Embed Size (px)
Citation preview

Pyrolysis of Organomercury Compounds 3241
United States Office of Naval Research. A. K. M. was a guestscientist at Rice University through the Exchange Programsponsored by the U.S.S.R. and U.S.A. Academics of Science.
1. Becker, E. D., and G. C. Pirnentel, J. Chem. Phys., 25, 224(1956).
2. Hastie, J. W., R. H. Hauge, and J. L. Margrave, in Spec-troscopy in Inorganic Chemistry, ed. C. N. R. Rao and J. R.Ferraro (Academic Press, New York, N.Y., 1970), pp.57-106.
3. Maltsev, A. K., R. G. 'Mikaeliari, and 0. M. Nefedov, Bull.Acad. Sci., USSR (Div. Chem. Sci., 1971), in press.
4. Seyferth, D., J. M. Burlitch, R. J. Minasz, J. Y. Mui, H.
D. Simmons, Jr., A. J. H. Treiber, and S. R. Dowd, J. Amer.Chem. Soc., 87, 4259 (1965).
.5. Milligan, D. E., and M. E. Jacox, J. Chem. Phys., 47, 703(1967).
6. Andrews, L., J. Chem. Phys., 48, 972 (1968).7. Andrews, L., J. Chem. Phys., 48, 979 (1968).8. Rogers, E. E., S. Abramowitz, M. E. Jacox, and D. E.
Milligan, J. Chem. Phys., 52, 2198 (1970).9. Logan, T. J., Org. Synth., 46, 98 (1966).
10. Maltsev, A. K., R. G. Mikaelian, and 0. M. Nefedov, Bull.Acad. Sci., USSR (Div. Chem. Sci., 1971), in press.
11. King, S. T., J. Chem. Phys., 49, 1321 (1968).12. Greeti, J. H. S., Spectrochim. Acta, Part A, 24, 863 (1968).13. Mathews, C. W., Can. J. Phys., 45, 23.55 (1967).
Addendum. The authors of the article entitled "CollagenBiosynthesis: Synthesis and Secretion of a High MolecularWeight Collagen Precursor (Procollagen)," R. L. Church,S. E. Pfeiffer, and M. L. Tanzer, which appeared in theNovember 1971 issue of Proc. Nat. Acad. Sci. USA, 68,2638-2642, wish to amplify the term "procollagen" asfollows.
Since several investigators have identified procollagenmolecules of various sizes, a consistent nomenclature isdesirable. By analogy with the conversion of chymotryp-sinogen into chymotrypsin, we suggest that the pro-
collagen intermediates be given a secondary designation,such as Procollagen I, Procollagen II, etc. (We havepurposely avoided Greek symbols because they alreadysignify tropocollagen subunits.) Furthermore, we proposethat the secondary numbering correspond to the size of theprocollagen molecule. Accordingly, Procollagen I woulddesignate the largest molecule, which is described in ourreport, while ProcoUagen II, III, etc., would be assignedto the smaller intermediates identified by others. Thisproposed nomenclature should minimize confusion andwould immediately denote the relative size of the pro-collagen molecule.
Proc. Nat. Acad. Sci. USA 68 (1971)

Proc. Nat. Acad. Sci. USAVol. 68, No. 11, pp. 2638-2642, November 1971
Collagen Biosynthesis: Synthesis and Secretion of a High MolecularWeight Collagen Precursor (Procollagen)
(fibroblasts/isotopic tracers/acrylamide gel electrophoresis/gel filtration)
R. L. CHURCH, S. E. PFEIFFER, AND M. L. TANZER
Departments of Biochemistry and Microbiology, University of Connecticut Health Center, Farmington, Conn. 06032
Communicated by Max Tishler, August 9, 1971
ABSTRACT Sodium dodecyl sulfate-acrylamide gelelectrophoresis and molecular-sieve chromatography on8% agarose demonstrate the existence of a very highmolecular weight (500,000-600,000), proline-rich protein incultured 3T6 fibroblasts that appears to be the precursormolecule (procollagen) of collagen. The kinetics of [3H]-proline uptake indicate that this precursor is synthesizedat a different rate than are other cell proteins and issecreted apparently unchanged into the medium, whereit undergoes modification; it then precipitates aroundthe cells as collagen fibrils that contain the characteristictropocollagen polypeptide chains. The solubility of thisprecursor in hot 5% CI3CCOOH, its hydroxyproline toproline ratio, and its sensitivity to highly-purified bac-terial collagenase all indicate that this molecule is ofcollagenous nature, but that it has considerable regionsof noncollagen peptide (about half of the molecule iscollagenase sensitive, and it has halfof the normal amountof hydroxylated proline residues).These results support the concept of a procollagen mole-
cule, of molecular weight about 500,000-60,000, that con-tains large regions of noncollagen peptides, which mightallow the collagen a-chain regions to associate in registerand that also can provide the correct chain compositionfor tropocollagen. Upon secretion of the procollagenmolecule, the intercollagen-peptide regions are cleavedand the finished tropocollagen molecule then polymerizesinto typical intercellular fibrils.
