Embed Size (px)
Citation preview

Oecologia (1996) 105:141-150 �9 Springer-Verlag 1996
Martha R. Downs �9 Knute J. Nadelhoffer Jerry M. Melillo �9 John D. Aber
Immobilization of a lSN-labeled nitrate addition by decomposing forest litter
Received: 9 February 1995 / Accepted: 21 July 1995
Abstract Effects of chronic HNO 3 and H2SO 4 additions on decomposition of senesced birch leaf, beech leaf, spruce needle, and wood chip litters were examined. Lit- ters were incubated for up to 4 years in fiberglass mesh (1 ram) bags on experimental plots in a mixed-species forest near the Bear Brooks Watershed Manipulation (BBWM)
' site in eastern Maine, United States. Plot treatments in- cluded HNO 3 additions at 28 and 56 kg N �9 ha -1 �9 year-l, H2SO 4 additions at 128 kg S - ha -1 �9 year -1, and a com- bined HNO 3 and HzSO 4 treatment at 28 kg N and 64 kg S �9 ha -1 . year-1. The 15N content of all NO 3 added was arti- ficially increased to 344%o 815N. Litter bags were collect- ed each fall and analyzed for organic matter loss, nitrogen concentration, and ~5N abundance throughout the 4-year experiment. Extractive (non-polar-soluble+water-solu- ble), cellulose (acid-soluble), and lignin (acid-insoluble) fractions were analyzed for the first 2 years. In wood chips, nitrogen additions increased mass loss and N con- centration, but not the mass of N after 4 years. Neither N nor S additions had large effects on mass loss, N concen- tration, or N content of leaf litters. All litters immobilized and mineralized N simultaneously, but we were able to place a lower bound on gross N immobilization by mass balancing 15N additions. Birch and spruce litters showed net mineralization, while beech leaf and wood chip litters showed net immobilization. Net immobilizing litters were those with the highest initial cellulose concentration (wood chips=80%, beech leaves=54%), and we attribute the higher capacity for immobilization to more readily available carbon. Lignin mass increased initially in all lit- ter types except spruce needles. Also, extractives in net immobilizing litters either increased initially (wood chips) or decreased at a slower rate than bulk litter (beech leaves). We calculate the potential of decomposing litter to immobilize exogenous nitrate in this system to be
M. R. Downs ( ~ ) �9 K. J. Nadelhoffer �9 J. M. Melillo The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA 02543, USA
J. D. Abet Complex Systems Research Center, University of New Hampshire, Durham, NH 03824, USA
1-1.5 kg N �9 ha -I �9 yea~ 1, which is about half of the usual NO 3 deposition at this site, but only a small fraction of the experimental addition�9
Key words Decomposition �9 N immobilization - N saturation �9 Nitrate - ~SN
Introduction
Over the past 10 years, the focus of research on atmo- spheric deposition has shifted from sulfuric acid to N de- position, with increased attention to the concept of N sat- uration. A system becomes N-saturated when N inputs exceed the capacity of the system to retain and use N (Aber et al. 1989; Schulze 1989). Nitrogen saturation is associated with increased rates of N cycling and losses of nitrate to drainage waters (Nihlgard 1985; Kahl et al. 1993). Because decomposition of recent plant litter is be- lieved to be N-limited in many situations (Aber and Mel- illo 1980; Melillo et al. 1982), it is possible that in- creased N availability will accelerate decomposition and N cycling rates.
Several studies have examined the impact of acid de- position (most often H2SO4) on decomposition. Sulfuric acid appears to reduce mass loss, at least at relatively high deposition rates, but the role of nitric acid with re- spect to mass loss is unclear. Only a few studies have ex- amined the effects of either nitric or sulfuric acid on ni- trogen dynamics in decomposing litter. Prescott and Par- kinson (1985) found lower rates of pine needle decompo- sition at a higher sulfur loading (28 vs. 23 and 12 kg S �9 ha -~ �9 year~l). B~Sth et al. (1980) found decreased decom- position of pine needles in the 2 years following the com- pletion of H2SO 4 additions at 50 or 150 kg H2804-S �9 ha -1 �9 year- 1. At higher rates of S addition (32 or 96 kg S - ha -1 - yea1-1), Berg (1986) found conflicting results de- pending on vegetation type, as did Neuvonen and Suo- mela (1990) using mixed HNO 3 and H2SO 4 at pH 3 and pH 4. H~gvar and Kjondal (1981) noted decreased de- composition of birch leaves with H2SO 4 additions at pH 2, but not at pH 3 or 4.

142 OECOLOGIA 105 (1996) �9
In an air-pol lut ion gradient study in the San Bernardi- no mounta ins , Fenn and D u n n (1989) found increased rates of decomposi t ion and increased fungal diversity in areas subjected to higher levels of ni t rogen deposi t ion and ozone. A similar result was found by Tie tema (1993) compar ing oak leaf litter decompos ing in an N-saturated and an N-l imi ted site. Both of these studies, however compared decomposi t ion rates of native litters, so they do not dis t inguish be tween the indirect effect of N depo- sit ion through improved litter quali ty and the direct ef- fect of addit ional N supply. Reviewing the evidence, Fog (1988) found that N addit ions f requent ly yielded no change in decomposi t ion rates and occasional ly inhibi ted decomposi t ion.
We know of no studies that have addressed the direct inf luence of N or S deposi t ion on N dynamics in decom- posing litter. In this study, we examine the direct effects of nitric and sulfuric acid addit ions on litter decomposi- t ion at a site with relatively low levels of background de- posit ion. Furthermore, we explore mechan i sms control- l ing N dynamics by examin ing the changes in l ignin, cel- lulose, and soluble carbon fractions dur ing the first 2 years of decomposi t ion. Finally, we est imate the im- portance of litter decomposi t ion in re ta ining nitrate addi- t ions to the forest in which our study was conducted.
Methods
Treatments
The study was carried out on replicate 15x15 m forest plots of the Watershed Manipulation Project (WMP) adjacent to the Bear Brooks watersheds in eastern Maine, United States (44~ ' ' N, 6806'25 ' ' W), approximately 50 km inland from the Gulf of Maine and 60 km west of New Brunswick, Canada. Our decomposition experiment was part of an interdisciplinary study of the effects of N and S deposition on forest biogeochemistry, which is described more fully elsewhere (Kahl et al. 1993; Norton et al. 1993; Rustad et al. 1993; Nadelhoffer et al. in press). The plots were located on the southeast-facing slope of Lead Mountain at 260_+20 m eleva- tion. The site is dominated by American beech (Fagus grandifolia Ehrh.), red spruce (Picea rubra Sarg.), yellow birch (Betula ail- eghaniensis Britt.), and red, sugar, and striped maples (Acer ru- bruin, L., A. saccharum Marsh, and A. pennsylvanicum L.) grow- ing on spodosols. Background rates of N deposition are low, with about 75% of the total wet deposition of 4 kg N �9 ha q . yeal -I oc- curring as nitrate (Kahl et al. 1993). Sulfur deposition for the treatment period averaged 10.9 kg S �9 ha q �9 year -1 (J. Scofield, personal communication).
