8 per group, P < 0.01).Pretreatment with the NT antagonist (n = 7) did not signif-icantly inhibit the stress-mediated increase in fecal pelletoutput after restraint stress (data not shown).
30,L.
S4L 250
0 20-U)
15
..
0 100.
Nw
0a. 5-X)
0-
+
T
T..
Control Stress SR48692+ Stress
FIG. 3. Effect of the NT antagonist SR 48692 on stress-inducedPGE2 release from rat colon. Rats were injected with either saline orSR 48692 as described in the Fig. 1 legend. Rats were then immobi-lized, whereas control rats moved freely in their cages. After 30 min,rats were killed, colon was removed, and colonic explants (n = 3) wereplaced in organ culture dishes and incubated (at 37°C for 2 h) inmodified Kreb's buffer (26). One hundred-microliter aliquots ofmedium were collected and PGE2 levels were measured by an immu-noenzymatic assay as described. Results are expressed as picograms ofPGE2 released in the medium per milligram of tissue wet weight. Eachbar represents the mean ± SEM of 6-10 different experiments, eachwith duplicate determinations. +, P < 0.05 versus control; *, P < 0.05versus stress.
Physiology: Castagliuolo et aL
12614 Physiology: Castagliuolo et al.
a
7N
° 54
'-3 21
0 41
CL
0--
Control
+
Stress SR 48692+ Stress
FIG. 4. Effect of the NT antagonist SR 48692 on immobilizationstress RMCPII release. Rats were injected with either saline or SR48692 as described the Fig. 1 legend. After 30 min, immobilization ratswere killed, colon was removed, and colonic explants (n = 3) wereplaced in organ culture dishes and incubated (at 37°C for 2 h) inmodified Kreb's buffer (26). RMCPII was measured in 100-,ul aliquotsof the organ culture medium by a sandwich enzyme-linked immu-nosorbent assay. Each bar represents the mean ± SEM of 6-10different experiments, each with duplicate determinations. +,P < 0.05versus control; *, P < 0.05 versus stress.
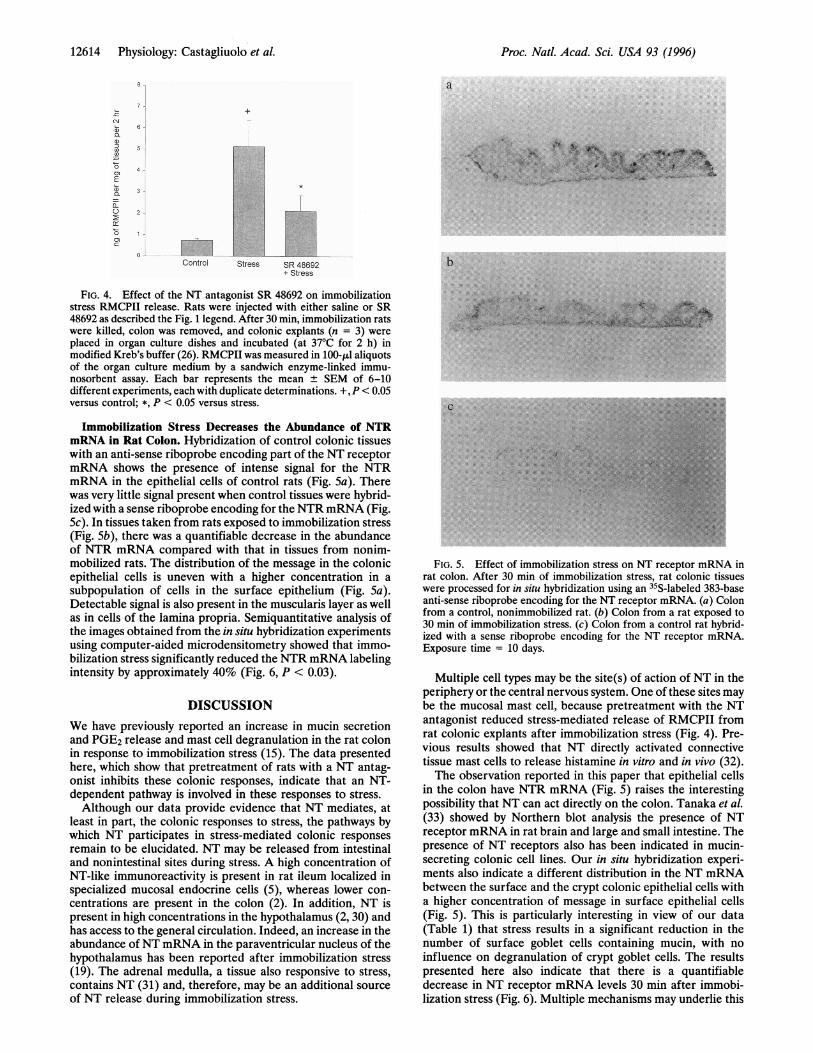
Immobilization Stress Decreases the Abundance of NTRmRNA in Rat Colon. Hybridization of control colonic tissueswith an anti-sense riboprobe encoding part of the NT receptormRNA shows the presence of intense signal for the NTRmRNA in the epithelial cells of control rats (Fig. Sa). Therewas very little signal present when control tissues were hybrid-ized with a sense riboprobe encoding for the NTRmRNA (Fig.Sc). In tissues taken from rats exposed to immobilization stress(Fig. Sb), there was a quantifiable decrease in the abundanceof NTR mRNA compared with that in tissues from nonim-mobilized rats. The distribution of the message in the colonicepithelial cells is uneven with a higher concentration in asubpopulation of cells in the surface epithelium (Fig. Sa).Detectable signal is also present in the muscularis layer as wellas in cells of the lamina propria. Semiquantitative analysis ofthe images obtained from the in situ hybridization experimentsusing computer-aided microdensitometry showed that immo-bilization stress significantly reduced the NTR mRNA labelingintensity by approximately 40% (Fig. 6, P < 0.03).
DISCUSSIONWe have previously reported an increase in mucin secretionand PGE2 release and mast cell degranulation in the rat colonin response to immobilization stress (15). The data presentedhere, which show that pretreatment of rats with a NT antag-onist inhibits these colonic responses, indicate that an NT-dependent pathway is involved in these responses to stress.Although our data provide evidence that NT mediates, at