Functional proteins are synthesized in animal cells by twogeneral pathways, either by direct formation of the finalproduct or by formation of an inactive precursor, which isselectively altered by proteolysis, to yield the active product.In certain cases, the proteolysis mechanism provides multi-chain molecules from single-polypeptide-chain precursors.Examples of this process include the formation of pancreaticenzymes (1), insulin (2), and the capsid proteins ofpoliovirus (3). An analogous pathway occurs in the case ofnucleic acids, most notably that of ribosomal-RNA formation(4).Two of the major problems in the formation of the helical,
three-stranded tropocollagen molecule are the provision forcorrect chain composition, namely two al and one a2 chains(5), and the necessity for helical folding of the chains in theproper register and with correct polarity (6). A unique solu-tion for these requirements would be provided by the synthesisof a single polypeptide chain that folded appropriately andthen underwent suitable proteolytic modification. We
present, in this report, data that support such a model, andindicate that the proteolysis step occurs extracellularly.
MATERIALS AND METHODS
Confluent 3T6 fibroblasts were maintained in Dulbecco-Vogt-modified Eagle's medium supplemented with 10% calfserum and 50 ,sg/ml of sodium ascorbate, at 370C, under10% C02-90% air. Cultures were labeled with [3,4-'H]i,proline (5.26 Ci/mmol; 10 ,Ci/ml), [2-8Hlglycine (6.8Ci/mmol; 10 IMCi/ml), or [U-14C]tyrosine (0.4lCi/mmol;10 MCi/ml) in 1.0 or 10 ml of medium, depending on the sizeof the culture dish.
After exposure to label, the medium was removed, the cellswere washed once with phosphate-buffered saline (pH 7.X),and 0.5-2.0 ml of 1% Nonidet P-40 detergent (Shell ChemicalCo.) in isotonic buffer, phosphate-saline, pH 7.6 (7) wasadded. The cells were allowed to lyse for 5 min in an ice bath,after which the lysate was centrifuged at 2000 X g for10 min. Three fractions were obtained: medium (centrifuged at5000 X g to remove dead cells and debris), cell supernatantor "cell sap" (2000 X g supernatant), and cell pellet (2000 Xg pellet).Sodium Dodecyl Sulfate (SDS)-Acrylamide Gel Electro-
phoreis. Samples from each fraction were denatured withSDS (1%), reduced with 2-mercaptoethanol (1%) or dithio-threitol (1%), heated at 100'C for 1 min (8), and electro-phoresed in 3% polyacrylamide-0.2% ethylene diacrylategels that contained 0.3% SDS. Radioactivity was measuredin 1.5-mm gel slices dissolved in 1.0 N NH4OH and countedin 10 ml of Triton-X100-toluene scintillation fluid. Ap-proximate molecular weights were determined by comparisonof the mobilities of cellular proteins with purified rat, guineapig, and calf-skin al, Ij, and y collagen chains (9).
Molecular-Sieve Chromatography. Culture medium andNP-40-supernatant fluids, made 0.1 N with EDTA to in-hibit collagenase, were chromatographed on 100 X 1.5 cmcolumns of Bio-Gel A-1.5 m equilibrated with 0.05 M Tris*HCl (pH 7.6)-0.01 M EDTA and eluted with the same bufferat 40C. 2-ml fractions were collected and their radioactivitywas measured in 0.1-ml aliquots in 10 ml of toluene-BeckmanBiosolv-3 scintillation fluid. Gel filtration on Bio-Gel A-15mwas done the same way.