Three rows of six plots (one of each treatment) were separated by 15 m within rows and 30 m between rows. The six treatments were: control (water only), low N (28 kg N �9 ha -1 . yeal-1), high N (56 kg N �9 h~ 1 . year-l), low S (64 kg S �9 ha q �9 year-l), high S (128 kg S - ha -1 . year-l), and N+S (28 kg N and 64 kg S �9 ha 1 . yem-1). Treatments were applied at approximately 10-day inter- vals from May through October. Water was drawn from a subsur- face interception system and acids and ~5N were added and mixed in six 4000-1 polyethylene tanks at the summit. Treatment solu- tions were then gravity fed through polyethylene lines to quarter- arc sprayers located at the corners of each plot. Feedwater SQand NO 3 concentrations averaged 103 and 15 geq �9 1-1, respectively, from 1988 through 1992 (Norton et al. 1994). Treatment concen- trations were within 6% of target values (Rustad et al., unpub- lished work). Our targeted 15N label was 0.495 atom % 15N or, us- ing per mil notation, 350%0 815N. The average measured label was
344%0 515N (SE=0.96%~ ~15N, n=18). All plots were tested for evenness of spray distribution (Rustad et.al. 1993), which was found to be similar to forest throughfall.
Litter decomposition
We chose beech leaves, yellow birch leaves, and red spruce nee- dles because these are the dominant types of foliage on the plots. We also chose red maple wood chips to represent a litter type with very low N concentration and to examine the potential for decom- posmg woody biomass to immobilize added N. Beech and red spruce litters were collected at the WMP site in the fall of 1987, air-dried, sorted and thoroughly mixed. Wood chips (approximate- ly lx2x2.5 cm, bark-free) were collected from disease-free trees which had been felled near Blackhawk Island, Wisconsin in the fall of 1980. Yellow birch leaves were collected as part of a sepa- rate project at Harvard Forest, Massachussetts, in the fall of 1986, air-dried, sorted and mixed, All litter was stored under cool, dry, dark conditions until placed in the field.
Our litterbags were approximately 25x25 cm and made from 1-mm mesh fiberglass window screening sewn with polyester thread. Each bag initially contained approximately 15 g air-dry lit- ter. Each bag was uniquely identified and the initial mass of the bag and the litter it contained was recorded. Samples of initial ma- terial were set aside for air- to oven-dry weight conversion and ini- tial litter chemistry. Bags were strung together by litter type and inserted just under the fresh litter layer in May 1988, just prior to the intended start of treatments. Bags were retrieved in October of 1988, 1989, 1990 and 1991. All species were not distributed in all treatments and bags could not always be retrieved due to loss and animal damage (see Results Table 2 for the number of bags of each litter type retrieved from each treatment). We scraped surface debris off of each bag, and pulled out extending roots. Bags were then dried at 50~ and any remaining debris removed by brushing. Bag weights were recorded and the contents were removed. When roots remained in the bags, they were removed and their mass was subtracted from the litter weight. We ground the decomposed litter to pass a 0.84-mm sieve.
Analytical methods
Total N was measured on a Perkin-Elmer 240C or 2400 elemental analyzer, using acetanilide as a standard. We analyzed lignin and cellulose using the forest products technique outlined in McClaugherty et al. (1985) and refined in Ryan et al. (1990). This technique employs sequential extractions in methylene chloride, water and 72% sulfuric acid (TAPPI 1969, 1975). All 15N analyses were conducted in the stable isotope laboratory at the Ecosystems Center in Woods Hole, Massachusetts. Samples were analyzed for 15N using a Finnegan MAT delta S isotope ratio mass spectrometer according to the methods of Fry et al. (1992). Analytical errors are generally less than 0.2%o 815N.
N dynamics
Net N immobilization was calculated as an increase in the mass of N remaining in litter and net N mineralization as a decrease. We also calculated immobilization of added nitrate-N for a given year using the initial I5N abundance, the abundance at the end of each year of decomposition, the sizes of the litter N pools, and the known 15N abundance of the labeled nitrate applications, using the following equation:
l= [(N t 1X S t 1) -(Nt0 x~t0) ]/~f
where: /=mass of added NO 3 immobilized by decomposing litter Ntl=mass of N in decomposing litter at the end of the time period Nt0=mass of N in decomposing litter at the beginning of a time pe- riod
The resulting value of N immobilization from added nitrate is far greater than net N immobilization, as it records only the move- ment of N from the enriched pool of added NO 3 into litter which is, at least initially, not enriched. It is not quite the same as gross immobilization, however, because after litter becomes slightly en- riched with tSN, some ~SN is lost along with 14N. Therefore, our measure of N immobilization is best defined as a lower bound on gross N immobilization, but, as N turnover in litter increases, it becomes a poorer estimate of gross immobilization.
Statistical analyses
100
Treatment differences were compared using two-way analysis of variance on log-transformed data for each litter type, with treat- ment and time as factors. When treatment differences were found to be significant, we used Fisher least significant difference tests (SYSTAT 5.01: Systat Inc., Evanston, Ill.) to separate means. Re- gressions on mass loss and percent N were estimated using least squares (SYSTAT 5.01: Systat Inc., Evanston, Ill.). Slopes of re-
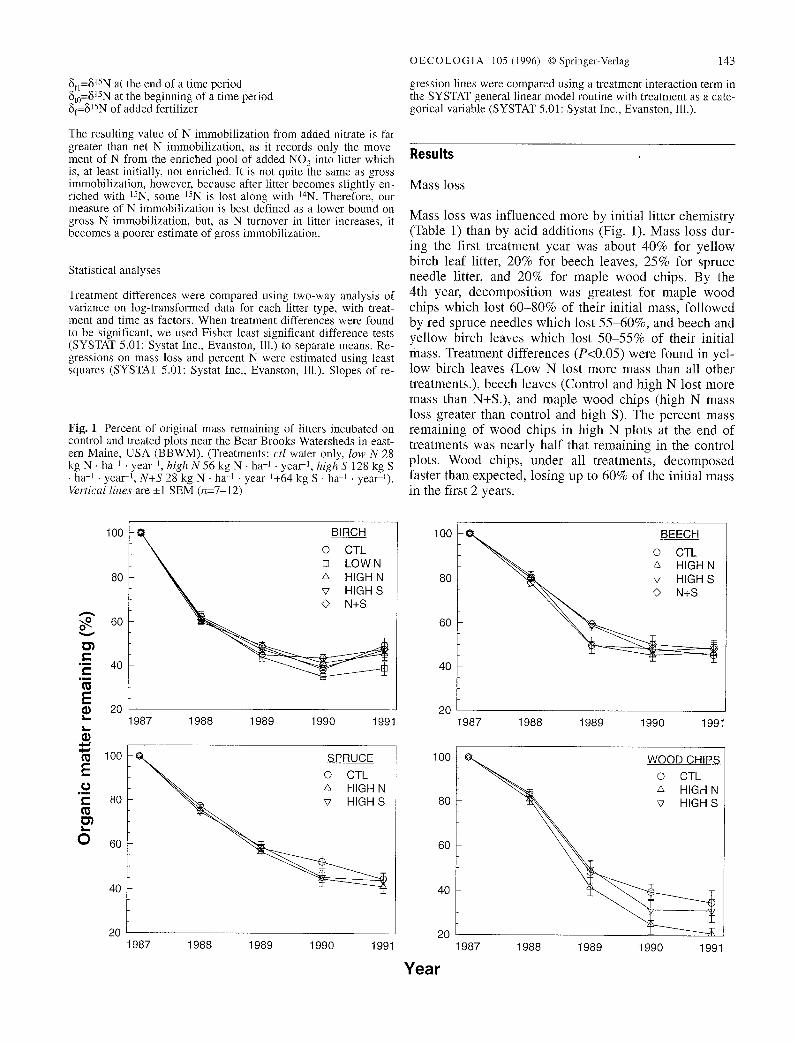
Fig. 1 Percent of original mass remaining of litters incubated on control and treated plots near the Bear Brooks Watersheds in east- ern Maine, USA (BBWM). (Treatments: ctI water only, low N 28 kg N �9 ha J . yeaF 1, high N 56 kg N �9 ha -~ - yeai -1, high S 128 kg S �9 ha -] �9 yea1-1, N+S 28 kg N �9 ha q . yeal-l+64 kg S �9 h ~ 1 �9 year-l). Vertical lines are _+1 SEM (n=7-12)
OECOLOG1A 105 (1996) �9 Springer-Verlag 143
gression lines were compared using a treatment interaction term in the SYSTAT general linear model routine with treatment as a cate- gorical variable (SYSTAT 5.01: Systat Inc., Evanston, Ill.).