least in part, the colonic responses to stress, the pathways bywhich NT participates in stress-mediated colonic responsesremain to be elucidated. NT may be released from intestinaland nonintestinal sites during stress. A high concentration ofNT-like immunoreactivity is present in rat ileum localized inspecialized mucosal endocrine cells (5), whereas lower con-centrations are present in the colon (2). In addition, NT ispresent in high concentrations in the hypothalamus (2, 30) andhas access to the general circulation. Indeed, an increase in theabundance ofNT mRNA in the paraventricular nucleus of thehypothalamus has been reported after immobilization stress(19). The adrenal medulla, a tissue also responsive to stress,contains NT (31) and, therefore, may be an additional sourceof NT release during immobilization stress.
.,,5,'.', ' ............
.;.. ........ ...s,s;,
.. .. .. " . . .~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~............ . ......~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~. ,.... . . .,:!. ..~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~. . .. ,...... .......: ...: ::::
c .... ....... .:
:: ..:* . ...... .. .. <M.. .............*... :.....
.....
:: ..
:........ .........
FIG. 5. Effect of immobilization stress on NT receptor mRNA inrat colon. After 30 min of immobilization stress, rat colonic tissueswere processed for in situ hybridization using an 35S-labeled 383-baseanti-sense riboprobe encoding for the NT receptor mRNA. (a) Colonfrom a control, nonimmobilized rat. (b) Colon from a rat exposed to30 min of immobilization stress. (c) Colon from a control rat hybrid-ized with a sense riboprobe encoding for the NT receptor mRNA.Exposure time = 10 days.
Multiple cell types may be the site(s) of action of NT in theperiphery or the central nervous system. One of these sites maybe the mucosal mast cell, because pretreatment with the NTantagonist reduced stress-mediated release of RMCPII fromrat colonic explants after immobilization stress (Fig. 4). Pre-vious results showed that NT directly activated connectivetissue mast cells to release histamine in vitro and in vivo (32).The observation reported in this paper that epithelial cells
in the colon have NTR mRNA (Fig. 5) raises the interestingpossibility that NT can act directly on the colon. Tanaka et al.(33) showed by Northern blot analysis the presence of NTreceptor mRNA in rat brain and large and small intestine. Thepresence of NT receptors also has been indicated in mucin-secreting colonic cell lines. Our in situ hybridization experi-ments also indicate a different distribution in the NT mRNAbetween the surface and the crypt colonic epithelial cells witha higher concentration of message in surface epithelial cells(Fig. 5). This is particularly interesting in view of our data(Table 1) that stress results in a significant reduction in thenumber of surface goblet cells containing mucin, with noinfluence on degranulation of crypt goblet cells. The resultspresented here also indicate that there is a quantifiabledecrease in NT receptor mRNA levels 30 min after immobi-lization stress (Fig. 6). Multiple mechanisms may underlie this
Proc. Natl. Acad. Sci. USA 93 (1996)

Proc. Natl. Acad. Sci. USA 93 (1996) 12615
0.030 -
0.025 -
0.020 -
0._
>,0.015-
a)_ 0.010
0.005
0.000
Control
(p<0.03)
T
Stress
FIG. 6. Densitometric analysis of the in situ hybridization studies(Fig. 5) of the abundance of NT mRNA in the rat colon followingimmobilization stress.