Proline and hydroxyproline were separated and assayedby conventional methods (10). Bacterial collagenase (Worth-ington) was purified and used by the method of Peterkofskyand Diegelmann (11).
2638
Abbreviations: SDS, sodium dodecyl sulfate; HMW, high mo-lecular weight (protein); LMW, low molecular weight (protein).
Procollagen Synthesis and Secretion 2639
RESULTS
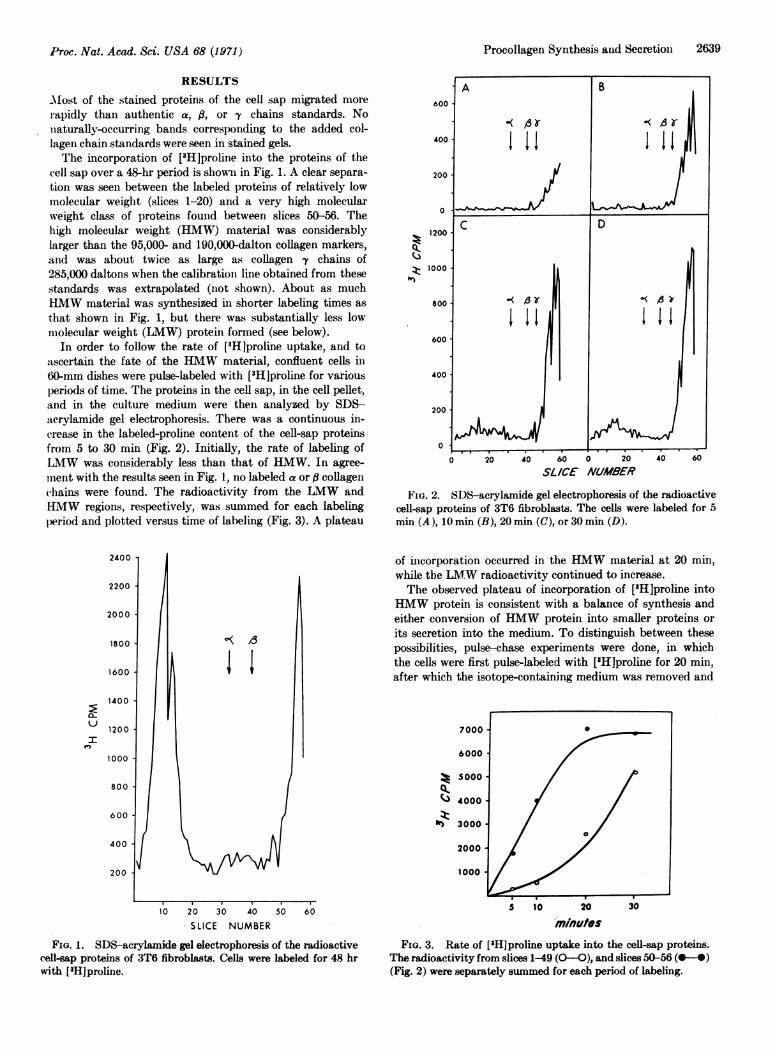
Most of the stained proteins of the cell sap migrated morerapidly than authentic a, ft, or fy chains standards. Noiiaturally-occurring bands corresponding to the added col-lagen chain standards were seen in stained gels.The incorporation of [3H]proline into the proteins of the
cell sap over a 48-hr period is shown in Fig. 1. A clear separa-tion was seen between the labeled proteins of relatively lowmolecular weight (slices 1-20) and a very high molecularweight class of proteins found between slices 50-56. Thehigh molecular weight (HMW) material was considerablylarger than the 95,000- and 190,000-dalton collagen markers,and was about twice as large as collagen y chains of285,000 daltons when the calibration line obtained from thesestandards was extrapolated (not shown). About as muchHMW material was synthesized in shorter labeling times asthat shown in Fig. 1, but there was substantially less lowmolecular weight (LMW) protein formed (see below).