Results
Mass loss
Mass loss was in f luenced more by ini t ial l i t ter chemis t ry (Table 1) than by ac id addi t ions (Fig. 1). Mass loss dur- ing the first t rea tment year was about 40% for ye l low bi rch l ea f litter, 20% for beech leaves, 25% for spruce needle litter, and 20% for map le wood chips. By the 4th year, decompos i t i on was greates t for map le wood chips which lost 6 0 - 8 0 % of their ini t ia l mass, fo l lowed by red spruce needles which lost 5 5 - 6 0 % , and beech and ye l low birch leaves which lost 5 0 - 5 5 % of their init ial mass. Trea tment d i f ferences (P<0.05) were found in yel- low birch leaves (Low N lost more mass than all other t reatments . ) , beech leaves (Control and high N lost more mass than N+S.) , and m a p l e wood chips (high N mass loss greater than control and high S). The percen t mass r ema in ing of wood chips in high N plots at the end o f t rea tments was near ly ha l f that r ema in ing in the control plots. W o o d chips, under all t rea tments , d e c o m p o s e d faster than expected , los ing up to 60% of the ini t ial mass in the first 2 years.
100
80
6o
r- �9 -- 40 C , m
E II) 2O s _ _
I1)
100 E O e- 8O
O') l _
O 6o
40
20
e k BIRCH
o CTL [] LOWN A HIGH N
" ~ v HIGH S
1987 1988 1989 1990 1991
% SPRUCE
~ . O CTL - ~ A HIGH N
987 1988 1989 1990 1991
80
60
40
20
100 WOOD CHIPS
O CTL A HIGH N
80
60
40
2O
Y e a r
- % BEECH
O CTL z~ HIGH N v HIGH S
1987 1988 1989 1990 1991
8tl=~lSN at the end of a time period ~t0=~15N at the beginning of a time period 8f-=815N of added fertilizer
1987 1988 1989 1990 1991
144 OECOLOGIA 105 (1996) �9 Springer-Verlag
Table 1 Chemistry of initial litter materials used in Bear Brooks Watershed Manipula- tion (BBWM) external plot de- composition study. [LCI=lig- nin/(lignin+cellulo se) from Aber et al. (1990)]
Initial material
Birch leaves Beech leaves Spruce needles Maple wood
Total nitrogen (g. kg -1) 12.1 7.4 5.9 1,2 Total carbon (g �9 kg -1) 481 469 506 469 C:N ratio 40 63 86 391 Total solubles (%) 41,6 22.8 26.0 6.5 Acid-soluble (cellulose) (%) 3 9.5 53.5 44.2 80.4 Acid-insoluble (lignin) (%) 19,0 23.7 29.9 13.1 LCI 0.33 0.31 0.40 0.14 Lignin:N ratio 16 32 51 109
25
I L -
= m
m
O'1
Z
20
15
10 0 CTL
[] LOW N A HIGH N v HIGH S 0 N+S
e-
1987
25
1988 1989 1990 1991
O = m
20 e'-
U 15 O O
Z 10
SPRUCE o CTL A HIGH N v HIGH S
1987 1988 1989 1990 1991
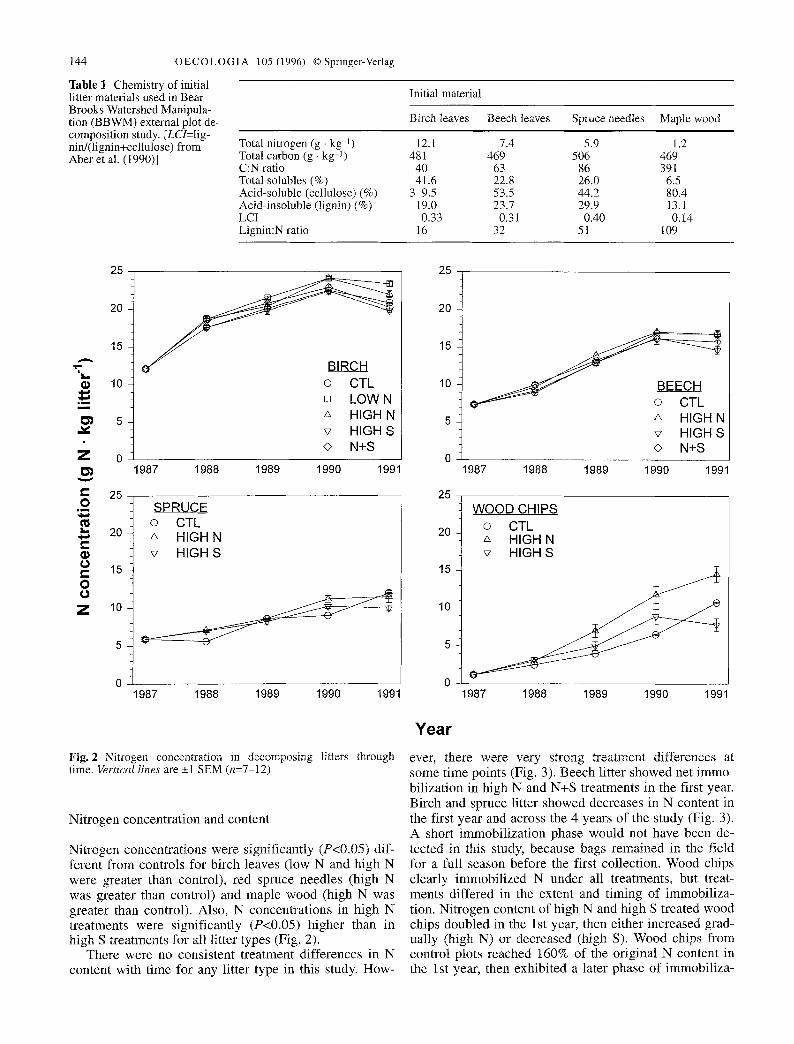
Fig. 2 Nitrogen concentration in decomposing litters through time. Vertical lines are -+1 SEM (n=7-12)
Nitrogen concentration and content
Nitrogen concentrations were significantly (P<0.05) dif- ferent from controls for birch leaves (low N and high N were greater than control), red spruce needles (high N was greater than control) and maple wood (high N was greater than control). Also, N concentrations in high N treatments were significantly (P<0.05) higher than in high S treatments for all litter types (Fig. 2).