decrease, including a change in the stability of the message ora change in its transcription.
Previous studies showed that administration of the peptidecorticotropin-releasing factor (CRF), either parenterally orinto the cerebral ventrical of the paraventricular nucleus of thehypothalamus, in freely moving rats mimicked the effect ofstress on intestinal motility (14, 23). Also, administration of aCRF antagonist either i.v. or into the cerebrospinal fluidreduces the increased colonic transit caused by stress (23, 34).We have shown that in addition to motility changes, restraintstress causes colonic mucin and PGE2 release and activatescolonic mucosal mast cells (15). These colonic changes weremimicked by i.v. or intracerebral injection ofCRF in conscious,nonstressed rats and inhibited by pretreatment with the CRFantagonist a-helical CRF9_41 (15). In this report, althoughadministration of a NT antagonist did not reduce colonicmotility as measured by fecal pellet output, it inhibited theother colonic responses caused by i.v. CRF administration(35). It may be that the NT-dependent pathway(s) in theseCRF-mediated responses occurs locally within the colon.However, in relation to the data presented here that the NTantagonist inhibits colonic responses to immobilization stress,the antagonist may act to prevent CRF secretion from thehypothalamus. In this context, it is important to know thatthere is a wealth of data showing anatomic localization of NTand its receptor in a number of hypothalamic sites (36, 37)critical for the regulation of CRF secretion.
We would like to thank Sanofi Recherche for generously providingthe neurotensin antagonist SR 48692. This work has been supportedby National Institutes of Health Grants DK 47-343 (C.P.) and DK29-876 (S.E.L.). I.C. is a recipient of a Research Fellowship Awardfrom the Crohn's and Colitis Foundation of America, Inc.
1. Carraway, R. & Leeman, S. E. (1973) J. Bio. Chem. 248, 6854-6861.
2. Carraway, R. & Leeman, S. E. (1976) J. Bio. Chem. 251, 7045-7052.
3. Holzer, P., Bucsics, A., Saria, A. & Lembeck, F. (1982) Neuro-science 7, 2919-2924.
4. Doyle, N., Greely, G. H., Jr., Mate, L., Sakamota, T., Townsend,C. M., Jr., & Thompson, J. (1985) Surgery 45, 337-341.
5. Frigerio, B., Ravazola, M., Ito, S., Buffa, R., Capella, C., Solcia,E. & Orci, L. (1977) Histochemistry 54, 123-131.
6. Polak, J., Sullivan, S. & Bloom, S. (1977) Nature (London) 270,183-184.
7. Izukura, M., Evers, B. M., Parekh, D., Yoshinaga, K., Uchida, T.,Townsend, C. M. & Thompson, J. C. (1992) Ann. Surg. 215,520-527.
8. Evers, B. M., Izukura, M., Chung, D. H., Parekh, D., Yoshinaga,K., Greely, G. H., Jr., Uchida, T., Townsend, C. M., Jr., &Thompson, J. C. (1992) Gastroenterology 103, 86-91.
9. Feurle, G. E., Muller, B. & Rix, E. (1987) Gut 28, Suppl. 1,19-23.
10. Spokes, R. A., Lee, Y. C., Yiangou, Y., Domin, J. & Bloom, S. R.(1990) Eur. J. Pharmacol. 175, 43-47.
11. Anderson, S., Rosell, S., Hjelmquist, U., Chang, D. & Folkers, K.(1977) Acta Physio. Scand. 100, 231-235.
12. Bardon, T. & Ruckebusch, Y. (1985) Regul. Pept. 10, 107-114.13. Williams, C. L., Peterson, J. M., Villar, R. G. & Burks, T. F.