In order to follow the rate of [3H]proline uptake, and toascertain the fate of the HMW material, confluent cells in60-mm dishes were pulse-labeled with [3H]proline for variousperiods of time. The proteins in the cell sap, in the cell pellet,and in the culture medium were then analyzed by SDS-acrylamide gel electrophoresis. There was a continuous in-crease in the labeled-proline content of the cell-sap proteinsfrom 5 to 30 min (Fig. 2). Initially, the rate of labeling ofLMW was considerably less than that of HMW. In agree-mnent with the results seen in Fig. 1, no labeled a or ft collagenchains were found. The radioactivity from the LMW andHMW regions, respectively, was summed for each labelingperiod and plotted versus time of labeling (Fig. 3). A plateau
2400
2200
2000
1800
1600
I
1400
1200
1000
800
600
400
200
600
400 O~~~~~~~~~ 11200
0
C D1200-
1000
gool 1 11 I 11600
400
200
0 20 40 60 0 20 40 60
SLICE' NUMBER
FIG. 2. SDS-acrylamide gel electrophoresis of the radioactivecell-sap proteins of 3T6 fibroblasts. The cells were labeled for 5min (A), 10 min (B), 20 min (C), or 30 min (D).
of incorporation occurred in the HMW material at 20 min,while the LMW radioactivity continued to increase.The observed plateau of incorporation of [8H]proline into
HMW protein is consistent with a balance of synthesis andeither conversion of HMW protein into smaller proteins orits secretion into the medium. To distinguish between thesepossibilities, pulse-chase experiments were done, in whichthe cells were first pulse-labeled with [3H]proline for 20 min,after which the isotope-containing medium was removed and
10 20 30 40 50
SLICE NUMBER60
FIG. 1. SDS-acrylamide gel electrophoresis of the radioactivecell-sap proteins of 3T6 fibroblasts. Cells were labeled for 48 hrwith [mHjproline.
5 10 20 30
m/nluesFiG. 3. Rate of [3H]proline uptake into the cell-sap proteins.
The radioactivity from slices 1-49 (O0O), and slices 50-56 (0-4)(Fig. 2) were separately summed for each period of labeling.
Proc. Nat. Acad. Sci. USA 68 (1971)

2640 Biochemistry: Church et al.
400
x 300'
% 200
k 100
0 10 611101020 30 40 50 600 30 40 50 60
SLICE NUMBER
FIG. 4. SDS-acrylamide gel electrophoresis of the radio-active cell-sap proteins (left) and culture-medium proteins(right) from a pulse-chase experiment: 20 min pulse, and 20 minchase.
fresh medium that contained an excess of nonradioactiveproline was added; the incubation was then continued for anadditional 20-40 min. The distribution of radioactivity inthe proteins of the medium and in the cell sap after 20 minof chase time is shown in Fig. 4. Virtually all of the radio-activity in the medium was in HMW protein, which ac-cumulated in the medium as it disappeared from the cells(Fig. 5). These results suggest that at least some of the HMIWprotein was secreted without reduction of its size (see Dis-cussion). Although labeled HMW protein accumulated inthe medium, no radioactive a or , chains were detected inthe acrylamide gels, as would have been expected if labeledtropocollagen molecules had been secreted by the cells. How-ever, upon prolongation of the pulse-chase experiment for1-2 days, very small amouts of a, #, and y components ap-peared in the medium.
Similar studies with ['H glycine substituted for ['H]prolinegave the same results as in Figs. 1-4. In contrast, when['4C]tyrosine or [14C]leucine was used (only small amountsof these amino acids occur in collagen), only slight incorpora-tion into the HMW proteins of the cells and culture mediumwas found, although the incorporation into LMW cell proteinwas unchanged.