There were no consistent treatment differences in N content with time for any litter type in this study. How-
25
20
15
10
z~ HIGH N v HIGH S O N+S
1987
25
1988 1989 1990 1991
20
15
10
WOOD CHIPS
0 CTL HIGH N
v HIGH S
1987 1988 1989 1990 1991
Year
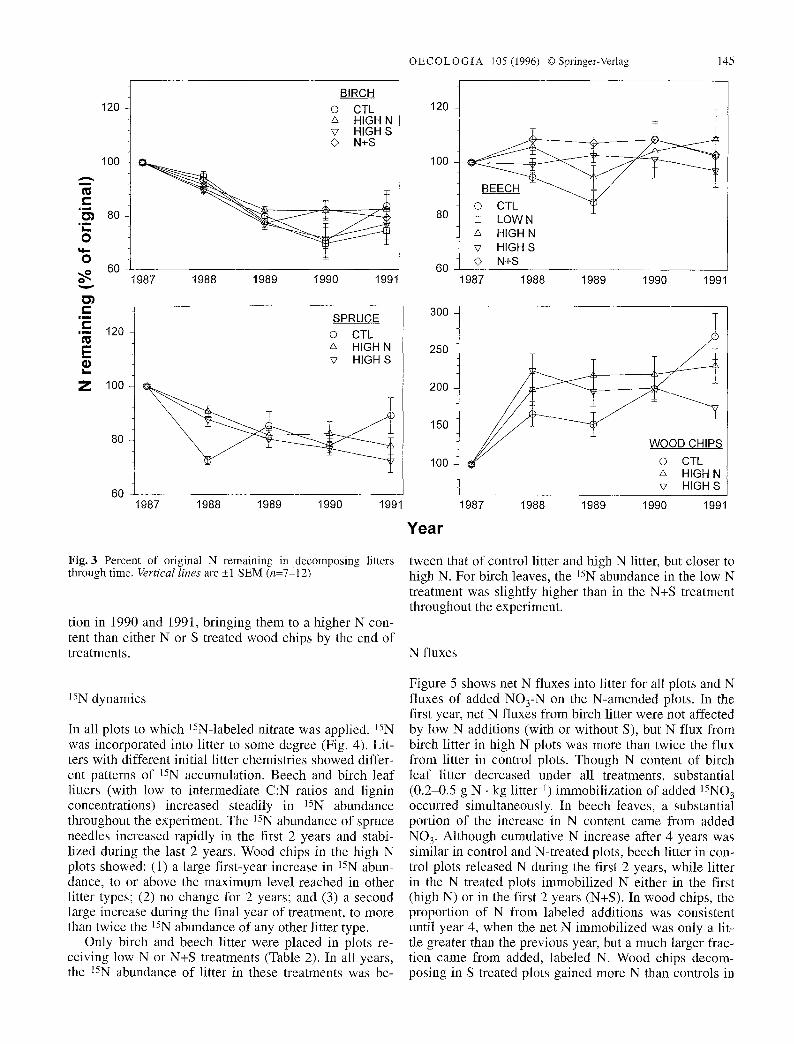
ever, there were very strong treatment differences at some time points (Fig. 3). Beech litter showed net immo- bilization in high N and N+S treatments in the first year. Birch and spruce litter showed decreases in N content in the first year and across the 4 years of the study (Fig. 3). A short immobilization phase would not have been de- tected in this study, because bags remained in the field for a full season before the first collection. Wood chips clearly immobilized N under all treatments, but treat- ments differed in the extent and timing of immobiliza- tion. Nitrogen content of high N and high S treated wood chips doubled in the 1st year, then either increased grad- ually (high N) or decreased (high S). Wood chips from control plots reached 160% of the original N content in the 1 st year, then exhibited a later phase of immobiliza-
OECOLOGIA 105 (1996) �9 Springer-Verlag 145
120
100
80
60
80
BIRCH �9 CTL
HIGH N HIGH S
O N+S
i,- =4
O
O
1987 1988 1989 1990 1991
=m
c I I 120
E tl) L _
Z 100
60
Year
SPRUCE �9 CTL /" HIGH N v HIGH S
1987 1988 1989 1990 199
Fig. 3 Percent of original N remaining in decomposing litters through time. Vertical lines are _+1 SEM (n=7-12)
tion in 1990 and 199l, bringing them to a higher N con- tent than either N or S treated wood chips by the end of treatments.
15N dynamics
In all plots to which 15N-labeled nitrate was applied, 15N was incorporated into litter to some degree (Fig. 4). Lit- ters with different initial litter chemistries showed differ- ent patterns of 15N accumulation. Beech and birch leaf litters (with low to intermediate C:N ratios and lignin concentrations) increased steadily in 15N abundance throughout the experiment. The 15N abundance of spruce needles increased rapidly in the first 2 years and stabi- lized during the last 2 years. Wood chips in the high N plots showed: (1) a large first-year increase in 15N abun- dance, to or above the maximum level reached in other litter types; (2) no change for 2 years; and (3) a second large increase during the final year of treatment, to more than twice the ]SN abundance of any other litter type.
Only birch and beech litter were placed in plots re- ceiving low N or N+S treatments (Table 2). In all years, the 15N abundance of litter in these treatments was be-
120
100
80
60
~O_ CTL [] LOW N /" HIGH N v HIGH S
N+S
1987 1988 1989 1990 1991
3oo t
25O
2O0
150
100 1
t
O CTL /', HIGH N v HIGH S
1987 1988 1989 1990 1991
tween that of control litter and high N litter, but closer to high N. For birch leaves, the 15N abundance in the low N treatment was slightly higher than in the N+S treatment throughout the experiment.
N fluxes
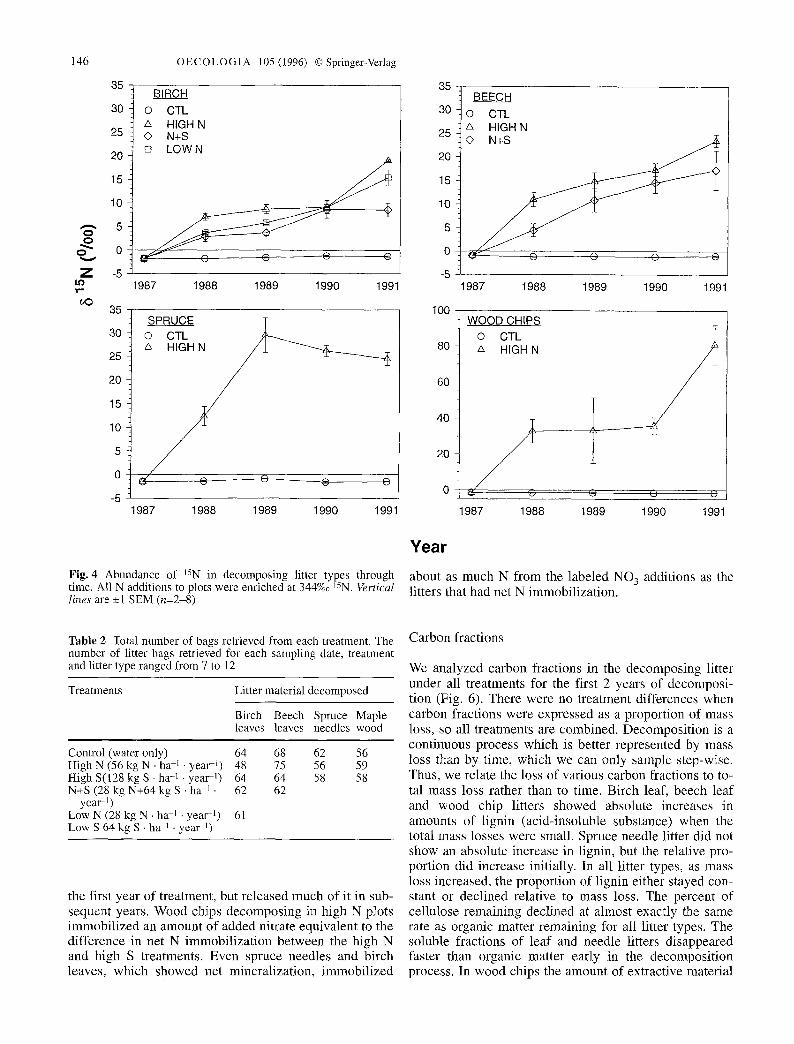
Figure 5 shows net N fluxes into litter for all plots and N fluxes of added NO3-N on the N-amended plots. In the first year, net N fluxes from birch litter were not affected by low N additions (with or without S), but N flux from birch litter in high N plots was more than twice the flux from litter in control plots. Though N content of birch leaf litter decreased under all treatments, substantial (0.2-0.5 g N �9 kg litter -1) immobilization of added 15NO 3 occurred simultaneously. In beech leaves, a substantial portion of the increase in N content came from added NO 3. Although cumulative N increase after 4 years was similar in control and N-treated plots, beech litter in con- trol plots released N during the first 2 years, while litter in the N treated plots immobilized N either in the first (high N) or in the first 2 years (N+S). In wood chips, the proportion of N from labeled additions was consistent until year 4, when the net N immobilized was only a lit- tle greater than the previous year, but a much larger frac- tion came from added, labeled N. Wood chips decom- posing in S treated plots gained more N than controls in
146 OECOLOGIA 105 (i996) �9 Springer-Verlag
35
s Z
30
25
20
15
10
5
0
-5
BIRCH
O CTL HIGH N
O N+S [] LOW N
I . ,O
t.,O 1987
35
1988 1989 1990 1991
30
25
20
15
10
5
0
-5
SPRUCE T o CTL
v
1987 1988 1989 1990 1991
Fig. 4 Abundance of 15N in decomposing litter types through time. All N additions to plots were enriched at 344%0 15N. Vertical lines are -+1 SEM (n=2-8)
35
30
25
20
15
10
5
0
-5
BEECH
O CTL A HIGH N
o o o o
1987
100
1988 1989 1990 1991
80
60
40
20
0
WOOD CHIPS O CTL A HIGH N
1987 1988
�9 �9 u
1989 1990 1991
Year
about as much N from the labeled NO 3 additions as the litters that had net N immobilization.