(1987) Am. J. Physio. 253, G582-G586.14. Williams, C. L., Villar, R. G., Peterson, J. M. & Burks, T. F.
(1988) Gastroenterology 94, 611-621.15. Castagliuolo, I., LaMont, J. T., Qiu, B., Fleming, S. M., Bhaskar,
K. R, Nikulasson, S. T., Kornetsky, C. & Pothoulakis, C. (1996)Am. J. Physiol., in press.
16. Fuxe, K., Agnati, L., Anderson, K., Eneroth, P., Horfstrand, A.& Golstein, Z. M. (1984) Neurochem. Int. 6, 737-750.
17. Gudelsky, G. A., Berry, S. A. & Meltzer, H. Y. (1989) Neuroen-docrinology 49, 604-609.
18. Rowe, W., Viau, V., Meaney, M. & Quirion, R. (1992)Ann. N.YAcad. Sci. 668, 365-367.
19. Ceccatelli, S., Vilar, M. J., Goldstein, M. & Hokfelt, T. (1989)Proc. Natl. Acad. Sci. USA 86, 9569-9573.
20. Augeron, C., Voisin, T., Maoret, J. J., Berthon, B., Laburthe, M.& Laboisse, C. L. (1992) Am. J. Physiol. 262, G470-G476.
21. Gully, D., Canton, M., Boigegrain, R., Jeanjean, F., Molimard,J. C., Poncelet, M., Gueudet, C., Heaulme, M., Leris, R.,Brouard, A., Pelaprat, D., Labbd-Jullie, C., Mazella, J., Maf-frand, J. P., Rostene, W., Kitabgi, P. & Le Fur, G. (1993) Proc.Natl. Acad. Sci. USA 90, 65-69.
22. Nicot, A., Berod, A., Gully, D., Rowe, W., Quirion, R., Ron deKloet, E. & Rostene, W. (1994) Neuroendocrinology 59, 527-578.
23. Monnikes, H., Schmidt, B. G. & Tache, Y. (1993) Gastroenter-ology 104, 716-723.
24. Bhaskar, K. R. & Greeth, J. I. (1974) Biochem. J. 143, 669-679.25. Gong, D., Turner, B. K., Bhaskar, K. R. & LaMont, J. T. (1990)
Am. J. Physio. 259, G681-G686.26. LaMorte, W. W., LaMont, J. T., Booker, M. L., Scott, T. E. &
Turner, B. K. (1986) Am. J. Physiol. 251, G701-G709.27. Castagliuolo, I., LaMont, J. T., Letourneau, R., Kelly, C. P.,
O'Keane, J. C., Jaffer, A., Theoharides, T. C. & Pothoulakis, C.(1994) Gastroenterology 107, 657-665.
28. Pothoulakis, C., Castagliuolo, I., LaMont, J. T., Jaffer, A.,O'Keane, A. J., Snider, R. M. & Leeman, S. E. (1994) Proc. Natl.Acad. Sci. USA 91, 947-951.
29. Miller, M. A., Kolb, P. E., Planas, B. & Raskind, M. A. (1994)Endocrinology 135, 1986-1995.
30. Emson, P. C., Goedert, M., Horsfield, P., Rioux, F. & St. Pierre,S. (1982) J. Neurochem. 38, 992-999.
31. Ferris, C. F., Carraway, R. E., Brandt, K. & Leeman, S. E. (1986)Neuroendocrinology 43, 352-358.
32. Carraway, R., Cochrane, D. E., Lansman, J. B., Leeman, S. E.,Paterson, B. M. & Welch, H. J. (1982) J. Physio. (London) 323,403-414.
33. Tanaka, K., Masu, M. & Nakanishi, S. (1990) Neuron 4,847-854.34. Lenz, H. J., Raedler, A., Greten, H., Vale, W. W. & Rivier, J. E.
(1988) Gastroenterology 95, 1510-1517.35. Castagliuolo, I., Qiu, B. S., Leeman, S. E., LaMont, J. T. &
Pothoulakis, C. (1996) Gastroenterology 110, 1062 (abstr.).36. Ceccatelli, S., Ericksson, M. & Hokfelt, T. (1989) Neuroendocri-
nology 49, 309-323.37. Young, W. S., III, & Kuhar, M. J. (1981) Brain Res. 206,
273-285.
Physiology: Castagliuolo et aL





