In order to further study the nature of the ['H proline-labeled HMW protein, the medium from several experimentswas pooled, lyophylized, and subjected to molecular-sievechromatography on 8% agarose (Fig. 6). ['H]Hydroxyprolinewas present both in the major peak (which eluted with thevoid volume of the column) and in the minor peak (whicheluted at 194-208 ml), where the majority of the serum pro-teins from the culture medium were found; the free ['H ]prolinein the medium eluted after 300 ml). More than half of thehydroxyproline, which eluted from 112-208 ml, was associatedwith the major peak. The ratio of ['H]hydroxyproline to['H]proline in the major peak was 0.42, which is about halfof the ratio of these imino acids in purified collagens (12)or in hydrolyzates of 3T6 cells (13). The material in the majorpeak was pooled, concentrated to near dryness by lyophyliza-tion, and incubated with N-ethylmaleimide and purifiedcollagenase; the reaction mixture was then adjusted to 5%Cl3CCOOH-0.5% tannic acid to precipitate both collagenousand noncollagenous proteins (11). The supernatant, contain-ing collagen-derived peptides, was assayed for radioactivity.41% of the HMW peak was digested in this assay. More than85% of the original HMW was soluble in hot 5% Cl3CCOOHin the absence of collagenase, as would be expected for acollagen-related protein (14). Moreover, if hot 5% Cl3CCQOOH-0.5% tannic acid was substituted for hot 5% C13CCOOH,
Proc. Nat. Acad. Sci. USA 68 (1971)
over 95% of the [3H]proline in the HMW peak was pre-cipitated.Treatment of the cells with ,3-aminopropionitrile fumarate
(50 Ag/ml), which inhibits collagen crosslink formation, orwith a,a-dipyridyl (1 mM), which inhibits hydroxylationof proline and lysine residues in collagen, had little effectupon proline uptake into the HMW protein in the cells.Similarly, exposure to puromycin (10 /Ag/ml) for 3-10 min,after 10 min of ['H]proline labeling, did not alter the sizeof the intracellular HMW protein. Incubation of the cellswith phenylmethylsulfonyl fluoride (50 ,ug/ml), a proteaseinhibitor, during or after ['H proline incorporation, also hadno effect on the size of the intracellular HMW protein. Inorder to evaluate the occurrence of nonspecific aggregation,samples of the cell sap and culture medium were treated withhigher concentrations of SDS (56o') and 2-mercaptoethanol(5%), or with dithiothreitol in place of the mercaptoethanol.The electrophoretic pattern was not changed by these treat-ments.
Since the 3T6 fibroblasts form typical extracellular fibrilsthat contain tropocollagen molecules (15), we examined thecell pellet (which contained cell nuclei, cell debris, and theextracellular collagen fibrils) and the culture medium for theircontent of the polypeptide chain components of tropocollagen(Fig. 7). The medium contained mostly HMW proteinand no collagen a or # chains, while the pellet contained pro-tein that migrated with collagen a and # chains (and smallerproteins) but little HMW protein. The small amount ofradioactive material migrating at higher molecular weightthan a or f3 chains may be more-highly crosslinked polymersof collagen chains (which were observed when fibrils of calf-skin collagen were dissolved in SDS and electrophoresed onSDS gels). Electron microscopy confirmed that 3T6 cellsmake collagen fibrils under these experimental conditions;radiochemical analyses showed that the labeled medium, cellsupernatant, and cell pellet all contained ['Hihydroxyproline.
DISCUSSIONThe experimental results indicate the presence in 3T6 cellcultures of a polypeptide (procollagen) much larger thantropocollagen, but nevertheless containing many of the ratherspecific properties of collagen molecules. The amount of
F I I. .
0 10 20 30 40 50 60
rminutesFIG. 5. Rate of disappearance of intracellular procollagen,
and its appearance in the medium. The procollagen radioactivitywas determined by measuring the area under the procollagenpeak in gel patterns such as those:shown in Fig. 4. These valueswere then plotted as a function of the length of chase; 0-O, intra-cellular; 0-0, culture medium.

Procollagen Synthesis and Secretion 2641
hydroxyproline in the polypeptide and its sensitivity topurified collagenase indicate that only 50-60% of the moleculehas amino-acid sequences characteristic of collagen. Thefinding that no a and # chains are present in the cells ormedium indicates that collagen is synthesized and secretedinto the medium as procollagen, and not as tropocollagen orfree a and ,B chains. Moreover, the hydroxyproline in themedium seems to be present mainly in procollagen anddialyzable hydroxyproline. The dialyzable hydroxyprolineaccounts for about 30% of the total ['Hihydroxyproline inthe medium. This result indicates that a large amount ofcollagen-like material is degraded in the cultures, perhapsby enzymes in the serum added to the medium, or by col-lagenases synthesized by the cells. Indeed, we find that if themedium that contains procollagen is subjected to gel filtra-tion at room temperature with calcium in place of EDTAin the elution buffer, most of the HMW peak is degraded tosmall molecules and free amino acids. As shown in Fig. 6,even gel filtration in the presence of EDTA at 4VC causessome breakdown. We do not know if this represents specificcleavage. Recent findings in other laboratories show that alarge, collagen-like protein is present in the medium of humanfibroblasts (16); pro a-1 chains are converted into typicalcollagen a-1 chains (17). These pro-al chains, of molecularweight 120,000-140,000 (17), may represent an intermediate
24
22
20
18
0
x
16
14
12
10
8
6
40 80 120 160 200 240EFFLUENT VOLUME,
280 320
ML360
FIG. 6. Gel filtration of the radioactive proteins of the culturemedium. Fractions were pooled as indicated by the bars, hy-drolyzed, and assayed for [8H]hydroxyproline. The numbersabove the bars are the cpm of [3H]hydroxyproline in the pooledfractions. The column void-volume is 130 ml. Similar results wereobtained when the column was eluted at room temperature withthe same buffer containing SDS (0.2%) and dithiothreitol (5mM). Moreover, the pooled material at the void volume was
found to migrate, on SDS-acrylamide gel electrophoresis, in thesame location as HMW protein (Fig. 4).