Table 2 Total number of bags retrieved from each treatment. The number of litter bags retrieved for each sampling date, treatment and litter type ranged from 7 to 12
Treatments Litter material decomposed
Birch Beech Spruce Maple leaves leaves needles wood
Control (water only) 64 68 High N (56 kg N - ha -1 �9 year t) 48 75 High S(128 kg S. ha -1 �9 year -1) 64 64 N+S (28 kg N+64 kg S �9 ha -1 �9 62 62
yeaF 1) Low N (28 kg N. ha -1 . year -1) 61 Low S 64 kg S �9 ha -1 - year -1)
62 56 56 59 58 58
the first year of treatment, but released much of it in sub- sequent years. Wood chips decomposing in high N plots immobilized an amount of added nitrate equivalent to the difference in net N immobilization between the high N and high S treatments. Even spruce needles and birch leaves, which showed net mineralization, immobilized
Carbon fractions
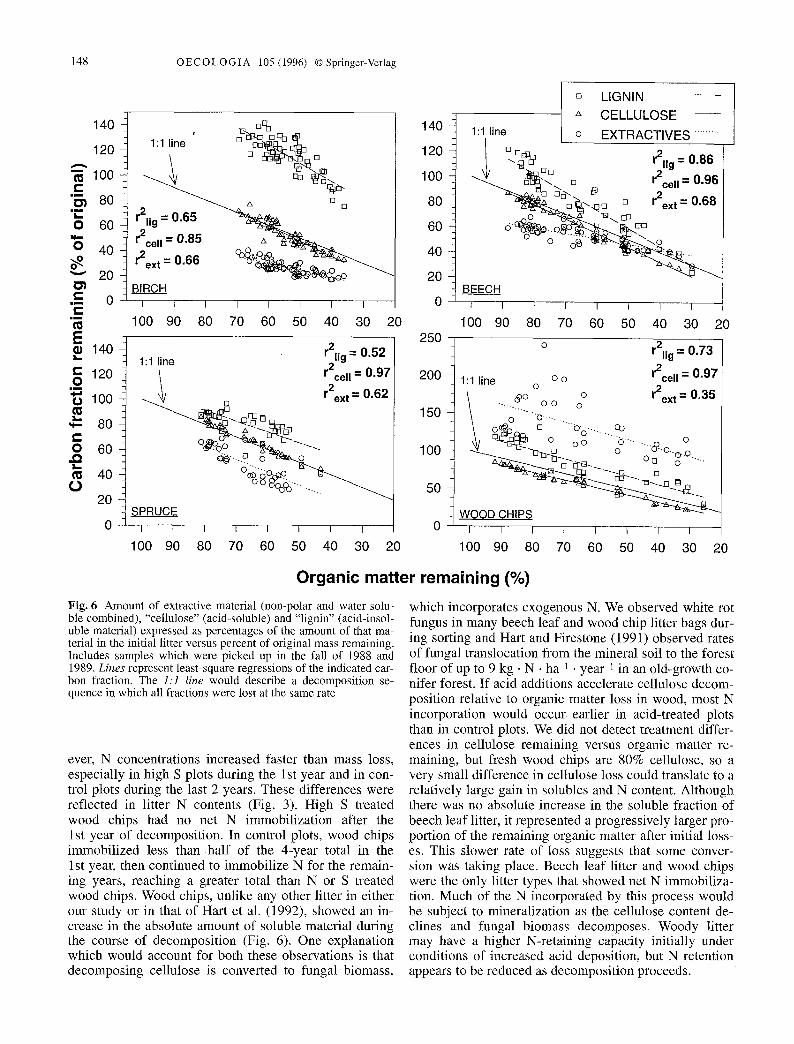
We analyzed carbon fractions in the decomposing litter under all treatments for the first 2 years of decomposi- tion (Fig. 6). There were no treatment differences when carbon fractions were expressed as a proportion of mass loss, so all treatments are combined. Decomposition is a continuous process which is better represented by mass loss than by time, which we can only sample step-wise. Thus, we relate the loss of various carbon fractions to to- tal mass loss rather than to time. Birch leaf, beech leaf and wood chip litters showed absolute increases in amounts of lignin (acid-insoluble substance) when the total mass losses were small. Spruce needle litter did not show an absolute increase in lignin, but the relative pro- portion did increase initially. In all litter types, as mass loss increased, the proportion of lignin either stayed con- stant or declined relative to mass loss. The percent of cellulose remaining declined at almost exactly the same rate as organic matter remaining for all litter types. The soluble fractions of leaf and needle litters disappeared faster than organic matter early in the decomposition process. In wood chips the amount of extractive material
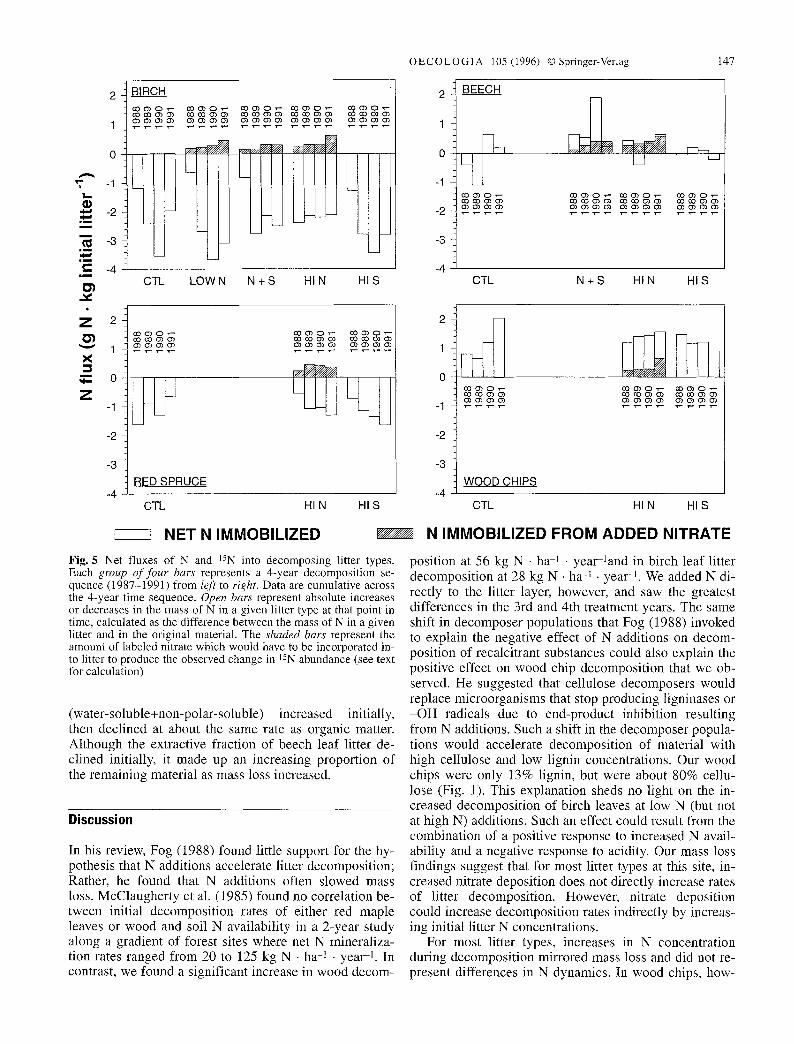
OECOLOGIA 105 (1996) �9 Springer-Verlag 147
' L -
o
m
m
= m
r'-
z
X m
Z
2
1
0
-1
-2
-3
-4
2
1
0
-1
-3
-4
B , . c .