0.~~~~~~~~~~~~~04 2
3-
2-
1020 30 40 50 60 20 30 60
SLICE NUMBER SLICE NUDBER
FIG. 7. SDS-acrylamide gel electrophoresis of the radioactiveproteins from the cell pellet (left) and culture medium (right) after48 hr of incubation in the presence of ['H] proline.
step in the sequential cleavage of the large procollagenmolecule into typical tropocollagen molecules. It is known thatboth the N-terminus and the C-terminus of the final alchains contain noncollagenous polypeptide appendages inpeptide-bond linkage (6, 18). Alternatively, the appearanceof pro-al chains may be due to nonspecific degradation ofprocollagen as a consequence of homogenization, prolongedextraction at low pH, or other preparation methods. In ourexperiments, the use of Nonidet P-40 detergent to rapidlylyse the cells, in conjunction with the rapid addition ofdenaturing agents, probably minimizes protein degradationthat would arise from proteases and from the shearing forcesof homogenization.The large size of procollagen and its high content of non-
collagenous regions could provide sufficient stretches ofintercollagen regions to allow correct alignment and polarityof the collagenous portions. Once the folding into the triplehelix of tropocollagen occurs, digestion by proteases otherthan specific collagenases would hardly affect the collagenregions (19). The surprising aspect of our results is that theproteolytic step occurs extracellularly; the absence oftropocollagen in the medium suggests that once it is formedthe tropocollagen either quickly aggregates into collagenfibrils or that it may also be rapidly destroyed (accountingfor the dialyzable hydroxyproline).
It has been reported that Dintzis-type experiments showthat the three polypeptide chains of collagen are initiated andsynthesized separately (20). However, close examination ofthe kinetics obtained, particularly the results after the earliestpulses, show considerable nonlinearity of the isotope gradientand allow great latitude in interpretation of the data. More-over, the experimental procedure was designed to select fortropocollagen molecules already present in extracellularfibrous collagen, a quite different approach than ours.The existence of a protropocollagen molecule was postulated
several years ago (21) and some evidence in its support wasfound (22). Moreover, the presence of large collagen-likeproteins circulating in human serum has been well documented(23), and at least one such protein has been well characterized(24); whether these proteins are related to procollagen can-not be ascertained with the available data.
Proc. Nat. Acad. Sci. USA 68 (1971)

2642 Biochemistry: Church et al.
Modification of certain amino-acid side chains in collagenoccurs after peptide-bond formation, analogous to the altera-tion of bases in the precursor of ribosomal RNA (4). Suchmodifications of collagen include hydroxylation of prolineand lysine (25), glycosylation of hydroxylysine (26), and theconversion of e-amino groups into aldehydes (27). In lightof our current findings, it would appear that such modifica-tions occur on the finished procollagen molecule, perhapseven on the extracellular form. In this regard, a recent reportindicates that collagen-aldehyde formation continues in re-constituted collagen fibrils (28), suggesting that enzymaticmodifications may even continue in the collagen fibers.
NOTE ADDED IN PROOFAfter the original submission of this manuscript, severalpapers have been published that report: (a) both a and pre-achains are synthesized in vivo and in vitro [Lazarides, E., andL. N. Lukens, Nature (London), 232, 37 (1971)]; (b) themedium of 3T6 fibroblasts contains a large, collagen-relatedprotein [Ramaley, P. B., and J. Rosenbloom, FEBS Lett., 15,59 (1971) ]; (c) pre-al chains give rise to ae 1 chains in extractsof chick calvaria in culture [Mifller, P. K., E. McGoodwin,and G. R. Martin, Biochem. Biophys. Res. Commun., 44, 110(1971)].We thank Drs. Richard Wang and Howard Green for supplying
the 3T6 fibroblasts. This work was partially supported by NIHGrant AM-12683-03, by a Merck Foundation Faculty Award(Steven E. Pfeiffer), and by the Life Insurance Medical ResearchFund. R. L. C. was a postdoctoral fellow of NIH (CA-52097-01)and M. L. T. was an Established Investigator of the AmericanHeart Association during the course of this work.
1. Dreyer, W. J., and H. Neurath, J. Biol. Chem., 217, 527(1955).
2. Chance, R. E., R. Al. Ellis, and W. W. Bromer, Science, 161,165 (1968).
3. Jacobson, M. F., J. Asso, and D. Baltimore, J. Mol. Biol.,49, 657 (1970).
4. Darnell, J. E., Jr., Bacteriol. Rev., 32, 262 (1968).
5. Piez, K. A., E. A. Eigner, and M. S. Lewis, Biochemistry, 2,58 (1963).
6. Kang, A. H., P. Bornstein, and H. A. Piez, Biochemistry, 6,788 (1967).
7. Borun, T. W., M. D. Scharff, and E. Robbins, Biochim.Biophys. Acta, 149, 302 (1967).
8. Maizel, J. V., Jr., in Fundamental Techniques in Virology, ed.K. Habel and N. P. Salzman (Academic Press, New York,1969), p. 343.
9. Furthmayr, H., and R. Timpl, Anal. Biochem., 41, 510(1971).
10. Peterkofsky, B., and S. Udenfriend, Biochem. Biophys. Res.Commun., 6, 184 (1961).
11. Peterkofsky, B., and R. Diegelmann, Biochemistry, 10, 988(1971).
12. Eastoe, J. E., in Treatise on Collagen, ed. C. M. Ramachan-dran (Academic Press, New York, 1967-1968), Vol. 1, p. 1.
13. Goldberg, B., and H. Green, J. Cell Biol., 22, 227 (1964).14. Fitch, S. M., M. L. R. Harkness, and R. D. Harkness,
Nature (London), 176,173 (1955).15. Levene, C. I., and C. J. Bates, J. Cell Sci., 7, 671 (1970).16. Layman, D. L., E. B. Goodwin, and G. R. Martin, Proc.
Nat. Acad. Sci., USA, 68, 454 (1971).17. Bellamy, G., and P. Bornstein, Proc. Nat. Acad. Sci., USA,
68, 1138 (1971).18. Stark, M., J. Rauterberg, and K. Kuhn, FEBS Lett., 13, 101
(1971).19. Seifter, S., and E. Harper, in Methods in Enzymology, ed.
G. E. Perlmann and L. Lorand (Academic Press, NewYork, 1970), Vol. 19, p. 613.
20. Vuust, J., and K. A. Piez, J. Biol. Chem., 245, 6201 (1970).21. Schmitt, F. O., Bull. N.Y. Acad. Med., 36, 725 (1960).22. Fessler, J. H., and L. A. Smith, in Chemistry and Molecular
Biology of the Intercellular Matrix, ed. E. A. Balazs (Aca-demic Press, New York, 1970), Vol. 1, p. 457.
23. Leroy, E. C., A. Kaplan, S. Undenfriend, and A. Sjoerdsma,J. Biol. Chem., 239, 3350 (1964).
24. Yonemasu, K., R. M. Stroud, W. Miedermeier, and W. T.Butler, Biochem. Biophys. Res. Commun., 43, 1388 (1971).
25. Udenfriend, S., Science, 152, 1335 (1966).26. Spiro, R. G., J. Biol. Chem., 243, 1923 (1967).27. Pinnell, S. R., and G. R. Martin, Proc. Nat. Acad. Sci.,
USA, 61, 708 (1968).28. Deshmukh, A., K. Deshmukh, and M. E. Nimni, Bio-
chemistry, 10, 2337 (1971 ).
Proc. Nat. Acad. Sci. USA 68 (1971)





