coo) 033)(3) (3303(3) 03{33{3303 030303 03(3)
;TL LOW N N + S HI N HI S
RED SPRUCE
CTL HI N HI S
2
1
0
-1
-2
-3
-4
BEECH
( : 0 ( 3 0 0 " ) 0 ) CO CO O) O) CO QO O ) O ) 1 ~ 0 0 Ob cr~ Ob Ob O ) 1:~ O ) O'~ O~ O ) (3) O3 1 ~ O3 O ) O~ O ) 13~
CTL N + S HI N HI S
2
1
0
-1
-2
-3
-4
QO~O) O~ (~3000)~ C0030) 0) 0303030~ 030~0~0") O'J~O~O3
WOOD CHIPS
CTL HI N HI S
1 ] NET N I M M O B I L I Z E D N I M M O B I L I Z E D F R O M A D D E D N I T R A T E
Fig. 5 Net fluxes of N and 15N into decomposing litter types. Each group o f f our bars represents a 4-year decomposition se- quence (1987-1991) from left to right. Data are cumulative across the 4-year time sequence. Open bars represent absolute increases or decreases in the mass of N in a given litter type at that point in time, calculated as the difference between the mass of N in a given litter and in the original material. The shaded bars represent the amount of Iabeled nitrate which would have to be incorporated in- to litter to produce the observed change in 15N abundance (see text for calculation)
(water-soluble+non-polar-soluble) increased initially, then declined at about the same rate as organic matter. Although the extractive fraction of beech leaf litter de- clined initially, it made up an increasing proportion of the remaining material as mass loss increased.
Discussion
In his review, Fog (1988) found little support for the hy- pothesis that N additions accelerate litter decomposition; Rather, he found that N additions often slowed mass loss. McClaugherty et al. (1985) found no correlation be- tween initial decomposition rates of either red maple leaves or wood and soil N availability in a 2-year study along a gradient of forest sites where net N mineraliza- tion rates ranged from 20 to 125 kg N - ha -1 - year -l. In contrast, we found a significant increase in wood decom-
position at 56 kg N �9 ha 1 . yearqand in birch leaf litter decomposition at 28 kg N �9 ha q �9 year -1. We added N di- rectly to the litter layer, however, and saw the greatest differences in the 3rd and 4th treatment years. The same shift in decomposer populations that Fog (1988) invoked to explain the negative effect of N additions on decom- position of recalcitrant substances could also explain the positive effect on wood chip decomposition that we ob- served. He suggested that cellulose decomposers would replace microorganisms that stop producing ligninases or - O H radicals due to end-product inhibition resulting from N additions. Such a shift in the decomposer popula- tions would accelerate decomposition of material with high cellulose and low lignin concentrations. Our wood chips were only 13% lignin, but were about 80% cellu- lose (Fig. 1). This explanation sheds no light on the in- creased decomposition of birch leaves at low N (but not at high N) additions. Such an effect could result f rom the combination of a positive response to increased N avail- ability and a negative response to acidity. Our mass loss findings suggest that for most litter types at this site, in- creased nitrate deposition does not directly increase rates of litter decomposition. However, nitrate deposition could increase decomposition rates indirectly by increas- ing initial litter N concentrations.
For most litter types, increases in N concentration during decomposition mirrored mass loss and did not re- present differences in N dynamics. In wood chips, how-
148 O E C O L O G I A 105 (1996) �9 Springer-Verlag
140
120
lOO c
80 O 60
O 40
20 03 C 0 C
E 140
120 O ,,i,.,I o 100
80
O 60 .O
40 o
20
1:1 line m~a.qz~ t~ n
) ~ . . . . . . . . j z x D O
_
eex,-- BIRCH
I I I I I I I I
100 90 80 70 60 50 40 30 20
r21ig -- 0.52 1:1 line r2cell
1 ~ . . ~ ~ ~A~ ~ r2ext = = 00.;9;
(~"" (3- .
SPRUCE I I I I I I I I
100 90 80 70 60 50 40 30 20
140 ]
120 ~
,oo 80
60 ]
40 ]
250
200
150
100
50
0
o LIGNIN
" CELLULOSE
1:1 line o EXTRACTIVES .........
I ~ "-. ~ r 2 ,. - 0 86 -~ _ I , ~ - "
D~'~-.. D [] r2cell = 0.96
N ~ . . ~ u ~ o n r " e x t = 0 . 6 8 ^ o ' ~ z ~ . . . z ~ ~ [33
"" " ~ [ 3 ~
�9 . . . . . .
BEECH I I I I I I I I
100 90 80 70 60 50 40 30 20
o r21ig = 0.73
1:1 line o o r2cell = 0.97 O
r2ext = 0.35 / ....... ~176 O ' " " e - - - . . , . .
\ o~ , [] '0-6... oo _~/ q~_~@ o o O ....... b .......... ~. o
"-D~ D_.~.._ 0 0 "O"e"q" "~,~--"-"x'---~s ~"-D--a-~ o o '
WOOD CHIPS I I I t I I I I
100 90 80 70 60 50 40 30 20
Organic matter remaining (%)
Fig. 6 Amount of extractive material (non-polar and water-solu- ble combined), "cellulose" (acid-soluble) and "lignin" (acid-insol- uble material) expressed as percentages of the amount of that ma- terial in the initial litter versus percent of original mass remaining. Includes samples which were picked up in the fall of 1988 and 1989. Lines represent least-square regressions of the indicated car- bon fraction. The I : l line would describe a decomposition se- quence in which all fractions were lost at the same rate
ever, N concentrations increased faster than mass loss, especially in high S plots during the 1 st year and in con- trol plots during the last 2 years. These differences were reflected in litter N contents (Fig. 3). High S treated wood chips had no net N immobilization after the 1st year of decomposition. In control plots, wood chips immobil ized less than h a l f of the 4-year total in the 1 st year, then continued to immobilize N for the remain- ing years, reaching a greater total than N or S treated wood chips. Wood chips, unlike any other litter in either our study or in that of Hart et al. (1992), showed an in- crease in the absolute amount of soluble material during the course of decomposition (Fig. 6). One explanation which would account for both these observations is that decomposing cellulose is converted to fungal biomass,
which incorporates exogenous N. We observed white rot fungus in many beech leaf and wood chip litter bags dur- ing sorting and Hart and Firestone (1991) observed rates of fungal translocation f rom the mineral soil to the forest floor of up to 9 k g . N �9 ha -1 �9 year -1 in an old-growth co- nifer forest. I f acid additions accelerate cellulose decom- position relative to organic matter loss in wood, most N incorporation would occur earlier in acid-treated plots than in control plots. We did not detect treatment differ- ences in cellulose remaining versus organic matter re- maining, but fresh wood chips are 80% cellulose, so a very small difference in cellulose loss could translate to a relatively large gain in solubles and N content. Although there was no absolute increase in the soluble fraction of beech leaf litter, it represented a progressively larger pro- portion of the remaining organic matter after initial loss- es. This slower rate of loss suggests that some conver- sion was taking place. Beech leaf litter and wood chips were the only litter types that showed net N immobiliza- tion. Much of the N incorporated by this process would be subject to mineralization as the cellulose content de- clines and fungal biomass decomposes. Woody litter may have a higher N-retaining capacity initially under conditions of increased acid deposition, but N retention appears to be reduced as decomposition proceeds.
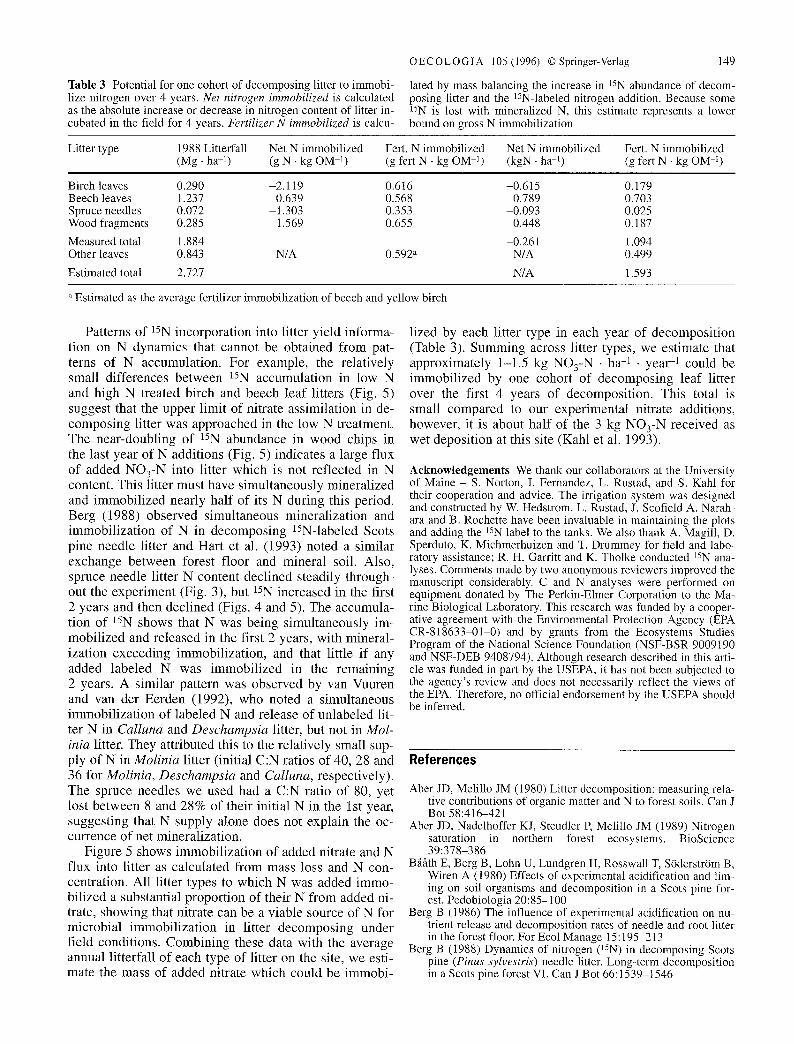
Table 3 Potential for one cohort of decomposing litter to immobi- lize nitrogen over 4 years. Net nitrogen immobilized is calculated as the absolute increase or decrease in nitrogen content of litter in- cubated in the field for 4 years. Fertilizer N immobilized is calcu-
OECOLOGIA 105 (1996) �9 Springer-Verlag 149
lated by mass balancing the increase in 15N abundance of decom- posing litter and the 15N-labeled nitrogen addition. Because some 15N is lost with mineralized N, this estimate represents a lower bound on gross N immobilization
Litter type 1988 Litterfall Net N immobilized Fert. N immobilized Net N immobilized Fert. N immobilized (Mg �9 ha -i) (g N �9 kg OM 1) (g fert N - kg OM 1) (kgN �9 ha 1) (g fert N - kg OM -l)
Birch leaves 0.290 -2.119 0.616 -0.615 0.179 Beech leaves 1.237 0.639 0.568 0.789 0.703 Spruce needles 0.072 -1.303 0.353 -0.093 0.025 Wood fragments 0.285 1.569 0.655 0.448 0.187
Measured total 1.884 -0.261 1.094 Other leaves 0.843 N/A 0.592 a N/A 0.499
Estimated total 2.727 N/A 1.593
a Estimated as the average fertilizer immobilization of beech and yellow birch
Patterns o f 15N incorporat ion into litter yield informa- tion on N dynamics that cannot be obtained f rom pat- terns of N accumulation. For example, the relatively small differences between 15N accumulat ion in low N and high N treated birch and beech leaf litters (Fig. 5) suggest that the upper limit o f nitrate assimilation in de- compos ing litter was approached in the low N treatment. The near-doubling o f 15N abundance in wood chips in the last year o f N additions (Fig. 5) indicates a large flux o f added NO3-N into litter which is not reflected in N content. This litter must have simultaneously mineral ized and immobi l ized nearly half o f its N during this period. Berg (1988) observed simultaneous mineralization and immobil izat ion o f N in decompos ing 15N-labeled Scots pine needle litter and Hart et al. (1993) noted a similar exchange between forest f loor and mineral soil. Also, spruce needle litter N content declined steadily through- out the experiment (Fig. 3), but 15N increased in the first 2 years and then declined (Figs. 4 and 5). The accumula- tion o f 15N shows that N was being s imultaneously im- mobi l ized and released in the first 2 years, with mineral- ization exceeding immobil izat ion, and that little if any added labeled N was immobi l ized in the remaining 2 years. A similar pattern was observed by van Vuuren and van der Eerden (1992), who noted a s imultaneous immobil izat ion o f labeled N and release o f unlabeled lit- ter N in CalIuna and Deschampsia litter, but not in Mol- inia litter. They attributed this to the relatively small sup- ply o f N in Molinia litter (initial C :N ratios o f 40, 28 and 36 for Molinia, Deschampsia and Calluna, respectively). The spruce needles we used had a C:N ratio o f 80, yet lost between 8 and 28% of their initial N in the 1st year, suggest ing that N supply alone does not explain the oc- currence o f net mineralization.
Figure 5 shows immobil izat ion o f added nitrate and N flux into litter as calculated f rom mass loss and N con- centration. All litter types to which N was added immo- bilized a substantial proport ion of their N f rom added ni- trate, showing that nitrate can be a viable source o f N for microbial immobi l izat ion in litter decompos ing under field conditions. Combin ing these data with the average annual litterfall o f each type of litter on the site, we esti- mate the mass o f added nitrate which could be immobi-
lized by each litter type in each year of decomposi t ion (Table 3). Summing across litter types, we estimate that approximately 1-1.5 kg NO3-N �9 ha -1 - year -1 could be immobi l ized by one cohort o f decompos ing leaf litter over the first 4 years of decomposi t ion. This total is small compared to our experimental nitrate additions, however, it is about half of the 3 kg NO3-N received as wet deposit ion at this site (Kahl et al. 1993).
Acknowledgements We thank our collaborators at the University of Maine - S. Norton, I. Fernandez, L. Rustad, and S. Kahl for their cooperation and advice. The irrigation system was designed and constructed by W. Hedstrom. L. Rustad, J. Scofield A. Narah- ara and B. Rochette have been invaluable in maintaining the plots and adding the 15N label to the tanks. We also thank A. Magill, D. Sperduto, K. Michmerhuizen and T. Drummey for field and labo- ratory assistance; R. H. Garritt and K. Tholke conducted 15N ana- lyses. Comments made by two anonymous reviewers improved the manuscript considerably. C and N analyses were performed on equipment donated by The Perkin-Elmer Corporation to the Ma- rine Biological Laboratory. This research was funded by a cooper- ative agreement with the Environmental Protection Agency (EPA CR-818633-01-0) and by grants from the Ecosystems Studies Program of the National Science Foundation (NSF-BSR 9009190 and NSF-DEB 9408794). Although research described in this arti- cle was funded in part by the USEPA, it has not been subjected to the agency's review and does not necessarily reflect the views of the EPA. Therefore, no official endorsement by the USEPA should be inferred.
References
Aber JD, Melillo JM (1980) Litter decomposition: measuring rela- tive contributions of organic matter and N to forest soils. Can J Bot 58:416-421
Aber JD, Nadelhoffer KJ, Steudler P, Melillo JM (1989) Nitrogen saturation in northern forest ecosystems. BioScience 39:378-386
Bfifith E, Berg B, Lohn U, Lundgren H, Rosswall T, S6derstr6m B, Wiren A (1980) Effects of experimental acidification and lim- ing on soil organisms and decomposition in a Scots pine for- est. Pedobiologia 20:85-100
Berg B (1986) The influence of experimental acidification on nu- trient release and decomposition rates of needle and root litter in the forest floor. For Ecol Manage 15:195-213
Berg B (1988) Dynamics of nitrogen (15N) in decomposing Scots pine (Pinus sylvestris) needle litter. Long-term decomposition in a Scots pine forest VI. Can J Bot 66:1539-1546

150 OECOLOGIA 105 (1996) �9 Springer-Verlag
Fenn ME, Dunn PH (1989) Litter decomposition across an air-pol- lution gradient in the San Bernardino mountains. Soil Sci Soc Am J 53:1560-1567
Fog K (1988) The effect of added nitrogen on the rate of decom- position of organic matter. Biol Rev 63:433-462
Fry BW, Brand W, Mersch FJ, Tholke K, Garritt R (1992) Auto- mated analysis system for coupled ~I3C and 615N measure- ments. Anal Chem 64:288-291
Hfigvar S, Kjondal BR (1981) Decomposition of birch leaves: dry weight loss, chemical changes and effects of artificial rain. Pedobiologia 22:232-245
Hart SC, Firestone MK (1991) Forest floor-mineral soil interac- tions in the internal nitrogen cycle of an old-growth forest. Biogeochemistry 12:103-127
Hart SC, Firestone MK, Paul EA (1992) Decomposition and nutri- ent dynamics of ponderosa pine needles in a Mediterranean- type climate. Can J For Res 22:306-314
Hart SC, Firestone MK, Paul EA, Smith JL (1993) Flow and fate of soil nitrogen in an annual grassland and a young mixed-co- nifer forest. Soil Biol Biochem 25:431-442
Kahl JS, Norton SA, Fernandez IJ, Nadelhoffer KJ, Driscoll CT, Aber JD (1993) Experimental inducement of nitrogen satura- tion at the watershed scale. Environ Sci Technol 27:565-568
McClaugherty CA, Pastor JA, Aber JD, Melillo JM (1985) Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 66:266-275
Melillo J, Aber J, Muratore J (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 63:621-626
Nadelhoffer KJ, Downs MR, Fry B, Aber JD, Magill AH, Melillo JM (1995) The fate of 15N-labeled nitrate additions to a north- ern hardwood forest in eastern Maine, USA. Oecologia, in press
Nihlgard B (1985) The ammonium hypothesis - an additional ex- planation to the forest dieback in Europe. Ambio 14:2-8
Neuvonen S, Suomela J (1990) The effect of simulated acid rain on pine needle and birch leaf litter decomposition. J Appl Ecol 27:857-872
Norton SA, Kahl JS, Fernandez IJ, Schofield JR Rustad LE, Haines TA, Lee J (1993) The Watershed Manipulation Project: two-year results at the Bear Brook Watershed in Maine (BBWM). In: Rasmussen L, Brydges T, Mathy P (eds) Experi- mental manipulations of biota and biogeochemical cycling in ecosystems: approach, methodologies, fndings (CEC ecosys- tems research report no. 4). ECSC-EEC-EAEC, Brussels, pp 55-63
Norton SAI Kahl JS, Fernandez I, Rustad LE, Scofield JR Haines TA (1994) Response of the West Bear Watershed, Maine, USA to the addition of (NH4)2SO 4 - three year results. For Ecol Manage 68:61-73
Prescott CE, Parkinson D (1985) Effects of sulphur pollution on rates of litter decomposition in a pine forest. Can J Bot 63:1436-1443
Rustad LE, Fernandez IJ, Fuller RD, David MB, Nodvin SC, Halt- eman WA (1993) Soil solution response to acidic deposition in a northern hardwood forest. Agric Ecosys Environ 47:117-134
Rustad LE, Fernandez IJ, David MB, Mitchell M J, Nadelhoffer KJ, Fuller RB (in prep) Experimental soil acidification and re- covery at the Bear Brook Watershed in Maine (BBWM). Soil Sci Soc Am J
Ryan MG, Melillo JM, Ricca A (1990) A comparison of methods for determining proximate carbon fractions of forest litter. Can J For Res 20:166-171
Schulze ED (1989) Air pollution and forest decline in a spruce (Picea abies) forest. Science 244:776-783
TAPPI (1969) Organic solvent extractives in pulp (T 204 os-69). Technical Association of the Pulp and Paper Industry, Atlanta
TAPPI (1975) Water solubles in wood and pulp (T 207 os-75). Technical Association of the Pulp and Paper Industry, Atlanta
Tietema A (1993) Mass loss and nitrogen dynamics in decompos- ing acid forest litter in the Netherlands at increased nitrogen deposition. Biogeochemistry 20:45-62
Vuuren MMI van, Eerden LJ van der (1992) Effects of three rates of atmospheric nitrogen deposition enriched with 15N on litter decomposition in a heathland. Soil Biol Biochem 24:527-532





























